WO1993020185A1 - Method for in vitro proliferation of dendritic cell precursors and their use to produce immunogens - Google Patents
Method for in vitro proliferation of dendritic cell precursors and their use to produce immunogens Download PDFInfo
- Publication number
- WO1993020185A1 WO1993020185A1 PCT/US1993/003141 US9303141W WO9320185A1 WO 1993020185 A1 WO1993020185 A1 WO 1993020185A1 US 9303141 W US9303141 W US 9303141W WO 9320185 A1 WO9320185 A1 WO 9320185A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- cells
- dendritic
- cell
- antigen
- csf
- Prior art date
Links
- 210000004443 dendritic cell Anatomy 0.000 title claims abstract description 581
- 239000002243 precursor Substances 0.000 title claims abstract description 156
- 238000000034 method Methods 0.000 title claims description 182
- 238000000338 in vitro Methods 0.000 title claims description 32
- 230000035755 proliferation Effects 0.000 title description 23
- 108091007433 antigens Proteins 0.000 claims abstract description 301
- 102000036639 antigens Human genes 0.000 claims abstract description 301
- 239000000427 antigen Substances 0.000 claims abstract description 298
- 210000001744 T-lymphocyte Anatomy 0.000 claims abstract description 83
- 230000002062 proliferating effect Effects 0.000 claims abstract description 55
- 238000011282 treatment Methods 0.000 claims abstract description 20
- 229960005486 vaccine Drugs 0.000 claims abstract description 13
- 201000010099 disease Diseases 0.000 claims abstract description 12
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims abstract description 12
- 208000023275 Autoimmune disease Diseases 0.000 claims abstract description 11
- 206010067584 Type 1 diabetes mellitus Diseases 0.000 claims abstract description 5
- 201000006417 multiple sclerosis Diseases 0.000 claims abstract description 4
- 210000004027 cell Anatomy 0.000 claims description 560
- 108010017213 Granulocyte-Macrophage Colony-Stimulating Factor Proteins 0.000 claims description 140
- 210000001185 bone marrow Anatomy 0.000 claims description 79
- 210000004369 blood Anatomy 0.000 claims description 73
- 239000008280 blood Substances 0.000 claims description 73
- 210000001519 tissue Anatomy 0.000 claims description 73
- 239000002609 medium Substances 0.000 claims description 48
- 108091003079 Bovine Serum Albumin Proteins 0.000 claims description 31
- 210000003719 b-lymphocyte Anatomy 0.000 claims description 30
- 108090000623 proteins and genes Proteins 0.000 claims description 30
- 108010017080 Granulocyte Colony-Stimulating Factor Proteins 0.000 claims description 23
- 102000004269 Granulocyte Colony-Stimulating Factor Human genes 0.000 claims description 23
- 239000001963 growth medium Substances 0.000 claims description 23
- 239000000203 mixture Substances 0.000 claims description 23
- 239000012980 RPMI-1640 medium Substances 0.000 claims description 21
- 238000012258 culturing Methods 0.000 claims description 20
- 230000008569 process Effects 0.000 claims description 20
- 102000004457 Granulocyte-Macrophage Colony-Stimulating Factor Human genes 0.000 claims description 18
- 239000012894 fetal calf serum Substances 0.000 claims description 17
- 239000012634 fragment Substances 0.000 claims description 17
- 108060008682 Tumor Necrosis Factor Proteins 0.000 claims description 16
- 102000000852 Tumor Necrosis Factor-alpha Human genes 0.000 claims description 16
- 108010002386 Interleukin-3 Proteins 0.000 claims description 14
- 230000000295 complement effect Effects 0.000 claims description 14
- 239000000758 substrate Substances 0.000 claims description 14
- 210000001616 monocyte Anatomy 0.000 claims description 13
- 239000000126 substance Substances 0.000 claims description 12
- 241001465754 Metazoa Species 0.000 claims description 11
- 230000003053 immunization Effects 0.000 claims description 11
- 241000894006 Bacteria Species 0.000 claims description 10
- 201000008827 tuberculosis Diseases 0.000 claims description 10
- 238000004113 cell culture Methods 0.000 claims description 9
- 210000002966 serum Anatomy 0.000 claims description 9
- MZOFCQQQCNRIBI-VMXHOPILSA-N (3s)-4-[[(2s)-1-[[(2s)-1-[[(1s)-1-carboxy-2-hydroxyethyl]amino]-4-methyl-1-oxopentan-2-yl]amino]-5-(diaminomethylideneamino)-1-oxopentan-2-yl]amino]-3-[[2-[[(2s)-2,6-diaminohexanoyl]amino]acetyl]amino]-4-oxobutanoic acid Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC(O)=O)NC(=O)CNC(=O)[C@@H](N)CCCCN MZOFCQQQCNRIBI-VMXHOPILSA-N 0.000 claims description 8
- 108010002352 Interleukin-1 Proteins 0.000 claims description 8
- 241000282412 Homo Species 0.000 claims description 7
- 230000003612 virological effect Effects 0.000 claims description 6
- 230000002147 killing effect Effects 0.000 claims description 5
- -1 G- CSF Proteins 0.000 claims description 4
- 206010012438 Dermatitis atopic Diseases 0.000 claims description 3
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 claims description 3
- 201000008937 atopic dermatitis Diseases 0.000 claims description 3
- 230000003394 haemopoietic effect Effects 0.000 claims description 3
- 230000011132 hemopoiesis Effects 0.000 claims description 2
- 206010028417 myasthenia gravis Diseases 0.000 claims description 2
- 230000001580 bacterial effect Effects 0.000 claims 2
- 239000013603 viral vector Substances 0.000 claims 1
- 230000001419 dependent effect Effects 0.000 abstract description 20
- 238000004519 manufacturing process Methods 0.000 abstract description 11
- 102100039620 Granulocyte-macrophage colony-stimulating factor Human genes 0.000 description 122
- 230000001464 adherent effect Effects 0.000 description 56
- 241000699666 Mus <mouse, genus> Species 0.000 description 50
- 210000003714 granulocyte Anatomy 0.000 description 45
- 108091054438 MHC class II family Proteins 0.000 description 43
- 210000002540 macrophage Anatomy 0.000 description 43
- 102000043131 MHC class II family Human genes 0.000 description 41
- 210000000612 antigen-presenting cell Anatomy 0.000 description 28
- 239000002245 particle Substances 0.000 description 27
- 108090000765 processed proteins & peptides Proteins 0.000 description 27
- 210000000265 leukocyte Anatomy 0.000 description 26
- 239000000047 product Substances 0.000 description 26
- 102000004169 proteins and genes Human genes 0.000 description 25
- 241000699670 Mus sp. Species 0.000 description 24
- 108700018351 Major Histocompatibility Complex Proteins 0.000 description 23
- 230000028993 immune response Effects 0.000 description 23
- 210000000952 spleen Anatomy 0.000 description 23
- 230000020382 suppression by virus of host antigen processing and presentation of peptide antigen via MHC class I Effects 0.000 description 23
- 230000001965 increasing effect Effects 0.000 description 22
- 238000011161 development Methods 0.000 description 21
- 230000004936 stimulating effect Effects 0.000 description 21
- 102000004196 processed proteins & peptides Human genes 0.000 description 20
- 230000004044 response Effects 0.000 description 20
- 230000018109 developmental process Effects 0.000 description 19
- 238000002474 experimental method Methods 0.000 description 19
- 210000001165 lymph node Anatomy 0.000 description 19
- 102000004127 Cytokines Human genes 0.000 description 17
- 108090000695 Cytokines Proteins 0.000 description 17
- 230000003389 potentiating effect Effects 0.000 description 17
- 102000007651 Macrophage Colony-Stimulating Factor Human genes 0.000 description 16
- 108010046938 Macrophage Colony-Stimulating Factor Proteins 0.000 description 16
- IQFYYKKMVGJFEH-XLPZGREQSA-N Thymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-XLPZGREQSA-N 0.000 description 16
- 230000030741 antigen processing and presentation Effects 0.000 description 16
- 230000006870 function Effects 0.000 description 16
- 238000011065 in-situ storage Methods 0.000 description 16
- 238000002360 preparation method Methods 0.000 description 15
- 229940098773 bovine serum albumin Drugs 0.000 description 14
- 230000002163 immunogen Effects 0.000 description 14
- 230000000242 pagocytic effect Effects 0.000 description 14
- 239000002671 adjuvant Substances 0.000 description 13
- 238000002372 labelling Methods 0.000 description 13
- 244000052616 bacterial pathogen Species 0.000 description 12
- 239000004816 latex Substances 0.000 description 12
- 229920000126 latex Polymers 0.000 description 12
- 230000035800 maturation Effects 0.000 description 12
- 239000002356 single layer Substances 0.000 description 12
- 238000010186 staining Methods 0.000 description 12
- NLXLAEXVIDQMFP-UHFFFAOYSA-N Ammonia chloride Chemical compound [NH4+].[Cl-] NLXLAEXVIDQMFP-UHFFFAOYSA-N 0.000 description 11
- 102000000646 Interleukin-3 Human genes 0.000 description 11
- 241001529936 Murinae Species 0.000 description 11
- 230000000890 antigenic effect Effects 0.000 description 11
- 239000008187 granular material Substances 0.000 description 11
- 238000001727 in vivo Methods 0.000 description 11
- 238000004062 sedimentation Methods 0.000 description 11
- CSCPPACGZOOCGX-UHFFFAOYSA-N Acetone Chemical compound CC(C)=O CSCPPACGZOOCGX-UHFFFAOYSA-N 0.000 description 10
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 10
- 206010057249 Phagocytosis Diseases 0.000 description 10
- 229910052799 carbon Inorganic materials 0.000 description 10
- 238000006243 chemical reaction Methods 0.000 description 10
- 230000003308 immunostimulating effect Effects 0.000 description 10
- 230000008782 phagocytosis Effects 0.000 description 10
- 239000000725 suspension Substances 0.000 description 10
- NHBKXEKEPDILRR-UHFFFAOYSA-N 2,3-bis(butanoylsulfanyl)propyl butanoate Chemical compound CCCC(=O)OCC(SC(=O)CCC)CSC(=O)CCC NHBKXEKEPDILRR-UHFFFAOYSA-N 0.000 description 9
- 102000043129 MHC class I family Human genes 0.000 description 9
- 108091054437 MHC class I family Proteins 0.000 description 9
- 230000015572 biosynthetic process Effects 0.000 description 9
- 230000000694 effects Effects 0.000 description 9
- 210000003426 epidermal langerhans cell Anatomy 0.000 description 9
- 210000003743 erythrocyte Anatomy 0.000 description 9
- 230000003834 intracellular effect Effects 0.000 description 9
- 210000002751 lymph Anatomy 0.000 description 9
- 102000013415 peroxidase activity proteins Human genes 0.000 description 9
- 108040007629 peroxidase activity proteins Proteins 0.000 description 9
- 239000002253 acid Substances 0.000 description 8
- 238000004458 analytical method Methods 0.000 description 8
- 238000000376 autoradiography Methods 0.000 description 8
- 239000012737 fresh medium Substances 0.000 description 8
- 208000015181 infectious disease Diseases 0.000 description 8
- 210000001911 interdigitating cell Anatomy 0.000 description 8
- 239000004033 plastic Substances 0.000 description 8
- 101150052863 THY1 gene Proteins 0.000 description 7
- 230000002121 endocytic effect Effects 0.000 description 7
- 210000004700 fetal blood Anatomy 0.000 description 7
- 210000001821 langerhans cell Anatomy 0.000 description 7
- 230000037361 pathway Effects 0.000 description 7
- 230000002093 peripheral effect Effects 0.000 description 7
- 238000012545 processing Methods 0.000 description 7
- 238000000926 separation method Methods 0.000 description 7
- 239000013598 vector Substances 0.000 description 7
- DGVVWUTYPXICAM-UHFFFAOYSA-N β‐Mercaptoethanol Chemical compound OCCS DGVVWUTYPXICAM-UHFFFAOYSA-N 0.000 description 7
- 238000011740 C57BL/6 mouse Methods 0.000 description 6
- 241000699800 Cricetinae Species 0.000 description 6
- 238000011765 DBA/2 mouse Methods 0.000 description 6
- BAQCROVBDNBEEB-UBYUBLNFSA-N Metrizamide Chemical compound CC(=O)N(C)C1=C(I)C(NC(C)=O)=C(I)C(C(=O)N[C@@H]2[C@H]([C@H](O)[C@@H](CO)OC2O)O)=C1I BAQCROVBDNBEEB-UBYUBLNFSA-N 0.000 description 6
- 101000884281 Rattus norvegicus Signal transducer CD24 Proteins 0.000 description 6
- 230000000735 allogeneic effect Effects 0.000 description 6
- 230000009851 immunogenic response Effects 0.000 description 6
- 238000002347 injection Methods 0.000 description 6
- 239000007924 injection Substances 0.000 description 6
- 210000004698 lymphocyte Anatomy 0.000 description 6
- 210000005210 lymphoid organ Anatomy 0.000 description 6
- 239000000463 material Substances 0.000 description 6
- 229960000554 metrizamide Drugs 0.000 description 6
- 210000001539 phagocyte Anatomy 0.000 description 6
- 210000000130 stem cell Anatomy 0.000 description 6
- 210000003934 vacuole Anatomy 0.000 description 6
- 102000000589 Interleukin-1 Human genes 0.000 description 5
- 241000283973 Oryctolagus cuniculus Species 0.000 description 5
- 230000005867 T cell response Effects 0.000 description 5
- 241000700605 Viruses Species 0.000 description 5
- 150000001413 amino acids Chemical class 0.000 description 5
- 235000019270 ammonium chloride Nutrition 0.000 description 5
- 210000000988 bone and bone Anatomy 0.000 description 5
- 238000001493 electron microscopy Methods 0.000 description 5
- 210000002950 fibroblast Anatomy 0.000 description 5
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 5
- 230000005847 immunogenicity Effects 0.000 description 5
- 238000004091 panning Methods 0.000 description 5
- 239000002953 phosphate buffered saline Substances 0.000 description 5
- 230000002000 scavenging effect Effects 0.000 description 5
- 230000004083 survival effect Effects 0.000 description 5
- 238000012546 transfer Methods 0.000 description 5
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 4
- 102100032912 CD44 antigen Human genes 0.000 description 4
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 4
- 229930182566 Gentamicin Natural products 0.000 description 4
- CEAZRRDELHUEMR-URQXQFDESA-N Gentamicin Chemical compound O1[C@H](C(C)NC)CC[C@@H](N)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](NC)[C@@](C)(O)CO2)O)[C@H](N)C[C@@H]1N CEAZRRDELHUEMR-URQXQFDESA-N 0.000 description 4
- 101000868273 Homo sapiens CD44 antigen Proteins 0.000 description 4
- 108010001336 Horseradish Peroxidase Proteins 0.000 description 4
- 102000004889 Interleukin-6 Human genes 0.000 description 4
- 108090001005 Interleukin-6 Proteins 0.000 description 4
- 206010028980 Neoplasm Diseases 0.000 description 4
- 239000006146 Roswell Park Memorial Institute medium Substances 0.000 description 4
- 238000003556 assay Methods 0.000 description 4
- 210000000601 blood cell Anatomy 0.000 description 4
- 238000009640 blood culture Methods 0.000 description 4
- 230000001413 cellular effect Effects 0.000 description 4
- 229960002518 gentamicin Drugs 0.000 description 4
- 150000004676 glycans Chemical class 0.000 description 4
- 230000012010 growth Effects 0.000 description 4
- 238000003306 harvesting Methods 0.000 description 4
- 238000002649 immunization Methods 0.000 description 4
- 230000001939 inductive effect Effects 0.000 description 4
- 210000003712 lysosome Anatomy 0.000 description 4
- 230000001868 lysosomic effect Effects 0.000 description 4
- 239000003550 marker Substances 0.000 description 4
- 230000000813 microbial effect Effects 0.000 description 4
- 210000000056 organ Anatomy 0.000 description 4
- 229920001282 polysaccharide Polymers 0.000 description 4
- 239000005017 polysaccharide Substances 0.000 description 4
- 230000004043 responsiveness Effects 0.000 description 4
- 210000003491 skin Anatomy 0.000 description 4
- 239000007787 solid Substances 0.000 description 4
- 210000004989 spleen cell Anatomy 0.000 description 4
- 230000003393 splenic effect Effects 0.000 description 4
- 230000000638 stimulation Effects 0.000 description 4
- 230000001225 therapeutic effect Effects 0.000 description 4
- 210000001541 thymus gland Anatomy 0.000 description 4
- 230000035899 viability Effects 0.000 description 4
- 238000005406 washing Methods 0.000 description 4
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Chemical compound O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 4
- 241000283707 Capra Species 0.000 description 3
- 108010051152 Carboxylesterase Proteins 0.000 description 3
- 102000013392 Carboxylesterase Human genes 0.000 description 3
- 102000009109 Fc receptors Human genes 0.000 description 3
- 108010087819 Fc receptors Proteins 0.000 description 3
- 102100031573 Hematopoietic progenitor cell antigen CD34 Human genes 0.000 description 3
- 102000018713 Histocompatibility Antigens Class II Human genes 0.000 description 3
- 101000777663 Homo sapiens Hematopoietic progenitor cell antigen CD34 Proteins 0.000 description 3
- 101000738771 Homo sapiens Receptor-type tyrosine-protein phosphatase C Proteins 0.000 description 3
- 102000009490 IgG Receptors Human genes 0.000 description 3
- 108010073807 IgG Receptors Proteins 0.000 description 3
- 206010025323 Lymphomas Diseases 0.000 description 3
- 239000004677 Nylon Substances 0.000 description 3
- 102100037422 Receptor-type tyrosine-protein phosphatase C Human genes 0.000 description 3
- 230000018199 S phase Effects 0.000 description 3
- 230000008901 benefit Effects 0.000 description 3
- 230000011712 cell development Effects 0.000 description 3
- 230000003915 cell function Effects 0.000 description 3
- 230000010261 cell growth Effects 0.000 description 3
- 210000000805 cytoplasm Anatomy 0.000 description 3
- 230000001086 cytosolic effect Effects 0.000 description 3
- 238000010790 dilution Methods 0.000 description 3
- 239000012895 dilution Substances 0.000 description 3
- 239000000839 emulsion Substances 0.000 description 3
- 239000011521 glass Substances 0.000 description 3
- 210000000987 immune system Anatomy 0.000 description 3
- 108010028930 invariant chain Proteins 0.000 description 3
- 238000002955 isolation Methods 0.000 description 3
- 208000032839 leukemia Diseases 0.000 description 3
- 210000004379 membrane Anatomy 0.000 description 3
- 239000012528 membrane Substances 0.000 description 3
- 238000000386 microscopy Methods 0.000 description 3
- 230000005012 migration Effects 0.000 description 3
- 238000013508 migration Methods 0.000 description 3
- 210000000440 neutrophil Anatomy 0.000 description 3
- 229920001778 nylon Polymers 0.000 description 3
- 239000013618 particulate matter Substances 0.000 description 3
- 239000008188 pellet Substances 0.000 description 3
- 210000003024 peritoneal macrophage Anatomy 0.000 description 3
- 230000002265 prevention Effects 0.000 description 3
- 230000037452 priming Effects 0.000 description 3
- 230000000750 progressive effect Effects 0.000 description 3
- 230000001177 retroviral effect Effects 0.000 description 3
- 210000002536 stromal cell Anatomy 0.000 description 3
- 238000003786 synthesis reaction Methods 0.000 description 3
- 210000002303 tibia Anatomy 0.000 description 3
- NALREUIWICQLPS-UHFFFAOYSA-N 7-imino-n,n-dimethylphenothiazin-3-amine;hydrochloride Chemical compound [Cl-].C1=C(N)C=C2SC3=CC(=[N+](C)C)C=CC3=NC2=C1 NALREUIWICQLPS-UHFFFAOYSA-N 0.000 description 2
- 210000001239 CD8-positive, alpha-beta cytotoxic T lymphocyte Anatomy 0.000 description 2
- 108010071942 Colony-Stimulating Factors Proteins 0.000 description 2
- WSFSSNUMVMOOMR-UHFFFAOYSA-N Formaldehyde Chemical compound O=C WSFSSNUMVMOOMR-UHFFFAOYSA-N 0.000 description 2
- SXRSQZLOMIGNAQ-UHFFFAOYSA-N Glutaraldehyde Chemical compound O=CCCCC=O SXRSQZLOMIGNAQ-UHFFFAOYSA-N 0.000 description 2
- 102100036242 HLA class II histocompatibility antigen, DQ alpha 2 chain Human genes 0.000 description 2
- 102000006354 HLA-DR Antigens Human genes 0.000 description 2
- 108010058597 HLA-DR Antigens Proteins 0.000 description 2
- 239000012981 Hank's balanced salt solution Substances 0.000 description 2
- HTTJABKRGRZYRN-UHFFFAOYSA-N Heparin Chemical compound OC1C(NC(=O)C)C(O)OC(COS(O)(=O)=O)C1OC1C(OS(O)(=O)=O)C(O)C(OC2C(C(OS(O)(=O)=O)C(OC3C(C(O)C(O)C(O3)C(O)=O)OS(O)(=O)=O)C(CO)O2)NS(O)(=O)=O)C(C(O)=O)O1 HTTJABKRGRZYRN-UHFFFAOYSA-N 0.000 description 2
- 101000930801 Homo sapiens HLA class II histocompatibility antigen, DQ alpha 2 chain Proteins 0.000 description 2
- 102100026878 Interleukin-2 receptor subunit alpha Human genes 0.000 description 2
- 102000015696 Interleukins Human genes 0.000 description 2
- 108010063738 Interleukins Proteins 0.000 description 2
- 101710160107 Outer membrane protein A Proteins 0.000 description 2
- 102000007079 Peptide Fragments Human genes 0.000 description 2
- 108010033276 Peptide Fragments Proteins 0.000 description 2
- 230000037453 T cell priming Effects 0.000 description 2
- 108010067390 Viral Proteins Proteins 0.000 description 2
- 230000002378 acidificating effect Effects 0.000 description 2
- 230000004913 activation Effects 0.000 description 2
- 230000003698 anagen phase Effects 0.000 description 2
- 238000013459 approach Methods 0.000 description 2
- 230000002902 bimodal effect Effects 0.000 description 2
- 230000004071 biological effect Effects 0.000 description 2
- 229960002685 biotin Drugs 0.000 description 2
- 235000020958 biotin Nutrition 0.000 description 2
- 239000011616 biotin Substances 0.000 description 2
- 210000005208 blood dendritic cell Anatomy 0.000 description 2
- 210000002798 bone marrow cell Anatomy 0.000 description 2
- 230000015556 catabolic process Effects 0.000 description 2
- 230000004663 cell proliferation Effects 0.000 description 2
- 238000005119 centrifugation Methods 0.000 description 2
- 239000003153 chemical reaction reagent Substances 0.000 description 2
- 239000000356 contaminant Substances 0.000 description 2
- 210000001151 cytotoxic T lymphocyte Anatomy 0.000 description 2
- 238000006731 degradation reaction Methods 0.000 description 2
- 238000013461 design Methods 0.000 description 2
- 230000004069 differentiation Effects 0.000 description 2
- 230000029087 digestion Effects 0.000 description 2
- 238000009826 distribution Methods 0.000 description 2
- 229940079593 drug Drugs 0.000 description 2
- 239000003814 drug Substances 0.000 description 2
- 230000012202 endocytosis Effects 0.000 description 2
- 210000002615 epidermis Anatomy 0.000 description 2
- 238000000605 extraction Methods 0.000 description 2
- 230000001605 fetal effect Effects 0.000 description 2
- 238000007667 floating Methods 0.000 description 2
- MHMNJMPURVTYEJ-UHFFFAOYSA-N fluorescein-5-isothiocyanate Chemical compound O1C(=O)C2=CC(N=C=S)=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 MHMNJMPURVTYEJ-UHFFFAOYSA-N 0.000 description 2
- 238000013467 fragmentation Methods 0.000 description 2
- 238000006062 fragmentation reaction Methods 0.000 description 2
- 239000003102 growth factor Substances 0.000 description 2
- 210000002443 helper t lymphocyte Anatomy 0.000 description 2
- 229960002897 heparin Drugs 0.000 description 2
- 229920000669 heparin Polymers 0.000 description 2
- 239000005556 hormone Substances 0.000 description 2
- 229940088597 hormone Drugs 0.000 description 2
- 238000009169 immunotherapy Methods 0.000 description 2
- 230000002779 inactivation Effects 0.000 description 2
- CGIGDMFJXJATDK-UHFFFAOYSA-N indomethacin Chemical compound CC1=C(CC(O)=O)C2=CC(OC)=CC=C2N1C(=O)C1=CC=C(Cl)C=C1 CGIGDMFJXJATDK-UHFFFAOYSA-N 0.000 description 2
- 206010022000 influenza Diseases 0.000 description 2
- 230000000977 initiatory effect Effects 0.000 description 2
- 108010044426 integrins Proteins 0.000 description 2
- 102000006495 integrins Human genes 0.000 description 2
- 230000003993 interaction Effects 0.000 description 2
- 210000005133 interdigitating dendritic cell Anatomy 0.000 description 2
- 229940076264 interleukin-3 Drugs 0.000 description 2
- 229940100601 interleukin-6 Drugs 0.000 description 2
- 238000001990 intravenous administration Methods 0.000 description 2
- 239000000644 isotonic solution Substances 0.000 description 2
- 239000007788 liquid Substances 0.000 description 2
- 238000009630 liquid culture Methods 0.000 description 2
- 210000003563 lymphoid tissue Anatomy 0.000 description 2
- 235000012054 meals Nutrition 0.000 description 2
- 230000007246 mechanism Effects 0.000 description 2
- 238000012544 monitoring process Methods 0.000 description 2
- 210000005087 mononuclear cell Anatomy 0.000 description 2
- 210000002864 mononuclear phagocyte Anatomy 0.000 description 2
- 210000000066 myeloid cell Anatomy 0.000 description 2
- 230000005868 ontogenesis Effects 0.000 description 2
- 210000003463 organelle Anatomy 0.000 description 2
- 230000001590 oxidative effect Effects 0.000 description 2
- 239000008194 pharmaceutical composition Substances 0.000 description 2
- 230000009696 proliferative response Effects 0.000 description 2
- 230000003134 recirculating effect Effects 0.000 description 2
- 239000000523 sample Substances 0.000 description 2
- 230000001235 sensitizing effect Effects 0.000 description 2
- YEENEYXBHNNNGV-XEHWZWQGSA-M sodium;3-acetamido-5-[acetyl(methyl)amino]-2,4,6-triiodobenzoate;(2r,3r,4s,5s,6r)-2-[(2r,3s,4s,5r)-3,4-dihydroxy-2,5-bis(hydroxymethyl)oxolan-2-yl]oxy-6-(hydroxymethyl)oxane-3,4,5-triol Chemical compound [Na+].CC(=O)N(C)C1=C(I)C(NC(C)=O)=C(I)C(C([O-])=O)=C1I.O[C@H]1[C@H](O)[C@@H](CO)O[C@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 YEENEYXBHNNNGV-XEHWZWQGSA-M 0.000 description 2
- 241000894007 species Species 0.000 description 2
- 238000010561 standard procedure Methods 0.000 description 2
- UCSJYZPVAKXKNQ-HZYVHMACSA-N streptomycin Chemical compound CN[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@](C=O)(O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](NC(N)=N)[C@H](O)[C@@H](NC(N)=N)[C@H](O)[C@H]1O UCSJYZPVAKXKNQ-HZYVHMACSA-N 0.000 description 2
- 239000006228 supernatant Substances 0.000 description 2
- 239000013589 supplement Substances 0.000 description 2
- 206010056873 tertiary syphilis Diseases 0.000 description 2
- 238000012360 testing method Methods 0.000 description 2
- 230000002992 thymic effect Effects 0.000 description 2
- 231100000331 toxic Toxicity 0.000 description 2
- 230000002588 toxic effect Effects 0.000 description 2
- 230000007306 turnover Effects 0.000 description 2
- 210000000689 upper leg Anatomy 0.000 description 2
- 210000005135 veiled cell Anatomy 0.000 description 2
- WWYNJERNGUHSAO-XUDSTZEESA-N (+)-Norgestrel Chemical compound O=C1CC[C@@H]2[C@H]3CC[C@](CC)([C@](CC4)(O)C#C)[C@@H]4[C@@H]3CCC2=C1 WWYNJERNGUHSAO-XUDSTZEESA-N 0.000 description 1
- IQFYYKKMVGJFEH-OFKYTIFKSA-N 1-[(2r,4s,5r)-4-hydroxy-5-(tritiooxymethyl)oxolan-2-yl]-5-methylpyrimidine-2,4-dione Chemical compound C1[C@H](O)[C@@H](CO[3H])O[C@H]1N1C(=O)NC(=O)C(C)=C1 IQFYYKKMVGJFEH-OFKYTIFKSA-N 0.000 description 1
- GOZMBJCYMQQACI-UHFFFAOYSA-N 6,7-dimethyl-3-[[methyl-[2-[methyl-[[1-[3-(trifluoromethyl)phenyl]indol-3-yl]methyl]amino]ethyl]amino]methyl]chromen-4-one;dihydrochloride Chemical compound Cl.Cl.C=1OC2=CC(C)=C(C)C=C2C(=O)C=1CN(C)CCN(C)CC(C1=CC=CC=C11)=CN1C1=CC=CC(C(F)(F)F)=C1 GOZMBJCYMQQACI-UHFFFAOYSA-N 0.000 description 1
- BZTDTCNHAFUJOG-UHFFFAOYSA-N 6-carboxyfluorescein Chemical compound C12=CC=C(O)C=C2OC2=CC(O)=CC=C2C11OC(=O)C2=CC=C(C(=O)O)C=C21 BZTDTCNHAFUJOG-UHFFFAOYSA-N 0.000 description 1
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 1
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 1
- 108090001008 Avidin Proteins 0.000 description 1
- 238000011752 CBA/J (JAX™ mouse strain) Methods 0.000 description 1
- 101100289995 Caenorhabditis elegans mac-1 gene Proteins 0.000 description 1
- 102000008186 Collagen Human genes 0.000 description 1
- 108010035532 Collagen Proteins 0.000 description 1
- 208000035473 Communicable disease Diseases 0.000 description 1
- 108010060123 Conjugate Vaccines Proteins 0.000 description 1
- 230000006820 DNA synthesis Effects 0.000 description 1
- 206010012442 Dermatitis contact Diseases 0.000 description 1
- 241001061260 Emmelichthys struhsakeri Species 0.000 description 1
- 108090000371 Esterases Proteins 0.000 description 1
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 1
- 108050001049 Extracellular proteins Proteins 0.000 description 1
- 238000002738 Giemsa staining Methods 0.000 description 1
- 241000720950 Gluta Species 0.000 description 1
- 102000016355 Granulocyte-Macrophage Colony-Stimulating Factor Receptors Human genes 0.000 description 1
- 108010092372 Granulocyte-Macrophage Colony-Stimulating Factor Receptors Proteins 0.000 description 1
- 102000008949 Histocompatibility Antigens Class I Human genes 0.000 description 1
- 108010088652 Histocompatibility Antigens Class I Proteins 0.000 description 1
- 108010027412 Histocompatibility Antigens Class II Proteins 0.000 description 1
- 101000935040 Homo sapiens Integrin beta-2 Proteins 0.000 description 1
- 101001057504 Homo sapiens Interferon-stimulated gene 20 kDa protein Proteins 0.000 description 1
- 101001055144 Homo sapiens Interleukin-2 receptor subunit alpha Proteins 0.000 description 1
- 101000946889 Homo sapiens Monocyte differentiation antigen CD14 Proteins 0.000 description 1
- 108090000144 Human Proteins Proteins 0.000 description 1
- 102000003839 Human Proteins Human genes 0.000 description 1
- 108091006905 Human Serum Albumin Proteins 0.000 description 1
- 102000008100 Human Serum Albumin Human genes 0.000 description 1
- 102000004157 Hydrolases Human genes 0.000 description 1
- 108090000604 Hydrolases Proteins 0.000 description 1
- 102100025390 Integrin beta-2 Human genes 0.000 description 1
- 102100020881 Interleukin-1 alpha Human genes 0.000 description 1
- 102000003816 Interleukin-13 Human genes 0.000 description 1
- 108090000176 Interleukin-13 Proteins 0.000 description 1
- 108010082786 Interleukin-1alpha Proteins 0.000 description 1
- 108010032774 Interleukin-2 Receptor alpha Subunit Proteins 0.000 description 1
- 102000007351 Interleukin-2 Receptor alpha Subunit Human genes 0.000 description 1
- 108010038453 Interleukin-2 Receptors Proteins 0.000 description 1
- 102000010789 Interleukin-2 Receptors Human genes 0.000 description 1
- 208000030729 Juvenile myasthenia gravis Diseases 0.000 description 1
- 108010013709 Leukocyte Common Antigens Proteins 0.000 description 1
- 102000017095 Leukocyte Common Antigens Human genes 0.000 description 1
- 108010030317 Macrophage-1 Antigen Proteins 0.000 description 1
- 102100025136 Macrosialin Human genes 0.000 description 1
- 101710156482 Macrosialin Proteins 0.000 description 1
- 241000124008 Mammalia Species 0.000 description 1
- 101150076359 Mhc gene Proteins 0.000 description 1
- 102000003979 Mineralocorticoid Receptors Human genes 0.000 description 1
- 108090000375 Mineralocorticoid Receptors Proteins 0.000 description 1
- 102100035877 Monocyte differentiation antigen CD14 Human genes 0.000 description 1
- 101000746372 Mus musculus Granulocyte-macrophage colony-stimulating factor Proteins 0.000 description 1
- 241000186359 Mycobacterium Species 0.000 description 1
- 108091028043 Nucleic acid sequence Proteins 0.000 description 1
- 240000007594 Oryza sativa Species 0.000 description 1
- 208000012868 Overgrowth Diseases 0.000 description 1
- 241001494479 Pecora Species 0.000 description 1
- 229930182555 Penicillin Natural products 0.000 description 1
- JGSARLDLIJGVTE-MBNYWOFBSA-N Penicillin G Chemical compound N([C@H]1[C@H]2SC([C@@H](N2C1=O)C(O)=O)(C)C)C(=O)CC1=CC=CC=C1 JGSARLDLIJGVTE-MBNYWOFBSA-N 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 208000034189 Sclerosis Diseases 0.000 description 1
- 108010032838 Sialoglycoproteins Proteins 0.000 description 1
- 102000007365 Sialoglycoproteins Human genes 0.000 description 1
- 241000282887 Suidae Species 0.000 description 1
- 206010052779 Transplant rejections Diseases 0.000 description 1
- SXEHKFHPFVVDIR-UHFFFAOYSA-N [4-(4-hydrazinylphenyl)phenyl]hydrazine Chemical compound C1=CC(NN)=CC=C1C1=CC=C(NN)C=C1 SXEHKFHPFVVDIR-UHFFFAOYSA-N 0.000 description 1
- GPKUGWDQUVWHIC-UHFFFAOYSA-N [4-(4-hydrazinylphenyl)phenyl]hydrazine tetrahydrochloride Chemical compound Cl.Cl.Cl.Cl.NNC1=CC=C(C=C1)C1=CC=C(NN)C=C1 GPKUGWDQUVWHIC-UHFFFAOYSA-N 0.000 description 1
- 230000003213 activating effect Effects 0.000 description 1
- 230000001154 acute effect Effects 0.000 description 1
- 239000013566 allergen Substances 0.000 description 1
- 229940037003 alum Drugs 0.000 description 1
- 210000003484 anatomy Anatomy 0.000 description 1
- 239000003242 anti bacterial agent Substances 0.000 description 1
- 230000001147 anti-toxic effect Effects 0.000 description 1
- 229940088710 antibiotic agent Drugs 0.000 description 1
- 230000005875 antibody response Effects 0.000 description 1
- 239000003146 anticoagulant agent Substances 0.000 description 1
- 229940127219 anticoagulant drug Drugs 0.000 description 1
- 238000000149 argon plasma sintering Methods 0.000 description 1
- JPIYZTWMUGTEHX-UHFFFAOYSA-N auramine O free base Chemical compound C1=CC(N(C)C)=CC=C1C(=N)C1=CC=C(N(C)C)C=C1 JPIYZTWMUGTEHX-UHFFFAOYSA-N 0.000 description 1
- 230000006472 autoimmune response Effects 0.000 description 1
- 229960000190 bacillus calmette–guérin vaccine Drugs 0.000 description 1
- 239000003855 balanced salt solution Substances 0.000 description 1
- 239000011324 bead Substances 0.000 description 1
- 230000000975 bioactive effect Effects 0.000 description 1
- 238000002306 biochemical method Methods 0.000 description 1
- 230000033228 biological regulation Effects 0.000 description 1
- 230000001851 biosynthetic effect Effects 0.000 description 1
- 230000000903 blocking effect Effects 0.000 description 1
- 150000001720 carbohydrates Chemical class 0.000 description 1
- 230000000747 cardiac effect Effects 0.000 description 1
- 238000012754 cardiac puncture Methods 0.000 description 1
- 210000001715 carotid artery Anatomy 0.000 description 1
- 239000000969 carrier Substances 0.000 description 1
- 230000021164 cell adhesion Effects 0.000 description 1
- 210000005056 cell body Anatomy 0.000 description 1
- 230000022131 cell cycle Effects 0.000 description 1
- 230000024245 cell differentiation Effects 0.000 description 1
- 239000006285 cell suspension Substances 0.000 description 1
- 230000003833 cell viability Effects 0.000 description 1
- 230000017455 cell-cell adhesion Effects 0.000 description 1
- 230000007969 cellular immunity Effects 0.000 description 1
- 238000012512 characterization method Methods 0.000 description 1
- 238000002512 chemotherapy Methods 0.000 description 1
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 1
- 229920001436 collagen Polymers 0.000 description 1
- 239000000084 colloidal system Substances 0.000 description 1
- 230000001332 colony forming effect Effects 0.000 description 1
- 229940031670 conjugate vaccine Drugs 0.000 description 1
- 238000011441 consolidation chemotherapy Methods 0.000 description 1
- 239000000470 constituent Substances 0.000 description 1
- 238000010276 construction Methods 0.000 description 1
- 208000010247 contact dermatitis Diseases 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 230000002596 correlated effect Effects 0.000 description 1
- 230000000875 corresponding effect Effects 0.000 description 1
- 239000012228 culture supernatant Substances 0.000 description 1
- 230000000093 cytochemical effect Effects 0.000 description 1
- 230000002380 cytological effect Effects 0.000 description 1
- 230000009089 cytolysis Effects 0.000 description 1
- 230000001461 cytolytic effect Effects 0.000 description 1
- 238000012303 cytoplasmic staining Methods 0.000 description 1
- 229940127089 cytotoxic agent Drugs 0.000 description 1
- 239000002254 cytotoxic agent Substances 0.000 description 1
- 231100000599 cytotoxic agent Toxicity 0.000 description 1
- 210000004544 dc2 Anatomy 0.000 description 1
- 230000004041 dendritic cell maturation Effects 0.000 description 1
- 230000000779 depleting effect Effects 0.000 description 1
- 239000002274 desiccant Substances 0.000 description 1
- 239000003599 detergent Substances 0.000 description 1
- 238000010586 diagram Methods 0.000 description 1
- 230000001079 digestive effect Effects 0.000 description 1
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 1
- 238000007598 dipping method Methods 0.000 description 1
- 230000008034 disappearance Effects 0.000 description 1
- 230000006806 disease prevention Effects 0.000 description 1
- 239000012153 distilled water Substances 0.000 description 1
- 208000015355 drug-resistant tuberculosis Diseases 0.000 description 1
- 230000009977 dual effect Effects 0.000 description 1
- 239000012645 endogenous antigen Substances 0.000 description 1
- 210000001163 endosome Anatomy 0.000 description 1
- 230000002708 enhancing effect Effects 0.000 description 1
- 210000000981 epithelium Anatomy 0.000 description 1
- 210000003414 extremity Anatomy 0.000 description 1
- 230000035611 feeding Effects 0.000 description 1
- 230000004927 fusion Effects 0.000 description 1
- 238000001415 gene therapy Methods 0.000 description 1
- 230000002068 genetic effect Effects 0.000 description 1
- 210000004565 granule cell Anatomy 0.000 description 1
- 230000005484 gravity Effects 0.000 description 1
- 230000036541 health Effects 0.000 description 1
- VKYKSIONXSXAKP-UHFFFAOYSA-N hexamethylenetetramine Chemical group C1N(C2)CN3CN1CN2C3 VKYKSIONXSXAKP-UHFFFAOYSA-N 0.000 description 1
- 210000004408 hybridoma Anatomy 0.000 description 1
- 230000001900 immune effect Effects 0.000 description 1
- 230000007235 immunity generation Effects 0.000 description 1
- 230000001976 improved effect Effects 0.000 description 1
- 230000000937 inactivator Effects 0.000 description 1
- 238000011534 incubation Methods 0.000 description 1
- 229960000905 indomethacin Drugs 0.000 description 1
- 239000000411 inducer Substances 0.000 description 1
- 239000012678 infectious agent Substances 0.000 description 1
- 230000002458 infectious effect Effects 0.000 description 1
- 230000002757 inflammatory effect Effects 0.000 description 1
- 238000001802 infusion Methods 0.000 description 1
- 230000002401 inhibitory effect Effects 0.000 description 1
- 239000002054 inoculum Substances 0.000 description 1
- 238000003780 insertion Methods 0.000 description 1
- 230000037431 insertion Effects 0.000 description 1
- 229940047122 interleukins Drugs 0.000 description 1
- 210000005204 intestinal dendritic cell Anatomy 0.000 description 1
- 238000007918 intramuscular administration Methods 0.000 description 1
- 238000007912 intraperitoneal administration Methods 0.000 description 1
- 239000010410 layer Substances 0.000 description 1
- 230000021633 leukocyte mediated immunity Effects 0.000 description 1
- 239000003446 ligand Substances 0.000 description 1
- 210000004185 liver Anatomy 0.000 description 1
- 230000004807 localization Effects 0.000 description 1
- 230000033001 locomotion Effects 0.000 description 1
- 210000003141 lower extremity Anatomy 0.000 description 1
- 230000002132 lysosomal effect Effects 0.000 description 1
- 230000014759 maintenance of location Effects 0.000 description 1
- 230000010534 mechanism of action Effects 0.000 description 1
- 230000001404 mediated effect Effects 0.000 description 1
- 210000003071 memory t lymphocyte Anatomy 0.000 description 1
- 244000005700 microbiome Species 0.000 description 1
- 210000003470 mitochondria Anatomy 0.000 description 1
- 230000011278 mitosis Effects 0.000 description 1
- 230000000394 mitotic effect Effects 0.000 description 1
- ZAHQPTJLOCWVPG-UHFFFAOYSA-N mitoxantrone dihydrochloride Chemical compound Cl.Cl.O=C1C2=C(O)C=CC(O)=C2C(=O)C2=C1C(NCCNCCO)=CC=C2NCCNCCO ZAHQPTJLOCWVPG-UHFFFAOYSA-N 0.000 description 1
- 238000002156 mixing Methods 0.000 description 1
- 230000004048 modification Effects 0.000 description 1
- 238000012986 modification Methods 0.000 description 1
- 239000003068 molecular probe Substances 0.000 description 1
- 210000000865 mononuclear phagocyte system Anatomy 0.000 description 1
- 230000004899 motility Effects 0.000 description 1
- 201000009671 multidrug-resistant tuberculosis Diseases 0.000 description 1
- 210000002487 multivesicular body Anatomy 0.000 description 1
- 210000003205 muscle Anatomy 0.000 description 1
- 210000003643 myeloid progenitor cell Anatomy 0.000 description 1
- VGKONPUVOVVNSU-UHFFFAOYSA-N naphthalen-1-yl acetate Chemical compound C1=CC=C2C(OC(=O)C)=CC=CC2=C1 VGKONPUVOVVNSU-UHFFFAOYSA-N 0.000 description 1
- 210000000822 natural killer cell Anatomy 0.000 description 1
- 235000015097 nutrients Nutrition 0.000 description 1
- 230000008506 pathogenesis Effects 0.000 description 1
- 230000001717 pathogenic effect Effects 0.000 description 1
- 229940049954 penicillin Drugs 0.000 description 1
- 210000005259 peripheral blood Anatomy 0.000 description 1
- 239000011886 peripheral blood Substances 0.000 description 1
- 210000003200 peritoneal cavity Anatomy 0.000 description 1
- 229940021222 peritoneal dialysis isotonic solution Drugs 0.000 description 1
- 210000001986 peyer's patch Anatomy 0.000 description 1
- 230000000704 physical effect Effects 0.000 description 1
- 239000002504 physiological saline solution Substances 0.000 description 1
- 229920000729 poly(L-lysine) polymer Polymers 0.000 description 1
- 239000002244 precipitate Substances 0.000 description 1
- 238000001556 precipitation Methods 0.000 description 1
- 210000004986 primary T-cell Anatomy 0.000 description 1
- 210000005238 principal cell Anatomy 0.000 description 1
- 210000001948 pro-b lymphocyte Anatomy 0.000 description 1
- 230000007425 progressive decline Effects 0.000 description 1
- 230000000069 prophylactic effect Effects 0.000 description 1
- 238000000159 protein binding assay Methods 0.000 description 1
- 210000001243 pseudopodia Anatomy 0.000 description 1
- 210000004879 pulmonary tissue Anatomy 0.000 description 1
- 239000000700 radioactive tracer Substances 0.000 description 1
- 238000011536 re-plating Methods 0.000 description 1
- 102000005962 receptors Human genes 0.000 description 1
- 108020003175 receptors Proteins 0.000 description 1
- 238000011084 recovery Methods 0.000 description 1
- 230000009467 reduction Effects 0.000 description 1
- 230000002829 reductive effect Effects 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 230000000284 resting effect Effects 0.000 description 1
- 210000003660 reticulum Anatomy 0.000 description 1
- PYWVYCXTNDRMGF-UHFFFAOYSA-N rhodamine B Chemical compound [Cl-].C=12C=CC(=[N+](CC)CC)C=C2OC2=CC(N(CC)CC)=CC=C2C=1C1=CC=CC=C1C(O)=O PYWVYCXTNDRMGF-UHFFFAOYSA-N 0.000 description 1
- 239000012679 serum free medium Substances 0.000 description 1
- 230000009450 sialylation Effects 0.000 description 1
- 210000004988 splenocyte Anatomy 0.000 description 1
- 239000007858 starting material Substances 0.000 description 1
- 229960005322 streptomycin Drugs 0.000 description 1
- 238000007920 subcutaneous administration Methods 0.000 description 1
- 230000009469 supplementation Effects 0.000 description 1
- 238000002560 therapeutic procedure Methods 0.000 description 1
- 210000001685 thyroid gland Anatomy 0.000 description 1
- 230000007838 tissue remodeling Effects 0.000 description 1
- 238000004627 transmission electron microscopy Methods 0.000 description 1
- 238000002054 transplantation Methods 0.000 description 1
- 210000004881 tumor cell Anatomy 0.000 description 1
- 210000002700 urine Anatomy 0.000 description 1
- 238000002255 vaccination Methods 0.000 description 1
- 210000005166 vasculature Anatomy 0.000 description 1
- 210000002845 virion Anatomy 0.000 description 1
- 239000011782 vitamin Substances 0.000 description 1
- 235000013343 vitamin Nutrition 0.000 description 1
- 229940088594 vitamin Drugs 0.000 description 1
- 229930003231 vitamin Natural products 0.000 description 1
- 230000029663 wound healing Effects 0.000 description 1
Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/0005—Vertebrate antigens
- A61K39/0008—Antigens related to auto-immune diseases; Preparations to induce self-tolerance
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/461—Cellular immunotherapy characterised by the cell type used
- A61K39/4615—Dendritic cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/462—Cellular immunotherapy characterized by the effect or the function of the cells
- A61K39/4621—Cellular immunotherapy characterized by the effect or the function of the cells immunosuppressive or immunotolerising
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/462—Cellular immunotherapy characterized by the effect or the function of the cells
- A61K39/4622—Antigen presenting cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/464—Cellular immunotherapy characterised by the antigen targeted or presented
- A61K39/4643—Vertebrate antigens
- A61K39/46433—Antigens related to auto-immune diseases; Preparations to induce self-tolerance
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/46—Cellular immunotherapy
- A61K39/464—Cellular immunotherapy characterised by the antigen targeted or presented
- A61K39/4648—Bacterial antigens
- A61K39/464817—Mycobacterium, e.g. Mycobacterium tuberculosis
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0634—Cells from the blood or the immune system
- C12N5/0639—Dendritic cells, e.g. Langherhans cells in the epidermis
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K35/00—Medicinal preparations containing materials or reaction products thereof with undetermined constitution
- A61K35/12—Materials from mammals; Compositions comprising non-specified tissues or cells; Compositions comprising non-embryonic stem cells; Genetically modified cells
- A61K2035/122—Materials from mammals; Compositions comprising non-specified tissues or cells; Compositions comprising non-embryonic stem cells; Genetically modified cells for inducing tolerance or supression of immune responses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K2239/00—Indexing codes associated with cellular immunotherapy of group A61K39/46
- A61K2239/31—Indexing codes associated with cellular immunotherapy of group A61K39/46 characterized by the route of administration
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K2239/00—Indexing codes associated with cellular immunotherapy of group A61K39/46
- A61K2239/38—Indexing codes associated with cellular immunotherapy of group A61K39/46 characterised by the dose, timing or administration schedule
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/20—Cytokines; Chemokines
- C12N2501/22—Colony stimulating factors (G-CSF, GM-CSF)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/20—Cytokines; Chemokines
- C12N2501/23—Interleukins [IL]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/20—Cytokines; Chemokines
- C12N2501/25—Tumour necrosing factors [TNF]
Definitions
- This invention relates to a method of culturing cells of the immune system.
- a method is provide f° r culturing proliferating dendritic cell precursors an for their maturation in vitro to mature dendritic cells.
- This invention also relates to dendritic cell modifie antigens which are T cell dependent, the method of makin them, and their use as immunogens.
- Vaccines, methods o immunizing animals and humans using the mature dendriti cells of the invention, and the modified antigens are als described.
- Dendritic cells process and present antigens most likely on abundant, newly synthesized MHC class II molecules, and then strong accessory and cell-cell adhesion functions are acquired (4-7) .
- Dendritic cells can migrate via the blood a nd lymph to lymphoid organs (8-10) . There, presumably as the "interdigitating 1 * cells of the T-area (8,11-13), antigens can be presented to T cells in the recirculating pool (14) .
- the progenitors of dendritic cells in the different compartments outlined above.
- dendritic cells in delivering antigens in such a way that a strong immune response ensues i.e., "immunogenicity", is widely acknowledged, but the use of these cells is hampered by the fact that there are very few 0 in any given organ.
- white cells In human blood, for example, about 0.1% of the white cells are dendritic cells (25) and these have not been induced to grow until this time.
- previous studies (20, 21) have not reported the development, in culture, of large numbers of dendritic cells from bone 5 marrow.
- dendritic cells can process foreign antigens into peptides that immunologically active T cells must recognize (4,6,7,14) i.e., dendritic cells accomplish the phenomenon of "antige presentation", the low numbers of dendritic cells prohibit c their use in identifying immunogenic peptides.
- Dendritic cells in spleen (15) and afferent lymph (16,17) are not in the cell cycle but arise from a proliferating precursor.
- dendritic cells emanate from the bone marrow (15,16,18,19), yet it has been difficult to generate these cells in culture except for two 5 reports describing their formation in small numbers (20,21) .
- Steinman, R. The Dendritic Cell System and Its Role In Immunogenicity", Ann. Rev. Immunol.. 0 9:271-96 (1991).
- polysaccharide generally elicit a T-cell independent immune response. There is no memory response and therefore no protection t subsequent infection with the polysaccharide antigen. 0 Proteins, however, do elicit a T-cell dependent response i infants.
- conjugate vaccines containin a polysaccharide covalently coupled to a protein convert the polysaccharide T-independent response to a T-dependen response. Unfortunately, little is known concerning th 5 sites on proteins which confer their T-cell dependen character, therefore hampering the design of more specifi immunogens.
- dendritic cells play a crucial role i the initiation of T-cell dependent responses.
- Dendriti o cells bind and modify antigens in a manner such that th modified antigen when presented on the surface of th dendritic cell can activate T-cells to participate in th eventual production of antibodies.
- the modification o antigens by dendritic cells may, for example, included 5 fragmenting a protein to produce peptides which have region which specifically are capable of activating T-cells.
- MHC Q The events whereby cells fragment antigens int peptides, and then present these peptides in associatio with products of the major histocompatibility complex, (MHC Q are termed "antigen presentation".
- the MHC is a region o highly polymorphic genes whose products are expressed on th surfaces of a variety of cells. MHC antigens are th principal determinants of graft rejection.
- Dendritic cells are specialized antigen presenting cells in the immune response of whole animals (14,31). Again however, the ability to use dendritic cells to identify and extract the " immunogenic peptides is hampered by the small numbers of these specialized antigen presenting 5 cells.
- Particle uptake is a specialized activity of mononuclear and polymorphonuclear phagocytes. Dead cells, immune complexes, and microorganisms all are avidly internalized. Following fusion with hydrolase-rich o lysosomes, the ingested particles are degraded (60,61). This degradation must be to the level of permeable amino acids (62,63) and saccharides, otherwise the vacuolar apparatus would swell with indigestible materials (64,65). Such clearance and digestive functions of phagocytes 5 contribute to wound healing, tissue remodeling, and host de ense.
- APCs antigen presenting cells
- processing requires the generation of peptides at least 8-18 amino acids in length (66,67), while scavenging entails digestion to amino acids (62,63).
- presentation requires the binding of peptides to MHC class II products (6,68), whereas scavenging does not require MHC 5 products.
- antigen presentation can function at a ° low capacity, since only a few hundred molecules of ligand need to be generated for successful stimulation of certain T-T hybrids (69,70) and primary T cell populations (71).
- This invention provides a method of producing a population of dendritic cell precursors from proliferating cell cultures.
- the method comprises (a) providing a tissue source comprising dendritic cell precursors; (b) treating the tissue source from (a) to increase the proportion of dendritic cell precursors to obtain a population of cells suitable for culture in vitro; (c) culturing the tissue source on a substrate in a culture medium comprising GM-CSF, or a biologically active derivative of GM-CSF, to obtain proliferating nonadherent cells and cell clusters; (d) subculturing the nonadherent cells and cell clusters to produce cell aggregates comprising proliferating dendritic cell precursors; and (e) serially subculturing the cell aggregates one or more times to enrich the proportion of dendritic cell precursors.
- This invention also provides a method of producing in vitro mature dendritic cells from proliferating cell cultures.
- the method comprises (a) providing a tissue source comprising dendritic cell precursor cells; (b) treating the tissue source from (a) to increase the proportion of dendritic cell precursors in order to obtain a population of cells suitable for culture in vitro; (c) culturing the tissue source on a substrate in a culture medium comprising GM-CSF, or a biologically active derivative of GM-CSF, to obtain non-adherent cells and cell clusters; (d) subculturing the nonadherent cells and cell clusters to produce cell aggregates comprising proliferating dendritic cell precursors; (e) serially subculturing the cell aggregates one or more times to enrich the proportion of dendritic cell precursors; and (f) continuing to culture the dendritic cell precursors for a period of time sufficient to allow them to mature into mature dendritic cells.
- This invention also provides a method of increasing the proportion of dendritic cells present in the tissue source by pretreating the individual with a substance to stimulate he atopoiesis.
- the pretreatment step comprises killing cells expressin antigens which are not expressed on dendritic precursor cells by contacting the bone marrow with antibodies specific for antigens not present on dendritic precursor cells in a medium comprising complement. Removal of undesirable non- dendritic cell precursors may also be accomplished b adsorbing the undesirable non-dendritic or their precurso cells onto a solid support.
- This invention also provides a method of preparing antigens from dendritic cell precursors comprising providing precursor dendritic cells from a population of precursor cells capable of proliferating, contacting the precursor cells with antigen for a period of time sufficient to allow the dendritic cell precursors to phagocytose the antigen and obtain antigen-containing dendritic cell precursors; culturing the antigen containing-dendritic cell precursors under conditions and for a period of time sufficient for the antigen to be processed and presented by dendritic cell precursors.
- the antigens processed by the dendritic cell precursors as a result of phagocytosis may themselves be used alone or in combination with adjuvants including dendritic cell precursors to evoke an immune response in an individual to the antigen.
- the invention provides self- peptide antigens produced by pulsing the dendritic cells of
- This invention also provides a method of treating autoimmune diseases by treating an individual with 0 therapeutically effective amounts of self-peptides produced according to the method of the invention to induce tolerance to the self-proteins.
- the treatment of autoimmune diseases comprising administering to an individual in need of treatment a 5 therapeutically effective amount of antigen-activate dendritic cells where the antigen is a self-protein o autoantigen is also provided.
- a further objective of the invention is to provide a method of immunizing individuals with T-cell dependen antigens for the prevention and treatment of disease.
- Fig. 3 FACS analyses of dendritic cell precursors tha could be dislodged by Pasteur pipetting of proliferatin aggregates, and dendritic cells released spontaneously i culture.
- the mAb are: Ml/42 anti-MHC class I [ATCC # TI 126]; NLDC145 anti-interdigitating cell (13); M5/114 anti MHC class II [ATCC # TIB 120]; 33D1 anti-dendritic cel [ATCC # TIB 227]; B5-5 anti-thy-1.
- the staining with anti MHC mAbs is bimodal, but the released cell fraction o dendritic cells is richest in expression of MHC class I an II.
- Fig. 6 Physical properties of the MLR stimulating cells that develop in GM-CSF supplemented bone marrow cultures [see text] .
- Fig. 8 Detailed cell cytofluorometric phenotype analysi 0 of the la-positive cells released from the growing dendriti cell aggregates. Contaminating, la-negative granulocyte were gated out on the basis of lower forward light scatter so that one could examine the expression of many surfac antigens on the larger cells using rat and hamster anti 5 mouse mAbs (7,17) as indicated.
- Fig. 9 Quantitation of developing cells that bear th dendritic cell restricted granule antigens 2A1 and M342.
- Fig. 10 Progenitor-progeny relationships in growin dendritic cells. Growing aggregates were separated at d 5 from bone marrow cultures and pulsed with 3 H-TdR at 0. ⁇ Ci/ l, 3 x 10 5 cells/well, for 12h. All wells were replaced with fresh medium and returned to culture for 1, 2, or 3 days of chase. The yields of released cells during the chase were 2.0, 2.9, and 3.0 x 10 s respectively per well. The content of Ia + cells was 28% after the pulse, and 47%, 55%, and 62% on days 1, 2, and 3 respectively. The data are shown as percentage of cells that were radiolabeled, with the filled in bars being cells that express the 2A1 granule cell antigen of mature dendritic cells.
- A An aggregate of developing dendritic cells cytospun after a 2Oh exposure to 2u latex spheres. Many cells in the aggregate are labeled with the uniform latex particles [arrows] .
- the sorted cells were cytospun, stained with auramine-rhodamine to visualize the cell- associated BCG, and double labeled with a different mAb an immunoperoxidase.
- the left and right panels of each pair are phase contrast and acid fast views respectively. Arrows on the left indicate the location of the bacilli on th right.
- the label for class II, [I-A and I-E, M5/114] outlines the cell processes better than the dendritic cell restricted NLDC-145 antibody.
- Fig 14 Electron microscopy of BCG in dendritic cells. As in Fig 2, BCG was added to GM-CSF stimulated d6 bon marrow cultures for a day. After washing and 2 more days o culture, the released cells were processed for electro microscopy. A,B. Low power views to show the typical dendriti cells with numerous processes and a few phagocytosed BC [arrows] .
- T cells were purified from lymph nodes that drain paws that had been primed with complete [CFA] or incomplete [IFA]
- Mature dendritic cells are d8 bone marrow cultures, and immature dendritic cells are from d5-6 cultures.
- Fig 16 Antigen presentation to naive lymph node T cells in situ.
- Growing cultures of bone marrow dendritic cells were pulsed with BCG at d5-6, and used immediately or after a 2d chase culture to activate T cells.
- the populations were injected into the paws of naive mice without artificial adjuvants.
- Five days later the draining lymph nodes were taken and stimulated in vitro with graded doses of PPD or BSA (the dendritic cells had been grown with fetal calf serum) , the BSA to serve as a nonparticulate antigen.
- Data are means and standard errors for groups of 5 mice, each studied separately. Control lymph nodes not exposed to BCG pulsed dendritic cells did not respond to PPD or to BSA ( ⁇ 2000 cpm) .
- Fig 17 Antigen presentation to naive spleen cells in situ.
- Growing cultures of bone marrow dendritic cells were pulsed with BCG at d5-6 (immature) , at d7-8 (mature) , or at d5-6 followed by a 2d chase.
- 10 6 cells of each group were injected i.v. into groups of mice. 5 or 10 days later, the spleen cells were cultured in vitro with graded doses of PPD or BSA as antigen. Since the dendritic cells were cultured in FCS, the use of BSA serves as control to ensure that all dendritic cell populations were comparably immunogenic in vivo. Unprimed spleen did not respond to either BSA or PPD.
- Figs 18A, B and C Mixed Leukocyte Reaction (MLR) assay of human dendritic cells produced according to the metho described in Example 6. Graded doses of irradiated cell (30 to 30,000 in serial 3 fold dilutions) were added to 2 10 5 accessory cell-depleted T cells.
- MLR Mixed Leukocyte Reaction
- tissue source comprising dendritic cell precursors whic precursor cells are capable of proliferating and maturing i vitro into dendritic cells when treated according to the method of the invention.
- Such precursor cells are nonadherent and typically do not label with mAb markers found on mature dendritic cells such as Ia antigens, 2A1 and M342 antigens (34, 44) and the NLDC145 interdigitating cell source antigen (13) .
- tissue sources are spleen, afferent lymph, bone marrow and blood. More preferred tissue sources are bone marrow and blood. Blood is also a preferred tissue source of precursor cells because it is easily accessible and could be obtained in relatively large quantities.
- hematopoietic factor to be administered may be determined by one skilled in the art by monitoring the cell differential of individuals to whom the factor is being administered.
- dosages of factors such as G-CSF and GM-CSF will b e similar to the dosage used to treat individuals recovering from treatment with cytotoxic agents.
- GM-CSF or G-CSF is administered for 4 to 7 days at standard doses prior to removal of source tissue to increase the proportion of dendritic cell precursors.
- the tissue source may be treated prior to culturing to enrich the proportion of dendritic precursor cells relative to other cell types. Such pretreatment may also remove cells whic may compete with the proliferation of dendritic precurso cells or inhibit their proliferation or survival Pretreatment may also be used to make the tissue source mor suitable for in vitro culture.
- the method of treatment wil likely be tissue specific depending on the particular tissu source. For example, spleen or bone marrow if used as tissue source would first be treated so as to obtain singl cells followed by suitable cell separation techniques t separate leukocytes from other cell types. Treatment o blood would involve cell separation techniques to separat leukocytes from other cells types including red blood cell (RBCs) which are toxic.
- RBCs red blood cell
- B cells ar removed prior to culturing of bone marrow in GM-CSF.
- Whil B cells and pre-B cells do not grow in response to GM-CSF they represent approximately 50% of the initial marro suspension and thereby preclude the use of staining wit anti-la monoclonal antibodies to quickly enumerate dendriti cells.
- granulocytes are GM-CSF responsive an readily proliferate in the presence of GM-CSF. As such, th B cells and granulocytes mask the presence of dendritic cel precursors.
- pretreatment cells which compete and mask the proliferation of precursor dendritic cells are killed.
- Such pretreatment comprises killing cells expressing antigens which are not expressed on dendritic precursor cells by contacting bone marrow with antibodies specific for antigens not present on dendritic precursor cells in a medium comprising complement.
- Another form of pretreatment to remove undesirable cells suitable for use with this invention is adsorbing the undesirable precursor cells or their precursors onto a solid support using antibodies specific for antigens expressed on the undesirable cells.
- Several methods of adsorbing cells to solid supports of various types are known to those skilled in the art and are suitable for use with this invention. For example, undesirable cells may be removed by "panning" using a plastic surface such as a petri dish.
- Any isotonic solution commonly used in tissue cultur may be used as the medium for separating blood leukocyte from platelets and red blood cells.
- suc isotonic solutions are phosphate buffered saline.
- RPMI 1640 is preferred.
- Cells obtained from treatment of the tissue source are cultured to form a primary culture on an appropriate substrate in a culture medium supplemented with GM-CSF or a 5 GM-CSF derivative protein or peptide having an amino acid sequence which sequence maintains biologic activity typical of GM-CSF.
- the appropriate substrate may be any tissue culture compatible surface to which cells may adhere.
- the substrate is commercial plastic treated for use in tissue culture. Examples include various flasks, roller bottles, petri dishes and multi-well containing plates made for use in tissue culture. Surfaces treated with a substance, for example collagen or poly-L-lysine, or antibodies specific for a particular cell type to promote 5 cell adhesion may also be used provided they allow for the differential attachment of cells as described below.
- Cells are preferably plated at an initial cell density of about 7.5 X 10 5 cells per cm 2 . At this dose, the surface is not fully covered by cells, but there are no big spaces (2-3 0 cell diameters) either.
- the la-negative marrow 5 nonlymphocytes comprising dendritic cell precursors are preferably cultured in high numbers, about 10 6 /well (5 x 10 5 cells/cm 2 ) Liquid marrow cultures which are set up for purposes other than culturing dendritic cell precursors are typically seeded at 1/lOth this dose, but it is then 0 difficult to identify and isolate the aggregates of developing dendritic cells.
- cytokines are produced from the human gene usin reco binant techniques (rhu) .
- TNF ⁇ at concentrations fro about 10-50 U/ml may be used to increase dendritic cell yields several fold.
- the primary cultures from the tissue source are allowe to incubate at about 37°C under standard tissue cultur conditions of humidity and pH until a population of cell has adhered to the substrate sufficiently to allow for th separation of nonadherent cells.
- the dendritic cel precursor in blood initially is nonadherent to plastic, i contrast to monocytes, so that the precursors can b separated after overnight culture.
- Monocytes an fibroblasts are believed to comprise the majority o adherent cells and usually adhere to the substrate withi about 6 to about 24 hours.
- the nonadherent cells from the primary culture are subcultured by transferring them to new culture flasks at a density sufficient to allow for survival of the cells and which results in the development over time of clusters of growing cells that are loosely attached to the culture surface or to the firmly adherent cells on the surface. These clusters are the nidus of proliferating dendritic cell precursors.
- culture flasks refers to any vessel suitable for culturing cells. It is desirable to subculture all of the nonadherent cells from the primary culture at a density of between about 2 X 10 5 cells and 5 X - 5 cells per cm 2 . Preferably at about 2.5 X 10 5 per cm 2 .
- the dendritic cells besides being identified by thei stellate shape may also be identified by detecting thei expression of specific antigens using monoclonal antibodies.
- a panel of monoclonal antibodies may be used t identify and characterize the cells in the GM-CSF expande cultures. The monoclonal antibodies are reviewed elsewher (23, 24 which are incorporated herein by reference).
- the pulse and chase protocol which may be used to charge developing dendritic cells with organisms according to our invention allows the two broad components of immunostimulation to take place sequentially. These components are a) antigen capture and presentation, here the capture of particulates by immature dendritic cells, and b) development of potent accessory or immunostimulatory functions during the chase period. The situation is comparable to that seen in the handling of soluble proteins (4,6) and particles (74) by epidermal Langerhans cells. Each of the two broad components of APC function entails many subcomponents.
- the novel antigens of the invention are prepared b combining substances to be modified or other antigens wit the dendritic cells prepared according to the method of th invention.
- the dendritic cells process or modify antigen in a manner which promotes the stimulation of T-cells by th processed or modified antigens.
- Such dendritic cel modified antigens are advantageous because they can be mor specific and have fewer undesirable epitopes than non- modified T-dependent antigens.
- the dendritic cell modified antigens may be purified by standard biochemical methods.
- MHC major histocompatibility complex
- dendritic cells are pulsed with the endogenous antigen t produce the relevant "self-peptide".
- the relevant self peptide is different for each individual because MH products are highly polymorphic and each individual MH molecules might bind different peptide fragments. Th "self-peptide” may then be used to design competing peptide or to induce tolerance to the self protein in the individua in need of treatment.
- B. Blood Preparation Blood was obtained by cardiac puncture or from the carotid artery. The blood was diluted in, or allowed to drip into, RPMI-1640 with 100 u/ml heparin [about 2 ml/mouse]. Blood cells were pelleted at 1000 rpm at 4°, resuspended in RPMI 1640, and sedi ented again.
- the adherent cells did not develop dendritic cell colonies, but during the next week, the nonadherent populations exhibited three changes. First, most of the lymphocytes and granulocytes died or could be removed by washing. Second, the surface of the well became covered with a monolayer of tightly adherent cells that included macrophages and fibroblasts. Third, affixed to scattered sites on the monolayer, there developed small aggregates of cells. The cultures were fed with GM-CSF (30 u/ml) at day 6-7 and then every 3 days by aspirating 0.5- 0.75 ml of the medium and adding back an equal volume of fresh medium with GM-CSF. The aggregates continued t expand in number and size. At about day 10, the cells were ready to be subcultured. Any residual loose cells could b rinsed off prior to dislodging the aggregates into fres medium and GM-CSF. About 0.8-1 million dislodged cells pe original well were divided into 3 subculture wells.
- the cytospin cells were stained for specific antigens with mAb and immunoperoxidase as above. Also, the slides were dipped in photographic emulsion [Kodak autoradiography emulsion type NTB2 #165-4433] for exposure [5 days] prior to development, staining with Giemsa, and mounting in Permount. For pulse chase experiments, a lower dose of 3H-TdR was used to maintain cell viability, but the cells were handled similarly otherwise. The pulse was applied at 0.1 uCi/ml for 2h or for 16h, the latter to provide higher initial labeling indices. The cells were washed and chased for 1-3 days prior to harvesting and analysis as above with immunoperoxidase, autoradiography, and Giemsa staining.
- Pulse chase experiments were then done to establish that labeled cells in the aggregate were giving rise to typical dendritic cells.
- Cultures were first exposed to a low dose of 3H-TdR, either for 2h or for 16h, the latter to label a larger percentage of the cells in the aggregates.
- the wells were washed free of radiolabel, and then the aggregates were dislodged and separated from free cells by lg sedimentation.
- the aggregates were transferred to fresh medium without radiolabel, and over the next 1-3 days of culture, many dendritic cells were released into the medium. When the "chased" cultures were examined, several findings were apparent.
- a second specialized feature of dendritic cells i their capacity to home to the T areas of peripheral lymphoi tissues (8,10).
- Dendritic cells or other cell types were labeled at 2-10 x 10 6 /ml with carboxyfluorescein for 10 mi on ice [Molecular Probes C-1157; 30 uM final concentration in Hanks balanced salt solution (HBSS) with 5% FCS], washed in RPMI 1640, and injected in a volume of 50 ul RPMI-1640 into the foot pads.
- HBSS Hanks balanced salt solution
- FCS 5% FCS
- the sections were applied to multiwell slides [Carlson Scientific microslides #111006], stored at -20°C, dried in a desiccator 30' prior to use [or left at room temp overnight], fixed in acetone, and stained with a peroxidase conjugated rabbit anti-FITC antibody [Dakopatts, P404].
- dendritic cells that had been generated with GM-CSF from blood, homing to the T area was observed with injections of 200,000 cells. The selective localization to the T areas was confirmed by double labeling the specimens with mAb that stain B cells or T cells. Therefore dendritic cells produced in culture have the key functional features of this lineage: homing to the T-dependent regions and strong accessory activity.
- the surface phenotype of the blood cell that gives rise to the dendritic cell colonies was assessed by treating the starting population with antibodies and complement. Treatment with either 33D1 anti-dendritic cell, anti-MH class II, or anti thy-1 did not eliminate the colony formin unit [not shown] . Instead, removal of thy-l + or Ia + cell enriched colony numbers several fold. CSF's other than GM CSF were also tested, either at the start of the 1-3 wee culture, or upon transfer of 2-3 week old aggregates to for veiled cells. None of the CSF's tested, i.e., IL-3, M-CSF G-CSF, SCF, supported the formation of colonies or matur dendritic cells.
- dendritic colonie are very much dependent upon GM-CSF.
- Dendritic cel precursors were not observed from neonatal epidermis, whi contains mainly la" Langerhans cells (29) .
- the precursors to the aggregates of proliferating dendritic cells were not typical monocytes or dendritic cells, because the number of aggregates that developed could 5 be increased substantially if one depleted monocytes by adherence or la-positive cells with antibody and complement. Without wishing to be bound by theory, we tentatively conclude that blood contains an la-negative precursor that forms a proliferating aggregate. In the aggregate, o dendritic cells mature and are released as nonproliferating progeny.
- the labeled progeny In pulse chase protocols, large numbers of labeled progeny were released from the aggregates, and these released cells were nonadherent, motile, and strongly stimulatory in the MLR. After combined autoradiography and immunoperoxidase labeling, the labeled progeny carried the granular antigens, the NLDC-145 antigen, and very high levels of MHC class II. Each of these cytologic and antigenic markers are largely restricted to dendritic cells.
- the functional maturation that occurred in the ° proliferating aggregate is striking.
- the dendritic cells that were generated in culture were potent MLR stimulators. 100 dendritic cells induced a much stronger primary MLR than 100,000 blood leukocytes. The increase in stimulating activity per la-positive cell was at least 2 logs between the time that the aggregates first appeared and the time that typical dendritic cells were released in large numbers. Over this time period, cell recovery increased 5-10 fold. Also the dendritic cell progeny homed in a precise way to the T cell area of lymph node, another functional property that was not detectable in blood cells [data not shown].
- mice Female BALB/C, male DBA/ 2 , and female C57BL/6 mice, 7 wks old, were purchased from Japan SLC [Hamamatsu, Shizuoka, Japan].
- the culture medium was RPMI-1640 [Nissui, Tokyo, Japan; GIBCO, Grand Island, NY] supplemented with 5% FCS, 50 ⁇ M 2-Mercaptoethanol, and 20 ⁇ g/ml genta icin.
- Murine rGM- CSF 10 8 U/mg protein] was kindly provided by Kirin Brewery 5 Co [Maebashi, Gumma, Japan].
- Kirin Brewery 5 Co [Maebashi, Gumma, Japan].
- a panel of rat and hamster mAbs to mouse leukocyte antigens is described elsewhere (23, 24) .
- FITC- and peroxidase-conjugated mouse anti-rat IgG were purchased from Boehringer Mannheim [Indianapolis, IN] and FITC- and peroxidase-conjugated goat anti-hamster lg [ ⁇ 0 and L-chain] were from Jackson Immunoresearch Lab [Westgrove, PA] and Caltag [San Francisco, CA] respectively.
- Bone marrow cultures After removing all muscle tissues with gauze from the mouse femurs and tibias, the 5 bones were placed in a 60 mm dish with 70% alcohol for 1 min, washed twice with PBS, and transferred into a fresh dish with RPMI-1640. Both ends of the bones were cut with scissors in the dish, and then the marrow was flushed out using 2 ml of RPMI-1640 with a syringe and 25G needle. The tissue was suspended, passed through nylon mesh to remove small pieces of bone and debris, and red cells were lysed with ammonium chloride. After washing, lymphocytes and la- positive cells were killed with a cocktail of mAbs and rabbit complement for 60 min at 37°C.
- the mAbs were GK 1.5 anti-CD4, HO 2.2 anti-CD8, B21-2 anti-la, and RA3-3A1/6.1 anti-B220/CD45R all obtained from the ATCC [TIB 207, 150, 229, and 146 respectively].
- 7.5-10 x 10 5 cells were placed in 24 well plates [Nunc, Naperville, IL] in 1 ml of medium supplemented with 500-1000 U/ml rGM-CSF. The cultures were usually fed every 2d for about 2 to 10 days, by gently swirling the plates, aspirating 3/4 of the medium, and adding back fresh medium with GM-CSF.
- Th aggregates were subcultured at 2-3 x 10 5 /ml in fresh maxim with GM-CSF, typically for 1 day in 16 mm wells. Afte overnight culture, large numbers of typical dendritic cell were released. Adherent macrophages also expanded in thes cultures, but most remained firmly adherent to the cultur surface. ° C. Cytological Comparison of Dendritic Cell Precursors and Ia-ne ative. Bone Marrow Nonlymphocytes
- dendritic profiles are in the released fraction [a hand lens is useful to detect cell 5 shape and contaminating granulocytes, in the Giemsa stain], while the adherent cells are for the most part typical vacuolated macrophages. Strong MHC class II expression occurs on all released cells but for a few typical granulocytes. Only a subset of the firmly adherent cells o express class II. Most, released cells express the 2A1 endocytic vacuole antigen, while the adherent cells are 2A1 weak or negative.
- Cell surface and intracellular antigens Cell surface 5 staining utilized cytofluorography [FACScan; Becton
- EDTA-PBS EDTA-PBS
- EDTA-PBS EDTA-PBS
- cytospins were fixed in acetone and stained with mAbs followed by peroxidase conjugated anti-rat or anti- hamster lg.
- the peroxidase was visualized with diaminobenzidine, and the nuclei counterstained with Giemsa.
- F. Mixed leukocyte reactions Cells from the bone marrow cultures were exposed to 15 Gy of X-ray irradiation and applied in graded doses to 3 x 10 5 syngeneic or allogeneic T cells in 96 well flat bottomed culture plates for 4d. The cells were prepared by passing spleen and lymph node suspensions through nylon wool and then depleting residual APCs with anti-la plus Jlld mAbs plus complement. 3H- thymidine uptake was measured at 80-94h after a pulse of 4 uCi/ml [222 GBq/mmol; American Radiolabeled Chemicals, Ine, St.Louis, MO].
- G Aggregates of proliferating dendritic cells from mouse bone marrow supplemented with GM-CSF.
- Attached to the surface of the culture wells were cells with the cytologic features of macrophages, and these also expanded in numbers during the first week of culture. These cells could be dislodged by pipetting after incubation at 37°C in the presence of 10 mM EDTA.
- the nonadherent cells which were mainly granulocytes in the first 4 days, were obtained by gently swirling the plates and harvesting the cells.
- the loosely adherent cells which contained the aggregates of presumptive dendritic cell precursors and dendritic cells at day 4 and later times, were dislodged by pipetting over the surface of firmly adherent stromal cells.
- the most potent stimulating activity was in the adherent fraction.
- the nonadherent fraction was very active. If one tested firmly adherent macrophages, there was no MLR stimulating activity [Fig 6B, open squares].
- Cytospins were prepared to further compare the released dendritic cells with the firmly adherent stromal population.
- Giemsa stain the cells that had released from the aggregates had the typical stellate shape of dendritic 5 cells, while the adherent cells were for the most part vacuolated macrophages.
- Many of the dendritic cells had a perinuclear spot of nonspecific esterase stain, while the more adherent populations had abundant cytoplasmic esterase.
- the released cells stained strongly for MHC class II 0 products, except for the contaminants with typical granulocyte nuclei.
- the strongly adherent cells contained a subpopulation of class II positive cells.
- the antibodies are termed M342 (34) and 2A1. Many of the dendritic cells had strong 2A1 stain, and a smaller number expressed M342. The adherent cells had a few profiles with weak 2A1.
- the released cells had many large veils or lamellipodia extending from several directions of the cell body.
- the cytoplasm had many mitochondria, few electron dense granules and lysosomes, but several electron lucent vesicles some with the cytologic features of multivesicular bodies.
- the numerous cell processes extending from the dendritic cells were evident in the semi-thin sections of our preparations.
- a bone marrow-derived dendritic cell at d5 of culture shows many cytoplasmic veils.
- a close up of the perinuclear region shows profiles of smooth reticulum and vacuoles. There are few lysosomal or phagocytic structures.
- mice BALB/C x DBA/2 Fl, C57BL/6 x DBA/2 Fl, and BALB/C male and female mice were purchased from the Trudeau Institute [Saranac Lake, NY] and Japan SLC [Hamamatsu] and used at 6-10 weeks of age.
- Bone marrow cultures As described in Example 2 above, bone marrow was flushed from the femus and tibias, depleted of red cells with 0.83% ammonium chloride, and cultured in 24 well plates [Nunc, Napaville, IL and Corning #25820, Corning NY] at 10° cells/well in 1 ml of RPMI-1640 supplemented with 5% fetal calf serum, 20 ug/ml gentamicin, and 1000 U/ml of recombinant murine GM-CSF [Kiren Brewery, Maebashi, Gumma, Japan; 9.7 x 10 7 U/mg] .
- cells in the washed cultures were dislodged and 3-4 x 10 6 cells transferred to a 60 mm Petri dish for a 1 or 2 day "chase" period in particle-free, fresh, GM-CSF supplemented medium.
- Class Il-rich, mature dendritic cells developed during the chase as described in Example 2, and these were isolated by cell sorting [below].
- particles were also administered to 7-8d bone marrow cultures that are rich in single nonproliferating mature dendritic cells.
- BCG mycobacteria [Trudeau Institute, 1.5-2.5 x 10 8 CFU/ml; Kyowa Pharmaceutical Industries, Tokyo] were administered at approximately 10 7 live BCG per 16mm diameter well. Uptake was assessed following an "acid fast" stain using an auramine-rhodamine procedure that is more sensitive than Ziehl " Neelsen and facilitates organism counts. Colloidal carbon [Pellikan Ink, Hannover, Germany] was added at 1:2000 dilution. The carbon was identified as a black granular stain in specimens stained with Diff-Quik R [Baxter Healthcare Corp, Miami, FL] . Suspensions of 2u latex particles [0.5% v/v; Seradyn, Indianapolis, IN] were applied to the cultures at 50 ul/well, a dose which covers the surface of the culture well with beads.
- D. Isolation of mature dendritic cells by cell sorting As noted before in Example 2, the dendritic cells that are produced in GM-CSF stimulated bone marrow cultures express very high levels of surface MHC class II products [monoclonals B21-2, TIB 227 and M5/114, TIB 120 from the ATCC] as well as moderate levels of a dendritic cell- restricted antigen recognized by monoclonal NLDC-145. Immediately after the pulse with BCG, or after an additional 2 days of "chase" culture, the cells were stained with biotin B21-2 and FITC-streptavidin [Tago, Burlingame, CA].
- Class Il-rich cells then were sorted [FACStar Plus, Becton Dickinson, Mountainview, CA] and cytospun onto glass slides [Shandon Inst. Sewicky, PA] .
- the sorted cells were stained with Diff Quick® which outlines the stellate shape of dendritic cells in cytospins and allows enumeration of profiles containing perinuclear depots of internalized colloidal carbon or latex spheres.
- the cytospins were fixed in absolute acetone for 10 min at room temperature and stained with M5/114 anti-class II, NLDC-145 anti-dendritic cell, or RA3-6B2 anti-B220 or anti-B cell [the latter as a control] followed by POX conjugated mouse anti-rat lg [Boehringer Mannheim, Indianapolis,IN] and diaminobenzidine tetraHCl [Polyscience Ine, Warrington, PA].
- the preparations were then double labeled for acid-fast bacilli with auramine rhodamine. Virtually all the cells in the preparation were rich in NLDC-145 and MHC class II products.
- the number of BCG bacilli in at least 400 cells were enumerated.
- mice were primed with complete Freunds' adjuvant [CFA, SIGMA, St.Louis, MO; 25 ul in the fore and rear paws] or as a control, mycobacteria- free incomplete Freunds' [ICFA]. 7-14d later, the draining lymph nodes were dissociated into a single cell suspension and depleted of APCs with mAbs to MHC class II, B220, and heat stable antigens [M5/114 anti-la, RA3-6B2 anti-B220, and Jlld anti-HSA; TIB 120, 146, and 183 from the ATCC respectively] and rabbit complement.
- CFA complete Freunds' adjuvant
- SIGMA St.Louis, MO
- ICFA mycobacteria- free incomplete Freunds'
- 3xl0 5 of these APC- depleted, primed T cells were cultured in 96 well flat- bottomed microtest wells [Corning #25860] in RPMI-1640 medium supplemented with 0.5% mouse serum and 50 uM 2- mercaptoethanol.
- Graded doses of BCG-pulsed, bone marrow or spleen APCs were added.
- 1 uCi of 3H-thymidine [NEN, Boston, MA; 20 Ci/mmol; 4 uCi/ml] uptake was added to monitor DNA synthesis at 72-88h. Data shown are means of triplicates in which standard errors were ⁇ 15% of the mean.
- APCs that had been pulsed with antigen in vitro were administered in vivo to unprimed CxD2 Fl mice.
- To prime T cells in draining lymph node 2x10 s dendritic cells were injected into the paws, and lymph node cells were prepared 5d later.
- To prime T cells in spleen 10 6 cells were injected i.v. , and splenocytes were prepared 5 or lOd later.
- bulk lymph node or spleen cells were cultured as above and challenged with graded doses of protein antigens, either purified protein derivative [PPD, from Statenserum Institute, Copenhagen, Denmark, or from Dr.
- proliferating cell aggregates When mouse bone marrow or blood is stimulated with GM- CSF, proliferating cell aggregates appear, and these give rise to large numbers of typical immunostimulatory dendritic cells.
- the proliferating aggregates are best identified by washing away the majority of nonadherent granulocytes that are also induced by GM-CSF in the cultures.
- the time point when the aggregates were first sizable [5-10 cells wide] we applied different particles over a 20-22h period.
- Fig 12A Aggregates that had been exposed to particles were recultured an additional 2 days. During this time, large numbers of cells were released into suspension. These primarily were mature dendritic cells with characteristic stellate shapes and high levels of MHC class II and NLDC-145 antigens. When the released cells were examined by light microscopy, many contained latex spheres and often around a clear perinuclear zone or centrosphere [Fig 12B] . We also studied colloidal carbon uptake in a similar manner.
- Live BCG mycobacteria were administered as the phagocytic meal over a 20-22h period using the protocol for administering latex particles described above.
- Cell- associated bacilli were visualized by a sensitive fluorescent acid-fast stain.
- the developing dendritic cell aggregates contained many organisms.
- the cells were resuspended and sorted those cells with high levels of MHC class II products.
- the BCG pulse about 20% of the sorted cells contained acid fast bacilli [Table 1] .
- the majority of MHC class II-weak cells were not studied further because of excessive stickiness during cell sorting.
- Quantitative data of dendritic cells containing BCG Mouse bone marrow cultures were stimulated in 16mm wells for 5d with GM-CSF, washed, and exposed to BCG organisms for 20h. The cultures were washed again and either examined immediately, or pooled and transferred to a 60 mm dish for an additional 2d chase culture. The dendritic cells in the cultures were selected as la-rich cells using a fluorescent activated cell sorter and then cytospun onto glass slides for staining for acid fast bacilli. During the chase period, the percentage of la-rich cells in the cultures increased 2-2.5 fold, and the total number of cells increased 2 fold, resulting in a 4-5 fold increase in the number of la-rich cells.
- the percentage of dendritic cells containing BCG also rose to 50% [Table 1, Fig. 13].
- Double labeling experiments verified that cells with acid fast bacilli expressed MHC class II and the dendritic cell-restricted NLDC 145 antigen Fig. 13. Because the total number of MHC class II and NLDC- 145 positive cells had increased 4-fold in just 2d, it is likely that these BCG-laden dendritic cells were derived from less mature but phagocytic progenitors in the aggregates.
- BSA bovine serum albumin
- the surface markers of the primed cells were tested by antibody and complement mediated lysis of the populations prior to measuring 3H-thymidine uptake [data not shown] .
- the proliferating cells were positive for thy-1, but negative for MHC class II, heat stable antigen, and B220.
- Anti-CD4 hybridoma culture supernatant blocked proliferation more than 85% i.e., the primed cells were helper-type T cells.
- Dendritic cells prepared according to the method described in Example 1 are plated at a concentration of approximately 1 x 10 s cells per well of a 24 well plastic culture plate. The cells are incubated in RPMI 1640 containing 5% fetal calf serum and GM-CSF (30 u/ml) . Antigen is added to the dendritic cell cultures and the cultures are incubated with antigen for approximately 4 hours or for sufficient time to allow the dendritic cells to handle the antigen in an immunologically relevant form, or in a form that can be recognized by T cells.
- Such handling of the antigen by the dendritic cells involves the dendritic cells 1) acquiring, 2) processing, and 3) presenting the antigen to the T cells in a form which is recognized by the T cells. Following binding of the antigen to the dendritic cells the cells are collected from the culture and used to immunize syngeneic mice. The activated dendritic cells are injected subcutaneously into the mice in an amount sufficient to induce an immune response to the antigen.
- Dendritic cells prepared as described in Example 1 are pulsed with a protein antigen for a time sufficient to allow the dendritic cells to acquire, process and present the modified antigen on the surface of the dendritic cells. The dendritic cells are then collected from the culture for extraction of the modified antigen.
- the dendritic cells are solubilized with detergent to extract the modified antigen bound to MHC molecules.
- the MHC molecules bound to modified antigen are purified by precipitation with antibodies which bind the MHC molecules such as MH2.
- the modified antigens are extracted from the precipitate for analysis.
- the isolated mononuclear cells were depleted of cells that were not dendritic cell progenitors. These contaminants were coated with monoclonal antibodies to CD3 and HLA-DR antigens and depleted on petri dishes coated with affinity-purified, goat anti-mouse IgG ("panning") .
- Cells were cultured under the following conditions: 1) without presence of additional cytokines; 2) GM-CSF, 400 or 800 U/ml; 3) GM-CSF, 400 or 800 U/ml, plus IL-l ⁇ , 50 LAF units/ml for the last 24 h of culture; 4) GM-CSF, 400 or 800 U/ml, plus TNF ⁇ , 50 U/ml; 5) GM-CSF, 400 or 800 U/ml, plus TNF- ⁇ , 50 U/ml, plus IL-l ⁇ , 50 LAF units/ml for the last 24 h of culture; 6) GM-CSF, 400 or 800 U/ml, plus IL-3, 100 U/ml; 7) GM-CSF, 400 or 800 U/ml, plus IL-3, 100 U/ml, plus IL-l ⁇ , 50 LAF units/ml for the last 24 h.
- GM-CSF is an essential cytokine. G-CSF, M-CSF, IL-3, or no cytokine do not permit the development of dendritic cell balls. GM-CSF at 400-800 U/ml is optimal, irregardless of whether donors had been treated with either GM-CSF or G-CSF to expand the number of myeloid progenitor cells in blood. Addition of TNF ⁇ at 10-50 U/ml usually but not always increased dendritic cell yields up to two-fold (cf. Caux et al..
- Dendritic cells are the principal cells in mouse spleen bearing immunogenic fragments of foreign proteins. J. Exp. Med. 172:383.
- HRP horseradish peroxidase
Abstract
Description
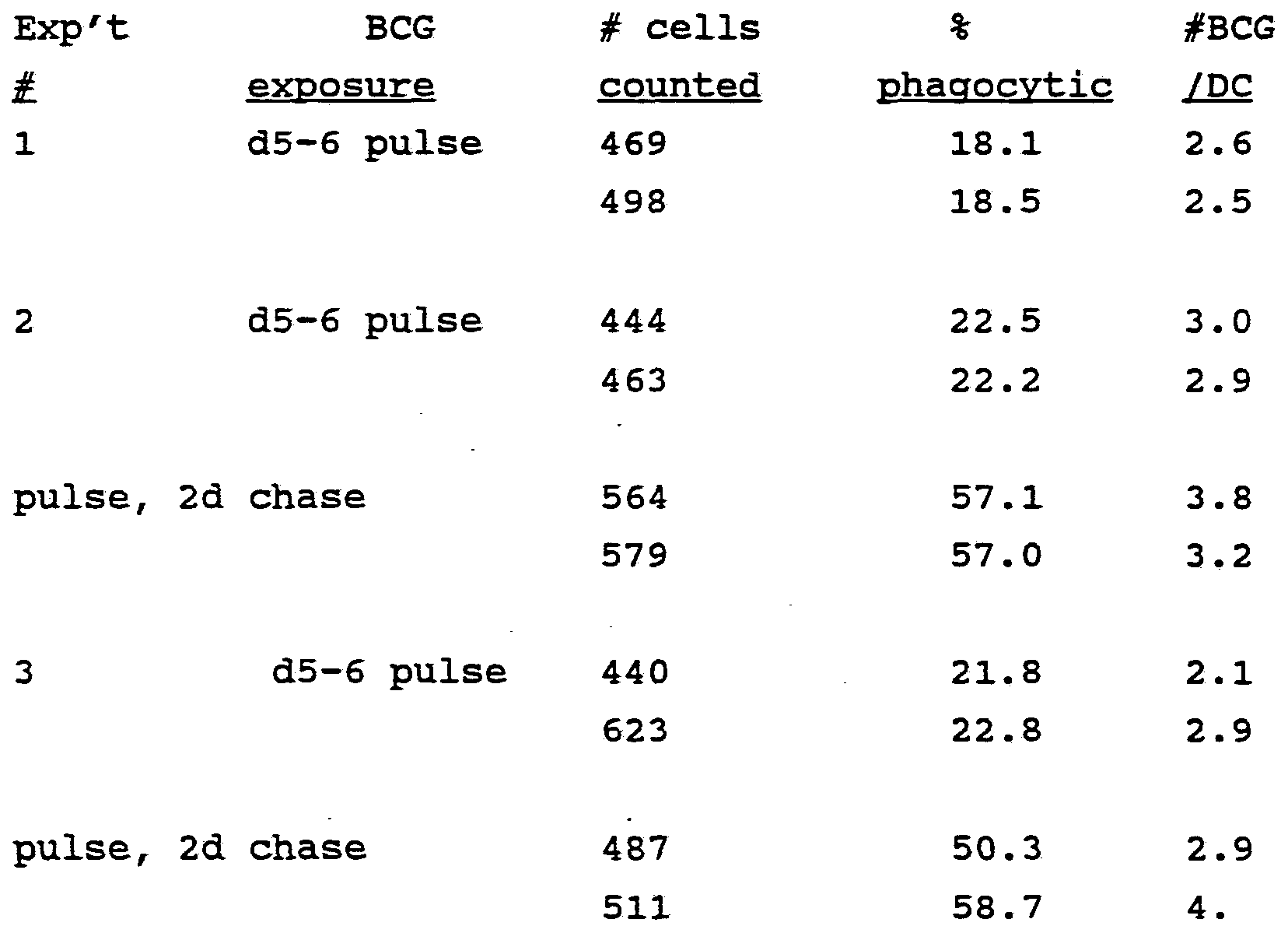
Claims
Priority Applications (6)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| AT93911581T ATE260971T1 (en) | 1992-04-01 | 1993-04-01 | METHOD FOR THE IN VITRO CULTIVATION OF DENDRITIC PRECURSOR CELLS AND THEIR USE FOR IMMUNOGENIC PRODUCTION |
| CA2133409A CA2133409C (en) | 1992-04-01 | 1993-04-01 | Method for in vitro proliferation of dendritic cell precursors and their use to produce immunogens |
| DE69333433T DE69333433T2 (en) | 1992-04-01 | 1993-04-01 | METHOD FOR THE VITRO CULTIVATION OF DENDRITIC PROCUREMENT CELLS AND THE USE THEREOF FOR IMMUNOGENOUS PRODUCTION |
| EP93911581A EP0633929B1 (en) | 1992-04-01 | 1993-04-01 | METHOD FOR $i(IN VITRO) PROLIFERATION OF DENDRITIC CELL PRECURSORS AND THEIR USE TO PRODUCE IMMUNOGENS |
| AU40461/93A AU687733B2 (en) | 1992-04-01 | 1993-04-01 | Method for in vitro proliferation of dendritic cell precursors and their use to produce immunogens |
| JP51773893A JP3649335B2 (en) | 1992-04-01 | 1993-04-01 | Method for in vitro expansion of dendritic cell precursors and their use in immunogen production |
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US86161292A | 1992-04-01 | 1992-04-01 | |
| US07/861,612 | 1992-04-01 | ||
| US98135792A | 1992-11-25 | 1992-11-25 | |
| US07/981,357 | 1992-11-25 | ||
| US08/040,677 | 1993-03-31 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO1993020185A1 true WO1993020185A1 (en) | 1993-10-14 |
Family
ID=27127636
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/US1993/003141 WO1993020185A1 (en) | 1992-04-01 | 1993-04-01 | Method for in vitro proliferation of dendritic cell precursors and their use to produce immunogens |
Country Status (2)
| Country | Link |
|---|---|
| AU (1) | AU687733B2 (en) |
| WO (1) | WO1993020185A1 (en) |
Cited By (47)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1995028479A1 (en) * | 1994-04-14 | 1995-10-26 | KLINIKUM DER ALBERT-LUDWIGS-UNIVERSITäT FREIBURG | Process for producing dendritic cells, cells thus produced and container for carrying out this process |
| WO1997003186A2 (en) * | 1995-07-12 | 1997-01-30 | Activated Cell Therapy, Inc. | Method for in vitro proliferation of dentritic cells, composition containing the cells entrapped in a three-dimensional matrix and use for immunization |
| EP0765386A1 (en) * | 1994-06-14 | 1997-04-02 | The Board Of Trustees Of The Leland Stanford Junior University | Methods for in vivo t cell activation by antigen-pulsed dendritic cells |
| US5648248A (en) * | 1994-12-30 | 1997-07-15 | Boehringer Ingelheim International Gmbh | Methods for producing differentiated cells from immature hematopoietic cells |
| US5648219A (en) * | 1995-06-07 | 1997-07-15 | Zymogenetics, Inc. | Immortalized dendritic cells |
| US5849589A (en) * | 1996-03-11 | 1998-12-15 | Duke University | Culturing monocytes with IL-4, TNF-α and GM-CSF TO induce differentiation to dendric cells |
| EP0896506A1 (en) * | 1996-03-22 | 1999-02-17 | Yale University | Methods for inducing immune responsiveness in a subject |
| US5994126A (en) * | 1992-04-01 | 1999-11-30 | The Rockefeller University | Method for in vitro proliferation of dendritic cell precursors and their use to produce immunogens |
| US6004807A (en) * | 1992-03-30 | 1999-12-21 | Schering Corporation | In vitro generation of human dendritic cells |
| WO2000018933A1 (en) * | 1998-09-30 | 2000-04-06 | American Foundation For Biological Research, Inc. | Immunotherapy of cancer through expression of truncated tumor or tumor-associated antigen |
| US6080409A (en) * | 1995-12-28 | 2000-06-27 | Dendreon Corporation | Immunostimulatory method |
| EP1030909A1 (en) * | 1998-09-11 | 2000-08-30 | KOHN, Leonard D. | Immune activation by double-stranded polynucleotides |
| US6274378B1 (en) | 1997-10-27 | 2001-08-14 | The Rockefeller University | Methods and compositions for obtaining mature dendritic cells |
| US6300090B1 (en) | 1994-07-29 | 2001-10-09 | The Rockefeller University | Methods of use of viral vectors to deliver antigen to dendritic cells |
| EP1176986A1 (en) * | 1999-04-20 | 2002-02-06 | Richard L. Edelson | Differentiation of monocytes into functional dendritic cells |
| US6355238B1 (en) | 1992-11-18 | 2002-03-12 | Yale University | Specific immune system modulation |
| US6440735B1 (en) | 1998-03-31 | 2002-08-27 | Geron Corporation | Dendritic cell vaccine containing telomerase reverse transcriptase for the treament of cancer |
| EP1237929A1 (en) * | 1999-12-03 | 2002-09-11 | The Corporation Of The Trustees Of The Order Of The Sisters Of Mercy In Queensland | Dendritic cell-specific antibodies |
| US6458585B1 (en) | 1996-08-14 | 2002-10-01 | Nexell Therapeutics Inc. | Cytokine-free culture of dendritic cells |
| WO2003030965A2 (en) * | 2001-09-27 | 2003-04-17 | Martin Scholz | Artificial lymph node |
| US6602709B1 (en) | 1998-02-20 | 2003-08-05 | The Rockefeller University | Methods for use of apoptotic cells to deliver antigen to dendritic cells for induction or tolerization of T cells |
| US6652848B1 (en) | 1997-04-15 | 2003-11-25 | Dana Farber Cancer Institute, Inc. | Dendritic cell hybrids |
| WO2006031870A2 (en) | 2004-09-14 | 2006-03-23 | Argos Therapeutics, Inc. | Strain independent amplification of pathogens and vaccines thereto |
| US7030228B1 (en) | 1999-11-15 | 2006-04-18 | Miltenyi Biotec Gmbh | Antigen-binding fragments specific for dendritic cells, compositions and methods of use thereof antigens recognized thereby and cells obtained thereby |
| US7067110B1 (en) * | 1999-07-21 | 2006-06-27 | Emd Lexigen Research Center Corp. | Fc fusion proteins for enhancing the immunogenicity of protein and peptide antigens |
| US7109031B2 (en) | 1999-04-20 | 2006-09-19 | Yale University | Methods for inducing the differentiation of monocytes into functional dendritic cells and immunotherapeutic compositions including such dendritic cells |
| US7198948B2 (en) | 1996-02-12 | 2007-04-03 | The Rockefeller University | Methods and compositions for obtaining mature dendritic cells |
| US7361346B1 (en) * | 1996-05-15 | 2008-04-22 | Altarex Corp. | Therapeutic compositions that produce an immune response |
| US7402307B2 (en) * | 1998-03-31 | 2008-07-22 | Geron Corporation | Method for identifying and killing cancer cells |
| US7419957B2 (en) | 2001-08-22 | 2008-09-02 | The United States Of America As Represented By The Secretary Of The Department Of Health And Human Services | Peptides of melanoma antigen and their use in diagnostic, prophylactic and therapeutic methods |
| US7534434B2 (en) | 2004-12-28 | 2009-05-19 | The Rockefeller University | Glycolipids and analogues thereof as antigens for NK T cells |
| US7601342B2 (en) | 1997-04-15 | 2009-10-13 | Dana Farber Cancer Institute | Cell fusions and methods of making and using the same |
| US7837990B2 (en) | 2005-03-28 | 2010-11-23 | The Rockefeller University | In vivo expanded NKT cells and methods of use thereof |
| US7867977B2 (en) | 2005-11-03 | 2011-01-11 | The United States Of America As Represented By The Department Of Health And Human Services | Immunogenic peptides and methods of use for treating and preventing cancer |
| US7923013B2 (en) | 2004-12-28 | 2011-04-12 | The Rockefeller University | Glycolipids and analogues thereof as antigens for NKT cells |
| EP2311952A2 (en) | 1994-04-22 | 2011-04-20 | The United States Of America,As Represented By The Secretary, Department Of Health & Human Services | Antibodies to MRT-1 protein or fragments thereof |
| US8038994B2 (en) | 1996-05-15 | 2011-10-18 | Quest Pharmatech Inc. | Combination therapy for treating disease |
| US8313945B2 (en) | 1999-04-20 | 2012-11-20 | Yale University | Methods for inducing the differentiation of blood monocytes into functional dendritic cells |
| WO2012164004A1 (en) | 2011-06-01 | 2012-12-06 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Novel antigen peptide and uses thereof |
| EP2606897A1 (en) | 2011-12-22 | 2013-06-26 | Laboratorios Del. Dr. Esteve, S.A. | Methods and compositions for the treatment of diseases caused by enveloped viruses |
| US8524495B2 (en) | 2007-05-16 | 2013-09-03 | Yale University | Methods for inducing the differentiation of blood monocytes into functional dendritic cells |
| WO2013175258A1 (en) | 2012-05-22 | 2013-11-28 | Inserm (Institut National De La Sante Et De La Recherche Medicale) | Novel melanoma antigen peptide and uses thereof |
| WO2013175256A1 (en) | 2012-05-22 | 2013-11-28 | Inserm (Institut National De La Sante Et De La Recherche Medicale) | Novel melanoma antigen peptide and uses thereof |
| US8926973B2 (en) | 2001-03-30 | 2015-01-06 | Merck Patent Gmbh | Reducing the immunogenicity of fusion proteins |
| WO2015150492A2 (en) | 2014-04-01 | 2015-10-08 | INSERM (Institut National de la Santé et de la Recherche Médicale) | An isolated donor mhc-derived peptide and uses thereof |
| WO2015150491A1 (en) | 2014-04-01 | 2015-10-08 | INSERM (Institut National de la Santé et de la Recherche Médicale) | An isolated donor mhc-derived peptide and uses thereof |
| US10272112B2 (en) | 2013-11-18 | 2019-04-30 | University of Pittsburgh—of the Commonwealth System of Higher Education | Microsphere-based delivery and ex vivo manipulation of dendritic cells for autoimmune therapies |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1991013632A1 (en) * | 1990-03-14 | 1991-09-19 | The Immune Response Corporation | Idiotypic vaccination against b cell lymphoma |
-
1993
- 1993-04-01 AU AU40461/93A patent/AU687733B2/en not_active Expired
- 1993-04-01 WO PCT/US1993/003141 patent/WO1993020185A1/en active IP Right Grant
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1991013632A1 (en) * | 1990-03-14 | 1991-09-19 | The Immune Response Corporation | Idiotypic vaccination against b cell lymphoma |
Non-Patent Citations (6)
Cited By (83)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6004807A (en) * | 1992-03-30 | 1999-12-21 | Schering Corporation | In vitro generation of human dendritic cells |
| US7923247B2 (en) | 1992-04-01 | 2011-04-12 | Argos Therapeutics, Inc. | Method for in vitro proliferation of dendritic cell precursors and their use to produce immunogens |
| US5994126A (en) * | 1992-04-01 | 1999-11-30 | The Rockefeller University | Method for in vitro proliferation of dendritic cell precursors and their use to produce immunogens |
| US6355238B1 (en) | 1992-11-18 | 2002-03-12 | Yale University | Specific immune system modulation |
| WO1995028479A1 (en) * | 1994-04-14 | 1995-10-26 | KLINIKUM DER ALBERT-LUDWIGS-UNIVERSITäT FREIBURG | Process for producing dendritic cells, cells thus produced and container for carrying out this process |
| US5866115A (en) * | 1994-04-14 | 1999-02-02 | Klinikum Der Albert-Ludwigs-Universitat Freiburg | Process for preparing dendritic cells, cells thus produced and containers for carrying out this process |
| EP2311952A2 (en) | 1994-04-22 | 2011-04-20 | The United States Of America,As Represented By The Secretary, Department Of Health & Human Services | Antibodies to MRT-1 protein or fragments thereof |
| EP0765386A1 (en) * | 1994-06-14 | 1997-04-02 | The Board Of Trustees Of The Leland Stanford Junior University | Methods for in vivo t cell activation by antigen-pulsed dendritic cells |
| EP0765386A4 (en) * | 1994-06-14 | 1998-01-07 | Univ Leland Stanford Junior | Methods for in vivo t cell activation by antigen-pulsed dendritic cells |
| US6300090B1 (en) | 1994-07-29 | 2001-10-09 | The Rockefeller University | Methods of use of viral vectors to deliver antigen to dendritic cells |
| US5648248A (en) * | 1994-12-30 | 1997-07-15 | Boehringer Ingelheim International Gmbh | Methods for producing differentiated cells from immature hematopoietic cells |
| US5648219A (en) * | 1995-06-07 | 1997-07-15 | Zymogenetics, Inc. | Immortalized dendritic cells |
| US6121044A (en) * | 1995-07-12 | 2000-09-19 | Dendreon Corporation | Potent antigen presenting cell composition |
| WO1997003186A3 (en) * | 1995-07-12 | 1997-05-09 | Activated Cell Therapy Inc | Method for in vitro proliferation of dentritic cells, composition containing the cells entrapped in a three-dimensional matrix and use for immunization |
| WO1997003186A2 (en) * | 1995-07-12 | 1997-01-30 | Activated Cell Therapy, Inc. | Method for in vitro proliferation of dentritic cells, composition containing the cells entrapped in a three-dimensional matrix and use for immunization |
| US6080409A (en) * | 1995-12-28 | 2000-06-27 | Dendreon Corporation | Immunostimulatory method |
| US6210662B1 (en) | 1995-12-28 | 2001-04-03 | Dendreon Corporation | Immunostimulatory composition |
| US7659119B2 (en) | 1996-02-12 | 2010-02-09 | Argos Therapeutics, Inc. | Method and compositions for obtaining mature dendritic cells |
| US7198948B2 (en) | 1996-02-12 | 2007-04-03 | The Rockefeller University | Methods and compositions for obtaining mature dendritic cells |
| US5849589A (en) * | 1996-03-11 | 1998-12-15 | Duke University | Culturing monocytes with IL-4, TNF-α and GM-CSF TO induce differentiation to dendric cells |
| EP0896506A1 (en) * | 1996-03-22 | 1999-02-17 | Yale University | Methods for inducing immune responsiveness in a subject |
| EP0896506A4 (en) * | 1996-03-22 | 2003-06-18 | Univ Yale | Methods for inducing immune responsiveness in a subject |
| US7361346B1 (en) * | 1996-05-15 | 2008-04-22 | Altarex Corp. | Therapeutic compositions that produce an immune response |
| US8038994B2 (en) | 1996-05-15 | 2011-10-18 | Quest Pharmatech Inc. | Combination therapy for treating disease |
| US6458585B1 (en) | 1996-08-14 | 2002-10-01 | Nexell Therapeutics Inc. | Cytokine-free culture of dendritic cells |
| US6652848B1 (en) | 1997-04-15 | 2003-11-25 | Dana Farber Cancer Institute, Inc. | Dendritic cell hybrids |
| US7601342B2 (en) | 1997-04-15 | 2009-10-13 | Dana Farber Cancer Institute | Cell fusions and methods of making and using the same |
| EP2058389A1 (en) | 1997-10-27 | 2009-05-13 | Rockefeller University | Defined dendritic cell maturation medium comprising TNF-alpha, IL-1beta, IL-6, GM-CSF and IL-4 |
| US6274378B1 (en) | 1997-10-27 | 2001-08-14 | The Rockefeller University | Methods and compositions for obtaining mature dendritic cells |
| US7129037B2 (en) | 1998-02-20 | 2006-10-31 | The Rockefeller University | Methods for use of apoptotic cells to deliver antigen to dendritic cells for induction or tolerization of T cells |
| US7989203B2 (en) | 1998-02-20 | 2011-08-02 | The Rockefeller University | Methods for use of apoptotic cells to deliver antigen to dendritic cells for induction or tolerization of T cells |
| US6602709B1 (en) | 1998-02-20 | 2003-08-05 | The Rockefeller University | Methods for use of apoptotic cells to deliver antigen to dendritic cells for induction or tolerization of T cells |
| US7402307B2 (en) * | 1998-03-31 | 2008-07-22 | Geron Corporation | Method for identifying and killing cancer cells |
| US7824849B2 (en) | 1998-03-31 | 2010-11-02 | Geron Corporation | Cellular telomerase vaccine and its use for treating cancer |
| US6440735B1 (en) | 1998-03-31 | 2002-08-27 | Geron Corporation | Dendritic cell vaccine containing telomerase reverse transcriptase for the treament of cancer |
| EP1030909A1 (en) * | 1998-09-11 | 2000-08-30 | KOHN, Leonard D. | Immune activation by double-stranded polynucleotides |
| EP1030909A4 (en) * | 1998-09-11 | 2005-11-16 | Medical Inc Sentron | Immune activation by double-stranded polynucleotides |
| US6387888B1 (en) | 1998-09-30 | 2002-05-14 | American Foundation For Biological Research, Inc. | Immunotherapy of cancer through expression of truncated tumor or tumor-associated antigen |
| WO2000018933A1 (en) * | 1998-09-30 | 2000-04-06 | American Foundation For Biological Research, Inc. | Immunotherapy of cancer through expression of truncated tumor or tumor-associated antigen |
| US8313945B2 (en) | 1999-04-20 | 2012-11-20 | Yale University | Methods for inducing the differentiation of blood monocytes into functional dendritic cells |
| US7109031B2 (en) | 1999-04-20 | 2006-09-19 | Yale University | Methods for inducing the differentiation of monocytes into functional dendritic cells and immunotherapeutic compositions including such dendritic cells |
| EP1176986A1 (en) * | 1999-04-20 | 2002-02-06 | Richard L. Edelson | Differentiation of monocytes into functional dendritic cells |
| EP1176986A4 (en) * | 1999-04-20 | 2004-06-16 | Edelson Richard Leslie | Differentiation of monocytes into functional dendritic cells |
| US6607722B2 (en) | 1999-04-20 | 2003-08-19 | Richard Leslie Edelson | Methods for inducing the differentiation of monocytes into functional dendritic cells |
| US6524855B2 (en) | 1999-04-20 | 2003-02-25 | Richard Leslie Edelson | Methods for inducing the differentiation of monocytes into functional dendritic cells |
| US7067110B1 (en) * | 1999-07-21 | 2006-06-27 | Emd Lexigen Research Center Corp. | Fc fusion proteins for enhancing the immunogenicity of protein and peptide antigens |
| US8183039B2 (en) | 1999-11-15 | 2012-05-22 | Miltenyi Biotec Gmbh | Method for isolating BDCA-4+ dendritic cells |
| US10407486B2 (en) | 1999-11-15 | 2019-09-10 | Miltenyi Biotech Gmbh | Ligating BDCA-2 protein with BDCA-2 specific antibody for detecting or modulating dendritic cells |
| US7030228B1 (en) | 1999-11-15 | 2006-04-18 | Miltenyi Biotec Gmbh | Antigen-binding fragments specific for dendritic cells, compositions and methods of use thereof antigens recognized thereby and cells obtained thereby |
| EP1237929A4 (en) * | 1999-12-03 | 2004-04-28 | Order Sisters Of Mercy Queensl | Dendritic cell-specific antibodies |
| AU779803B2 (en) * | 1999-12-03 | 2005-02-10 | Corporation Of The Trustees Of The Order Of The Sisters Of Mercy In Queensland, The | Dendritic cell-specific antibodies |
| EP1237929A1 (en) * | 1999-12-03 | 2002-09-11 | The Corporation Of The Trustees Of The Order Of The Sisters Of Mercy In Queensland | Dendritic cell-specific antibodies |
| US8926973B2 (en) | 2001-03-30 | 2015-01-06 | Merck Patent Gmbh | Reducing the immunogenicity of fusion proteins |
| US7749967B2 (en) | 2001-08-22 | 2010-07-06 | The United States Of America As Represented By The Department Of Health And Human Services. | Peptides of a melanoma antigen and their use in diagnostic, prophylactic, and therapeutic methods |
| US8252545B2 (en) | 2001-08-22 | 2012-08-28 | The United States Of America, As Represented By The Secretary Department Of Health And Human Services | Peptides of a melanoma antigen and their use in diagnostic, prophylactic, and therapeutic methods |
| US7419957B2 (en) | 2001-08-22 | 2008-09-02 | The United States Of America As Represented By The Secretary Of The Department Of Health And Human Services | Peptides of melanoma antigen and their use in diagnostic, prophylactic and therapeutic methods |
| WO2003030965A2 (en) * | 2001-09-27 | 2003-04-17 | Martin Scholz | Artificial lymph node |
| WO2003030965A3 (en) * | 2001-09-27 | 2004-04-01 | Martin Scholz | Artificial lymph node |
| WO2006031870A2 (en) | 2004-09-14 | 2006-03-23 | Argos Therapeutics, Inc. | Strain independent amplification of pathogens and vaccines thereto |
| EP2742951A2 (en) | 2004-09-14 | 2014-06-18 | Argos Therapeutics, Inc. | Strain independent amplification of HIV polynucleotides |
| US8163290B2 (en) | 2004-12-28 | 2012-04-24 | The Rockefeller University | Glycolipids and analogues thereof as antigens for NKT cells |
| US7534434B2 (en) | 2004-12-28 | 2009-05-19 | The Rockefeller University | Glycolipids and analogues thereof as antigens for NK T cells |
| US7923013B2 (en) | 2004-12-28 | 2011-04-12 | The Rockefeller University | Glycolipids and analogues thereof as antigens for NKT cells |
| US8586051B2 (en) | 2004-12-28 | 2013-11-19 | The Rockefeller University | Glycolipids and analogues thereof as antigens for NKT cells |
| US7837990B2 (en) | 2005-03-28 | 2010-11-23 | The Rockefeller University | In vivo expanded NKT cells and methods of use thereof |
| US8614304B2 (en) | 2005-11-03 | 2013-12-24 | The United States of America, as represented by the Secretary, Department of Health and Human Serives | Immunogenic peptides and methods of use for treating and preventing cancer |
| US9206245B2 (en) | 2005-11-03 | 2015-12-08 | The United States Of America, As Represented By The Secretary, Department Of Health And Human Services | Immunogenic peptides and methods of use for treating and preventing cancer |
| US7867977B2 (en) | 2005-11-03 | 2011-01-11 | The United States Of America As Represented By The Department Of Health And Human Services | Immunogenic peptides and methods of use for treating and preventing cancer |
| US8524495B2 (en) | 2007-05-16 | 2013-09-03 | Yale University | Methods for inducing the differentiation of blood monocytes into functional dendritic cells |
| US9321991B2 (en) | 2010-04-30 | 2016-04-26 | Yale University | Methods for inducing the differentiation of blood monocytes into functional dendritic cells |
| US10087418B2 (en) | 2010-04-30 | 2018-10-02 | Yale University | Methods for inducing the differentiation of blood monocytes into functional dendritic cells |
| US10934526B2 (en) | 2010-04-30 | 2021-03-02 | Yale University | Methods for inducing the differentiation of blood monocytes into functional dendritic cells |
| WO2012164004A1 (en) | 2011-06-01 | 2012-12-06 | INSERM (Institut National de la Santé et de la Recherche Médicale) | Novel antigen peptide and uses thereof |
| WO2013092509A1 (en) | 2011-12-22 | 2013-06-27 | Laboratorios Del Dr. Esteve, S.A. | Inhibitors of sialoadhesin for the treatment of diseases caused by enveloped viruses |
| EP2606897A1 (en) | 2011-12-22 | 2013-06-26 | Laboratorios Del. Dr. Esteve, S.A. | Methods and compositions for the treatment of diseases caused by enveloped viruses |
| US9789130B2 (en) | 2011-12-22 | 2017-10-17 | Laboratorios Del Dr. Esteve, S.A. | Method for blocking HIV entry into dendritic cells by inhibiting sialoadhesin-sialyllactose binding interactions with gangliosides |
| WO2013175258A1 (en) | 2012-05-22 | 2013-11-28 | Inserm (Institut National De La Sante Et De La Recherche Medicale) | Novel melanoma antigen peptide and uses thereof |
| WO2013175256A1 (en) | 2012-05-22 | 2013-11-28 | Inserm (Institut National De La Sante Et De La Recherche Medicale) | Novel melanoma antigen peptide and uses thereof |
| US10940164B2 (en) | 2013-11-18 | 2021-03-09 | University of Pittsburgh—of the Commonwealth System of Higher Education | Microsphere-based delivery and ex vivo manipulation of dendritic cells for autoimmune therapies |
| US10272112B2 (en) | 2013-11-18 | 2019-04-30 | University of Pittsburgh—of the Commonwealth System of Higher Education | Microsphere-based delivery and ex vivo manipulation of dendritic cells for autoimmune therapies |
| WO2015150491A1 (en) | 2014-04-01 | 2015-10-08 | INSERM (Institut National de la Santé et de la Recherche Médicale) | An isolated donor mhc-derived peptide and uses thereof |
| EP3495377A1 (en) | 2014-04-01 | 2019-06-12 | INSERM (Institut National de la Santé et de la Recherche Médicale) | An isolated donor mhc-derived peptide and uses thereof |
| WO2015150492A2 (en) | 2014-04-01 | 2015-10-08 | INSERM (Institut National de la Santé et de la Recherche Médicale) | An isolated donor mhc-derived peptide and uses thereof |
Also Published As
| Publication number | Publication date |
|---|---|
| AU4046193A (en) | 1993-11-08 |
| AU687733B2 (en) | 1998-03-05 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| EP0633929B1 (en) | METHOD FOR $i(IN VITRO) PROLIFERATION OF DENDRITIC CELL PRECURSORS AND THEIR USE TO PRODUCE IMMUNOGENS | |
| AU687733B2 (en) | Method for in vitro proliferation of dendritic cell precursors and their use to produce immunogens | |
| US10226519B2 (en) | Cancer vaccines and vaccination methods | |
| Inaba et al. | Dendritic cell progenitors phagocytose particulates, including bacillus Calmette-Guerin organisms, and sensitize mice to mycobacterial antigens in vivo. | |
| US7659119B2 (en) | Method and compositions for obtaining mature dendritic cells | |
| US20180346887A1 (en) | Immunogenic peptides for use in the prevention and/or treatment of infectious diseases, autoimmune diseases, immune responses to allofactors, allergic diseases, tumors, graft rejection and immune responses against viral vectors used for gene therapy or gene vaccination | |
| Bernhard et al. | Generation of immunostimulatory dendritic cells from human CD34+ hematopoietic progenitor cells of the bone marrow and peripheral blood | |
| US6274378B1 (en) | Methods and compositions for obtaining mature dendritic cells | |
| WO1997029182A9 (en) | Method and compositions for obtaining mature dendritic cells | |
| JPH11505512A (en) | Method for preparing macrophages, and kits and compositions therefor | |
| US8563308B2 (en) | Culture-expanded T suppressor cells and methods of use thereof | |
| WO2020038299A1 (en) | Exosome-based antitumor vaccine | |
| US20030124091A1 (en) | Endothelial cell derived hematopoietic growth factor | |
| JP2000503545A (en) | Novel antigen presenting cells, methods for their preparation, and their use as cellular vaccines | |
| US20020085993A1 (en) | Method for in vitro proliferation of dendritic cell precursors and their use to produce immunogens | |
| Vuckovic et al. | Growth factors, cytokines and dendritic cell development | |
| JP2003508047A (en) | Use of cytokines, cells and mitogens to suppress graft-versus-host disease | |
| Robinson et al. | Dendritic cell protocols | |
| Denis et al. | cells from peripheral blood |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| AK | Designated states |
Kind code of ref document: A1 Designated state(s): AU CA FI JP |
|
| AL | Designated countries for regional patents |
Kind code of ref document: A1 Designated state(s): AT BE CH DE DK ES FR GB GR IE IT LU MC NL PT SE |
|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application | ||
| DFPE | Request for preliminary examination filed prior to expiration of 19th month from priority date (pct application filed before 20040101) | ||
| WWE | Wipo information: entry into national phase |
Ref document number: 1993911581 Country of ref document: EP |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 2133409 Country of ref document: CA |
|
| WWP | Wipo information: published in national office |
Ref document number: 1993911581 Country of ref document: EP |
|
| WWG | Wipo information: grant in national office |
Ref document number: 1993911581 Country of ref document: EP |