DESCRIPTION
Development of Novel Anti-Microbial Agents Based on Bacteriophage Genomics
BACKGROUND OF THE INVENTION
The present invention relates to the field of antibacterial agents and the treatment of infections of animals or other complex organisms by bacteria.
The frequency and spectrum of antibiotic-resistant infections have, in recent years, increased in both the hospital and community. Certain infections have become essentially untreatable and are growing to epidemic proportions in the developing world as well as in institutional settings in the developed world. The staggering spread of antibiotic resistance in pathogenic bacteria has been attributed to microbial genetic characteristics, widespread use of antibiotic drugs, and changes in society that enhance the transmission of drug-resistant organisms. This spread of drug resistant microbes is leading to ever increasing morbidity, mortality and health-care costs.
Ironically, it is the very success of antibiotics, resulting in their widespread use, that has contributed the most to rising numbers of drug resistant bacterial strains. The longer a bacterial strain is exposed to a drug, the more likely it is to acquire resistance. Today, a total of 160 antibiotics, all based on a few basic chemical structures and targeting a small number of metabolic pathways, have found their way to market. Over-prescription of these drugs, as well as the failure of patients to comply with the complete antibiotic regimen, has lead to the rapid emergence of antibiotic resistant strains. Such misuse of prescriptions, careless use of antibiotics in virtually all commercial production of beef and fowl, and changing societal conditions, such as the growth of day-care centers, increased long-term care in hospitals, and increased mobility of the population, has provided an environment where drug-resistant microbes can emerge and spread. Thus, virtually all common infectious bacteria are becoming, or have already become, resistant to one or more groups of antibiotics. Such resistance now reaches all classes of antibiotics currently in use, including: β-lactams, fluoroquinolones, aminoglycosides, macrolide peptides, chloramphenicol, tetracyclines, rifampicin, folate inhibitors, glycopeptides, and - mupirocin. Over the last 45 years bacteria have adapted genetically to avoid the destruction/alteration of the essential pathways that these chemotherapeutic agents
target. Antibiotic resistant bacterial strains are now emerging at a higher rate than the rate at which new antibiotics are being developed. The consequence of this dilemma has been a dramatic increase in the cost of treating infections what would otherwise easily succumb to routine antibiotic therapy. Furthermore, and perhaps most importantly, the emergence of multiple drug resistant pathogenic bacteria has led to a significant increase in morbidity and mortality, particularly in institutional settings.
Most major pharmaceutical companies have on-going drug discovery programs for novel anti-microbials. These are based on screens for small molecule inhibitors (natural products, bacterial culture media, libraries of small molecules, combinatorial chemistry) of crucial metabolic pathways of the micro-organism of interest (e.g., bacteria, fungi, parasites, worms). The screening process is largely for cytotoxic compounds and in most cases is not based on a known mechanism of action of the compounds. Pharmaceutical companies have large programs in this area. Classical drug screening programs are being exhausted and many of these pharmaceutical companies are looking towards rational drug design programs. Several small to mid-size biotechnology companies as well as large pharmaceutical companies have developed systematic high-throughput sequencing programs to decipher the genetic code of specific micro-organisms of interest. The goal is to identify, through sequencing, unique biochemical pathways or intermediates that are unique to the microorganism. Knowledge of this may, in turn, form the rationale for a drug discovery program based on the mechanism of action of the identified enzymes/proteins. Genome Therapeutics Corp., The Institute for Genome Research, Human Genome Sciences Inc., and other companies have such sequencing programs in place. However, one of the most critical steps in this approach is the ascertainment that the identified proteins and biochemical pathways are 1) non- redundant and essential for bacterial survival, and 2) constitute suitable and accessible targets for drug discovery.
SUMMARY OF THE INVENTION
While animals such as humans are, on occasion, infected by pathogenic bacteria, bacteria also have natural enemies. A number of host-specific viruses, known as bacteriophages or phages, infect and kill bacteria in the natural environment. Such bacteriophages generally have small compact genomes and bacteria are their exclusive hosts. Many known bacteria are host to a large number of bacteriophages that have been described in the literature. During the 1940's - 1960's, phage biology was an area of active research. As a testimony to this, the study of phages which infect and inhibit the enteric bacterium Escherichia coli (E. coli) contributed much to the early understanding of molecular biology and virology.
As is generally understood, bacteriophage (or phages) are viruses that infect and kill bacteria. They are natural enemies of bacteria and, over the course of evolution, have developed proteins (products of DNA sequences) which enable them to infect a host bacteria, replicate their genetic material, usurp host metabolism, and ultimately kill their host. The scientific literature well documents the fact that many known bacteria have a large number of such bacteriophages (Ackermann and DuBow, 1987) that can infect and kill them (for example, see the ATCC bacteriophage collection at http://www.atcc.org). This invention utilizes the observation that bacteriophages successfully infect and inhibit or kill host bacteria, targeting a variety of normal host metabolic and physiological traits, some of which are shared by all bacteria, pathogenic and nonpathogenic alike. The term "pathogenic" as used herein denotes a contribution to or implication in disease or a morbid state of an infected organism. The invention thus involves identifying and elucidating the molecular mechanisms by which phages interfere with host bacterial metabolism, an objective being to provide novel targets for drug design. Whether the phage blocks bacterial RNA transcription or translation, or attacks other important metabolic pathways, such as cell wall assembly or membrane integrity, the basic blueprint for a phage 's bacteria-inhibiting ability is encoded in its genome and can be unlocked using bioinformatics, functional genomics, and proteomics. By these means, the invention utilizes sequence information from the genomics of bacteriophage to identify novel antimicrobials that can be further used to actively and/or prophylactically treat bacterial infection.
Two important components of the invention thus are: i) the identification of bacteria-inhibiting phage open reading frames ("ORF's) and corresponding products that can be used to develop antibiotics based on amino acid sequence and secondary structural characteristics of the ORF products, and ii) the use of bacteriophages to map
out essential bacterial target genes and homologs, which can in turn lead to the development of suitable anti-microbial agents. These two avenues represent new and general methods for developing novel antimicrobials.
The invention thus concerns the identification of bacteriophage ORFs that supply bacteria-inhibiting functions. In this regard, use of the terms "inhibit", "inhibition", "inhibitory", and "inhibitor" all refer to a function of reducing a biological activity or function. Such reduction in activity or function can, for example, be in connection with a cellular component, e.g., an enzyme, or in connection with a cellular process, e.g., synthesis of a particular protein, or in connection with an overall process of a cell, e.g., cell growth. In reference to bacterial cell growth, for example, an inhibitory effect (i.e., a bacteria-inhibiting effect) may be bacteriocidal (killing of bacterial cells) or bacteriostatic (i.e., stopping or at least slowing bacterial cell growth). The latter slows or prevents cell growth such that fewer cells of the strain are produced relative to uninhibited cells over a given period of time. From a molecular standpoint, such inhibition may equate with a reduction in the level of, or elimination of, the transcription and/or translation of a specific bacterial target(s), or reduction or elimination of activity of a particular target biomolecule.
It is particularly advantageous to evaluate a plurality of different phage ORFs for inhibitory activity that may be from one, but is preferably from a plurality of different phage. For example, evaluating ORFs from a number of different phage of the same bacterial host provides at least two advantages. One is that the multiple phages will provide identification of a variety of different targets. Second, it is likely that multiple phage will utilize the same cellular target As used herein, the terms "bacteriophage" and "phage" are used interchangeably to refer to a virus which can infect a bacterial strain or a number of different bacterial strains.
In the context of this invention, the term "bacteriophage ORF" or ""phage ORF" or similar term refers to a nucleotide sequence in or from a bacteriophage. In connection with a particular ORF, the terms refer an open reading frame which has at least 95% sequence identity, preferably at least 97% sequence identity, more preferably at least 98% sequence identity with an ORF from the particular phage identified herein (e.g., with an ORF as identified herein) or to a nucleic acid sequence which has the specified sequence identify percentage with such an ORF sequence. A first aspect of the invention thus provides a method for identifying a _ bacteriophage nucleic acid coding region encoding a product active on an essential bacterial target by identifying a nucleic acid sequence encoding a gene product which
provides a bacteria-inhibiting function when the bacteriophage infects a host bacterium, preferably one that is an animal or plant pathogen, more preferably a bird or mammalian pathogen, and most preferably a human pathogen. The bacteriophage is an uncharacterized bacteriophage. Thus, the method excludes, for example, phage λ, φxl74, ml 3 and other E. cob-specific bacteriophage that have been studied with respect to gene number and/or function. It also excludes, for example, the nucleic acid coding regions described in Tables 12-14, and in preferred embodiments, excludes the phage in which those regions are naturally located.
In connection with bacteriophage, the term "uncharacterized" means that a certain bacteriophage's genome has not yet been fully identified such that the genes having function involved in inhibiting host cells have not been identified. In particular, phage for which the description of genomic or protein sequence was first provided herein are uncharacterized. Phage sequences for which host bacteria- inhibiting functions have been identified prior to the filing of the present application (or alternatively prior to the present invention) are specifically excluded from the aspects involving utilization of sequences from uncharacterized bacteriophage, except that aspects may involve a plurality of phage where one or more of those phage are uncharacterized and one or more others have been characterized to some extent. A number of different bacteria-inhibiting phage ORFs are indicated in Tables 11-14. The phage ORFs or sequences identified therein are not within the term
"uncharacterized; alternatively, in preferred embodiments the phage containing those ORFs are excluded from this term. Further, any additional phage ORFs (or alternatively the phage which contain those ORFs) which have previously been described in the art as bacteria-inhibiting ORFs are expressly excluded; those ORFs or phage are known to those skilled in the art and the exclusion can be made express by specifically naming such ORFs or phage as needed (likewise for uncharacterized targets as described below). For the sake of brevity, such a listing is not expressly presented, as such information is readily available to those skilled in the art.
Stating that an agent or compound is "active on" a particular cellular target, such as the product of a particular gene, means that the target is an important part of a cellular pathway which includes that target and that the agent acts on that pathway.
Thus, in some cases the agent may act on a component upstream or downstream of the stated target, including on a regulator of that pathway or a component of that pathway. By "essential", in connection with a gene or gene product, is meant that the host cannot survive without, or is significantly growth compromised, in the ∑ ssπce depletion, or alteration of functional product. An "essential gene" is thus one that encodes a product that is beneficial, or preferably necessary, for cellular growth in
vitro in a medium appropriate for growth of a strain having a wild-type allele corresponding to the particular gene in question. Therefore, if an essential gene is inactivated or inhibited, that cell will grow significantly more slowly, preferably less than 20%), more preferably less than 10%, most preferably less than 5% of the growth rate of the uninhibited wild-type, or not at all, in the growth medium. Preferably, in the absence of activity provided by a product of the gene, the cell will not grow at all or will be non-viable, at least under culture conditions similar to the in vivo conditions normally encountered by the bacterial cell during an infection. For example, absence of the biological activity of certain enzymes involved in bacterial cell wall synthesis can result in the lysis of cells under normal osmotic conditions, even though protoplasts can be maintained under controlled osmotic conditions. In the context of the invention, essential genes are generally the preferred targets of antimicrobial agents. Essential genes can encode target molecules directly or can encode a product involved in the production, modification, or maintenance of a target molecule. A "target" refers to a biomolecule that can be acted on by an exogenous agent, thereby modulating, preferably inhibiting, growth or viability of a cell. In most cases such a target will be a nucleic acid sequence or molecule, or a polypeptide or protein. However, other types of biomolecules can also be targets, e.g., membrane lipids and cell wall structural components. The term "bacterium" refers to a single bacterial strain, and includes a single cell, and a plurality or population of cells of that strain unless clearly indicated to the contrary. In reference to bacteria or bacteriophage, the term "strain" refers to bacteria or phage having a particular genetic content. The genetic content includes genomic content as well as recombinant vectors. Thus, for example, two otherwise identical bacterial cells would represent different strains if each contained a vector, e.g., a plasmid, with different phage ORF inserts.
In preferred embodiments, the phage is Staphylococcus aureus phage 77, 3A, 96, or 44 AHJD, Enterococcus sp. phage 182, or Streptococcus pneumoniae phage Dp-1. In preferred embodiments, the phage is selected from. Preferred embodiments involve expressing at least one recombinant phage ORF(s) in a bacterial host followed by inhibition analysis of that host. Inhibition following expression of the phage ORF is indicative that the product of the ORF is active on an essential bacterial target. Such evaluation can be carried out in a variety of different formats, such as on a support matrix such as a solidified medium in a petri dish, or in liquid culture.
Preferably a plurality of phage ORFs are expressed in at least one bacterium. The plurality of phage ORFs can be from one or a plurality of phage. With respect to a single phage or at least one phage in a plurality of phages, the plurality of expressed ORFs preferably represents at least 10%>, more preferably at least 20%, 40%, or 60%, still more preferably at least 80% or 90%, and most preferably at least 95% of the ORFs in the phage genome. Preferably, for a plurality of phage, the plurality of expressed ORFs preferably represents at least 10%, more preferably at least 20%, 40%), or 60%), still more preferably at least 80%> or 90%, and most preferably at least 95%) of the ORFs in the phage genome of each phage. The plurality of phage ORFs can be expressed in a single bacterium, or in a plurality of bacteria where one ORF is expressed in each bacterium, or in a plurality of bacteria where a plurality of ORFs are expressed in at least one or in all of the plurality of bacteria, or combinations of these.
In embodiments of the above aspect (as well as in other aspects herein) in which a plurality of phage are utilized, a plurality of phage have the same bacterial host species; have different bacterial host species; or both. The plurality of phage includes at least two different phage, preferably at least 3,4,5,6,8,10,15,20, or more different phage. Indeed, more preferably, the plurality of phage will include 50, 75, 100, or more phage. As described herein, the larger number of phage is useful to provide additional target and target evaluation information useful in developing antibacterial agents, for example, by providing identification of a larger range of bacterial targets, and/or providing further indication of the suitability of a particular target (for example, utilization of a target by a number of different unrelated phage can suggest that the target is particularly stable and accessible and effective) and/or can indicate alternate sites on a target which interact with different inhibitors. Further embodiments involve confirmation of the inhibitor function of the phage ORF, such as by utilizing or incorporating a control(s) designed to confirm the inhibitory nature of the ORF(s) being evaluated. The control can, for example, be provided by expression of an inactive or partially inactive form of the ORF or ORF product, and/or by the absence of expression of the ORF or ORF product in the same or a closely comparable bacterial strain as that used for expression of the test ORF. The reduced level of activity or the absence of active ORF product in the control will thus not provide the inhibition provided by a corresponding inhibitory ORF, or will provide a distinguishably lower level of inhibition. An inactivated or partially inactivated control has a mutation(s), e.g., in the coding region or in flanking regulatory elements, that reduce(s) or eliminate(s) the normal function of the ORF. Thus, the inhibition of a bacterium following expression of a phage ORF is determined by comparison with the effects of expression of an inactivated ORF or the
response of the bacteria in the absence of expression in the same or similar type bacterium. Such determination of inhibition of the bacterium following expression of the ORF is indicative of a bacteria-inhibiting function. These manipulations are routinely understood and accomplished by those of skill in the art using standard techniques. In embodiments utilizing absence of expression of the ORF, the bacteria can, for example, contain an empty vector or a vector which allows expression of an unrelated sequence which is preferably non-inhibitory. Alternatively, the bacteria may have no vector at all. Combinations of such controls or other controls may also be utilized as recognized by those skilled in the art. In embodiments involving expression of a phage ORF in a bacterial strain, in preferred embodiments that expression is inducible.
By "inducible" is meant that expression is absent or occurs at a low level until the occurrence of an appropriate environmental stimulus provides otherwise. For the present invention such induction is preferably controlled by an artificial environmental change, such as by contacting a bacterial strain population with an inducing compound (i.e., an inducer). However, induction could also occur, for example, in response to build-up of a compound produced by the bacteria in the bacterial culture, e.g., in the medium. As uncontrolled or constitutive expression of inhibitory ORFs can severely compromise bacteria to the point of eradication, such expression is therefore undesirable in many cases because it would prevent effective evaluation of the strain and inhibitor being studied. For example, such uncontrolled expression could prevent any growth of the strain following insertion of a recombinant ORF, thus preventing determination of effective transfection or transformation. A controlled or inducible expression is therefore advantageous and is generally provided through the provision of suitable regulatory elements, e.g. , promoter/operator sequences that can be conveniently transcriptionally linked to a coding sequence to be evaluated. In most cases, the vector will also contain sequences suitable for efficient replication of the vector in the same or different host cells and/or sequences allowing selection of cells containing the vector, i.e., "selectable markers." Further, preferred vectors include convenient primer sequences flanking the cloning region from which PCR and/or sequencing may be performed. As knowledge of the nucleotide sequence of phage ORFs is useful, e.g., for assisting in the identification of phage proteins active against essential bacterial host targets, preferred embodiments involve the sequencing of at least a portion of the phage genome in combination with the above methods. This can be done either-before or after or independent of expression and inhibition of the ORF in the bacteria, and provides information on the nature and characteristics of the ORF. Such a portion is
preferably at least 10%, 20%, 40%, 80%, 90%, or 100% of the phage genome. For embodiments in which a plurality of phage are utilized, preferably each phage is sequenced to an extent as just specified.
Such sequencing is preferably accompanied by computer sequence analysis to define and evaluate ORF(s), ORF products, structural motifs or functional properties of ORF products, and/or their genetic control elements. Thus, certain embodiments incorporate computer sequence analyses or nucleic acid and/or amino acid sequences. Further, existing data banks can provide phage sequence and product information which can be utilized for analysis and identification of ORFs in the sequence. Computer analysis may further employ known homologous sequences from other species that suggest or indicate conserved underlying biochemical function(s) for the inhibitory or potentially inhibitory ORF sequence(s) being evaluated. This can include the sequences of signature motifs of identified classes of inhibitors.
In the context of the phage nucleic acid sequences, e.g., gene sequences, of this invention, the terms "homolog" and "homologous" denote nucleotide sequences from different bacteria or phage strains or species or from other types of organisms that have significantly related nucleotide sequences, and consequently significantly related encoded gene products, preferably having related function. Homologous gene sequences or coding sequences have at least 70%> sequence identity (as defined by the maximal base match in a computer-generated alignment of two or more nucleic acid sequences) over at least one sequence window of 48 nucleotides, more preferably at least 80 or 85%, still more preferably at least 90%>, and most preferably at least 95%o. The polypeptide products of homologous genes have at least 35% amino acid sequence identity over at least one sequence window of 18 amino acid residues, more preferably at least 40%>, still more preferably at least 50% or 60%>, and most preferably at least 70%>, 80%>, or 90%. Preferably, the homologous gene product is also a functional homolog, meaning that the homolog will functionally complement one or more biological activities of the product being compared. For nucleotide or amino acid sequence comparisons where a homology is defined by a % sequence identity, the percentage is determined using BLAST programs ( with default parameters (Altschul et al., 1997, "Gapped BLAST and PSI-BLAST: a new generation of protein database search programs, Nucleic Acid Res. 25:3389-3402). Any of a variety of algorithms known in the art which provide comparable results can also be used, preferably using default parameters. Performance characteristics for three different algorithms in homology searching is described in Salamov et al^ 1999, "Combining sensitive database searches with multiple intermediates to detect distant
homologues." Protein Eng. 12:95-100. Another exemplary program package is the GCG™ package from the University of Wisconsin.
Homo logs may also or in addition be characterized by the ability of two complementary nucleic acid strands to hybridize to each other under appropriately stringent conditions. Hybridizations are typically and preferably conducted with probe-length nucleic acid molecules, preferably 20-100 nucleotides in length. Those skilled in the art understand how to estimate and adjust the stringency of hybridization conditions such that sequences having at least a desired level of complementarity will stably hybridize, while those having lower complementarity will not. For examples of hybridization conditions and parameters, see, e.g.,. Maniatis, T. et al. (1989)
Molecular Cloning: A Laboratory Manual. Cold Spring Harbor University Press, Cold Spring, N.Y.; Ausubel, F.M. et al. (1994) Current Protocols in Molecular Biology. John Wiley & Sons, Secaucus, NJ. Homologs and homologous gene sequences may thus be identified using any nucleic acid sequence of interest, including the phage ORFs and bacterial target genes of the present invention.
A typical hybridization, for example, utilizes, besides the labeled probe of interest, a salt solution such as 6xSSC (NaCl and Sodium Citrate base) to stabilize nucleic acid strand interaction, a mild detergent such as 0.5%> SDS, together with other typical additives such as Denhardt's solution and salmon sperm DNA. The solution is added to the immobilized sequence to be probed and incubated at suitable temperatures to preferably permit specific binding while minimizing nonspecific binding. The temperature of the incubations and ensuing washes is critical to the success and clarity of the hybridization. Stringent conditions employ relatively higher temperatures, lower salt concentrations, and/or more detergent than do non-stringent conditions. Hybridization temperatures also depend on the length, complementarity level, and nature (ie, "GC content") of the sequences to be tested. Typical stringent hybridizations and washes are conducted at temperatures of at least 40°C, while lower stringency hybridizations and washes are typically conducted at 37°C down to room temperature (~25°C). One of skill in the art is aware that these conditions may vary according to the parameters indicated above, and that certain additives such as formamide and dextran sulphate may also be added to affect the conditions.
By "stringent hybridization conditions" is meant hybridization conditions at least as stringent as the following: hybridization in 50%> formamide, 5X SSC, 50 mM NaH2PO4, pH 6.8, 0.5% SDS, 0.1 mg/mL sonicated salmon sperm DNA, and 5X Denhart's solution at 42°C overnight; washing with 2X SSC, 0.1% SDS at 45°G; and washing with 0.2X SSC, 0.1% SDS at 45°C.
In sequence comparison analyses, an ORF, or motif, or set of motifs in a bacteriophage sequence can be compared to known inhibitor sequences, e.g., homologous sequences encoding homologous inhibitors of bacterial function. Likewise, the analysis can include comparison with the structure of essential bacterial gene products, as structural similarities can be indicative of similar or replacement biological function. Such analysis can include the identification of a signature, or characteristic motif(s) of an inhibitor or inhibitor class.
Also, the identification of structural motifs in an encoded product, based on nucleotide or amino acid sequence analysis, can be used to infer a biochemical function for the product. A database containing identified structural motifs in a large number of sequences is available for identification of motifs in phage sequences. The database is PROSITE, which is available at www.expasy.ch/cgi~bin scanprosite. The identification of motifs can, for example, include the identification of signature motifs for a class or classes of inhibitory proteins. Other such databases may also be used. In aspects and preferred embodiments described herein, in which a bacterium or host bacterium is specified, the bacterium or host bacterium is preferably selected from a pathogenic bacterial species, for example, one selected from Table 1. Preferably, an animal or plant pathogen is used. For animals, preferably the bacterium is a bird or mammalian pathogen, still more preferably a human pathogen. In aspects and preferred embodiments involving a bacteriophage or sequences from a bacteriophage, one or more bacteriophage are preferably selected from those listed in Table 1. Those exemplary bacteriophge are readily obtained from the indicated sources.
In some cases, it is advantageous to utilize phage with non-pathogenic host bacteria. The genome, structural motif, ORF, homolog, and other analyses described herein can be performed on such phage and bacteria. Such analysis provides useful information and compositions. The results of such analyses can also be utilized in aspects of the present invention to identify homologous ORFs, especially inhibitor ORFs in phage with pathogenic bacterial hosts. Similarly, identification of a target in a non-pathogenic host can be used to identify homologous sequences and targets in pathogenic bacteria, especially in genetically closely related bacteria. Those skilled in the art are familiar with bacterial genetic relationships and with how to determine relatedness based on levels of genomic identity or other measures of nucleotide sequence and/or amino acid sequence similarity, and/or other physical and culture characteristics such as morphology, nutritional requirements, or minimal media-to support growth.
Also in preferred embodiments, an embodiments of this aspect is combined with an embodiment of the following aspect.
A related aspect of the invention provides methods for identifying a target for antibacterial agents by identifying the bacterial target(s) of at least one uncharacterized or untargeted inhibitor protein or RNA from a bacteriophage. Such identification allows the development of antibacterial agents active on such targets. Preferred embodiments for identifying such targets involve the identification of binding of target and phage ORF products to one another. The phage ORF products may be subportions of a larger ORF product that also binds the host target. In preferred embodiments, the phage protein or RNA is from an uncharacterized bacteriophage in Table 1. This aspect preferably includes the identification of a plurality of such targets in one or a plurality of different bacteria, preferably in one or a plurality of bacteria listed in Table 1.
In preferred embodiments of this aspect and other aspects of this invention involving particular phage ORFs or phage sequences, the ORF is Staphylococcus aureus phage 77 ORF 17, 19, 43, 102, 104, or 182 as identified in U.S. application 09/407,804, S. aureus phage 44AHJD ORF 1, 9, or 12, Streptococcus pneumoniae phage Dp-1 ORF 001, 002, 004, 008, 010, 013, 016, 021, 029, 030, 038, or 041, or Enterococcus sp. phage 182 ORF 002, 008, or 014. As indicated for the above aspect, preferably the method involves the use of a plurality of different phage, and thus a plurality of different phage inhibitors and/or inhibitor ORFs.
In addition to uncharacteized phage ORF products, it is also useful to identify the targets of phage ORF products which are known to be inhibitors of host bacteria, but where the target has not been identified. Thus, such inhibitors can likewise be utilized as "untargeted" inhibitor phage ORFs and ORF products, e.g., proteins or RNAs.
In the context of inhibitor proteins or RNAs from a phage, the term "uncharacterized" means that a bacteria-inhibiting function for the protein has not previously been identified. Preferably, but not necessarily, the sequence of the protein or the corresponding coding region or ORF was not described in the art before the filing of the present application for patent (or alternatively prior to the present invention). Thus, this term specifically excludes any bacteria-inhibiting phage protein and its associated bacterial target which has been identified as inhibitory before the present invention or alternatively before the filing of the present application, for, example those identified in Tables 12-14 or otherwise identified herein. For example, from E. coli, phage T7 genes 0.7 and 2.0 target the host RNA polymerase, phage T4
gp55/gp33 alter the specificity of host RNA polymerase. The T4 regB gene product also targets the host translation apparatus. As with the uncharacterized bacteriophage ORFs or bacteriophage above, for such identified proteins, the sequences encoding those proteins are excluded from the uncharacterized inhibitor proteins. The term "fragment" refers to a portion of a larger molecule or assembly. For proteins, the term "fragment" refers to a molecule which includes at least 5 contiguous amino acids from the reference polypeptide or protein, preferably at least 8, 10, 12, 15, 20, 30, 50 or more contiguous amino acids. In connection with oligo- or polynucleotides, the term "fragment" refers to a molecule which includes at least 15 contiguous nucleotides from a reference polynucleotide, preferably at least 24, 30, 36, 45, 60, 90, 150, or more contiguous nucleotides.
Preferred embodiments involve identification of binding that include methods for distinguishing bound molecules, for example, affinity chromatography, immunoprecipitation, crosslinking, and/or genetic screen methods that permit proteimprotein interactions to be monitored. One of skill in the art is familiar with these techniques and common materials utilized (see, e.g., Coligan, J. et al. (eds.) (1995) Current Protocols in Protein Science. John Wiley & Sons, Secaucus, N.J.).
Genetic screening for the identification of proteimprotein interactions typically involves the co-introduction of both a chimeric bait nucleic acid sequence (here, the phage ORF to be tested) and a chimeric target nucleic acid sequence that, when co- expressed and having affinity for one another in a host cell, stimulate reporter gene expression to indicate the relationship. A "positive" can thus suggest a potential inhibitory effect in bacteria. This is discussed in further detail in the Detailed Description section below. In this way, new bacterial targets can be identified that are inhibited by specific phage ORF products or derivatives, fragments, mimetics, or other molecules.
Other embodiments involve the identification and/or utilization of mutant targets by virtue of their host's relatively unresponsive nature in the presence of expression of ORFs previously identified as inhibitory to the non-mutant or wild-type strain. Such mutants have the effect of protecting the host from an inhibition that would otherwise occur and indirectly allow identification of the precise responsible target for follow-up studies and anti-microbial development. In certain embodiments, rescue from inhibition occurs under conditions in which a bacterial target or mutant target is highly expressed. This is performed, for example, through coupling of the sequence with regulatory element promoters, e.g., as known in the art, which regulate expression at levels higher than wild-type, e.g., at a level sufficiently higher that the
inhibitor can be competitively bound to the highly expressed target such that the bacterium is detectably less inhibited.
Identification of the bacterial target can involve identification of a phage- specific site of action. This can involve a newly identified target, or a target where the phage site of action differs from the site of action of a previously known antibacterial agent or inhibitor. For example, phage T7 genes 0.7 and 2.0 target the host RNA polymerase, which is also the cellular target for the antibacterial agent, rifampin. To the extent that a phage product is found to act at a different site than previously described inhibitors, aspects of the present invention can utilize those new, phage- specific sites for identification and use of new agents. The site of action can be identified by techniques well-known to those skilled in the art, for example, by mutational analysis, binding competition analysis, and/or other appropriate techniques.
Once a bacterial host target protein or nucleic acid or mutant target sequence has been identified and/or isolated, it too can be conveniently sequenced, sequence analyzed (e.g., by computer), and the underlying gene(s), and corresponding translated product(s) further characterized. Preferred embodiments include such analysis and identification. Preferably such a target has not previously been identified as an appropriate target for antibacterial action. Certain embodiments include the identification of at least one inhibitory phage
ORF or ORF product, e.g., as described for the above aspect, and thus are a combination of the two aspects.
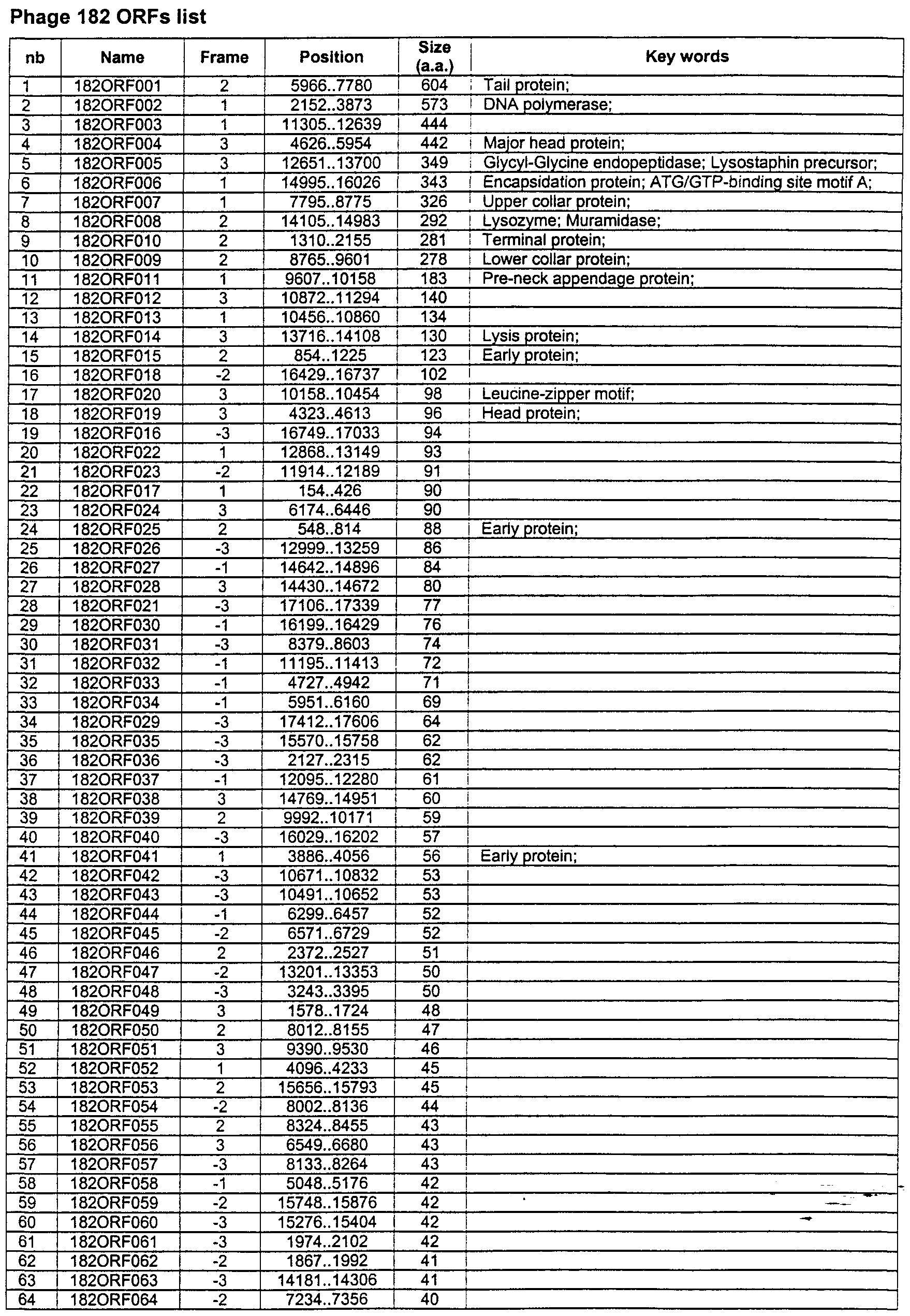
Additionally, the invention provides methods for identifying targets for antibacterial agents by identifying homologs of a bacterial target e.g., S. aureus, Enterococcus faecalis or other Enterococci, and Streptococcus pneumoniae of a bacteriophage inhibitory ORF product. Such homologs may be utilized in the various aspects and embodiments described herein as describded for the host Enterococcus sp. for bacteriophage 182.
Other aspects of the invention provide isolated, purified, or enriched specific phage nucleic acid and amino acid sequences, subsequences, and homologs thereof for phage selected from uncharacterized phage listed in Table 1, preferably from bacteriophage 77, 3 A, 96, 44AHJD (Staphylococcus aureus host bacterium), Dp-1 (Streptococcus pneumoniae host), or 182 (Enterococcus host) or other phage listed in Table 1 for those bacteria. For example, such sequences do not include sequences identified in any of Tables 11-14. Nucleotide sequences of this aspect are at least 15 nucleotides in length, preferably at least 18, 21, 24, or 27 nucleotides in length, more preferably at least 30, 50, or 90 nucleotides in length. In certain embodiments, longer
nucleic acids are preferred, for example those of at least 120, 150, 200, 300, 600, 900 or more nucleotides. Such sequences can, for example, be amplification oligonucleotides (e.g., PCR primers), oligonucleotide probes, sequences encoding a portion or all of a phage-encoded protein, or a fragment or all of a phage-encoded protein. In preferred embodiments, the nucleic acid sequence contains a sequence which is within a length range with a lower length as specified above, and an upper length limit which is no more than 50, 60, 70, 80, or 90%> of the length of the corresponding full-length ORF. The upper length limit can also be expressed in terms of the number of base pairs of the ORF (coding region). In preferred embodiments, the nucleic acid sequence is from Staphylococcus aureus phage 77 ORF 17, 19, 43, 102, 104, or 182 as identified in U.S. application 09/407,804, S. aureus phage 44 AHJD ORF 1, 9, or 12, Streptococcus pneumoniae phage Dp-1 ORF 001, 002, 004, 008, 010, 013, 016, 021, 029, 030, 038, or 041, or Enterococcus sp. phage 182 ORF 002, 008, or 014. As it is recognized that alternate codons will encode the same amino acid for most amino acids due to the degeneracy of the genetic code, the sequences of this aspect includes nucleic acid sequences utilizing such alternate codon usage for one or more codons of a coding sequence. For example, all four nucleic acid sequences GCT, GCC, GCA, and GCG encode the amino acid, alanine. Therefore, if for an amino acid there exists an average of three codons, a polypeptide of 100 amino acids in length will, on average, be encoded by 3100 , or 5 x 1047 , nucleic acid sequences. Thus, a nucleic acid sequence can be modified (e.g., a nucleic acid sequence from a phage as specified above) to form a second nucleic acid sequence encoding the same polypeptide as encoded by the first nucleic acid sequence using routine procedures and without undue experimentation. Thus, all possible nucleic acid sequences that encode the specified amino acid sequences are also fully described herein, as if all were written out in full, taking into account the codon usage, especially that preferred in the host bacterium. The alternate codon descriptions are available in common texbooks, for example, Stryer, BIOCHEMISTRY 3rd ed., and Lehninger, BIOCHEMISTRY 3rd ed., along wth many others. Codon preference tables for various types of organisms are available in the literature. Sequences with alternate codons at one or more sites can also be utilized in the computer-related aspects and embodiments herein. Because of the number of sequence variations involving alternate codon usage, for the sake of brevity, individual sequences are not separately listed herein. Instead the alternate sequences are described by reference to the natural sequence with replacement of one or more (up to all e.g., up to 3, 5, 10, 15, 20, 30, 40, 50, or more) of the degenerate codons with alternate codons from the alternate codon
table (Table 6), or a modified table applicable to a particular organism that has differing codon usage, preferably with selection according to preferred codon usage for the normal host organism or a host organism in which a sequence is intended to be expressed. Those skilled in the art also understand how to alter the alternate codons to be used for expression in organisms where certain codons code differently than shown in the "universal" codon table.
For amino acid sequences or polypeptides, sequences contain at least 5 peptide- linked amino acid residues, and preferably at least 6, 7, 10, 15, 20, 30, or 40, amino acids having identical amino acid sequence as the same number of contiguous amino acid residues in a particular phage ORF product. In some cases longer sequences may be preferred, for example, those of at least 50, 60, 70, 80, or 100 amino acids in length. In preferred embodiments, the amino acid sequence contains a sequence which is within a length range with a lower length as specified above, and an upper length limit which is no more than 50, 60, 70, 80, or 90%> of the length of the corresponding full-length ORF product. The upper length limit can also be expressed in terms of the number of amino acid residues of the ORF product. In preferred embodiments, the amino acid sequence or polypeptide has bacteria-inhibiting function when expressed or otherwise present in a bacterial cell which is a host for the bacteriophage from which the sequence was derived. By "isolated" in reference to a nucleic acid is meant that a naturally occurring sequence has been removed from its normal cellular (e.g., chromosomal) environment or is synthesized in a non-natural environment (e.g., artificially synthesized). Thus, the sequence may be in a cell-free solution or placed in a different cellular environment. The term does not imply that the sequence is the only nucleotide chain present, but that it is essentially free (about 90-95% pure at least) of non-nucleotide material naturally associated with it, and thus is distinguished from isolated chromosomes.
The term "enriched" means that the specific DNA or RNA sequence constitutes a significantly higher fraction (2-5 fold) of the total DNA or RNA present in the cells or solution of interest than in normal or diseased cells or in cells from which the sequence was originally taken. This could be caused by a person by preferential reduction in the amount of other DNA or RNA present, or by a preferential increase in the amount of the specific DNA or RNA sequence, or by a combination of the two. However, it should be noted that enriched does not imply that there are no other DNA or RNA sequences present, just that the relative amount of the sequence of interest has been significantly increased.
The term "significant" is used to indicate that the level of increase is useful to the person making such an increase and an increase relative to other nucleic acids of about at least 2-fold, more preferably at least 5- to 10-fold or even more. The term also does not imply that there is no DNA or RNA from other sources. The other source DNA may, for example, comprise DNA from a yeast or bacterial genome, or a cloning vector such as pUC19. This term distinguishes from naturally occurring events, such as viral infection, or tumor type growths, in which the level of one mRNA may be naturally increased relative to other species of mRNA. That is, the term is meant to cover only those situations in which a person has intervened to elevate the proportion of the desired nucleic acid.
It is also advantageous for some purposes that a nucleotide sequence be in purified form. The term "purified" in reference to nucleic acid does not require absolute purity (such as a homogeneous preparation). Instead, it represents an indication that the sequence is relatively more pure than in the natural environment (compared to the natural level, this level should be at least 2-5 fold greater, e.g., in terms of mg/mL). Individual clones isolated from a cDNA library may be purified to electrophoretic homogeneity. The claimed DNA molecules obtained from these clones could be obtained directly from total DNA or from total RNA. The cDNA clones are not naturally occurring, but rather are preferably obtained via manipulation of a partially purified naturally occurring substance (messenger RNA). The construction of a cDNA library from mRNA involves the creation of a synthetic substance (cDNA) and pure individual cDNA clones can be isolated from the synthetic library by clonal selection of the cells carrying the cDNA library. Thus, the process which includes the construction of a cDNA library from mRNA and isolation of distinct cDNA clones yields an approximately 106-fold purification of the native message. Thus, purification of at least one order of magnitude, preferably two or three orders, and more preferably four or five orders of magnitude is expressly contemplated.
The terms "isolated", "enriched", and "purified" as respect nucleic acids, above, may similarly be used to denote the relative purity and abundance of polypeptides ( multimers of amino acids joined one to another by -carboxyl:α-amino group (peptide) bonds). These, too, may be stored in, grown in, screened in, and selected from libraries using biochemical techniques familiar in the art. Such polypeptides may be natural, synthetic or chimeric and may be extracted using any of a variety of methods, such as antibody immunoprecipitation, other "tagging" - techniques, conventional chromatography and/or electrophoretic methods. Some of the above utilize the corresponding nucleic acid sequence.
As indicated above, aspects and embodiments of the invention are not limited to entire genes and proteins. The invention also provides and utilizes fragments and portions thereof, preferably those which are "active" in the inhibitory sense described above. Such peptides or oligopeptides and oligo or polynucleotides have preferred lengths as specified above for nucleic acid and amino acid sequences from phage; corresponding recombinant constructs can be made to express the encoded same.
Also included are homologous sequences and fragments thereof.
Nucleic acid sequences of the present invention can be isolated using a method similar to those described herein or other methods known to those skilled in the art. In addition, such nucleic acid sequences can be chemically synthesized by well- known methods. Also, by having particular phage ORFs, e.g., the phage ORFs identified herein (e.g., anti-bacterial ORFs of the present invention, portions thereof, or oligonucleotides derived therefrom as described), other antimicrobial sequences from other bacteriophage sources can be identified and isolated using methods described here or other methods, including methods utilizing nucleic acid hybridization and/or computer-based sequence alignment methods.
The invention also provides bacteriophage antimicrobial DNA segments from other phages based on nucleic acids and sequences hybridizing to the presently identified inhibitory ORF under high stringency conditions or sequences that are highly homologous. The bacteriophage segment from a specific phage, e.g., an antimicrobial DNA segment, can be used to identify a related segment from another unrelated phage based on stringent conditions of hybridization or on being a homolog based on nucleic acid and/or amino acid sequence comparisons. As with identified inhibitory sequences, such homologous coding sequences and products can be used as antimicrobials, to construct active portions or derivatives, to construct peptidomimetics, and to identify bacterial targets.
The nucleotide and amino acid sequences identified herein are believed to be correct, however, certain sequences may contain a small percentage of errors, e.g., 1- 5%. In the event that any of the sequences have errors, the corrected sequences can be readily provided by one skilled in the art using routine methods. For example, the nucleotide sequences can be confirmed or corrected by obtaining and culturing the relevant phage, and purifying phage genomic nucleic acids. A region or regions of - interest can be amplified, e.g., by PCR from the appropriate genomic template, using primers based on the described sequence. The amplified regions can then be sequenced using any of the available methods (e.g., a dideoxy termination method).
This can be done redundantly to provide the corrected sequence or to confirm that the described sequence is correct. Alternatively, a particular sequence or sequences can be identified and isolated as an insert or inserts in a phage genomic library and isolated, amplified, and sequenced by standard methods. Confirmation or correction of a nucleotide sequence for a phage gene provides an amino acid sequence of the encoded product by merely reading off the amino acid sequence according to the normal codon relationships and/or expressed in a standard expression system and the polypeptide product sequenced by standard techniques. The sequences described herein thus provide unique identification of the corresponding genes, coding sequences, and other sequences, allowing those sequences to be used in the various aspects of the present invention.
In other aspects, the invention provides recombinant vectors and cells harboring at least one of the phage ORFs or portion thereof, or bacterial target sequences described herein. As understood by those skilled in the art, vectors may be provided in different forms, including, for example, plasmids, cosmids, and virus- based vectors. See, e.g.. Maniatis. T. et al. ( 1989-) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor University Press, Cold Spring, N.Y.; See also, Ausubel, F.M. et al. (eds.) (1994) Current Protocols in Molecular Biology. John Wiley & Sons, Secaucus, NJ. In preferred embodiments, the vectors will be expression vectors, preferably shuttle vectors that permit cloning, replication, and expression within bacteria. An "expression vector" is one having regulatory nucleotide sequences containing transcriptional and translational regulatory information that controls expression of the nucleotide sequence in a host cell. Preferably the vector is constructed to allow amplification from vector sequences flanking an insert locus. In certain embodiments, the expression vectors may additionally or alternativley support expression, and/or replication in animal, plant and/or yeast cells due to the presence of suitable regulatory sequences, e.g., promoters, enhancers, 3' stabilizing sequences, primer sequences, etc. In preferred embodiments, the promoters are inducible and specific for the system in which expression is desired, e.g., bacteria, animal, plant, or yeast. The vectors may optionally encode a "tag" sequence or sequences to facilitate protein purification. Convenient restriction enzyme cloning sites and suitable selective marker(s) are also optionally included. Such selective markers can be, for example, antibiotic resistance markers or markers which supply an essential nutritive growth factor to an otherwise deficient mutant host, e.g., tryptophan, histidine, or leucjng"tn the Yeast Two-Hybrid systems described below.
The term "recombinant vector" relates to a single- or double-stranded circular nucleic acid molecule that can be transfected into cells and replicated within or independently of a cell genome. A circular double-stranded nucleic acid molecule can be cut and thereby linearized upon treatment with appropriate restriction enzymes. An assortment of nucleic acid vectors, restriction enzymes, and the knowledge of the nucleotide sequences cut by restriction enzymes are readily available to those skilled in the art. A nucleic acid molecule encoding a desired product can be inserted into a vector by cutting the vector with restriction enzymes and ligating the two pieces together. Preferably the vector is an expression vector, e.g., a shuttle expression vector as described above.
By " recombinant cell" is meant a cell possessing introduced or engineered nucleic acid sequences, e.g., as described above. The sequence may be in the form of or part of a vector or may be integrated into the host cell genome. Preferably the cell is a bacterial cell. In another aspect, the invention also provides methods for identifying and/or screening compounds "active on" at least one bacterial target of a bacteriophage inhibitor protein or RNA. Preferred embodiments involve contacting such a bacterial target or targets (e.g., bacterial target proteins) with a test compound, and determining whether the compound binds to or reduces the level of activity of the bacterial target (e.g., a bacterial target protein). Preferably this is done either in vivo (i.e., in a cell- based assay) or in vitro, e.g., in a cell-free system under approximately physiological conditions.
The compounds that can be used may be large or small, synthetic or natural, organic or inorganic, proteinaceous or non-pro teinaceous. In preferred embodiments, the compound is a peptidomimetic, as described herein, a bacteriophage inhibitor protein or fragment or derivative thereof, preferably an "active portion", or a small molecule.
In preferred embodiments, the bacterial target is a target of a phage ORF identified herein, e.g., S. aureus phage 44AHJD ORF 1, 9, or 12, Streptococcus pneumoniae p age Dp-1 ORF 001, 002, 004, 008, 010, 013, 016, 021, 029, 030, 038, or 041, or Enterococcus sp. phage 182 ORF 002, 008, or 014.
In particular embodiments, the methods include the identification of bacterial targets or the site of action of an inhibitor on a bacterial target as described above or otherwise described herein. In embodiments involving binding assays, preferably binding is to a fragment or portion of a bacterial target protein, where the fragment includes less than 90%, 80%, 70%, 60%, 50%, 40%, or 30% of an intact bacterial target protein. Preferably,
the at least one bacterial target includes a plurality of different targets of bacteriophage inhibitor proteins, preferably a plurality of different targets. The plurality of targets can be in or from a plurality of different bacteria, but preferably is from a single bacterial species. A "method of screening" refers to a method for evaluating a relevant activity or property of a large plurality of compounds (e.g., a bacteria-inhibiting activity), rather than just one or a few compounds. For example, a method of screening can be used to conveniently test at least 100, more preferably at least 1000, still more preferably at least 10,000, and most preferably at least 100,000 different compounds, or even more.
In the context of this invention, the term "small molecule" refers to compounds having molecular mass of less than 2000 Daltons, preferably less than 1500, still more preferably less than 1000, and most preferably less than 600 Daltons. Preferably but not necessarily, a small molecule is not an oligopeptide. In a related aspect or in preferred embodiments, the invention provides a method of screening for potential antibacterial agents by determining whether any of a plurality of compounds, preferably a plurality of small molecules, is active on at least one target of a bacteriophage inhibitor protein or RNA. Preferred embodiments include those described for the above aspect, including embodiments which involve determining whether one or more test compounds bind to or reduce the level of activity of a bacterial target, and embodiments which utilize a plurality of different targets as described above.
The identification of bacteria-inhibiting phage ORFs and their encoded products also provides a method for identifying an active portion of such an encoded product. This also provides a method for identifying a potential antibacterial agent by identifying such an active portion of a phage ORF or ORF product. In preferred embodiments, the identification of an active portion involves one or more of mutational analysis, deletion analysis, or analysis of fragments of such products. The method can also include determination of a 3-dimensional structure of an active portion, such as by analysis of crystal diffraction patterns. In further embodiments, the method involves constructing or synthesizing a peptidomimetic compound, where the structure of the peptidomimetic compound corresponds to the structure of the active portion. In this context, "corresponds" means that the peptidomimetic compound structure has sufficient similarities to the structure of the active portion that the peptidomimetic will interact with the same molecule as the phage protein and preferably will elicit at least one cellular response in common which relates to the inhibition of the cell by the phage protein.
~>l
In preferred embodiments, the ORF or ORF product is or is derived or obtained from S. aureus phage 44AHJD ORF 1, 9, or 12, Streptococcus pneumoniae phage Dp-1 ORF 001, 002, 004, 008, 010, 013, 016, 021, 029, 030, 038, or 041, or Enterococcus sp. phage 182 ORF 002, 008, or 014 or product thereof. The methods for identifying or screening for compounds or agents active on a bacterial target of a phage-encoded inhibitor can also involve identification of a phage-specific site of action on the target.
Preferably in the methods for identifying or screening for compounds active on such a bacterial target, the target is uncharacterized; the target is from an uncharacterized bacterium from Table 1 ; the site of action is a phage-specfic site of action.
Further embodiments include the identification of inhibitor phage ORFs and bacterial targets as in aspects above.
An "active portion" as used herein denotes an epitope, a catalytic or regulatory domain, or a fragment of a bacteriophage inhibitor protein that is responsible for, or a significant factor in, bacterial target inhibition. The active portion preferably may be removed from its contiguous sequences and, in isolation, still effect inhibition.
By "mimetic" is meant a compound structurally and functionally related to a reference compound that can be natural, synthetic, or chimeric. In terms of the present invention, a "peptidomimetic," for example, is a compound that mimics the activity- related aspects of the 3-dimensional structure of a peptide or polyeptide in a non- peptide compound, for example mimics the structure of a peptide or active portion of a phage- or bacterial ORF-encoded polypeptide.
A related aspect provides a method for inhibiting a bacterial cell by contacting the bacterial cell with a compound active on a bacterial target of a bacteriophage inhibitor protein or RNA, where the target was uncharacterized. In preferred embodiments, the compound is such a protein, or a fragment or derivative thereof; a structural mimetic, e.g., a peptidomimetic, of such a protein or fragment; a small molecule; the contacting is performed in vitro, the contacting is performed in vivo in an infected or at risk organism, e.g., an animal such as a mammal or bird, for example, a human, or other mammal described herein; the bacterium is selected from a genus and or species listed in Table 1 ; the bacteriophage inhibitor protein is uncharacterized; the bacteriophage inhibitor protein is from an uncharacterized phage listed in Table 1 ; the phage inhibitor protein is from one of S. aureus phage 44AHJD ORF 1, 9, or 12, Streptococcus pneumoniae phage Dp-1 ORF 001, 002, 004, 008, 010, 013, 016^02ϊ 029, 030, 038, or 041, ox Enterococcus sp. phage 182 ORF 002, 008, or 014.
In the context of targets in this invention, the term "uncharacterized" means that the target was not recognized as an appropriate target for an antibacterial agent prior to the filing of the present application or alternatively prior to the present invention. Such lack of recognition can include, for example, situations where the target and/or a nucleotide sequence encoding the target were unknown, situations where the target was known, but where it had not been identified as an appropriate target or as an essential cellular component, and situations where the target was known as essential but had not been recognized as an appropriate target due to a belief that the target would be inaccessible or otherwise that contacting the cell with a compound active on the target in vitro would be ineffective in cellular inhibition, or ineffective in treatment of an infection. Methods described herein utilizing bacterial targets, e.g., for inhibiting bacteria or treating bacterial infections, can also utilize "uncharacterized target sites", meaning that the target has been previously recognized as an appropriate target for an antibacterial agent, but where an agent or inhibitor of the invention is used which acts at a different site than that at which the previously utilized antibacterial agent, i.e., a phage-specific site. Preferably the phage-specific site has different functional characteristics from the previously utilized site. In the context of targets or target sites, the term "phage-specific" indicates that the target or site is utilized by at least one bacteriophage as an inhibitory target and is different from previously identified targets or target sites.
In the context of this invention, the term "bacteriophage inhibitor protein" refers to a protein encoded by a bacteriophage nucleic acid sequence which inhibits bacterial function in a host bacterium. Thus, it is a bacteria-inhibiting phage product. In the context of this invention, the phrase "contacting the bacterial cell with a compound active on a bacterial target of a bacteriophage inhibitor protein" or equivalent phrases refer to contacting with an isolated, purified, or enriched compound or a composition including such a compound, but specifically does not rely on contacting the bacterial cell with an intact phage which encodes the compound. Preferably no intact phage are involved in the contacting. Related aspects provide methods for prophylactic or therapeutic treatment of a bacterial infection by administering to an infected, challenged or at risk organism a therapeutically or prophylactically effective amount of a compound active on a target of a bacteriophage inhibitor protein or RNA, or as described for the previous aspect. Preferably the bacterium involved in the infection or risk of infection produces the identified target of the bacteriophage inhibitor protein or alternatively produces-a homologous target compound. In preferred embodiments, the host organism is a plant or animal, preferably a mammal or bird, and more preferably, a human or other
mammal described herein. Preferred embodiments include, without limitation, those as described for the preceding aspect.
Compounds useful for the methods of inhibiting, methods of treating, and pharmaceutical compositions can include novel compounds, but can also include compounds which had previously been identified for a purpose other than inhibition of bacteria. Such compounds can be utilized as described and can be included in pharmaceutical compositions.
In preferred embodiments of this and other aspects of the invention utilizing bacterial target sequences of a bacteriiophage inhibitory ORF product, the target sequence is encoded by a Staphylococcus nucleic acid coding sequence, preferably S. aureus, a Streptococcus nucleic acid coding sequence, preferably Streptococcus pneumoniae, or Enterococcus nucleic acid coding sequence. Possible target sequences are described herein by reference to sequence source sites.
The amino acid sequence of a polypeptide target is readily provided by translating the corresponding coding region. For the sake of brevity, the sequences are not reproduced herein. For the sake of brevity, the sequences are described by reference to the GenBank entries instead of being written out in full herein. In cases where the TIGR or GenBank entry for a coding region is not complete, the complete sequence can be readily obtained by routine methods, e.g., by isolating a clone in a phage host genomic library, and sequencing the clone insert to provide the relevant coding region. The boundaries of the coding region can be identified by conventional sequence analysis and/or by expression in a bacterium in which the endogenous copy of the coding region has been inactivated and using subcloning to identify the functional start and stop codons for the coding region. In the context of nucleic acid or amino acid sequences of this invention, the term "corresponding" indicates that the sequence is at least 95% identical, preferably at least 97% identical, and more preferably at least 99%> identical to a sequence from the specified phage genome, a ribonucleotide equivalent, a degenerate equivalent (utilizing one or more degenerate codons), or a homologous sequence, where the homolog provides functionally equivalent biological function.
By "treatment" or "treating" is meant administering a compound or pharmaceutical composition for prophylactic and/or therapeutic purposes. The term "prophylactic treatment" refers to treating a patient or animal that is not yet infected but is susceptible to or otherwise at risk of a bacterial infection. The term "therapeutic treatment" refers to administering treatment to a patient already suffering from, infection.
The term "bacterial infection" refers to the invasion of the host organism, animal or plant, by pathogenic bacteria. This includes the excessive growth of bacteria which are normally present in or on the body of the organism, but more generally, a bacterial infection can be any situation in which the presence of a bacterial population(s) is damaging to a host organism. Thus, for example, an organism suffers from a bacterial population when excessive numbers of a bacterial population are present in or on the organism's body, or when the effects of the presence of a bacterial population(s) is damaging to the cells, tissue, or organs of the organism.
The terms "administer", "administering", and "administration" refer to a method of giving a dosage of a compound or composition, e.g., an antibacterial pharmaceutical composition, to an organism. Where the organism is a mammal, the method is, e.g., topical, oral, intravenous, transdermal, mtraperitoneal, intramuscular, or intrathecal. The preferred method of administration can vary depending on various factors, e.g., the components of the pharmaceutical composition, the site of the potential or actual bacterial infection, the bacterium involved, and the infection severity.
The term "mammal" has its usual biological meaning referring to any organism of the Class Mammalia of higher vertebrates that nourish their young with milk secreted by mammary glands, e.g., mouse, rat, and, in particular, human, bovine, sheep, swine, dog, and cat.
In the context of treating a bacterial infection a "therapeutically effective amount" or "pharmaceutically effective amount" indicates an amount of an antibacterial agent, e.g., as disclosed for this invention, which has a therapeutic effect. This generally refers to the inhibition, to some extent, of the normal cellular functioning of bacterial cells that renders or contributes to bacterial infection. The dose of antibacterial agent that is useful as a treatment is a "therapeutically effective amount." Thus, as used herein, a therapeutically effective amount means an amount of an antibacterial agent that produces the desired therapeutic effect as judged by clinical trial results and/or animal models. This amount can be routinely determined by one skilled in the art and will vary depending on several factors, such as the particular bacterial strain involved and the particular antibacterial agent used.
In connection with claims to methods of inhibiting bacteria and therapeutic or prophylactic treatments, "a compound active on a target of a bacteriophage inhibitor protein" or terms of equivalent meaning differ from administration of or contactwTth an intact phage naturally encoding the full-length inhibitor compound. While an intact phage may conceivably be incoφorated in the present methods, the method at
least includes the use of an active compound as specified different from a full length inhibitor protein naturally encoded by a bacteriophage and/or a delivery or contacting method different from administration of or contact with an intact phage encoding the full-length protein. Similarly, pharmaceutical compositions described herein at least include an active compound different from a full-length inhibitor protein naturally encoded by a bacteriophage or such a full-length protein is provided in the composition in a form different from being encoded by an intact phage. Preferably the methods and compositions do not include an intact phage.
In accord with the above aspects, the invention also provides antibacterial agents and compounds active on bacterial targets of bacteriophage inhibitor proteins or RNAs, where the target was uncharacterized as indicated above. As previously indicated, such active compounds include both novel compounds and compounds which had previously been identified for a purpose other than inhibition of bacteria. Such previously identified biologically active compounds can be used in embodiments of the above methods of inhibiting and treating. In preferred embodiments, the targets, bacteriophage, and active compound are as described herein for methods of inhibiting and methods of treating. Preferably the agent or compound is formulated in a pharmaceutical composition which includes a pharmaceutically acceptable carrier, excipient, or diluent. In addition, the invention provides agents, compounds, and pharmaceutical compositions where an active compound is active on an uncharacterized phage-specific site.
In preferred embodiments, the target is as described for embodiments of aspects above.
Likewise, the invention provides a method of making an antibacterial agent. The method involves identifying a target of a bacteriophage inhibitor polypeptide or protein or RNA, screening a plurality of compounds to identify a compound active on the target, and synthesizing the compound in an amount sufficient to provide a therapeutic effect when administered to an organism infected by a bacterium naturally producing the target. In preferred embodiments, the identification of the target and identification of active compounds include steps or methods and/or components as described above (or otherwise herein) for such identification. Likewise, the active compound can be as described above, including fragments and derivatives of phage inhibitor proteins, peptidomimetics, and small molecules. As recognized by those skilled in the art, peptides can be synthesized by expression systems and purified, or can be synthesized artificially. In preferred embodiments the inhibitory phage ORF- products is from S. aureus phage 44AH D ORF 1, 9, or 12, Streptococcus
pneumoniae phage Dp-1 ORF 001, 002, 004, 008, 010, 013, 016, 021, 029, 030, 038, or 041, or Enterococcus sp. phage 182 ORF 002, 008, or 014.
As indicated above, sequence analysis of nucleotide and/or amino acid sequences can beneficially utilize computer analysis. Thus, in additional aspects the invention provides computer-related hardware and media and methods utilizing and incorporating sequence data from uncharacterized phage, e.g., uncharacterized phage listed in Table 1, preferably at least one of Staphylococcus aureus phage S. aureus phage 44AHJD ORF 1, 9, or 12, Streptococcus pneumoniae phage Dp-1 ORF 001, 002, 004, 008, 010, 013, 016, 021, 029, 030, 038, or 041, or Enterococcus sp. phage 182 ORF 002, 008, or 014, or 44 AHJD, Enterococcus sp. phage 182, or
Streptococcus pneumoniae phage Dp-1. In general, such aspects can facilitate the above-described aspects. Various embodiments involve the analysis of genetic sequence and encoded products, as applied to the evaluating bacteriophage inhibitor ORFs and compounds and fragments related thereto. The various sequence analyses, as well as function analyses, can be used separately or in combination, as well as in preceding aspects and embodiments. Use in combination is often advantageous as the additional information allows more efficient prioritizing of phage ORFs for identification of those ORFs that provide bacteria-inhibiting function.
In one aspect, the invention provides a computer-readable device which includes at least one recorded amino acid or nucleotide sequence corresponding to one of the specified phage and a sequence analysis program for analyzing a nucleotide and/or amino acid sequence. The device is arranged such that the sequence information can be retrieved and analyzed using the analysis program. The analysis can identify, for example, homologous sequences or the indicated %s of the phage genome and structural motifs. Preferably the sequence includes at least 1 phage ORF or encoded product, more preferably at least 10%, 20%, 30%, 40%, 50%, 70%, 90%, or 100%o of the genomic phage ORFs and/or equivalent cDNA, RNA, or amino acid sequences. Preferably the sequence or sequences in the device are recorded in a medium such as a floppy disk, a computer hard drive, an optical disk, computer random access memory (RAM), or magnetic tape. The program may also be recorded in such medium. The sequences can also include sequences from a plurality of different phage.
In this context, the term "corresponding" indicates that the sequence is at least 95% identical, preferably at least 97% identical, and more preferably at least 99% identical to a sequence from the specified phage genome, a ribonucleotide equivalent, a degenerate equivalent (utilizing one or more degenerate codons), or a homologous sequence, where the homolog provides functionally equivalent biological function.
Similarly, the invention provides a computer analysis system for identifying biologically important portions of a bacteriophage genome. The system includes a data storage medium, e.g., as identified above, which has recorded thereon a nucleotide sequence corresponding to at least a portion of at least one uncharacterized bacteriophage genome, a set of program instructions to allow searching of the sequence or sequences to analyze the sequence, and an output device where the portion includes at least the sequence length as specified in the preceding aspect. The output device is preferably a printer, a video display, or a recording medium. More one than one output device may be included. For each of the present computer-related asepcts, the bacteriophage are preferably selected from the uncharacterized phage listed in Table 1, more preferably from bacteriophage 77, 3 A, 96, 44 AHJD (S. aureus), Dp-1 (Streptococcus pneumoniae), or 182 (Enterococcus).
In keeping with the computer device aspects, the invention also provides a method for identifying or characterizing a bacteriophage ORF by providing a computer-based system for analyzing nucleotide or amino acid sequences, e.g., as describe above. The system includes a data storage medium which has recorded a sequences or sequences as described for the above devices, a set of instructions as in the preceding aspect, and an output device as in the preceding aspect. The method further involves analyzing at least one sequence, and outputting the analysis results to at least one output device.
In preferred embodiments, the analysis identifies a sequence similarity or homology with a sequence or sequences selected from bacterial ORFs encoding products with related biological function; ORFs encoding known inhibitors; and essential bacterial ORFs. Preferably the analysis identifies a probable biological function based on identification of structural elements or characteristic or signature motifs of an encoded product or on sequence similarity or homology. Preferably the uncharacterized bacteriophage is from Table 1 , more preferably at least one of bacteriophage 77, 3 A, 96, 44 AHJD (S. aureus), Dp-1 (Streptococcus pneumoniae), or 182 (Enterococcus). In preferred embodiments, the method also involves determining at least a portion of the nucleotide sequence of at least one uncharacterized bacteriophage as indicated, and recording that sequence on data storage medium of the computer-based system. In preferred embodiments, the analysis identifies a sequence similarity of homology with a S. aureus phage 44AHJD ORF 1, 9, or 12, Streptococcus pneumoniae phage Dp-1 ORF 001, 002, 004, 008, 010, 013, 016, 021, 029, 030, 038, or 041, or Enterococcus sp. phage 182 ORF 002, 008, or 014. , ~~
As used in the claims to describe the various inventive aspects and embodiments, "comprising" means including, but not limited to, whatever follows the word "comprising". Thus, use of the term "comprising" indicates that the listed elements are required or mandatory, but that other elements are optional and may or may not be present. By "consisting of is meant including, and limited to, whatever follows the phrase "consisting of. Thus, the phrase "consisting of indicates that the listed elements are required or mandatory, and that no other elements may be present. By "consisting essentially of is meant including any elements listed after the phrase, and limited to other elements that do not interfere with or contribute to the activity or action specified in the disclosure for the listed elements. Thus, the phrase "consisting essentially of indicates that the listed elements are required or mandatory, but that other elements are optional and may or may not be present depending upon whether or not they affect the activity or action of the listed elements.
Further embodiments will be apparent from the following Detailed Description and from the claims.
BRIEF DESCRIPTION OF THE DRAWINGS
FIGURE 1 A and IB are flow schematics showing the manipulations used to convert pT0021, an arsenite inducible vector containing the luciferase gene, into pTHA or pTM, two ars inducible vectors. Vector pTHA contains BamH I, Sal I, and Hind III cloning sites and a downstream HA epitope tag. Vector pTM contains Bam HI and Hind III cloning sites and no HA epitope tag.
FIGURE 2 is a schematic representation of the cloning steps involved to place the DNA segments of any of ORFs 17/ 19/ 43/ 102/104/182 or other sequences into pTHA to assess inhibitory potential. For subcloning into pTM or pT0021, Individual ORFs were amplified by the PCR using oligonucleotides targeting the ATG and stop codons of the ORFs. Using this strategy, Bam HI and Hind III sites were positioned immediately upstream or downstream, respectively of the start and stop codons of each ORF. Following digestion with Bam HI and Hind III, the PCR fragments were _ subcloned into the same sites of pT0021 or pTM. Clones were verified by PCPTand direct sequencing.
FIGURE 3 shows a schematic representation of the functional assays used to characterize the bactericidal and bacteriostatic potential of all predicted ORFs (>33 amino acids) encoded by bacteriophage 77. Fig. 3A) Functional assay on semi-solid support media. Fig. 3B) Functional assay in liquid culture.
FIGURE 4A, B, and C is a bar graph showing the results of a screen in liquid media to assess bacteriostatic or bactericidal activity of 93 predicted ORFs (>33 amino acids) encoded by bacteriophage 77. Growth inhibition assays were performed as detailed in the Detailed Description. The relative growth of Staphylococcus aureus transformants harboring a given bacteriophage 77 ORF (identified on the bottom of the graph), in the absence or presence of arsenite, is plotted relative to growth of a Staphylococcus aureus transformant containing ORF 5, a non-toxic bacteriophage 77 ORF (which is set at 100%). Each bar represents the average obtained from three Staph A transformants grown in duplicate. Bacteriophage 77 ORFs showing significant growth inhibition consist of ORFs 17, 19, 102, 104, and 182.
FIGURE 5 shows a block diagram of major components of a general purpose computer.
FIGURE 6 shows an ORF map for Streptococcus pneumoniae bacteriophage Dp-1 showing the ORF identifiers, genomic locations, and orientations of the 85 identified ORFs that were found to have ribosomal binding sites and thus are expected to be expressed.
FIGURE 7 shows a schematic representation of the arsenite-inducible expression system present in a shuttle vector designed to express individual Streptococcus bacteriophage Dp-1 ORFs in Streptococcus. Various modifications can be readily made to such a vector, or other vectors can be readily constructed to provide inducible expression of ORFs in a particular host bacterium using well-known techniques.
DETAILED DESCRIPTION OF THE PREFERRED EMBODIMENTS
The invention may be more clearly understood from the following description. The tables will first be briefly described.
Table 1 is a listing of a large number of available bacteriophage that can be readily obtained and used in the present invention.
Table 2 shows the complete nucleotide sequence of the genome of Staphylococcus aureus bacteriophage 77. Table 3 shows a list of all the ORFs from Bacteriophage 77 that were screened in the functional assay to identify those with anti-microbial activity.
Table 4 shows the predicted nucleotide sequence, predicted amino acid sequence, and physiochemical parameters of ORF 17/ 19/ 43/ 102/ 104/ 182]. These include the primary amino acid sequence of the predicted protein, the average molecular weight, amino acid composition, theoretical pi, hydrophobicity map, and predicted secondary structure map.
Table 5 shows homology search results. BLAST analysis was performed with ORFs 17/ 19/ 43/ 102/ 104/ 182 against NCBI non-redundant nucleotide and Swissprot databases. The results of this search indicate that: I) ORF 17 has no significant homology to any gene in the NCBI non-NCBI non-redundant nucleotide database, II) ORF 19 has significant homology to one gene in the NCBI non- redundant nucleotide database - the gene encoding ORF 59 of bacteriophage phi PVL, III) ORF 43 has significant homology to one gene in the NCBI non-redundant nucleotide database - the gene encoding ORF 39 of phi PVL, IV) ORF 102 has significant homology to one gene in the NCBI non-redundant nucleotide database - the gene encoding ORF 38 of phi PVL, V) ORF 104 has no significant homology to any gene in the NCBI non-redundant nucleotide database, VI) ORF 182 has significant homology to one gene in the NCBI non-redundant nucleotide database - the gene encoding ORF 39 of phi PVL. Table 6 is a table from Alberts et al., MOLECULAR BIOLOGY OF THE
CELL 3rd ed., showing the redundancy of the "universal" genetic code. ~ —
Table 7 shows the complete nucleotide sequence of Staphylococcus aureus bacteriophage 3A.
Table 8 is a listing of the ORFs identified in Staphylococcus aureus bacteriophage 3A.
Table 9 shows the complete nucleotide sequence of Staphylococcus aureus bacteriophage 96. Table 10 is a listing of the ORFs identified in Staphylococcus aureus bacteriophage 96.
Table 11 is a listing of sequences deposited in the NCBI public database (GeneBank) for bacteriophage listed in Table 1.
Table 12 is a listing of phage which encode a known lysis function , including the identified lysis gene.
Table 13 is a listing of bacteriophage which encode holin genes, where holin genes encode proteins which form pores and eventually enable other enzymes to kill the host bacterium.
Table 14 is a listing of bacteriophage which encode kil genes. Table 15 is a list of Staphylococcus aureus sequences identified by accession number which may include sequences from genes coding for target sequences for the phage 77-encoded antimicrobial proteins or peptides. The sequences were obtained by searching GenBank for listings.
Table 16 shows the nucleotide sequence of the genome of Staphylococcus aureus phage 44 AHJD.
Table 17 lists and shows the sequence position of the 73 ORFs predicted to be encoded by Staphylococcus aureus bacteriophage 44 AHJD that are greater than 33 amino acids.
Table 18 shows the ORF sequences and putative amino acid sequences for the Staphylococcus aureus bacteriophage 44AHJD ORFs greater than 33 amino acids.
Table 19 shows the similarities in sequence identified between predicted
Staphylococcus aureus bacteriophage 44 AHJD ORFs and sequences present in public databases.
Table 20 shows the homology alignments between predicted Staphylococcus aureus bacteriophage 44 AHJD ORFs and the corresponding protein sequences present in public sequence databases.
Table 21 shows the complete nucleotide sequence of the genome of
Enterococcus bacteriophage 182. - ~
Table 22 lists and shows the sequence position of the 80 ORFs identified in bacteriophage 182 and that are greater than 33 amino acids.
Table 23 shows the nucleotide and predicted amino acid sequence of all 80 ORFs identified in bacteriophage 182.
Table 24 shows the similarities identified to date in sequence between Enterococcus phage 182 ORFs greater than 33 amino acids and sequences present in public sequence databases.
Table 25 shows the predicted amino acid sequence as well as the predicted secondary structures map for two Enterococcus bacteriophage 182 ORFs.
Table 26 shows the homology alignments between predicted Enterococcus bacteriophage 182 ORFs and the corresponding protein sequences present in public sequence databases.
Table 27 list Enterococcus sequences listed in GenBank providing possible Enterococcal target sequences for inhibitory Enterococcus bacteriophage 182 ORFs and other compounds with antibacterial activity.
Table 28 shows the complete nucleotide sequence of the genome of Streptococcus bacteriophage Dp- 1.
Table 29 lists and shows sequence position of the 273 ORFs identified in Pneumococcal bacteriophage Dp-1 that are greater than 33 amino acids, 85 of which are predicted to be expressed in Dp-1 as having a ribosomal binding site. That set of 85 ORFs is shown in the attached drawings. Table 30 shows the nucleotide and predicted amino acid sequence of all 273
ORFs identified in bacteriophage Dp-1 that are identified as being expressed.
Table 31 shows the similarities identified in sequence between Streptococcus phage Dp-1 ORFs greater than 33 amino acids and sequences present in public sequence databases. Table 32 shows the 4731 bp sequence of Dp-1 published by Sheehan et al.,
1997).
Table 33 lists Streptococcus pneumoniae sequences listed in GenBank providing possible target sequences for inhibitory Streptococcus pneumoniae bacteriophage Dp-1 ORFs and other compounds with antibacterial activity
Background:
As indicated above, the present invention is concerned, in part, with the use of bacteriophage coding sequences and the encoded polypeptides or RNA transcripts to _ identify bacterial targets for potential new antibacterial agents. Thus, the invention concerns the selection of relevant bacteria. Particularly relevant bacteria are those which are pathogens of a complex organism such as an animal, e.g., mammals,
reptiles, and birds, and plants. Examples include Stapylococcus aureus, Enterococcus species, and Streptococcus pneumoniae. However, the invention can be applied to any bacterium (whether pathogenic or not) for which bacteriophage are available or which are found to have cellular components closely homologous to components targeted by phage of another bacterium.
Thus, the invention also concerns the bacteriophage which can infect a selected bacterium. Identification of ORFs or products from the phage which inhibit the host bacterium both provides an inhibitor compound and allows identification of the bacterial target affected by the phage-encoded inhibitor. Such targets are thus identified as potential targets for development of other antibacterial agents or inhibitors and the use of those targets to inhibit those bacteria. As indicated above, even if such a target is not initially identified in a particular bacterium, such a target can still be identified if a homologous target is identified in another bacterium. Usually, but not necessarily, such another bacterium would be a genetically closely related bacterium. Indeed, in some cases, a phage-encoded inhibitor can also inhibit such a homologous bacterial cellular component.
The demonstration that bacteriophage have adapted to inhibiting a host bacterium by acting on a particular cellular component or target provides a strong indication that that component is an appropriate target for developing and using antibacterial agents, e.g., in therapeutic treatments. Thus, the present invention provides additional guidance over mere identification of bacterial essential genes, as the present invention also provides an indication of accessability of the target to an inhibitor, and an indication that the target is sufficiently stable over time (e.g., not subject to high rates of mutation) as phage acting on that target were able to develop and persist. Thus, the present invention identifies a subset of essential cellular components which are particularly likely to be appropriate targets for development of antibacterial agents.
The invention also, therefore, concerns the development or identification of inhibitors of bacteria, in addition to the phage-encoded inhibitory proteins (or RNA transcripts), which are active on the targets of bacteriophage-encoded inhibitors. As described herein, such inhibitors can be of a variety of different types, but are preferably small molecules.
The following description provides preferred methods for use in the various aspects of the invention. However, as those skilled in the art will readily recognize, other approaches can be used to obtain and process relevant information. Thus-the- invention is not limited to the specifically described methods. In addition, the following description provides a set of steps in a particular order. That series of steps
describes the overall development involved in the present invention. However, it is clear that individual steps or portions of steps may be usefully practiced separately, and, further, that certain steps may be performed in a different order or even bypassed if appropriate information is already available or is provided by other sources or methods.
Selecting and Growing Phage. and Isolating DNA
Conceptually, the first step involves selecting bacterial hosts of interest. Preferably, but not necessarily, such hosts will be pathogens of clinical importance. Alternatively, because bacteria all share certain fundamental metabolic and structural features, these features can be targeted for study in one strain, for example a nonpathogenic one, and extrapolated to similarly succeed in pathogenic ones. Nonpathogenic strains may also exhibit initial advantages in being not only less dangerous, but also, for example, in having better growth and culturing characteristics and/or better developed molecular biology techniques and reagents. Consequently, advantageously the invention provides the ability target virtually any bacteria, but preferably pathogenic bacteria, with antimicrobial compounds designed and/or developed using bacteriophage inhibitory proteins and peptides from phage with nonpathogenic and/or pathogenic hosts. We have selected Staphylococcus aureus, Streptococcus pneumoniae, various
Enterococci, and Pseudomonas aeruginosa as initial exemplary pathogens. These bacteria are a major cause of morbidity and mortality in hospital-based infections, and the appearance of antibiotics resistance in all three organisms makes it increasingly difficult to treat benign infections involving these organisms. Such infections can include, for example, otitis media, sinusitis, and skin, and airway infections (Neu, H.C. (1992). Science 257, 1064-1073). However, the approach described below is clearly applicable to any human bacterial pathogens including but not restricted to Mycobacterium tuberculosis, Nesseria gonorrhoeae, Haemophilus influenza, Acinobacter, Escherichia coli, Shigella dysenteria, Streptococcus pyogenes, Helicobacter pylori, and Mycoplasma species. This invention can also be applied to the discovery of anti-bacterial compounds directed against pathogens of animals other than humans, for example, sheep, cattle, swine, dogs, cats, birds, and reptiles. Similarly, the invention is not limited to animals, but also applies to plants and plant pathogens. In general, the bacteria are grown according to standard methodologies -, employed in the art, including solid, semi-solid or liquid culturing, which procedures can be found in or extrapolated from standard sources such as Maloy, S.R., Stewart,
V.J., and Taylor, R.K. Genetic Analysis of Pathogenic Bacteria (1996) Cold Spring Harbor Laboratory Press, or Maniatis, T. et al. (1989) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor University Press, Cold Spring, N.Y.; or Ausubel, F.M. et al. (1994) Current Protocols in Molecular Biology. John Wiley & Sons, Secaucus, N J. Culture conditions are selected which are adapted to the particular bacterium generally using culture conditions known in the art as appropriate, or adaptations of those conditions.
Nucleic acids within these bacteria can be routinely extracted through common procedures such as described in the above-referenced manuals and as generally known to those skilled in the art. Those nucleic acid stocks can then be used to practice the other inventive aspects described below.
Selection and Growth of Bacteriophage. and Isolation of DNA
The second step involves assembling a group of bacteriophages (phage collection) for one or more of the targeted bacterial hosts. While the invention can be utilized with a single bacteriophage for a pathogen or other bacterium, it is preferable to utilize a plurality of phage for each bacterium, as comparisons between a plurality of such phage provides useful additional information. Non-limiting examples of phage and sources for some of the above-mentioned pathogenic bacteria are found in Table 1. The criteria used to select such phages is that they are infectious for the microbe targeted, and replicate in, lyse, or otherwise inhibit growth of the bacterium in a measurable fashion. These phages can be very different from one another (representing different families), as judged by criteria such as morphology (head, tail, plate, etc.), and similarity of genome nucleotide sequence (cross-hybridization). Since such diverse bacteriophages are expected to block bacterial host metabolism and ultimately inhibit by a variety of mechanisms, their combined study will lead to the identification of different mechanisms by which the phages independently inhibit bacterial targets. Examples include degradation of host DNA (Parson K.A., and Snustad, D.P. (1975). J. Virol. 15, 221-444) and inhibition of host RNA transcription (Severinova, E., Severinov, K. and Darst, S.A. (1998;. J.Mol. Biol. 279, 9-18). This, in turn, yields novel information on phage proteins that can inhibit the targeted microbe. As explained below, this 1) forms the basis of novel drug discovery efforts based on knowledge of the primary amino acid sequence of the phage inhibitor protein (e.g., peptide fragments or peptidomimetics) and/or 2) leads to the identification of bacterial biochemical pathways, the proteins of which are essentiaTor significant for survival of the targeted microbe, and which enzymatic steps or
chemical reactions can be targeted by classical drug discovery methods using molecular inhibitors, for example, small molecule inhibitors.
Bacteriophage are generally either of two types, lytic or filamentous, meaning they either outright destroy their host and seek out new hosts after replication, or else continuously propogate and extrude progeny phage from the same host without destroying it. Regardless of the phage life cycle and type, preferred embodiments incorporate phage which impede cell growth in measurable fashion and preferably stop cell growth. To this end, lytic phage are preferred, although certain nonlytic species may also suffice, e.g., if sufficiently bacteriostatic. Various procedures that are commonly understood by those of skill in the art can be routinely employed to grow, isolate, and purify phage. Such procedures are exemplified by those found in such common laboratory aids such as Maloy, S.R., Stewart, V.J., and Taylor, R.K. Genetic Analysis of Pathogenic Bacteria (1996) Cold Spring Harbor Laboratory Press; Maniatis, T. et al. (1989) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor University Press, Cold Spring, N.Y.; and Ausubel, F.M. et al. (eds.) (1994) Current Protocols in Molecular Biology. John Wiley & Sons, Secaucus, NJ. The techniques generally involve the culturing of infected bacterial cells that are lysed naturally and/or chemically assisted, for example, by the use of an organic solvent such as chloroform that destroys the host cells thereby liberating the phage within. Following this, the cellular debris is centrifuged away from the supernatant containing the phage particles, and the phage then subsequently and selectively precipitated out of the supernatant using various methods usually employing the use of alcohols and/or other chemical compounds such as polyethylene glycol (PEG). The resulting phage can be further purified using various density gradient/centrifugation methodologies. The resulting phage are then chemically lysed, thereby releasing their nucleic acids that can be conveniently precipitated out of the supernatant to yield a viral nucleic acid supply of the phage of interest.
Exemplary bacteriophage are indicated in Table 1, along with sources where those phage may be obtained.
Exemplary bacteria include the reference bacteria for the identified bacteriophage, available from the same sources.
Characterizing Bacteriophage Genomes for ORFs The third step involves systematically characterizing the genetic information contained in the phage genome. Within this genetic information is the sequence of all RNAs and proteins encoded by the phage, including those that are essential or
instrumental in inhibiting their host. This characterization is preferably done in a systematic fashion. For example, this can be done by first isolating high molecular weight genomic DNA from the phage using standard bacterial lysis methods, followed by phage purification using density gradient ultracentrifugation, and extraction of nucleic acid from the purified phage preparation. The high molecular weight DNA is then analyzed to determine its size and to evaluate a proper strategy for its sequencing. The DNA is broken down into smaller size fragments by sonication or partial digestion with frequently cutting restriction enzymes such as Sau3A to yield predominantly 1 to 2 kilobase length DNA, which DNA can then be resolved by gel electrophoresis followed by extraction from the gel.
The ends of the fragments are enzymatically treated to render them suitable for cloning and the pools of fragments are cloned in a bacterial plasmid to generate a library of the phage genome. Several hundred of these random DNA fragments contained in the plasmid vector are isolated as clones after introduction into an appropriate bacterium, usually Escherichia coli. They are then individually expanded in culture and the DNA from each individual clone is purified. The nucleotide sequences of the inserts of these clones are determined by standard automated or manual methods, using oligonucleotide primers located on either side of the cloning site to direct polymerase mediated sequencing (e.g., the Sanger sequencing method or a modification of that method). Other sequencing methods can also be used. The sequence of individual clones is then deposited in a computer, and specific software programs (for example, Sequencher™, Gene Codes Corp.) are used to look for overlap between the various sequences, resulting in ordering of contig sequences and ultimately providing the complete sequence of the entire bacteriophage genome (one such example is given in Table 2 for Staphylococcus aureus bacteriophage 77; others are also provided herein). This complete nucleotide sequence is preferably determined with a redundancy of at least 3- to 5-fold (number of independent sequencing events covering the same region) in order to minimize sequencing errors. Preferably, the bacterial strain used as a phage host should not possess any other innate plasmids, transposons, or other phage or incompatible sequences that would complicate or otherwise make the various manipulations and analyses more difficult.
Commercially available computer software programs are used to translate the nucleotide sequence of the phage to identify all protein sequences encoded by tb_e phage (hereafter called open reading frames or ORFs). (Customized software can clearly also be used.) As phages are known to transcribe their genome into RNA from
both strands, in both directions, and sometimes in more than one frame for the same sequence, this exercise is done for both strands and in all six possible reading frames. As evolutionary constraints have forced the phage to conserve all of its vital protein sequences in as small a genome as possible, it is straightforward to identify all the proteins encoded by the phage by simple examination of the 6 translation frames of the genome. Once these ORFs are identified, they are cataloged into a phage proteome database (Table 3 lists ORFs identified from phage 77; ORF lists are also provided for other exemplary phage). This analysis is preferably performed for each phage under study. The process of ORF identification can be varied depending on the desired results. For example, the minimum length for the putative encoded polypeptide can be varied, and/or putative coding regions that have an associated Shine-Dalgarno sequence can be selected. In the case of phage 77 ORFs, such parameter adjustment was performed and resulted in the identification of ORFs as listed herein. Different parameters had resulted in the identification of the ORFs listed in the preceding U.S. Provisional Application 60/110,992, filed December 3, 1998, which is hereby incorporated by reference in its entirety.
Exemplary phage 77 ORFs identified in that provisional application and as identified herein are shown in the following table:
Identifying and Characterizing Inhibitory Phage ORFs
The fourth step entails identifying the phage protein or proteins or RNA transcripts that have the ability to inhibit their bacterial hosts. This can be accomplished, for example, by either or both of two non-mutually exclusive methods. The first method makes use of bioinformatics. Over the past few years, a large amount of nucleotide sequence information and corresponding translated products have become available through large genome sequencing projects for a variety of organisms including mammals, insects, plants, unicellular eukaryotes (yeast and fungi), as well as several bacterial genomes such as E. coli, Mycobacterium tuberculosis, Bacillus subtilis, Staphylococcus aureus and many others. Such sequences have been deposited in public databases (for example, non-redundant
sequence database at GenBank and SwissProt protein sequence database) (http://www.ncbi.nlm.nih.gov)) and can be freely accessed to compare any specific query sequence to those present in such databases. For example, GenBank contains over 1.6 billion nucleotides corresponding to 2.3 million sequence records. Several computer programs and servers (e.g., TBLASTN) have been created to allow the rapid identification of homology between any given sequence from one organism to that of another present in such databases, and such programs are public and available free of charge.
In addition, it has been well established that basic biochemical pathways can be conserved in very distant organisms (for example bacteria and man), and that the proteins performing the various enzymatic steps in these pathways are themselves conserved at the amino acid sequence level. Thus, proteins performing similar functions (e.g. DNA repair, RNA transcription, RNA translation) have frequently preserved key structural signatures, identifiable by similarities across regions of proteins (domains and motifs). The antimicrobials of the present invention will preferably target features and targets that are highly characteristic or conserved in microbes, and not higher organisms.
Most genomes encode individual proteins or groups of proteins that can be assembled into protein families that have been evolutionarily conserved. Therefore, similarity between a new query sequence and that of a member of a protein family (reference sequences from public databases) can immediately suggest a biochemical function for the novel query sequence, which in our case is a phage ORF.
The sequence homology between individual members of evolutionarily distant members of a protein family is usually not randomly distributed along the entire length of the sequence but is often clustered into "motifs" and "domains". These correspond to key three-dimensional folds that form key catalytic and/or regulatory structures that perform key biochemical function(s) for the group of proteins. Commercially available computer software programs can identify such motifs in a new query sequence, again providing functional information for the query sequence. Such structural and functional motifs have also been derived from the combined analysis of primary sequence databases (protein sequences) and protein structure databases (X-ray crystallography, nuclear magnetic resonance) using so-called "threading" methods (Rost B,l and Sander C. (1996) Ann. Rev. Biophy. Biomol. Struct. 25, 113-136). Such motifs and folds are themselves deposited in public databases whichϋan be directly accessed (for example, SwissProt database; 3D-ALI at EMBL, Heidelberg; PROSITE). This basic exercise leads to a structural homology map in which each of
the phage ORFs has been probed for such similarities, and where initial structural and functional hits are identified (selected examples of sequence homologies detected between individual ORFs from the genome of Staphylococcus aureus bacteriophage 77 and sequences deposited in public databases are shown in Table 5 for ORFs 17/19/43/102/104/182).
This analysis can point out phage proteins with similarity to proteins from other phages (such as those for E. coli) playing an important role in the basic biochemical pathways of the phage (such as DNA replication, RNA transcription, tRNAs, coat protein and assembly). Selected examples of such proteins include integrase and capsid protein. Therefore, this analysis enables identification and elimination of non-essential ORFs as candidates for an inhibitor function, as well as the identification of (potentially) useful ones.
In addition, this analysis can point out specific ORFs as possible inhibitor ORFs. For example these ORFs may encode proteins or enzymes that alter bacterial cell structure, metabolism or physiology, and ultimately viability. Examples of such proteins present in the genome of Staphylococcus aureus bacteriophage 77 include orfl4 (deoxyuridine triphosphatase from bacteriophage T5), and orfl5 (sialidase). (These ORF identifications are as listed in provisional application 60/110,992.) Other examples include ORFs 9 and 12 of S. aureus phage 44 AHJD, which encode the putative lysis functions found in many bacteriophages - a "holin" and an "amidase". In addition, it is well known that bacterial and eukaryotic viruses can usurp pathways from their host in order to use them to their advantage in blocking host cellular pathways upon infection. The phage can achieve this by 1) directly producing an inhibitor of a key host pathway (e.g. T7 gene 0.5 and 2), 2) directly producing a novel activity (e.g. T4 DNA polymerase), and 3) altering concentrations of cell components by producing similar functions (e.g. T4 transfer RNAs). The identification of sequence similarity between phage ORFs and bacterial host genome sequences will be highly indicative of such a mechanism. (Selected examples of such homologies are listed in Figure 4 of the provisional application 60/110,992 and include orf4 (homologous to autolysin), orf20 (hypothetical protein from
Staphyloccus aureus) and orf29 (hypothetical protein from Staphyloccus aureus.)) These ORFs can be analyzed by a standard biochemical approach to directly test their inhibitor functions (e.g., as described below).
Alternatively, a homology search may reveal that a given phage ORF is related to a protein present in the databases having an activity known to be inhibitory, (e.gA inhibitor of host RNA polymerase by E. coli bacteriophage T7. Such a finding would implicate the phage ORF product in a related activity. This will also suggest that a
new antimicrobial could be derived by a mimetic approach (e.g., peptidomimetic) imitating this function or by a small molecule inhibitor to the bacterial target of the phage ORF, or any steps in the relevant host metabolic pathway, e.g., high throughput screening of small molecule libraries. Selected examples of such similarity between ORFs of Staphyloccus aureus bacteriophage 77 and proteins with inhibitor functions for bacterial hosts are listed in Figure 4 of the provisional application 60/110,992. These include orf9 (similar to bacteriophage PI kilA function), and orf4 (autolysin of Staphylococcus aureus, amidase enzymatic activity).
A reason for the biochemical study of individual ORFs for inhibitor function is that their expression or overexpression will block cellular pathways of the host, ultimately leading to arrest and/or inhibition of host metabolism. In addition, such ORFs can alter host metabolism in different ways, including modification of pathogenicity. Therefore, individual ORFs identified above are expressed, preferably overexpressed, in the host and the effect of this expression or overexpression on host metabolism and viability is measured. This approach can be systematically applied to every ORF of the phage, if necessary, and does not rely on the absolute identification of candidate ORFs by bioinformatics. Individual ORFs are resynthesized from the phage genomic DNA, e.g., by the polymerase chain reaction (PCR), preferably using oligonucleotide primers flanking the ORF on either side. These single ORFs are preferably engineered so that they contain appropriate cloning sites at their extremities to allow their introduction into a new bacterial expression plasmid, allowing propagation in a standard bacterial host such as E. coli, but containing the necessary information for plasmid replication in the target microbe such as S. aureus (hereafter referred to as shuttle vector). Shuttle vectors and their use are well known in the art. Such shuttle vectors preferably also contain regulatory sequences that allow inducible expression of the introduced ORF. As the candidate ORF may encode an inhibitor function that will eliminate the host, it is beneficial that it not be expressed prior to testing for activity. Thus, screening for such sequences when expressed in a constitutive fashion is less likely to be successful when the inhibitor is lethal. In the exemplary inducible system presented in Figure 1A, IB, 2, and 7, regulatory sequences from the ars operon of S. aureus are used to direct individual ORF expression in S. aureus (or other bacteria in which the ars system is functional). The ars operon encodes a series of proteins which normally mediate the extrusion of arsenite and other trivalent oxyanions from the cells when they are exposed to such toxic substances in their environment. The operon encoding this detoxifying _ mechanism is normally silent and only induced when arsenite-related compounds are
present. (Tauriainen, S. et al. (1997) App. Env. Microb., Vol. 63, No. 11, p. 4456- 4461.)
Therefore, individual phage ORFs can be expressed in S. aureus in an inducible fashion by adding to the culture medium non-toxic arsenite concentrations during the growth of individual S. aureus clones expressing such individual phage ORFs. Toxicity of the phage inhibitor ORF for the host is monitored by reduction or arrest of growth under induction conditions, as measured by optical density in liquid culture or after plating the induced cultures on solid medium. Subsequently, interference of the phage ORF with the host biochemical pathways ultimately leading to reduced or arrested host metabolism can be measured by pulse-chase experiments using radiolabeled precursors of either DNA replication, RNA transcription, or protein synthesis. Similar constructs can be made and used for other bacteria using well- known techniques.
Those skilled in the art are familiar with a variety of other inducible systems which can also be used for the controlled expression of phage ORFs, including, for example, lactose (see e.g., Stratagene' s LacSwitch™II system; La Jolla, CA) and tetracycline-based systems (see, e.g. Clontech's Tet On/Tet Off™ system; Palo Alto, CA). The arsenite-inducible system described is further depicted in Figures 1, 2 and 7.
The selection or construction of shuttle vectors and the selection and use of inducible systems are well known and thus other shuttle vectors appropriate for other bacteria can be readily provided by those skilled in the art, e.g., for use in other bacterial species.
Standard methodologies for expressing proteins from constructs, and isolating and manipulating those proteins, for example in cross-linking and affinity chromatography studies, may be found in various commonly available and known laboratory manuals. See, e.g., Current Protocols in Protein Science. John Wiley & Sons, Secaucus, N.J., and Maniatis, T. et al. H989 Molecular Cloning: A Laboratory Manual. Cold Spring Harbor University Press, Cold Spring, N. Y.
It has been found that certain phage or other viruses inhibit host cells, at least in part, by producing an antisense RNA which binds to and inhibits translation from a bacterial RNA seqeunce. Thus, in the case of potentially inhibitor RNA transcripts encoded by the phage genome, a strong indicator of a possible inhibitory function is provided by the identification of phage sequence which is the identical to or fully complementary (or with only a small percentage of mismatch, e.g., <10%, preferably less than 5%, most preferably less than 3%, to a bacterial sequence. This approaches convenient in the case of bacteria that have been essentially completely sequenced, as the comparison can be performed by computer using public database information.
The inhibitory effect of the transcript can be confirmed using expression of the phage sequence in a host bacterium. If needed, such inhibitory can also be tested by transfecting the cells with a vector that will transcribe the phage sequence to form RNA in such manner that the RNA produced will not be translated into a polypeptide. Inhibition under such conditions provides a strong indication that the inhibition is due to the transcript rather than to an encoded polypeptide.
In an alternative, the expression of an ORF in a host bacterium is found to be inhibitory, but the inhibition is found to be due to an RNA product of the genomic coding region. For antisense inhibition, the sequence of the bacterial target nucleic acid sequence can be identified by inspection of the phage sequence, and the full sequence of the relevant coding region for the bacterial product can be found from a database of the bacterial genomic sequence or can be isolated by standard techniques (e.g., a clone in a genomic library can be isolated which contains the full bacterial ORF, and then sequenced). In either case, the identification of a target which is inhibited by an RNA transcript produced by a phage provides both the possible inhibition of bacteria naturally containing the same target nucleic acid sequence, as well as the ability to use the target sequence in screening for other types of compounds which will act directly on the target nucleic acid sequence or on a polypeptide product expressed or regulated, at least in part, by the target of the inhibitory phage RNA.
In some cases it will be found that the target of an inhibitory phage RNA or protein has previously been found to be a target of an inhibitory phage RNA or protein has previously been found to be a target for an antibacterial agent. In such cases, the phage inhibitor can still provide useful information if it is found that the phage-encoded product acts at a different site than the previously identified antibacterial agent or inhibitor, i.e., acts at a phage-specific site. For many targets, action at a different site provides highly beneficial characteristics and/or information. For example, an alternate site of inhibitor action can at least partially overcome a resistance mechanism in a bacterium. As an illustration, in many cases, resistance is due, in large part, to altered binding characteristics of the immediate target to the antibacterial agent. The altered binding is due to a structural change which prevents or destabilizes the binding. However, the structural change is frequently quite local, so that compounds which bind at different local sites will b unaffected or affected to a much lesser degree. Indeed, in some cases the local sites will be on a different molecule and so may be completely unaffected by the local structural change creating resistance to the original agent(s). An example of resistance due to altered binding is
provided by methicillin-resistant Staphylococcus aureus, in which the resistance is due to an altered penicillin-binding protein.
In other cases, a new site of action can have improved accessibility as compared to a site acted on by a previously identified agent. This can, for example, assist in allowing effective treatment at lower doses, or in allowing access by a larger range of types of compounds, potentially allowing identification of more potential active agents.
Another advantage is that the structural characteristics of a different site of action will lead to identification and/or development of inhibitors with different structures and different pharmacological parameter. This can allow a greater range of possibilities when selecting an antibacterial agent.
Yet further, different sites often produce different inhibitory characteristics in the target organism. This is commonly the case for multi-domain target proteins. Thus, inhibition targeting an alternate site can produce more efficacious action, e.g., faster killing, slower development of resistance, lower numbers of surviving cells, and different secondary effects (for example, different nutrient utilization).
Staphylococcus aureus phage 77
As indicated above, the present invention is concerned, in part, with the use of bacteriophage 77 coding sequences and the encoded polypeptides or RNA transcripts to identify bacterial targets for potential new antibacterial agents.
As described, phage 77 ORFs 17, 19, 43, 102, 104, and 182 have been found to have bacteria inhibiting function. Identification of ORFs 17, 19, 43, 102, 104, and 182 and products from the phage which inhibit the host bacterium both provides an inhibitor compound and allows identification of the bacterial target affected by the phage-encoded inhibitor. Such a target is thus identified as a potential target for development of other antibacterial agents or inhibitors and the use of those targets to inhibit those bacteria. As indicated above, even if such a target is not initially identified in a particular bacterium, such a target can still be identified if a homologous target is identified in another bacterium. Usually, but not necessarily, such another bacterium would be a genetically closely related bacterium. Indeed, in some cases, an inhibitor encoded by phage 77 ORF 17, 19, 43, 102, 104, or 182 can also inhibit such a homologous bacterial cellular component. -
Possible bacterial target sequences are described herein by reference to sequence source sites. In preferred embodiments, the sequence encoding the target corresponds
to a S. aureus nucleic acid sequence available from numerous sources including S. aureus sequences deposited in GenBank, S. aureus sequences found in European Patent Application No. 97100110.7 to Human Genome Sciences, Inc. filed January 7, 1997, S. aureus sequences available from TIGR at http://www.tigr.org/tdb/mdb/mdb.html. and S. aureus sequences available from the Oklahoma University S. aureus sequencing project at the following URL: http://www.genome.ou.edu/staph new.html. Such possible targets are particularly applicable to S aureus phages 77, 3A, 96, and 44 AHJD.
The amino acid sequence of a polypeptide target is readily provided by translating the corresponding coding region. For the sake of brevity, the sequences are not reproduced herein. Also, in preferred embodiments, a target sequence corresponds to a S. aureus coding sequence corresponding to a sequence listed in Table 15 herein. The listing in Table 15 describes S. aureus sequences currently listed with GenBank. Again, for the sake of brevity, the sequences are described by reference to the database accession numbers instead of being written out in full herein. In cases where an entry for a coding region is not complete, the complete sequence can be readily obtained by routine methods, e.g., by isolating a clone in a phage host S. aureus genomic library, and sequencing the clone insert to provide the relevant coding region. The boundaries of the coding region can be identified by conventional sequence analysis and/or by expression in a bacterium in which the endogenous copy of the coding region has been inactivated and using subcloning to identify the functional start and stop codons for the coding region.
Staphyloccus aureus phage 44 AHJD The present invention also can utilize the identification of naturally occuring
DNA sequence elements within Staphylococcus aureus bacteriophage 44AHJD which encode proteins with antimicrobial activity.
Such identification can utilize bioinformatics identification of specific proteins
(ORFs) utilized by Staphylococcus aureus bacteriophage 44AHJD during the viral life cycle, resulting in a slowing or arrest of growth of the bacterial host, or in death, of the Staphylococcus aureus host including lysis of the infected bacteria. Thus, some of the bacteriophage 44AHJD DNA sequences encoding these proteins (ORFs) are predicted to encode antimicrobial functions. Information derived from these DNA sequences and translated ORFs can, in turn, be utilized to develop inhibitory __ compounds by peptidomimetics that can also function as antimicrobials. In addition, the identification of the host bacterial proteins that are targeted and inhibited by the
antimicrobial bacteriophage ORFs can themselves provide novel targets for drug discovery.
The methodology described above is used to identify and characterize DNA sequences from Staphylococcus sp. bacteriophage 44 AHJD that have antimicrobial activity. As described in the Examples, the Staphylococcus aureus propagating strain (PS 44A), obtained from the Felix d'Herelle Reference Centre (#HER 1101), was used as a host to propagate its phage 44AHJD, also obtained from the Felix d'Herelle Reference Centre (#HER 101). By sequencing, we found that bacteriophage 44AHJD consists of 16,668 bp (Table 16) predicted to encode 73 ORFs greater than 33 amino acids (Tables 17 & 18). Computational analysis of the predicted protein products of Staphylococcus aureus bacteriophage 44AHJD identified homolgs in public sequence databases as listed inTable 19 and 20, along with the accompanying list of related proteins.
From this analysis, it is apparent that 3 genes (ORF 3, 7, and 8) are related to structural proteins found in other bacteriophages. These include genes predicted to encode a tail protein (ORF 3), an upper collar/connector protein of the phage virion (ORF 7), and a lower collar protein (ORF 8). Bioinformatics has also identified one gene whose product is likely involved in phage DNA synthesis. One gene (ORF 1) shows significant homology to DNA polymerases of a number of bacteriophages, bacteria and fungi, and the product of this gene is likely responsible for replicating the genetic material of bacteriophage 44 AHJD. ORF 2 encodes a protein with homology to the dinC gene of Bacillus subtilis that encodes a protein involved in teichoic acid biosynthesis. Teichoic acid is a polyphosphate polymer found in some, but not all, Gram positive organisms (and not in Gram negative organisms), where it is attached to the peptidoglycan layer. The phage protein may thus be involved in the synthesis of this material for incorporation into the cell wall, allowing enhanced lysis by the phage lysis enzymes or, as many enzymes can function in "reverse reactions", may be involved in its degradation allowing for penetration of the peptidoglycan and phage genome entry into the cell following adsorption. The similarity between Staphylococcus aureus bacteriophage 44AHJD and E. coli phage T7 indicates that they may share similar mechanisms of replication and growth. Both phages belong!0 the Pododviridae Family of bacteriophages and are members of the "T7-like" Genus of this Family (Ackermann and DuBow; Vlth ICTV Report).
Two genes, ORF 9 and 12, were identified with the potential to encode antimicrobial protein products. The homology alignments are shown in Tables 19 and 20. The predicted product of ORF 9 is related to a class of genes which encodes lysozyme-like functions, enzymes which cleave linkages in the mucopolysaccharide cell wall structure of a variety of micro-organisms, including that from the
Staphylococcus aureus bacteriophage Twort. ORF 12 of Staphylococcus aureus bacteriophage 44AHJD shows homology to a set of lysis proteins from several bacteriophages. These lysis proteins are also referred to as holins, and represent phage-encoded lysis functions required for transit of the phage murein hydrolases (lysozyme) to the periplasm, where it can digest the cell wall and thus lyse the bacterium.
Thus, in particular embodiments, the present invention provides a nucleic acid sequence isolated from Staphylococcus aureus bacteriophage 44AHJD comprising at least a portion of one of the genes described above with antimicrobial activity. For example, ORF 1 encodes a DNA polymerase function. This polymerase may utilize host-derived accessory proteins for its activity when replicating the phage template, sequestering such proteins from use by the bacterial polymerase, resulting in inhibition of DNA replication, cell division, and cell growth. Alternatively, ORF 9 directly encodes a polypeptide with antimicrobial activity. ORF 9 is predicted to encode an amidase, a protein known to act as a cell wall degrading enzyme. ORF 12 likely encodes a holin function required for transit of the phage amidase (gene 9 product) to the periplasm. When this type of gene product from Bacillus phage phi 29 (gene 14), was cloned in Escherichia coli, cell death ensued (Steiner et al., 1993). Thus, production of proteins from Bacillus phage phi 29 gene 14 in E. coli resulted in cell death, whereas production of protein from Bacillus phage phi 29 gene 14 concomitantly with the phi 29 lysozyme or unrelated murein-degrading enzymes led to lysis, suggesting that membrane-bound protein 14 induces a nonspecific lesion in the cytoplasmic membrane (Steiner et al., 1993).
The present invention also provides the use of the Staphylococcus bacteriophage 44 AHJD antimicrobial ORFs or ORF products as pharmacological agents, either wholly or in part and derivatives, as well as the use of conespαndThg peptidomimetics, developed from amino acid or nucleotide sequence knowledge derived from Staphylococcus bacteriophage 44 AHJD killer ORFs.
Enterococcus phage 182
Bacteriophage 182 was obtained from the Felix D'Herelle phage collection
(Ste. Foy, Quebec) and infects Enterococcus sp. Group D. The genome of Enterococcus bacteriophage 182 consists of 17,833 bp (Table 21) and is predicted to encode 80 ORFs greater than 33 amino acids (Tables 22 and 23). Computational analysis of the predicted protein products of Enterococcus bacteriophage 182 was performed in order to identify protein products related to those deposited in public databases. Bacteriophage 182 protein products which detected sequences with significant sequence similarity in public databases are listed in Table 24 and 26, along with the accompanying list of related proteins.
From this analysis, it is apparent that 5 genes (ORF 001, 004, 007, 009, and 011) are related to structural proteins of several Bacillus phages - Bacillus bacteriophage PZA, phi-29, and B103. These include genes predicted to encode a tail protein (ORF 001), a head protein (ORF 004), and upper collar protein (ORF 007), a lower collar protein (ORF 009), and a pre-neck appendage protein (ORF 011). Two gene products are predicted to encode genes which direct phage morphogenesis - these are ORF 005 and 019.
Bioinformatics has also identified three genes whose products are likely involved in phage DNA synthesis. One gene, ORF 002 shows significant homology to DNA polymerases of a number of bacteriophages, and the product of this gene is likely responsible for replicating the genetic material of bacteriophage 182. ORF 006 encodes a protein with homology to the encapsidation proteins of several other bacteriophages, including Bacillus phage phi-29 (PI 1014), PZA (P07541), and B 103 (X99260) and Streptococcus phage CP-1 (Z47794). These gene products catalyze the in vivo and in vitro genome-encapsidation reaction (Garvey et al., 1985). Proteins involved in genome packaging have been shown to have additional activities that affect biochemical reactions in other phages and their hosts. For example, the coat protein of the RNA bacteriophage MS2 interacts with viral RNA to translationally repress replicase synthesis (Pickett and Peabody, 1993). This protein-RNA interaction also plays a role in genome encapsidation, enveloping a single copy of the viral " genome in a protein shell composed of many molecules of coat protein. In addition, the bacteriophage λ terminase enzyme can be lethal to E. coli when expressed,
suggesting cleavage of packaging sites in the bacterial chromosome. Also present within bacteriophage 182 is a gene, ORF 010, that encodes a protein that is related to the terminal proteins of Bacillus phage Nf (P06812), Bacillus phage GA-1 (X96987) and Bacillus phage B103 (X99260). DNA terminal proteins are linked to the 5' ends of both strands of the genome and are essential for DNA replication playing a role in initial priming of DNA replication. The similarity between Enterococcus bacteriophage 182 and Bacillus phages phi-29, PZA, and B103 indicates that they may share similar mechanisms of replication and growth. Protein-primed DNA replication is a well described phenomenon, and in the phi-29-like phages, the ends of the DNA serve as origins and termini of replication (Gutierrez et al., 1986; Yoshikawa et al., 1985).
There is also a gene (ORF 015) that encodes a protein showing homology to an early protein product of Bacillus bacteriophage PZA and the single-strand nucleic acid binding protein of bacteriophage B103. Two genes, ORF 008 and 014, were identified with the potential to encode anti-microbial protein products. The homology alignments are shown in Tables 24 & 26 and biochemical features of the predicted polypeptides shown in Table 25. The predicted product of ORF 008 is related to a class of genes which encodes lysozyme- like functions, enzymes which cleave linkages in the mucopolysaccharide cell wall structure of a variety of micro-organisms. ORF 014 of Enterococcus 182 shows homology to a set of lysis proteins from Bacillus bacteriophage phi-29, PZA, and B103. These lysis proteins are also referred to as holins and represent phage encoded lysis functions required for transit of the phage murein hydrolases (lysozyme) to the periplasm, where it can digest the outer cell wall and thus lyse the bacterium. Thus, the present invention provides a nucleic acid sequence obtained from
Enterococcus bacteriophage 182 comprising at least a portion of a phage 182 ORF, preferably an inhibitory ORF, and more preferably at least a portion of one of the genes described above with anti-microbial activity. For example, ORF 002 encodes a DNA polymerase function. This polymerase may utilize host-derived accessory proteins for its activity when replicating the phage template, sequestering such proteins from use by the bacterial polymerase, resulting in inhibition of DNA replication, cell division, and cell growth. Alternatively, ORFs 008 or 014 directly encode polypeptides with anti-microbial activity. ORF 008 is predicted to encode an
autolytic lysozyme, a protein known to have anti-microbial activity (Martin et al, 1998). ORF 014 likely encodes a holin function required for transit of the phage murein hydrolases to the periplasm. When the related product from Bacillus phage phi 29 (gene 14), was cloned in Escherichia coli, cell death ensued (Steiner et al, 1993). Thus, production of proteins from Bacillus phage phi 29 gene 14 in E. coli resulted in cell death, whereas production of protein from Bacillus phage phi 29 gene 14 concomitantly with the phi 29 lysozyme or unrelated murein-degrading enzymes led to lysis, suggesting that membrane-bound protein 14 induces a nonspecific lesion in the cytoplasmic membrane (Steiner et al, 1993). The present invention also provides the use of the Enterococcus bacteriophage
182 anti-microbial ORFs as pharmacological agents, either wholly or in part and derivatives, as well as the use of corresponding peptidomimetics, developed from amino acid or nucleotide sequence knowledge derived from Enterococcus bacteriophage 182 killer ORFs. This can be done where the structure of the peptidomimetic compound corresponds to the structure of the active portion of a product of an ORF. In this analysis, the peptide backbone is transformed into a carbon based hydrophobic structure that can retain cytostatic or cytocidal activity for the bacterium. This is done by standard medicinal chemistry methods, measuring growth inhibition of the various molecules in liquid cultures or on solid medium. These mimetics also represent lead compounds for the development of novel antibiotics. In this context, "corresponds" means that the peptidomimetic compound structure has sufficient similarities to the structure of the active portion of a product of one of the Enterococcus ORFs listed, that the peptidomimetic will interact with the same molecule as the product of the ORF, and preferably will elicit at least one cellular response in common which relates to the inhibition of the cell by the phage protein.
To validate the identity of an ORF as a killer ORF, it is preferably expressed in the host or other test bacterial organism and the effect of this expression on bacterial growth and replication is assessed. Therefore, all individual ORFs identified herein, e.g., those identified above, can be expressed, preferably overexpressed, in a suitable host bacterium e.g., a host Enterococcus and the effect of this expression or overexpression on host metabolism and viability can be measured. _
Individual ORFs can be resynthesized from the phage genomic DNA by the polymerase chain reaction (PCR) using oligonucleotide primers flanking the ORF on
either side. Those skilled in the art are familiar with the design and synthesis of appropriate primer sequences. These single ORFs are preferably engineered so that they contain appropriate cloning sites at their extremities to allow their introduction into a new bacterial expression plasmid, allowing propagation in a standard bacterial host such as E. coli, but containing the necessary information for plasmid replication in the target microbe, Enterococcus sp. (hereafter referred to as a shuttle vector).
This shuttle vector also preferably contains regulatory sequences that allow inducible expression of the introduced ORF. As the candidate ORF may encode a killer function that will eliminate the host, it is highly advantageous that it not be expressed (or at least not expressed at a substantial level) prior to testing for activity; thus screening for such sequences in a constitutive fashion is less likely to be successful (lethality). In an example presented in Fig. 7, regulatory sequences from the ars operon are used to direct individual ORF expression in Enterococcus. The ars operon encodes a series of proteins which normally mediate the extrusion of arsenite and several other trivalent oxyanions from the cells when they are exposed to such toxic substances in their environment. The operon encoding this detoxifying mechanism is normally silent and only induced when arsenite-related compounds are present.
Therefore, individual phage ORFs can be expressed in Enterococcus or other suitable host in an inducible fashion by adding to the culture medium non-toxic arsenite concentrations during the growth of individual Enterococcus (or other host cells) clones expressing such individual phage ORFs. Toxicity of the phage killer ORF for the host is monitored by reduction or arrest of growth under induction conditions, as measured by optical density in liquid culture or after plating the induced cultures on solid medium. Subsequently, interference of the phage ORF with the host biochemical pathways ultimately leading to reducing or arresting host metabolism can be measured by pulse chase experiments using radiolabeled precursors of either DNA replication, RNA transcription, or protein synthesis.
Of course, other inducible regulatory sequences (e.g., promoters, operators, etc.) may be used (e.g., systems using positive induction of expression or systems using release of repression). A variety of such systems are known to those- skilled in the art and can be utilized in the present invention.
Nucleic acid sequences of the present invention can be isolated using a method similar to those described herein or other methods known to those skilled in the art. In addition, such nucleic acid sequences can be chemically synthesized by well- known methods. Having the phage 182 ORFs, e.g., anti-bacterial ORFs of the present invention, portions thereof, or oligonucleotides derived therefrom as described, other anti-microbial sequences from other bacteriophage sources can be identified and isolated using methods described here or other methods, including methods utilizing nucleic acid hybridization and/or computer-based sequence alignment methods.
The invention also provides bacteriophage anti-microbial DNA segments from other phages based on nucleic acids and sequences hybridizing to the presently identified inhibitory ORF under high stringency conditions or sequences which are highly homologous. The bacteriophage anti-microbial DNA segment from bacteriophage 182 can be used to identify a related segment from another unrelated phage based on stringent conditions of hybridization or on being a homolog based on nucleic acid and/or amino acid sequence comparisons. As with the phage 182 inhibitory sequences, such homologous coding sequences and products can be used as antimicrobials, to construct active portions or derivatives, to construct peptidomimetics, and to identify bacterial targets.
Enterococcus sequences are listed in Table 27 by accession number, providing identification of possible targets of Enterococcus phage inhibitory ORF products, e.g., from phage 182.
Streptococcus pneumoniae
As indicated in the Summary above, the present invention is concerned with the use of Streptococcus sp. bacteriophage Dp-1 coding sequences and the encoded polypeptides or RNA transcripts to identify bacterial targets for potential new antibacterial agents.
Streptococcus pneumoniae is an important cause of community-acquired pneumonia and a major cause of otitis media, sinusitis, and meningitis in children and adults. In Spain and other Mediterranean countries, the majority of S. pneumoniae are relatively resistant to penicillin (Klugman, 1990; Fenoll et al., 1991; Jorgenserret al., 1990). These strains also have decreased susceptibility to broad-spectrum cephaloporins, which are frequently used in the empiric treatment of meningitis and
other serious invasive bacterial infections. High-level resistance of pneumococci has been encountered in Hungary where 10% of children who were colonized with S. pneumoniae carried penicillin resistant strains that were also resistant to tetracycline, erythromycin, trimethoprim/sulfamethoxazole, and 30% resistant to chloramphenicol (Neu, 1992). The resistance of pneumococci to macrolides such as erythromycin averages 20-25% in France, -20% in Japan, and <10% in Spain (Neu, 1992).
The antimicrobial susceptibilities and distribution of serotypes of the 42 isolates of S. pneumoniae in southern Taiwan from invasive infections have been recently determined (Hseuh et al., 1996). Resistance rates among these isolates were: erythromycin, 61.9%; clindamycin, 47.6%; chloramphenicol, 19%>; and tetracycline, 73.8%). Resistance to three or more classes of antibiotics was found in 33.3% of the isolates. Bacteremic pneumonia and primary bacteremia accounted for 64.3%> of the infections and mortality was 42.6%>. Given the severity of these infections despite adequate antibiotic therapy, there is clearly a need for introduction of new therapeutic options to prevent mortality due to invasive S. pneumoniae infections.
Pneumococcal phages belong to four families and they present a great variety in morphology, including lytic and temperate phages (for a review, see Garcia et al., 1997). Examples of lytic phages are Cρ-1 and Dp-1, whereas examples of temperate phages are HB-3, EJ-1, and HB-746. The complete nucleotide sequence and functional organization of Cp-1 has been reported (Martin et al., 1996). Cp-1 has a 19,345 bp double-stranded DNA genome, with a terminal protein covalently linked to its 5' ends, that replicates by a protein primed mechanism. The phage contains 29 ORFs, 23 on one strand and 6 on the opposite. When these predicted proteins were compared to sequences compiled in GenBank EMBL databases, to ORFs showed significant similarity to proteins of bacteriophage 29 that infects B. subtilis (Martin et al., 1996). The similar proteins corresponded to those involved in DNA replication (terminal protein and DNA polymerase), structural and morphogenic proteins (major head, collar, connector, tail, and encapsidation proteins), and proteins involved in lysis function (holin and lysozyme). In its strategy of lysis, the holin gene product inserts itself into the cell membrane, allowing access of the lysozyme to the peptidoglycan.. _ Expression of the Cp-1 holin protein in E. coli results in cell death after 2- hours of induction, but did not lead to lysis (Garcia et al., 1997). Cells harboring a plasmid construction with holin and lysozyme genes together did lyse after induction and the
viability loss was similar to that of the culture expressing holin alone. Cloning of these lytic genes in S. pneumoniae showed that both genes had the same effect as in E. coli. That is, holin itself did not lyse the culture but the viability loss was noticeable, whereas both holin and lysozyme together were capable of lysing M31, an amidase deleted mutant (Garcia et al., 1997).
Recently, a small portion (~4 kbp) of a second S. pneumoniae phage, Dp-1, has been sequenced (Sheehan et al., 1997). This portion contains the genes coding for the lytic system (Sheehan et al., 1997) and shows a modular organization similar to that described for Cp-1. However, in this case, a single chimeric protein appears to be made in which the N-terminal domain is highly similar to that of the murein hydrolase coded by a gene found in the phage BK5-T that infects Lactococcus lactis, and the C- terminal domain is homologous to holins. Thus, both functions appear to have been combined in a novel chimeric protein.
Bacteriophage Dp-1 was obtained from Dr. P. Garcia (Departamento de Microbiologia Molecular, Centro de Departamento de Investigaciones Biologicas, Consejo Superior de Investigaciones Cientificas, Velazquez, Madrid, Spain). We found that Dp-1 has a double-stranded DNA genome of 56,506 bp, predicted to encode 85 ORFs greater than 33 amino acids and with upstream Shine-Dalgarno motifs for translation initiation (Tables 28 & 30, and Fig. 6). Computational analysis of the predicted protein products of Streptococcus bacteriophage Dp-1 protein products, which detected homologs in public databases, are listed inTable 31, along with the accompanying list of related proteins.
From this analysis, it is apparent that several predicted genes of Dp-1 encode polypeptides that are related to structural proteins. ORFs 001, 002, 004, and 030 are predicted to encode tail proteins, minor structural proteins, and minor capsid proteins (Table 31). We also note the identification of several gene products that are likely involved in DNA synthesis. These include ORF 3 which encodes DNA polymerase, ORF 8 which encodes a S WI/SNF helicase-related protein, ORF 10 encodes a protein showing homology to recA, and ORF 13 encodes a dnaZX-like ORF. In E. coli, RapA encodes an RNA polymerase (RNAP)-associated protein with -
ATPase activity and which is a homolog of the eukaryotic SWI/SNF family, a set of proteins whose members are involved are involved in transcription activation, nucleosome remodeling, and DNA repair. RapA forms a stable complex with RNAP,
as if it were a subunit of RNAP and it is possible that the ORF 8 product behaves similarly or in a dominant-negative fashion to inhibit the activity of RapA. Mutation of the essential E. coli dnaZX results in a block in DNA chain elongation during replication (Maki et al., 1988). The dnaZX gene has only one open reading frame for a 71 -kDa polypeptide from which the two distinct DNA polymerase III holoenzyme subunits, tau (71 kDa) and gamma (47 kDa), are produced. The tau subunit is the precursor of the gamma subunit, and the gamma subunit is produced by a -1 frameshift causing early termination of translation (Tsuchihashi et al., 1990). These proteins show single-strand DNA binding properties that is ATPase (and dATPase) dependent and are thought to increasing the processivity of the core DNA polymerase enzyme (Lee et al., 1987).
There are several Dp-1 ORFs which encode proteins predicted to play a role in cellular metabolic pathways. These include polypeptides involved in coenzyme PQQ synthesis (ORFs 20, 29, 38). Pyrrolo-quinoline quinone (PQQ) is the non-covalently bound prosthetic group of many quinoproteins catalysing reactions in the periplasm of Gram-negative bacteria. Most of these involve the oxidation of alcohols or aldose sugars. Interestingly, ORFs 20, 29, and 30 also show homology to the exoenzyme S regulon (Frank, 1997). Proteins encoded by the P. aeruginosa exoenzyme S regulon may be involved in a contact-mediated translocation mechanism to transfer anti-host factors directly into eukaryotic cells disrupting eukaryotic signal transduction through ADP-ribosylation (Frank, 1997).
There is also a protein with similarity to GTP cyclohydrolase I (ORF 21) and ORF 41 which shows homology to dUTPase (Table 31). GTP cyclohydrolase I is an enzyme that catalyzes the first reaction in the pathway for the biosynthesis of the pteridine, a cofactor of the monooxygenases of the aromatic amino acids. Disruption of the homologous gene in Saccharomyces cerevisiae leads to a recessive conditional lethality due to folinic acid auxotrophy, that can be complemented with the mammalian or bacterial GTP cyclohydrolase I enzymes (Nardese et al., 1996; Mancini et al., 1999). ORF 16 shows high homology to autolysin. This region of the phage sequence was previously reported (Sheehan et al., 1997) and encompasses ~ 4 kbp of our sequence. The sequence published by (Sheehan et al., 1997) is shown in Table 32.
Thus, the present invention provides a nucleic acid sequence obtained from
Streptococcus bacteriophage Dp-1 comprising at least a portion of a phage Dp-1 QRF; - preferably an inhibitory ORF, and more preferably at least a portion of one of the genes described above with anti-microbial activity. For example, ORF 013 encodes a
protein with homology to the gamma subunit of DNA polymerase (dnaX gene). This protein may act in a dominant-negative fashion to sequester the host DNA polymerase for its own replication, thus inhibiting host DNA replication. The dnaX gene product is essential for E. coli replication (Kodaira et al., 1983). In certain preferred embodiments of the present invention, the bacterial target of a bacteriophage inhibitor ORF product, e.g., an inhibitory protein or polypeptide, is encoded by a Streptococcus nucleic acid coding sequence from a host bacterium for bacteriophage Dp-1. As above, possible target sequences are described herein by reference to sequence source sites. The sequence encoding the target preferably corresponds to a Streptococcus nucleic acid sequence available from The Institute for Genomic Research (TIGR), or available from GenBank or other public database. The TIGR Streptococcus sequences are publicly available at The Institute for Genomics Research at URL: http://www.tigr.org
The amino acid sequence of a polypeptide target is readily provided by translating the corresponding coding region. For the sake of brevity, the sequences are not reproduced herein. Also, in preferred embodiments, a target sequence corresponds to a Streptococcus pneumoniae coding sequences corresponding to a sequence listed in Table 33 herein. Sequences for other Streptococcal species are also available from TIGR and./or from GenBank. The listing in Table 33 describes Streptococcus sequences currently deposited in GenBank. Again, for the sake of brevity, the sequences are described by reference to the GenBank entries instead of being written out in full herein. In cases where the TIGR or GenBank entry for a coding region is not complete, the complete sequence can be readily obtained by routine methods, e.g., by isolating a clone in a phage Dp-1 host Streptococcus sp. genomic library, and sequencing the clone insert to provide the relevant coding region. The boundaries of the coding region can be identified by conventional sequence analysis and/or by expression in a bacterium in which the endogenous copy of the coding region has been inactivated and using subcloning to identify the functional start and stop codons for the coding region. In the various aspects of this invention involving Dp-1 sequences, preferably the sequence is preferably not contained in the sequence described in Sheehan et al., 1997 (Table 32).
Validating Identified Inhibitory Phage ORFs _ — "" A fifth step involves validating the identified phage inhibitor ORF by independent methods, and delineating further possible smaller segments of the ORFs
that have inhibitory activity. Several methods exist to validate the role of the identified ORF as an inhibitor ORF.
One example utilizes the creation of a mutant variant of the phage ORF in which the candidate ORF carries a partial or complete loss-of-function mutation that is measurable as compared with the non-mutant ORF. Comparison of the effects of expression of the loss of function mutant with the normal ORF provides confirmation of the identification of an inhibitor ORF where the loss-of-function mutant provides a measurably lower level of inhibition, preferably no inhibition. The loss of function may be conditional, e.g., temperature sensitive. Once validation of the inhibitor ORF is achieved, a bi-directional deletion analysis can be carried out using the same experimental system to identify the minimal polypeptide segment that has inhibitor activity. This may be carried out by a variety of means, e.g., by exonuclease or PCR methodologies, and is used to determine if a relatively small segment of the ORF (i.e., the product of the ORF) still possesses inhibitory activity when isolated away from its native sequence. If so, a portion of the ORF encoding this "active portion" can be used as a template for the synthesis of novel anti-microbial agents and further allowing derivation of the peptide sequence, e.g., using modified peptides and or peptidomimetics.
In creation of certain peptidomimetics, the peptide backbone is transformed into a carbon-based hydrophobic structure that can retain inhibitor activity against the bacterium. This is done by standard medicinal chemistry methods, typically monitored by measuring growth inhibition of the various molecules in liquid cultures or on solid medium. These mimetics can also represent lead compounds for the development of novel antibiotics. Recently, a major effort has been undertaken by the pharmaceutical industry and their biotechnology partners for the sequencing of bacterial pathogen genomes. The rationale is that the systematic sequencing of the genome will identify all of the bacterial proteins and therefore this proteome will be the target for designing novel inhibitor antibiotics. Although systematic, this approach has several major problems. The first is that analysis of primary amino acid sequences of bacterial proteins does not immediately reveal which protein will be essential for viability of the bacterium, and target validation is thus a major issue. The second problem is one of redundancy, as several biochemical pathways are either structurally duplicated in bacteria (different iso forms of the same enzyme), or functionally duplicated by the presence of salvage pathways in the event of a metabolic block in one pathway (different nutritional conditions). The third is that even a valid target may not be structurally or
functionally amenable to inhibition by small molecules because of inaccessibility (sequestration of target).
Therefore, there is considerable interest within the pharmaceutical and biotechnology industry in identifying key targets for drug discovery amongst the mass of novel targets generated by large-scale genomic sequencing projects.
On the other hand, and underscoring the instant invention, the phages herein described have, over millions of years, evolved specific mechanisms to target such key biochemical pathways and proteins. In the few cases where inhibition by phages has been elucidated (e.g., see ref. 3), such bacterial targets are invariably rate-limiting in their respective biochemical pathways, are not redundant, and/or are readily accessible for inhibition by the phage (or by another inhibitory compound). Therefore, the sixth step of this invention involves identifying the host biochemical pathways and proteins that are targeted by the phage inhibitory mechanisms.
Identifying. Validating, and Characterizing Bacterial Host Target Proteins and Affected Pathways
A rationale for this step is that the inhibitor ORF product from the phage physically interacts with and/or modifies certain microbial host components to block their function. Exemplary approaches which can be used to identify the host bacterial pathways and proteins that interact with, and preferably also are inhibited by, phage ORF product(s) are described below.
One approach is a genetic screen to determine physiological protein:protein interaction, for example, using a yeast two hybrid system. In this assay, the phage ORF is fused to the carboxyl terminus of the yeast Gal4 activation domain II (amino acids 768-881) to create a bait vector. A cDNA library of cloned S. aureus sequences which have been engineered into a plasmid where the S. aureus sequences are fused to the DNA binding domain of Gal4 is also generated. These plasmids are introduced alone, or in combination, into yeast strain Y190 - previously engineered with chromosomally integrated copies of the E. coli lacZ and the selectable HIS3 genes, both under Gal4 regulation (Durfee, T., Becherer, K., Chen, P.-L., Yeh, S.-H., Yang, Y., Kilburn, A.E., Lee, W.-H., and Elledge, S J. (1993). Genes & Dev. 1, 555-569). If the two proteins expressed in yeast interact, the resulting complex will activate transcription from promoters containing Gal4 binding sites. A lacZ and His3 gene, each driven by a promoter containing Gal4 binding sites, have been integrated into the. . genome of the host yeast system used for measuring protein-protein interactions. Such a system provides a physiological environment in which to detect potential protein interactions. This system has been extensively used to identify novel protein-protein
interaction partners and to map the sites required for interaction (for example, to identify interacting partners of translation factors (Qiu, H., Garcia-Barrio, M.T., and Hinnebusch, A.G. (1998). Mol & Cell Biology 18, 2697-2711), transcription factors (Katagiri, T., Saito, H., Shinohara, A., Ogawa, H., Kamada, N., Nakamura ,Y., and Miki, Y. (1998). Genes, Chromosomes & Cancer 21, 217-222), and proteins involved in signal transduction (Endo, T.A., Masuhara, M., Yokouchi, M., Suzuki, R., Sakamoto, H., Mitsui, K., Matsumoto, A., Tanimura, S., Ohtsubo, M., Misawa, H., Miyazaki, T., Leonor N., Taniguchi, T., Fujita, T., Kanakura, Y., Komiya, S., and Yoshimura, A. Nature. 387, 921-924). This approach has also been used in many published reports to identify interaction between mammalian viral and mammalian cell proteins.
For example, the non-structural protein NS1 of parvovirus is essential for viral DNA amplification and gene expression and is also the major cytopathic effector of these viruses. A yeast two-hybrid screen with NS 1 identified a novel cellular protein of unknown function that interacts with NS- 1 , called SGT, for small glutamine-rich tetratricopeptide repeat (TPR)-containing protein (Cziepluch C. Kordes E. Poirey R. Grewenig A. Rommelaere, J, and Jauniaux JC. (1998) J Virol. 72, 4149-4156). In another screen, the adenovirus E3 protein was recently shown to interact with a novel tumor necrosis factor alpha-inducible protein and to modulate some of the activities of E3 (Li Y. Kang J. and Horwitz M.S. (1998). Mol & Cell Biol. 18, 1601-1610). In yet another recent screen, the herpes simplex virus 1 alpha regulatory protein ICP0 was found to interact with (and stabilize) the cell cycle regulator cyclin D3 (Kawaguchi Y. Van Sant C. and Roizman B. (1997). J Virol. 71,7328-7336).
Another two-hybrid system for identifying proteimprotein interactions is commercially available from STRATEGENE™ as the CYTO-TRAP™ system (Chang et al., Strategies Newsletter 11(3), 65-68 (1998)(from Stratagene)). The system is a yeast-based method for detecting proteimprotein interactions in vivo, using activation of the Ras signal transduction cascade by localizing a signal pathway component, human Sos (hSos), to its activation site in the yeast plasma membrane. The system uses a temperature-sensitive Saccharomyces cerevisiae mutant, strain cdc25H, which contains a point mutation at amino acid residue 1328 of the cdc25 gene. This gene encodes a guanyl nucleotide exchange factor which binds and activates Ras, leading to cell growth. The mutation in the cdc25 gene prevents host growth at 37°C, but at a permissive temperature of 25°C, growth is normal. The system utilizes the ability of (hSos) to complement the cdc25 defect and activate Fhe yeast Ras signaling pathway. Once (hSos) is expressed and localized to the plasma membrane, the cdc25H yeast strain grows at 37°C. Localizing hSos to the plasma
membrane occurs through a protei protein interaction. A protein of interest, or bait, is expressed as a fusion protein with hSos. The library, or target proteins are expressed with the myristylation membrane-localization signal. The yeast cells are then incubated under restrictive conditions (37°C). If the bait and the target protein interact, the hSos protein is recruited to the membrane, activating the Ras signaling pathway and allowing the cdc25H yeast strain to grow at the restrictive temperature.
The protein targets of phage inhibitory ORFs can also be identified using bacterial genetic screens. One approach involves the overexpression of a phage inhibitory protein in mutagenized bacterial host species, followed by plating the cells and searching for colonies that can survive the antimicrobial activity of the inhibitory ORF. These colonies are then grown, their DNA extracted, and cloned into an expression vector that contains a replicon of a different incompatibility group from the plasmid expressing the original ORF. This library is then introduced into a wild- type host bacterium in conjunction with an expression vector driving synthesis of the phage ORF, followed by selection for surviving bacteria. Thus, bacterial DNA fragments from the survivors presumably contain a DNA fragment from the original mutagenized host bacterial genome that can protect the cell from the antimicrobial activity of the inhibitory phage ORF. This fragment can be sequenced and compared with that of the bacterial host to determine in which gene the mutation lies. This approach enables one to determine the targets and pathways that are affected by the killing function.
A second approach is based on identifying proteimprotein interactions between the phage ORF product and bacterial S. aureus, e.g., proteins using a biochemical approach based, for example, on affinity chromatography. This approach has been used, for example, to identify interactions between lambda phage proteins and proteins from their E. coli host (Sopta, M., Carthew, R.W., and Greenblatt, J. (1985) J. Biol. Chem. 260, 10353-10369). The phage ORF is fused to a peptide tag (e.g. glutathione-S-transferase ("GST"), 6xHIS, ("HIS") and/or calmodulin binding protein ("CPB")) within a commercially available plasmid vector that directs high level expression on induction of a suitably responsive promoter driving the fusion's expression. The translated fusion protein is expressed in E. coli, purified, and immobilized on a solid phase matrix via, for example the tag. Total cell extracts from the host bacterium, e.g., S. aureus, are then passed through the affinity matrix containing the immobilized phage ORF fusion protein; host proteins retained on the . . column are then eluted under different conditions of ionic strength, pH, detergents etc., and characterized by gel electrophoresis and other techniques. Appropriate controls are run to guard against nonspecific binding to the resin. Target proteins thus
recovered should be enriched for the phage protein/peptide of interest and are subsequently electrophoretically or otherwise separated, purified, sequenced, or biochemically analyzed. Usually sequencing entails individual digestion of the proteins to completion with a protease (e.g.-trypsin), followed by molecular mass and amino acid composition and sequence determination using, for example, mass spectrometry, e.g., by MALDI-TOF technology (Qin, J., Fenyo, D., Zhao, Y., Hall, W.W., Chao, D.M., Wilson, C.J., Young, R.A. and Chait, B.T. (1997). Anal. Chem. 69, 3995-4001).
The sequence of the individual peptides from a single protein are then analyzed by the bioinformatics approach described above to identify the S. aureus protein interacting with the phage ORF. This analysis is performed by a computer search of the S. aureus genome for an identified sequence. Alternatively, all tryptic peptide fragments of the S. aureus genome can be predicted by computer software, and the molecular mass of such fragments compared to the molecular mass of the peptides obtained from each interacting protein eluted from the affinity matrix. The responsible gene sequence can be obtained, for example by using synthetic degenerate nucleic acid sequences to pull out the corresponding homologous bacterial sequence. Alternatively, antibodies can be generated against the peptide and used to isolate nascent peptide/mRNA transcript complexes, from which the mRNA can be reverse transcribed, cloned, and further characterized using the procedures discussed herein. A variety of other binding assay methods are known in the art and can be used to identify interactions between phage proteins and bacterial proteins or other bacterial cell components. Such methods that allow or provide identification of the bacterial component can be used in this invention for identifying putative targets. Validation of the interaction between the phage ORF product and the bacterial proteins or other components can be obtained by a second independent assay (e.g., co-immunoprecipitation or protein-protein crosslinking experiments (Qiu, H., Garcia- Barrio, M.T., and Hinnebusch, A.G. (1998). Mol & Cell Biology 18, 2697-2711 ; Brown, S. and Blumenthal, T. (1976). Proc. Natl. Acad. Sci. USA 73, 1131-1135)). Finally, the essential nature of the identified bacterial proteins is preferably determined genetically by creating a constitutive or inducible partial or complete loss- of-function mutation in the gene encoding the identified interacting bacterial protein. This mutant is then tested for bacterial survival and replication.
The protein target of the phage inhibitor function can also be identified using a. _ genetic approach. Two exemplary approaches will be delineated here. The first ~ approach involves the overexpression of a predetermined phage inhibitor protein in mutagenized host bacteria, e.g., S. aureus, followed by plating the cells and searching
for colonies that can survive the inhibitor. These colonies will then be grown, their DNA extracted and cloned into an expression vector that contains a replicon of a different incompatibility group, and preferably having a different selectible marker than the plasmid expressing the phage inhibitor. Thus, host DNA fragments from the mutant that can protect the cell from phage ORF inhibition can be sequenced and compared with that of the bacterial host to determine in which gene the mutation lies. This approach allows rapid determination of the targets and pathways that are affected by the inhibitor.
Alternatively, the bacterial targets can be determined in the absence of selecting for mutations using an approach known as "multicopy suppression". In this approach, the DNA from the wild type host is cloned into an expression vector that can coexist, as previously described, with one containing a predetermined phage inhibitor. Those plasmids that contain host DNA fragments and genes that protect the host from the phage inhibitor can then be isolated and sequenced to identify putative targets and pathways in the host bacteria.
Regardless of the specific mode of identification, screening assays may additionally utilize gene fusions to specific "reporter genes" to identify a bacterial gene(s) whose expression is affected when the host target pathway is affected by the phage inhibitor. Such gene fusions can be used to search a number of small molecule compounds for inhibitors that may affect this pathway and thus cause cell inhibition. This approach will allow the screening of a large number of molecules on petri dishes or 96-well format by monitoring for a simple color change in the bacterial colonies. In this manner, we can validate host targets and classes of compounds for further study and clinical development. These inhibitors also represent lead compounds for the development of other antibiotics.
Bioinformatics and comparative genomics are preferably then applied to the identified bacterial gene products to predict biochemical function. The biochemical activity of the protein can be verified in vitro in cell free assays or in vivo in intact cells. In vitro biochemical assays utilizing cell-free extracts or purified protein are established as a basis for the screening and development of inhibitors.
These inhibitors, preferably small molecule inhibitors, may comprise peptides, antibodies, products from natural sources such as fungal or plant extracts or small molecule organic compounds. In general, small molecule organic compounds are preferred. These compounds may, for example, be identified within large compound libraries, including combinatorial libraries. For example, a plurality of compounds, preferably a large number of compounds can be screened to determine whether any of the compounds binds or otherwise disrupts or inhibits the identified bacterial target.
Compounds identified as having any of these activities can then be evaluated further in cell culture and/or animal model systems to determine the pharmacological properties of the compound, including the specific anti-microbial ability of the compound. For mixtures of natural products, including crude preparations, once a preparation or fraction of a preparation is shown the have an anti-microbial activity, the active substance can be isolated and identified using techniques well known in the art, if the compound is not already available in a purified form.
Identified compounds possessing anti-microbial activity and similar compounds having structural similarity can be further evaluated and, if necessary, derivatized according to synthesis and/or modification methods available in the art selected as appropriate for the particular starting molecule.
Derivatization of identified anti-microbials In cases where the identified anti-microbials above might represent peptidal compunds, the in vivo effectiveness of such compounds may be advantageously enhanced by chemical modification using the natural polypeptide as a starting point and incorporating changes that provide advantages for use, for example, increased stability to proteolytic degradation, reduced antigenicity, improved tissue penetration, and/or improved delivery characteristics.
In addition to active modifications and derivative creations, it can also be useful to provide inactive modifications or derivatives for use as negative controls or introduction of immunologic tolerance. For example, a biologically inactive derivative which has essentially the same epitopes as the corresponding natural antimicrobial can be used to induce immunological tolerance in a patient being treated. The induction of tolerance can then allow uninterrupted treatment with the active anti-microbial to continue for a significantly longer period of time.
Modified anti-microbial polypeptides and derivatives can be produced using a number of different types of modifications to the amino acid chain. Many such methods are known to those skilled in the art. The changes can include, for example, reduction of the size of the molecule, and/or the modification of the amino acid sequence of the molecule. In addition, a variety of different chemical modifications of the naturally occurring polypeptide can be used, either with or without modifications to the amino acid sequence or size of the molecule. Such chemical modifications can, for example, include the incoφoration of modified or non-natural amino acids or^iόn- amino acid moieties during synthesis of the peptide chain, or the post-synthesis modification of incoφorated chain moieties.
The oligopeptides of this invention can be synthesized chemically or through an appropriate gene expression system. Synthetic peptides can include both naturally occurring amino acids and laboratory synthesized, modified amino acids.
Also provided herein are functional derivatives of anti-microbial proteins or polypeptides. By "functional derivative" is meant a "chemical derivative,"
"fragment," "variant," "chimera," or "hybrid" of the polypeptide or protein, which terms are defined below. A functional derivative retains at least a portion of the function of the protein, for example reactivity with a specific antibody, enzymatic activity or binding activity. A "chemical derivative" of the complex contains additional chemical moieties not normally a part of the protein or peptide. Such moieties may improve the molecule's solubility, absoφtion, biological half-life, and the like. The moieties may alternatively decrease the toxicity of the molecule, eliminate or attenuate any undesirable side effect of the molecule, and the like. Moieties capable of mediating such effects are disclosed in Alfonso and Gennaro (1995). Procedures for coupling such moieties to a molecule are well known in the art. Covalent modifications of the protein or peptides are included within the scope of this invention. Such modifications may be introduced into the molecule by reacting targeted amino acid residues of the peptide with an organic derivatizing agent that is capable of reacting with selected side chains or terminal residues, as described below.
Cysteinyl residues most commonly are reacted with alpha-haloacetates (and corresponding amines), such as chloroacetic acid or chloroacetamide, to give carboxymethyl or carboxyamidomethyl derivatives. Cysteinyl residues also are derivatized by reaction with bromotrifluoroacetone, chloroacetyl phosphate, N- alkylmaleimides, 3-nitro-2-pyridyl disulfide, methyl 2-pyridyl disulfide, p-chloro- mercuribenzoate, 2-chloromercuri-4-nitrophenol, or chloro-7-nitrobenzo-2-oxa-l,3- diazole.
Histidyl residues are derivatized by reaction with diethylprocarbonate at pH 5.5-7.0 because this agent is relatively specific for the histidyl side chain. Para- bromophenacyl bromide also is useful; the reaction is preferably performed in 0.1 M sodium cacodylate at pH 6.0.
Lysinyl and amino terminal residues are reacted with succinic or other carboxylic acid anhydrides. Derivatization with these agents has the effect of reversing the charge of the lysinyl residues. Other suitable reagents for derivatizing primary amine- containing residues include imidoesters such as methyl _ picolinimidate; pyridoxal phosphate; pyridoxal; chloroborohydride;
trinitrobenzenesulfonic acid; O-methylisourea; 2,4 pentanedione; and transaminase- catalyzed reaction with glyoxylate.
Arginyl residues are modified by reaction with one or several conventional reagents, among them phenylglyoxal, 2,3-butanedione, 1,2-cyclohexanedione, and ninhydrin. Derivatization of arginine residues requires that the reaction be performed in alkaline conditions because of the high p , of the guanidine functional group. Furthermore, these reagents may react with the groups of lysine as well as the arginine alpha-amino group.
Tyrosyl residues are well-known targets of modification for introduction of spectral labels by reaction with aromatic diazonium compounds or tetranitromethane. Most commonly, N-acetylimidizol and tetranitromethane are used to form O-acetyl tyrosyl species and 3-nitro derivatives, respectively.
Carboxyl side groups (aspartyl or glutamyl) are selectively modified by reaction carbodiimide (R'-N-C-N-R') such as l-cyclohexyl-3-(2-moφholinyl(4-ethyl) carbodiimide or l-ethyl-3-(4-azonia-4,4-dimethylpentyl) carbodiimide. Furthermore, aspartyl and glutamyl residues are converted to asparaginyl and glutaminyl residues by reaction with ammonium ions.
Glutaminyl and asparaginyl residues are frequently deamidated to the corresponding glutamyl and aspartyl residues. Alternatively, these residues are deamidated under mildly acidic conditions. Either form of these residues falls within the scope of this invention.
Derivatization with bifunctional agents is useful, for example, for cross- linking component peptides to each other or the complex to a water-insoluble support matrix or to other macromolecular carriers. Commonly used cross-linking agents include, for example, 1,1 -bis (diazoacetyl)-2-phenylethane, glutaraldehyde, N- hydroxysuccinimide esters, for example, esters with 4-azidosalicylic acid, homobi- functional imidoesters, including disuccinimidyl esters such as 3,3'- dithiobis(succinimidylpropionate), and bifunctional maleimides such as bis-N- maleimido-l,8-octane. Derivatizing agents such as methyl-3-[p-azidophenyl) dithiolpropioimidate yield photoactivatable intermediates that are capable of forming crosslinks in the presence of light. Alternatively, reactive water-insoluble matrices such as cyanogen bromide-activated carbohydrates and the reactive substrates described in U.S. Patent Nos. 3,969,287; 3,691,016; 4,195,128; 4,247,642; 4,229,537; and 4,330,440 are employed for protein immobilization. Other modifications include hydroxylation of proline and lysine, phosphorylation of hydroxyl groups of seryl or threonyl residues, methylation of the alpha-amino groups of lysine, arginine, and histidine side chains (Creighton, T.E.,
Proteins: Structure and Molecular Properties, W.H. Freeman & Co., San Francisco, pp. 79-86 (1983)), acetylation of the N-terminal amine, and, in some instances, amidation of the C-terminal carboxyl groups.
Such derivatized moieties may improve the stability, solubility, absoφtion, biological half life, and the like. The moieties may alternatively eliminate or attenuate any undesirable side effect of the protein complex. Moieties capable of mediating such effects are disclosed, for example, in Alfonso and Gennaro (1995).
The term "fragment" is used to indicate a polypeptide derived from the amino acid sequence of the protein or polypeptide having a length less than the full-length polypeptide from which it has been derived. Such a fragment may, for example, be produced by proteolytic cleavage of the full-length protein. Preferably, the fragment is obtained recombinantly by appropriately modifying the DNA sequence encoding the proteins to delete one or more amino acids at one or more sites of the C-terminus, N-terminus, and or within the native sequence. Another functional derivative intended to be within the scope of the present invention is a "variant" polypeptide that either lacks one or more amino acids or contains additional or substituted amino acids relative to the native polypeptide. The variant may be derived from a naturally occurring polypeptide by appropriately modifying the protein DNA coding sequence to add, remove, and/or to modify codons for one or more amino acids at one or more sites of the C-terminus, N-terminus, and/or within the native sequence.
A functional derivative of a protein or polypeptide with deleted, inserted and/or substituted amino acid residues may be prepared using standard techniques well-known to those of ordinary skill in the art. For example, the modified components of the functional derivatives may be produced using site-directed mutagenesis techniques (as exemplified by Adelman et al., 1983, DNA 2:183; Sambrook et al., 1989) wherein nucleotides in the DNA coding sequence are modified such that a modified coding sequence is produced, and thereafter expressing this recombinant DNA in a prokaryotic or eukaryotic host cell, using techniques such as those described above. Alternatively, components of functional derivatives of complexes with amino acid deletions, insertions and/or substitutions may be conveniently prepared by direct chemical synthesis, using methods well-known in the art.
Insofar as other anti-microbial inhibitor compounds identified by the invention _ described herein may not be peptidal in nature, other chemical techniques exisftδ allow their suitable modification, as well, and according the desirable principles discussed above.
Administration and Pharmaceutical Compositions
For the therapeutic and prophylactic treatment of infection, the preferred method of preparation or administration of anti-microbial compounds will generally vary depending on the precise identity and nature of the anti-microbial being delivered. Thus, those skilled in the art will understand that administration methods known in the art will also be appropriate for the compounds of this invention.
The particularly desired anti-microbial can be administered to a patient either by itself, or in pharmaceutical compositions where it is mixed with suitable carriers or excipient(s). In treating an infection, a therapeutically effective amount of an agent or agents is administered. A therapeutically effective dose refers to that amount of the compound that results in amelioration of one or more symptoms of bacterial infection and or a prolongation of patient survival or patient comfort.
Toxicity, therapeutic and prophylactic efficacy of anti-microbials can be determined by standard pharmaceutical procedures in cell cultures and/or experimental organisms such as animals, e.g., for determining the LD50 (the dose lethal to 50%> of the population) and the ED50 (the dose therapeutically effective in 50%) of the population). The dose ratio between toxic and therapeutic effects is the therapeutic index and it can be expressed as the ratio LDJ0/ED50. Compounds that exhibit large therapeutic indices are preferred. The data obtained from these cell culture assays and animal studies can be used in formulating a range of dosage for use in humans. The dosage of such compounds lies preferably within a range of circulating concentrations that include the ED50 with little or no toxicity. The dosage may vary within this range depending upon the dosage form employed and the route of administration utilized.
For any compound identified and used in the method of the invention, the therapeutically effective dose can be estimated initially from cell culture assays. Such information can be used to more accurately determine useful doses in organisms such as plants and animals, preferably mammals, and most preferably humans. Levels in plasma may be measured, for example, by HPLC or other means appropriate for detection of the particular compound.
The exact formulation, route of administration and dosage can be chosen by the individual physician in view of the patient's condition (see e.g. Fingl et. al., in The Pharmacological Basis of Therapeutics, 1975, Ch. 1 p.l). It should be noted that the attending physician would know how' and when to terminate, interrupt, or adjust administration due to toxicity, organ dysfunction, or other systemic malady. Conversely, the attending physician would also know to adjust treatment to higher levels if the clinical response were not adequate (precluding
toxicity). The magnitude of an administered dose in the management of the disorder of interest will vary with the severity of the condition to be treated and the route of administration. The severity of the condition may, for example, be evaluated, in part, by standard prognostic evaluation methods. Further, the dose and perhaps dose frequency, will also vary according to the age, body weight, and response of the individual patient. A program comparable to that discussed above also may be used in veterinary or phyto medicine.
Depending on the specific infection target being treated and the method selected, such agents may be formulated and administered systemically or locally, i.e., topically. Techniques for formulation and administration may be found in Alfonso and Gennaro (1995). Suitable routes may include , for example, oral, rectal, transdermal, vaginal, transmucosal, intestinal, parenteral, intramuscular, subcutaneous, or intramedullary injections, as well as intrathecal, intravenous, or intraperitoneal injections. For injection, the agents of the invention may be formulated in aqueous solutions, preferably in physiologically compatible buffers such as Hanks' solution, Ringer's solution, or physiological saline buffer. For transmucosal administration, penetrants appropriate to the barrier to be permeated are used in the formulation. Such penetrants are generally known in the art. Use of pharmaceutically acceptable carriers to formulate identified antimicrobials of the present invention into dosages suitable for systemic administration is within the scope of the invention. With proper choice of carrier and suitable manufacturing practice, the compositions of the present invention, in particular those formulated as solutions, may be administered parenterally, such as by intravenous injection. Appropriate compounds can be formulated readily using pharmaceutically acceptable carriers well known in the art into dosages suitable for oral administration. Such carriers enable the compounds of the invention to be formulated as tablets, pills, capsules, liquids, gels, syrups, slurries, suspensions and the like, for oral ingestion by a patient to be treated. Agents intended to be administered intracellularly may be administered using techniques well known to those of ordinary skill in the art. For example, such agents may be encapsulated into liposomes, then administered as described above. Liposomes are spherical lipid bilayers with aqueous interiors. All molecules present in an aqueous solution at the time of liposome formation are incoφorated into the aqueous interior. The liposomal contents are both protected from the external microenvironment and, because liposomes fuse with cell membranes, are efficiently
delivered into the cell cytoplasm. Additionally, due to their hydrophobicity, small organic molecules may be directly administered intracellularly.
Pharmaceutical compositions suitable for use in the present invention include compositions wherein the active ingredients are contained in an effective amount to achieve the intended puφose. Determination of the effective amounts is well within the capability of those skilled in the art.
In addition to the active ingredients, these pharmaceutical compositions may contain suitable pharmaceutically acceptable carriers comprising excipients and auxiliaries which facilitate processing of the active compounds into preparations which can be used pharmaceutically. The preparations formulated for oral administration may be in the form of tablets, dragees, capsules, or solutions, including those formulated for delayed release or only to be released when the pharmaceutical reaches the small or large intestine.
The pharmaceutical compositions of the present invention may be manufactured in a manner that is itself known, e.g., by means of conventional mixing, dissolving, granulating, dragee-making, levitating, emulsifying, encapsulating, entrapping or lyophilizing processes.
Pharmaceutical formulations for parenteral administration include aqueous solutions of the active anti-microbial compounds in water-soluble form. Alternatively, suspensions of the active compounds may be prepared as appropriate oily injection suspensions. Suitable lipophilic solvents or vehicles include fatty oils such as sesame oil, or synthetic fatty acid esters, such as ethyl oleate or triglycerides, or liposomes. Aqueous injection suspensions may contain substances which increase the viscosity of the suspension, such as sodium carboxymethyl cellulose, sorbitol, or dextran. Optionally, the suspension may also contain suitable stabilizers or agents which increase the solubility of the compounds to allow for the preparation of highly concentrated solutions.
Pharmaceutical preparations for oral use can be obtained by combining the active compounds with solid excipient, optionally grinding a resulting mixture, and processing the mixture of granules, after adding suitable auxiliaries, if desired, to obtain tablets or dragee cores. Suitable excipients are, in particular, fillers such as sugars, including lactose, sucrose, mannitol, or sorbitol; cellulose preparations such as, for example, maize starch, wheat starch, rice starch, potato starch, gelatin, gum tragacanth, methyl cellulose, hydroxypropylmethyl-cellulose, sodium carboxymethylcellulose, and/or polyvinylpyrrolidone (PVP). If desired, -, disintegrating agents may be added, such as the cross-linked polyvinyl pyrrolidone, agar, or alginic acid or a salt thereof such as sodium alginate.
Dragee cores are provided with suitable coatings. For this puφose, concentrated sugar solutions may be used, which may optionally contain gum arabic, talc, polyvinyl pyrrolidone, carbopol gel, polyethylene glycol, and/or titanium dioxide, lacquer solutions, and suitable organic solvents or solvent mixtures. Dyestuffs or pigments may be added to the tablets or dragee coatings for identification or to characterize different combinations of active compound doses.
Pharmaceutical preparations which can be used orally include push-fit capsules made of gelatin, as well as soft, sealed capsules made of gelatin and a plasticizer, such as glycerol or sorbitol. The push-fit capsules can contain the active ingredients in admixture with filler such as lactose, binders such as starches, and/or lubricants such as talc or magnesium stearate and, optionally, stabilizers. In soft capsules, the active compounds may be dissolved or suspended in suitable liquids, such as fatty oils, liquid paraffin, or liquid polyethylene glycols. In addition, stabilizers may be added. The above methodologies may be employed either actively or prophylactically against an infection of interest.
Computer-related Aspects and Embodiments
In addition to the provision of compounds as chemical entities, nucleotide sequences, or fragments thereof at least 95%, preferably at least 97%, more preferably at least 99%, and most preferably at least 99.9%> identical to phage inhibitor sequences can also be provided in a variety of additional media to facilitate various uses.
Thus, as used in this section, "provided" refers to an article of manufacture, rather than an actual nucleic acid molecule, which contains a nucleotide sequence of the present invention; e.g., a nucleotide sequence of an exemplary bacteriophage or a sequence encoding a bacterial target or a fragment thereof, preferably a nucleotide sequence at least 95%, more preferably at least 99%> and most preferably at least 99.9%o identical to such a bacteriophage or bacterial sequence, for example, to a polynucleotide of an unsequenced phage listed in Table 1, preferably of bacteriophage 77 (S. aureus host) or bacteriophage 3A (S.aureus host) or bacteriophage 96 (S. aureus host). Such an article provides a large portion of the particular bacteriophage genome or bacterial gene and parts thereof (e. ., a bacteriophage open reading frame (ORF)) in a form which allows a skilled artisan to examine and/or analyze the sequence using means not directly applicable to examining the actual genome or gene. _ or subset thereof as it exists in nature or in purified form as a chemical entity. In one application of this aspect, a nucleotide sequence of the present invention can be recorded on computer readable media. As used herein, "computer
readable media" refers to any medium that can be read and accessed directly by a computer. Such media include, but are not limited to: magnetic storage media, such as floppy discs, hard disc storage medium, magnetic tape; optical storage media such as CD-ROM; electrical storage media such as RAM and ROM; and hybrids of these categories, such as magnetic/optical storage media. A skilled artisan can readily appreciate how any of the presently known computer readable mediums can be used to create an article of manufacture which includes one or more computer readable media having recorded thereon a nucleotide sequence or sequences of the present invention. Likewise, it will be clear to those of skill how additional computer readable media that may be developed also can be used to create analogous manufactures having recorded thereon a nucleotide sequence of the present invention.
As used herein, "recorded" refers to a process for storing information on computer readable medium. A skilled artisan can readily adopt any of the presently known methods for recording information on computer readable medium to generate manufactures comprising the nucleotide sequence information of the present invention.
A variety of data storage structures are available to a skilled artisan for creating a computer readable medium having recorded thereon a nucleotide sequence of the present invention. The choice of the data storage structure will generally be based on the means chosen to access the stored information. In addition, a variety of data processor programs and formats can be used to store the nucleotide sequence information of the present invention on computer readable medium. The sequence information can, for example, be presented in a word processing test file, formatted in commercially available software such as WordPerfect and Microsoft Word, or represented in the form of an ASCII file, stored in a database application, such as DB2, Sybase, Oracle, or the like. A skilled artisan can readily adapt any number of data processor structuring formats (e.g., text file or database) in order to obtain computer readable medium having recorded thereon the nucleotide sequence information of the present invention. Computer software is publicly available which allows a skilled artisan to access sequence information provided in a computer readable medium. Thus, by providing in computer readable form a nucleotide sequence of an unsequenced bacteriophage, such as an exemplary bacteriophage listed in Table 1 or of a sequence encoding a bacterial target or a fragment thereof, preferably a nucleotide sequence at - - least 95%>, more preferably at least 99%> and most preferably at least 99.9%> identical to such a bacteriophage or bacterial sequence, for example, to a polynucleotide of bacteriophage 77 (S. aureus host) or bacteriophage 3A (S.aureus host) bacteriophage
96 (S. aureus host), bacteriophage 44 AHJD (S. aureus host), bacteriophage Dp-1 (Streptococcus pneumoniae host), or bacteriophage 182 (Enterococcus host) the present invention enables the skilled artisan to routinely access the provided sequence information for a wide variety of puφoses. Those skilled in the art understand that software can implement a variety of different search or analysis software which implement sequence search and analysis algorithms, e.g., the BLAST (Altschul et al., J. Mol. Biol. 215:403410 (1990) and BLAZE (Brutlag et al., Comp. Chem 17:203-207 (1993)) search algorithms. For example, such search algorithms can be implemented on a Sybase system and used to identify open reading frames (ORFs) within the bacteriophage genome which contain homology to ORFs or proteins from other viruses, e.g, other bacteriophage, and other organisms, e.g., the host bacterium. Among the ORFs discussed herein are protein encoding fragments of the bacteriophage genomes which encode bacteria-inhibiting proteins or fragments. The present invention further provides systems, particularly computer-based systems, which contain the sequence information described. Such systems are designed to identify, among other things, useful fragments of the bacteriophage genomes.
As used herein, "a computer-based system" refers to the hardware, software, and data storage media used to analyze the nucleotide sequence information of the present invention. The minimum hardware of the computer-based systems of the present invention comprises a central processing unit (CPU), input device, output device, and data storage medium or media. A skilled artisan will readily recognize that any of the currently available general puφose computer-based system are suitable for use in the present invention, as well as a variety of different specialized or dedicated computer-based systems.
As stated above, the computer-based systems of the present invention comprise data storage media having stored therein a nucleotide sequence of the present invention and the necessary hardware and software for supporting and implementing a search and/or analysis program.
As used herein, "data storage media" refers to memory which can store nucleotide sequence information of the present invention, or a memory access means which can access manufactures having recorded thereon the nucleotide sequence information of the present invention. As used herein, "search program" refers to one or more programs which are implemented on the computer-based system to compare a target sequence or target structural motif with the sequence information stored within the data storage means.
Search means are used to identify fragments or regions of the present gnomic sequences which match a particular target sequence or target motif. A variety of known algorithms are disclosed publicly and a variety of commercially available software for conducting search means are and can be used in the computer-based systems of the present invention. Examples of such software includes, but is not limited to, MacPattern (EMBL), BLASTN and BLASTX (NCBIA). A skilled artisan can readily recognize that any one of the available algorithms or implementing software packages for conducting homology searches and/or sequence analyses can be adapted for use in the present computer-based systems. As used herein in connection with sequence searches and analyses, a "target sequence" can be any DNA or amino acid sequence of six or more nucleotides or two or more amino acids. A skilled artisan can readily recognize that the longer a target sequence is, the less likely a target sequence will be present as a random occurrence in the database. Also, the target sequence length is preferably selected to include sequence corresponding to a biologically relevant portion of an encoded product, for example a region which is expected to be conserved across a range of source organisms. Preferably the sequence length of a target polypeptide sequence is from 5- 100 amino acids, more preferably 7-50 or 7-100 amino acids, and still more preferably 10-80 or 10-100 amino acids. Preferably the sequence length of a target polynucleotide sequence is from 15-300 nucleotide residues, more preferably from 21- 240 or 21-300, and still more preferably 30-150 or 30-300 nucleotide residues. However, it is well recognized that searches for commercially important fragments, such as sequence fragments involved in gene expression and protein processing, may be of shorter length. Likewise, it may be desirable to search and/or analyze longer sequences.
As used herein, "a target structural motif," or "target motif," refers to any rationally selected sequence or combination of sequences in which the sequence(s) are chosen based on a three-dimensional configuration which is formed upon the folding of the target motif. There are a variety of target motifs known in the art. Protein target motifs include, but are not limited to, enzymatic active sites and signal sequences. Nucleic acid target motifs include, but are not limited to promoter sequences, haiφin structures and inducible expression elements (protein binding sequences).
A variety of structural formats for the input and output devices can be used to_ input and output the information in the computer-based systems of the preser_r invention. A preferred format for an output device ranks fragments of the bacteriophage or bacterial sequences possessing varying degrees of homology to the
target sequence or target motif. Such presentation provides a skilled artisan with a ranking of sequences which contain various amounts of the target sequence or target motif and identifies the degree of homology contained in the identified fragment.
A variety of comparing methods and/or devices and/or formats can be used to compare a target sequence or target motif with the sequence stored in data storage media to identify sequence fragments of the bacteriophage or bacterium in question. One skilled in the art can readily recognize that any one of the publicly available homology search programs can be used as the search program for the computer-based systems of the present invention. Of course, suitable proprietary systems that may be known to those of skill, or later developed, also may be employed in this regard. Figure 6 provides a block diagram of a computer system illustrative of embodiments of this aspect of present invention. The computer system 102 includes a processor 106 connected to a bus 104. Also connected to the bus 104 are a main memory 108 (preferably implemented as random access memory, RAM) and a variety of secondary storage devices 110, such as a hard drive 112 and a removable medium storage device 114. The removable medium storage device 114 may represent, for example, a floppy disk drive, a CD-ROM drive, a magnetic tape drive, etc. A removable storage medium 116 (such as a floppy disk, a compact disk, a magnetic tape, etc.) containing control logic and/or data recorded therein may be inserted into the removable medium storage device 114. The computer system 102 includes appropriate software for reading the control logic and/or the data from the removable medium storage device 114, once it is inserted into the removable medium storage device 114.
A nucleotide sequence of the present invention may be stored in a well-known manner in the main memory 108, any of the secondary storage devices 110, and/or a removable storage medium 116. During execution, software for accessing and processing the sequence (such as search tools, comparing tools, etc.) reside in main memory 108, in accordance with the requirements and operating parameters of the operating system, the hardware system and the software program or programs. The data storage medium in which the sequence is embodied and the central processor need not be part of a single stand-alone computer, but may be separated so long as data transfer can occur. For example, the processor or processors being utilized for a search or analysis can be part of one general puφose computer, and the data storage medium can be part of a second general puφose computer connected to_a_ network, or the data storage medium can be part of a network server. As another example the data storage medium can be part of a computer system or network accessible over telephone lines or other remote connection method.
EXAMPLES
Example 1. Growth of Staph A bacteriophage 77 and purification of genomic DNA. The Staphylococcus aureus propagating strain (PS 77; ATCC #27699) was used as a host to propagate its respective phage 77 (ATCC # 27699-B1). Two rounds of plaque purification of phage 77 were performed on soft agar essentially as described in Sambrook et al (1989). Briefly, the PS 77 strain was grown overnight at 37°C in Nutrient broth [NB: 0.3% Bacto beef extract, 0.5% Bacto peptone (Difco Laboratories) and 0.5% NaCl (w/v)].The culture was then diluted 20x in NB and incubated at 37°C until the OD540= .2 (early log phase) with constant agitation. In order to obtain single plaques, phage 77 was subjected to 10-fold serial dilutions using phage buffer (1 mM MgSO4, 5 mM MgCl2, 80 mM NaCl and 0.1% Gelatin (w/v)) and 10 μl of each dilution was used to infect 0.5 ml of the cell suspension in the presence of 400 μg/ml CaCl2. After incubation of 15 min at room temperature (RT), 2 ml of melted soft agar kept at 45°C (NB supplemented with 0.6%» agar) was added to the mixture and poured onto the surface of 100 mm nutrient agar plates (0.3% Bacto Beef extract, 0.5% Bacto peptone, 0.5%> NaCl and 1.5% Bacto agar (w/v)). After overnight incubation at 30°C, a single plaque was isolated, resuspended in 1 ml of phage buffer by end over end rotation for 2 hrs at 20°C, and the phage suspension was diluted and used for a second infection as described above. After overnight incubation at 30°C, a single plaque was isolated and used as a stock.
The propagation procedure for bacteriophage 77 was modified from the agar layer method of Swanstδrm and Adams (1951). Briefly, the PS 77 strain was grown to stationary phase overnight at 37°C in Nutrient broth. The culture was then diluted twenty-fold in NB and incubated at 37°C until the OD540= .2. The suspension (15xl07 Bacteria) was then mixed with 15xl05 plaque forming units (pfu) to give a ratio of 100-bacteria/phage particle in the presence of 400 μg/ml of CaCl2. After incubation for 15 min at 20°C, 7.5 ml of melted soft agar (NB plus 0.6%> agar) were added to the mixture and poured onto the surface of 150 mm nutrient agar plates and incubated 16 hrs at 30°C. To collect the phage plate lysate, 20 ml of NB were added to each plate and the soft agar layer was collected by scrapping off with a clean microscope slide followed by shaking of the agar suspension for 5 min to break up the agar. The mixture was then centrifuged for 10 min at 4,000 RPM (2,830xg) in a JA-10 rotor- * ' (Beckman) and the supernatant fluid (lysate) was collected and subjected to~a treatment with 10 μg /ml of DNase I and RNase A for 30 min at 37°C. To precipitate the phage particles, the phage suspension was adjusted to 10%> (w/v) PEG 8000 and
0.5 M of NaCl followed by incubation at 4°C for 16 hrs. The phage was recovered by centrifugation at 4,000 φm (3,500xg) for 20 min at 4°C on a GS-6R table top centrifuge (Beckman). The pellet was resuspended with 2 ml of phage buffer (1 mM MgSO4, 5 mM MgCl2, 80 mM NaCl and 0.1 %> Gelatin). The phage suspension was extracted with 1 volume of chloroform and further purified by centrifugation on a cesium chloride step gradient as described in Sambrook et al. (1989), using a TLS 55 rotor centrifuged in an Optima TLX ultracentrifuge (Beckman) for 2 h at 28,000 φm (67,000xg) at 4°C. Banded phage was collected and ultracentrifuged again on an isopycnic cesium chloride gradient (1.45 g/ml) at 40,000 rpm (64,000xg) for 24 h at 4°C using a TLV rotor (Beckman). The phage was harvested and dialyzed for 4 h at room temperature against 4 L of dialysis buffer consisting of 10 mM NaCl, 50 mM Tris-HCl [pH 8] and 10 mM MgCl2. Phage DNA was prepared from the phage suspension by adding 20 mM EDTA, 50 mg/ml Proteinase K and 0.5% SDS and incubating for 1 h at 65°C, followed by successive extractions with 1 volume of phenol, 1 volume of phenol-chloroform and 1 volume of chloroform. The DNA was then dialyzed overnight at 4°C against 4 L of TE (10 mM Tris pH 8.0, lmM EDTA).
Example 2. DNA sequencing of Bacteriophage 77 genome
Four micrograms of phage 77 DNA was diluted in 200 μl of TE (10 mM Tris, [pH 8.0], 1 mM EDTA) in a 1.5 ml eppendorf tube and sonication was performed (550 Sonic Dismembrator™, Fisher Scientific). Samples were sonicated under an amplitude of 3 μm with bursts of 5 s spaced by 15 s cooling in ice/water for 3 to 4 cycles. The sonicated DNA was then size fractionated by electrophoresis on 1%> agarose gels utilizing TAE (1 x TAE is: 40 mM Tris-acetate, 1 mM EDTA [pH 8.0]) as the running buffer. Fractions ranging from 1 to 2 kbp were excised from the agarose gel and purified using a commercial DNA extraction system according to the instructions of the manufacturer (Qiagen), with a final elution of 50 μl of 1 mM Tris (pH 8.5).
The ends of the sonicated DNA fragments were repaired with a combination of T4 DNA polymerase and the Klenow fragment of E. coli DNA polymerase I, as follows. Reactions were performed in a reaction mixture (final volume, 100 μl) containing sonicated phage DNA, 10 mM Tris-HCl [pH 8.0], 50 mM NaCl, 10 mM MgCl2, 1 mM DTT, 50 μg/ml BSA, 100 μM of each dNTP and 15 units of T4 DNA polymerase (New England Biolabs) for 20 min at 12°C followed by addition of 12.5 units of Klenow large fragment (New England Biolabs) for 15 min at room- temperature. The reaction was stopped by two phenol chloroform extractions and the
DNA was precipitated with ethanol and the final DNA pellet was resuspended in 20 μl of H2O.
Blunt-ended DNA fragments were cloned by ligation directly into the Hinc II site of pKSII+ vector (New England Biolabs) dephosphorylated by treatment with calf intestinal alkaline phosphatase (New England Biolabs)-treated pKS 11+ vector
(Stratagene). A typical ligation reaction contained 100 ng of vector DNA, 2 to 5 μl of repaired sonicated phage DNA (50-100 ng) in a final volume of 20 μl containing 800 units of T4 DNA ligase (New England Biolabs) and was incubated overnight at 16°C. Transformation and selection of bacterial clones containing recombinant plasmids was performed in E. coli DHlOβ according to standard procedures (Sambrook et al., 1989).
Recombinant clones were picked from agar plates into 96-well plates containing 100 μl LB and 100 μg/ml ampicillin and incubated at 37°C. The presence of phage DNA insert was confirmed by PCR amplification using T3 and T7 primers flanking the Hinc II cloning site of the pKS 11+ vector. PCR amplification of foreign insert was performed in a 15 μl reaction volume containing 10 mM Tris (pH 8.3), 50 mM KCl, 1.5 mM MgCl2, 0.02% gelatin, 1 μM primer, 187.5 μM each dNTP, and 0.75 units Taq polymerase (BRL). The thermocycling parameters were as follows: 2 min initial denaturation at 94°C for 2 min, followed by 20 cycles of 30 sec denaturation at 94°C, 30 sec annealing at 57°C, and 2 min extension at 72°C, followed by a single extension step at 72°C for 10 min. Clones with insert sizes of 1 to 2 kbp were selected and plasmid DNA was prepared from the selected clones using QIAprep™ spin miniprep kit (Qiagen).
The nucleotide sequence of the extremities of each recombinant clone was determined using an ABI 377-36 automated sequencer with two types of chemistry: ABI prism Big Dye™ primer or ABI prism Big Dye™ terminator cycle sequencing ready reaction kit (Applied Biosystems). To ensure co-linearity of the sequence data and the genome, all regions of phage genome were sequenced at least once from both directions on two separate clones. In areas that this criteria was not initially met, a sequencing primer was selected and phage DNA was used directly as sequencing template employing ABI prism Big Dye™ terminator cycle sequencing ready reaction kit.
Example 3. Bioinformatic management of primary nucleotide sequence from Phage 77. " _ —• *" "
Phage 77 sequence contigs were assembled using Sequencher™ 3.1 software (GeneCodes). To close contig gaps, sequencing primers were selected near the edge of
the contigs. Phage DNA was used directly as sequencing template employing ABI prism BIG DYE™ terminator cycle sequencing ready reaction kit. The complete sequence of bacteriophage 77 is shown in Table 2.
A software program was developed and used on the assembled sequence of bacteriophage 77 to identify all putative ORFs larger than 33 codons. Other ORF identification software can also be utilized, preferably programs which allow alternative start codons. The software scans the primary nucleotide sequence starting at nucleotide #1 for an appropriate start codon. Three possible selections can be made for defining the nature of the start codon; I) selection of ATG, II) selection of ATG or GTG, and III) selection of either ATG, GTG, TTG, CTG, ATT, ATC, and ATA. This latter initiation codon set corresponds to the one reported by the NCBI (http://www.ncbi.nlm.nih.gov/htbin-post/Taxonomv/wprintgc ?mode=c. for the bacterial genetic code.
When an appropriate start codon is encountered, a counting mechanism is employed to count the number of codons (groups of three nucleotides) between this start codon and the next stop codon downstream of it. If a threshold value of 33 is reached, or exceeded, then the sequence encompassed by these two codons (start and stop codons) is defined as an ORF. This procedure is repeated, each time starting at the next nucleotide following the previous stop codon found, in order to identify all the other putative ORFs. The scan is performed on all three reading frames of both DNA strands of the phage sequence.
Sequence homology (BLAST) searches for each ORF are then carried out using an implementation of BLAST programs, although any of a variety of different sequence comparison and matching programs can be utilized as known to those skilled in the art. Downloaded public databases used for sequence analysis include: i) non-redundant GenBank (ftp://ncbi.nlm.nih.gOv/blast/db/nr.Z), ii) Swissprot (ftp://ncbi.nlm.nih.gOv/blast/db/swissprot.Z); iii) vector (ftp://ncbi.nlm.nih.gOv/blast/db/vector.Z); iv) pdbaa databases (ftp://ncbi.nlm.nih.gOv/blast/db/pdbaa.Z); v) S. aureus NCTC 8325 (ftp://ftp.genome.ou.edu/pub/staph/staph-lk.fa); vi) streptococcus pyogenes (ftp://ftp.genome.ou.edu/pub/strep/strep-lk.fa); vii) Streptococcus pneumoniae
(ftp://ftp.tigr.org/pub/data/s_pneumoniae/gsp.contigs.112197.Z); viii) Mycobacterium tuberculosis CSU#9 (ftp://ftp.tigr.Org/pub/data/m_tuberculosis/TB_091097.Z) and ix) pseudomonas aeruginosa Chttp://www.genome.washington.edu/pseudo/data.html..
The results of the homology searches performed on the ORFs is shown in Table 5.
Example 4. Subcloning of Bacteriophage 77 ORFs into a Staph A inducible expression system.
The shuttle vector pT0021, in which the firefly luciferase (lucFF) expression is controlled by the ars (arsenite) promoter/operator (Tauriainen et al., 1997), was modified in the following fashion. Two oligonucleotides corresponding to a short antigenic peptide derived from the heamaglutinin protein of influenza virus (HA epitope tag) were synthesized (Field et al., 1988). The sense strand HA tag sequence (with BamHI, Sail and Hindlll cloning sites) is:
5 '-gatcccggtcgaccaagcttTACCCATACGACGTCCCAGACTACGCCAGCTGA-3 ' (where upper case letters denote the nucletotide sequence of the HA tag); the antisense strand HA tag sequence (with a Hindlll cloning site) is: 5 '-agctTCAGCTGGCGTAGTCTGGGACGTCGTATGGGTAaagcttggtcgaccgg-3 ' (where upper case letters denote the sequence of the HA tag). The two HA tag oligonucleotides were annealed and ligated into pT0021 vector which had been digested with BamHI and Hindlll. This manipulation resulted in replacement of the lucFF gene by the HA tag. This modified shuttle vector containing the arsenite inducible promoter, the arsR gene, and HA tag was named pTHA. A diagram outlining our modification of pT0021 to generate pTHA is shown in Fig. 1A.
Each ORF, encoded by Bacteriophage 77, larger than 33 amino acids and having a Shine-Dalgarno sequence upstream of the initiation codon was selected for functional analysis for bacterial inhibition. In total, 98 ORFs were selected and screened as detailed below. A list of these is presented in Table 3. Each individual ORF, from initiation codon to last codon (excluding the stop codon), was amplified from phage genomic DNA using the polymerase chain reaction (PCR). For PCR amplification of ORFs, each sense strand primer targets the initiation codon and is preceded by a BamHI restriction site (5cgggatcc3'. and each antisense oligonucleotide targets the pentultimate codon (the one before the stop codon) of the ORF and is preceded by a Sal I restriction site 'gcgtcgaccg3). The PCR product of each ORF was gel purified and digested with BamHI and Sail. The digested PCR product was then gel purified using the Qiagen kit as described, ligated into BamHI and Sail digested pTHA vector, and used to transform E. coli bacterial strain DH10β(as described _ - - above). As a result of this manipulation, the HA tag is set inframe with the ORF and is positioned at the carboxy terminus of each ORF (pTHA ORF clones). Recombinant pTHA/ORF clones were picked and their insert sizes were confirmed by PCR analysis
using primers flanking the cloning site. The names and sequences of the primers that were used for the PCR amplification were: HAF:
5TATTATCCAAAACTTGAACA3'; HAR: 5 CGGTGGTATATCCAGTGATT3'. The sequence integrity of cloned ORFs was verified directly by DNA sequencing using primers HAF and HAR. In cases where verification of ORF sequence could not be achieved by one pass with the sequencing primers, additional internal primers were selected and used for sequencing.
Staphylococcus aureus strain RN4220 (Kreiswirth et al., 1983) was used as a recipient for the expression of recombinant plasmids. Electoporation was performed essentially as previously described (Schenk and Laddaga, 1992). Selection of recombinant clones was performed on Luria-Broth agar (LB-agar) plates containing 30 μg/ml of kanamycin.
For each ORF introduced in the pTHA plasmid, 3 independent transformants were isolated and used to individually inoculate cultures in 5 ml of TSB containing 30μg/ml kanamycin, followed by growth to saturation (16 hrs at 30°C). An aliquot of this stationary phase culture was used to generate a frozen glycerol stock of the transformant ( stored at - 80°C). The remaining culture was used for plasmid DNA extraction. Bacterial cells were harvested by centrifugation at 3000 x g at 22°C for 5 min. The pellet was resuspended in 200 μl 25%) sucrose containing 25U/ml of lysostaphin and incubated for 15 min at 37°C. Then, 400μl of alkaline SDS solution (3%) SDS, 0.2N NaOH) were added, well mixed and incubated for 7 min at room temperature. After the alkaline SDS treatment, 300μl of ice-cold 3M sodium acetate pH 4.8 were added, and the mix is immediately spun at 13000g for 15 min at room temperature. The supernatant was transferred to a new 1.5 ml conical centrifuge tube and 650μl of isopropanol (stored at room temperature) were added. The mix was then centrifuged at 13,000 x g for 5 min. The supernatant fluid was discarded, the pellet washed with 70% ethanol, and resuspended in 320 μl sterile distilled water.
The presence of individual phage 77 ORF DNA inserts in the plasmid was verified by PCR amplification using 1.5 μl transformant miniprep DNA in a PCR with primers flanking the cloning site of ORF in pTHA vector (HAF and HAR). The composition of the PCR reaction and the cycling parameters are identical to those employed for library screening described above.
Example 5. Functional assay for bacterial inhibitory activity of bacteriophage 77 ORFs. "
The anti-microbial activity of individual phage 77 ORFs was monitored by two growth inhibitory assays, one on solid agar medium, the other in liquid medium.
In general, Staphylococcus bacteria transformed with expression plasmids containing individual ORFs were grown in normal TSA medium and stored in 19% glycerol. At pre-determined times, arsenite was added to the culture to induce transcription of the phage 77 ORFs cloned immediately downstream from an arsenite-inducible promoter in the pTHA expression plasmid.
The effect of ORF induction on bacterial growth characteristics was then monitored and quantitated. The growth inhibition assay on solid medium was performed by streaking pTHA ORF containing S. aureus transformant onto LB-Kn and TSA-Kn plates containing increasing concentrations of sodium arsenite (0; 2.5; 5; and 7.5 μM). Arsenite is used to induce the expression of cloned DNA in pTHA vector. In parallel, 3 μl of 1/10 and 1/100 dilutions of the frozen cultures of the pTHA/ORF transformants were spotted as single drops onto LB-Kn and TSA-Kn plates containing increasing concentration of sodium arsenite (0; 2.5; 5; and 7.5 μM). The plates were then incubated 16 hrs at 37°C, and the effect of arsenite-induced ORF expression on bacterial growth was monitored and quantitated by comparing the extent to that seen in control plates. As positive controls for growth inhibition,the holin/lysin genes of the Sthaphylococcus aureus phage Twort (Loessner et al., 1998) was subcloned into the pTHA ars inducible vector and used.
For the growth inhibition assay in liquid medium, stationary phase cultures were prepared by inoculating 2.5ml TSB-Kn with frozen S. aureus RN4220 transformants containing phage 77 ORFs cloned in pTHA vector followed by incubation for 16 hrs at 37°C. These cultures were then diluted 1/100 in the same medium, and the bacteria were allowed to grow for 2 hrs at 37°C to reach early log phase. 150 μl of such culture were then mixed with 2.35 ml TSB-Kn medium with or without arsenite (the final concentration of arsenite in the medium was 0 or 5 μM arsenite). After 3.5 hrs incubation at 37°C with shaking at 250 φm, 100 μl of bacterial culture was removed from each tube for OD565 measurement. Serial ten-fold dilutions of the culture in buffered saline solution (0.85%> NaCl) were then spotted onto TSB-Kn plates. The plates were incubated at 37°C 16 hrs and the number of surviving colonies counted the following day. The growth inhibitory property of individual ORFs was then quantitated by comparing CFU numbers under normal or arsenite-induction conditions. A schematic flow of the inhibition analysis is shown in Fig. 3 (also applicable to inhibition analysis for the other phage and bacteria pointed out herein). Inhibition results are shown in Figures 4A-C.
Example 6: Itentification of Cecropin Signature Motif in Staphylococcus aureus Bacteriophage 3A ORF
The genome for S. aureus bacteriophage 3A was determined and the sequence was analyzed essentially as described for bacteriophage 77 in the examples above. Upon blast analysis of the identified open reading frames of phage 3A, the presence of an amino acid sequence corresponding to a cecropin signature motif was observed. This motif (WDGHKTLEK) is located at position aa 481 -489. Cecropins were originally identified in proteins from the cecropia moth and are recognized as potent antibacterial proteins that constitute an important part of the cell-free immunity of insects. Cecropins are small proteins (31-39 amino acid residues) that are active against both Gram-positive and Gram-negative bacteria by disrupting the bacterial membranes. Although the mechanisms by which the cecropons cause cell death are not fully understood, it is generally thought to involve channel formation and membrane destabilization.
The identification of a motif corresponding to a known inhibitor suggests that the product of ORF002 is also an inhibitory compound. Such inhibitory activity can be confirmed as described herein or by other methods known in the art. Confirmation of the inhibitory activity would indicate that the ORF product could serve as the basis for construction of mimetic compounds and other inhibitors directed to the target of the ORF002 product.
Boman & Hultmark, 1987, Ann. Rev. Microbiol. 41:103-126. Boman, 1991, Cell 65:205-207.
Boman et al., 1991, Eur. J. Bioichem. 201:23-31.
Wang et al., J. Biol. Chem. 273:27438-27448.
Example 7. Growth of Staphylococcus aureus bacteriophage 44 AHJD: Staphylococcus aureus propagating strain (PS 44A) (Felix d'Herelle Reference
Centre #HER 1101) was used as a host to propagate its respective phage 44 AHJD (Felix d'Herelle Reference Centre #HER 101). Two rounds of plaque purification of phage 44AHJD were performed on soft agar essentially as described in Sambrook et al. (1989). Briefly, the Staphylococcus aureus PS strain was grown overnight at 37°C in Nutrient Broth [NB: 3 g Bacto Beef Extract, 5 g Bactopeptone per liter, (Difco Laboratories # 0003-17-8), supplemented with 0.5% NaCl]. The culture was then diluted 20 fold in NB and incubated at 37°C until an OD540 of 0.2. In order to obtain single plaques, phage 44 AHJD was subjected to 10-fold serial dilutions using the phage buffer (1 mM MgSO4, 5 mM MgCl2, 80 mM NaCl and 0.1% Gelatin) -ancflO μl were used to infect 0.5 ml of the cell suspension in the presence of 400 μg/ml of
CaCl2. After incubation of 15 min at room temperature, 2 ml of melted soft agar (NB supplemented with 0.6% of agar) were added to the mixture and poured onto the surface of 100 mm nutrient agar plates (3 g Bacto Beef extract, 5 g Bactopeptone, 0.5% NaCl and 15 g of Bacto agar per liter (Difco Laboratories # 0001-17-0). After overnight incubation at 37°C, a single plaque was isolated, resuspended in 1ml of phage buffer by end over end rotation for 2 h at room temperature and the phage suspension was diluted and used for a second infection as described above. After overnight incubation at 37°C, a single plaque was isolated and used as a stock.
Large scale purification of bacteriophage and preparation of phage DNA was as follows.
The propagation method was carried out by using the agar layer method described by Swanstόrm and Adams (1951). Briefly, the PS 44A strain was grown to stationary phase overnight at 37°C in Nutrient Broth. The culture was then diluted 20x in NB and incubated at 37°C until the A540= 0.2. The suspension (15xl07 Bacteria) was then mixed with 15xl05 phage particles to give a ratio of 100-bacteria/phage particle in the presence of 400 μg/ml of CaCl2. After incubation of 15 min at room temperature, 7.5 ml of melted soft agar were added to the mixture and poured onto the surface of 150 mm nutrient agar plates and incubated overnight at 37°C. To collect the lysate, 20 ml of NB were added to each plate and the soft agar layer was collected by scrapping off with a clean microscope slide and shaken vigorously for 5 min to break up the agar. The mixture was then centrifuged for 10 min at 4,000 φm (2,830 xg) using a JA-10 rotor (Beckman) and the supernatant (lysate) is collected and subjected to a treatment with 10 μg/ml of DNase I and RNase A for 30 min at 37°C. To precipitate the phage particles, 10% (w/v) of PEG 8000 and 0.5 M of NaCl were added to the lysate and the mixture was incubated on ice for 16 h. The phage was recovered by centrifugation at 4,000 φm (3,500 xg) for 20 min at 4°C on a GS-6R table top centrifuge (Beckman).
The pellet was resuspended with 2 ml of phage buffer (1 mM MgSO4, 5 mM MgCl2, 80 mM NaCl and 0.1 %> Gelatin). The phage suspension was extracted with 1 volume of chloroform and further purified by centrifugation on a preformed cesium chloride step gradient as described in Sambrook et al. (1989), using a TLS 55,roior and centrifuged in an Optima TLX ultracentrifuge (Beckman) for 2 h at 28,000 φm (67,000 xg) at 4°C. Banded phage was collected and ultracentrifuged again on an
isopycnic cesium chloride gradient (1.45 g/ml) at 40,000 φm (64,000 x g) for 24 h at 4°C using a TLV rotor (Beckman). The phage was harvested and dialyzed for 4 h at room temperature against 4 L of dialysis buffer consisting of 10 mM NaCl, 50 mM Tris-HCl [pH 8] and 10 mM MgCl2. Phage DNA was prepared from the phage suspension by adding 20 mM EDTA, 50 μg/ml Proteinase K and 0.5% SDS and incubating for 1 h at 65°C, followed by successive extractions with 1 volume of phenol, 1 volume of phenol-chloroform and 1 volume of chloroform. The DNA was then dialyzed overnight at 4°C against 4 L of TE (10 mM Tris-HCl [pH 8.0], lmM EDTA).
Example 8. DNA sequencing of the Bacteriophage 44 AHJD genome.
Four mg of phage DNA was diluted in 200 μl of TE pH 8.0 in a 1.5 ml eppendorf tube and sonication was performed (550 Sonic Dismembrator, Fisher Scientific). Samples were sonicated under an amplitude of 3 μm with bursts of 5 s spaced by 15 s cooling in ice/water for 3 to 4 cycles and size fractionated on 1% agarose gels. The sonicated DNA was then size fractionated by gel electrophoresis. Fractions ranging from 1 to 2 kbp were excised from the agarose gel and purified using a coommercial DNA extraction system according to the instructions of the manufacturer (Qiagen) and eluted in 50 μl of lmMTris-HCl [ pH 8.5]. The ends of the sonicated DNA fragments were repaired with a combination of
T4 DNA polymearse and the Klenow fragment of E. coli DNA polymerase 1 as follows. Reactions were performed in a final volume of 100 μl containing DNA, 10 mM Tris-HCl pH 8.0, 50 mM NaCl, 10 mM MgCl2, 1 mM DTT, 5 μg BSA, 100 μM of each dNTP and 15 units of T4 DNA polymerase (New England Biolabs) for 20 min at 12°C followed by addition of 12.5 units of Klenow fragment (New England Biolabs) for 15 min at room temperature. The reaction was stopped by two phenol/chloroform extractions and the DNA was ethanol precipitated and resuspended in 20 μl of H2O. Cloning of the sonicated phage DNA into pKSII vector and transformation: Blunt-ended DNA fragments were cloned by ligation directly into the-HrøcII_ ~ site of the pkSII vector (Stratagene) dephosphorylated with calf intestinal alkaline phosphatase (New England Biolabs). A typical reaction contained 100 ng of vector, 2
to 5 μl of repaired sonicated phage DNA (50-100 ng) in a final volume of 20 μl containing 800 units of T4 DNA ligase (New England Biolabs) overnight at 16°C. Transformation and selection of positive clones was performed in the host strain DH10 β of E. coli using ampicillin as a selective antibiotic as described in Sambrook er a/. (1989).
Recombinant clones were picked from agar plates into 96-well plates containing 100 ml LB and 100 μg/ml ampicillin and incubated at 37°C. The presence of phage DNA insert was confirmed by PCR amplification using T3 and T7 primers flanking the Hindi cloning site of the pKS vector. PCR amplification of the potential foreign inserts was performed in a 15 μl reaction volume containing 10 mM Tris-HCl (pH 8.3), 50 mM KCl, 1.5 mM MgCl2, 0.02% gelatin, 1 mM primer, 187.5 μM each dNTP, and 0.75 units Taq polymerase (BRL). The thermocycling parameters were as follows: 2 min initial denaturation at 94°C for 2 min, followed by 20 cycles of 30 sec denaturation at 94°C, 30 sec annealing at 58C, and 2 min extension at 72°C, followed by a single extension step at 72°C for 10 min. Clones with insert sizes of 1 to 2 kbp were selected and plasmid DNA was prepared from the selected clones using the QIAprep™ spin miniprep kit (Qiagen).
The nucleotide sequence of the extremities of each recombinant clone was determined using an ABI 377-36 automated sequencer with two types of chemistry: ABI prism BigDye™ primer cycle sequencing (21M13 primer: #403055)(M13REV primer: #403056) or ABI prism BigDye™ terminator cycle sequencing ready reaction kit (Applied Biosystems; #4303152). To ensure co-linearity of the sequence data and the genome, all regions of the phage genome were sequenced at least once from both directions on two separate clones. In areas that this criteria was not initially met, a sequencing primer was selected and phage DNA was used directly as sequencing template employing ABI prism BigDye™ terminator cycle sequencing ready reaction kit.
Example 9. Bioinformatic management of primary nucleotide sequence. Sequence contigs were assembled using Sequencher™ 3.1 software " — ""
(GeneCodes). To close contig gaps, sequencing primers were selected near the edge of the contigs. Phage DNA was used directly as sequencing template employing ABI
prism BigDye™ terminator cycle sequencing ready reaction kit (Applied Biosystems;
#4303152). The complete sequence oϊ Staphylococcus aureus bacteriophage 44AHJD is shown in Table 16.
A software program was used on the assembled sequence of bacteriophage 44AHJD to identify all putative ORFs larger than 33 codons. The software scans the primary nucleotide sequence starting at nucleotide #1 for an appropriate start codon.
Three possible selections can be made for defining the nature of the start codon; I) selection of ATG, II) selection of ATG or GTG, and III) selection of either ATG,
GTG, TTG, CTG, ATT, ATC, and ATA. This latter initiation codon set corresponds to the one reported by the NCBIfhttρ://www.ncbi.nlm.nih.gov/htbin- post/Taxonomv/wprintgc?mode=c. for the bacterial genetic code. When an appropriate start codon is encountered, a counting mechanism is employed to count the number of codons (groups of three nucleotides) between this start codon and the next stop codon downstream of it. If a threshold value of 33 is reached, or exceeded, then the sequence encompassed by these two codons is defined as an ORF. This procedure is repeated, each time starting at the next nucleotide following the previous stop codon found, in order to identify all the other putative ORFs. The scan is performed on all three reading frames of both DNA strands of the phage sequence.
The predicted ORFs for bacteriophage 44AHJD are listed in Tables 17 & 18. Sequence homology searches for each ORF were carried out using an implementation of blast programs. Downloaded public databases used for sequence analysis include:
(i) non-redundant GenBank (ftp://ncbi.nlm.nih.gOv/blast/db/nr.Z), ii) Swissprot (ftp://ncbi.nlm.nih.gOv/blast/db/swissprot.Z); iii) vector (ftp://ncbi.nlm.nih.gOv/blast/db/vector.Z); iv) pdbaa databases (ftp://ncbi.nlm.nih.gOV/Wast/db/pdbaa.Z); v) Staphylococcus aureus NCTC 8325 (ftp://ftp.genome.ou.edu/pub/staph/staph- lk.fa); vi)Stα/7/j >/ococcM-: o ene5(ftp://ftp.tigr.org/pub/data/s_pneumoniae/gsp.contigs.1121 97.Z); vii)PRODOM(ftp://ftp.toulouse.ir_ra.fr/pub/prodom/current_release/prodom99__l.forbl ast.gz); viii) DOMO (ftp://ftp.infobiogen.fr/pub/db/domo/);
ix) TREMBL (ftp://www.expasy.ch/databases/sp_tr_nrdb/fasta/)
The results of the homology searches performed on the ORFs of bacteriophage 44AHJD are shown in Tables 19 & 20.
Example 10. Sub-Cloning of Bacteriophage 44 AHJD ORFs.
Expression preferably utilizes a shuttle expression vector which is arranged such that expression of the exogenous bacteriophage 44 AHJD ORF sequence is inducible. For example, the shuttle vector pT0021, in which the firefly luciferase (lucFF) expression is controlled by the ars (arsenite) promoter/operator (Tauriainen et al., 1997), can be modified in the following fashion. Two oligonucleotides corresponding to a short antigenic peptide derived from the heamaglutinin protein of influenza virus (HA epitope tag) were synthesized (Field et al., 1988). The sense strand HA tag sequence (with BamHI, Sail and Hindlll cloning sites) is: 5 '-gatcccggtcgaccaagcttTACCCATACGACGTCCCAGACTACGCCAGCTGA-3 ' (where upper case letters denote the nucletotide sequence of the HA tag); the antisense strand HA tag sequence (with a Hindlll cloning site) is:
5'-agctTCAGCTGGCGTAGTCTGGGACGTCGTATGGGTAaagcttggtcgaccgg-3' (where upper case letters denote the sequence of the HA tag). The two HA tag oligonucleotides were annealed and ligated into pT0021 vector which had been digested with BamHI and Hindlll. This manipulation resulted in replacement of the lucFF gene by the HA tag. This modified shuttle vector containing the arsenite inducible promoter, the arsR gene, and HA tag was named pTHA. A diagram outlining our modification of pT0021 to generate pTHA is shown in Fig. 1A (another userful vector construct is shown in Fig. IB). Each ORF, encoded by Bacteriophage 44 AHJD, larger than 33 amino acids and having a Shine-Dalgarno sequence upstream of the initiation codon can be selected for functional analysis for bacterial inhibition. Each individual ORF, from initiation codon to last codon (excluding the stop codon), can be amplified from phage genomic DNA using the polymerase chain reaction (PCR). For PCR amplification of ORFs, each sense strand primer targets the initiation codon and is preceded by a BamHI restriction site (5'cgggatcc3') and each antisense oligonucleotide targets the pentultimate codon (the one before the stop codon) of the ORF and is preceded by a Sal I restriction site (5'gcgtcgaccg3). The PCR product of each ORF can be gel
purified and digested with BamHI and Sail. The digested PCR product can then be gel purified using the Qiagen kit as described, ligated into BamHI and Sail digested pTHA vector, and used to transform E. coli bacterial strain DH10β(as described above). As a result of this manipulation, the HA tag is set inframe with the ORF and is positioned at the carboxy terminus of each ORF (pTHA/ORF clones). Recombinant pTHA ORF clones will be picked and their insert sizes were confirmed by PCR analysis using primers flanking the cloning site. The following primers can be used for PCR amplification: HAF: 5TATTATCCAAAACTTGAACA3'; HAR: 5 CGGTGGTATATCCAGTGATT3'. The sequence integrity of cloned ORFs can be verified directly by DNA sequencing using primers HAF and HAR. In cases where verification of ORF sequence can not be achieved by one pass with the sequencing primers, additional internal primers will be selected and used for sequencing.
Staphylococcus aureus strain RN4220 (Kreiswirth et al., 1983) will be used as a recipient for the expression of recombinant plasmids. Εlectoporation will be performed essentially as previously described (Schenk and Laddaga, 1992). Selection of recombinant clones will be performed on Luria-Broth agar (LB-agar) plates containing 30 μg/ml of kanamycin.
Alternatively, a constitutive promoter can be used to drive expression of the introduced ORF, and compare cell growth to control bacterial cells containing the parental vector lacking any introduced phage ORF. Recombinant plasmids will be introduced into Staphylococcus aureus strain RN4220 (Kreiswirth et al., 1983) using electoporation as previously described (Schenk and Laddaga, 1992). Cloning of ORFs with a Shine-Dalgarno sequence
ORFs with a Shine-Dalgarno sequence are selected for functional analysis of bacterial killing. Each ORF, from initiation codon to last codon (excluding the stop codon), can be amplified by PCR from phage genomic DNA. For PCR amplification of ORFs, each sense strand primer starts at the initiation codon and is preceded by a restriction site and each antisense strand starts at the last codon (excluding the stop codon) and is preceded by a different restriction site. The PCR product of each ORF will be gel purified and digested with the restriction enzymes with sites contained on the PCR oligonucleotides. The digested PCR product is then gel purified using4he ~ Qiagen kit, ligated into the modified shuttle vector, and used to transform bacterial strain DH10. Recombinant clones are then picked and their insert sizes confirmed by
PCR analysis using primers flanking the cloning site as well as restriction digestion. The sequence fidelity of cloned ORFs can be verified by DNA sequencing using the same primers as used for PCR. In the cases that the verification of ORFs can not be achieved by one path of sequencing using primers flanking the cloning site internal primers can be selected and used for sequencing. Recombinant plasmids can be introduced into Staphylococcus aureus strain RN4220 (Kreiswirth et al., 1983) using electoporation as previously described (Schenk and Laddaga, 1992). Induction of gene expression from the ars promoter.
If an inducible promoter is used, e.g., the ars promoter, induction can be assessed, for example, in either of the two methods.
1. Screening on agar plates
The functional identification of killer ORFs can be performed by spreading an aliquot of S. aureus transformed cells containing phage 44 AHJD ORFs onto agar plates containing different concentrations of sodium arsenite (0; 2.5; 5; and 7.5 μM). The plates are incubated overnight at 37°C, after which a growth inhibition of the ORF transformants on plates that contain arsenite are compared to plates without arsenite.
2. Quantification of growth inhibition in liquid medium
Cells containing different recombinant plasmids can be grown for overnight at 37°C in LB medium supplemented with the appropriate antibiotic selection. These are then diluted to the mid log phase (OD540= 2) with fresh media containing antibiotic and transferred to 96-well microtitration plates (100 μl/well). Inducer is then added at different final concentrations (ranging from 2.5 to 10 μM) and the culture incubated for an additional 2 hrs at 37°C. The effect of expression of the phage 44 AHJD ORFs on bacterial cell growth is then monitored by measuring the OD540 and comparing the rate of growth to the culture not containing inducer. [As positive controls for growth inhibition, the kilA gene of phage lambda (Reisinger, GR., Rietsch, A., Lubitz, W. and Blasi, U. 1993 Virology #193: 1033-1036), and the holin/lysin genes of the Sthaphylococcus aureus phage Twort (Loessner, MJ., Gaeng, S., Wendlinger, G., Maier, SK. and Scherer, S. 1998. FEMS Microbiology Letters #162:265-274) can be subcloned into the ars inducible vector. An aliquot of the induced and uninduced culture can also be plated out on agar plates containing an appropriate antibiotic- selection but lacking inducer. Following incubation overnight at 37°C, the number of
colonies is counted. Any ORF showing bacteriostatic activity will show a lower, but detectable, number of colonies on the agar plates when grown in the presence of inducer as compared to when grown in the absence of inducer. Any ORF showing full bacteriocidal activity will show no colonies on the agar plates, when grown in the presence of inducer as compared to when grown in the absence of inducer.
REFERENCES
Ackermann, H-W. and DuBow, M. S. (1987). Viruses of Prokaryotes. Volumes I and II. CRC Press, Boca Raton, Florida.
Tenover, F.C. and McGowan Jr., J.E. (1998). Bacterial Infections of Humans. Epidemiology and Control.(A.S. Evans and P.S. Brachman, eds.) Plenum Medical Book Company, New York, N.Y. pp. 83-93.
Rusterholtz, K., and Pohlschroder, M. ( 1999). Cell 96, 469-470.
Gray, B.M. (1998). Bacterial Infections of Humans. Epidemiology and Control.(A.S. Evans and P.S. Brachman, eds.) Plenum Medical Book Company, New York, N.Y. pp. 673 D 711.
Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989). Molecular cloning: A laboratory manual. Cold Spring Harbor Laboratory, New York. Cold Spring Harbor Laboratory Press.
Ausubel, F.M. et al. (1994) Current Protocols in Molecular Biology. John Wiley & Sons, Secaucus, NJ.
Rost B,l and Sander C. (1996). Ann. Rev. Biophy. Biomol. Struct. 25, 113-136.
Martin, A.C., Lopez, R., Garcia, P. (1998). J Bacteriol 180, 210-217.
Steiner, M., Lubitz, W., Blasi, U. (1993). J. Bacteriol. 175, 1038-1042.
Durfee, T., Becherer, K., Chen, P.-L., Yeh, S.-H., Yang, Y., Kilburn, A.E., Lee, W, H., and Elledge, S J. (1993). Genes Dev. 7, 555-569.
Qiu, H., Garcia-Barrio, M.T., and Hinnebusch, A.G. ( 1998). Mol Cell Biol. 18, 2697-2711.
Katagiri, T., Saito, H., Shinohara, A., Ogawa, H., Kamada, N., Nakamura ,Y., and Miki, Y. (1998). Genes, Chromosomes & Cancer 21, 217-222.
Endo, T.A., Masuhara, M., Yokouchi, M., Suzuki, R., Sakamoto, H., Mitsui, K., Matsumoto, A., Tanimura, S., Ohtsubo, M., Misawa, H., Miyazaki, T., Leonor N., Taniguchi, T., Fujita, T., Kanakura, Y., Komiya, S., and Yoshimura, A. (1997). Nature 387, 921-924.
Karimova, G., Pidoux, J., Ullmann, A., Ladant, D. (1998) Proc. Natl. Acad. Sci. 95, 5752-5756.
Sopta, M., Carthew, R.W., and Greenblatt, J. (1995) J. Biol. Chem. 260, 10353- 10369.
Qin, J., Fenyo, D., Zhao, Y., Hall, W.W., Chao, D.M., Wilson, C.J., Young, R.A. and Chait, B.T. (1997). Anal. Chem. 69, 3995-4001.
Swanstrόm, M. and Adams, M.H. (1951). Proc. Soc. Exptl. Biol. & Med. 78: 372- 375.
Røder, B.L., Wandall, D. A., Frimødt-Moller, N., Epersen, F., Skinhøj, P. and Rosdahl, T. (1999). Arch. Intern. Med. 159: 462-469.
Sanabria, T.J., Albert, J.S., Goldberg, R., Pape, L.A. and Cheeseman, S.H. (1990). Arch. Intern. Med. 150: 1305-1309.
Frimødt-Moller, N., Epersen, F., Skinhøj, P. and Rosdahl, V.T. (1997). Clin. Microbiol. Infect. 3: 297-305.
Harbath, S., Rutschmann, O., Sudre, P. and Pittet, D. (1998). Arch. Intern. Med. 158: 182-189.
Steinberg, J.P., Clark, C.C. and Hackman, B.O. (1996). Clin. Infect. Dis. 23: 255-259.
Field, J., Nikawa, J.-L, Broek, D., MacDonald, B., Rodgers, L., Wilson, LA., Lemer, R.A., and Wigler, M. (1988). Purification of a RAS -responsive adenylyl cyclase complex from Saccharomyces cerevisiae by use of an epitope addition method. Mol. Cell. Biol. 8: 2159-2165.
Kreiswirth, BN., Lofdahl, S., Belley, MJ., O'Reilly, M., Shlievert, PM., Bergdoll, MS. and Novicks, RP. (1983) Nature 305: 709-712.
Schenk, S. and Laddaga, RA. (1992) FEMS Microbiology Letters 94: 133-138.
Cohen, M.L. (1992) Science 257, 1050-1055.
Example 11. Growth of Enterococcus bacteriophage 182 and purification of genomic DNA.
The Enterococcus propagating strain (PS) (Enterococcus sp. Group D, Felix d'Herelle Reference Centre #HER 1080) was used as host to propagate its respective phage 182 (Felix d'Herelle Reference Centre #HER 80). Two rounds of plaque purification of phage 182 were performed on soft agar essentially as described in Sambrook et al. (1989). Briefly, the Enterococcus sp. PS strain was grown overnight at 37°C in Tryptic Soy Broth [TSB: 17 g Bacto tryptone, 3 g Bacto soytone, 2.5 g Bacto dextrose, 5 g Sodium chloride, and 2.5 g Dipotassium phosphate per liter (Difco Laboratories (#0370-17-3)]. The culture was then diluted 20 fold in TSB and incubated at 37°C until the OD540= 0.2 (early log phase) with constant agitation. In order to obtain single plaques, phage 182 was subjected to 10 fold serial dilutions using the phage buffer (1 mM MgSO4, 5 mM MgCl2, 80 mM NaCl and 0.1% Gelatin (w/v)) and 10 1 of each dilution was used to infect 0.5 ml of the bacterial cell suspension. After incubation at 15 min at 37°C, 2 ml of melted soft agar (TSB supplemented with 0.6% agar) was added to the mixture and poured onto the surface of 100 mm Trytic Soy Agar plates [TSA: 15 g Tryptone peptone, 5 g Soytone peptone, 5 g Sodium chloride and 15 g of Agar per liter (Difco Laboratories #0369- 17)]. After overnight incubation at 37°C, a single plaque was isolated, resuspended in 1 ml of phage buffer by end over end rotation for 2 hrs at room temperature, and the phage suspension was diluted and used for a second infection as described above. After overnight incubation at 37°C, a single plaque was isolated and used as a stock for all subsequent manipulations.
The propagation procedure for bacteriophage 182 was modified from the agar layer method of Swanstδrm and Adams (1951). Briefly, the Enterococcus sp. PS strain was grown to stationary phase overnight at 37°C in TSB. The culture was then ~ - - diluted 20 fold in TSB and incubated at 37°C until the A540= 0.2. The suspension (15xl07 Bacteria) was then mixed with 15xl05 plaque forming units (pfu) to give a
ratio of 100-bacteria/pfu. After incubation of 15 min at 37°C, 7.5 ml of melted soft agar (TSB plus 0.6% agar) were added to the mixture and poured onto the surface of 150 mm TSA plates and incubated 16 hrs at 37°C. To collect the plate lysate, 20 ml of TSB were added to each plate and the soft agar layer was collected by scrapping off with a clean microscope slide followed by vigorous shaking of the agar suspension for 5 min to break up the agar. The mixture was then centrifuged for 10 min at 4,000 φm (2,830 xg) using a JA-10 rotor (Beckman) and the supernatant fluid (lysate) is collected and subjected to a treatment with 10 μg /ml of DNase I and RNase A for 30 min at 37°C. To precipitate the phage particles, the phage suspension was adjusted to 10% (w/v) of PEG 8000 and 0.5 M of NaCl followed by incubation at 4°C for 16 hrs. The phage was recovered by centrifugation at 4,000 φm (3,500 xg) for 20 min at 4°C on a GS-6R table top centrifuge (Beckman). The pellet was resuspended with 2 ml of phage buffer (1 mM MgSO4, 5 mM MgCl2, 80 mM NaCl and 0.1% Gelatin). The phage suspension was extracted with 1 volume of chloroform and further purified by centrifugation on a cesium chloride step gradient as described in Sambrook et al. (1989), using a TLS 55 rotor and centrifuged in an Optima TLX ultracentrifuge (Beckman) for 2 hrs at 28,000 φm (67,000 xg) at 4°C. Banded phage was collected and ultracentrifuged again on an isopycnic cesium chloride gradient (1.45 g/ml) at 40,000 φm (64,000 xg) for 24 hrs at 4°C using a TLV rotor (Beckman). The phages were harvested and dialyzed for 4 hrs at room temperature against 4 L of dialysis buffer consisting of 10 mM NaCl, 50 mM Tris-HCl [pH 8] and 10 mM MgCl2. Phage DNA was prepared from the phage suspension by adding 20 mM EDTA, 50 g/ml Proteinase K and 0.5% SDS and incubating for 1 hr at 65°C, followed by successive extractions with 1 volume of phenol, 1 volume of phenol-chloroform and 1 volume of chloroform. The DNA was then dialyzed overnight at 4°C against 4 L of TE (10 mM Tris-HCl [pH 8.0], lmM EDTA).
Example 12. DNA sequencing of the Bacteriophage 182 genome.
Four micrograms of phage DNA was diluted in 200 μl of TE (10 mM Tris, [pH 8.0], 1 mM EDTA) in a 1.5 ml eppendorf tube and sonication was performed (550 Sonic Dismembrator, Fisher Scientific). Samples were sonicated under an, amplitude of 3 μm with bursts of 5 s spaced by 15 s cooling in ice/water for 3 to 4
cycles. The sonicated DNA was then size fractionated by electrophoresis on 1% agarose gels utilizing TAE (1 x TAE is: 40 mM Tris-acetate, 1 mM EDTA [pH 8.0]) as the running buffer. Fractions ranging from 1 to 2 kbp were excised from the agarose gel and purified using a commercial DNA extraction system according to the instructions of the manufacturer (Qiagen), with a final elution of 50 μl of 1 mM Tris [pH 8.5].
The ends of the sonicated DNA fragments were repaired with a combination of T4 DNA polymerase and the Klenow fragment ofE. coli DNA polymerase I, as follows. Reactions were performed in a reaction mixture (final volume, 100 μl) containing sonicated phage DNA, 10 mM Tris-HCl [pH 8.0], 50 mM NaCl, 10 mM MgCl2, 1 mM DTT, 50 μg/ml BSA, 100 μM of each dNTP and 15 units of T4 DNA polymerase (New England Biolabs) for 20 min at 12°C followed by addition of 12.5 units of the Klenow large fragment of DNA polymerase I(New England Biolabs) for 15 min at room temperature. The reaction was stopped by two phenol/chloroform extractions and the DNA was precipitated with ethanol and the final DNA pellet resuspended in 20 μl of H2O.
Blunt-ended DNA fragments were cloned by ligation directly into the Hinc II site of the pKSII+ vector (New England Biolabs) dephosphorylated by treatment with calf intestinal alkaline phosphatase (New England Biolabs). A typical ligation reaction contained 100 ng of vector DNA, 2 to 5 μl of repaired sonicated phage DNA (50-100 ng) in a final volume of 20 μl containing 800 units of T4 DNA ligase (New England Biolabs) and was incubated overnight at 16°C. Transformation and selection of bacterial clones containing recombinant plasmids was performed in E. coli DHlOβ according to standard procedures (Sambrook et al, 1989). Recombinant clones were picked from agar plates into 96-well plates containing 100 μl LB and 100 μg/ml ampicillin and incubated at 37°C. The presence of phage DNA insert was confirmed by PCR amplification using T3 and T7 primers flanking the Hinc II cloning site of the pKS vector. PCR amplification of the potential foreign inserts was performed in a 15 μl reaction volume containing 10 mM Tris (pH 8.3), 50 mM KCl, 1.5 mM MgCl2, 0.02% gelatin, 1 μM primer, 187.5 μM each dNT-P,~ and 0.75 units Taq polymerase (BRL). The thermocycling parameters were as follows: 2 min initial denaturation at 94°C for 2 min, followed by 20 cycles of 30 sec
denaturation at 94°C, 30 sec annealing at 58°C, and 2 min extension at 72°C, followed by a single extension step at 72°C for 10 min. Clones with insert sizes of 1 to 2 kbp were selected and plasmid DNA was prepared from the selected clones using the QIAprep™ spin miniprep kit (Qiagen). The nucleotide sequence of the extremities of each recombinant clone was determined using an ABI 377-36 automated sequencer with two types of chemistry: ABI prism Big Dye™ primer cycle sequencing (21M13 primer: #403055)(M13REV primer: #403056) or ABI prism Big Dye™ terminator cycle sequencing ready reaction kit (Applied Biosystems; #4303152). To ensure co-linearity of the sequence data and the genome, all regions of the phage genome were sequenced at least once from both directions on two separate clones. In areas that this criteria was not initially met, a sequencing primer was selected and phage DNA was used directly as sequencing template employing ABI prism BigDye™ terminator cycle sequencing ready reaction kit.
Example 13. Bioinformatic management of primary nucleotide sequence. Sequence contigs were assembled using Sequencher™ 3.1 software (GeneCodes). To close contig gaps, sequencing primers were selected near the edge of the contigs. Phage DNA was used directly as sequencing template employing ABI prism BigDye™ terminator cycle sequencing ready reaction kit (Applied Biosystems; #4303152). The complete sequence oϊ Enterococcus bacteriophage 182 is shown in Table 21.
A software program was used on the assembled sequence of bacteriophage 182 to identify all putative ORFs larger than 33 codons. The software scans the primary nucleotide sequence starting at nucleotide #1 for an appropriate start codon. Three possible selections can be made for defining the nature of the start codon; I) selection of ATG, II) selection of ATG or GTG, and III) selection of either ATG, GTG, TTG, CTG, ATT, ATC, and ATA. This latter initiation codon set corresponds to the one reported by the NCBI(http://www.ncbi.nlm.nih.gov/htbin- post/Taxonomv/wprintgc?mode=c. for the bacterial genetic code. When an appropriate start codon is encountered, a counting mechanism is employed to count the number of codons (groups of three nucleotides) between this start codon and the
next stop codon downstream of it. If a threshold value of 33 is reached, or exceeded, then the sequence encompassed by these two codons is defined as an ORF. This procedure is repeated, each time starting at the next nucleotide following the previous stop codon found, in order to identify all the other putative ORFs. The scan is performed on all three reading frames of both DNA strands of the phage sequence.
The predicted ORFs for bacteriophage 182 are listed in Tables 22 & 23.
Sequence homology searches for each ORF were carried out using an implementation of BLAST programs. Downloaded public databases used for sequence analysis include: (i) non-redundant GenBank (ftp://ncbi.nlm.mh.gOv/blast db/nr.Z), ii) Swissprot (ftp://ncbi.nlm.nih.g0v/blast/db/swissprot.Z); iii) vector (ftp://ncbi.nlm.nih.gOv/blast/db/vector.Z); iv) pdbaa databases (ftp://ncbi.nlm.r_ih.g0v/blast/db/pdbaa.Z); v) staphylococcus aureus NCTC 8325 (ftp://ftp.genome.ou.edu/pub/staph/staph- lk.fa); vi) streptococcus pyrogenes
(ftp://ftp.tigr.org/pub/data/s_pneumoniae/gsp.contigs.112197.Z); vii) PRODOM fftp://ftp.toulouse.inra.fr/pub/prodom/current release/prodom99.1.forblast.gz .: viii) DOMO (ftp://ftp.infobiogen.fr/pub/db/domo/.: ix) TREMBL (ftp://www.expasy.ch databases/sp_tr_nrdb/fasta/)
The results of the homology searches performed on the ORFs of bacteriophage
182 are shown in Tables 24 & 26.
Example 14. Sub-Cloning of Bacteriophage 182 ORFs. Preparation of the shuttle expression vector
Expression preferably utilizes a shuttle expression vector which is arranged such that expression of the exogenous bacteriophage 182 ORF sequence is inducible. For example, the plasmid pND50 replicates in E. coli, E.faecalis, and S. aureus (Yamagishi, J., Kojima, T., Oyamada, Y., Fujimoto, K., Hattori, H., Nakamura, S., and Inoue, M. 1996. Antimocrob. Agents Chemother. 40, 1157-1163). This plasmid— " can be modified by conventional techniques to insert the inducible arsenite promoter, derived from the shuttle vector pT0021, in which the firefly luciferase (lucFF)
expression is controlled by the ars promoter/operator from a S. aureus plasmid (Tauriainen, S., Kaφ, M., Chang, W and Virta, M. (1997). Recombinant luminescent bacteria for measuring bioavailable arsenite and antimonite. Appl. Environ. Microbiol. 63:4456-4461). This modified shuttle vector will contain the ars promoter, arsR gene and a cloning site for introduction of individual phage ORFs downstream from a shine-delgarno sequence.
Other inducible regulatory sequences can be utilized instead of the arsenite- inducible system. An example is a nisin-inducible system The nisA promoter activity is dependent on the proteins NisR and NisK, which constitute a two-component signal transduction system that responds to the extracellular inducer nisin. The nisin sensitivity and inducer concentration required for maximal induction varies among the strains, but is functional in Streptococcus pyogenes, Streptococcus agalactiae, Streptococcus pneumoniae, Enterococcus faecalis, and Bacillus subtilis. Significant induction of the nisA promoter (10- to 60-fold induction) can be obtained in all of the species. A vector containing this promoter was published as Eichenbaum Z, Federle MJ, Marra D, de Vos WM, Kuipers OP, Kleerebezem M, and Scott JR (1998) Appl Environ Microbiol 64, 2763-2769. Other vectors, e.g., plasmids, can also be utilized which will allow replication and transciption in Enterococcus.
Alternatively, a constitutive promoter can be used (e.g„ the β-lactamase promoter is constitutive in E. faecalis - see ref. 1) to drive expression of the introduced ORF, and compare cell growth to control bacterial cells containing the parental vector lacking any introduced phage ORF. Recombinant plasmids are introduced into E. faecalis strain FA2-2 by electroporation, as previously described (Yamagishi, J., Kojima, T., Oyamada, Y., Fujimoto, K., Hattori, H., Nakamura, S., and Inoue, M. 1996. Antimicrob. Agents Chemother. 40, 1157-1163). Cloning of ORFs with a Shine-Dalgarno sequence
ORFs with a Shine-Dalgarno sequence are selected for functional analysis of bacterial killing. Each ORF, from initiation codon to last codon (excluding the stop codon), will be amplified by PCR from phage genomic DNA. For PCR amplification of ORFs, each sense strand primer starts at the initiation codon and is preceded by a restriction site and each antisense strand starts at the last codon (excluding the sto r~ codon) and is preceded by a different restriction site. The PCR product of each ORF will be gel purified and digested with the restriction enzymes with sites contained on
the PCR oligonucleotides. The digested PCR product is then gel purified using the Qiagen kit, ligated into the modified shuttle vector, and used to transform bacterial strain DHlOβ. Recombinant clones are then picked and their insert sizes confirmed by PCR analysis using primers flanking the cloning site as well as restriction digestion. The sequence fidelity of cloned ORFs will be verified by DNA sequencing using the same primers as used for PCR. In the cases that the verification of ORFs can not be achieved by one path of sequencing using primers flanking the cloning site internal primers will be selected and used for sequencing. Recombinant plasmids will be introduced into E. faecalis strain FA2-2 by electroporation, as previously described (Yamagishi, J., Kojima, T., Oyamada, Y., Fujimoto, K., Hattori, H., Nakamura, S., and Inoue, M. 1996. Antimicrob. Agents Chemother. 40, 1157-1163). Induction of gene expression from the ars promoter.
If an inducible promoter is used, e.g., the ars promoter, induction can be assessed, for example, in either of the two methods. 1. Screening on agar plates
The functional identification of killer ORFs can be performed by spreading an aliquot of E. faecalis transformed cells containing phage 182 ORF onto agar plates containing different concentrations of sodium arsenite (0; 2.5; 5; and 7.5 μM). The plates are incubated overnight at 37°C, after which a growth inhibition of the ORF transformants on plates that contain arsenite are compared to plates without arsenite. 2. Quantification of growth inhibition in liquid medium
Cells containing different recombinant plasmids can be grown for overnight at 37°C in LB medium supplemented with the appropriate antibiotic selection. These are then diluted to the mid log phase (OD540=.2) with fresh media containing antibiotic and transferred to 96-well microtitration plates (100 μl/well). Inducer is then added at different final concentrations (ranging from 2.5 to 10 μM) and the culture incubated for an additional 2 h at 37°C. The effect of expression of the phage 182 ORFs on bacterial cell growth is then monitored by measuring the OD540 and comparing the rate of growth to the culture not containing inducer. As positive controls for growth inhibition, the kilA gene of phage lambda (Reisinger, GR., Rietsch, A., Lubitz, W. and - . Blasi, U. 1993 Virology #193: 1033-1036), and the holin/lysin genes Of the Sthaphylococcus aureus phage Twort (Loessner, MJ., Gaeng, S., Wendlinger, G.,
Maier, SK. and Scherer, S. 1998. FEMS Microbiology Letters #162:265-274) were subcloned into the ars inducible vector. An aliquot of the induced and uninduced culture can also be plated out on agar plates containing an appropriate antibiotic selection but lacking inducer. Following incubation overnight at 37°C, the number of colonies is counted. Any ORF showing bacteriostatic activity will show a lower, but detectable, number of colonies on the agar plates when grown in the presence of inducer as compared to when grown in the absence of inducer. Any ORF showing bacteriocidal activity will show no colonies on the agar plates, when grown in the presence of inducer as compared to when grown in the absence of inducer.
REFERENCES
1. Cohen, MX. (1992). Science 257, 1050-1055.
2. Tenover, F.C. and McGowan Jr., J.E. (1998). Bacterial Infections of Humans. Epidemiology and ControliA.S. Evans and P.S. Brachman, eds.) Plenum Medical Book Company, New York, N.Y. pp. 83-93.
3. Rusterholtz, K., and Pohlschroder, M. (1999). Cell 96, 469-470.
4. Neu, H.C. (1992). Science 257, 1064-1073.
5. Murray, B.E. (1990). Clin. Microbiol. Rev. 3, 46-65.
6. Gray, B.M. (1998). Bacterial Infections of Humans. Epidemiology and Control.(A.S. Evans and P.S. Brachman, eds.) Plenum Medical Book Company, New
York, N.Y. pp. 673 - 711.
• Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989). Molecular cloning: A laboratory Manual. Cold Spring Harbor Laboratory, New York. Cold Spring Harbor
Laboratory Press. 7. Ausubel, F.M. et al. (1994) Current Protocols in Molecular Biology. John Wiley
& Sons, Secaucus, NJ.
8. Rost B,l and Sander C. (1996). Ann. Rev. Biophy. Biomol. Struct. 25, 113-136.
9. Garvey, KJ., Saedi, M.S., and Ito, J. (1985). Gene 40, 311-316.
10. Pickett, G.G. and Peabody, D.S. (1993). Nucl. Acids Res. 21, 4621-4626. 11. Gutierrez, J., Vinos, J., Prieto, I., Mendez, E., Hermoso, J., and Salas, M. (1986). Virology 155, 474-483.
12. Yoshikawa, H., Garvey, K.J., and Ito, J. (1985). Gene 37, 125-130.
13. Martin, A.C., Lopez, R., Garcia, P. (1998). J Bacteriol 180, 210-217.
14. Steiner, M., Lubitz, W., Blasi, U. (1993). J. Bacteriol. 175, 1038-1042.
• Durfee, T., Becherer, K., Chen, P.-L., Yeh, S.-H., Yang, Y., Kilburn, A.E., Lee, W.-H., and Elledge, S.J. (1993). Genes Dev. 7, 555-569.
• Qiu, H., Garcia-Barrio, M.T., and Hinnebusch, A.G. ( 1998). Mol Cell Biol. 18, 2697-2711.
• Katagiri, T., Saito, H., Shinohara, A., Ogawa, H., Kamada, N., Nakamura ,Y., and Miki, Y. (1998). Genes, Chromosomes & Cancer 21, 217-222.
• Endo, T.A., Masuhara, M., Yokouchi, M., Suzuki, R., Sakamoto, H., Mitsui, K., Matsumoto, A., Tanimura, S., Ohtsubo, M., Misawa, H., Miyazaki, T., Leonor N., Taniguchi, T., Fujita, T., Kanakura, Y., Komiya, S., and Yoshimura, A. (1997). Nature 387, 921-924.
• Karimova, G., Pidoux, J., Ullmann, A., Ladant, D. (1998) Proc. Natl. Acad. Sci. 95, 5752-5756.
• Sopta, M., Carthew, R.W., and Greenblatt, J. (1995) J. Biol. Chem. 260, 10353- 10369.
• Qin, J., Fenyo, D., Zhao, Y., Hall, W.W., Chao, D.M., Wilson, C.J., Young, R.A. and Chait, B.T. (1997). Anal. Chem. 69, 3995-4001.
• Swanstrom, M. and Adams, M.H. (1951). Proc. Soc. Exptl. Biol. Med. 78, 372- 375.
Example 15. Growth of Streptococcus bacteriophage Dp-1 and purification of genomic DNA.
The Streptococcus pneumoniae R6 propagating strain (PS) (Tomasz, 1966) was used as host to propagate its respective phage Dp-1 (McDonnell et al., 1975). (Alternatively, Streptococcus (Diplococcus) pneumoniae R36A could be used. Strain R36A is available from ATCC as #11733 or 27336. Streptococcus pneumoniae is also available from Felix d'Herelle Reference Center in Quebec, Canada as catalog number HER 1054. Other S. pneumoniae strains are also available from ATCC.) Two rounds of plaque purification of phage Dp-1 were performed on soft agar essentially as described in Sambrook et al (1989). Briefly, the Streptococcus R6 PS strain was grown overnight at 37°C in K-Cat media [K-Cat: 10 g Bacto casitone, 5 g _. Bacto tryptone, 1 g Yeast extract, 5g Potassium chloride, 0.2% Glucose, 30mM~ Potassium phosphate buffer [pH 8] and 250,000 Units Catalase per liter (Boehringer Mannheim #10683600). The culture was then diluted 20 fold in K-CAT and
incubated at 37°C until the OD540= 0.2 (early log phase) with constant agitation. In order to obtain single plaques, Dp-1 phage was subjected to 10-fold serial dilutions using the phage buffer (100 mM Tris-HCl [pH 7.5], 100 mM NaCl and 10 mM MgCl2)and 10 μl of each dilution was used to infect 0.5 ml of the cell suspension. After incubation of 15 min at 37°C, 2 ml of melted soft agar (K-CAT supplemented with 0.8%) of agar) were added to the mixture and poured onto the surface of 100 mm K-CAT agar plates [K-CAT supplemented with 1.2 % of agar]. After solidification of the soft agar layer, an additional 5 ml of melted soft agar was added to visualize distinct plaques (Ronda et al., 1978). After overnight incubation at 37°C, a single plaque was isolated, resuspended in 1 ml of phage buffer by end over end rotation for 2 hrs at room temperature, and the phage suspension was diluted and used for a second infection as described above. After overnight incubation at 37°C, a single plaque was isolated and used as a stock for all subsequent manipulations.
The propagation procedure for bacteriophage Dp-1 was modified from the agar layer method of Swanstδrm and Adams (1951). Briefly, the R6 strain of
Streptococcus pneumoniae was grown to stationary phase overnight at 37°C in K- CAT. The culture was then diluted 20 fold in K-CAT and incubated at 37°C until the OD540= 0.2. The suspension (15xl07 Bacteria) was then mixed with 15xl05 plaque forming units (pfu) to give a ratio of lOO-bacteria/pfu. After incubation of 15 min at 37°C, 7.5 ml of melted soft agar (K-CAT plus 0.8%> agar) were added to the mixture and poured onto the surface of 150 mm K-CAT agar plates and incubated 16 hrs at 37°C. After solidification of the soft agar layer, 7.5 ml of melted soft agar were added to each plate. To collect the plate lysate, 20 ml of K-CAT media were added to each plate and the soft agar layers were collected by scrapping off with a clean microscope slide followed by vigorous shaking of the agar suspension for 5 min to break up the agar. The mixture was then centrifuged for 10 min at 4,000 φm (2,830 xg) using a JA-10 rotor (Beckman) and the supernatant (lysate) was collected and subjected to a treatment with 10 μg /ml of DNase I and RNase A for 30 min at 37°C. To precipitate the phage particles, the phage suspension was adjusted to 10%. (w/v) of PEG 8000 and 0.5 M of NaCl followed by incubation at 4°C for 16 hrs. The phage was recovered by centrifugation at 4,000 φm (3,500 xg) for 20 min at 4°C on a GS-6R table top centrifuge (Beckman). The pellet was resuspended with 2 ml of phage buffer (100 mM Tris-HCl [pH 7.5], 100 mM NaCl and 10 mM MgCl2). The phage suspension was extracted with 1 volume of chloroform and further purified by centrifugation on a cesium chloride step gradient as described in Sambrook et al. (1989), using a TLS-55 * rotor and centrifuged in an Optima TLX ultracentrifuge (Beckman) for 2 hrs at 28,000 φm (67,000 xg) at 4°C. Banded phage was collected and ultracentrifuged again on an
isopycnic cesium chloride gradient (1.45 g/ml) at 40,000 φm (64,000 xg) for 24 hrs at 4°C using a TLV rotor (Beckman). The phage was harvested and dialyzed for 4 hrs at room temperature against 4 L of dialysis buffer consisting of 10 mM NaCl, 50 mM Tris-HCl [pH 8] and 10 mM MgCl2. Phage DNA was prepared from the phage suspension by adding 20 mM EDTA, 50 μg/ml Proteinase K and 0.5% SDS and incubating for 1 hr at 65°C, followed by successive extractions with 1 volume of phenol, 1 volume of phenol-chloroform and 1 volume of chloroform. The DNA was then dialyzed overnight at 4°C against 4 L of TE (10 mM Tris-HCl [pH 8.0], lmM EDTA).
Example 16. DNA sequencing of the Bacteriophage Dp-1 genome.
Four micrograms of phage DNA was diluted in 200 μl of TE (10 mM Tris, [pH 8.0], 1 mM EDTA) in a 1.5 ml eppendorf tube and sonication was performed (550 Sonic Dismembrator, Fisher Scientific). Samples were sonicated under an amplitude of 3 μm with bursts of 5 sec spaced by 15 sec cooling in ice/water for 3 to 4 cycles. The sonicated DNA was then size fractionated by electrophoresis on 1%> agarose gels utilizing TAE (1 x TAE is: 40 mM Tris-acetate, 1 mM EDTA [pH 8.0]) as the running buffer. Fractions ranging from 1 to 2 kbp were excised from the agarose gel and purified using a commercial DNA extraction system according to the instructions of the manufacturer (Qiagen), with a final elution of 50 μl of 1 mM Tris [pH 8.5].
The ends of the sonicated DNA fragments were repaired with a combination of T4 DNA polymerase and the Klenow fragment ofE. coli DNA polymerase I, as follows. Reactions were performed in a reaction mixture (final volume, 100 μl) containing sonicated phage DNA, 10 mM Tris-HCl [pH 8.0], 50 mM NaCl, 10 mM MgCl2, 1 mM DTT, 50 μg/ml BSA, 100 μM of each dNTP and 15 units of T4 DNA polymerase (New England Biolabs) for 20 min at 12°C followed by addition of 12.5 units of the Klenow large fragment of DNA polymerase I (New England Biolabs) for 15 min at room temperature. The reaction was stopped by two phenol/chloroform extractions and the DNA was precipitated with ethanol and the final DNA pellet resuspended in 20 μl of H2O.
Blunt-ended DNA fragments were cloned by ligation directly into the Hinc II site of the pKSII+ vector (New England Biolabs) dephosphorylated by treatment with calf intestinal alkaline phosphatase (New England Biolabs). A typical ligation reaction contained 100 ng of vector DNA, 2 to 5 μl of repaired sonicated phage PNA (50-100 ng) in a final volume of 20 μl containing 800 units of T4 DNA ligase (New England Biolabs) and was incubated overnight at 16°C. Transformation and selection
of bacterial clones containing recombinant plasmids was performed in E. coli DHlOβ according to standard procedures (Sambrook et al, 1989).
Recombinant clones were picked from agar plates into 96-well plates containing 100 μl LB and 100 μg/ml ampicillin and incubated at 37°C. The presence of phage DNA insert was confirmed by PCR amplification using T3 and T7 primers flanking the Hinc II cloning site of the pKS vector. PCR amplification of the potential foreign inserts was performed in a 15 μl reaction volume containing 10 mM Tris (pH 8.3), 50 mM KCl, 1.5 mM MgCl2, 0.02% gelatin, 1 μM primer, 187.5 μM each dNTP, and 0.75 units Taq polymerase (BRL). The thermocycling parameters were as follows: 2 min initial denaturation at 94°C for 2 min, followed by 20 cycles of 30 sec denaturation at 94°C, 30 sec annealing at 58°C, and 2 min extension at 72°C, followed by a single extension step at 72°C for 10 min. Clones with insert sizes of 1 to 2 kbp were selected and plasmid DNA was prepared from the selected clones using the QIAprep™ spin miniprep kit (Qiagen). The nucleotide sequence of the extremities of each recombinant clone was determined using an ABI 377-36 automated sequencer with two types of chemistry: ABI prism Big Dye™ primer cycle sequencing (21M13 primer: #403055)(M13RΕV primer: #403056) or ABI prism Big Dye™ terminator cycle sequencing ready reaction kit (Applied Biosystems; #4303152). To ensure co-linearity of the sequence data and the genome, all regions of the phage genome were sequenced at least once from both directions on two separate clones. In areas that this criteria was not initially met, a sequencing primer was selected and phage DNA was used directly as sequencing template employing ABI prism Big Dye™ terminator cycle sequencing ready reaction kit.
Example 17. Bioinformatic management of primary nucleotide sequence.
Sequence contigs were assembled using Sequencher™ 3.1 software (GeneCodes). To close contig gaps, sequencing primers were selected near the edge of the contigs. Phage DNA was used directly as sequencing template employing ABI prism BigDye™ terminator cycle sequencing ready reaction kit (Applied Biosystems; #4303152). The complete sequence oϊ Streptococcus bacteriophage Dp-1 is shown in Table 28.
A software program was used on the assembled sequence of bacteriophage Dp-1 to identify all putative ORFs larger than 33 codons. The software scans the primary nucleotide sequence starting at nucleotide #1 for an appropriate start codorrr Three possible selections can be made for defining the nature of the start codon; I) selection of ATG, II) selection of ATG or GTG, and III) selection of either ATG,
GTG, TTG, CTG, ATT, ATC, and ATA. This latter initiation codon set corresponds to the one reported by the NCBI(http://www.ncbi.nlm.nih.gov/htbin- post/Taxonomv/wprintgc?mode=:c') for the bacterial genetic code. When an appropriate start codon is encountered, a counting mechanism is employed to count the number of codons (groups of three nucleotides) between this start codon and the next stop codon downstream of it. If a threshold value of 33 is reached, or exceeded, then the sequence encompassed by these two codons is defined as an ORF. This procedure is repeated, each time starting at the next nucleotide following the previous stop codon found, in order to identify all the other putative ORFs. The scan is performed on all three reading frames of both DNA strands of the phage sequence. The predicted ORFs for bacteriophage Dp-1 are listed in Tables 29 and 30, and Fig. 6.
Sequence homology searches for each ORF were carried out using an implementation of BLAST programs. Downloaded public databases used for sequence analysis include: (i) non-redundant GenBank (ftp://ncbi.nlm.nih.gOv/blast/db/nr.Z), ii) Swissprot (ftp://ncbi.nlm.nih.gOv/blast/db/swissprot.Z); iii) vector (ftp://ncbi.nlm.nih.gOv/blast/db/vector.Z); iv) pdbaa databases (ftp://ncbi.nlm.nih.gOv/blast/db/pdbaa.Z); v) staphylococcus aureus NCTC 8325 (ftp://ftp.genome.ou.edu/pub/staph staph-lk.fa); vi) streptococcus pyogenes (ftp://ftp.tigr.org/pub/data s_pneumoniae/gsp.contigs.112197.Z); vii) PRODOM (ftp://ftp.toulouse.inra.fr/pub/prodom/current release/prodom99.1.forblast.gz .: viii) DOMO rftp://ftp.infobiogen.fr/pub/db/domo : ix) TREMBL (ftp://www.expasy.ch/databases/sp_tr_nrdb/fasta/)
The results of the homology searches performed on the ORFs of bacteriophage
Dp-1 are shown in Table 31.
Example 18. Sub-Cloning of Bacteriophage Dp-1 ORFs.
Preparation of the shuttle expression vector
Expression preferably utilizes a shuttle expression vector which is arranged such that expression of the exogenous bacteriophage Dp-1 ORF sequence is inducible. For example, the plasmid pLSE4 replicates in E. coli, and S. pneumoniae (Diaz- and Garcia, 1990). This plasmid can be modified by conventional techniques to insert the inducible arsenite promoter, derived from the shuttle vector pT0021, in which the
firefly luciferase (lucFF) expression is controlled by the ars promoter/operator from a S. aureus plasmid (Tauriainen, S., Kaφ, M., Chang, W and Virta, M. (1997). Recombinant luminescent bacteria for measuring bioavailable arsenite and antimonite. Appl. Environ. Microbiol. 63:4456-4461). This modified shuttle vector will contain the ars promoter, arsR gene and a cloning site for introduction of individual phage ORFs downstream from a shine-dalgarno sequence.
Other inducible regulatory sequences can be utilized instead of the arsenite- inducible system. An example is a nisin-inducible system The nisA promoter activity is dependent on the proteins NisR and NisK, which constitute a two-component signal transduction system that responds to the extracellular inducer nisin. The nisin sensitivity and inducer concentration required for maximal induction varies among the strains, but is functional in Streptococcus pyogenes, Streptococcus agalactiae, Streptococcus pneumoniae, Enterococcus faecalis, and Bacillus subtilis. Significant induction of the nisA promoter (10- to 60-fold induction) can be obtained in all of the species. A vector containing this promoter was published as Eichenbaum Z, Federle MJ, Marra D, de Vos WM, Kuipers OP, Kleerebezem M, and Scott JR (1998) Appl Environ Microbiol 64, 2763-2769. Other vectors, e.g., plasmids, can also be utilized which will allow replication and transcription in Streptococcus.
Alternatively, a constitutive promoter can be used to drive expression of the introduced ORF, and compare cell growth to control bacterial cells containing the parental vector lacking any introduced phage ORF. Recombinant plasmids are introduced into S. pneumoniae R6 as previously described (Diaz and Garcia, 1990)
Cloning of ORFs with a Shine-Dalgarno sequence ORFs with a Shine-Dalgarno sequence are selected for functional analysis of bacterial killing. Each ORF, from initiation codon to last codon (excluding the stop codon), will be amplified by PCR from phage genomic DNA. For PCR amplification of ORFs, each sense strand primer starts at the initiation codon and is preceded by a restriction site and each antisense strand starts at the last codon (excluding the stop codon) and is preceded by a different restriction site. The PCR product of each ORF will be gel purified and digested with the restriction enzymes with sites contained on the PCR oligonucleotides. The digested PCR product is then gel purified using the Qiagen kit, ligated into the modified shuttle vector, and used to transform bacterial strain DHlOβ. Recombinant clones are then picked and their insert sizes confirmed by PCR analysis using primers flanking the cloning site as well as restriction- — digestion. The sequence fidelity of cloned ORFs will be verified by DNA sequencing using the same primers as used for PCR. In the cases that the verification of ORFs can not be achieved by one path of sequencing using primers flanking the cloning site
internal primers will be selected and used for sequencing. Recombinant plasmids will be introduced into S. pneumoniae R6 as previously described (Diaz and Garcia, 1990). Induction of gene expression from the ars promoter.
If an inducible promoter is used, e.g., the ars promoter, induction can be assessed, for example, in either of the two methods.
1. Screening on agar plates
The functional identification of killer ORFs can be performed by spreading an aliquot of S. pneumoniae transformed cells containing phage Dp-1 ORFs onto agar plates containing different concentrations of sodium arsenite (0; 2.5; 5; and 7.5 μM). The plates are incubated overnight at 37°C, after which a growth inhibition of the ORF transformants on plates that contain arsenite are compared to plates without arsenite.
2. Quantification of growth inhibition in liquid medium
Cells containing different recombinant plasmids can be grown for overnight at 37°C in LB medium supplemented with the appropriate antibiotic selection. These are then diluted to the mid log phase (OD540=.2) with fresh media containing antibiotic and transferred to 96-well microtitration plates (100 μl/well). Inducer is then added at different final concentrations (ranging from 2.5 to 10 μM) and the culture incubated for an additional 2 hrs at 37°C. The effect of expression of the phage Dp-1 ORFs on bacterial cell growth is then monitored by measuring the OD540 and comparing the rate of growth to the culture not containing inducer. [As positive controls for growth inhibition, the kilA gene of phage lambda (Reisinger, GR., Rietsch, A., Lubitz, W. and Blasi, U. 1993 Virology #193: 1033-1036), and the holin/lysin genes of the Sthaphylococcus aureus phage Twort (Loessner, MJ., Gaeng, S., Wendlinger, G., Maier, SK. and Scherer, S. 1998. FEMS Microbiology Letters #162:265-274) can be subcloned into the ars inducible vector. An aliquot of the induced and uninduced culture can also be plated out on agar plates containing an appropriate antibiotic selection but lacking inducer. Following incubation overnight at 37°C, the number of colonies is counted. Any ORF showing bacteriostatic activity will show a lower, but detectable, number of colonies on the agar plates when grown in the presence of inducer as compared to when grown in the absence of inducer. Any ORF showing full bacteriocidal activity will show no colonies on the agar plates, when grown in the presence of inducer as compared to when grown in the absence of inducer.
REFERENCES
15. Cohen, M.L. (1992) Science 257, 1050-1055.
16. Tenover, F.C. and McGowan Jr., J.E. (1998) Bacterial Infections of Humans. Epidemiology and ControUA.S. Evans and P.S. Brachman, eds.) Plenum Medical Book Company, New York, N.Y. pp. 83-93. 17. Rusterholtz, K., and Pohlschroder, M. (1999) Cell 96, 469-470.
18. Klugman, K.P. (1990) Clin. Microbiol. Rev. 3, 171-196.
19. Fenoll, A., Martin Bourgon, C, Munoz, R., Vicioso, D., Casal, J. (1991) Rev. Infect. Disease 13, 56-60.
20. Jorgensen, J.H., Doern, G. V., Maher, L. A., Howell, A. W., Redding, J. S. (1990) Antimicrob. Agents Chemother. 34, 2075-2080. 21. Neu, H.C. (1992) Science 257, 1064-1073.
Hsueh, P. R., Wu, J. J., Hsiue, T. R. (1996) J Formos Med Assoc5, 364-371.
Garcia, P., Martin, A.C., and Lopez, R. (1997) Microbial Drug Res. 3, 165-176.
Martin, A.C., Lopez, R., and Garcia, P. (1996) J. Virol. 70, 3678-3687.
Sheehan, M.M., Garcia, J.L., Lopez, R., and Garcia, P. (1997) Mol. Microbiol. 25,
717-725.
Kodaira, M., Biswas, S.B., and Kornberg, A. (1983) Mol. Gen. Genet. 192, 80-96.
Maki, S. and Kornberg, A. (1988) J. Biol. Chem.263, 6547-6554.
Tsuchihashi Z, Kornberg A. (1990) Proc. Natl. Acad. Sci. USA. 87, 2516-2520.
Lee, S.H. and Walker, J.R. (1987) Proc Natl Acad Sci USA 84, 2713-2717.
Smidt, C.R., Steinberg, F.M., Rucker, R. (1991) Proc Soc Exp Biol Med 197, 19-
26.
Frank, D.W, (1997) Mol Microbiol. 26, 621-629.
Nardese, V., Gutlich, M., Brambilla, A., Carbone, M.L.(1996) Biochem Biophys Res Commun 218, 273-279.
Mancini, R., Saracino, F., Buscemi, G., Fischer, M., Schramek, N., Bracher, A., Bacher, A., Gutlich, M., Carbone, M.L. (1999) Biochem Biophys Res Commun 255,521-527.
Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) Molecular cloning. *A laboratory Manual. Cold Spring Harbor Laboratory, New York. Cold* Spring Harbor Laboratory Press.
22. Ausubel, F.M. et al. (1994) Current Protocols in Molecular Biology. John Wiley & Sons, Secaucus, NJ.
23. Rost B,l and Sander C. (1996) Ann. Rev. Biophy. Biomol. Struct. 25, 113-136.
24. Garvey, K.J., Saedi, M.S., and Ito, J. (1985) Gene 40, 311-316.
25. Pickett, G.G. and Peabody, D.S. (1993) Nucl. Acids Res. 21, 4621-4626. 26. Gutierrez, J., Vinos, J., Prieto, I., Mendez, E., Hermoso, J., and Salas, M. (1986) Virology 155, 474-483.
27. Yoshikawa, H., Garvey, K.J., and Ito, J. (1985) Gene 37, 125-130. 28. Martin, A.C., Lopez, R., Garcia, P. (1998) J Bacteriol 180, 210-217.
29. Steiner, M., Lubitz, W., Blasi, U. (1993) J. Bacteriol. 175, 1038-1042.
• Durfee, T., Becherer, K., Chen, P.-L., Yeh, S.-H., Yang, Y., Kilburn, A.E., Lee, W.-H., and Elledge, S J. (1993). Genes Dev. 7, 555-569.
• Qiu, H., Garcia-Barrio, M.T., and Hinnebusch, A.G. (1998) Mol Cell Biol. 18, 2697-2711. • Katagiri, T., Saito, H., Shinohara, A., Ogawa, H., Kamada, N., Nakamura ,Y., and Miki, Y. (1998) Genes, Chromosomes & Cancer 21, 217-222.
• Endo, T.A., Masuhara, M., Yokouchi, M., Suzuki, R., Sakamoto, H., Mitsui, K., Matsumoto, A., Tanimura, S., Ohtsubo, M., Misawa, H., Miyazaki, T., Leonor N., Taniguchi, T., Fujita, T., Kanakura, Y., Komiya, S., and Yoshimura,
A. (1997) Nature 387, 921-924.
• Karimova, G., Pidoux, J., Ullmann, A., Ladant, D. (1998) Proc. Natl. Acad. Sci. 95, 5752-5756.
• Sopta, M., Carthew, R.W., and Greenblatt, J. (1995) J. Biol. Chem. 260, 10353- 10369.
• Qin, J., Fenyo, D., Zhao, Y., Hall, W.W., Chao, D.M., Wilson, C J., Young, R.A. and Chait, B.T. (1997) Anal. Chem. 69, 3995-4001.
• Tomasz, A. (1966) Journal of Bacteriology 91, 1050-1061.
• McDonnell, M., Ronda, LC and Tomasz, A. (1975) Virology 63, 577-582.
• Ronda C, Lopez, R, Tomasz, A. and Portoles A. (1978) 26, 221-225.
• Swanstrόm, M. and Adams, M.H. (1951) Proc. Soc. Exptl. Biol. Med. 78, 372- 375. Diaz E and Garcia JL. (1990) Gene 90, 163-167. • Tauriainen, S., Kaφ, M., Chang, W and Virta, M. (1997). Recombinant luminescent bacteria for measuring bioavailable arsenite and antimonite. Appl. Environ. Microbiol. 63:4456-4461.
All patents and publications mentioned in the specification are indicative of the levels of skill of those skilled in the art to which the invention pertains. All references cited in this disclosure are incoφorated by reference to the same extent as if each reference had been incoφorated by reference in its entirety individually.
One skilled in the art would readily appreciate that the present invention is well adapted to carry out the objects and obtain the ends and advantages mentioned, as well as those inherent therein. The specific methods and compositions described herein as presently representative of preferred embodiments are exemplary and are not intended as limitations on the scope of the invention. Changes therein and other uses will occur to those skilled in the art which are encompassed within the spirit of the invention are defined by the scope of the claims. It will be readily apparent to one skilled in the art that varying substitutions and modifications may be made to the invention disclosed herein without departing from the scope and spirit of the invention. For example, those skilled in the art will recognize that the invention may suitably be practiced using a variety of different bacteria, bacteriophage, and sequencing methods within the general descriptions provided.
The invention illustratively described herein suitably may be practiced in the absence of any element or elements, limitation or limitations which is not specifically disclosed herein. Thus, for example, in each instance herein any of the terms "comprising," "consisting essentially of and "consisting of may be replaced with either of the other two terms. The terms and expressions which have been employed are used as terms of description and not of limitation, and there is not intention that in the use of such terms and expressions of excluding any equivalents of the features shown and described or portions thereof, but it is recognized that various modifications are possible within the scope of the invention claimed. Thus, it should be understood that although the present invention has been specifically disclosed by preferred embodiments and optional features, modification and variation of the _ - - - concepts herein disclosed may be resorted to by those skilled in the art, and thafsuch modifications and variations are considered to be within the scope of this invention as defined by the appended claims.
In addition, where features or aspects of the invention are described in terms of Markush groups or other grouping of alternatives, those skilled in the art will recognize that the invention is also thereby described in terms of any individual member or subgroup of members of the Markush group or other group. For example, if there are alternatives A, B, and C, all of the following possibilities are included: A separately, B separately, C separately, A and B, A and C, B and C, and A and B and C. Thus, for example, for the bacteria and phage specified herein, the embodiments expressly include any subset or subgroup of those bacteria and/or phage. While each such subset or subgroup could be listed separately, for the sake of brevity, such a listing is replaced by the present description.
Thus, additional embodiments are within the scope of the invention and within the following claims.
Table 1
Phages against human and animal pathogenic bacteria
Infec. Immun. 1982. 35: 343-349
Mol.Gen.Genet 1998.258: 323-325
Aaφ247 Oral Micriol. Immunol 1997.12: 40-46
Actinomyces viscosus 43146-B1 The American Type Culture Collection
Infect.Immun.l985.48:228-233
Infect.Immun.l988.56:54-59
Plasmid 1997.37: 141-153
Aeromonas hydrophila PM2** & PM3 FEMS Microbiol.Lett. 1990.57:277-282
Aehl Felix d'Herelle Reference
Aeh2 Centre.Quebec.Quebec
PM4
PM5
PM6
T7-ah
Bordetella Felix d'Herelle Reference parapertussis Centre,Quebec,Quebec
Mol. Gen. Mikrobiol. Virusol. 1988.4: 22-25
Zh.Mi__robiol.Epidemiol.I___muno. 1987.5:9- 13
41405 Zh.MikrobioI.Epidemiol.Immuno. 1987.5:9- 13
Brucella abortus Felix d'Herelle Reference Centre,Quebec,Quebec
23448-B1 The American Type Culture Collection 23448-B2 23448-B3 17385-B1 17385-B2
10/1
24 11
212/XV
BK-2, TB & Zh.Mikrobiol.Epidemiol.Immunobiol.1983.2: Fi** 48-52
R/c & R/O Dev. Biol. Stand. 1984.56: 55-62
Brucella canis R/c Dev. Biol. Stand. 1984.56: 55-62
Brucella melitensis BK-2 23456-B1 The American Type Culture Collection
Brucella suis Wb Zentralbl. Veterinarmed.1975.22 : 866-867
11.
N4* Vet.Microbiol.l992.30:203-212
Phi 80 trp Ann.Inst.Pasteur.1971.120:121-125
Obeta 1 J.Bacteriol.l978.133:172-177
P1CM J.Gen.Microbiol.1978.107:73-83
PA-2* J.Bacteriol.1990.172: 1660- 1662
186* Mol.Gen.Genet.1982.187:87-95
186.IX.B Mol.Microbiol.1992.6:2629-2642
21" Virology 1983.129:484-489
P4* MicrobiolRev.1993.57:683-702
82* J.Biol.Chem.1987.262:11721-11725
PSP3 J.Bacteriol.1996.178:5668-5675
HK022* Nucleic Acids Res.1994.22:354-356
D108* Nucleic Acids Res.l986.14:3813-3825
Escherichia coli Rb49 J.Mol.Biol.1997.267:237-249 (Cont'd) Ike** J.Mol.Biol.l 985.181:27-39
P22dis Mol.Gen.Genet.1978.166:233-243
N15* J.Bacteriol.1996.178 : 1484-1486
Ifl" Proc.R.Soc.Lond.B.Biol.Sci.l991.245:23-30
Stx2Phi-I & Infec..Immun.l998.66:4100-4107 Stx2Phi-II
18 Virology 1987.156:122-126
X J.Gen.Microbiol.1981.126:389-396
AC3 Mol.Microbiol.1991.5:715-725
HER 317 Felix d'Herelle Refrence HER 330 Centre,Quebec,Quebec HER 333 HER 335 HER 334 HER 331 HER 316
Legendre Leo Roy Sedge
Mol.Microbiol.1993.7:395-405
J.Mol.Biol.l998.279:143-164
Proc.Natl.Acad.Sci USA.1988.84:2833-2837
Mol.Biol.Rep. 1981.30:11-15
Proc.Natl.Acad.Sci.USA 1997.94: 10961- 10966
29M, 31M, 122, Arch. Virol.1993.133:39-49 & 154, 37, 29D, 46, Am.Rev.Respir.Dis.1975.112: 17-22 139,110, 141, 74D, AG1 & DS6A
Mycobacterium 23052-B1 The American Type Culture Collection fortuitum 27207-B1
Bo 4 27207-B2
Bo 7
Mycobacterium leprae Ann.Microbiol. (Paris) 1982.133:93-97
Mycobacterium 25618-B1 The American Type Culture Collection tuberculosis 25618-B2
DS6A 4243-B1
110, 139 & 33D Arch. Virol.1993.133 :39-49
AG1.GS4E, The Biology of Mycobacteria. Academic BG1, Press,Toronto 1982 (Ratledge & Stanford) PH & BK1 1982.309-351
Mycobacterium sp Phagus pellegrini 11760-B1 The American Type Collection Culture
NN 11761-B1
Bl 23239-B1
Pseudomonas 12175-B1 The American Type Culture Collection aeruginosa 2 12175-B2
2A 12175-B3
2B 12175-B4
11 14205-B1
16 14206-B1
24 14207-B1
27 14208-B1
44 14209-B1
73 14210-B1
95 14211-B1
109 14212-B1
113 14213-B1
249 14214-B1
B3 15692-B1
Hoff2 14203-B1
Hoff3 14204-B1
Pa 12055-B1
Pb 12055-B2
PB-1 15692-B3
Pc 12055-B3
Pf 25102-B1
PP7** 15692-B2
Felix d'Herelle Reference Centre,Quebec,Quebec
7 & 31
PD" J.Virol.1983.47:221-223 φ-MC Can.J.Microbiol.l969.15:1179-1186
Pfl" J.Mol.Biol.1991.218 :349-364
PR4* J.Gen. Virol.1979.43 :583-592
A7 J.Bacteriol.l992.174:2407-2411
KF1 J.Biochem.1983.93:61-71
( TX** Mol.Microbiol.1993.4: 1703- 1709 f_* J.Virol.l977.24:135-141
Pseudomonas 297,309,318, Arch.Virol.1993.131:141-151 aeruginosa 11,
(Cont'd)
HER 101 Felix d'Herelle Reference HER 239 Centre,Quebec,Quebec HER 283 HER 49
Twort** φll* J.Bacteriol.1988.170:2409-2411
φl3** & φ42* J.Gen..Microbiol.l989.135:1679-1697
L54a* J.Bcteriol.1986.166:385-391
80α* Can.J.Microbiol.l996.43:612-616
94,95 & 96 J.Clin.Microbiol.1988.26:2395-2401 φl31,A3 & A5 Staphylococci & Staphylococcal Infections.1997. Voll:503-508 (Karger.Basel)
Phi PVL** Gene 1998.215:57-67
Staphylococcus BaSTC2 Felix d'Herelle Reference carnosus Centre,Quebec,Quebec
Staphylococcus la, 2b, 3a, 4b, Can.J.Microbiol.l988.34:1358-1361 epidermidis 5a, 6b, 7b, 8c, 9a, 10a, l ib, 12a & 13b
41, 63, 11811, Res.Virol.l994.145:l l l-121
138,
245, 336, 392 &
550
Staphylococcus 1154A, 1405, Res.Virol.1990.141: 625-635 & saprophyticus 1314, 1139 & Res.Virol.l994.145:l l l-121 1259
Staphylococcus sp. Phi 812, Phi 131, Virology 1998.246:241-252 SK311 & U16
Streptococcus faecalis VD13 HER44 Felix d'Herelle Reference Centre,Quebec,Quebec
Streptococcus faecium PE1 Zentralbl.Bakteriol.l975.231:421-425
Streptococcus oralis Cp-l** & Cp- FEMS Microbiol.Lett.l989.65:187-192
7**
13.
Table 2
>Bacteriophage 77, complete genome sequence, 1708 nucleotides
1 gatcaaaata cttggggaac ggttagggag taaacttcgc gataatttta aaaattcatg
61 tataaccccc ctcttataac cattttaagg caggtgatga aatggagatt atagtcgatg
121 aaaatttagt gcttaaagaa aaagaaaggc tacaagtatt atataaagac atacctagca
181 ataaattaaa agtagttgat ggtttaatta ttcaagcagc aaggctacgt gtaatgcttg
2 1 attacatgtg ggaagacata aaagaaaaag gtgattatga tttatttact caatctgaaa
301 aggcgccacc atatgaaagg gaaagaccag tagccaaact atttaatgct agagatgctg
361 catatcaaaa aataatcaaa caattatcgg atttattgcc cgaagagaaa gaagacacag 21 aaacgccatc tgatgattac ctatgattag taataaatac gttgatgaat atataaattt
481 gtggaaacaa ggaaagataa ttttaaataa agaaagaatt gatctcttta attatctaca
541 aaaacatata tattcacgag atgatgtata ttttgatgaa cagaaaatcg aggattgtat
601 caaatttatt gaaaaatggt attttccaac attaccattt caaaggttta tcatagctaa
661 tatatttctt atagataaaa atacagatga agctttcttt acagaatttg ctattttcat
721 gggacgtgga ggcgggaaaa acggtctaat aagtgctatt agtgattttc tttctacgcc
781 cttacacgga gttaaagaat atcacatctc cattgttgct aatagtgaag atcaagcaaa
841 aacatcgttt gatgaaatca gaaccgtttt aatggataac aaacgaaata agacgggtaa
901 aacgccaaaa gctccttatg aagttagtaa agcaaaaata ataaaccgtg caactaaatc
961 ggttattcga tataacacat caaacacaaa aaccaaagac ggtggacgtg aggggtgtgt
1021 tatttttgat gaaattcatt atttctttgg tcctgaaatg gtaaacgtca aacgtggtgg
1081 attaggtaaa aagaaaaata gaagaacgtt ttatataagt actgatggtt ttgttagaga
1141 gggttatatc gatgcaatga agcacaaaat tgcaagtgta ttaagtggca aggttaaaaa
1201 tagtagattg tttgcttttt attgtaagtt agacgatcca aaagaagttg atgacagaca
1261 gacgtgggaa aaggcgaacc caatgttaca taaaccgtta tcagaatacg ctaaaacact
1321 gctaagcacg attgaagaag aatataacga tttaccattc aaccgttcaa ataagcccga
1381 attcatgact aagcgaatga atttgcctga agttgacctt gaaaaagtaa tagcaccatg
1441 gaaagaaata ctagcgacta atagagagat accaaattta gataatcaaa tgtgtattgg
1501 tggtttagac tttgcaaaca ttcgagattt tgcaagtgta gggctattat tccgaaaaaa
1561 cgatgattac atttggttag gacattcgtt tgtaagacaa gggtttttgg atgatgtcaa
1621 attagaacct cctattaaag aatgggaaaa aatgggatta ttgaccattg tcgatgatga
1681 tgtcattgaa attgaatata tagttgattg gtttttaaag gctagagaaa aatatgggct
1741 tgaaaaagtc atagctgata attatagaac tgatattgta agacgtgcgt ttgaggatgc
1801 tggcataaaa cttgaagtac ttagaaatcc aaaagcaata catggattac ttgcaccacg
1861 tatcgataca atgtttgcga aacataacgt aatatatgga gacaatcctt tgatgcgttg
1921 gtttactaat aatgttgctg taaaaatcaa gccggatgga aataaagagt atatcaaaaa
1981 agatgaagtc agacgtaaaa cggatggatt catggctttt gttcacgcat tatatagagc
2041 agacgatata gtagacaaag acatgtctaa agcgcttgat gcattaatga gtatagattt
2101 ctaatagagg aggtgagaca tgagtattct agaaaagata tttaaaacta ggaaagatat
2161 aacatatatg cttgatttag atatgataga agatctatca caacaagcgt atgtgaaacg
2221 tttagcgatt gatagttgta ttgaatttgt tgcgcgagct gtcgctcaaa gtcattttaa
2281 agtattggaa ggtaatagaa ttcaaaagaa tgatgtttac tacaagttaa atataaaacc
2341 aaatactgac ttatcaagcg atagtttttg gcaacaagtt atatataaac taatttatga
2401 taacgaggtt ttaatcgtag taagtgacag caaagaatta cttatcgcag atagctttta
2461 cagagaagag tacgctttgt atgatgatat attcaaagat gtaacggtta aagattatac
2521 ttatcaacgt actttcacaa tgcaagaggt catatattta aagtacaaca acaataaagt
2581 gacacacttt gtagaaagtc tattcgaaga ttacgggaaa atattcggaa gaatgatagg
2641 tgcacaatta aaaaactatc aaataagagg gattttgaaa tctgcctcta gcgcatatga
2701 cgaaaagaat atagaaaaat tacaagcgtt cacaaataaa ttattcaata cttttaataa
2761 aaatcaacta gcaatcgcgc ctttgataga aggttttgat tatgaggaat tatctaatgg
2821 tggtaagaat agtaacatgc ctttttctga attgagtgag ctaatgagag atgcaataaa
2881 aaatgttgcg ttgatgattg gtatacctcc aggtttgatt tacggagaaa cagctgattt
2941 ggaaaaaaac acgcttgtat ttgagaagtt ctgtttaaca cctttattaa aaaagattca
3001 gaacgaatta aacgcgaaac tcataacaca aagcatgtat ttgaaagata caagaataga
3061 aattgtcggt gtgaataaaa aagacccact tcaatatgct gaagcaattg acaaacttgt
3121 aagttctggt tcatttacaa ggaatgaggt gcggattatg ttaggtgaag aaccatcaga
3181 caatcctgaa ttagacgaat acctgattac taaaaactac gaaaaagcta acagtggtga
3241 aaatgatgaa aaagaaaaag atgaaaacac tttgaaaggt ggtgatgaag atgaaagcg -
3301 agattaaagg cgtcatcgtt tccaacgaag ataaatgggt ttacgaaatg cttggtatgg
3361 attcgacttg tcctaaagat gttttaacac aactagaatt tagtgatgaa gatgttgata
3421 ttataattaa ctcaaatggt ggtaacctag tagctggtag tgaaatatat acacatttaa
3481 gagctcataa aggcaaagtg aatgttcgta tcacagcaat agcagcaagt gcggcatcgc
3541 ttatcgcaat ggctggtgac cacatcgaaa tgagtccggt tgctagaatg atgattcaca
3601 atccttcaag tattgcgcaa ggagaagtga aagatctaaa tcatgctgca gaaacattag
3661 aacatgttgg tcaaataatg gctgaggcat atgcggttag agctggtaaa aacaaacaag
3721 aacttataga aatgatggct aaggaaacgt ggctaaatgc tgatgaagcc attgaacaag
3781 gttttgcgga tagtaaaatg tttgaaaacg acaatatgca aattgtagca agcgatacac
3841 aagtgttatc gaaagatgta ttaaatcgtg taacagcttt ggtaagtaaa acgccagagg
3901 ttaacattga tattgacgca atagcaaata aagtaattga aaaaataaat atgaaagaaa
3961 aggaatcaga aatcgatgtt gcagatagta aattatcagc aaatggattt tcaagattcc
4021 ttttttaata caaaaatagg aggtcataaa atgactataa atttatcgga aacattcgca
4081 aatgcgaaaa acgaatttat taatgcagta aacaacggtg aaccgcaaga aagacaaaat
4141 gaattgtacg gtgacatgat taaccaacta tttgaagaaa ctaaattaca agcaaaagca
4201 gaagctgaaa gagtttctag tttacctaaa tcagcacaaa ctttgagtgc aaaccaaaga
4261 aatttcttta tggatatcaa taagagtgtt ggatataaag aagaaaaact tttaccagaa
4321 gaaacaattg atagaatctt cgaagattta acaacgaatc atccattatt agctgactta
4381 ggtattaaaa atgctggttt gcgtttgaag ttcttaaaat ccgaaacttc tggcgtggct
4441 gtttggggta aaatctatgg tgaaattaaa ggtcaattag atgctgcgtt cagtgaagaa
4501 acagcaattc aaaataaatt gacagcgttt gttgttttac caaaagattt aaatgatttt
4561 ggtcctgcgt ggattgaaag atttgttcgt gttcaaatcg aagaagcatt tgcagtggcg
4621 cttgaaactg cgttcttaaa aggtactggt aaagaccaac cgattggctt aaaccgtcaa
4681 gtacaaaaag gtgtatcggt aactgatggt gcttatccag agaaagaaga acaaggtacg
4741 cttacatttg ctaatccgcg cgctacggtt aatgaattga cgcaagtgtt taaataccac
4801 tcaactaacg agaaaggtaa atcagtagcg gttaaaggta atgtaacaat ggttgttaat
4861 ccgtccgatg cttttgaggt tcaagcacag tatacacatt taaatgcaaa tggcgtatat
4921 gttactgctt taccatttaa tttgaatgtt attgagtcta cagttcaaga agcaggtaag
4981 gttttaacgt acgttaaagg tctatatgat ggttatttag ctggtggtat taatgttcag
5041 aaatttaaag aaacacttgc gttagatgat atggatttat acactgcaaa acaatttgct
5101 tacggcaaag cgaaagataa taaagttgct gctgtttgga aattagattt aaaaggacat
5161 aaaccagctt tagaagatac cgaagaaaca ctataaaatt ttatgaggtg ataaaatggt
5221 gaaatttaaa gttgttagag aatttaaaga catagagcac aatcaacaca agtacaaagt
5281 aggggagttg tatccagctg aagggtataa caatcctcgt gttgaattgt tgacaaatca
5341 aatcaaaaat aagtacgaca aagtttatat cgtaccttta gataagctga caaaacaaga
5401 attattagaa ctatgcgaat cattacaaaa aaaagcgtct agttcaatgg ttaaaagtga
5461 aatcatcgac ttattgaatg gtgaagacaa tgacgattga tgatttgctt gtcaaattta
5521 aatcacttga aaagattgac cataattcag aggatgagta cttaaagcag ttgttaaaaa
5581 tgtcgtacga gcgtataaaa aatcagtgcg gagtttttga attagagaat ttaataggtc
5641 aagaattgat acttatacgc gctagatatg cttatcaaga tttattagaa cacttcaacg
5701 acaattacag acctgaaata atagattttt cgttatctct aatggaggta tcagaagatg
5761 aagaaagtgt ttaagaaacc tagaattaca actaaacgtt taaatacgcg tgttcatttt
5821 tataagtata ctgaaaataa tggtccagaa gctggagaaa aagaagaaaa attattatat
5881 agctgttggg cgagtattga tggtgtctgg ttacgtgaat tagaacaagc tatctcaaac
5941 ggaacgcaaa atgacattaa attgtatatt cgtgatccgc aaggtgatta tttacccagt
6001 gaagaacatt atcttgaaat tgaatcaaga tatttcaaaa atcgtttgaa tataaagcaa
6061 gtatcaccag atttggataa taaagacttt attatgattc gcggaggata tagttcatga
6121 gtgtgaaagt gacaggtgat aaagcattag aaagagaatt agaaaaacat tttggcataa
6181 aagagatggt aaaagttcaa gataaggcgt taatagctgg tgctaaggta attgttgaag
6241 aaataaaaaa acaactcaaa ccttcagaag actcaggagc actgattagt gagattggtc
6301 gtactgaacc tgaatggata aaggggaaac gtactgttac aattaggtgg cgtgggcctt
6361 ttgaacgatt tagaatagta catttaattg aaaatggtca tgttgagaaa aagtcaggaa
6421 aatttgtaaa acctaaagct atgggtggga ttaatagagc aataagacaa gggcaaaata
6481 agtattttga gacgctaaaa agggagttga aaaaattgtg attgatattt tgtacaaagt
6541 tcatgaagtg attagtcaag acagaattat tagagagcac gtaaatatca ataatattaa
6601 gttcaataaa taccctaatg taaaagatac tgatgtacct tttattgtta ttgacgatat
6661 cgacgaccca atacctacaa cttatactga cggagatgag tgtgcatata gttatattgt
6721 ccaaatagat gtttttgtta agtacaatga tgaatataat gcgagaatca taagaaataa
6781 gatatctaat cgcattcaaa agttattatg gtctgaacta aaaatgggaa atgtttcaaa
6841 tggaaaaccg gaatatatag aagaatttaa aacatataga agctctcgcg tttacgaggg
6901 cattttttat aaggaggaaa attaaatggc agtaaaacat gcaagtgcgc caaaggcgta
6961 tattaacatt actggtttag gtttcgctaa attaacgaaa gaaggcgcgg aattaaaata
7021 tagtgatatt acaaaaacaa gaggattaca aaaaattggt gttgaaactg gtggagaact
7081 aaaaacagct tatgctgatg gcggtccaat tgaatcaggg aatacagacg gagaaggtaa
7141 aatctcatta caaatgcatg cgttccctaa agagattcgc aaaattgttt ttaatgaaga
7201 ttatgatgaa gatggcgttt acgaagagaa acaaggtaaa caaaacaatt acgtagctgt
7261 atggttcaga caagagcgta aagacggtac atttagaaca gttttattac ctaaagttat-
7321 gtttacaaat cctaaaatcg atggagaaac ggctgagaaa gattgggatt tctcaagtga
7381 agaggttgaa ggtgaggcac ttttcccttt agttgataat aaaaagtcag tacgtaagta
7441 tatctttgat tcagctaaca tgacaaatca tgatggagac ggtgaaaaag gcgaagaggc
7501 tttcttaaag aaaattttag gcgaagaata tactggaaac gtgacagagg gtaacgaaga
7561 aactttgtaa caaaaccggc ttcatcggaa actgcggtaa agtcggttaa tataccagat
7621 agcattaaaa cacttaaagt tggcgacaca tacgatttaa atgttgtagt agagccatct
7681 aatcaaagta agttattgaa atacacaaca gatcaaacga atattgtatc aatcaatagt
7741 gatggtcaag ttactgcgga agcacaaggc attgctacgg ttaaagcaac agttggtaat
7801 atgagtgaca ctataacaat aaatgtagaa gcataagagg gggcaacccc tctattttat
7861 ttgaaaataa ggagagtatt ataaaatggc aaaattaaaa cgtaacatta ttcaattagt
7921 agaagatcca aaagcaaatg aaattaaatt acaaacgtac ttaacaccac acttcatttc
7981 atttgaaatt gtatacgaag caatggattt aatcgatgat attgaggacg aaaatagcac
8041 gatgaagcca agagaaatcg ctgacagatt gatggatatg gttgtaaaaa tttacgataa
8101 ccaattcaca gttaaagacc taaaagaacg tatgcatgca cctgatggaa tgaatgcact
8161 tcgtgaacaa gtgattttca ttactcaagg tcaacaaact gaggaaacta gaaattttat
8221 ccagaacatg aaataaagcc tgaagattta acatataaag caatgttgaa aaatatggat
8281 actctcatga tggacttaat tgaaaatggt aaagacgcta acgaagtttt aaaaatgcca
8341 tttcattatg tgctttccat atatcaaaat aaaaataatg acatttctga agaaaaagca
8401 gaggctttaa ttgatgcatt ttaaccttaa ccgtttggtt agggttattt ttttgaactt
8461 ttttagaaag gaggtaaaaa atgggagaaa gaataaaagg tttatctata ggtttggatt
8521 tagatgcagc aaatttaaat agatcatttg cagaaatcaa acgaaacttt aaaactttaa
8581 attctgactt aaaattaaca ggcaacaact tcaaatatac cgaaaaatca actgatagtt
8641 acaaacaaag gattaaagaa cttgatggaa ctatcacagg ttataagaaa aacgttgatg
8701 atttagccaa gcaatatgac aaggtatctc aagaacaggg cgaaaacagt gcagaagctc
8761 aaaagttacg acaagaatat aacaaacaag caaatgagct gaattattta gaaagagaat
8821 tacaaaaaac atcagccgaa tttgaagagt tcaaaaaagc tcaagttgaa gctcaaagaa
8881 tggcagaaag tggctgggga aaaaccagta aagtttttga aagtatggga cctaaattaa
8941 caaaaatggg tgatggttta aaatccattg gtaaaggttt gatgattggt gtaactgcac
9001 ctgttttagg tattgcagca gcatcaggaa aagcttttgc agaagttgat aaaggtttag
9061 atactgttac tcaagcaaca ggcgcaacag gcagtgaatt aaaaaaattg cagaactcat
9121 ttaaagatgt ttatggcaat tttccagcag atgctgaaac tgttggtgga gttttaggag
9181 aagttaatac aaggttaggt tttacaggta aagaacttga aaatgccaca gagtcattct
9241 tgaaattcag tcatataaca ggttctgacg gtgtgcaagc cgtacagtta attacccgtg
9301 caatgggcga tgcaggtatc gaagcaagtg aatatcaaag tgttttggat atggtagcaa
9361 aagcggcgca agctagtggg ataagtgttg atacattagc tgatagtatt actaaatacg
9421 gcgctccaat gagagctatg ggctttgaga tgaaagaatc aattgcttta ttctctcaat
9481 gggaaaagtc aggcgttaat actgaaatag cattcagtgg tttgaaaaaa gctatatcaa
9541 attggggtaa agctggtaaa aacccaagag aagaatttaa gaagacatta gcagaaattg
9601 aaaagacgcc ggatatagct agcgcaacaa gtttagcgat tgaagcattt ggtgcaaagg
9661 caggtcctga tttagcagac gctattaaag gtggtcgctt tagttatcaa gaatttttaa
9721 aaactattga agattcccaa ggcacagtaa accaaacatt taaagattct gaaagtggct
9781 ccgaaagatt taaagtagca atgaataaat taaaattagt aggtgctgat gtatgggctt
9841 ctattgaaag tgcgtttgct cccgtaatgg aagaattaat caaaaagcta tctatagcgg
9901 ttgattggtt ttccaattta agtgatggtt ctaaaagatc aattgttatt ttcagtggta
9961 ttgctgctgc aattggtcct gtagtttttg ggttaggtgc atttataagt acaattggca
10021 atgcagtaac tgtattagct ccattgttag ctagtattgc aaaggctggt ggattgatta
10081 gttttttatc gactaaagta cctatattag gaactgtctt cacagcttta actggtccaa
10141 ttggcattgt attaggtgta ttggctggtt tagcagtcgc atttacaatt gcttataaga
10201 aatctgaaac atttagaaat tttgttaatg gtgcaattga aagtgttaaa caaacattta
10261 gtaattttat tcaatttatt caacctttcg ttgattctgt taaaaacatc tttaaacaag
10321 cgatatcagc aatagttgat ttcgcaaaag atatttggag tcaaatcaat ggattcttta
10381 atgaaaacgg aatttccatt gttcaagcac ttcaaaatat atgcaacttt attaaagcga
10441 tatttgaatt tattttaaat tttgtaatta aaccaattat gttcgcgatt tggcaagtga
10501 tgcaatttat ttggccggcg gttaaagcct tgattgtcag tacttgggag aacataaaag
10561 gtgtaataca aggtgcttta aatatcatac ttggcttgat taagttcttc tcaagtttat
10621 tcgttggtga ttggcgagga gtttgggacg ccgttgtgat gattcttaaa ggagcagttc
10681 aattaatttg gaatttagtt caattatggt ttgtaggtaa aatacttggt gttgttaggt
10741 actttggcgg gttgctaaaa ggattgatag caggaatttg ggacgtaata agaagtatat
10801 tcagtaaatc tttatcagca atttggaatg caacaaaaag tatttttgga tttttattta
10861 atagcgtaaa atcaattttc acaaatatga aaaattggtt atctaatact tggagcagta
10921 tccgtacgaa tacaatagga aaagcgcagt cattatttag tggcgtcaaa tcaaaattta
10981 ctaatttatg gaatgcgacg aaagaaattt ttagtaattt aagaaattgg atgtcaaata
11041 tttggaattc cattaaagat aatacggtag gaattgcaag ccgtttatgg agtaaggtac
11101 gtggaatttt cacaaatatg cgcgatggct tgagttccat tatagataag attaaaagtc
11161 atatcggcgg tatggtaagc gctattaaaa aaggacttaa taaattaatc gacggtttaa
11221 actgggtcgg tggtaagttg ggaatggata aaatacctaa gttacacact ggtacagagc
11281 acacacatac tactacaaga ttagttaaga acggtaagat tgcacgtgac acattcgcta
11341 cagttgggga taagggacgc ggaaatggtc caaatggttt tagaaatgaa atgattgaat _
11401 tccctaacgg taaacgtgta atcacaccta atacagatac taccgcttat ttacctaaag
11461 gctcaaaagt atacaacggt gcacaaactt attcaatgtt aaacggaacg cttccaagat
11521 ttagtttagg tactatgtgg aaagatatta aatctggtgc atcatcggca tttaactgga
11581 caaaagataa aataggtaaa ggtaccaaat ggcttggcga taaagttggc gatgttttag
11641 attttatgga aaatccaggc aaacttttaa attatatact tgaagctttt ggaattgatt
11701 tcaattcttt aactaaaggt atgggaattg caggcgacat aacaaaagct gcatggtcta
11761 agattaagaa aagtgctact gattggataa aagaaaattt agaagctatg ggcggtggcg 11821 atttagtcgg cggaatatta gaccctgaca aaattaatta tcattatgga cgtaccgcag 11881 cttataccgc tgcaactgga agaccatttc atgaaggtgt cgattttcca tttgtatatc 11941 aagaagttag aacgccgatg ggtggcagac ttacaagaat gccatttatg tctggtggtt 12001 atggtaatta tgtaaaaatt actagtggcg ttatcgatat gctatttgcg catttgaaaa 12061 actttagcaa atcaccacct agtggcacga tggtaaagcc cggtgatgtt gttggtttaa 12121 ctggtaatac cggatttagt acaggaccac atttacattt tgaaatgagg agaaatggac 12181 gacattttga ccctgaacca tatttaagga atgctaagaa aaaaggaaga ttatcaatag 12241 gtggtggcgg tgctacttct ggaagtggcg caacttatgc cagtcgagta atccgacaag 12301 cgcaaagtat tttaggtggt cgttataaag gtaaatggat tcatgaccaa atgatgcgcg 12361 ttgcaaaacg tgaaagtaac taccagtcaa atgcagtgaa taactgggat ataaatgctc 12421 aaagaggaga cccatcaaga ggattattcc aaatcatcgg ctcaactttt agagcaaacg 12481 ctaaacgtgg atatactaac tttaataatc cagtacatca aggtatctca gcaatgcagt 12541 acattgttag acgatatggt tggggtggtt ttaaacgtgc tggtgattac gcatatgcta 12601 caggtggaaa agtttttgat ggttggtata acttaggtga agacggtcat ccagaatgga 12661 ttattccaac agatccagct cgtagaaatg atgcaatgaa gattttgcat tatgcagcag 12721 cagaagtaag agggaaaaaa gcgagtaaaa ataagcgtcc tagccaatta tcagacttaa 12781 acgggtttga tgatcctagc ttattattga aaatgattga acaacagcaa caacaaatag 12841 ctttattact gaaaatagca caatctaacg atgtgattgc agataaagat tatcagccga 12901 ttattgacga atacgctttt gataaaaagg tgaacgcgtc tatagaaaag cgagaaaggc 12961 aagaatcaac aaaagtaaag tttagaaaag gaggaattgc tattcaatga tagacactat 13021 taaagtgaac aacaaaacaa ttccttggtt gtatgtcgaa agagggtttg aaataccctc 13081 ttttaattat gttttaaaaa cagaaaatgt agatggacgt tcggggtcta tatataaagg 13141 gcgtaggctt gaatcttata gttttgatat acctttggtg gtacgtaatg actatttatc 13201 tcacaacggc attaaaacac atgatgacgt cttgaatgaa ttagtaaagt tttttaacta 13261 cgaggaacaa gttaaattac aattcaaatc taaagattgg tactggaacg cttatttcga 13321 aggaccaata aagctgcaca aagaatttac aatacctgtt aagttcacta tcaaagtagt 13381 actaacagac ccttacaaat attcagtaac aggaaataaa aatactgcga tttcagacca 13441 agtttcagtt gtaaatagtg ggactgctga cactccttta attgttgaag cccgagcaat 13501 taaaccatct agttacttta tgattactaa aaatgatgaa gattatttta tggttggtga 13561 tgatgaggta accaaagaag ttaaggatta catgcctcct gtttatcata gtgagtttcg 13621 tgatttcaaa ggttggacta agatgattac tgaagatatt ccaagtaatg acttaggtgg 13681 taaggtcggc ggtgactttg tgatatccaa tcttggcgaa ggatataaag caactaattt 13741 tcctgatgca aaaggttggg ttggtgctgg cacgaaacga gggctcccta aagcgatgac 13801 agattttcaa attacctata aatgtattgt tgaacaaaaa ggtaaaggtg ccggaagaac 13861 agcacaacat atttatgata gtgatggtaa gttacttgct tctattggtt atgaaaataa 13921 atatcatgat agaaaaatag gacatattgt tgttacgttg tataaccaaa aaggagaccc 13981 caaaaagata tacgactatc agaataaacc gataatgtat aacttggaca gaatcgttgt 14041 ttatatgcgg ctcagaagag taggtaataa attttctatt aaaacttgga aatttgatca 14101 cattaaagac ccagatagac gtaaacctat tgatatggat gagaaagagt ggatagatgg 14161 cggtaagttt tatcagcgtc cagcttctat catagctgtc tatagtgcga agtataacgg 14221 ttataagtgg atggagatga atgggttagg ttcattcaat acggagattc taccgaaacc 14281 gaaaggcgca agggatgtca ttatacaaaa aggtgattta gtaaaaatag atatgcaagc 14341 aaaaagtgtt gtcatcaatg aggaaccaat gttgagcgag aaatcgtttg gaagtaatta 14401 tttcaatgtt gattctgggt acagtgaatt aatcatacaa cctgaaaacg tctttgatac 14461 gacggttaaa tggcaagata gatatttata gaaaggagat gagagtgtga tacatgtttt 14521 agattttaac gacaagatta tagatttcct ttctactgat gacccttcct tagttagagc 14581 gattcataaa cgtaatgtta atgacaattc agaaatgctt gaactgctca tatcatcaga 14641 aagagctgaa aagttccgtg aacgacatcg tgttattata agggattcaa acaaacaatg 14701 gcgtgaattt attattaact gggttcaaga tacgatggac ggctacacag agatagaatg 14761 tatagcgtct tatcttgctg atataacaac agctaaaccg tatgcaccag gcaaatttga 14821 gaaaaagaca acttcagaag cattgaaaga tgtgttgagc gatacaggtt gggaagtttc 14881 tgaacaaacc gaatacgatg gcttacgtac tacgtcatgg acttcttatc aaactagata 14941 tgaagtttta aagcaattat gtacaaccta taaaatggtt ttagattttt atattgagct 15001 tagctctaat accgtcaaag gtagatatgt agtactcaaa aagaaaaaca gcttattcaa 15061 aggtaaagaa attgaatatg gtaaagattt agtcgggtta actaggaaga ttgatatgtc 15121 agaaatcaaa acagcattaa ttgctgtggg acctgaaaat gacaaaggga agcgtttaga 15181 gctagttgtg acagatgacg aagcgcaaag tcaattcaac ctacctatgc gctatatttg 15241 ggggatatat gaaccacaat cagatgatca aaatatgaat gaaacacgat taagttcttt 15301 agccaaaaca gagttaaata aacgtaagtc ggcagttatg tcatatgaga ttacttctac 15361 tgatttggaa gttacgtatc cgcacgagat tatatcaatt ggcgatacag tcagagtaaa 15421 acatagagat tttaacccgc cattgtatgt agaggcagaa gttattgctg aagaatataa _ 15481 cataatttca gaaaatagca catatacatt cggtcaacct aaagagttca aagaatcaga 15541 attacgagaa gagtttaaca agcgattgaa cataatacat caaaagttaa acgataatat 15601 tagcaatatc aacactatag ttaaagatgt tgtagatggt gaattagaat actttgaacg 15661 caaaatacac aaaagtgata caccgccaga aaatccagtc aatgatatgc tttggtatga 15721 tacaagtaac cctgatgttg ctgtcttgcg tagatattgg aatggtcgat ggattgaagc 15781 aacaccaaat gatgttgaaa aattaggtgg tataacaaga gagaaagcgc tattcagtga
15841 attaaacaat atttttatta atttatctat acaacacgct agtcttttgt cagaagctac 15901 agaattactg aatagcgagt acttagtaga taatgatttg aaagcggact tacaagcaag 15961 tttagacgct gtgattgatg tttataatca aattaaaaat aatttagaat ctatgacacc 16021 cgaaactgca acgattggtc ggttggtaga tacacaagct ttatttcttg agtatagaaa 16081 gaaattacaa gatgtttata cagatgtaga agatgtcaaa atcgccattt cagatagatt 16141 taaattatta cagtcacaat acactgatga aaaatataaa gaagcgttgg aaataatagc 16201 aacaaaattt ggtttaacgg tgaatgaaga tttgcagtta gtcggagaac ctaatgttgt 16261 taaatcagct attgaagcag ctagagaatc cacaaaagaa caattacgtg actatgtaaa 16321 aacatcggac tataaaacag acaaagacgg tattgttgaa cgtttagata ctgctgaagc 16381 tgagagaacg actttaaaag gtgaaatcaa agataaagtt acgttaaacg aatatcgaaa 16441 cggattggaa gaacaaaaac aatatactga tgaccagtta agtgatttgt ccaataatcc 16501 tgagattaaa gcaagtattg aacaagcaaa tcaagaagcg caagaagctt taaaatcata 16561 cattgatgct caagatgatc ttaaagagaa ggaatcgcaa gcgtatgctg atggtaaaat 16621 ttcggaagaa gagcaacgcg ctatacaaga tgctcaagct aaacttgaag aggcaaaaca 16681 aaacgcagaa ctaaaggcta gaaacgctga aaagaaagct aatgcttata cagacaacaa 16741 ggtcaaagaa agcacagatg cacagaggaa aacattgact cgctatggtt ctcaaattat 16801 acaaaatggt aaggaaatca aattaagaac tactaaagaa gagtttaatg caaccaatcg 16861 tacactttca aatatattaa acgagattgt tcaaaatgtt acagatggaa caacaatcag 16921 atatgatgat aacggagtgg ctcaagcttt gaatgtgggg ccacgtggta ttagattaaa 16981 tgctgataaa attgatatta acggtaatag agaaataaac cttcttatcc aaaatatgcg 17041 agataaagta gataaaaccg atattgtcaa cagtcttaat ttatcaagag agggtcttga 17101 tatcaatgtt aatagaattg gaattaaagg cggtgacaat aacagatatg ttcaaataca 17161 gaatgattct attgaactag gtggtattgt gcaacgtact tggagaggga aacgttcaac 17221 agacgatatt tttacgcgac tgaaagacgg tcacctaaga tttagaaata acaccgctgg 17281 cggttcactt tatatgtcac attttggtat ttcgacttat attgatggtg aaggtgaaga 17341 cggtggttca tctggtacga ttcaatggtg ggataaaact tacagtgata gtggcatgaa 17401 tggtataaca atcaattcct atggtggtgt cgttgcacta acgtcagata ataatcgggt 17461 tgttctggag tcttacgctt catcgaatat caaaagcaaa caggcaccgg tgtatttata 17521 tccaaacaca gacaaagtgc ctggattaaa ccgatttgca ttcacgctgt ctaatgcaga 17581 taatgcttat tcgagtgacg gttatattat gtttggttct gatgagaact atgattacgg 17641 tgcgggtatc aggttttcta aagaaagaaa taaaggtctt gttcaaattg ttaatggacg 17701 atatgcaaca ggtggagata caacaatcga agcagggtat ggcaaattta atatgctgaa 17761 acgacgtgat ggtaataggt atattcatat acagagtaca gacctactgt ctgtaggttc 17821 agatgatgca ggagatagga tagcttctaa ctcaatttat agacgtactt attcggccgc 17881 agctaatttg catattactt ctgctggcac aattgggcgt tcgacatcag cgcgtaaata 17941 caagttatct atcgaaaatc aatataacga tagagatgaa caactggaac attcaaaagc 18001 tattcttaac ttacctatta gaacgtggtt tgataaagct gagtctgaaa ttttagctag 18061 agagctgaga gaagatagaa aattatcgga agacacctat aaacttgata gatacgtagg 18121 tttgattgct gaagaggtgg agaatttagg attaaaagag tttgtcacgt atgatgacaa 18181 aggagaaatt gaaggtatag cgtatgatcg tctatggatt catcttatcc ctgttatcaa 18241 agaacaacaa ctaagaatca agaaattgga ggagtcaaag aatgcaggat aacaaacaag 18301 gattacaagc taatcctgaa tatacaattc attatttatc acaggaaatt atgaggttaa 18361 cacaagaaaa cgcgatgtta aaagcgtata tacaagaaaa taaagaaaat caacaatgtg 18421 ctgaggaaga gtaatcctta gcactatttt tatacaaaaa tttaaggagg tcatttaatt 18481 atggcaaaag aaattatcaa caatacagaa aggtttattt tagtacaaat cgacaaagaa 18541 ggtacagaac gtgtagtata tcaagatttc acaggaagtt ttacaacttc tgaaatggtt 18601 aaccatgctc aagattttaa atctgaagaa aacgctaaga aaattgcgga gacgttaaat 18661 ttgttatatc aattaactaa caaaaaacaa cgtgtgaaag tagttaaaga agtagttgaa 18721 agatcagatt tatctccaga ggtaacagtt aacactgaaa cagtatgaaa agctatgagt 18781 tagatactca tagtctttat tcttttagaa agcgggtgta ctgaattggg gtggttcaaa 18841 aaacacgaac atgaatggcg catcagaagg ttagaagaga atgataaaac aatgctcagc 18901 acactcaacg aaattaaatt aggtcaaaaa acccaagagc aagttaacat taaattagat 18961 aaaaccttag atgctattca aaaagaaaga gaaatagatg aaaagaataa gaaagaaaat 19021 gataagaaca tacgtgatat gaaaatgtgg gtgcttggtt tagttgggac aatatttggg 19081 tcgctaatta tagcattatt gcgtatgctt atgggcatat aagagaggtg attaccatgt 19141 tcggattaaa ttttggagct tcgctgtgga cgtgtttctg gtttggtaag tgtaagtaat 19201 agttaagagt cagtgcttcg gcactggctt tttattttgg ataaaaggag caaacaaatg 19261 gatgcaaaag taataacaag atacatcgta ttgatcttag cattagtaaa tcaattctta 19321 gcgaacaaag gtattagccc aattccagta gacgatgaaa ctatatcatc aataatactt 19381 actgtagtcg ctttatatac aacgtataaa gacaatccaa catctcaaga aggtaaatgg 19441 gcaaatcaaa aattaaagaa atataaagct gaaaataagt atagaaaagc aacagggcaa 19501 gcgccaatta aagaagtaat gacacctacg aatatgaacg acacaaatga tttagggtag 19561 gtggttgata tatgttaatg acaaaaaatc aagcagaaaa atggtttgac aattcattag 19621 ggaaacaatt caacccagat ggttggtatg gatttcagtg ttatgattac gccaatatgt 19681 tctttatgtt agcgacaggc gaaaggctgc aaggtttata tgcttataat atcccgtttg 19741 ataataaagc aaagattgaa aaatatggtc aaataattaa aaactatgac agctttttac 19801 cgcaaaagtt ggatattgtc gttttcccgt caaagtatgg tggcggagct ggacacgttg 19861 aaattgttga gagcgcaaat ttaaatactt tcacatcatt tggtcaaaac tggaacggta
19921 aaggttggac taatggcgtt gcgcaacctg gttggggtcc tgaaactgtg acaagacatg
19981 ttcattatta tgacaatcca atgtatttta ttaggttaaa cttccctaac aacttaagcg
20041 ttggcaataa agctaaaggt attattaagc aagcgactac aaaaaaagag gcagtaatta
20101 aacctaaaaa aattatgctt gtagccggtc atggttataa cgatcctgga gcagtaggaa
20161 acggaacaaa cgaacgcgat tttatacgta aatatataac gcctaatatc gctaagtatt
20221 taagacatgc aggacatgaa gttgcattat acggtggctc aagtcaatca caagatatgt
20281 atcaagatac tgcatacggt gttaatgtag gcaataaaaa agattatggc ttatattggg
20341 ttaaatcaca ggggtatgac attgttctag aaatacattt agacgcagca ggagaaagcg
20401 caagtggtgg gcatgttatt atctcaagtc aattcaatgc agatactatt gataaaagta
20461 tacaagatgt tattaaaaat aacttaggac aaataagagg tgtgacacct cgtaatgatt
20521 tactaaatgt taatgtatca gcagaaataa atataaatta tcgtttatct gaattaggtt
20581 ttattactaa taaaaatgat atggattgga ttaagaaaaa ctatgacttg tattctaaat
20641 taatagccgg tgcgattcat ggtaagccta taggtggttt ggtagctggt aatgttaaaa
20701 catcagctaa aaacaaaaaa aatccaccag tgccagcagg ttatacactc gataagaata
20761 atgtccctta taaaaaagaa caaggcaatt acacagtagc taatgttaaa ggtaataatg
20821 taagagacgg ttattcaact aattcaagaa ttacaggggt attacccaac aacacaacaa
20881 ttacgtatga cggtgcatat tgtattaatg gttatagatg gattacttat attgctaata
20941 gtggacaacg tcgttatata gcgacaggag aggtagacaa ggcaggtaat agaataagta
21001 gttttggtaa gtttagcacg atttagtatt tacttagaat aaaaattttg ctacattaat
21061 tatagggaat cttacagtta ttaaataact atttggatgg atgttaatat tcctatacac
21121 tttttaacat ttctctcaag atttaaatgt agataacagg caggtacttc ggtacttgcc
21181 tattttttta tgttatagct agccttcggg ctagtttttt gttatgatgt gttacacatg
21241 catcaactat ttacatctat ccttgttcac ccaagcatgt cactggatgt tttttcttgc
21301 gatagagagc atagttttca tactactccc cgtagtatat atgactttag cattcccgta
21361 taacagttta cggggtgctt ttatgttata attgctttta tatagtagga gtgaactata
21421 tagccgggca gaggccatgt atctgactgt tggtcccaca ggagacatct tccttgtcat
21481 cactcgatac atatatctta acaacataga aatgttacat tcgctataac cgtatcttaa
21541 tcgatacggt tatatttatt cccctacaac caacaaaacc acagatccta ttaatttagg
21601 attgtggtta ttttttgcgt ttttttgggg caaaaaaagg gcagattatt tgaaaaaggg
21661 caaacgcttg tggaaaagct aaaaggttaa aaatgacaaa aaccttgata caacagtgtt
21721 tttggacgct cgtgtacgtt agagaatgac cggtttacca tcatacaagg gtgggattaa
21781 cttgtgttaa aaagccttta atatcagttg ttacaaagga tttgtagcgt ctttaaaaat
21841 aaaaaagggc agaaaaaggg cagatacctt ttagtacaca agtttttcta atttttgctc
21901 taactctctg tccattttct ctgttacatg tgtatacacc tttatagtcg ttttttcatc
21961 tgtatgtcct actcttttca taattgcttt taacgatata ttcatttccg ccaataaact
22021 tatgtgtgta tgccttagtg tgtgagtagt aactttttta tttatattta atgattctgc
22081 agctgaggac aatcgtttgt ttatcctact gccttgcata ggatttcctt ggcaagttgt
22141 gaatataaac cctctatcaa catagcttgg ttcccattgt tgcatctttt tattttctaa
22201 cattattttt ttcaatacat ttgctatcct tgaattgatg gcgatttttc ttcttgaacc
22261 tgcggtctta gtagtatctt tgtgaccaaa tccagcatta catttgattc tgtgaatagt
22321 gccattaata gcgatcgttt tatttttgag gtcaacatct ttaacttgga gagctaataa
22381 ctcacctatg cgcatacctg ttaaagcttg aacttctaca gccccagcaa ctaaaatacg
22441 agctctatac tgcatgttat tatcgttcag tataaaatcg cgtatctgta ttacctgttc
22501 catctctaaa tagttataca ttttcgcttc ttctttttct atatcttcta tcgtcttact
22561 cttctttggt agtgtgacgc tatttaatat gtgttcgttt ggataattgt aaaatttaac
22621 ggcgtattta atagcttctt tcatatgtcc aagttgacgc tttacctgat ttgcagaata
22681 tacgtttgat aatttgttaa taaatgtttg catgtacttt gtatcaattt tgtttaaaag
22741 taaattttga gaactgttct ttttgatgtt tttgattctt gttttcaaat tatcaagcgt
22801 cgttacttta aagccagatg tttttatatg atattcaagc cattcatcta ataacgcgtg
22861 aaaagtcaaa gtttttaatt cgcttgacga cttgttgttt agtttttctt ttattttttc
22921 ttctaaacga aacattgcct ctttttgcga ttgctttgta ttcttattca agacaacact
22981 tacacgtttc catttatctg tatacggatc tttgtatttc tcgtagtatc tatacttcgt
23041 ttcattgttc ttatttttaa atttttcaaa ccacatttta catccctcct caaaattggc
23101 aaaaaataat aagggtaggc gggctaccca tgaaaattgt ataaaaaaag acgcctgtat
23161 aaaatacaga cgccacttat aattataaga ttacatggtt aattaccaaa aatggtaacg
23221 aatatatacg tgttttaaag gataaacctt taatatatta aaattatatc atcttatatc
23281 agggatctgc aatatattat tattaattct atttatcagt aacataatat ccgaagaatc
23341 tattactgga tttttaattt tttggggtaa aacttttctt atgcgaaact tactaatcgg
23401 ctggaaagaa tttatgcaag cgtaactatt accttttaat ttttttacct tatcaattgc
23461 tgatactatg ttattaatgt ttctgtcaat tttatttaat ttattttcaa tttctaaact
23521 atcagatata aattcaataa aataatcttt agtgatgaat tctgtgttgt ttttttggta
23581 ttttttatcg aaaacttctt ttaatatagc tgaattattt tgcgcgctaa ttaaatttaa _
23641 aaacaatctt aaataatact cccatttcaa atcaaaattc atctttaaat actttttgtt
23701 ttctttagag gataagggaa taacatttac tatatcctcc gtattagaat catttttatt
23761 catcactatt gcaaagtgtg aattagaaaa ttctttatta acgtttatac cgaaatctac
23821 aaaaactatt tctccttgtt taaactttgg ataaaaacct ttatggtttt tttcaccttc
23881 aaatctcttg agtaaatagt gaatatctga atctaacttt ttaaattttg gatttccaga
23941 agtttttaat ttattaatgc gtttttctat attatgcgtc atcatttctc ctttattctc
24001 gctcacactc tcaccaccat tcaacgtcta cacttgtagg cgttttttga ttagtaaaat
24061 cataatgaat cttctttggt taacttatcg ccatctattt tttgtgaaat aaattccaag
24121 tatttacgcg cattatgtga cgataaatct ttaggtaact cataagtgaa tggttgatta
24181 ccactagtta aaacttcata tactatagtt tcttttttta ttttgcaatt agttattttc
24241 attataaact ccttttaaac actgctgaaa tagacgtctt tttcaaataa gcatgattaa
24301 tactttaatt ctttaatcca catatattta aaagtgaggt agtaggtaat aaatataaga
24361 cttaaagtta agattgcttt tttcatgtca atttctcctt tgtttatatt tatattaaag
24421 cgctaaatat acgttattaa tcacaataca actttgccca ttactttaat atcactaaac
24481 gaagcgactt tgatatcatc atacttcgga tttagagata ccaaattaat atagtcttcg
24541 catatatcta cacgcttgat aagacttact ccatctaata caacgagtgc aattgtacca
24601 tctttaatag aatcttcttt cttaataaaa gcgtatgttc cttgttttaa cataggttcc
24661 attgaatcac cattaactaa aatacaaaaa tcagcatttg atggcgtttc gtcttcttta
24721 aaaaatactt cttcatgcaa tatgtcatca tataattctt ctcctatgcc agcaccagtt
24781 gcaccacatg caatatacga tactagttta gactctttat attcatctat agaagtgact
24841 ttattctgtt catctaattg ctcatttgca tagttaagta cgttttcttg gcggggaggt
24901 gtgagttgag aaaatatgtt attgattttt gacattatcg tttcatcttg acgttcttcg
24961 tcaggaactc gataagaatc tacatcatac cccataagcc acgcttcacc gacatttaaa
25021 gttttagata ataagaataa tttatgttgg tctggagaag accttccatt aacatactgg
25081 gataagtgac tttttgacat tttaatattc aattcttttt gaaagggttt cgacttttct
25141 agaatatcta cttgacgcaa gttcctatct ttcataattt gttttaatct ttcagaagtg
25201 ttttgcattg gtaatgcctc cttgaaattc attatatagg aagggaaata aaaatcaata
25261 caaaagttca acttttttaa ctttttgtgt tgacattgtt caaaattggg gttatagtta
25321 ttatagttca aatgtttgaa cttaggaggt gattatttga atactaatac aacttttgat
25381 ttttcgttat tgaacggtaa gatagtcgaa gtgtactcga cacaatttaa ctttgctata
25441 gctttaggtg tatcagaaag aactttgtct ttgaagttga acaacaaagt accatggaaa
25501 acaacagaca ttattaaagc ttgtaagtta ttgggaatac ctataaaaga tgttcacaaa
25561 tattttttta aacagaaagt tcaaatgttt gaacttaata agtaaaggag gcataacaca
25621 tgcaagaacg agaaaaggtt aataaaagta acacatcttc aaatgaagca tcaaaacctt
25681 ttaggacaaa ttgaagctta cgacaaaacg cttaaagaaa taaagtacac tcgagacctt
25741 tacaacaaac acctaagcat gaacaacgaa gacgcattcg ctggtttgga aatggtagag
25801 gatgaaatta ctaaaaagct acgaagtgct atcaaagagt tccaaaaagt agtgaaagcg
25861 ttagacaagc ttaacggtgt tgaaagcgat aacaaagtta ctgatttaac agagtggcgg
25921 aaagtgaatc agtaacattc acttcttaat ataaccacgc ttatcaacat ccacattgag
25981 cagatgtgag cgagagctgg cgatgatatg agccgcgttt aaatacattc gatagtcatt
26041 gcgataaccg tctgctgaat gtgggtgttg aggaaaaagg aggatactca aatgcaagca
26101 ttacaaacat ttaattttaa agagctacca gtaagaacag tagaaattga aaacgaacct
26161 tattttgtag gaaaagatat tgctgagatt ttaggatatg caagatcaaa caatgccatt
26221 agaaatcatg ttgatagcga ggacaagctg acgcaccaat ttagtgcatc aggtcaaaac
26281 agaaatatga tcattatcaa cgaatcagga ttatacagtc taatcttcga tgcttctaaa
26341 caaagcaaaa acgaaaaaat tagagaaacc gctagaaaat tcaaacgctg ggtaacatca
26401 gatgtcctac cagctattcg caaacacggt atatacgcaa cagacaatgt aattgaacaa
26461 acattaaaag atccagacta catcattaca gtgttgactg agtataagaa agaaaaagag
26521 caaaacttac ttttacaaca gcaagtagaa gttaacaaac caaaagtatt attcgctgac
26581 tcggtagctg gtagtgataa ttcaatactt gttggagaac tagcgaaaat acttaaacaa
26641 aacggtgttg atataggaca aaacagattg ttcaaatggt taagaaataa tggatatctc
26701 attaaaaaga gtggagaaag ttataactta ccaactcaaa agagtatgga tctaaaaatc
26761 ttggatatca aaaaacgaat aattaataat ccagatggtt caagtaaagt atcacgtaca
26821 ccaaaagtaa caggcaaagg acaacaatac tttgttaata agtttttagg agaaaaacaa
26881 acatcttaaa aggaggaaca caatggaaca aatcacatta accaaagaag agttgaaaga
26941 aattatagca aaagaagtta gagaggctat aaatggcaag aaaccaatca gttcaggttc
27001 aattttcagt aaagtaagaa tcaataatga cgatttagaa gaaatcaata aaaaactcaa
27061 tttcgcaaaa gatttgtcgc taggaagatt gaggaagctc aatcatccga ttccgctaaa
27121 aaagtatcag catggcttcg aatcaattca tcaaaaagct tatgtacaag atgttcatga
27181 ccatattaga aaattaacat tatcaatttt tggagtgaca cttaattcag acttgagtga
27241 aagtgaatac aacctagcag caaaagttta tcgagaaatc aaaaactatt atttatacat
27301 ctatgaaaag agagtttcag aattaactat cgatgatttc gaataaagga ggaacaacaa
27361 atgttacaaa aatttagaat tgcgaaagaa aaaaataaat taaaactcaa attactcaag
27421 catgctagtt actgtttaga aagaaacaac aaccctgaac tgttgcgagc agttgcagag
27481 ttgttgaaaa aggttagcta aattcaacgg taaggatttg ccctgcctcc acacttagag
27541 tttgagatcc aacaaacaca taagttttag tagggtctag aaaaaatgtt tcgatttcct
27601 cttttgtaac agtttcaatt ccttcatatc ctggaaaaac aattttcttt aaatccgaaa
27661 catgtttttt tgaaccatcc tttaaagtaa ctagaagttt catacttatc acctccttag_
27721 gttgataaca acattataca cgaaaggagc ataaacaata tgcaagcatt acaaacaaat
27781 tcgaacatcg gagaaatgtt caatattcaa gaaaaagaaa atggagaaat cgcaatcagc
27841 ggtcgagaac ttcatcaagc attagaagtt aagacagcat ataaagattg gtttccaaga
27901 atgcttaaat acggatttga agaaaataca gattacacag ctatcgctca aaaaagagca
27961 acagctcaag gcaatatgac tcactatatt gaccacgcac tcacactaga cactgcaaaa
28021 gaaatcgcaa tgattcaacg tagtgaacct ggcaaacgtg caagacaata tttcatccaa
28081 gttgaaaaag catggaacag cccagaaatg attatgcaac gtgctttaaa aattgctaac 28141 aacacaatca atcaattaga aacaaagatt gcacgtgaca aaccaaaaat tgtatttgca 28201 gatgcagtag ctactactaa gacatcaatt ttagttggag agttagcaaa gatcattaaa 28261 caaaacggta taaacatcgg gcaacgcaga ttgtttgagt ggttacgtca aaacggattc 28321 cttattaaac gcaagggtgt ggattataac atgcctacac agtattcaat ggaacgtgag 28381 ttattcgaaa ttaaagaaac atcaatcaca cattcggacg gtcacacatc aattagtaag 28441 acgccaaaag taacaggtaa aggacaacaa tactttgtta acaagttttt aggagaaaaa 28501 caaacaactt aataggagga attacaaatg aacgcactat acaaaacaac cctcctcatc 28561 acaatggcag ttgtgacgtg gaaggtttgg aagattgaga agcacactag aaaacctgtg 28621 attagtagca gggcgttgag tgactatcta aacaacaaat ctttaaccat accgaaagat 28681 gctgaaaatt ctactgaatc tgctcgtcgc cttttgaagt tcgccgaaca aactattagc 28741 aaataacaac attatacacg aaaggaaaga tagaaatgcc aaaaatcata gtaccaccaa 28801 caccagaaaa cacatataga ggcgaagaaa aatttgtgaa aaagttatac gcaacaccta 28861 cacaaatcca tcaattgttt ggagtatgta gaagtacagt atacaactgg ttgaaatatt 28921 accgcaaaga taatttaggt gtagaaaatt tatacattga ttattcacca acaggcactc 28981 tgattaatat ttctaaattg gaagagtatt tgatcagaaa gcataaaaaa tggtattagg 29041 aggatattaa atgagcaaca tttataaaag ctacctagta gcagtattat gcttcacagt 29101 cttagcgatt gtacttatgc cgtttctata cttcactaca gcatggtcaa ttgcgggatt 29161 cgcaagtatc gcaacattca tgtactacaa agaatgcttt ttcaaagaat aaaaaaactg 29221 ctacttgttg gagcaagtaa cagtatcaaa cacttaagaa aaaattcatg ttcaatataa 29281 aacgaaaaac ggaggaagtc aagatgtatt acgaaatagg cgaaatcata cgcaaaaata 29341 ttcatgttaa cggattcgat tttaagctat tcattttaaa aggtcatatg ggcatatcaa 29401 tacaagttaa agatatgaac aacgtaccaa ttaaacatgc ttatgtcgta gatgagaatg 29461 acttagatat ggcatcagac ttatttaacc aagcaataga tgaatggatt gaagagaaca 29521 cagacgaaca ggacagacta attaacttag tcatgaaatg gtaggaggtc gctatgaagc 29581 agactgtaac ttatatcatt cgtcataggg atatgccaat ttatataact aacaaaccaa 29641 ctgataacaa ttcagatatt agttactcca caaatagaaa tagagctagg gagtttaacg 29701 gtatggaaga agcgagtatc aatatggatt atcacaaagc aatcaagaaa acagtgacag 29761 aaactattga gtacgaggag gtagaacatg actgaggaaa aacaagaacc acaagaaaaa 29821 gtaagcatac tcaaaaaact aaagataaat aatatcgctg agaaaaataa aaggaaattc 29881 tataaatttg cagtatacgg aaaaattggc tcaggaaaaa ccacgtttgc tacaagagat 29941 aaagacgctt tcgtcattga cattaacgaa ggtggaacaa cggttactga cgaaggatca 30001 gacgtagaaa tcgagaacta tcaacacttt gtttatgttg taaatttttt acctcaaatt 30061 ttacaggaga tgagagaaaa cggacaagaa atcaatgttg tagttattga aactattcaa 30121 aaacttagag atatgacatt gaatgatgtg atgaaaaata agtctaaaaa accaacgttt 30181 aatgattggg gagaagttgc tgaacgaatt gtcagtatgt acagattaat aggaaaactt 30241 caagaagaat acaaattcca ctttgttatt acaggtcatg aaggtatcaa caaagataaa 30301 gatgatgaag gtagcactat caaccctact atcactattg aagcgcaaga acaaattaaa 30361 aaagctatta cttctcaaag tgatgtgtta gctagggcaa tgattgaaga atttgatgat 30421 aacggagaaa agaaagctag atatattcta aacgctgaac cttctaatac gtttgaaaca 30481 aagattagac attcaccttc aataacaatt aacaataaga aatttgcaaa tcctagcatt 30541 acggacgtag tagaagcaat tagaaatgga aactaaaaat taattaaaag gacggtattt 30601 aattatgaaa atcacaggac aagcgcaatt tactaaagaa acaaatcaag aaaagtttta 30661 taacggctca gcagggtttc aagctggaga attcacagtg aaagttaaaa atattgaatt 30721 caatgataga gaaaatagat atttcacaat cgtatttgaa aatgatgaag gcaaacaata 30781 taaacataat caatttgtac cgccgtataa atatgatttc caagaaaaac aattgattga 30841 attagttact cgattaggta ttaagttaaa tcttcctagc ttagattttg ataccaatga 30901 tcttattggt aagttttgtc acttggtatt gaaatggaaa ttcaatgaag atgaaggtaa 30961 gtattttacg gatttttcat ttattaaacc ttacaaaaag ggcgatgatg ttgttaacaa 31021 acctattccg aagacagata agcaaaaagc tgaagaaaat aacggggcac aacaacaaac 31081 atcaatgtct caacaaagca atccatttga aagcagtggc caatttggat atgacgacca 31141 agatttagcg ttttaaggtg tggtttaaat gcaatacatt acaagatacc agaaagataa 31201 cgacggtact tattccgtcg ttgctactgg tgttgaactt gaacaaagtc acattgactt 31261 actagaaaac ggatatccac taaaagcaga agtagaggtt ccggacaata aaaaactatc 31321 tatagaacaa cgcaaaaaaa tattcgcaat gtgtagagat atagaacttc actggggcga 31381 accagtagaa tcaactagaa aattattaca aacagaattg gaaattatga aaggttatga 31441 agaaatcagt ctgcgcgact gttctatgaa agttgcaagg gagttaatag aactgattat 31501 agcgtttatg tttcatcatc aaatacctat gagtgtagaa acgagtaagt tgttaagcga 31561 agataaagcg ttattatatt gggctacaat caaccgcaac tgtgtaatat gcggaaagcc 31621 tcacgcagac ctggcacatt atgaagcagt cggcagaggc atgaacagaa acaaaatgaa 31681 ccactatgac aaacatgtat tagcgttatg tcgcgaacat cacaacgagc aacatgcgat 31741 tggcgttaag tcgtttgatg ataaatacca cttgcatgac tcgtggataa aagttgatga 31801 gaggctcaat aaaatgttga aaggagagaa aaaggaatga atagactaag aataataaaa 31861 atagcactcc taatcgtcat cttggcggaa gagattagaa atgctatgca tgctgtaaaa 31921 gtggagaaaa ttttaaaatc tccgtttagt taatacaggt ttttacaaaa gctttaccat 31981 aggcggacaa actaattgag ccttttttga tgtctattac ccaggggctg taatgtaact 32041 ttaatacttc aaattcaatg ccagaaagtt tacttattgt ttctaggttg tgtcctgact 32101 ttaacattct tttaacaaat tctaatcccg aaacaaatct ttgtttttct ataatcttat
32161 taaagtgatt taaaaactga ggagcataaa acttattata aattcctttt tttgttaagt 32221 aagacatgtc aaaagtttca tttaaaaccc ctaaccttac taggttatta attgaaattt 32281 cggttgattc tatatctaac ggagagtctt ttattaacgt gtccgatata ttcataccgt 32341 cattctttgg gtttaaaacc gctctatatt taacggcagg atgtacttcg tgattcttta 32401 aatgttttaa aagaatagca tcatttgggg ataattgttt aattatttca acaaatgaat 32461 ggtgggttaa tgagtttttt ctgtcatcca tagatgatgc tattagtttt gcgaacatat 32521 tacttaaagt tttttcacta atgtaaaact ttgaagcttc tagagcagga cctagaagag 32581 aaaattgtgg ttcttgtaaa ttatttttag gtacagaaga tatttctttt ttaaattgtt 32641 ctttgaattt ttcaaattct acttctcttt gataaataac tttatccaca taaaggtgga 32701 atttcccaaa gacaagttcc caagttttag agaatgtttc tacaggccct tttgatgcgc 32761 cttcaataat tttatcaata cctttaccta aaataggatc cataattatt cacccccaat 32821 ctaacgcaat agcgataata aaattatacc agaaaggaga atcaacatga ctgaccaacc 32881 aagttactac tcaataatta cagcaaatgt cagatacgat aaccgactta ctgacagcga 32941 aaagttactt tttgcagaaa taacatcttt aagtaacaaa tacggatact gcacagcaag 33001 taatggttac tttgcaactt tatacaacgt tgttaaggaa actatatctc gtagaatttc 33061 gaaccttacc aactttggtt atctaaaaat cgaaattatc aaagaaggta atgaagttaa 33121 acaaaggaag atgtacccct tgacgcaaac gtcaatacct attgacgcaa aaatcaatac 33181 ccctattgat aattctgtca atacccctat tgacgcaaat gtcaaagaga atattacaag 33241 tattaataat acaagtaata acaatataaa tagaatagat atattg cgg gcaacccgac 33301 agcatcttct ataccctata aagaaattat cgattactta aacaaaaaag cgggcaagca 33361 ttttaaacac aatacagcta aaacaaaaga ttttattaaa gcaagatgga atcaagattt 33421 taggttggag gattttaaaa aggtgattga tatcaaaaca gctgagtggc taaacacgga 33481 tagcgataaa taccttagac cagaaacact ttttggcagt aaatttgagg ggtacctcaa 33541 tcaaaaaata caaccaactg gcacggatca attggaacgc atgaagtacg acgaaagtta 33601 ttgggattag ggggatatta tgaaaccact attcagcgaa aagataaacg aaagcttgaa 33661 aaaatatcaa cctactcatg tcgaaaaagg attgaaatgt gagagatgtg gaagtgaata 33721 cgacttatat aagtttgctc ctactaaaaa acacccgaat ggttacgagt ataaagacgg 33781 ttgcaaatgt gaaatctatg aggaatataa gcgaaacaag caacggaaga taaacaacat 33841 attcaatcaa tcaaacgtta atccgtcttt aagagatgca acagtcaaaa actacaagcc 33901 acaaaatgaa aaacaagtac acgctaaaca aacagcaata gagtacgtac aaggcttctc 33961 tacaaaagaa ccaaaatcat taatattgca aggttcatac ggaactggta aaagccacct 34021 agcatacgct atcgcaaaag cagtcaaagc taaagggcat acggttgctt ttatgcacat 34081 accaatgttg atggatcgta tcaaagcgac atacaacaaa aatgcagtag agactacaga 34141 cgagctagtc agattgctaa gtgatattga tttacttgta ctagatgata tgggtgtaga 34201 aaacacagag cacactttaa ataaactttt cagcattgtt gataacagag taggtaaaaa 34261 caacatcttt acaactaact ttagtgataa agaactaaat caaaatatga actggcaacg 34321 tataaattcg agaatgaaaa aaagagcaag aaaagtaaga gtaatcggag acgatttcag 34381 ggagcgagat gcatggtaac caaagaattt ttaaaaacta aacttgagtg ttcagatatg 34441 tacgctcaga aactcataga tgaggcacag ggcgatgaaa ataggttgta cgacctattt 34501 atccaaaaac ttgcagaacg tcatacacgc cccgctatcg tcgaatatta aggagtgtta 34561 aaaatgccga aagaaaaata ttacttatac cgagaagatg gcacagaaga tattaaggtc 34621 atcaagtata aagacaacgt aaatgaggtt tattcgctca caggagccca tttcagcgac 34681 gaaaagaaaa ttatgactga tagtgaccta aaacgattca aaggcgctca cgggcttcta 34741 tatgagcaag aattaggttt acaagcaacg atatttgata tttagaggtg gacgatgagt 34801 aaatacaacg ctaagaaagt tgagtacaaa ggaattgtat ttgatagcaa agtagagtgt 34861 gaatattacc aatatttaga aagtaatatg aatggcacta attatgatca tatcgaaata 34921 caaccgaaat tcgaattatt accaaaacta gataaacaac gaaagattga atatattgca 34981 gacttcgcgt tatatctcga tggcaaactg attgaagtta tcgacattaa aggtatgcca 35041 accgaagtag caaaacttaa agctaagatt ttcagacata aatacagaaa cataaaactc 35101 aattggatat gtaaagcgcc taagtataca ggtaaaacat ggattacgta cgaggaatta 35161 attaaagcaa gacgagaacg caaaagagaa atgaagtgat ctaatgcaac aacaagcata 35221 tataaatgca acgattgata taaggatacc tacagaagtt gaatatcagc attttgatga 35281 tgtggataaa gaaaaagaag cgctggcaga ttacttatat aacaatcctg acgaaatact 35341 agagtatgac aatttaaaaa ttagaaacgt aaatgtagag gtggaataaa tgggcagtgt 35401 tgtaatcatt aataataaac catataaatt taacaatttt gaaaaaagaa ataatggcaa 35461 agcgtgggat aaatgctgga attgtttcta aacgtgttag aggttgttgg gagttttcag 35521 aagctttaga cgcgccttat ggcatgcacc taaaagaata tagagaaatg aaacaaatgg 35581 aaaagattaa acaagcgaga ctcgaacgtg aattggaaag agagcgaaag aaagaggctg 35641 agctacgtaa gaagaagcca catttgttta atgtacctca aaaacattca cgtgatccgt 35701 actggttcga tgtcacttat aaccaaatgt tcaagaaatg gagtgaagca taatgagcat 35761 aatcagtaac agaaaagtag atatgaacaa aacgcaagac aacgttaagc aacctgcgca 35821 ttacacatac ggcgacattg aaattataga ttttattgaa caagttacgg cacagtaccc_ 35881 accacaatta gcattcgcaa taggtaatgc aattaaatac ttgtctagag caccgttaaa 35941 gaatggtcat gaggatttag caaaggcgaa gttttacgtc gatagagtat ttgacttgtg 36001 ggagtgatga ccatgacaga tagcggacgt aaagaatact taaaacattt tttcggctct 36061 aagagatatc tgtatcagga taacgaacga gtggcacata tccatgtagt aaatggcact 36121 tattactttc acggtcatat cgtgccaggt tggcaaggtg tgaaaaagac atttgataca 36181 gcggaagagc ttgaaacata tataaagcaa agtgatttgg aatatgagga acagaagcaa
36241 ctaactttat tttaaaaggg cggaaacaat gaaaatcaaa attgaaaaag aaatgaattt
36301 acctgaactt atccaatggg cttgggataa ccccaagtta tcaggtaata aaagattcta
36361 ttcaaatgat gttgagcgca actgttttgt gacttttcat gttgatagca tcttatgtaa
36421 tgtgactgga tatgtatcaa ttaacgataa atttactgtt caagaggaga tataacaatg
36481 aaaatcaaag ttaaaaaaga aatgagatta gatgaattaa ttaaatgggc gcgagaaaat
36541 ccggatctat cacaaggaaa aatatttttt tcaacaggat ttagtgatgg attcgttcgt
36601 tttcatccaa atacaaataa gtgttcgacg tcaagtttta ttccaattga tatccccttc
36661 atagttgata ttgaaaaaga agtaacggaa gagactaagg ttgataggtt gattgaatta
36721 ttcgagattc aagaaggaga ctataactct acactatatg agaacactag tataaaagaa
36781 tgtttatatg gcagatgtgt gcctaccaaa gcattctaca tcttaaacga tgacctaact
36841 atgacgttaa tctggaaaga tggggagttg ctagtatgat gttgaaattt aaagcttggg
36901 ataaagataa aaaagttatg agtattattg acgaaatcga ttttaatagt gggtacattt
36961 tgatttcaac aggttataaa agtttcaatg aagtaaaact attacaatac acaggattta
37021 aagatgtgca cggtgtggag atttatgaag gggatattgt tcaagattgt tattcgagag
37081 aagtaagttt tatcgagttt aaagaaggag ccttttatat aacttttagc aatgtaactg
37141 aattactaag tgaaaatgac gatattattg aaattgttgg aaatattttt gaaaatgaga
37201 tgctattgga ggttatgaga tgacgttcac cttatcagat gaacaatata aaaatctttg
37261 tactaactct aacaagttat tagataaact tcacaaagca ttaaaagatc gtgaagagta
37321 caagaagcaa cgagatgagc ttattgggga tatagcgaag ttacgagatt gtaacaaaga
37381 tctagagaag aaagcaagcg catgggatag gtattgcaag agcgttgaaa aagatttaat
37441 aaacgaattc ggtaacgatg atgaaagagt taaattcgga atggaattaa acaataaaat
37501 ttttatggag gatgacacaa atgaataatc gcgaaaaaat cgaacagtcc gttattagtg
37561 ctagtgcgta taacggtaat gacacagagg ggttgctaaa agagattgag gacgtgtata
37621 agaaagcgca agcgtttgat gaaatacttg agggaatgac aaatgctatt caacattcag
37681 ttaaagaagg tattgaactt gatgaagcag tagggattat ggcaggtcaa gttgtctata
37741 aatatgagga ggaataggaa aatgactaac acattacaag taaaactatt atcaaaaaat
37801 gctagaatgc ccgaacgaaa tcataagacg gatgcaggtt atgacatatt ctcagctgaa
37861 actgtcgtac tcgaaccaca agaaaaagca gtgatcaaaa cagatgtagc tgtgagtata
37921 ccagagggct atgtcggact attaactagt cgtagtggtg taagtagtaa aacgtattta
37981 gtgattgaaa caggcaagat agacgcggga tatcatggca atttagggat taatatcaag
38041 aatgatgaag aacgtgatgg aatacccttt ttatatgatg atatagacgc tgaattagaa
38101 gatggattaa taagcatttt agatataaaa ggtaactatg tacaagatgg aagaggcata
38161 agaagagttt accaaatcaa caaaggcgat aaactagctc aattggttat cgtgcctata
38221 tggacaccgg aactaaagca agtggaggaa ttcgaaagtg tttcagaacg tggagcaaaa
38281 ggcttcggaa gtagcggagt gtaaagacat cttagatcga gttaaggagg ttttggggaa
38341 gtgacgcaat acttagtcac aacattcaaa gattcaacag gacgaccaca tgaacatatt
38401 actgtggcta gagataatca gacgtttaca gttattgagg cagagagtaa agaagaagcg
38461 aaagagaagt acgaggcaca agttaaaaga gatgcagtta ttaaagtggg tcagttgtat
38521 gaaaatataa gggagtgtgg gaaatgacgg atgttaaaat taaaactatt tcaggtggag
38581 tttattttgt aaaaacagct gaaccttttg aaaaatatgt tgaaagaatg acgagtttta
38641 atggttatat ttacgcaagt actataatca agaaaccaac gtatattaaa acagatacga
38701 ttgaatcaat cacacttatt gaggagcatg ggaaatgaat cagctgagaa ttttattaca
38761 tgacggtagt agtttgatat tacatgaaga tgaattattt aacgaaatag tatttgtttt
38821 ggacaatttt agaaatgatg atgactattt aacgatagaa aaagattatg gcagagaact
38881 tgtattgaac aaaggttata tagttgggat caatgttgag gaggcagatg atgattaaca
38941 tacctaaaat gaaattcccg aaaaagtaca ctgaaataat caaaaaatat aaaaataaag
39001 cacctgaaga aaaggctaag attgaagatg attttattaa agaaattaaa gataaagaca
39061 gtgaatttta cagtcctacg atggctaata tgaatgaata tgaattaagg gctatgttaa
39121 gaatgatgcc tagtttaatt gatactggag atgacaatga tgattaaaaa acttaaaaat
39181 atggatgggt tcgacatctt tattgttgga atactgtcat tattcggtat attcgcattg
39241 ctacttgtta tcacattgcc tatctataca gtggctagtt accaacacaa agaattacat
39301 caaggaacta ttacagataa atataacaag agacaagata aagaagacaa gttctatatt
39361 gtattagaca acaaacaagt cattgaaaat tccgacttat tattcaaaaa gaaatttgat
39421 agcgcagata tacaagctag gttaaaagta ggcgataagg tagaagttaa aacaatcggt
39481 tatagaatac actttttaaa tttatatccg gtcttatacg aagtaaagaa ggtagataaa
39541 caatgattaa acaaatacta agactattat tcttactagc aatgtatgag ttaggtaagt
39601 atgtaactga gcaagtgtat attatgatga cggctaatga tgatgtagag gcgccgagtg
39661 attacgtctt tcgagcggag gtgagtgaat aatgagaata tttatttatg atttgatcgt
39721 tttgctgttt gctttcttaa tatccatata tattattgat gatggagtga taataaatgc
39781 attaggaatt tttggtatgt ataaaattat agattccttt tcagaaaata ttataaagag
39841 gtagataaaa atgaacgagc aaataatagg aagcatatat actttagcag gaggtgttgt
39901 gctttattca gttaaagaga tttttaggta ttttacagat tctaact.tac aacgtaaaaa_ _. - -
39961 aatcaattta gaacaaatat atccgatata tttagattgt tttaaaaagg ctaaaaagat ~
40021 gattggagct tatattattc caacagaaca gcatgaattt ttagattttt ttgatattga "~
40081 agtctttaat aatttagata agcaaagtaa aaaagcgtat gaaaatgtta ttggatttag
40141 acaaatgatt aatttatcaa atagagttaa ggcaatggaa gattttaaga tgagtttcaa
40201 caatgaattt agtacaaatc agattttttt taatccttct tttgttatgg aaacaattgc
40261 tattataaat gaatatcaaa aagatatatc ttatttaaaa aatataatta ataaaatgaa
40321 tgaaaataga gcttataatc atattgatag ttttatcact tcagagtacc gacgaaaaat
40381 aaacgattat aatctttatc ttgataaatt tgaagaacag tttagtcaaa agtttaaaat
40441 aaacagaact tcgataaaag aaagaattat tattaattta aacaagagga gatttaaatg
40501 atgtggatta ctatgactat tgtatttgct atattgctat tagtttgtat cagtattaat
40561 agtgatcgtg caagagagat acaagcactt agatatatga atgattatct acttgatgaa
40621 gtagttaaaa ctaaagggta caacgggtta gaagaataca ggattgaatt gaagcgaatg
40681 aataacgata ttaaaaagta atttatatta tcggaggtat tgcattgaat gataaagatt
40741 gagaaacacg atatcaaaaa gcttgaagaa tacattcagc acatcgataa ctatcgaaga
40801 gagttgaaga tgcgagaata tgaattactt gaaagtcatg aaccagataa tgcgggagct
40861 ggcaaaagta atttgccggg taacccgatt gaacgatgtg caataaagaa gtttagtgat
40921 aacaggtaca atacattaag aaatatagtt aacggtgtag atagattgat aggtgaaagt
40981 gatgaggata cgcttgagtt attaaggttt agatattggg attgtcctat tggttgttat
41041 gaatgggaag atatagcaca ttactttggt acaagtaaga caagtatatt acgtagaagg
41101 aatgcactga tcgataagtt agcaaagtat attggttatg tgtagcggac ttttacccta
41161 tgtaagtccg cattaaaaca gtttattatg ttagtatcag attaatattc aaagttatta
41221 aatgctaata cgacgcatga acaagaggcg catcactatg tgatgtgtcc ttttatttat
41281 gaggtatgaa catgttcaaa ctaattgtaa atacattact acacatcaag tatagatgag
41341 tcttgatact acttaagtta tataaggtga aacattatga tgactaaaga cgaacgtata
41401 cgattctata agtctaaaga atggcaaata acaagaaaaa gagtgctaga aagagataat
41461 tatgaatgtc aacaatgtaa gagagacggc aagttaacga catatgacaa aagcaagcgt
41521 aagtcgttgg atgtagatca tatattatcg ctagaacatc atccggagtt tgctcatgac
41581 ttaaacaatt tagaaacact gtgtattaaa tgtcacaaca aaaaagaaaa gagatttata
41641 aaaaaagaaa ataaatggaa agacgaaaaa tggtaaatac ccccgggtca aaaaaatcaa
41701 aagcgatc
Table 3
Name Position Name Position
77ORF005 19572..21026 48 77ORF052 1762..2013
77ORF006 3976..5196 49 77ORF053 37521.37757
77ORF007 21871..23076 50 77ORF054 22818..23060
77ORF008 2120..3307 51 77ORF055 17546..17788
77ORF009 31946..32803 52 77ORF058 18892..19122
77ORF010 26092..26889 53 77ORF059 34564.34785
77ORF011 24441..25208 54 77ORF064 29574..29795
77ORF012 29788..30576 55 77ORF065 28528-28746
77ORF013 33620..34399 56 77ORF066 27494..27703
77ORF014 27760..28512 57 77ORF069 38341.38547
77ORF015 3291..4028 58 77ORF070 36269.36475
77ORF016 32867..33610 59 77ORF071 40498..40701
77ORF017 23269.-23982 60 77ORF072 38735.38938
77ORF018 31169..31840 61 77ORF073 30945.31148
77ORF019 39851..40501 62 77ORF074 38544.38738
77ORF020 6926..7570 63 77ORF075 13673-13870
77ORF021 37762..38304 64 77ORF077 25357..25605
77ORF022 30605..31156 65 77ORF079 29089..29280
77ORF023 26903..27346 66 77ORF080 35204.35389
77ORF024 10700..11140 67 77ORF085 24060..24242
77ORF025 9707..10147 68 77ORF092 39706.39876
77ORF026 40729..41145 69 77ORF094 32226.32393
77ORF027 6518..6925 70 77ORF096 13606..13773
77ORF028 34795..35199 71 77ORF098 7092..7256
77ORF029 6117..6521 72 77ORF102 29051-29212
77ORF030 36478.36879 73 77ORF104 34393.34551
77ORF031 39151..39546 74 77ORF109 18282..18434
77ORF032 33892..34266 75 770RF112 39543.39692
77ORF033 5758..6120 76 770RF117 27361-27501
77ORF034 7886.-8236 77 770RF118 38390.38530
77ORF035 19258..19560 78 77ORF120 36059.36199
77ORF036 36876..37223 79 770RF124 33699.33833
77ORF037 102..446 80 770RF128 14221..14355
77ORF038 34908..35219 81 77ORF130 15675..15806
77ORF039 37220..37528 82 770RF133 8414..8542
77ORF040 41377..41676 83 77ORF140 13113-13235
77ORF041 35454.35753 84 770RF147 7029-7148
77ORF042 5490..5774 85 770RF149 30668..30787
77ORF043 29304..29564 86 770RF151 31837.31953
77ORF044 18481..18768 87 770RF155 30278.30391
77ORF045 5216..5500 88 770RF157 4044..4157
77ORF046 25603..25935 89 770RF167 20692-20799
77ORF047 11159..11425 90 770RF175 35717.35821
77ORF048 28776-29039 91 770RF176 6836..6940
77ORF049 36013.36255 92 770RF178 35390.35491
77ORF050 35753.36007 93 770RF179 8318..8419
77ORF051 38931.39167 94 77QRF182 29268-29564
Table 4
77ORF017 sequence
23982 atgacgcataatatagaaaaacgcattaataaattaaaaac tct
1 M T H N I E K R I N K K T S
23937 ggaaatccaaaatttaaaaagttagattcaga attcacta tta
16 G N P K F K K L D S D I H Y L
23892 ctcaagagatttgaaggtgaaaaaaaccataaaggtttttatcca
31 L K R F E G E K N H K G F Y P
23847 aagtttaaacaaggagaaatagtttttgtagatttcggtataaac
46 K F K Q G E I V F V D F G I N
23802 gttaataaagaattttctaattcacactttgcaatagtgatgaat
61 V N K E F S N S H F A I V N
23757 aaaaatgattctaatacggaggatatagtaaatgt at ccctta
76 K N D S N T E D I V N V I P L
23712 tcctctaaagaaaacaaaaagtatttaaagatgaattttgatttg 91 S S K E N K K Y L K M N F D L 23667 aaatgggagtattatttaagattgtttttaaatttaattagcgcg 106 K E Y Y L R L F L N L I S A 23622 caaaataattcagctatattaaaagaagttttcgataaaaaatac 121 Q N N S A I L K E V F D K K Y 23577 caaaaaaacaacacagaattcatcactaaagattattttattgaa 136 Q K N N T E F I T K D Y F I E
23532 tttatatctgatagtttagaaattgaaaataaattaaataaaatt
151 F I S D S L E I E N K L N K I
23487 gacagaaacattaataacatagtatcagcaattgataaggtaaaa
166 D R N I N N I V S A I D K V K
23442 aaattaaaaggtaatagttacgcttgcataaattctttccagccg
181 K L K G N S Y A C I N S F Q P
23397 attagtaagtttcgcataagaaaagttttaccccaaaaaattaaa
196 I S K F R I R K V L P Q K I K
23352 aatccagtaatagattcttcggatattatgttactgataaataga
211 N P V I D S S D I M L L I N R
23307 attaataataatatattgcagatccctgatataagatga 23269
226 I N N N I L Q I P D I R *
Physico-chemical parameters of ORF 77ORF017
1 MTHNIEKRIN KLKTSGNPKF KKLDSDIHYL LKRFEGEKNH KGFYPKFKQG EIVFVDFGIN
61 VNKEFSNSHF AIVMNKNDSN TEDIVNVIPL SSKENKKYLK MNFDLKWEYY LRLFLNLISA
121 QNNSAILKEV FDKKYQKNNT EFITKDYFIE FISDSLEIEN KLNKIDRNIN NIVSAIDKVK
181 KLKGNSYACI NSFQPISKFR IRKVLPQKIK NPVIDSSDIM LLINRINNNI LQIPDIR
Number of amino acids: 237
Average molecular weight (Daltons): 27887.38
Mean amino acid weight (Daltons): 117.67
Monoisotopic molecular weight (Daltons): 27869.83
Mean amino acid monoisotopic weight (Daltons): 117.59
Amino acid composition
Number of acidic (negative) amino acids (ED): 27
11.39%
Number of basic (positive) amino acids (KR): 41
17.30%
Total charge (KRED): 68
28.69%
Net charge (KR - ED): 14
5.91%
Theoritical pi: 10.01
Total linear charge density: 0.30
Average hydrophobicity: -5.37
Ratio of hydrophilicity to hydrophobicity: 1.41
Percentage of hydrophilic amino acid: 57.81%
Percentage of hydrophobic amino acid: 42.19%
Ratio of %hydrophilic to %hydrophobic: 1.37
133
77ORF019 sequence
39851 atgaacgagcaaataataggaagcatatatactttagcaggaggt
1 M N E Q I I G S I Y T L A G G
39896 gttgtgctttattcagttaaagagatttttaggtattttacagat
16 V V L Y S V K E I F R Y F T D
39941 tctaacttacaacgtaaaaaaatcaatttagaacaaatat ccg
31 S N L Q R K K I N L E Q I Y P
39986 atatatttagattgttttaaaaaggctaaaaagatgattggagct 46 I Y L D C F K K A K K M I G A
40031 tatattattccaacagaacagcatgaatttttagatttttttgat 61 Y I I P T E Q H E F L D F F D 40076 attgaagtctttaataatttagataagcaaagtaaaaaagcgtat 76 I E V F N N L D K Q S K K A Y
40121 gaaaatgtta ggatttagacaaatgattaatttatcaaataga
91 E N V I G F R Q M I N L S N R
40166 gttaaggcaatggaagattttaagatgagtttcaacaatgaattt
106 V K A M E D F K M S F N N E F
40211 agtacaaatcagattttttttaatccttcttttgttatggaaaca
121 S T N Q I F F N P S F V M E T
40256 attgctattataaatgaa atcaaaaagatatatcttatttaaaa
136 I A I I N E Y Q K D I S Y L K
40301 aata aattaataaaatgaatgaaaatagagcttataatcatatt
151 N I I N K M N E N R A Y N H I
40346 gatagttttatcacttcagagtaccgacgaaaaataaacga at
166 D S F I T S E Y R R K I N D Y
40391 aatctt atcttgataaatttgaagaacagtttagtcaaaagttt
181 N L Y L D K F E E Q F S Q K F
40436 aaaataaacagaacttcgataaaagaaagaattattattaattta
196 K I N R T S I K E R I I I N L
40481 aacaagaggagatttaaatga 40501
211 N K R R F K *
Physico-chemical parameters of ORF 77ORF019
1 MNEQIIGSIY TLAGGWLYS VKEIFRYFTD SNLQRKKINL EQIYPIYLDC FKKAKKMIGA
61 YIIPTEQHEF LDFFDIEVFN NLDKQSKKAY ENVIGFRQMI NLSNRVKAME DFKMSFNNEF
121 STNQIFFNPS FVMETIAIIN EYQKDISYLK NIINKMNENR AYNHIDSFIT SEYRRKINDY
181 NLYLDKFEEQ FSQKFKINRT SIKERIIINL NKRRFK
Number of amino acids: 216
Average molecular weight (Daltons): 26026.06
Mean amino acid weight (Daltons): 120.49
Monoisotopic molecular weight (Daltons): 26009.34
Mean amino acid monoisotopic weight (Daltons): 120.41
Amino acid composition
Number of acidic (negative) amino acids (ED): 26
12.04%
Number of basic (positive) amino acids (KR): 33
15.28%
Total charge (KRED): 59
27.31%
Net charge (KR - ED): 7
3.24%
Theoritical pi: 9.52
Total linear charge density: 0.28
Average hydrophobicity: -4.84
Ratio of hydrophilicity to hydrophobicity: 1.37
Percentage of hydrophilic amino acid: 54.17%
Percentage of hydrophobic amino acid: 45.8-3%
Ratio of %hydrophilic to %hydrophobic: 1.18 -
77ORF043 sequence
29304 atgtattacgaaataggcgaaatcatacgcaaaaatatt catgtt
1 M Y Y E I G E I I R K N I H V
29349 aacggattcgattttaagctattcattttaaaaggtcatatgggc 16 N G F D F K L F I L K G H M G
29394 a a caatacaagttaaagatatgaacaacgtaccaattaaacat 31 I S I Q V K D M N N V P I K H 29439 gcttatgtcgtagatgagaatgacttagatatggcatcagactta 46 A Y V V D E N D L D M A S D L 29484 tttaaccaagcaatagatgaatggattgaagagaacacagacgaa 61 F N Q A I D E I E E N T D E 29529 caggacagactaattaacttagtcatgaaatggtag 29564 76 Q D R L I N L V M K W *
Physico-chemical parameters of ORF 77ORF043
1 MYYEIGEIIR KNIHVNGFDF KLFILKGHMG ISIQVKDMNN VPIKHAYWD ENDLDMASDL 61 FNQAIDEWIE ENTDEQDRLI NLVMKW
Number of amino acids: 86
Average molecular weight (Daltons): 10186.68
Mean amino acid weight (Daltons): 118.45
Monoisotopic molecular weight (Daltons): 10180.02
Mean amino acid monoisotopic weight (Daltons): 118.37
Amino acid composition
Number of acidic (negative) amino acids (ED): 16
18.60%
Number of basic (positive) amino acids (KR): 8
9.30%
Total charge (KRED): 24
27.91%
Net charge (KR - ED):
9.30%
Theoritical pi: 4.38
Total linear charge density: 0.30
Average hydrophobicity: -2.80
Ratio of hydrophilicity to hydrophobicity: 1.19
Percentage of hydrophilic amino acid: 48.84%
Percentage of hydrophobic amino acid: 51.16%
Ratio of %hydrophilic to %hydrophobic: 0.95
77ORF102 sequence
29051 atgagcaacatttataaaagc acc agtagcagtat atgcttc
1 M S N I Y K S Y L V A V L C F
29096 acagtcttagcgattgtacttatgccgtttctatacttcactaca 16 T V L A I V L M P F L Y F T T
29141 gcatggtcaattgcgggattcgcaagtatcgcaacattcatgtac 31 A S I A G F A S I A T F M Y 29186 tacaaagaatgctttttcaaagaataa 29212 46 Y K E C F F K E *
Physico-chemical parameters of ORF 77ORF102
MSNIYKSYLV AVLCFTVLAI VLMPFLYFTT AWSIAGFASI ATFMYYKECF FKE
Number of amino acids: 53
Average molecular weight (Daltons): 6155.42
Mean amino acid weight (Daltons): 116.14
Monoisotopic molecular weight (Daltons): 6151.07
Mean amino acid monoisotopic weight (Daltons): 116.06
Amino acid composition
Number of acidic (negative) amino acids (ED): 2
3.77%
Number of basic (positive) amino acids (KR): 3
5.66%
Total charge (KRED): 5
9.43%
Net charge (KR - ED): 1
1.89%
Theoritical pi: 8.18
Total linear charge density: 0.13 -
Average hydrophobicity: ιo.8f
Ratio of hydrophilicity to hydrophobicity: 0.40
Percentage of hydrophilic amino acid: 28.30%
Percentage of hydrophobic amino acid: 71.70%
Ratio of %hydrophi!ic to %hydrophobic: 0.39
77ORF104 sequence
34393 atggtaaccaaagaatttttaaaaactaaacttgagtgttcagat
1 M V T K E F L K T K L E C S D
34438 atgtacgctcagaaactcatagatgaggcacagggcgatgaaaat
16 M Y A Q K L I D E A Q G D E N
34483 aggttgtacgacc atttatccaaaaacttgcagaacgtcataca
31 R L Y D L F I Q K L A E R H T
34528 cgccccgctatcgtcgaatattaa 34551
46 R P A I V E Y *
Physico-chemical parameters of ORF 77ORF104
1 MVTKEFLKTK LECSDMYAQK LIDEAQGDEN RLYDLFIQKL AERHTRPAIV EY
Number of amino acids: 52
Average molecular weight (Daltons): 6193.13
Mean amino acid weight (Daltons): 119.10
Monoisotopic molecular weight (Daltons): 6189.12
Mean amino acid monoisotopic weight (Daltons): 119.02
Amino acid composition
Number of acidic (negative) amino acids (ED): 10
19.23%
Number of basic (positive) amino acids (KR): 8
15.38%
Total charge (KRED): 18
34.62%
Net charge (KR - ED): -2
3.85%
Theoritical pi: 5.03
Total linear charge density: 0.38
Average hydrophobicity: -5.81 "
Ratio of hydrophilicity to hydrophobicity: 1.47
Percentage of hydrophilic amino acid: 53.85%
Percentage of hydrophobic amino acid: 46.15%
Ratio of "/ohydrophilic to %hydrophobic: 1.17
77ORF182 sequence
29268 atgttcaatataaaacgaaaaacggaggaagtcaagatgtattac
1 M F N I K R K T E E V K M Y Y 29313 gaaataggcgaaatcatacgcaaaaa a catgttaacggattc 16 E I G E I I R K N I H V N G F
29358 gattttaagctattcattttaaaaggtcatatgggcatatcaata 31 D F K L F I L K G H M G I S I
29403 caagttaaagatatgaacaacgtaccaattaaacatgcttatgtc
46 Q V K D M N N V P I K H A Y V
29448 gtagatgagaatgacttagatatggcatcagacttatttaaccaa
61 V D E N D L D M A S D L F N Q
29493 gcaatagatgaatgga tgaagagaacacagacgaacaggacaga
76 A I D E I E E N T D E Q D R
29538 ctaattaacttagtcatgaaatggtag 29564
91 L I N L V M K W *
Physico-chemical parameters of ORF 770RF182
1 MFNIKRKTEE VKMYYEIGEI IRKNIHVNGF DFKLFILKGH MGISIQVKDM NNVPIKHAYV 61 VDENDLDMAS DLFNQAIDEW lEENTDEQDR LINLVMKW
Number of amino acids: 98
Average molecular weight (Daltons): 11691.50
Mean amino acid weight (Daltons): 119.30
Monoisotopic molecular weight (Daltons): 11683.84
Mean amino acid monoisotopic weight (Daltons): 119.22
Amino acid composition
Number of acidic (negative) amino acids (ED): 18
18.37%
Number of basic (positive) amino acids (KR): 12
12.24%
Total charge (KRED): 30
30.61%
Net charge (KR - ED): -6
6.12%
Theoritical pi: 4.76 -
Total linear charge density: 0.33
Average hydrophobicity: -3.89
Ratio of hydrophilicity to hydrophobicity: 1.28
Percentage of hydrophilic amino acid: 51.02%)
Percentage of hydrophobic amino acid: 48.98%
Ratio of "/ohydrophilic to %hydrophobic: 1.04
Table 5
B ASTP 2.0.8 [Jan-05-1999]
Query= sid| 100017 | lan| 77ORF017 Phage 77 ORF | 23269-23982 | -3 (237 letters)
Database: nr
393,678 sequences; 120,452,765 total letters
Score E
Sequences producing significant alignments: (bits) Value
4493986|emb|CAB39045.l| (AL034559) predicted using hexExon; . 41 0.010 730607|sp|P23250|RPIl_YEAST NEGATIVE RAS PROTEIN REGULATOR P. 38 0.053 3097044|emb|CAA75299| (Y15035) KIR [Cowpox virus] 38 0.090 2146245 jpirj |S73794 hypothetical protein H91_orfl80 - Mycopl. 38 0.090 83910 |pir I |S04682 ribosomal protein varl - yeast (Candida gl . 37 0.15 133135 |sp|P21358|RMAR_CANGA MITOCHONDRIAL RIBOSOMAL PROTEIN . 37 0.15 2128843 |pir| |H64475 hypothetical protein MJ1409 - Methanococ. 36 0.20 5107017|gb|AAD39926.l|AF126285_2 (AF126285) RNA polymerase [. 36 0.35 2146210 I pir I |S73342 hypothetical protein E07_orfl66 - Mycopl. 35 0.60
Database: swissprot
79,449 sequences; 28,874,452 total letters
Score E
Sequences producing significant alignments: (bits) Value sp P23250 RPI1_ _YEAST NEGATIVE RAS PROTEIN REGULATOR PROTEIN. 38 0.014 sp P21358 RMAR" "CANGA MITOCHONDRIAL RIBOSOMAL PROTEIN VAR1. 37 0.040 sp Q21444 LDLC] "CAEEL LDLC PROTEIN HOMOLOG. 34 0.35 sp P27240 RFAY] "ECOLI LIPOPOLYSACCHARIDE CORE BIOSYNTHESIS PROT. 33 0.46 sp P53192 YGCO^ ~YEAST HYPOTHETICAL 27.1 KD PROTEIN IN ALK1-CKB1. 33 0.60 sp P32908 SMC1~ ~YEAST CHROMOSOME SEGREGATION PROTEIN SMC1 (DA-B. 33 0.60 sp P54683 TAGB" "DICDI PRESTALK-SPECIFIC PROTEIN TAGB PRECURSOR . 32 0.78 sp Q03100 CYAA" "DICDI ADENYLATE CYCLASE, AGGREGATION SPECIFIC (. 32 0.78
BLASTP 2.0.8 [Jan-05-1999]
Query= sid| 100019 | lan| 77ORF019 Phage 77 ORF| 39851-40501 | 2 (216 letters)
Database: nr
373,355 sequences; 114,214,446 total letters
Score E
Sequences producing significant alignments: (bits) Value gi 3341966 |dbj | BAA3 19321 (AB009866) orf 59 [bacteriophage phi PVL] 437 e-122 gi 2689911 (AE00079 2) B. burgdorferi predicted coding region BB 38 0.058 gi 1171589 |emb|CAA645741 (X95275) frameshift [Plasmodium falcip 37 0.10 gi 4493986 |emb|CAB39045.11 (AL034559) predicted using hexExon; 36 23 gi 141257|sp|P18019 |YPI9_CL0PE HYPOTHETICAL 14.5 KD PROTEIN (OR 36 29 gi 133412 jsp|P27059 |RP0B_ASTL0 DNA-DIRECTED RNA POLYMERASE BETA 35 51 i 312223l|sp|Q58851|HISX_METJA HISTIDINOL DEHYDROGENASE (HDH) 35 51 gi 3649757 |emb|CABl1106.11 (Z98547) predicted using hexExon; MA 34 0.66 gi 2688313 (AE001146) sensory transduction histidine kinase, pu 34 0.87
Database: swissprot
79,449 sequences; 28,874,452 total letters
Score E
Sequences producing significant alignments: (bits ) Value sp P18019 YPI9_CLOPE HYPOTHETICAL 14.5 KD PROTEIN (ORF9) . 36 0 079 sp Q58851 HISX_METJA HISTIDINOL DEHYDROGENASE (EC 1.1.1.23) (H. 35 0 14 sp P27059 RPOB_ASTLO DNA-DIRECTED RNA POLYMERASE BETA CHAIN (E. 35 0 14 sp Q02224 CENE_HUMAN CENTROMERIC PROTEIN E (CENP-E PROTEIN) . 34 0 31 sp P04931 ARP_PLAFA ASPARAGINE-RICH PROTEIN (AG319) (ARP) (FRA. 33 0 53 sp P18011 IPAB_SHIFL 62 KD MEMBRANE ANTIGEN. 32 0 69 sp P18709 VTA2_XENLA VITELLOGENIN A2 PRECURSOR (VTG A2) [CONTA. 32 0 90 sp Q64409 CP3H_CAVPO CYTOCHROME P450 3A17 (EC 1.14.14.1) (CYPI. 32 0 90 sp P21358 RMAR_CANGA MITOCHONDRIAL RIBOSOMAL PROTEIN VAR1. 32 0 90 sp Q03945 IPAB_SHIDY 62 KD MEMBRANE ANTIGEN. 32 1 2
70
BLASTP 2.0.8 [Jan-05-1999]
Query= sid| 100043 | lan| 77ORF043 Phage 77 ORF | 29304-29564 | 3 (86 letters)
Database: nr
373,355 sequences; 114,214,446 total letters
Score E
Sequences producing significant alignments: (bits) Value gi 3341947 |dbj |BAA31913 | (AB009866) orf 39 [bacteriophage phi PVL]] 182 6e-46 gi 744518 |prf I I 2014422A FKBP-rapamycin-associated protein [Homo. .. 32 0.84 gi 1169736 |sp|P42346|FRAP_RAT FKBP-RAPAMYCIN ASSOCIATED PROTEIN. .. 32 0.84 gi 1169735 |sp|P42345|FRAP_HUMAN FKBP-RAPAMYCIN ASSOCIATED PROTE . .. 32 0.84 gi 3282239 (U88966) rapamycin associated protein FRAP2 [Homo sa. .. 32 0.84 i 3875402|emb|CAA98122 I (Z73906) cDNA EST EMBL:D64544 comes fr. .. 31 2.5 gi 1084792 jpirj |S54091 hypothetical protein YPR070w - yeast (Sa. .. 30 4.2
Database: swissprot
79,449 sequences; 28,874,452 total letters
Score E
Sequences producing significant alignments: (bits) Value sp P42345 FRAP_HUMAN FKBP-RAPAMYCIN ASSOCIATED PROTEIN (FRAP) . 32 0.24 sp P42346 FRAP RAT FKBP-RAPAMYCIN ASSOCIATED PROTEIN (FRAP) (R. 32 0.24 sp P34554 YNP1_CAEEL HYPOTHETICAL 42.2 KD PROTEIN T05G5.1 IN C. 28 3.5 sp Q24118 LIO_DROME LINOTTE PROTEIN. 28 3.5 sp P80034 ACH2_BOMMO ANTICHYMOTRYPSIN II (ACHY-II) . 28 3.5 sp P22922 AlAT_BOMMO ANTITRYPSIN PRECURSOR (AT) . 28 3.5 sp Q44363 TRAA_AGRT6 CONJUGAL TRANSFER PROTEIN TRAA. 28 3.5 sp P38255 YBU5_YEAST HYPOTHETICAL 51.3 KD PROTEIN IN PH05-VPS1. 27 6.0 sp P55822 SH3B_HUMAN SH3BGR PROTEIN (21-GLUTAMIC ACID-RICH PRO. 27 7.9 sp Q58482 YA82_METJA HYPOTHETICAL PROTEIN MJ1082. 27 7.9 sp P34252 YKK8_YEAST HYPOTHETICAL 52.3 KD PROTEIN IN HAP4-AAT1. 27 7.9
BLASTP 2.0.8 [Jan-05-1999]
Query= sid| 100102 | Ian | 77ORF102 Phage 77 ORF| 29051-29212 | 2 (53 letters)
Database: nr
373,355 sequences; 114,214,446 total letters
Score E
Sequences producing significant alignments: (bits) Value gi|3341946|dbj |BAA31912| (AB009866) orf 38 [bacteriophage phi PVL] 96 3e-20 gij 4325288 |gb|AAD17315| (AF123593) voltage-dependent sodium cha ... 28 7.1 gi|2649684 (AE001040) A. fulgidus predicted coding region AF092... 28 9.3
Database: swissprot
79,449 sequences; 28,874,452 total letters
Score E
Sequences producing significant alignments: (bits) Value sp|P42087 HUTM_BACSU PUTATIVE HISTIDINE PERMEASE . 26 7.1 sp|P04775 CIN2_RAT SODIUM CHANNEL PROTEIN, BRAIN II ALPHA SUBU... 26 9.2 sp|P42619 YQJF_ECOLI HYPOTHETICAL 17.2 KD PROTEIN IN EXUR-TDCC... 26 9.2
BLASTP 2.0.8 [Jan-05-1999]
Query= sid| 100104 | Ian | 77ORF104 Phage 77 ORF | 34393-34551 | 1 (52 letters)
Database: nr
373,355 sequences; 114,214,446 total letters
Score E
Sequences producing significant alignments: (bits) Value gi 12315523 (AF016452) similar to the leucine-rich domains found... 29 4.2 gi|4377168|gb|AAD18990| (AE001666) CT711 hypothetical protein [... 29 5.4 gij 3882171 j dbj |BAA34445| (AB018268) KIAA0725 protein [Homo sapi... 28 9.3
Database: swissprot
79,449 sequences; 28,874,452 total letters
Score E
Sequences producing significant alignments: (bits) Value sp|P04879 RRPP_VSVIG RNA POLYMERASE ALPHA SUBUNIT (EC 2.7.7.48. 27 5.4 sp|P04880 RRPP_VSVIM RNA POLYMERASE ALPHA SUBUNIT (EC 2.7.7.48. 27 5.4 sp|Q13946 CN7A_HUMAN HIGH-AFFINITY CAMP-SPECIFIC 3', 5 '-CYCLIC . 26 7.1 sp|P35381 ATPA_DROME ATP SYNTHASE ALPHA CHAIN, MITOCHONDRIAL P. 26 9.3 sp|P54659 MVPB_DICDI MAJOR VAULT PROTEIN BETA (MVP-BETA) . 26 9.3 sp|P40397 YHXC_BACSU HYPOTHETICAL OXIDOREDUCTASE IN APRE-COMK . 26 9.3
[ 73
BLASTP 2.0.8 [Jan-05-1999]
Query= sid| 122748 | lan| 770RF182 Phage 77 ORF| 29268-29564 | 3 (98 letters)
Database: nr
393,678 sequences; 120,452,765 total letters
Score E
Sequences producing significant alignments: (bits) Value gi I 3341947 I dbj |BAA31913.l| (AB009866) orf 39 [bacteriophage phi .. 182 8e-46 gi|l084792 jpirj |S54091 hypothetical protein YPR070w - yeast (Sa.. 35 0.13 gi|H69736|sp|P42346|FRAP_RAT FKBP-RAPAMYCIN ASSOCIATED PROTEIN.. 32 1.1 gi j 744518 |prf j I 2014422A FKBP-rapamycin-associated protein [Homo.. 32 1.1 gij 5051381 |emb|CAB44736.l| (AL049653) dJ647M16.2 (FK506 binding .. 32 1.1 gi|4826730|ref |NP_004949.1 |pFRAPl | FK506 binding protein 12-rap.. 32 1.1 gi|3282239 (U88966) rapamycin associated protein FRAP2 [Homo sa.. 32 l.i
Database: swissprot
79,909 sequences; 29,054,478 total letters
Score E
Sequences producing significant alignments: (bits) Value
)|P42345 FRAP_HUMAN FKBP-RAPAMYCIN ASSOCIATED PROTEIN (FRAP) . 32 0.29
)|P42346 FRAP_RAT FKBP-RAPAMYCIN ASSOCIATED PROTEIN (FRAP) (R. 32 0.29
)|P40557 YIA5_YEAST PUTATIVE DISULFIDE ISOMERASE YIL005 PREC. 29 3.3
)|Q24118 LIO_DROME LINOTTE PROTEIN. 28 4.4
)|Q44363 TRAA_AGRT6 CONJUGAL TRANSFER PROTEIN TRAA. 28 4.4
)|P80034 ACH2_BOMMO ANTICHYMOTRYPSIN II (ACHY-II). 28 4.4
)|P34554 YNP1_CAEEL HYPOTHETICAL 42.2 KD PROTEIN T05G5.1 IN C. 28 4.4
)|P22922 AlAT_BOMMO ANTITRYPSIN PRECURSOR (AT). 28 4.4
Table 6
Table 7
Bacteriophage 3A, complete genome sequence
1 caaacgctag caacgcggat aaatttttca tgaaaggggg tctttatatg aagttaacaa aaaaacagct
71 aaaagaatat atagaagatt acaaaaaatc tgatgacata ttaattaatt tgtatataga aacatatgaa
141 ttttattgtc ggttaagaga tgaacttaaa aatagtgatt taatgataga gcatacaaac aaggctggtg
211 cgagcaatat tattaagaat ccattaagca tagaactgac aaaaacagtt caaacactaa ataacttact
281 caagtctatg ggtttaactg cagcacaaag aaaaaagata gttcaagaag aaggtggatt cggtgactat
351 taaagtttta aatgaacctt caccaaaact at aacaaca tggtatgcag agcaagtcac tcaagggaaa
421 ataaaaacaa gcaaatatgt tagaaaagaa tgtgagagac atcttagata tctagaaaat ggaggtaaat
491 gggtatttga tgaagaatta gcgcatcgtc ctattcgatt tatagaaaag ttttgtaaac cttccaaagg
561 atctaaacgt caacttgtat tacagccatg gcaacatttt attatcggca gtttgtttgg ttgggttcat
631 aaagaaacaa aactgcgcag gtttaaagaa gctttgatat ttatggggcg aaaaaatggt aaaacaacca
701 ctatttctgg ggttgctaac tatgctgtat cacaagatgg agaaaatggt gcagaaattc atttgttagc
771 aaacgtaatg aaacaagcta ggattctatt tgatgaatct aaggcgatga ttaaagctag cccaaagctt
841 ga aaaatt tcagaacatt aagagatgaa atccattatg acgcaacgat atcaaaaatt atgccccaag
911 catcagatag cgataagtta gatggattga atacacacat ggggattttt gatgaaattc atgaatttaa
981 agactataaa ttgatttcag ttataaaaaa ctcaagagct gcaaggttac aacctcttct catctacatt
1051 acgacagcag ggtatcaatt agatggtcca cttgttgata tggtagaagc gggaagagac accttagatc
1121 aaatcataga agacgaaaga actttttatt atttagcatc tttggatgat gacgatgata ttaatgattc
1191 gtcgaactgg ataaaagcaa atcccaactt aggtgtctct ataaatttag atgagatgaa agaagagtgg
1261 gaaaaagcta agagaacacc agctgaacgt ggagatttta taaccaaaag gtttaatatc tttgctaata
1331 atgacgagat gagttttatt gattacccaa cactccaaaa aaataatgaa attgtttctt tagaagagct
1401 ggaaggcaga ccgtgcacga ttggttatga tttatcagaa acagaggact ttacagccgc gtgtgctact
1471 tttgcgttag ataatggtaa agttgcagtt ttatcgcatt catggattcc taagcacaaa gttgaatatt
1541 ctaacgaaaa aataccctat agagaatggg aagaagatgg cttattaaca gtgcaagata agccttatat
1611 tgactaccaa gatgttttaa attggataat taagatgaat gagcattatg tagtagaaaa aattacttat
1681 gatagagcga acgcattcaa actaaatcaa gagttaaaaa attacgggtt tgaaacggaa gaaacaagac
1751 aaggagcttt gaccttgagc cctgcattga aggatttaaa agaaatgttt ttagatggga aaataatatt
1821 taataataat cctttaatga aatggtatat caataatgtt cagttgaaac tagacagaaa cggaaactgg
1891 ttgccgtcta agcaaagcag atatcgtaaa atagatggct ttgcagcatt tttaaacaca tatacagata
1961 ttatgaataa agttgtttct gatagtggtg aaggaaacat agagttta t agtattaaag acataatgcg
2031 ttaaggaggt gaatgttatc gcaaaagaga atattgtcac acgcataaag aaaaaattga tagacaattg
2101 gattgatcag tcaacttcta agctttatga ctttagccca tggaaaaata gatctttttg gggtgtaatt
2171 aataatacgc ttgaaactaa tgaaacgata ttttcagcta ttacaaagtt atctaattcg atggctagtt
2241 tgcccttgaa aatgtatgaa gattataaag tagttaatac agaagtatct gatttactta cagtgtcacc
2311 gaataattct ctgagcagtt ttgattttat taatcaaatt gaaacaatca gaaatgaaaa aggtaatgca
2381 tatgtgctaa ttgaacgaga catctatcat caaccatcaa agcttttctt attaaatcca gatgttgttg
2451 aaatgttaat tgaaaaccaa tcacgtgaac tttattattc cattcatgct gcaactggaa ataaattgat
2521 tgttcataat atggacatgt tgcattttaa acacatcgtg gcatctaata tggtgcaagg cattagtccg
2591 attgatgtgt tgaagaatac aactgatttt gataatgcag taagaacctt taatcttaca gaaatgcaaa
2661 aacctgattc tttcatgctt aaatatggtt ccaatgtagg taaagaaaaa aggcagcaag tgttagaaga
2731 tttcaaacag tactatgaag aaaacggtgg aatattattc caagagcctg gtgttgaaat cgaaccgtta
2801 cctaaaaaat atgtctctga agatatagtg gcaagcgaga atttaacaag agaaagagta gctaacgttt
2871 ttcaattgcc ctcagtattc ttaaatgcaa gatcaaatac aaatttcgcg aaaaatgaag agttaaacag
2941 attttacttg cagcatacct tattgccaat cgtcaaacag tatgaagaag aatttaatcg gaaactactt
3011 actaaaacag acagagaaaa aaataggtat tttaaattta acgttaaatc ttatttaagg gctgatagtg
3081 caacacaagc agaagtgtac tttaaagcag ttcgtagtgg ttactacact ataaatgaca ttagagagtg
3151 ggaagattta ccaccagttg aaggtggaga taagccgcta ataagcggtg atttataccc aattgacacg
3221 ccacttgaat taagaaaatc tttgaaaggt ggtgataaaa atgtcaatga aagctaagta ttttcaaatg
3291 aaaagaaaat caaaaagtaa aggtgaaata tttatttatg gtgatattgt aagtgataaa tggtttgaaa
3361 gtgatgtaac tgctacagat ttcaaaaata aactagatga actaggagac atcagtgaaa tagatgttca
3431 tataaattca tctggaggca gtgtatttga agggcatgca atatacaata tgctaaaaat gcatcctgca
3501 aaaattaata tctatgtcga tgccttagcg gcatcaattg ctagtgttat cgctatgagt ggtgacacta
3571 tttttatgca caaaaatagt tttttaatga ttcataattc atgggttatg actgtaggta atgcagaaga
3641 gttaagaaag acagcggatt tacttgaaaa aacagatgct gttagtaatt cagcttattt agataaagca
3711 aaagatttag atcaagaaca cttaaaacag atgttagatg cagaaacttg gcttactgca gaagaagcct
3781 tgtctttcgg cttgatagat gaaattttag gagctaatga aataactgct agtatctcta aagagcaata
3851 taagcgtttc gagaacgtcc cagaagattt aaagaaagat gtagacaaaa tcactaaaat cgatgatgta
3921 gatacgtttg aattggttga aacacctaaa gaaagtatgt cactagaaga aaaagaaaaa agagaaaaaa
3991 ttaaacgcga atgcgaaatt ttaaaaatga caatgagtta ttaggaggaa atgaaatgcc gacattatat
4061 gaattaaaac aatccttagg tatgattgga caacaattaa aaaataaaaa tgatgaattg agtcagaaag
4131 caacagaccc aaatattgat atggaagaca tcaaacaact agaaacagaa aaagcaggct tacaacaaag
4201 atttaacatt gttgaaagac aagtaaaaga cattgaagaa aaagaaaaag cgaaagttaa agacacagga
4271 gaagcttatc aatctttaaa tgatcatgag aagatggtta aagctaaggc agagttttat cgtcacgcga
4341 ttttaccaaa tgaatttgaa aaaccttcaa tggaggcaca acgtttatta cacgctttac caacaggtaa
4411 tgattcaggt ggtgataagc tcttaccaaa aacactttct aaagaaattg tttcagaacc atttgctaaa
4481 aaccaattac gtgaaaaagc tcgtctaact aacattaaag gtttagagat tccaagagtt tcatatactt
4551 tagacgatga tgacttcatt acagatgtag aaacagcaaa agaattaaaa ttaaaaggtg atacagttaa
4621 attcactact aataaattca aagtatttgc tgcaatttca gatactgtaa ttcatggatc agatgtagat
4691 ttagtaaact gggttgaaaa cgcactacaa tcaggtctag cagctaaaga acgtaaagat gccttagcag
4761 taagtcctaa atctggatta gatcacatgt cattttacaa tggatctgtt aaagaagttg agggagcaga
4831 catgtatgat gctattatta acgctttagc agatttacat gaagattacc gtgataacgc aacaatttat
4901 atgcgatatg cggattatgt caaaattatt agtgttcttt caaatggaac aacaaatttc tttgacacac 4971 cagcagaaaa agtatttggc aaaccagtag tatttacaga tgcagcagtt aaacctattg tgggagattt 5041 caattatttt ggaattaact atgatggaac aacttatgac actgataaag atgttaaaaa aggcgaatat 5111 ttgtttgtat taactgcatg gtatgatcag caacgtacat tagacagtgc attcagaatt gcaaaagcaa 5181 aagaaaatac aggttcatta cccagctaag ccccaaaagg ttaatgtaac agctaaggct aaatcagctg 5251 taatatcagc cgaatagggg tgatgaaatg agtttagaag aaattaaatt gtggttgaga attgactata 5321 atttcgaaaa tgatttaatt gaaggtctca ttcaatcggc taagtctgaa ttactattaa gtggggttcc 5391 agattatgac aaagatgact tggaataccc gcttttttgt acagcgatta gatatatcat tgcaagagat 5461 tatgaaagtc gtgggtactc aaatgaccaa tctagaagca aggtttttaa tgaaaaggga ttgcaaaaaa 5531 tgattctgaa attaaaaaag tggtaggtga tttttaaatg gaatttaatg aatttaaaga tcgcgcatat 5601 ttttttcaat atgtaaataa agggccgtat ccagatgaag aggaaaaaat gaagttgtat agttgctttt 5671 gtaaaatata taatccttct atgaaagata gagaaatttt aaaagcgact gaatcaaagt caggactaac 5741 cataattatg aggtcttcta aaattgaata tctaccacaa acaaatcact tagttaaaat tgacagaggc 5811 ttatattccg ataaattatt caacattaaa gaaataagaa ttgatacacc agatattggc tataatacag 5881 tggttttatc agaaaaatga gtgtagaaat taaagggata cctgaagtgt tgaagaaatt agaatcggta
5951 tacggtaaac aatcaatgca agctaagagt gatagagctt taaatgaagc atctgaattt tttataaagg 6021 ctttaaagaa agaattcgag agttttaaag atacgggtgc tagcatagaa gaaatgacta aatctaagcc 6091 ttatacaaaa gtaggaagtc aagaaagagc tgttttaatt gaatgggtag gccctatgaa tcgcaaaaac 6161 attattcact tgaatgaaca tggttataca agagatggaa aaaaatatac accaagaggt tttggagtta 6231 ttgcaaaaac attagctgct aatgaacgga agtatagaga aattataaaa aaggagttgg ccagataaat 6301 gaatatatta aacaccataa aagaaatttt attatctgat gcagagctcc aaacatatat aaattctaga 6371 atatactatt ataaagtcac tgaaaatgct gaaacttcca aaccttttgt tgttattaca cctatttatg 6441 atttaccttc agacttcatg tctgataaat atcttagtga agaatactta attcaaatag atgtagaatc 6511 ttcaaataat cagaaaacaa ttgatataac aaaacgaata agatatctgt tatatcaaca aaatttaatt 6581 caagcatcta gtcagttaga tgcttatttt gaagaaacta aacgttatgt gatgtcgaga cgttatcaag 6651 gcataccaaa aaatatatat tataaaaatc agcgcatcga ataggtgtgc tttttaattt ttaaggagga 6721 aataagcaat ggcagaagga caaggttctt ataaagtagg ttttaaaaga ttatacgttg gagtttttaa 6791 cccagaagca acaaaagtag ttaaacgcat gacatgggaa gatgaaaaag gtggtacagt tgatctaaat 6861 atcacaggtt tagcaccaga tttagtagat atgtttgcat ctaacaaacg tgtttggatg aaaaaacaag 6931 gtactaatga agttaagtct gacatgagta tttttaatat tccaagtgaa gatctaaata cagttattgg 7001 tcgttctaaa gataaaaatg gtacatcttg ggtaggagag aatacaagag caccatacgt aacagttatt 7071 ggagaatctg aagatggttt aacaggtcaa ccagtgtacg ttgcgctact taaaggtact tttagcttgg 7141 attcaattga atttaaaaca cgaggagaaa aagcagaagc accagagcca acaaaattaa ctggtgactg 7211 gatgaacaga aaagttgatg ttgatggtac tccacaaggt attgtatacg ggtatcatga aggtaaagaa 7281 ggagaagcag aattcttcaa aaaagtattc gttggataca cggacagtga agatcattca gaggattctg 7351 caagttcgtt acccagctaa cccccaaaat gttgaagtag cagttaattc aaaatctgca acagtttcag 7421 cagaataggg gctttcaaaa taaatcaaag gagaataatt tatgactaaa actttaaagg tttataaagg 7491 agacgacgtc gtagcttctg aacaaggtga aggcaaagtg tcagtaactt tatctaattt agaagcggat 7561 acaacttatc caaaaggtac ttaccaagtg gcatgggaag aaaatggtaa agaatctagt aaagttgatg 7631 tacctcaatt caaaaccaat ccaattctag tctcaggcgt atcatttaca cccgaaacta aatcaatcac 7701 ggtaaatgct gatgacaatg ttgaaccaaa cattgcacca agtacagcaa cgaataaaac gttgaaatat 7771 acaagtgaac atccagagtt tgttactgtt gatgagagaa caggagcaat tcacggtgta gctgagggaa 7841 cttcagttat cactgctacg tctactgacg gaagtgacaa gtctggacaa attacagtaa cagtaacaaa 7911 tggataatta tttgagacgc agaatatctg cgtctttttt atttgaataa aaggagctaa tacaatgatt 7981 aaatttgaaa ttaaagaccg taaaacagga aaaacagaga gctatacaaa agaagatgtg acaatgggcg 8051 aagcagaaaa atgctatgag tatttagaat tagtaaatca agagaataaa aaagaagtac ctaacgcaac 8121 aaaaatgaga caaaaagagc gacagttatt agtagattta tttaaagatg aaggattgac tgaagaagat 8191 gttttgaaca agatgagcac taaaacttat acaaaagcct tgaaagatat atttcgagaa atcaatggtg 8261 aagatgaaga agattcagaa actgaaccag aagagatggg aaagacagaa gaacaatctc aataaaagat 8331 attttatcga acattaagaa aatacaacgt ttctgtatgg agcagtatgg gtggacatta actgaagtca 8401 gaaaacagcc gtatgtaaaa cttttagaaa tacttaatga agagaataaa gaagagactg aagaaaaaca 8471 aagtgaacaa aaagtcatta caggtacgga tttaagaaaa ctttttggaa gctagaaagg aggttaatat 8541 gaatgaaaaa gtagaaggca tgaccttgga gctgaaatta gaccatttag gtgtccaaga aggcatgaag 8611 ggtttaaagc gacaattagg tgttgttaat agtgaaatga aagctaatct gtcatcattt gataagtctg 8681 aaaaatcaat ggaaaagtat caggcgagaa ttaaggggtt aaatgataag cttaaagttc aaaaaaagat 8751 gtattctcaa gtagaagatg agcttaaaca agttaacgct aattatcaaa aagctaaatc tagtgtaaaa 8821 gatgttgaga aagcatattt aaagctagta gaagctaata aaaaagaaaa attagctctt gataaatcta 8891 aagaagcctt aaaatcttcg aatacagaac ttaaaaaagc tgaaaatcaa tataaacgta caaatcaacg 8961 taaacaagat gcatatcaaa aacttaaaca gttgagagat gcagaacaaa agcttaagaa tagtaaccaa 9031 gctactactg cacaactaaa aagagcaagt gacgcagtac agaagcagtc cgctaagcat aaagcacttg 9101 ttgaacaata taaacaagaa ggcaatcaag ttcaaaaact aaaagtacaa aatgataatc tttcaaaatc 9171 aaacgaaaaa atagaaaatt cttacgctaa aactaatact aaattaaagc aaacagaaaa agaatttaat 9241 gatttaaata atactattaa gaatcatagc gctaatgtcg caaaagctga aacagctgtt aacaaagaaa 9311 aagctgcttt aaataattta gagcgttcaa tagataaagc ttcatccgaa atgaagactt ttaacaaaga 9381 acaaatgata gctcaaagtc atttcggcaa acttgctagt caagcggatg tcatgtcaaa gaaatttagt 9451 tctattggag ataaaatgac ttccctagga cgtacgatga cgatgggcgt atctacaccg attactttag 9521 ggttaggtgc agcattaaaa acaagtgcag acttcgaagg gcaaatgtct cgagttggag cgattgcaea 9591 agcaagcagt aaagacttaa aaagcatgtc taatcaagcg gttgacttag gcgctaaaac aagtaaaagt 9661 gctaacgaag ttgctaaagg tatggaagaa ttggcagctt taggctttaa tgccaaacaa acaatggagg 9731 ctatgccggg tgttatcagt gcagcagaag caagcggtgc agaaatggct acaactgcaa ctgtaatggc 9801 atcagcaatt aattctttcg gtttaaaagc atctgatgca aaccatgttg ctgatttact tgcgagatca 9871 gctaatgata gtgctgcaga tattcaatac atgggagatg cattaaaata tgcaggtact ccagcaaaag 9941 cattaggagt ttcaatagag gacacttctg cagcaattga agttttatct aactcagggt tagaggggtc 10011 tcaagcaggt actgcattaa gagcttcgtt tattaggcta gctaatccaa gtaaaagtac agctaaggaa
10081 atgaaaaaat taggtattca tttgtctgat gctaaaggtc aatttgttgg catgggtgaa ttgattagac 10151 agttccaaga caacatgaaa ggcatgacga gagaacaaaa actagcaaca gtggctacaa tagttggcac 10221 tgaagcagca agtggatttt tagccttgat tgaagcgggt ccagataaaa ttaatagcta tagcaaatca 10291 ttgaagaact ctaatggtga aagtaaaaaa gcagctgatt tgatgaaaga caacctcaaa ggtgctctgg 10361 aacaattagg tggcgctttt gaatcgttag caattgaagt tggtaaagat ttaacgccta tgattagagc 10431 aggtgcggaa ggattaacaa aattagttga tggatttaca catcttcctg gttggtttag aaaggcttcg 10501 gtaggtttag cgatttttgg tgcatctatt ggccctgctg ttcttgctgg tggcttatta atacgtgcag 10571 ttggaagcgc ggctaaaggc tatgcatcat taaatagacg cattgctgaa aatacaatac tgtctaatac 10641 caattcaaaa gcaatgaaat ctttaggtct tcaaacctta tttcttggtt ctacaacagg aaaaacgtca 10711 aaaggcttta aaggattagc cggagctatg ttgtttaatt taaaacctat aaatgttttg aaaaattctg 10781 caaagctagc aattttaccg ttcaaacttt tgaaaaacgg tttaggatta gccgcaaaat ccttatttgc 10851 agtaagtgga ggcgcaagat ttgctggtgt agccttaaag tttttaacag gacctatagg tgctacaata 10921 actgctatta caattgcata taaagttttt aaaaccgcat atgatcgtgt ggaatggttc agaaacggta 10991 ttaacggttt aggagaaact ataaagtttt ttggtggcaa aattattggc ggtgctgtta ggaagctagg 11061 agagtttaaa aattatcttg gaagtatagg caaaagcttc aaagaaaagt tttcaaagga tatgaaagat 11131 ggttataaat ctttgagtga cgatgacctt ctgaaagtag gagtcaacaa gtttaaagga tttatgcaaa 11201 ccatgggcac agcttctaaa aaagcatctg atactgtaaa agtgttgggg aaaggtgttt caaaagaaac 11271 agaaaaagct ttagaaaaat acgtacacta ttctgaagag aacaacagaa catggaaaa agtacgttta 11341 aactcgggtc aaataacaga agacaaagca aaaaaacttt tgaaaattga agcggattta tctaataacc 11411 ttatagctga aatagaaaaa agaaataaaa aggaactcga aaaaactcaa gaacttattg ataagtatag 11481 tgcgttcgat gaacaagaaa agcaaaacat tttaactaga actaaagaaa aaaatgactt gcgaattaaa 11551 aaagagcaag aactcaatca gaaaatcaaa gaattgaaag aaaaagcttt aagtgatggt cagatttcag 11621 aaaatgaaag aaaagaaatt gaaaagcttg aaaatcaaag acgtgacatc actgttaaag aattgagtaa 11691 gactgaaaaa gagcaagagc gtattttagt aagaatgcaa agaaacagaa atgcttattc aatagacgaa 11761 gcgagcaaag caattaaaga agcagaaaaa gcaagaaaag caagaaaaaa agaagtggac aagcaatatg 11831 aagatgatgt cattgctata aaaaataacg tcaacctttc taagtctgaa aaagataaat tattagctat 11901 tgctgatcaa agacataagg atgaagtaag aaaggcaaaa tctaaaaaag atgctgtagt agacgttgtt 11971 aaaaagcaaa ataaagatat tgataaagag atggatttat ccagtggtcg tgtatataaa aatactgaaa 12041 agtggtggaa tggccttaaa agttggtggt ctaacttcag agaagaccaa aagaagaaaa gtgataagta 12111 cgctaaagaa caagaagaaa cagctcgtag aaacagagaa aat taaaga aatggtttgg aaatgcttgg 12181 gacggcgtaa aaactaaaac tggcgaagct tttagtaaaa tgggcagaaa tgctaatcat tttggcggcg 12251 aaatgaaaaa aatgtggagt ggaatcaaag gaattccaag caaattaagt tcaggttgga gctcagccaa 12321 aagttctgta ggatatcaca ctaaggctat agctaatagt actggtaaat ggtttggaaa agcttggcaa 12391 tctgttaaat cgactacagg aagtatttac aatcaaacta agcaaaagta ttcagatgcc tcagataaag 12461 cttgggcgca ttcaaaatct atttggaaag ggacatcaaa atggtttagc aatgcatata aaagtgcaaa 12531 gggctggcta acggatatgg ctaataaatc gcgctcgaaa tgggataata tttctagtac agcatggtcg 12601 aatgcaaaat ccgtttggaa aggaacatcg aaatggttta gtaactcata caaatcttta aaaggttgga 12671 ctggagatat gtattcaaga gcccacgatc gttttgatgc aatttcaagt tcggcatggt ctaacgctaa 12741 atcagtattt aatggtttta gaaaatggct atcaagaaca tatgaatgga ttagagatat tggtaaagac 12811 atgggaagag ctgcggctga tttaggtaaa aatgttgcta ataaagctat tggcggttta aatagcatga 12881 ttggcggtat taataaaata tctaaagcca ttactgataa aaatctcatc aagccaatac ctacattgtc 12951 tactggtact ttagcaggaa agggtgtagc taccgataat tcgggagcat taacgcaacc gacatttgct 13021 gtattaaatg atagaggttc tggaaacgcc ccaggtggtg gagttcaaga agtaattcac agggctgacg 13091 gaacattcca tgcaccccaa ggacgagatg tggttgttcc actaggagtt ggagatagtg taataaatgc 13161 caatgacact ctgaagttac agcggatggg tgttttgcca aaattccatg gtggtacgaa aaagaaagat 13231 tggctagacc aacttaaagg taatataggt aaaaaagcag gagaatttgg agctacagct aaaaacacag 13301 cgcataatat caaaaaaggt gcagaagaaa tggttgaagc agcaggcgat aaaatcaaag atggtgcatc 13371 ttggttaggc gataaaatcg gcgatgtgtg ggattacgta caacatccag ggaaactagt aaataaagta 13441 atgtcaggtt taaatattaa ttttggaggc ggactaacgc tacagtaaaa attgctaaag gcgcgtactc 13511 attgctcaaa aagaaattaa tagacaaagt aaaatcgtgg tttgaagatt ttggtggtgg aggcgatgga 13581 agctatctat ttgaatatcc aatctggcaa agatttggac gctacacagg tggacttaac tttaatgacg 13651 gtcgtcacta tggtatagac tttggtatgc ctactggaac aaacgtttat gccgttaaag gtggtatagc 13721 agataaggta tggactgatt acggtggcgg taattctata caaattaaga ccggtgctaa cgaatggaac 13791 tggtatatgc atttatctaa gcaattagca agacaaggcc aacgtattaa agctggtcaa ctgataggga 13861 aatcaggtgc tacaggtaat ttcgttagag gagcacactt acatttccaa ttgatgcaag ggtcacatcc 13931 agggaatgat acagctaaag atccagaaaa atggttgaag tcacttaaag gtagtggcgt tcgaag ggt 14001 tcaggtgtta ataaggctgc atctgcttgg gcaggcgata tacgtcgtgc agcaaaacga atgggtgtta 14071 atgttacttc gggtgatgta ggaaatatca ttagcttgat tcaacacgaa tcaggaggaa atgcaggtat 14141 aactcaatct agttcgctta gagacatcaa cgttttacag ggcaatccag caaaaggatt gcttcaatat 14211 atcccacaaa catttagaca ttatgctgtt agaggtcaca acaatatata tagtggttac gatcagttat 14281 tagcgttctt taacaacaga tattggcgct cacagtttaa cccaagaggt ggttggtctc caagtggtcc 14351 aagaagatat gcgaatggtg gtttgattac aaagcatcaa cttgctgaag tgggtgaagg agataaacag 14421 gagatggtta tccctttaac tagacgtaaa cgagcaattc aattaactga acaggttatg cgcatcatcg 14491 gtatggatgg caagccaaat aacatcactg taaataatga tacttctaca gttgaaaaat tgttgaaaca 14561 aattgttatg ttaagtgata aaggaaataa attaacagat gcattgattc aaactgtttc ttctcaggat 14631 aataacttag gttctaatga tgcaattaga ggtttagaaa aaatattgtc aaaacaaagt gggcatagag 14701 caaatgcaaa taattatatg ggaggtttga ctaattaatg caatcttttg taaaaatcat agatggttac 14771 aaggaagaag taataacaga ttttaatcag cttatatttt tagatgcaag ggctgaaagt ccaaacacca 14841 atgataacag tgtaactatt aacggagtag atggtatttt accgggcgca attagttttg cgcctttttc 14911 attagtatta aggtttggct atgatggtat agatgttata gatttaaatt tatttgagca ttggtttaga 14981 tctgtgttta atcgcagaca tccttattat gttattactt ctcaaatgcc tggtgttaaa tatgcagtga 15051 atacagctaa tgttacatct aatttaaaag atggttcttc aactgaaatt gaagtaagtt taaatgttta 15121 taaagggtat tctgaatcag ttaattggac cgatagcgag ttcttattcg actctaattg gatgtttgaa 15191 aatggaattc ctcttgattt cacacctaaa tatactcata catcaaatca atttactatt tggaacggtt 15261 ctactgatac gataaatcca cgattcaagc acgatttgaa aatattaatt aatttaaatg cgagtggagg 15331 atttgaactg gttaactata caacaggtga tatttttaag tacaacaaaa gtatagataa aaacactgat
15401 tttgttttag atggtgtgta tgcatatcga gatataaata gagtgggaat tgatacaaat agaggcatta 15471 taacattagc gccaggtaaa aatgaattta agattaaagg agacatcagt gatattaaaa ctacatttaa 15541 gtttcctttt atttataggt aggtgattta atggattatc atgatcattt atcagtaatg gattttaatg 15611 aattgatttg tgaaaattta ctagatgtag attatggttc ttttaaagaa tattatgaac tgaatgaagc 15681 taggtacatc acttttacag tttatagaac tactcataat agttttgttt tcgatttact aatttgtgaa 15751 aacttcataa tttatcatgg tgaaaaatac acaattaagc agacagcgcc aaaggttgaa ggtgataaag 15821 tttttattga agttacggca tatcacataa tgtatgaatt tcaaaatcac tcagtggaat caaataagct 15891 tgatgacgac agtagcgaaa ctggtaaaac gccagaatac tctttagatg agtacttaag atatggattt 15961 gcaaatcaaa aaacttcggt caaaatgacc tataaaataa ttggaaattt taagcgaaaa gtaccgattg 16031 acgaattagg taacaaaaac ggcttagaat actgtaaaga agcggtagac ctatttggct gtataattta 16101 cccaaatgat acggagatat gtttttattc tcctgaaaca ttttatcaaa gaagcgagaa agtgattcga 16171 tatcaatata atactgatac tgtatctgca actgtcagta cattggaatt aagaacagct ataaaagttt 16241 ttggaaaaaa gtatacagct gaggaaaaga aaaattataa tcctattaga acaactgaca ttaaatattc 16311 aaatggtttt ataaaagaag gtacttatcg taccgcaaca attgggtcta aagctactat taactttgat 16381 tgcaagtatg gtaatgaaac agttagattt acaataaaaa agggctctca aggtggaata tataagttga 16451 ttttagacgg caagcaaatt aagcaaattt cttgttttgc taagtcggtt cagtctgaaa caatagattt 16521 aataaaaaat attgataaag gcaagcacgt tttagaaatg atatttttag gagaagaccc caaaaataga 16591 attgatatat cttcaaataa aaaagctaag ccttgtatgt atgttggaac tgaaaaatca acagtcttaa 16661 atttaattgc tgacaactca ggtcgcaatc aatacaaagc aattgttgac tacgtcgcag atagtgcaaa 16731 gcagtttggg attcgatatg ctaatacgca aacaaatgaa gatatcgaaa cacaggataa gctgttagaa 16801 tttgcaaaaa agcaaataaa tgatactcct aagactgaat tagatgttaa ttatataggt tatgaaaaaa 16871 tagagccaag agatagcgta ttctttgttc atgaattaat gggatataac actgaattaa aggttgttaa 16941 acttgatagg tcacatccat ttgtaaacgc aatagatgaa gtgtctttca gcaatgaaat aaaggatatg 17011 gtacaaattc aacaagcgct taacagacga gttattgcac aagataatag atataactat caagcaaatc 17081 gtataaatca tttatacact agtactttga attctccttt cgagacaatg gatataggga gtgtattaat 17151 ataatggcaa cagaagaagt taaaatcaaa gcgctacttg aaaacgataa acagtacttt ccagctacac 17221 attggaaagc tataaatggg ataccttatg caggcagtag tgatattgat ggattgcctc aagacggtat 17291 catttcggta gatgataaaa ataaattaga taatttaaaa ataggcgaag caggaattat tcaaaatagc 17361 attgtacaga aatccccaaa cggtaaattg tggaaaataa cagttgacga tagtgggaaa cttggtacag 17431 tgctatttta ttagaaagga aggtgcatta tggaaaattt gtatttaata aaggatttgg gagctttagc 17501 aggtcgagat tatagagcta aggaaataca aaacttacaa agaatagagc aatttgcgct tggcttgaca 17571 acagagttta agttgcatca gaaagctaaa acaattcaac acttcgctga gcaaatttat tataatggta 17641 gatcgcaagc agcagtaaac aaatctttac aaagtcaaat taacgcactt gttgtggcac cacgtaataa 17711 cagtgctaat gagattgttc aagctcgagt taatgtaaac ggcgaaacct ttgacacatt aaaagaacat 17781 ttagacgatt gggaaaccca aactcaaatt aataaagagg aaactataag agaattaaat aagaccaaac 17851 aagaaattct tgatatcgag tatcgttttg aacctgataa gcaagaattt ttatttgtga cagaacttgc 17921 acctcttaca aatgcagtaa tgcaatcctt ctggtttgat aatagaacag gcatagtata catgacacaa 17991 gctagaaata atggctatat gctaagtcgt ctaagaccta atggtcaatt tatagacagc tcattgattg 18061 taggtggggg tcatggtaca cataacggtt atagatatat tgatgatgag ttatggattt atagttttat 18131 cttaaatggt aataatgaga atacattagt tcgtttcaag tatacgccta atgtggaaat tagctatggc 18201 aagtatggta tgcaagatgt atttacagga cacccagaaa aaccctacat cacccctgtc ataaatgaaa 18271 aagaaaataa aattctatac agaattgaga gacctagaag tcactgggaa cttgaaaact caatgaatta 18341 tatagagata agaagtttag acgatgttga taaaaatatt gataaagttt tgcataaaat cagtatccct 18411 atgagactaa caaacgaaac ccaaccaatg cagggtgtga cttttgatga aaaatacttg tattggtata 18481 caggagacag taatccaaat aatagaaact atttaacggc tttcgattta gaaacaggag aagaagcgta 18551 tcaggttaat gctgactatg gtggaacact agattcattt cctggcgaat ttgcggaagc agaaggtttg 18621 caaatatact atgacaaaga tagtggtaaa aaagctttga tgctaggtgt tactgtcggt ggtgatggaa 18691 atagaacaca tcgtattttc atgattgggc aaagaggtat tttagaaata cttcactcaa gaggcgttcc 18761 ttttatcatg agtgacacag gtggtagagt taaaccttta ccaatgaggc ctgataaact taagaatctt 18831 gggatgttaa cagagccagg tctttactat ttatacactg atcatacagt tcaaatcgat gatttcccat 18901 taccaagaga atggcgtgat gcaggttggt tc tggaagt taagccacca caaactggcg gtgatgtaat 18971 tcagatattg acgcgtaata gttatgcaag gaatatgatg acttttgaaa gggtgctttc tggaagaact 19041 ggagacattt cggactggaa ttatgtgcct aaaaatagtg gtaaatggga gagagtacct tcattcatca 19111 caaaaatgtc agatattaac atagtaggca tgtcgtttta tttaactacg gatgatacaa aacgttttac 19181 agattttcca actgaacgta aaggggtagc tggttggaac ttatatgtag aagcttcaaa cacaggtggc 19251 tttgttcata ggctagttcg taatagtgtt acagcatctg ctgagatact attgaaaaat tatgatagta 19321 aaacaagttc agggccatgg actttacacg aagggagaat tataagttaa tgagtaattt agagaaatct 19391 gtagctataa atttagaaaa cacagcgcat tatgaaaata tttcaaatct agatataact tttagaacag 19461 gagagagtga ttcttctgtt cttcttttta atatcactaa aaataatcaa ccgttattat tgagtgaaga 19531 aaatatcaaa gcacgaatag cgattcgagg taaaggagtc atggtagttg ctccactaga aatattagat 19601 ccatttaaag gtattttaaa atttcaatta cctaatgatg taattaaacg agatggaagt tatcaagctc 19671 aagtttcggt tgcagaatta ggtaattcag acgtggtagt tgtcgagaga actatcacat ttaacgttga 19741 aaaaagtttg tttagcatga ttccatctga aacaaaatta cactatattg ttgaatttca ggaattagaa 19811 aaaactatta tggatcgtgc gaaagcaatg gacgaggcta taaaaaatgg tgaagattat gcgagtctga 19881 ttgaaaaagc taaagaaaaa ggtctatcag atattcaaat agcaaaatct tcaagtatag atgaattaaa 19951 gcaacttgct aatagccata tatctgattt ggaaaataaa gcgcaagcat attcaagaac attcgatgag 20021 caaaagcgat atatggatga gaaacatgaa gccttcaagc agtcagtgaa tagtggtggt ttagtcacaa 20091 gtggttctac ttcaaattgg caaaaagcta agattactaa agatgatggt aagataatgc agattactgg 20161 atttgatttt aataatccag aacaaagaat aggtgattca acccaattta tttatgtttc gcaagctata 20231 aattatccaa gaggtgttag tactaacggt actgtcgaat atttagtagt aacttcagat tacaagcgta - 20301 tgacttatcg accgaacggt acaaataaag tgtttgttaa aagaaaagaa gcgggttcat ggtctgagtg 20371 gtcagaatta gctattaatg attacaatac accttttgaa actgttcaaa gtgcccaatc aaaagctaat 20441 atggccgaaa gtaacgctaa attatacgca gatgacaagt ttaataaaag gtattcggtt atttttgatg 20511 gaacagcaaa tggtgtgggc tctacattgt acttaaatga gagtttagac caatttattt tattaatttt 20581 ttatgggact tttccaggtg gtgactttac agagtttggc agtccttttg gaggaggaaa gatttcattg 20651 aatccctcaa atcttccaga tggtgatgga aatggtggag gtgtttatga gtttggatta actaaatcta
20721 gtcgtacatc tttaactata tcaaacgatg tctatttcga cttaggaagt caaagaggct ctggtgcgaa 20791 cgcaaataga gggacaatta acaaaattat aggagtgaga aaataatgca aatattagtt aacaagcgta 20861 atgagataat ttcatacgct atcattggtg gctttgaaga aggtattgat attgaaaatt taccagaaaa 20931 tttctctcaa gtttttagac ctaaagcctt taaatattca aatggggaaa tagtttttaa cgaagattat 21001 tcagaagaaa aagatgactt gcatcaacag attgacagtg aagaacaaaa cacagtcgct tctgatgaca 21071 tcttacgaaa aatggttgct agtatgcaga aacaagttgt tcaaagtaca aagttatcga tgcaagttaa 21141 taagcaaaat gcactaatgg caaaacaact tgtgacactt aataaaaaat tagaagaggt taaaggagag 21211 actgaaaatg cttaaattaa tttcaccaac attcgaagat attaaaacat ggtatcaatt gaaagaatat 21281 agtaaagaag atatagcgtg gtatgtagat atggaagtta tagataaaga ggaatatgca attattacag 21351 gagaaaagta tccagaaaat ctagagtcat aggttataat cttatggctt tttaatttga ataaagtggg 21421 tggtgtaatg tttggattta ccaaacgaca cgaacaagat tggcgtttaa cgcgattaga agaaaatgat 21491 aagactatgt ttgaaaaatt cgacagaata gaagacagtc tgagaacgca agaaaaaatt tatgacaagt 21561 tagatagaaa tttcgaagaa ctaaggcgtg acaaagaaga agatgaaaaa aataaagaga aaaatgctaa 21631 aaatattaga gacatcaaga tgtggattct aggattaata gggacgattc taagtacatt tgttatagcc 21701 ttgttaaaaa ctatttttgg catttaaagg aggtgattac catgcttaag ggaattttag gatatagctt 21771 ttggtcgtgt ttctggttta gtaagtgtaa gtaatagtta agagtcagtg cttcggcact ggctttttat 21841 tttggaaaaa aggagcaaac aaatggatgc aaaagtaata acaagataca tcgtattgat cttagcatta 21911 gtaaatcaat tcttagcgaa caaaggtatt agcccgattc cagtagacga tgagaatata tcatcaataa 21981 tacttactgt tgttgcttta tatactacgt ataaagacaa tccaacatct caagaaggta aatgggcaaa 22051 tcaaaagcta aagaaatata aagctgaaaa caagtataga aaagcaacag ggcaagcgcc aattaaagaa 22121 gtaatgacac ctacgaatat gaacgacaca aatgatttag ggtaggtgtt gaccaatgtt gataacaaaa 22191 aaccaagcag aaaaatggtt tgataattca ttagggaagc agttcaatcc tgatttgttt tatggatttc 22261 agtgttacga ttacgcaaat atgtttttta tgatagcaac aggcgaaagg ttacaaggtt tatacgctta 22331 taatattcca tttgataata aagcaaggat tgaaaaatac gggcaaataa ttaaaaacta tgatagcttt 22401 ttaccgcaaa agttggacat tgtcgttttc ccgtcaaagt atggtggcgg agctggacat gttgaaattg 22471 ttgagagcgc taatctaaac actttcacat cgtttggcca aaattggaat ggtaaaggtt ggacaaatgg 22541 cgttgcgcaa cctggttggg gtcccgaaac cgttacaaga catgttcatt attacgatga cccaatgtat 22611 tttattagat taaatttccc agataaagta agtgttggag ataaagctaa aagcgttatt aagcaagcaa 22681 ctgccaaaaa gcaagcagta attaaaccta aaaaaatt t gcttgtagcc ggtcatggtt ataacgatcc 22751 tggagcagta ggaaacggaa caaacgaacg cgattttata cgtaaatata taacgccaaa tatcgctaag 22821 tatttaagac atgccggtca tgaagtcgca ttatatggtg gctcaagtca atcacaagac atgtatcaag 22891 atacagcata cggtgttaat gtaggtaata aaaaagatta tggcttatat tgggttaaat cacaggggta 22961 tgacattgtt ctagaaatac atttagacgc agcaggagaa agcgcaagtg gtgggcatgt tattatctca 23031 agtcaattca atgcagatac tattgataaa agtatacaag atgttattaa aaataactta ggacaaataa 23101 gaggtgtaac acctcgtaac gatttactaa atgttaacgt atcagcagaa ataaatataa attatcgctt 23171 atctgaatta ggttttatca ctaataaaaa tgatatggat tggattaaga aaaactatga cttgtattct 23241 aaattaatag ccggtgcgat tcatggtaag cctatcggtg gtgtgatatc tagtgaggtt aaaacaccag 23311 ttaaaaacga aaagaatccg ccagtgccag caggttatac acccgataaa aataatgtac cgtataaaaa 23381 agaaactggt tattacacag ttgccaatgt taaaggtaat aacgtaaggg acggctattc aactaattca 23451 agaattactg gtgtattacc taataacgca acaatcaaat atgacggcgc atattgtatc aatggctata 23521 gatggattac ttatattgct aatagtggac aacgtcgtta tattgctaca ggagaggtag acaaggcagg 23591 taatagaata agcagttttg gtaagtttag tgcagtttga taattgtata tgatgaatct taggcaggta 23661 cttcggtact tgcctattat ttaaaattaa taaacagtta atttttacat gaatatatta aattttaaaa 23731 aaacaaacgt ttttagtata taaattattt tgtgttcgta ttgtgtgcta tgattaaaaa gttgttatgg 23801 tcaactatat cgtggtttta tgtttattat caatcaaaat ataaattatt tataatttgt ttggtaatga 23871 acgggttttt ttcgaaataa tagtaaaaaa acacatttgt agatatttta aactcggtaa atcttttaat 23941 aaatatttaa ttttattaaa agttaaaaag gtttaatata aaaatgtaat aaaatttata aagaaaggaa 24011 atgattttta tggtcaaaaa aagactatta gctgcaacat tgtcgttagg aataatcact cctattgcta 24081 cttcgtttca tgaatctaaa gctgataaca atattgagaa tattggtgat ggcgctgagg tagtcaaaag 24151 aacagaagat acaagtagcg ataagtgggg ggtcacacaa aatattcagt ttgattttgt taaagataaa 24221 aagtataaca aagacgcttt gattttaaaa atgcaaggtt ttatcaattc aaagactact tattacaatt 24291 acaaaaacac agatcatata aaagcaatga ggtggccttt ccaatacaat attggtctca aaacaaatga 24361 ccccaatgta gatttaataa attatctacc taaaaataaa atagattcag taaatgttag tcaaacatta 24431 ggttataaca taggtggtaa ttttaatagt ggtccatcaa caggaggtaa tggttcattt aattattcaa 24501 aaacaattag ttataataaa ataaaaagta ggtgataaga tgactcaatt tctaggggcg cttcttctta 24571 caggagtttt aggttacata ccatataaat atctaacaat gataggttta gttagtgaaa aaaacaaggt 24641 tatcaatact cctgtattat tgattttttc tattgaaaca tgtttgatat ggttttatag ttttataatt 24711 tttaataatg ttgatttaaa aaatttgaat ttaattcagt tgcttacagg tctaaaagca aatattttgt 24781 ttctatttat ttttgtttta acagtgtttg tatttaatcc tttaattgtt aaatttatta tctggttaat 24851 taatataacc agaaagttta tgaaattgga ttgtataagc ttattagaca aaagagacaa gttgtttaat 24921 aacaacggta aaccagtatt tatagttata aaagactttg aaaacagaat cattgaagag ggtgaactta 24991 aaacctataa ttcagctggt agcgatttcg atttactaga agttgagcga caagatttca aagtatctga 25061 tttaccgtca aacgatgaat tgtatattaa acatacactt gtagacctta aacaacaaat taaattggat 25131 ttatatttaa tgaatgaata ctaatctttt ttcttagctt tttctgataa agtgcttttt aatttttcgc 25201 tggcgcccgg cttttcaaaa cttttgttta ttgggttact acgagtagct tcttgttttt tgtttttatc 25271 cgccataaaa ttctcaccac cattcaacgt ctacacttgt aggcgttttt ttatttagta aagtcataat 25341 gaatcttctt tggttaactt atctccatct attttttgtg aaataaattc caagtattta cgcgcattat 25411 gtgacgataa atctttaggt aactcataag tgaatggttg attaccacta gttaaaactt catatactat 25481 agtttctttt tttattttgc aattagttat tttcattata aacttccttt caaacactgc tgaaatagac 25551 gtcttttata ttaaagcgcc acacaggcgc tgttaatcac aatacaactt tgcccattac tttaatatta, 25621 ctaaacgaag cgactttgat atcatcatac ttcggattta gagataccaa attaa at g tcttcgcata 25691 tatctacacg cttgataaga cttactccat ctaatacaac gagtgcaatt gtaccatctt taatagaatc 25761 ttctttctta ataaaagcgt atgttccttg ttttaacata ggttccattg aatcaccatt aactaaaata 25831 caaaaatcag catttgatgg cgtttcgtct tctttaaaaa atacttcttc atgcaatatg tcatcatata 25901 attcttctcc tatgccagca ccagttgcac cacatgcaat atacgatact agtttagact ctttatatcc 25971 atctatagaa gtgactttat tctgttcttc caattgttca tttgcatagt taagtacgtt ttcttggcgg
26041 ggaggtgtga gtttgttgta tatggaagtg atgtcgttat cgtctttgta tgtagtattt gattcactat 26111 acaaatcatt aatcttcaca ttgaagtact cagccaaaat tttggcagtt gataatcgag gttcttcctt 26181 ttcattttcc cattttgata tcttgccttt cgttaatttc attaagtcgg gatatttatt attaagatca 26251 gttgctaatt gttccatagt catattttta tttttttctt agcttcttta aaccttcacc aatacccata 26321 cgaaaccctc cttatataag ataatttcat tataaaagtt tcgaaaacga aacgcaagga aaatattatt 26391 gcaaaagttg ttgacatcga aacttttatg atgtattctt aaatcaagtt gttacaaacg aaacaaaagg 26461 agggggttca atgacaacta gtgtagcaga taaaccatac ttaaaaataa aaagcttgat tgcacttaaa 26531 ggaactaacc aaaaagaagt tgctaaagca atcggaatga gtagaagttt attgagtata aagataaatc 26601 gaattaatgg cagagatttt acaacttcag aagctaaaaa attagcagat catttaaatg ttaaagttga 26671 tgattttttt taaactttaa gtttcgaaag tgacaactaa ataaaaataa ggaggacact atggaacaaa 26741 taacgttaac caaagaagag ttgaaagaaa ttatagcgaa agaagttaga aatgctataa aaggcgagaa 26811 accaatcagc tcaggtgcaa ttttcagtaa agtaagaatc aataatgacg atttagaaga aatcaataaa 26881 aaactcaatt tcgcaaaaga tttgtcgcta ggaagattga ggaagctcaa tcatccgatt ccgctaaaaa 26951 agtatcagca tggcttcgaa tcaattcatc aaaaagctta tgtacaagat gttcatgacc at ttagaaa 27021 attaacatta tcaatttttg gagtgacact taattcagac ttgagtgaaa gtgaatacaa cctagcagca 27091 aaaatttata gagatatcaa aaactattat ttatatatct atgaaaagag agtttcagaa ttaactatcg 27161 atgatttcga atgaaggagg aactacaaat gaaactacta agaaggctat tcaataaaaa acacgaaaac 27231 ttaattgacg tgtggcatgg aaatcaatgg ttaaaagtga aagaaagcaa attaaaaaaa tataaagtgg 27301 tctcggatag agaaggtaag aaatatctaa ttaaataagc gcacttaatt agtgcaag a atcaagtgcg 27371 ctattgcctt acaatcctaa atcttttctg cttttttctt cttcttgtaa tcccaataac acagaagagt 27441 aaatgctgaa atagtcacga gcaacgctat ctttagcgaa tgcaattacg tcatcaccga cttcttgcca 27511 ttcgttatga atcttatgtc tatctagagc tctaggtaat agcgagattg taatatcgtg agcaattttc 27581 tctaaatcca taaatttcac ctccttccac tgggagataa ctaaattata taacaaaaca acttaaagga 27651 ggaacgacaa atgcaagctc aaaacaaaaa agtcatctat tactactatg acgaagaagg taataggcga 27721 ccattagata ttcaaattaa tgacggatat gaactgatgg tccgatctca tttcatcaac aacaccattg 27791 aagaaatacc atacgtaaat aataacttat atgccttggt tgatggttat gaatttaagt tagattgaat 27861 ttttgagaaa gatattgaaa agctaatttc cccataagat taagagacat actggatgtt ttgttaacga 27931 ctcttttaac ttcgttccaa gttttattgt ctctaatatt atcgagaaat tcatggccag accaagtgat 28001 gtcatcaata atccaagaaa cgaccctgcc ttcgatgaat ttcagatcgc aacaaataaa tttagcttct 28071 tctaatttta aaagtgagta cattactgtt tcaaaatcat atttatcaaa aataatatta tcgttgaaat 28141 tatgtcgagt aagtggttca cctattttct tattagattc tatttctaag agcaagagtc taacgcaatc 28211 gtgattaagt ttcatcctat cacctccata acaggagtat agcagaaagg atcataaaca tcttaaaagg 28281 aggaataaca aatgaacatt caagaagcaa ctaagatagc tacaaaaaat cttgtctcta tgacacggaa 28351 agattggaaa gaaagtcatc gaactaagat attaccaaca aatgatagtt ttttacaatg catcatttca 28421 aatagcgatg ggacaaacct tatcagatat tggcaacctt cagccgatga cctcatggca aatgattggg 28491 aagttataaa cccaactaga gaccaggaat tattgaagca attttagaaa tgctatcaat gatacttttt 28561 aaattgtttt taaactcatt ttcaaagtaa acaacagtct tgtctgaaat tgttacatga taaatagtgt 28631 tactagcata cacgcσgttt aggaacccag agtttttaag tttatttaaa tcgtatttta catcttcgaa 28701 atgtagtttt tgaaaatact ttgtatgtat atctttagca cttccaaaat tattgcaggt taatttaacc 28771 gaacctaact ttacacattc taaataatct ttgtagagta cggacaagat atattgttgg tctttagtaa 28841 gtgtatcaaa ttcatcagat atcaagggca tgttatcacc tccttaggtt gataacaaca ttatacacga 28911 aaggagcata aacaaatgaa cacaagatca gaaggattgc gtataggcgt cccacaagtt tctagcaaag 28981 ctgatgcttc ttcatcctat ttaacggaaa aggaacgtaa cttaggagcg gaaa attag agcttattaa 29051 aaaaagtgat tacagctact tagaaataaa caaagttttc tatgcattag atagagaact tcaatacagg 29121 gcgaataata acaaacttta acatttatct aaaggagtga tagagatgcc aaaaatcata ataccaccaa 29191 caccagaaaa cacatatcga ggcgaagaaa aatttgtgaa aaagt a ac gcaacaccta cacaaatcca 29261 tcaattgttt ggagtatgta gaagtacagt atacaactgg ttgaaatatt accgtgaaga taatttaggt 29331 gtagaaaatt tatacattga ttattcagca acgggaacat tgattaatat ttctaaatta gaagagtatt 29401 tgatcagaaa gcataaaaaa tggtattagg aggattatca aatgagcgac acatataaaa gctacctatt 29471 agcagtgttg tgcttcacgg tcttagcgat tgtactcatg ccgtttctat acttcactac agcatggtca 29541 attgcgggat tcgcaagtat cgcaacattc atattttata aggaatactt ttatgaagaa taaagaaact 29611 gctacttgtt ggagcaagta acagtgcaag atgagcaatt gtcttaaata attatataag gagttattaa 29681 tatgacctta caacaaaaaa tactatcaca ttttgcaaca tatgacaatt tcaattctga tgatgttgtt 29751 gaagtttttg ggatatctaa aacacatgca aaatccacac tttcaagact taagaaaaaa ggaaagattg 29821 aattggaaag ttggggtatc tggcgtgttg ttgaaccgca gttacattta actgttgtag aacgtaagaa 29891 agagatatta gaagaacaat tcgagttatt ggcaagatta aacgaacaaa gtgatgaccc tagagaaata 29961 gaagaacgca tcaagttaat gattcgttta gccaaccaat tttaaggagg agttaatcaa tggcaatatt 30031 agaaggtatt tttgaagaat taaaactatt aaataagaat ttacgtgtgc taaatactga actatcaact 30101 gtagattcat caattgtaca agagaaagtt aaagaagcac caatgccaaa agatgaaaca gctcaactgg 30171 aatcagttga agaagttaag gaaacttctg ctgatttaac taaaga tat gttttatcag taggaaaaga 30241 gttccttaaa aaagcagata cttctgataa gaaagaattt agaaataaac ttaacgaact tggtgcggat 30311 aagctatcta ctatcaaaga agagcattat gaaaaaattg ttgattttat gaatgcgaga ataaatgcat 30381 gaagctagat cactcaaata gagctcatgc aaagcttagt gcaagtggag caaaacaatg gctaaactgt 30451 ccaccgagta ttaaggcaag tgaaggtatt gcagataaaa gttcagtttt tgctgaagaa ggtacattcg 30521 ctcatgagtt aagtgagtta tatttcagtc ttaaatatga aggcctaaca cagtttgagt ttaataaagc 30591 ttttcaaaat tataagcgaa atcaatatta cagtgaagag ttgcgcgaat atgttgaaga gtacgtagct 30661 aatgtagaag aaaaatataa cgaagctttg agtagagatg acgatgtaat agctttattt gaaacaaaat 30731 tggatttagg taaatacgtc cctgaatctt ttggtactgg tgatgtcatt atattttcag gtggtgtact 30801 tgaaattatt gaccttaaat acggtaaagg cattgaagtt tcagctatag ataatcctca acttagatta 30871 tatggcttgg gcgcatatga actgcttagt ttaatgtatg acattcatac agttcgcatg actatcatac __ 30941 aaccacgaat agataacttt tctactgaag agttaccaat atcaagatta cttcaatggg gaaccgattt 31011 tgttaaacca ttagccagac ttgcttataa cggtgaaggt gagtttaaag caggtagtca ttgtagattc 31081 tgtaagataa agcattcatg tagaacacgt gcagaataca tgcaaaatgt gcctcaaaag ccaccacatt 31151 tgttgagtga tgaagagatt gcagaacttt tatataaact gcctgacatc aaaaaatggg ctgatgaagt 31221 agaaaaa at gcactagatc aagcgaaaga aaatgataaa aactattctg gttggaagct tgtagaaggt 31291 cgctcgcgaa gaatgataac tgatacaaat gcaacgcttg aaaagttagt tgaagcaggt tataaacctg
31361 aagatattac agaaaccaag ttacttagca ttacgaattt agaaaaatta atcggcaaaa aagcattttc
31431 taaaattgca gaaggcttta tagaaaagcc acaaggtaaa ttaacacttg ctaccgagtc tgataaacga
31501 ccagctataa agcaatctgc tgaagatgat tttgacaaac tataaaaatt aaaaaggacg gtatataaac
31571 atgaaagcaa aagtattaaa taaaactaaa gtgattacag gaaaagtaag agcatcatat gcacatattt
31641 ttgaacctca cagtatgcaa gaagggcaag aagcaaagta ttcaatcagt ttaatcattc ctaaatcaga
31711 tacaagtacg ataaaagcca ttgaacaagc tatagaagct gctaaagaag aaggaaaagt tagtaagttt
31781 ggaggcaaag ttcctgcaaa tctgaaactt ccattacgtg atggagatac tgaaagagaa gatgatgtga
31851 attatcaaga cgcttatttt attaacgcat caagcaaaca agcacctggt attattgacc aaaacaaaat
31921 tagattaacg gattctggaa ctattgtaag tggtgactat attagagctt caatcaattt atttccattc
31991 aacacaaatg gtaataaggg tatcgcagtt ggattgaaca acattcaact tgtagaaaaa ggcgaacctc
32061 ttggcggtgc aagtgcagca gaagatgatt tcgatgaatt agacactgat gatgaggatt tcttataagt
32131 caataggtgg ggtttttagc cccactttaa ttttaaagaa attgaggtgt caagaatttg aaatttatga
32201 atatagatat tgaaacatat agcagtaacg atatttcgaa atgtggtgtc tataaataca cagaagctga
32271 agatttcgaa atcttaatta tagcttattc aatagatggt ggaccgatta gtgcgattga catgactaaa
32341 gtagataatg agcctttcca cgctgattat gagacgttta aaattgctct atttgaccct gctgtaaaaa
32411 agtatgcatt caatgctaat ttcgaaagaa cttgtcttgc taaacatttt aataaacaga tgccacctga
32481 agaatggatt tgcacaatgg ttaattcaat gcgtattggc ttacctgctt cgcttgataa agttggagaa
32551 gttttaagac tacaaaacca aaaagataaa gcaggtaaaa atttaattcg ttatttctct ataccttgta
32621 agccaacaaa agttaatgga ggaagaacaa gaaatttgcc tgaacatgat cttgaaaaat ggcaacaatt
32691 tatagattac tgtattcgag atgtagaagt agaaatgaca attgctaata aaattaaaga ctttccagta
32761 actgtaattg aacaagcata ttgggttttt gaccaacata taaacgacag aggtattaag ctttctaaat
32831 cattgatgtt aggagctaat gtgctcgata agcagagtaa agaagaattg cttaaacaag ctaaacatat
32901 aacaggttta gaaaatccta atagtcctac acagttattg gcttggttaa aggatgaaca aggattagat
32971 atacctaatt tacaaaagaa aacggttcag gattacttaa aagtagcaac aggaaaagct aaaaaaatgc
33041 tagaaattag attgcaaatg tctaaaacca gtgtgaaaaa atacaacaaa atgcatgaca tgatgtgcag
33111 tgatgaacgg gtaagaggtc tgtttcaatt ctacggtgcc ggtactggaa gatgggcagg tagaggtgta
33181 caacttcaga atttaacaaa gcattatatt tcagatactg aattagaaat agcaagagat cttattaaag
33251 aacaacgttt tgacgattta gatttattac tcaatgttca tcctcaagac ttattaagtc aattagttag
33321 gacgacattt actgctgaag aaggtaatga actagcagta agtgattttt ctgcaataga ggcaagagtc
33391 atagcatggt atgcaaaaga acaatggcgt ttagatgtgt tcaacacaca cggaaagata tatgaagcat
33461 cggcttctca aatgtttaat gtaccggtag aaagcataac taaaggcgac cctctcagac aaaaaggaaa
33531 agtgtccgaa ttagctttag gctatcaagg tggcgctgga gctttaaaag caatgggtgc attggaaatg
33601 ggcattgaag aaaacgagtt acaaggttta gttgatagtt ggcgtaacgc aaatcctaac atagttaatt
33671 tttggaaggc ttgccaagag gctgcaatta atactgtaaa atcccgaaag acgcatcata cacatggact
33741 tagattttat atgaaaaaag gttttctaat gattgaactg cctagtggaa gagctttagc ttatccaaaa
33811 gctttagttg gtgaaaatag ttggggtagt caagttgttg aatttatggg gttagatctt aaccgtaaat
33881 ggtcaaagtt aaaaacgtat ggtgggaagt tagtcgagaa tattgttcaa gcaactgcaa gggatttact
33951 tgcgatttct atagcaaggc ttgaagcatt aggttttaaa atagttggcc atgtccatga tgaagtaatt
34021 gtagaaatac ctagaggttc aaatggactt aaggaaatcg aaactatcat gaataagcct gttgattggg
34091 caaaaggatt gaatttgaat agtgacgggt ttacttctcc gttttatatg aaggattagg agtgtgattg
34161 catgcaacat caagcttata tcaatgcttc tgttgacatt agaattccta cagaagtcga aagtgttaat
34231 tacaatcaga ttgataaaga aaaagaaaat ttggcggact atttatttaa taatccaggt gaactattaa
34301 aatataacgt tataaatatt aaggttttag atttagaggt ggaatgatgg ctagaagaaa agttataaga
34371 gtgcgtatca aaggaaaact aatgacattg agagaagttt cagaaaaata tcacatatct ccagaacttc
34441 ttagatatag atacaaacat aaaatgcgcg gcgatgaatt attgtgtgga agaaaagact caaaatctaa
34511 agatgaagtt gaatatatgc agagtcaaat aaaagatgaa gaaaaagaga gagaaaaaat cagaaaaaaa
34581 gcgattttga acctatacca acgaaatgtg agagcggaat atgaagaaga aagaaagaga agattgagac
34651 catggcttta tgatggaacg ccacaaaaac attcacgtga tccgtactgg ttcgatgtca cttataacca
34721 aatgttcaag aaatggagtg aagcataatg agcgtaatca gtaacagaaa agtagatatg aacgaagcgc
34791 aagacaatgt taagcaacca gcgcactaca catacggcga cattgaaatt atagatttta tcgaacaggt
34861 tacggcacag tatccacctc aactagcatt cgcaataggt aatgcaataa aatacttgtc tagagcacct
34931 ttaaagaatg gtcatgagga tttagcaaag gcgaagtttt acgtccaaag agcttttgac ttgtgggagt
35001 gatgaccatg acagatagcg catgtaaaga atacttaaac caatttttcg gatctaagag atatctgtat
35071 caggataacg aacgagtggc acatatccat gtagtgaatg gcacttatta ctttcacggg catatcgtac
35141 caggctggca aggcgtgaaa aagacatttg atacagcgga agagctcgaa acatatataa agcaacatgg
35211 tttggaatac gaggaacaga agcaactaac tttattttaa ggagatagaa atgatgaaaa tcaaagttga
35281 aaaaataatg aaaatagacg aattaattaa gtgggcgcga gaaaatccgg agctatcatt tggcagaaaa
35351 tattatacaa cagacaaaaa tgatgaaaac tttatttact tcggtgtttt taaaaattgt tttaaaataa
35421 gcgattttat attagttaat gctactttta gtgtcaaagt tgaagaagaa gtaaccgaag aaactaagtt
35491 tgataggttg tttgaagtgt acgagattca agaaggagtc tataaatctg catcatatga gaatgctagt
35561 ataaacgaac gtttaaaaaa tgacagaatt tttcttgcta aagcattcta catcttaaac gacgacctaa
35631 ctatgacgtt aatttggaaa gaaggagagt tgattaaata atggaacacg gttcaaaaga atattacgaa
35701 aagcaaagtg aatactggtt tgatgaagca agcaagtttt tgaagcaacg tgatgagctt attggagata
35771 tagctaagtt aagagagtgc aacaaagagc tggagaagaa agcaagtgca tgggataggt attgcaagag
35841 cgttgaaaaa gatttaataa acgaatttgg caaagatggt gaaagagtta aatttggaat ggaattaaac
35911 aataaaattt ttatggagga agacgcaaat gaataaccgc gaacaaatcg aacaatcagt tattagtgct
35981 agcgcgtata acggcaatga cacagaggga ttattaaaag agattgagga cgtgtataag aaagcgcaag
36051 cgtttgatga aatacttgag ggtttaccta atgctatgca agatgcaatc aaagaagata ttggtcttga
36121 tgaagcagta ggaattatga cgggtcaagt tgtctataaa tatgaggagg agcaggaaaa tgactaacat
36191 attacaagtg aaactattat caaaagacgc tagaatgcca gaacgaaatc ataagacgga tgcaggttat
36261 gacatatttt cagctaaaac tgtcgtactt gagccacaag aaaaggcagt gatcaaaaca gatgtagctg
36331 taagcattcc agagggctat gtcggtttat taactagccg tagtggtgta agtagtaaaa cgcatttagt
36401 gattgaaaca ggcaagatag acgcgggata tcatggtaat ttagggatta atatcaagaa tgataatgaa
36471 acgttagaga gtgaggatat gagtaacttt ggtcggagtc cttctggtat agatggaaaa tacaccctac
36541 tacctgtaac agataaattt ttatgtatga atggtagtta tgtcataaat aaaggcgaca aactagctca
36611 attggttatc gtgcctatat ggacacctga actaaagcaa gtggaggaat tcgagagtgt ttcagaacgt
36681 ggagcaaaag gcttcggaag tagcggagtg taaagacata ttagatcgag tcaaggaggt tttggggaag 36751 tgagtgacat gttagaaata tttttcatag ggtttggtgt ttatctattt tgtcgcatag gtattatttt 36821 tctcaagagt aaaaagacta tacacacaaa cctatatgaa atgttgttga ttgctactat ctttgtgaca 36891 tctacatttg ctgataaaca tcaaaagacg catatcttaa tagcattttt agtaatgttt tttatgagta 36961 agctcaaaca agttcaaggg agctatgagg aatgacacaa tacctagtca caacatttaa agattcaaca 37031 ggacgtaagc atacacacat aactaaagct aagagcaatc aaaggtttac agttgttgat gcggagagta 37101 aagaagaagc gaaagagaag tacgaggcac aagttaaaag aaatgcagtt at aaattag ggcagttgtt 37171 tgaaaatata agggagtgtg ggaaatgact aaacaaatac taagattatt attcttacta gcgatgtatg 37241 agctaggcaa gtatgtaact gagcaagtat atattatgat gacggctaat gatgatgcag aggcgccgag 37311 tgactttgaa aaaatcagag ctgaagtttc atggtaatag ctattatcat ttttgaatta attatattaa 37381 tgtgtttagc aatagcactg gaggtgttgt aaatatgtgg attgtcattt caattgtttt atctatattt 37451 ttattgatct tgttaagtag catttctcat aagatgaaaa ccatagaagc attggagtat atgaatgctt 37521 atcttttcaa gcagttagta aaaaataatg gtgttgaagg tatagaagat tatgaaaatg aagttgaacg 37591 aattagaaaa agatttaaaa gctaaagaga ggcgttggct tctctgttct atttaaaat atgaaaggag 37661 ccgaacatgt tagacaaagt cactcaaata gaaacaatta aatatgatcg tgatgtttca tattcttatg 37731 ctgctagtcg tttatctaca cattggacta atcacaatat ggcttggtct gactttatgc agaagctagc 37801 acaaacagtt agaactaaag aagatttaac tgagtacaat aaaatgtcta agtctgaaca agccgatata 37871 aaagatgttg gcggatttgt cggtggttat ttaaaagaag gcaaacgacg tgctggtcaa gtcatgaatc 37941 gttcaatgtt aacacttgat atcgattatg ctgctcaaga tatgactgac atattatcta tgttttatga 38011 ttttgcatat tgtttatatt caacacataa gcatagagag ataagtccaa gactgcgttt agtgattcct 38081 ttaaaacgaa atgtaaatgc agatgagtat gaagctattg ggcgtaaagt cgcagatatc gttggcatgg 38151 attacttcga tgatacaact tatcaaccac ataggttaat gtattggcct tcaactagta acgatgcgga 38221 atttttcttt acctatgaag atttaccttt gttagaccca gataaaatat taaatgaata tgttgattgg 38291 actgacacat tagaatggcc aacgtcttca agggaagaga gtaagactaa aagattagca gataagcaag 38361 gcgacccaga agaaaagccg ggaattgttg gtgcattttg tagagcctat acgatagaag aagctataga 38431 aacttttatt cctgatttat acgaaaaaca ttctactaac cgttatacct atcatgaagg ttcaactgca 38501 ggtggattgg tgttatacga aaataacaag tttgcctatt ctcatcataa tacggatccc gtaagcggta 38571 tgcttgtgaa cagttttgat ttagtacgca tacacttata tggtgctcaa gatgaagacg ctaaaacaga 38641 tactccggtt aatcgactac ctagttataa agcaatgcag caaagagcgc aaaatgatga agttgttaaa 38711 aagcaattaa ttaacgacaa aatgtctgat gcaatgcagg atttcgatga aatagtaaat agcgatgatg 38781 catggtctga gacgttagaa attacttcga aaggtacttt caaagctagt atcccaaata tagaaattat 38851 attgcgtaat gatccaaatt taaaaggaaa aatagcattt aatgaattta caaaacaaat tgaatgctta 38921 gggaaaatgc catggaataa taattttaaa atacgtcaat ggcaagacgg tgatgatagc agtttaagaa 38991 gttatatcga aaagatttat gacatacacc attcaggcaa aacaaaagat gccattataa gcgtagcaat 39061 gcaaaatgcc tatcatccag taagagatta tctaaataaa atatcgtggg atggacataa acgtcttgaa 39131 aagttattta tcaaatactt aggtgttgaa gacactgaag tgaatagaac aactaccaaa aaggcattga 39201 ctgctggaat cgctcgagta atggagccag gatgtaaatt tgactatatg cttacacttt atggtcctca 39271 aggtgtaggt aaatctgctt tgctaaaaaa aataggtggt gcatggtttt ctgacagttt agtttctgtt 39341 actggtaagg aagcatatga ggcattacaa ggcgtttggt taatggaaat ggcagaactt gcagctacaa 39411 gaaaagctga agttgaagct attaagcatt tcatatctaa acaagttgac cggtttcgtg ttgcttatgg 39481 acattatatt gaagattttc caaggcaatg tattttcatt ggtacaacta ataaagttga tttcttaaga 39551 gatgaaactg gtggaagacg tttttggcca atgactgtaa atccagagag agttgaagtg aactggtcta 39621 aactaaccaa agaagagatc gaccaaatct gggcagaagc taaatactat tatgaacaag gagaagagtt 39691 gttccttaac cctgaactag aagaagaaat gcgttcaatc caaagtaaac atactgagga atctccatat 39761 acaggtatta ttgatgaata tcttaacacg ccaatcccaa gcaattggga agacttaact atctttgaaa 39831 gaagacgatt ttatcaaggt gatgttgata tgttaccaac aggaaatgta gattacattg aaagagacaa 39901 ggtctgtgcg cttgaagtgt ttgttgaatg ttttggtaaa gataagggag atagtagagg atctatggaa 39971 attagaaaga tttctaacgt cttaagacaa ttagacaatt ggtctgtata tgaaggcaat aaaagtggga 40041 aaattcgatt tggaaaagat tatggtgtac agatagcgta tgtaagagat gaaagtttag aggatttaat 40111 ataagaaata ttgaataaat atacattttt agatgttgta tcaaatgttg catcattttt tgagtgatgc 40181 aacacggtgg tgtaaaaagt aatcgtaggt gttgtatcat ttttggtgat gcaacattga tgcaacaaat 40251 gatacaacac ctctttccct tctcgctgta aggttcaacc ctgtttgttt ccaatgttgc atcaaattca 40321 ctataaagtt taaaaagtag tgttagggag taaaggggta taggggtaac cctctaacag ctatttttaa 40391 aagtttggca agaattgatg caacatcgga acacaaatat aaattttgta tacaaggtga ataaatgaaa 40461 gaatcgacat tagaaaaata tttagtgaaa gagataacaa agttaaatgg attatgttta aaatgggtcg 40531 cacctggaac aagaggtgta ccagatagaa ttattattat gccagaagga aaaacatatt ttgtagaaat 40601 gaagcaagaa aagggaaagt tacatccttt acaaaaatat gtgcatcggc aatttgaaaa cagagatcat 40671 acagtgtatg tgttatggaa taaagaacaa gtaaatactt ttataagaat ggtaggtgga acatttggcg 40741 attgatttca aaccacatag ctatcaaaag tatgcaatag ataaagtgat tgataatgag aaatacggtt 40811 tgtttttaga tatggggcta gggaaaacag tatcaacact tacagcattt agtgaattgc agttgttaga 40881 cactaaaaaa atgttagtca tagcacctaa acaagttgct aaagatacat gggttgatga agttgataag 40951 tggaaccatt taaatcatct gaaagtgtct ttagtcttag gaacacctaa agaaagaaat gatgcattaa 41021 acacagaggc tgatatctat gtaaccaata aagaaaatac taaatggtta tgtgatcaat ataaaaaaga 41091 atggccattt gacatggttg taattgatga actgtctaca tttaaaagtc ctaagagtca aaggtttaaa 41161 tctattaaaa agaaattacc actcattaat agatttatag gattaacagg aacacctagt ccaaatagtt 41231 tacaggattt atgggctcaa gtttatttga tagacagagg cgaaagactt gagtcttcat tcagtcgtta 41301 tcgagaaagg tactttaaac caacacatca agttagcgaa catgttttta actgggagct aagagacgga 41371 tctgaagaaa agatatatga acgaatagaa gatatatgtt taagcatgaa agcgaaagat tatctggata 41441 tgcctgacag agttgatact aaacaaacag tagtcttatc tgaaaaagaa agaaaagtat atgaagaatt 41511 agaaaaaaac tatattttag aatcggaaga agaaggaaca gttgtagctc agaatggggc atcattaagt 41581 caaaaactac ttcaactatc taacggtgca gtttatacag atgatgaaga tgtaagactt atacatgata^ 41651 agaagttaga taagttagag gaaattatag aggagtctca aggccaacca atattattgt tttataactt 41721 caaacatgat aaagaaagaa tacttcaaag gtttaaggaa gcaaccacat tagaggattc aaactataaa 41791 gaacgttgga atagtggaga cattaagctg cttatagcac atccagcaag tgcagggcat ggattaaact 41861 tacaacaagg tgggcacatt attgtttggt ttggacttac atggtcattg gaattatacc aacaagcaaa 41931 tgcaagatta tatagacaag gacaaaatca tacgactatt attcatcaca tcatgaccga taacacaata
1 S3
42001 gatcaaagag tatataaagc tttacaaaat aaagaactaa cgcaagaaga attgatgaaa gctattaaag
42071 caagaatagc taagcataag taatggaggt ataagatggg aaaggcgtca tatgatatta agccaggaac
42141 atttaaatat attgaatcag aaatatataa tttaaatgag aacaagaaag agataaatag attgagaatg
42211 gagatactta acccaacgaa agaactagac accaacattg tgtatggacc gttacaaaaa ggagagccag
42281 ttagaacaac tgagttaatg gcgacaaggt tattgactaa taagatgtta cgtaacttag aagagatggt
42351 tgaagcagtt gaaagtgagt acttaaagtt acctgaagat cataagaaag taataaggtt aaagtattgg
42421 aataaagata agaagctaaa gatagaacaa ataggggatg cttgtcacat gcatcgcaat acagttacta
42491 caatacgaaa gaactttgtt aaagcgatag cgtatcatgc aggtatcaaa taacattgtg caaagattgt
42561 gcaaaaggcc tacaaatctg tagtaatatg atagtatcgg aaagatgtat aaagttatct gaaagttata
42631 cgacataaat acatgaggca catcgctaag cggtgtgtct tttgttatgc aatcaaagag gtgtaagaga
42701 tgaccaagca taataacatt tataagcatg gtcgtaagtc atatcaatac gattggttct atcattcaaa
42771 agcatggaag aagttaagag agatagcatt agatagagat aattatcttt gtcaaatgtg tttacgcgaa
42841 gatattataa cagatgcaaa gattgtgcat cacattattt atgttgatga agattttaac aaagctttag
42911 acttagataa tctaatgtca gtttgttata gctgtcataa caaaattcat gcaaatgata atgacaaaag
42981 taatcttaag aaaattagag ttctaaaaat ttaaataaaa aaattattta aataaaattt tatgcccccc
43051 tgcccatcgg cttaaaatgt tttttcgccg ggtaccggag aggcc
Table 8
Bacteriophage 3A ORFs list
Table 9
Bacteriophage 96, complete genome sequence
1 catagttata ggettttcag ctatataeca agataagatt tatcccgccg tctceataaa aatatgettg
71 gaaaccttga tttaatgggg ttttaateta geaagtgtca aatatgtgtc aagaaaataa ttttetgaca
141 cgttgacctt gctctttttt atgttcatca agtaagtgag agtaggtgtc taaagttata gatatattat
211 aatggectaa tcttttgcta atatattcaa taggtatacc tttagaaagt aggaaagatg tatgcgtgtg
281 tcttaatgaa taaggtgtta ttgtagtatc atttagtcet atttgactct tagcatggtt aaatgacttt
351 ttaacggcat tatgactcaa tttaaacaac ttattatetg tacgttttgg taattttgat aatttagctt
421 taatatgttg tatatccttt tttggtacct ccacaagtct gtccgcgtta actgtttttg ttccacgaag
491 atgtattgta ccctcttttt cgtttagatc gataggcaac atattaatta catcgctgta tcttgcacca
561 gtgatagcta ggatgaataa aaaaatataa ctcgattcgt ctctagattt aaagtattct atcaattgca
631 agtattgttc tatggtgatg aatttagagt gttcgtcttt tgattttttt gtaccacgaa tatctatttg
701 atagctaggg tctttcttta aatagccctc atatactgca tctctgaagc attgtgataa acaactgttt
771 aatttacgaa ccgtttcatt agtacgacct cgaccgaatt cgttcaaaaa cttttgatac tccgaacgtt
841 tgatgttttt tattaaaaaa tcactcccga aatattcgtt aaataatttt aatgaacgtt gataccaata
911 gaattgttgt gaagcgacat gtttcttatt ttttgaatct aaccaatcat tgtaatattc ttcaaacttt
981 ttattttcat ctaaattgtt tccatcatcc aaatctctaa gcagttgttg agcagcgttg gttgcctcag
1051 ctttagtttt gaatcctgac tttettttct ttcctgattt gaaagacgga tgttttacgt cgtactgcca
1121 agatgctgtt gctttattct tcctttttgt aattgtaaat gacgccattt tacttttcct cctcaaaatt
1191 ggcaaaaaat aataagggta ggcgagctac ccgaaatttt attgttgaac aactattgct tcacttcttg
1261 cttttcctac ttcttttcta aaactatcat atgattgatt agggtgtgtt aacgacattc ctggaccacc
1331 tceagcatgt tggtttttgt ccggattatt ttccatttct tcagtggctc ttttagcatt taaatattct
1401 tcgtaactag gttcgtttgg gtcgcgtggt tgtgcttgtt gtccattatt ggtagctgga agattcttct
1471 gtacctgttg cttagatgtg ttattggttt gttgattgtt gttaatgttt gtgttgttct cgttgtttac
1541 ttgattattg ttatcgtttt gattactatt ttcttttttc gcttctgctt tatetttagt ttctttcttt
1611 ttgtctttgt tctctttctt tgtttcggtt ttcttgcttt cctctttctt atcgccgtcg ttgetaccgc
1681 atgcacctaa cactaacgca ctagctaata ataaaactaa taatcttttc atgttttaca ctcctttatt
1751 tgctatttgt tttaataaat ctatgatttc attgttttgt tctatgattt tgttttcatt tttaagatgt
1821 tcgtctaaca tctctattaa gacgaaattt tgatttatca tttcgtaagt aaacatttga cctgtgttgt
1891 taggattaga aaacgaacta ctgaaacgcg ttgaaaagct atctataaat tgaccaactt tattttttaa
1961 taacatatct ttaccgctct cagacattgt atttagttcg cgcttattta aagttttttc tataattttg
2031 tattttgttt cctgatttct ttcgatttct tctacttcaa aagggatatt gttattaaat ttttcgataa
2101 tatcacgttt ttcagaaact gacatacgat caaatacttg tttttgacct ttatttaact tccctcgaat
2171 ttttccggca gtccaagact ctttaactgt taacttatca ttaggaactt gattcatctt ttatatgaet
2241 ccttttctca tatttcttta tatttaaaaa ctctcaacgg ctcaaatgta atcgaatact cgccatagtg
2311 agttccaata ccgtatatct tcttatattg ttctattgcc tccaatatgt attcttcgct taattgtaga
2381 tactcagaca actcatacaa gttacgtacg ccataattgt aagettctac aatttcgcgt aacgggactg
2451 ctgagataaa gccgtgtcgt cttgcgtaat tttcgaactt gcgattgttg aatttcgatt gatctaaaat
2521 gttgccatac gtcaacttgt ggtgggcaag ttcttcatat aatacttcta atttgttcct ttcggataag
2591 gaaggtctaa taaaaatttc tccttcttga taccaaccat cgaatcotcg aggtactctt tgtgtttctt
2661 tcacttcaac ttcacatttc ataagcaatt cttcgtattt tcceatgegc caaacccctt tggtgtctta
2731 tttctttcta tctctaaccc attgcataaa attttcgatt tcttcccatt ettcgggagt aaattcatct
2801 ttatttgcat gaccggct t agtttcttga tgaatacttc tttcttctgt aattctcgat ttaggtacat
2871 taaagtaatc tgctaattgt tggacttttg atattctagg atatttaagt tctttaagcc agttagagat
2941 tgttgattga cttaccccga ttgcttcaga caattctact tgagtaatgt tgttctcttt cataagttgt
3011 tctaagttct ctgataaaat ttttctagca ctcttatatt ccataatttt ctcctttagt attacttaat
3081 gtaatactaa tttaccataa gtaatatcac ttttcaatac aaaatattao ttttttgaaa taaatatcac
3151 tttaggtgtt gacatattac tttaagtgat agtatagttg taaatgtcaa cgggaggtga tacgaaatgc
3221 cagaaaattt taaagagttc tctgtaaagg tctggagaac taattcgaat atgacacaac aagatgtcgc
3291 tgataaatta ggcgttacta aacaatctgt aataagatgg gaaaaagatg acgcagaatt aaaaggctta
3361 caattgtatg ctttagccaa attattcaac acagaagttg attatataaa ggctaaaaaa atttaacatt
3431 aatatcactt taagtgataa aggaggaaac tgaaatgcaa gaattacaaa catttaattt tgaagaatta
3501 ccagtaagga aaattgaagt ggaaggagaa cccttctttt taggtaagga tgttgctgaa attttagggt
3571 atgcacgagc agataacgec atacgcaatc atgttgatag tgaagatagg ctgatgcacc aaattagtgc
3641 gtcaggtcaa aacagaaata tgatcatcat caacgaatct ggattataca gtttaatctt tgacgcttct
3711 aaacaaagta aaaacgaaaa cattagagaa accgctagga aattcaaacg ctgggtaact tcggaagttt
3781 taccgacgtt aagaaaaact ggtgcttacc aagtacctag tgacccaatg caagcattga gattaatgtt
3851 tgaagctaca gaagaaacaa aacaagaaat taaaaacgtg aaagatgatg ttattgattt gaaagaaaat
3921 caaaaactgg atgcgggaga ctacaatttc ttaactagaa caatcaatca aagagtagct catatacaaa
3991 gactacatgc gataacaaac caaaaacaac gtagcgaatt attcagggat attaattcag aagtgaaaaa
4061 gatgactggt gcgagttcaa gaacgaacgt aagacaaaaa ca tcgacg atgtaattga aatgattgct
4131 aattggttcc cgtcacaagc tactttatac agaatcaagc aaattgaaat gaaattttaa aacgaaatat
4201 aggagaggct gaatatggaa tacatoggat atgcagacgc aaatgcgttt gtaaaaataa gtggcatttc
4271 aaaagatgat ctagagaaaa aagtctactc gaacaaagag tttcaaaaag aatgcatgta cagatttggt
4341 cgaggacaaa agcgttatat aaaaattgac aaagctattc aatttatcgg taccaattta atgattaatg*
4411 aatacgaatt ataggaggag ttatcaaatg agtaaaactt ataaaagcta cctagtagca gtactatgct
4481 tcacagtctt agcgattgta cttatgccgt ttctatactt cactacagcg tggtcaattg caggattcgc
4551 aagtatcgca acattcatat actacaaaga atacttttat gaagaataaa aaaactgcta cttgcgtcaa
4621 caagtaacag tgacaaacat ttatcaaaat atacaactta attaaatcaa aatatacgga ggtagtcaac
4691 tatggctgaa aatattaaaa ctgaacaaca ttattacact aaagatttct caggatacag aaatgaagaa
4761 gataactttg tageaaatca agaattgaca gtaacaatea eattgaacga gtacagaaaa cttattgaaa 4831 taaaggctgt taaagataaa gaagaagata cctacagagg taagtatttt gcggaagaaa gaaaaaacga 4901 aaaattggaa aaagaaaata taaaactaaa aaacaaaatt tatgaattac aaaacgaaga agataacgag 4971 gaggacgaag aagaeaagga ggacgagaac gatgtattac aaaattggtg agataaaaaa caaaattata 5041 agctttaacg ggtttgaatt taaagtgtct gtgatgaaga gacatgacgg tatcagtata caaatcaagg 5111 atatgaataa tgttccactt aaatcgtttc atgtcataga tttaagcgaa ctatatattg cgacggatgc 5181 aatgcgtgac gttataaacg aatggattga aaataacaca gatgaacagg acaaactaat taacttagtc 5251 atgaaatggt aggaggtatg aaaagtgaat gatttacaag agagagaatt agaaacattc gaacaagacg 5321 accgattcaa agtaactgat ctagacagtg ctaactgggt ttttaagaaa ctggatgcaa tcacaactaa 5391 agagaatgaa atcaacgatt tagcaaataa agaaattgaa cgcataaacg aatggaaaga taaagaagta 5461 gaaaaattac agagtggcaa agaatattta caaagccttg taattgaata ttacagaata caaaaagaac 5531 aagatagcaa attcaagttg aatacacctt acggaaaagt gacagccaga aaaggttcaa aagtcattca 5601 agttagcaat gagcaagaag tcattaaaca acttgagcaa cgaggttttg acaactatgt aaaagtaact 5671 aaaaaactta gccaatcaga cattaagaaa gatttcaatg taactgaaaa cggcacattg attgacgcaa 5741 acggcgaagt tttagagggt gctagcattg tggagaaacc aacgtcatac acggtaaagg tgggagaata 5811 gatgactgaa aaaactaatc aagatgtcga tattttaacg caactaggtg taaaagacat cagcaaacaa 5881 aatgcaaaca agttttataa atttgcgata tacggcaagt tcggtactgg taaaactacg tttttaacaa 5951 aagataacaa taccttagta ctagatataa atgaggacgg aacaacggta acagaagatg gggcagttgt 6021 gcagattaag aattataagc attttagtgc agtgattaaa atgctgccta aaattattga acaactaaga 6091 gaaaacggaa aacaaattga tgttgtagtg attgaaacaa tccaaaagtt acgtgatatc actatggacg 6161 acatcatgga cggtaaatca aagaaaccga catttaatga ttggggcgag tgtgctacac gcattgtaag 6231 tatttatcgt tatatttcta aattacaaga acattatcaa tttcatcttg ctataagcgg acacgagggc 6301 attaacaaag acaaagatga tgagggaagt actatcaatc caacaatcac gatagaggca caagaccaaa 6371 taaaaaaagc agtcatcagt caatctgacg tgttagcaag aatgacaata gaagaacatg agcaagacgg 6441 cgaaaaaact tatcaatatg tacttaacgc tgaaccatca aatttattcg agacaaagat aagacactca 6511 agcaacatca aaattaacaa caaacgtttc attaatccaa gtattaacga tgttgtacaa gcaattagaa 6581 atggtaatta aaaattaatt aaaaggacgg tataaaaatt atgaaaatca ctggtagaac acaatacatt 6651 caagaaacta atcaagaggc attcatgaaa ggtggggact ttttaggagc tggagaattt acagtaaaag 6721 ttgcaaatgt cgagtttaac gacagagaaa acagatactt caegattgtt tttgaaaaca acgaaggtaa 6791 acaatacaaa cacaaccaat tcgtcccacc attccaacaa gattatcaag aaaaacaata tatcgagtta 6861 cttagtagat taggaattaa attgaactta ccagatttaa cttttgacac agatcaatta attaacaaaa 6931 tcggaactat tgtacttaaa aataaattta acgaggaaca aggcaagtat tttgtaagac tctcatatgt 7001 aaaagtttgg aataaagacg atgaagtagt taataaacca gaacctaaaa ctgatgagat gaaacaaaaa 7071 gaacagcaag caaatggtaa acagacacct atgagtcaac aatcaaaccc attcgctaat gctaatggtc 7141 caatagaaat caatgatgat gatttaccgt tctaggacgt ggtttaaatg caatacatta caagatacca 7211 gaaagacaat gacggtactt attccgtcgt tgctactggt gttgaacttg aacaaagtca cattgattta 7281 ctagaaaacg gatatccgct aaaagcagaa gtagaggttc cggacaataa aaaactatct atagaacaac 7351 gcaaaaaaat attcgcaatg tgtagagata tagaaettca ctggggcgaa ccagtagaat caactagaaa 7421 attattacaa acagaattgg aaattatgaa aggttatgaa gaaatcagtc tgcgtgactg ttcaatgaaa 7491 gttgcgagag agttaataga actgattata tcgtttatgt ttcatcatca aatacctatg agtgtagaaa 7561 cgagtaagtt gttaagcgaa gataaagcgt tattatattg ggctaeaatc aaccgcaact gtgtaatatg 7631 cggaaagcct cacgcagacc tggcacatta tgaagcagtc ggcagaggta tgaacagaaa caagatgaat 7701 cactacgaca aacatgtgtt agcactgtgt agacaacatc ataatgaaca gcacgcaatt ggtgttaagt 7771 cgtttgatga taaatatcaa ttgcatgact cgtggataaa agttgatgag aggctcaata aaatgttgaa 7841 aggagagaaa aatgaataag ttactaatag atgactatcc gatacaagta ttaccgaaat tagctgaatt 7911 aatagggtta aacgaagcaa tagtattgca acaaattcat tattggctaa acaactcaaa acataaatac 7981 gatggcaaaa cttggatttt taattcttat ccagaatggc aaaaacaatt tccattttgg agcgagagaa 8051 c ataaaaag gacatttggg agtttagaaa aacaaaattt attgeatgta ggtaactaca acaaggctgg 8121 atttgaccgt acaaaatggt attcaatcaa ttatgaaaca ttaaacaaac tagtggcacg accatcggga 8191 caaaatggcc cgacgatgag gacaaattgg cacgatgcaa gaggacaaaa tgacccgacc aataccatag 8261 actacacaga gactaacaaa catagagaga cagacgacgt ctcaaagtca tttaagtata ttagtaccaa 8331 tttagaaatt atacaaaacc ctttaaaagc agaacagtta gaacacgaaa ttaaatcatt taagcaagat 8401 cagttcgaaa tagtaaaagt cgctaccgat tactgcaaag aaaacaacaa aggtctgaat tacttactaa 8471 ctgtattaaa gaactggaat aaagaaggcg tttcagataa agaaagtgct gaaaacaaat tgaaacctcg 8541 taactctaaa aaagaaacta ctgatgatgt catagcacaa atggaaaaag aattgagtga tgactaatgc 8611 cgatgagcaa aacacaagca ttagaaatta ttaaaaaagt taggtacgta tacaacatcg attttgataa 8681 accaaagtta gaaatgtgga ttgatgtatt aagtcaaaac ggggattatc aaccaactgt aaaagctgta 8751 gatggatata tcaacagtaa caacccgtac ccgcctaacc taccagcaat catgcgtaag gcacctaaaa 8821 aagtatctat tgagccggta gacaacgaaa ccgctacaea ccaatggaaa atgcagaatg accccgaata 8891 tgtcagacaa agaaaaatag cgctagataa cttcatgaat aagttggcag aatttggggg cgataacgaa 8961 tgaattacgg tcaatttgaa attgaaagca caataatcgc tacgctactt aaacaaccgg acgtactaga 9031 aaagataaga gttaaagatt aeatgtttac gaacgaaaag tttaaaacet ttttcaatta tgtaatggac 9101 gtcggaaaga tagatcatca agaaatctat ttaaaagcaa ctaaagataa agagttttta gatgcagata 9171 ctataactaa actttacaac tccgatttca ttggatacgg attctttgaa cgttatcaac aagaattatt 9241 ggaaagttat caaatcaaca aagcgaaaga attggtaact gagttcaaac aacaacctac gaaccaaaat 9311 tttaataact tgattgatga actcaaggat ttaaaaacaa ttactaacag aaaagaagac ggaaccaaga 9381 agtttgttga ggagtttgtc gatgagttat acagcgatag ccctaagaag caaattaaga cgggttataa 9451 gctcatggat tacaaaatag ggggattgga gccgtcgcaa ttaatcgtca tcgcagcgcg tccctcagtg 9521 ggtaagacag gttttgcatt aaacatgatg ctgaacatag cacaaaatgg atacaaaaca tctttcttta 9591 gtctcgaaac aactggcaca tcagtattga aacgtatgtt atcaacaatt actggtattg agttaacaaa^ 9661 gataaaagaa atcaggaact taacgccgga tgacttaaca aagttaacga atgcgatgga taaaatcatg 9731 aaattaggca tcgatatttc tgataaaagt aatatcacac cgcaagatgt gcgagcgcaa gcaatgaggc 9801 attcagacag gcaacaagtt atttttatag attatcttca actgatggat actgatgcga aagttgatag 9871 acgtgtagca gtagaaaaga tatcacgtga cttaaagata atcgctaacg agacaggcgc aatcatcgta 9941 ctactttcac aactgaatcg tggtgtcgag tctagacagg ataaaagacc aatgctatcg gacatgaaag 10011 aatcaggcgg aatagaagea gatgegagtt tagegatget actttacegt gatgattatt ataacegtga
10081 cgaagatgac agtatcactg gcaaatctat tgttgaatgt aacatagcca aaaacaaaga cggcgaaacc
10151 ggaataattg aatttgagta ttacaagaag actcagaggt ttttcacatg aatataatge aatteaaaag
10221 cttattgaaa tcgatgtatg aagagacaaa gcaaagcgac ccgattgtag caaatgtata tatcgagact
10291 ggttgggcgg tcaatagatt gttggacaat aacgagttat cgcctttcga tgattacgac agagttgaaa
10361 agaaaatcat gaatgaaatc aactggaaga aaacacacat taaggagtgt taaaaaatgc cgaaagaaaa
10431 atattactta taccgagaag atggcacgga agatattaag gtcatcaagt ataaagacaa cgtaaatgaa
10501 gtttattcgc tcacaggagc ccatttcagc gacgaaaaga aaattatgac tgatagtgac ctaaaacgat
10571 ttaaaggegc tcacgggctt ctatatgagc aagagetagg attgeaagca acgatatttg atatttagag
10641 gtggcacaat gagtaaatac aatgctaaga aagttgagta caaaggaatt gtatttgata gcaaagtaga
10711 gtgcgaatat taccaatatt tagaaagtaa tatgaatggc actaactatg atcgtatcga aatacaaccg
10781 aaatttgaat tacaacctaa attcgggaaa caaagaccga ttacgtatat agccgatttc tctttgtgga
10851 aggaagggaa actggttgaa gttatagacg ttaaaggtaa ggcgactgaa gttgccaaca tcaaagcgaa
10921 gatattcaga tatcagtata gagatgtgaa tttaacgtgg atatgtaaag cgcctaaata cacaggtcaa
10991 gaatggatgg tatatgagga cttagtgaaa gtcagacgta aaagaaaaag agaaatgaag tgatctaatg
11061 caacaacaag catatataaa cgcaaeaatt gatataagaa tacctacaga agttgaatat cagcattacg
11131 atgatgtgga taaagaaaaa gatacgctgg caaagcgctt agatgacaat ccggacgaat tactaaagta
11201 tgacaacata acaataagac atgcatatat agaggtggaa taaatgaagt tgaacgaagt attcgcaact
11271 aatttaaggg taatcatggc tagagataac gtaagtgtcc aagatttgca caatgaaact ggcgtatcaa
11341 gatcaactat tagtggatat aaaaacggaa aagctgagat ggttaactta aatgtattag ataaattggc
11411 agatgctcta ggtgttaatg taagtgaact atttactaga aatcacaaca cgcacaaatt agaggattgg
11481 attaaaaaag taaatgtata gaggtggaat aaatgagtat cgtaaagatt aacggtaaac catataaatt
11551 taccgaacat gaaaatgaat tgataaaaaa gaacggttta actccaggaa tggttgcaaa aagagtacga
11621 ggtggctggg cgttgttaga agccttacat gcaccttatg gtatgcgctt agctgagtat aaagaaattg
11691 tgttatccaa aatcatggag cgagagagca aagagcgtga aatggttagg caacgacgta aagaggctga
11761 actacgtaag aagaagccac atttgtttaa tgtgcctcaa aaacattctc gtgatccgta ctggttcgat
11831 gtcacttata accaaatgtt caagaaatgg agtgaagcat aatgagcata atcagtaaca gaaaagtaga
11901 tatgaacaaa acgcaagaca atgttaaaca accggcgcat tacacatacg gcaacattga aattatagat
11971 tttatcgaac aggttacggc acagtatcca cctcaactag cattcgcaat aggtaatgca atcaaatact
12041 tgtctagagc accgttaaag aatggtcatg aggatttagc aaaggcgaag ttttacgtcc aaagagcttt
12111 tgacttgtgg gagggttaac gatggcaacg caaaaacaag ttgattacgt aatgtcatta caggaacaat
12181 tgggattaga agactgtgaa aaatatacag acgaacaagt taaagctatg agtcataaag aagttagcaa
12251 tgtgattgaa aactataaga caagcatatg ggatgaagag ctatataacg aatgcatgtc gtttggtctg
12321 cctaattgtt aaaaggagtg atgaccatga acgatagcgc acgcaaagaa tacttaaacc aatttttcag
12391 ctctaagaga tatctgtatc aagacaacga gcgagtggca catatccatg tagtaaatgg cacttattac
12461 tttcacggac attataaaac gatgtttaaa ggcgtgaaaa agacatttga tactgctgaa gagctcgaaa
12531 tatatataaa gcaacatgat ttggaatatg aggaacagaa gcaaccaact ttattttaga ggagatggaa
12601 ataatggcaa agattaaaag aaaaaagaag atgacgctac tcgaactggt ggaatgggca tggaacaatc
12671 ctgaacaagt tgaaagtaaa gtgtttcaat cagatagaat gggcacgctt ggagaatgta gcgaagtaca
12741 tttttcaact gatgggcatg ggttttatac aaaagtagta acagataaag atatttttac tgtagaaatc
12811 acagaggaag tcactgaaga tactgagttt gattgtctag tagaactaaa cgatattgaa ggttttgaaa
12881 tatatgaaaa tgattcaatc agagagttga tagacggtac ttccagagcg ttttatatac taaacgaaga
12951 taaaactatg acattaattt ggaaagatgg ggagttggta gtatgatgca aacctataaa gtatgtcttt
13021 gtatcaagtt ctttgcatct aaatgtgatt ataaattaaa gaaacattat ttcgtgaaaa gtacgaatga
13091 ggaaaaagcc acgaacatgg tattaaaact gattcgtaaa aagctcccgt tcgaaactgc aagcatagaa
13161 gtcgaaaaag tggaggcaat ataatgatac aaccaacaag agaagaatta attaatttca tgaaaaaaca
13231 tggagctgaa aatgttgact ctatcactga tgagcaaagt gcaataagac actttagagc tcaatcaaaa
13301 gtttttaaag acgaacgtga tgagtacaag aagcaacgag atgagcttat cgaggatata gctaagttaa
13371 gaaaacgtaa cgaagagctg gagaacatgt ggcgcacagt caaaaatgaa ttgcttggaa gatacgaaca
13441 ttactgtttt aaaattagag aactacaccc tgagagcaaa gcgaacagga taggagctct ctatatagga
13511 ggtaaaagca ctgcagatat tatactgtcg cgaatggaag aactagacgg aacaaatgag ttctacgaat
13581 ttttagggca aatggaggca gacacaaatg aataaccgtg aacaaataga acaatcagtg atcagtacta
13651 gtgcgtataa cggtaatgac acagaggggt tactaaaaga gattgaggac gtgtataaga aagcgcaagc
13721 gtttgatgaa atacttgagg gaatgacaaa tgctattcaa cattcagtta aagaaggtat tgaacttgat
13791 gaagcagtag gggttatggc aggtcaagtt gtctataaat atgaggagga gcaggaaaat gagtattagt
13861 gtaggagata aagtatataa ccatgaaaca aacgaaagtc tagagattgt gcaattggtc ggagatatta
13531 gagatacaca ttataaactg tctgatgatt cagttattag cattatagat tttattacta aaccaattta
14001 tctaattaag ggggacgagt gagtggaatg gaaacgatta aaaaatgtgg tgccgcaccc agttatcaaa
14071 aataaaaatt taaagtcggt atacgtaaca aaagataatg tgaaagaggt tcaaaaagaa ttaggtttct
14141 ttgaaatttt taatgaagaa gtgttattaa ctggattttt atcatttcaa aggataccta tttacattat
14211 ttggattaat cctaaatctc ataagacgcc tagatattac tttgctaacg agcatgagat tgaaagatat
14281 tttgaatttt tggaggacga gtaaatgctt gaaatcatcg accaacgtga tgcattgcta gaagaaaagt
14351 atttaaacga cgactggtgg tacgagctag attattggtt gaataaacgc aagtcagaaa atgaacagat
14421 tgatattgat agagtgctta aatttattga ggaattaaaa cgataggaga taacgaataa atgaataatt
14491 taacagtaga tcaattaaaa gaacttttac aaatacaaaa ggagttcgac gatagaatac cgactagaaa
14561 tttaaatgac acagtagcta gtatgattat tgaatttgcg gagtgggtta acacacttga gttttttaaa
14631 aattggaaga aacaaccagg taagccatta gatacacaat tagatgagat tgctgattac ttagctttca
14701 gtttgcaatt aactctgact attgttgatg aagaagattt ggaagagact actgaggtta tggttgattt
14771 gattgaaaat gaagttactt tacctaaact acattcagtt tattttgttc atgtaatgca tacactaaca
14841 gaacaatttg taaaaggtat tgataatagt attgtacaag ttttaataat gccttttttg tacgccaata
14911 cttactatac aatcgaccaa ctcattgacg catacaaaaa gaaaatgaaa aggaaccacg aaagacaaga
14981 tggaacagca gacgcaggaa aaggatacgt gtaaagacat cttagatcga gtcaaggagg ttttggggaa ~"
15051 gtgacgcaat acttagtcac aacattcaaa gattcaacag gacaaccaca tgaacatttt actgctgcta
15121 gagataatca gacgtttaca gttgttgagg cggagagtaa agaaggagcg aaagagaagt acgagaaaca
15191 agttaagata aggagagatg gagatgccaa agaaaacggt aacgattgat gtagatgaaa acttattagt
15261 agtagctagt aatgaaatat cagaactatt atatgaatat gacagtgagt taatgtcagc tgatgaagat
15331 ggcgataata gagatatcga aaaaaaaaga gacgcattaa aacaagctat acaaattatc gataaattaa
15401 catgtcgagg aggcagacga tgattaacat acctaaaatg aaattcccga aaaagtacac tgaaataatc
15471 aagaaatata aaaataaaac acctgaagaa aaagctaaga ttgaagatga tttcattaaa gaaattaatg
15541 ataaagacag tgaattttac agtcctatga tggctaatat gaatgaacat gaattaaggg ctatgttaag
15611 aatgatgcct agtttaattg atactggaga tggcaatgat gattaaaaaa cttaaaaata tggattggtt
15681 cgatatcttt attgctggaa tactgcgatt attcggcgta atcgcactga tgcttgttgt catatcgcct
15751 atctatacag tggctagtta ccaaaacaaa gaagtatatc aagggacaat tacagataaa tataacaaga
15821 gacaagataa agaagacaag ttctatattg tgttagacaa caagcaagtc atcgaaaact ctgacttact
15891 attcaaaaag aaatttgata gcgcagacat acaagctagg ttaaaagtag gcgacaaagt agaagttaaa
15961 acgattggtt atagaataca ctttttaaat ttatatccgg tcttatacga agtaaagaag gtagataaat
16031 aatgattaaa caaatattaa gactattatt cttactagcg atgtatgagc taggtaagta tgtaactgag
16101 aaagtatata ttatgacgac ggctaatgat gatgtagagg cgccgagtga cttcgcaaag ttgagcgatc
16171 agtctgattt gatgagggcg gaggtgtcag agtagatgta tagcaaagag tcaattgtta atatgatagg
16241 cacacataaa atgaagtgta atgtattagc tgatgtaata ccggaatatg atagcaattc aattgcacag
16311 tatggcatac aagcaacgtt gccgaaacca caaggggaaa actcaagtaa agttgaagat gttgttgtga
16381 ggcttgagag agcaaataaa aggtatgctc agatgttaaa agaggttgag tttataaatc aatcgcaaca
16451 gagattggga cacgttgact tttgcttctt agagttattg aagaaaggtt ataacaggga tgcgattatc
16521 aagaagatgc ctaactctaa attaaataga aacaacttct tagcgcgccg tgatgagtta gcagaaaaga
16591 tttatctact acagtgacga aaatgacaaa aatgacagaa atgacgaaaa tgacactatt tttaaactgt
16661 gaattaattt tatataattg atttgtaaga attatcttaa gacgtggggt aatagccaca ttagatgttc
16731 tcatcgatgt gattgagaag tgacaaacat ataaaagatg atatgttacg ctattaatca cctactacct
16801 gcctatatgg tgggtagttt aattcttgca ttttgagtca taactatttt cctcctttca catttattga
16871 acgtagctcc tgcacaagat gtaggggcat tttttatatt taaataacta gagtaattaa cgtaaaggcg
16941 tgtgatacag tgaaaacaat tgattaaatt aacaccgaag caagaaaagt ttgtgctagg actcatagag
17011 ggcaagagcc aacggaaagc atatattgac gcagggtatt cgactaaagg taagagtggg gaatatctag
17081 ataaagaagc gagtacactt tttaaaaatc ggaaggtttc cggaaggtac gaaaaattgc gtcaagaagt
17151 agctgaacaa tcaaaatgga cacgccaaaa ggcctttgaa gaatatgagt ggctaaagaa tgtagctaag
17221 aatgacattg aaatagaggg agtgaagaaa gcgacagctg atgcattcct cgctagttta gatggtatga
17291 atagaatgac gttaggtaac gaagttttag ctaaaaagaa aatagaaact gaaattaaga tgcttgagaa
17361 gaagattgaa caaatagata aaggtgacag tggaacagaa gataaaatca aacaacttca cgacgcaata
17431 acggaagtga tcgtcaatga ataaacttaa atctttatat acggacaaac aaattgaaat attgaagcaa
17501 acgcaaaaac aagattggtt tatgttaatt aatcacggag caaagcgtac aggtaaaaca atattaaaca
17571 atgacttatt tttacgtgag ttaatgcgtg tgcgaaagat agcagacgaa gaaggaattg agacacctca
17641 atatatactt gctggtgcaa cattaggtac gattcaaaaa aacgtactaa tagagttaac taacaaatat
17711 ggcattgagt ttaattttga taaatataat tcattcatgt tatttggcgt tcaagtggtt cagacaggtc
17781 acagtaaagt aagtggtata ggagctatac gtggtatgac atcgtttggt gcatatatca atgaagcgtc
17851 gttagcgcat gaagaggtgt ttgacgagat taagtcacgt tgtagtggaa ctggtgcaag aatattggta
17921 gataccaacc ctgaccatcc cgagcattgg ttgttgaaag attatattga aaatacagat cctaaagcag
17991 gtatactgag tcaccaattt aagctcgatg acaataactt tcttaatgat agatataaag agtctattaa
18061 ggcttcaaca ccatcaggta tgttctatga acgtaatatc aacggtatgt gggtgtctgg tgacggtgta
18131 gtatatgccg actttgattt gaatgagaat acgattaaag cagatgaact ggacgacata cctatcaaag
18201 aatactttgc tggtgtcgac tggggttacg agcactatgg atctattgtg ttaataggac gaggtataga
18271 tggtaacttt tattttattg aggagcacgc acaccaattt aagtttattg atgattgggt ggttattgca
18341 aaagatattg taagtagata tggcaatatt aatttttact gcgatactgc acgacctgaa tacatcactg
18411 aatttagaag acatagatta cgtgcaatta acgctgataa aagtaaacta tcgggtgtgg aggaagttgc
18481 taagttgttc aaacaaaaca agttacttgt tctttatgat aatatggata ggtttaagca agaggtattt
18551 aaatatgttt ggcaccctac aaacggagag cctataaaag aatttgatga cgtgttggac tcgttaagat
18621 atgccatata cacacatact aaacctgaac gattaaggag ggggaaatga cattgtataa gttaatagat
18691 gatattgaag cacaaggaat attgcctaag catattgagg ctctaataga gtcacataaa gacgatagag
18761 agagaatggt taatctctat aatagataca agacacatat tgactatgta ccaatattca aacgtcgacc
18831 aattgaagaa aaagaagatt ttgaaactgg tggaaatgta aggcgattag acgtgtctgt taataacaaa
18901 cttaacaact cttttgacag cgaaattgtt gatacacgtg ttggttattt acatggtgtt cctgttactt
18971 atgatttaga tgaaaacgca gaaaaaaacg aaaagttgaa aaagtttata accaactttg ccattagaaa
19041 tagtgttgat gatgaggatt ctgaaatagg taaaatggca gcaatttgcg gatatggtgc taggttagca
19111 tatattgata cgaatggtga tattaggatt aagaatatag atccctataa tgttattttt gttggcgaca
19181 atattttaga acctacatac tcattgcgct acttttatga aaaagatgat gataatggca ctgattatgt
19251 gtacgcagag ttttacgata atgcttatta ttatgtattt cgaggagaag gtattgacgc tttgcaagaa
19321 gttggacgat atgaacattt atttgattac aatccattgt ttggtgtacc taacaacaaa gagatgatag
19391 gagatgctga aaaggttatt cacttaattg acgcatatga tttaacaatg agcgatgcat caagtgagat
19461 tagtcagaca cgtttagcat accttgtgtt acgcggtatg ggtatgagtg aagaaatgat tcaagaaaca
19531 caaaagagtg gcgcatttga gttgttcgac aaagatatgg acgttaaata cttaacaaaa gatgtaaatg
19601 acacaatgat tgagaaccat ttagatcgaa tcgaaaagaa tatcatgcgt tttgcaaagt cagtaaactt
19671 taattctgac gagtttaacg gaaatgtacc tatcattgga atgaaactta aacttatggc tttagagaac
19741 aagtgtatga cgtttgagcg taagatgaca gctatgttga ggtatcaatt caaagttatt ttatctgcat
19811 taaagcgtaa agggtacaac ttggatgatg atagttattt aaacctgata tttaagttca ctcgtaacat
19881 tccagttaat aagttagaag aatcacaagt gctaattaac ctgaagggac aagtttcaga acgaacaagg
19951 ttaggacaat cacaactagt tgatgatgtt gattacgaat tagacgaaat ggaaaaagaa agtcttgaat
20021 ttaatgacaa attacctgac atagatgaag gtgacgcaaa tgacaaatcc caaaataacc aatcagaatg
20091 atattgatga gtatatcgag ggtttaatct ctaaagcaga aaaaccaata gaacaactat ttgctaatcg
20161 acttaaagag ataaaacaaa tcatcgcaga tatgtttgag aaatatcaaa atgatgatgt gtatgtt-aca
20231 tggactgaat tcaataaata caacaggctc aataaggagt taactcgtat aggtacaatg ttgacttatg,
20301 actataggca agtagctaag atgattcaga agtcacaaga agatgcttat atagaaaaat tcσttatgag
20371 cctttattta tatgaaatgg cgagtcaaac atctatgcag tttgatgttc cgagtaaaga ggtaatcaaa
20441 tcagctattg aacaacctat tgagttcatt cgtttaatgc caacactaca aaaacatcgt gatgaagtat
20511 tgaaaaagat acgtatgcac attacacaag gtattatgag tggagagggt tactctaaga tagctaaagc
20581 aatacgtgat gatgtcggca tgtctaaagc tcaatcattg cgtgtggctc gtacagaagc aggcagagca
20651 atgtcacaag ctggacttga tagcgcaatg gttgctaaag ataacggttt gaatatgaag aaacgttggc
20721 atgctactaa agatacacga acacgtgata ctcatcgtca tttagatggg gaatcagtgg aaatagatca
20791 gaattttaaa tcaagtgggt gtgttgggca ggcgcccaag ctatttattg gtgtaaacag tgcgaaagag
20861 aatattaatt gtcgttgcaa attactttat tatattgatg aaaatgaatt gccaactgta atgagagcac
20931 gtaaagacga tggtaaaaat gaagttatcc cattcatgac ttatcgtgag tgggagaaat ataagcgaaa
21001 aggtggtaat tgatatggat tttaaaataa aagtaaatgt tgatactggc gaagctatag aaaagttaga
21071 acgcattaaa tccttgtacg aagagataat agagttacaa aacgaaaaag ttgttgtaaa cgtaacagtt
21141 aaaaatgaag ctgatttaga tatggttaaa acatctatta gcgaagaaaa tgctaaaaat aatgatttca
21211 cactttttta gttgtctctt tgctactcga ccttagcatg tcgttaaact gctttttatt atgcactttt
21281 cggactgtta gggtacgcga agggcaaaaa ggagttttga tatatgaata tcgaagaagt taagtctttt
21351 tttgaagaac acaaagacga taaagaagta aaagattatc taaagggact taagacggtg tctgttgatg
21421 acgttaaagg ctttttagat acagaagaag gtaaacgatt cattcaacct gaattagatc gttatcattc
21491 gaaaggatta gaatcatgga aagagaaaaa tcttgaggat ctaatcgaac aagaagtacg gaagcgtaat
21561 cctgagcaat cagaagaaca aaaacgtatt agtgctcttg aacaagagtt agaaaaacgc gacgcagagg
21631 caaaacgtga gaagttaaga agtaacgcgc taggtaaagc gcaggaacta aatttaccaa catccttagt
21701 tgatagattt ttaggcgatt ctgatgaaga tactgagcaa aacttaaaag ctttaaaaga aacctttgac
21771 aagtatgttc aaaaaggcgt tgagtctaaa tttaaatcga gtggaagaga tgttaaagaa tcacgaaatc
21841 aagatttaga cccttcaaat gtaaagtcca ttgaagaaat ggcgaaagaa atcaatatta gaaaataaag
21911 tgaggtaata aaatatggca actccaacat acacgccagg caatgttatt ttatcggatt ttaaaaacgg
21981 cgttattcca gcagaacaag gtactttaat catgaaagac attatggcta attcagcaat tatgaaatta
22051 gctaaaaatg agccaatgac agcacaaaag aaaaaattta cttacttagc aaaaggtgta ggcgcctact
22121 gggtatcaga aacggaacgt attcaaactt ctaagcctga atatgcgcaa gcagaaatgg aagctaagaa
22191 aattggtgta attattccgt tatcaaaaga gtttcttaaa tggactgcaa aagatttctt taatgaggtt
22261 aaacctctaa ttgcagaggc attttacaaa gcgtttgacc aagctgttat ctttggtact aaatcacctt
22331 acaacacttc aactagtggt aaaccgcttg ttgaaggcgc agaagagaaa ggtaacgttg ttacagatac
22401 taataattta tacgtagacc tttcggcatt aatggctact attgaagatg aagagttaga tccaaacgga
22471 gtattaacta cacgttcatt cagaagtaaa atgcgtaatg ctttagatgc taatgacaga ccattatttg
22541 atgctaacgg gaacgagatt atgggattac cactatctta tactggagcg gatgtatacg acaaaaagaa
22611 atcgttagca ctaatgggtg attgggatta cgcacgttac ggtatcttac aaggtattga gtatgcaatt
22681 tctgaagatg ccacgttaac gacgttacaa gcatcagatg cttctggcca accagtatca ttatttgaac
22751 gtgatatgtt cgctttacgt gcgacgatgc atattgcata catgaacgtt aaaccagaag cgttcgcaac
22821 gcttaaacca actgaatagg aggagatatg atggctaatc ctgcagaaga gattaaggta aaaaaagaca
22891 atatgactat tactgttaca aagaaggcat ttgactctta ttacagtctt gtcggttaca aagaggttaa
22961 atcacgtcgt actacgtctg ataagagcga gtgataaaaa tgactcttta tgaagatgtt aaacttttac
23031 tcaagaaaaa tggagtggaa gttaaaagtg atgaagaaga aatatttaag atggaagttg acggaatact
23101 agaagatgtt agggatataa caaacaatga ttttatgaaa gatggtcaag tcatttatcc ttactcaatc
23171 aaaaagtatg tcgcagatgt cctagagtat tatcaacgac ctgaagttaa aaagaattta aagtcaagaa
23241 gtatggggac agtgtcgtac acttataacg atggtgtccc tgattacatt agtggagtat taaacaggta
23311 taaacgagca aagtttcatc cgtttaaacc aataaggtag aggtgttgtt tgtgtttaac ccatacgacg
23381 aattccctca cactatttct attggaagta tcaaaaaagt aggagagtat ccaattatac aagagcgctt
23451 tgtaagcgat aaaacaatta aaggatttat ggatacgcct actacatctg aacaactaaa atttcatcaa
23521 atgtcacaag aatatgacag aaacctatat gtaccttatg acttgccaat atctaaaaac aatttatttg
23591 agtatgaggg tagaatcttt agtattgaag gtgattctgt agatcagggc ggacaacatg aaattaagtt
23661 actacgactt aagcaggtgc catatggcaa aagttaagta cggtgctgat agcatggttg ttgaattgga
23731 taagttcgat aagaaaatag aagagtgggt taaaaaaggt attgctaaaa caacgacgaa gatttacaac
23801 actgctgtag cattagctcc tgttgactta ggttttttag aagaaagtat tgactttaaa tatttcgatg
23871 gtgggttatc cagtgttata agtgtcggcg cagattatgc aatatacgtt gaatacggta ctggtatata
23941 tgctactggt cctggtggta gtcgtgctac aaagattccg tggagtttta aaggtgatga cggcgaatgg
24011 tacaccacat atggtcaagc gccacagcca ttttggaacc ctgcaattga cgcaggacgc aagacattcg
24081 agcagtattt ttcatagagg tggttaaata tgtgggtatc agttgagcct gaacttacaa atcaaatata
24151 taaaagatta atctcagacc ctaacattaa caaactagtt gatgataggg tttttgacgt tgttcaagat
24221 gacgctgttt acccatatat tgttgtgggt gaatcaaacg tcactaacaa cgaatctagc gcaacaatga
24291 gagaaacagt cggtattgtc atacatgtgt attcacagtt cgctacacaa tacgaggcta agctcatttt
24361 aagcgcgata ggttatgtgc ttaacagacc tatagaaata gataattacg agtttcaatt tagccgtatc
24431 gatagtcaag cagtattccc tgatatagac aggtttacta agcatggcac gatacggctt ttatttaagt
24501 acagacataa aaagaaaaac gaaggagtgt attaaatggc gcaaaaaaac tatttagcag ttgtacgtcc
24571 agctgaaact gacttagatc cagtagaatc tttattatta gctgacttac aagaaggtgg acatacgatt
24641 gaaaatgatt tagctgaaat agtacgaggc ggtaaaacgg actattctcc caatgcaatg tcagaatcat
24711 ttaaattaac aattggtaat gtgcctggag ataaaggaat tgaagcagtg aaacacgctg tacaaacagg
24781 tggacagttg cgtatatggc tttatgagcg taataaacgt gcagacggta aacatcacgg aatgtttggt
24851 tatgttgttc cagaatcatt tgaaatgtca tttgatgatg aaagtgacaa aatcgaacta tcattaaaag
24921 ttaaatggaa tacagcagaa ggtgctgaag ataacttgcc gaaagagtgg tttgaagctg caggtgcgcc
24991 tacagttgaa tacgaaaaat tcggcgaaaa agtcggaaca ttcgagaatc aaaagaaagc tagtgttgta
25061 tctgattcac acacggaaga ccattctatg taaactaata gatcaagggg gcgtaagctc cctatttttt
25131 tataaaaaaa ttgaaaagag gtatatattt tgactgaatt taatccaatt acaacattaa aaattaatga
25201 cggagaaaaa gattacgaag tagaagcaaa agtaacattt gcatttgacc gaaaagctga aaaattctca
25271 gaagatagcg aagatgggag aaaaggagca atgccaggat tcaatgttat ctttaacggt ttgctagaat
25341 ctagaaacaa agcgatttta caattttggg aatgtgctac tgcttattta aaaaacccac caactcgaga
25411 acaattagaa aaagcaattg atgatttcat cactgaaaac gaggatactt tgccgttatt acaaggggct
25481 ttggacaaac ttaacaatag tggttttttc aagagggaga gtcgctcgta ctggatgaca ttgaacaaag
25551 caccgaatat ggccaaaagc gaggacaaag aaatgacgaa agcaggcata gaaatgatga aagagaatta _,
25621 caaggaaatc atgggcgcag aaccttacac gattactcaa aaataaggca actgacagct agatatttag
25691 gatatatccc tgaacatgaa ttgttagcac taacacctgc tgaatggcgt gattggctta ttggtggtca
25761 ggataggtac ctagatcaaa gacaattatt aattgaacaa gcgcaagcta acggcttagt acaagcttct
25831 aagaggctaa ctagtatgat tcgtgacatt gagaaacaac gttacgaaat aagagaacct ggtagctatg
25901 ctcgtgtaca aaaagctaga ttagaagaag aaaaaagaag acgtgaactc ttcaaagaag gtacaagaaa
25971 attccttgaa tcgaaaggag gttagccttt ggatactcat tttatggcaa agattatggc caatattaga
26041 gatttccaaa gcaacgtaag gaaagctcaa cgattagcaa agacgtctgt accaaacgaa attgaaacag
26111 atgtaaaagc agatatttca agattccaaa gagctttaca acgcgctaaa tcaatggctc aacgatggcg
26181 agagcattct gttaaattat tcatgaaaac agatgagtat aaagcgaatt tagaacgcgc taaagctcaa
26251 gtagagcgat ttaaacaaca taaagtagat ttgaaactaa gtaacactga attaatggcc aaatataatg
26321 caactaaagc tactgtcgaa gcttggagaa aacatgttgt taagttggat ttagatgcaa accccgctaa
26391 aatggcggtt aaagggttta aagaagattt aatagatctt agcaggcata gttttgatat tgattccagc
26461 agatggaaat taggaaataa attcacaaaa gaattcaatg aagtcgaagg agcagttaaa cgttctttcg
26531 gaagaattgg tcagattatg agaaaagaag taaatggaac aagtgatatt tggggtaaac ttaacaactc
26601 attgaaagat tacggcgaga aaatggacgc cttagctact aaaatccgaa ctttcggtac tatcttcgcg
26671 caacaggtca aaggcttaat gattgctagt atacaagcat tgataccagt gattgccgga ttagtacctg
26741 caataatggc agtacttaat gcggttggtg tattaggtgg tggcgtttta ggtttagttg gcgcattctc
26811 tgtcgcaggt cttggagttg ttggctttgg tgcaatggct attagcgctc ttaaaatggt tgaagatgga
26881 acattggcag taacaaaaga agttcaaaac tttagagatg cgagcgatca gttaaaaact acatggcgtg
26951 atattgttaa agagaatcaa gcaagtatct ttaatgcgat gtcagcaggt atcagaggcg ttacaagtgc
27021 gatgtctcaa ttaaaaccat tcttatccga agtatctatg ctagttgaag caaacgcacg cgagtttgag
27091 aattgggtta aacattccga aacagctaag aaagcgtttg aagcattgaa tagcataggt ggcgcaatct
27161 tcggagattt attgaacgct gcaggacgat ttggcgacgg attagttaac attttcactc aattaatgcc
27231 gttgttcaaa tttgtgtctc aaggactaca gaacatgtct atagctttcc aaaattgggc taatagtgta
27301 gctggtcaga atgctattaa agcgtttatt gactacacta ccactaactt acctaagatt ggtcagatat
27371 ttggtaatgt gttcgctggt attggtaatt taatgattgc ttttgcacaa aacagttcca acatttttga
27441 ttggttggtt aaattaactt ctcaatttag agcatggtca gaacaagtag gacaatcaca agggtttaaa
27511 gactttatca gttatgttca agagaatggt cctactatta tgcagttaat cggtaatatc gtaaaagcat
27581 tagttgcttt tggtactgca atggctccta tagctagtaa attgttagac tttatcacta atctagctgg
27651 atttatcgct aaactattcg aaacacaccc agctatagca caagttgctg gcgttatggg tattttaggc
27721 ggtgtatttt gggctttaat ggctccgatt gttgctataa gtagtgtact tacaaatgtg tttggtttga
27791 gcttattcag cgtcactgaa aagattttag acttcgttag aacatcaagt ttagttactg gagctacgga
27861 agcattaata ggtgcattcg gttcgatttc agcacctatt ttagcagttg ttgcagtaat tggtgcattc
27931 attggtgtcc tcgtttattt atggaaaaca aacgagaact ttagaaatac tattactgaa gcgtggaacg
28001 gtgttaaaac ggcagtttct ggtgcgattc aaggtgtagt cggctggtta actgaattgt ggggcaaaat
28071 ccaatctacc ttacaaccga taatgcctat attgcaagta ttaggacaaa tattcatgca agttttaggt
28141 gttttggtaa taggcatcat tacaaacgtt atgaatatca tacaaggttt gtggacttta attacaattg
28211 cgttccaagc cataggaaca gtgatatccg tagcagtcca aatcatagta ggtttgttca ctgctttaat
28281 tcagttgctt actggcgact tctcaggtgc ttgggagact attaaaacta cggttaccaa tgtgcttgat
28351 acgatttggc aatacatgca atcagtttgg gagtcaatta tcggcttttt aactggcgta atgaatcgaa
28421 cactttctat gtttggtaca agttggtcac agatatggag tacaatcact aattttgtta gcagtatttg
28491 gaacactgtt acaagttggt tcagtcgagt ggcttcgagt gtagctgaaa aaatggggca agcactaaac
28561 tttattatca caaaaggttc tgaatgggtt tctaacattt ggaatacagt tacaagtttc gcgagtaaag
28631 tagctgatgg gtttaaaaga gttgtctcaa atgtaggtga cggtatgagt gatgcacttg gtaagattaa
28701 aagtttcttc agtgatttct taaatgccgg agcggaatta atcggcaaag tagctgaggg tgtagccaaa
28771 tctgcgcaca aagtagtcag cgcggtaggc gatgcgattt catcagcttg ggactctgta acttcattcg
28841 taagtggaca cggtggaggt agtagcttag gtaaaggttt agcggtatca caagcaaaag taattgctac
28911 agactttggc agtgccttta ataaagagct atcctctact ttgacagata gtatagtaaa tcctgtaagt
28981 acttctatag acagacacat gactagcgat gttcaacata gcttaaaaga aaataataga cctattgtga
29051 atgtaacgat tagaaatgag ggcgaccttg atttaattaa atcacgcatt gatgacatga acgctataga
29121 cggaagtttc aacttattat aagggaggtt tgttagttga tagcgcacga tatagaagta ataaggaatg
29191 gttcacagta tcgcgtcagt gacaatcctt tcacttataa tcacttggaa gtagttgaat ataacgttac
29261 aggcgcagga tatcatcgta actattctga tatagagggt attgatggta gatttcataa ttacgctaaa
29331 gaagaactta aaaaagtaga gcttaagata aggtataaag tacctaaaat tgcttatgct tcacatttaa
29401 agtcagacgt ccaagcacta tttgctggac gtttttattt aagggaatta gctacaccag acaattcaat
29471 taagtatgag catatattag atataσcaaa agacaaacaa gcatttgagc ttgattatgt tgatggacga
29541 caactttttg taggactagt aagtgaagtt tcttttgaca caacacaaac atcaggggaa ttttctttgt
29611 cgtttgaaac aaccgaacta ccatactttg aaagtgtcgg ttatagtact gatcttgaaa gtaataacga
29681 ccctgaaaaa tggtcggtac ctgatagatt gcctacaaac gaaggtgata agaggcgtca aatgacattt
29751 tacaacacta actcaggaga agtttattat aacggtgatg ttcctttaac acagtttaat cagtttaatg
29821 ttgttgaaat agagttagct gaagatgtta aagctaatga taaggatgga ttcactttct atacagataa
29891 aggaaatatc tcagttatta aggaagttga tttaaaagcc ggagataaaa taatcttcga cggtaaacat
29961 acctatagag gttatttaaa tatagattct tttaataaaa ctttagaaca accggtttta tatccaggct
30031 ggaatcgatt caagtctaat aaagtaatga aacaaattac atttagacac aaattatatt ttagataagg
30101 agtagcctat gccaatttta ttaaaaagtc tacagggtgt agggcacgct attaatgtta gtacaaaggt
30171 aagtaaaaag ctaaatgaag atagttcttt ggatctaact attatcgaga acgcgagtac gtttgacgca
30241 ataggtgcta taactaaaat gtggacgatc actcatgttg aaggtgaaga tgatttcaac gaatatgtaa
30311 ttgtcatact tgataagtct actattggcg aaaaaataag gcttgatatc aaagctaggc aaaaagaact
30381 tgatgacctt aacaattcta ggatttacca agagtataac gaaagtttta caggcgttga gttcttcaat
30451 actgtcttta aaggaacggg ttataagtat gtattacatc caaaagtaga tgcatctaaa ttcgagggat
30521 taggcaaagg agatacacga ttagaaatct ttaaaaaagg acttgagcgt tatcatctcg aatatgaata
30591 cgatgcaaag actaaaacgt ttcatttgta tgatgaatta tctaagtttg ccaattatta cattaaagct
30661 ggtgtgaatg ctgataacgt caaaatacaa gaagatgcat ctaaatgtta tacctttatt aaaggttatg
30731 gtgattttga tggacaacag acttttgcag aagcgggact acaaattgaa ttcactcatc cattagcaca
30801 attgataggt aaaagagaag cgccaccgct tgttgatgga cgtattaaaa aagaagatag tttaaaaaaa
30871 gcaatggagt tattgataaa gaaaagtgtc actgcttcta tttccttaga ctttgtagcg ttacgtgaac
30941 atttcccaga agctaaccct aaaataggtg atgttgttag agtggtggat tctgccatag gatataacga
31011 cttagtgaga atagtcgaaa tcactacaca tagagatgcg tacaataata tcactaagca agatgtagta
31081 ttaggagact ttacaaggcg taatcgttat aacaaagcag ttcatgatgc tgcaaattat gttaaaagcg
31151 taaaatctac aaaatccgac ccatctaaag aactaaaagc attaaacgca aaagttaacg caagtttatc
31221 tataaataat gaattggtta agcagaatga aaaaataaac gctaaagtcg ataagatgaa tactaaaaca
31291 gttacaactg ctaatggtac gatcatgtac gactttacta gtcaatcaag tataagaaac atcaaatcaa
31361 ttggaacgat tggcgactct gtagctagag ggtcgcacgc aaaaactaat ttcacagaaa tgttaggcaa 31431 gaaattgaaa gctaaaacga ctaatcttgc aagaggtggc gcaacaatgg caacagttcc aataggtaaa 31501 gaagcggtag aaaacagcat ttatagacaa gcagagcaaa taagaggaga cctaatcata ttacaaggca 31571 ctgatgatga ctggttacac ggttattggg caggcgtacc gataggcact gataaaacgg atacaaaaac 31641 gttttacggt gccttttgtt ctgcaattga agttattaga aagaataatc cagattcaaa aatactagtg 31711 atgacagcta caagacaatg ccctatgagt ggtacaacaa tacgccgtaa agacacggac aaaaacaaac 31781 tagggttaac acttgaggac tatgtaaacg ctcaaatatt agcttgtagt gagttagatg taccagtgtt 31851 tgacgcatat cacacagatt actttaagcc atacaatcca gcttttagga aagcgagcat ggaggacggc 31921 ttacacccta acgaaaaagg tcacgaggtt attatgtacg agttaatcaa ggattattac agtttttacg 31991 actaaaggag gcaaccaatg gcttacggat taattacaag tttacattca atgacaggtc ggaaaatagt 32061 tgctcaacat gagtataact atcgcttgtt agatgaaggt atgagcaaac ttgagaaaat gtttatatac 32131 catcaaaaag aagaaatata cgcacactca gcgaaacaaa ttaaatactt gaatgacagt gttgaagatt 32201 atttaacgta tttaaatagc cgttttagca atatgattct aggccataac ggcgacggta tcaatgaagt 32271 aaaagacgcg cgtattgata atacaggtta tggtcataag acattgcaag atcgtttgta tcatgattat 32341 tcaacactag atgctttcac taaaaaggtt gagaaagctg tagatgaaca ctataaagaa tatcgagcga 32411 cagaataccg attcgaacca aaagagcaag aaccggaatt tatcactgat ttatcgccat atacaaatgc 32481 agtaatgcaa tcattttggg tagaccctag aacgaaaatt atttatatga cgcaagctcg tccaggtaat 32551 cattacatgt tatctagatt gaagcccaac ggacaattta ttgatagatt gcttgttaaa aacggcggtc 32621 acggtacaca caatgcgtat agatacattg atggagaatt atggatttat tcagctgtat tggacagtaa 32691 caaaaacaac aagtttgtac gtttccaata tagaactgga gaaataactt atggtaatga aatgcaagat 32761 gtcatgccga atatatttaa cgacagatat acgtcagcga tttataatcc tatagaaaat ttaatgattt 32831 tcagacgtga atataaagct tctgaaagac aagctaagaa ttcattgaat ttcattgaag taagaagtgc 32901 tgacgatatt gataaaggta tagacaaagt attgtatcaa atggatatac ctatggaata cacttcagat 32971 acacaaccta tgcaaggtat cacttatgat gcaggtatct tatattggta tacaggtgat tcgaatacag 33041 ccaaccctaa ctacttacaa ggtttcgata taaaaacaaa agaattgtta tttaaacgac gtatcgatat 33111 tggcggtgtg aataataact ttaaaggaga cttccaagaa gctgagggtc tagatatgta ttacgatcta 33181 gaaacaggac gtaaagcact tttaataggg gtaactattg gacctggtaa taacagacat cactcaattt 33251 attctatcgg ccaaagaggt gttaaccaat tcttaaaaaa cattgcacct caagtatcga tgactgattc 33321 aggtggacgt gttaaaccgt taccaataca gaacccagca tatctaagtg atattacgga agttggtcat 33391 tactatatct atacgcaaga cacacaaaat gcattagatt tcccgttacc gaaagcgttt agagatgcag 33461 ggtggttctt ggatgtactg cctggacact ataatggtgc tctaagacaa gtacttacca gaaacagcac 33531 aggtagaaat atgcttaaat tcgaacgtgt cattgacatt ttcaataaga aaaacaacgg agcatggaat 33601 ttctgtccgc aaaacgccgg ttattgggaa catatcccta agagtattac aaaattatca gatttaaaaa 33671 tcgttggttt agatttctat atcactactg aagaatcaaa acgatttact gattttccta aagactttaa 33741 aggtattgca ggttggatat tagaagtaaa atcgaataca ccaggtaaca caacacaagt attaagacgt 33811 aataacttcc cgtctgcaca tcaattttta gttagaaact ttggtactgg tggcgttggt aaatggagtt 33881 tattcgaagg aaaggtggtt gaataatgat agtagataat ttttcgaaag acgataactt aatcgagtta 33951 caaacaacat cacaatataa tccaattatt gacacaaaca tcagtttcta tgaatcagat agaggaactg 34021 gtgttttaaa ttttgcagta actaagaata acagaccgtt atctataagt tctgaacatg ttaaaacatc 34091 tatcgtgtta aaaaccgatg attataacgt agatagaggc gcttatattt cagacgaatt aacgatagta 34161 gacgcaatta atgggcgttt gcagtatgtg ataccgaatg aatttttaaa acattcaggc aaggtgcatg 34231 ctcaggcatt ctttacacaa aacgggagta ataatgttgt tgttgaacgt caatttagct tcaatattga 34301 aaatgattta gttagtgggt ttgatggtat aacaaagctt gtttatatca aatctattca agatactatc 34371 gaagcagtcg gtaaagactt taaccaatta aagcaagata tggatgatac acaaacgtta atagcaaaag 34441 tgaatgatag tgcgacaaaa ggcattcaac aaatcgaaat caagcaaaac gaagctatac aagctattac 34511 tgcgacgcaa actagtgcaa cacaagctgt tacagctgaa gtcgataaaa tagttgaaaa agagcaagcg 34581 atttttgaac gtgttaacga agttgaacaa caaatcaatg gcgctgacct tgttaaaggt aattcaacaa 34651 caaattggca aaagtctaaa cttacagatg attacggtaa agcaattgaa tcgtatgagc agtccataga 34721 tagcgtttta agcgcagtta acacatctag gattattcat attactaatg caacagatgc gccagaaaag 34791 acggatatag gcacgttaga gaagcctgga caagatggtg ttgatgacgg ttcttcgttc gatgaatcaa 34861 cttatacatc aagcaaatct ggtgtgttag ttgtttatgt tgttgataat aatactgctc gtgcaacatg 34931 gtacccagac gattcaaacg atgagtacac aaaatacaaa atctacggca catggtaccc gttttataaa 35001 aagaatgatg gaaacttaac taagcaattt gttgaagaaa cgtctaacaa cgctttaaat caagctaagc 35071 agtatgtaga tgataaattc ggaacaacga gctggcaaca acataagatg acagaggcga atggtcaatc 35141 aattcaagtt aacttaaata atgcgcaagg cgatttggga tatttaactg ctggtaatta ctatgcaaca 35211 agagtgccgg atttaccagg tagtgttgaa agttatgagg gttatttatc ggtattcgtt aaagacgata 35281 caaacaagct atttaacttc acgccttata actctaaaaa gatttacaca cgatcaatca caaacggcag 35351 acttgagcaa cagtggacag ttcctaatga acataagtca acggtattgt tcgacggtgg agcaaatggt 35421 gtaggtacaa caatcaatct aaccgaacca tacacaaact attctatttt attagtaagt ggaacttatc 35491- caggtggcgt tattgaggga ttcggactaa ccacattacc taatgcaatt caattaagta aagcgaatgt 35561 agttgactca gacggtaacg gtggcggtat ttatgagtgt ttactatcca aaacaagtag cactacttta 35631 agaatcgata acgatgtgta ctttgattta ggtaaaacat caggttctgg agcgaatgcc aacaaagtta 35701 ctataactaa aattatgggg tggaaataat gaaaatcaca gtaaatgata aaaatgaagt tatcggatac 35771 gttaatactg gcggtttacg caatagttta gatgtagacg ataacaatgt gtctatcaaa ttcaaagaag 35841 agttcgaacc tagaaagttc gttttcacta acggcgaaat taaatacaat agcaatttcg aaaaagaaga 35911 cgtaccgaat gcatcaaacc aacaaagtgc gtcagattta agtgatgagg aacttcgcgg aatggttgca 35981 agtatgcaaa tgcagatgac gcaagtgaac atgttgacaa tgcaattgac gcaacaaaac gctatgttaa 36051 cacaacagtt gaccgaactg aaaactaaca aaacaaatac tgagggggac gtttaaatga tgaagatgat 36121 ttatccaact tttaaagaca ttaaaacttt ttatgtgtgg ggttgctata aaaatgagca aattaagtgg 36191 tacgtagaca tgggtgtaat cgacaaagaa gaatatgcat tgatcactgg tgaaaaatat ccagaggcaa 36261 aagatgaaaa gtcacaggtg taatgcttga ggctttttaa tttaacacaa agtaggtggc gtaatgtttg 36331 gatttaccaa acggcacgaa catgaatggc gaattagaag attagaagag aatgataaaa caatgcttag 36401 cactctcaat gagattaaat taggtcaaaa aactcaagag caagttaaca ttaaattaga taaaacttta 36471 gatgctatcc agagggaaag acagatagac gaaaaaaata agaaagaaaa cgacaaaaat atacgcgata 36541 tgaaaatgtg gattctcggt ttgataggga ctatcttcag tacgattgtc atagctttac taagaactat 36611 ttttggtatt taaaggaggt gattaccatg cttaaaggga ttttaggata tagcttctgg gcgtgcttct
36681 ggtttggtaa atgtaaataa cagttaagag tcagtgcttc ggcactggct ttttattttg attgaaatga
36751 ggtgcataca tgggattacc taacccaaag actagaaagc ctacagctag tgaagtggtg gagtgggcaa
36821 agtcgaatat tggtaagagg attaatatag ataattatcg gggcagtcaa tgttgggata cacctaactt
36891 tatttttaaa agatattggg gttttgtaac atggggcaat gctaaggata tggctaatta cagatatcct
36961 aagggtttcc gattctatcg ttattcatct ggatttgtac cggaacctgg agacatcgca gtttggcacc
37031 ctggcaacgg aataggttcg gacggacaca ccgcaatagt agtaggacca tctaataaaa gttattttta
37101 tagcgttgac caaaactggg ttaattctaa tagttggaca ggttctccag gaagattagt aagacaccct
37171 tatgtaagtg ttacaggctt tgttaggcct ccatactcaa aagatactag caaacctagt agtactgata
37241 caagttcagc atcaaaagcc aatgactcaa caattactgg cgaagcgaag aaaccgcaat ttaaagaagt
37311 taaaacagta aaatacactg cttacagcaa tgttttagat aaagaagagc acttcattga tcatatagtt
37381 gtaatgggtg atgaacgctc agatattcaa ggattatata taaaagaatc aatgcatatg cgttctgtag
37451 acgaactgta tacgcaaaga aataagttta taagcgatta tgaaataccg catttatatg tcgatagaga
37521 ggctacatgg cttgctagac caaccaattt tgatgacccg cgtcacccta attggctagt tattgaagta
37591 tgtggtggtc aaacagatag caaacgacaa ttcttattga atcaaataca agcgttaata cgtggtgttt
37661 ggttattgtc agggattgat aaaaacttat ctgaaacgac gttaaaggta gaccctaata tttggcgtag
37731 tatgaaagat ttaattaatt acgacttgat taagcaaggt ataccggata acgcaaagta tgagcaagtt
37801 aaaaagaaaa tgcttgagac atacattaaa cgagatatat tgacacgaga aaatataaaa gaagtaacga
37871 caaaaacaac aataagaatt agtgataaaa catcagttga cagtgcgtcc acacgaggcc ctactccatc
37941 agacgaaaaa ccaagcatcg ttactgaaac aagtccattc acattccagc aagcactgga tagacaaatg
38011 tctaggggta acccgaaaaa atctcataca tggggctggg ctaatgcaac acgagcacaa acgagctcgg
38081 caatgaatgt taagcgaata tgggaaagta acacgcaatg ctatcaaatg cttaatttag gcaagtatca
38151 aggcatttca gttagtgcgc ttaacaaaat acttaaagga aaaggaacgc tcgacggaca aggcaaagca
38221 ttcgcggaag cttgtaagaa aaacaacatt aacgaaattt atttgatcgc gcacgctttc ttagaaagtg
38291 gatacggaac aagtaacttc gctagtggta gatacggtgc atataattac ttcggtattg gtgcattcga
38361 caacgaccct gattatgcaa tgacgtttgc taaaaataaa ggttggacat ctccagcaaa agcaatcatg
38431 ggcggtgcta gcttcgtaag aaaggattac atcaataaag gtcaaaacac attgtaccga attagatgga
38501 atcctaagaa tccagctacc caccaatacg ctactgctat agagtggtgc caacatcaag caagtacaat
38571 cgctaagtta tataaacaaa tcggcttaaa aggtatctac ttcacaaggg ataaatataa ataaagaggt
38641 gtgtaaatgt acaaaataaa agatgttgaa acgagaataa aaaatgatgg tgttgactta ggtgacattg
38711 gctgtcgatt ttacactgaa gatgaaaata cagcatctat aagaataggt atcaatgaca aacaaggtcg
38781 tatcgatcta aaagcacatg gcttaacacc tagattacat ttgtttatgg aagatggctc tatattcaaa
38851 aatgagσccc ttattatcga cgatgttgta aaagggttcc ttacctacaa aatacctaaa aaggttatca
38921 aacacgctgg ttatgttcgc tgtaagctgt ttttagagaa agaagaagaa aaaatacatg tcgcaaactt
38991 ttctttcaat atcgttgata gtggtattga atctgctgta gcaaaagaaa tcgatgttaa attggtagat
39061 gatgctatta cgagaatttt aaaagataac gcgacagatt tattgagcaa agactttaaa gagaaaatag
39131 ataaagatgt catttcttac atcgaaaaga atgaaagtag atttaaaggt gcgaaaggtg ataaaggcga
39201 accgggacaa cctggtgcga aaggtgatac aggtaaaaaa ggagaacaag gcgcacccgg taaaaacggt
39271 actgtagtat caatcaatcc tgacactaaa atgtggcaaa ttgatggtaa agatacagat atcaaagcag
39341 aacctgagtt attggacaaa atcaatatcg caaatgttga agggttagaa gataaattgc aagaagttaa
39411 aaaaatcaaa gatacaactc tcaacgactc taaaacgtat acggattcaa aaattgctga actagttgat
39481 agcgcgcctg aatctatgaa tacattaaga gaattagcag aagcaataca aaacaactct atttcagaaa
39551 gtgtattgca acagattggc tcaaaagtta gtacagaaga ttttgaggaa ttcaaacaaa cactaaacga
39621 tttatatgct ccaaaaaatc ataatcatga tgagcggtat gttttgtcat ctcaagcttt tactaaacaa
39691 caagcggata atttatatca actaaaaagc gcatctcaac cgacggttaa aatttggaca ggaacagaaa
39761 atgaatataa ctatatatat caaaaagacc ctaatacact ttacttaatt aaggggtgat ttttatggaa
39831 ggtaatttta aaaatgtaaa gaagtttatt tacgaaggtg aagaatatac aaaagtatat gctggaaata
39901 tccaagtatg gaaaaagcct tcatcttttg taataaaacc cttacctaaa aataaatatc cggatagcat
39971 agaagaatca acagcaaaat ggacaataaa tggagttgaa cctaataaaa gttatcaggt gacaatagaa
40041 aatgtacgta gcggtataat gagggtttcg caaactaatt taggttcaag tgatttagga atatcaggag
40111 tcaatagcgg agttgcaagt aaaaatatca actttagtaa tccttcaggg atgttgtatg tcactataag
40181 tgatgtttat tcaggatctc caacattgac cattgaataa ttttaaacga ctaatttttt agtcgttttt
40251 tattttggat aaaaggagca aacaaatgga tgcaaaagta ataacaagat acatcgtatt gatcttagca
40321 ttagtaaatc aattcttagc gaacaaaggt attagcccga ttccagtaga cgatgagact atatcatcaa
40391 taatacttac tgttgttgct ttatatacta cgtataaaga caatccaaca tctcaagaag gtaaatgggc
40461 aaatcaaaag ctaaagaaat ataaagctga aaacaagtat agaaaagcaa cagggcaagc gccaattaaa
40531 gaagtaatga cacctacgaa tatgaacgac acaaatgatt tagggtaggt gttgaccaat gttgataaca
40601 aaaaaccaag cagaaaaatg gtttgataat tcattaggga agcagttcaa tcctgatttg ttttatggat
40671 ttcagtgtta cgattacgca aatatgtttt ttatgatagc aacaggcgaa aggttacaag gtttatacgc
40741 ttataatatt ccatttgata ataaagcaag gattgaaaaa tacgggcaaa taattaaaaa ctatgatagc
40811 tttttaccgc aaaagttgga tattgtcgtt ttcccgtcaa agtatggtgg cggagctgga catgttgaaa
40881 ttgttgagag cgcaaattta aacactttca catcatatgg gcaaaattgg aatggtaaag gttggacaaa
40951 tggcgttgcg caacctggtt ggggtcctga aactgttaca agacatgttc attattacga tgacccaatg
41021 tattttatta gattaaattt cccagataaa gtaagtgttg gagataaagc taaaagcgtt attaagcaag
41091 caactgccaa aaagcaagca gtaattaaac ctaaaaaaat tatgcttgta gccggtcatg gttataacga
41161 tcctggagca gtaggaaacg gaacaaacga acgcgatttt atccgtaaat atataacgcc aaatatcgct
41231 aagtatttaa gacatgcagg tcatgaagtt gcattatatg gtggctcaag tcaatcacaa gacatgtatc
41301 aagatactgc atacggtgtt aatgtaggaa ataataaaga ttatggatta tattgggtta aatcacaggg
41371 gtatgacatt gttctagaga ttcatttaga cgcagcagga gaaaatgcaa gtggtgggca tgttattatc
41441 tcaagtcaat tcaatgcgga tactattgat aaaagtatac aagatgttat taaaaataac ttaggacaaa
41511 taagaggtgt aacacctcgt aatgatttac tgaacgttaa tgtatcagca gaaataaata tcaattatcg
41581 tttatctgaa ttaggtttta ttactaataa aaaagatatg gattggatta agaagaatta tgacttgtat
41651 tctaaattaa tagctggtgc gattcatggt aagcctatag gtggtttggt agctggtaat gttaaaacat
41721 cagctaaaaa ccaaaaaaat ccaccagtgc cagcaggtta tacacttgat aagaataatg tgccttataa
41791 aaaagagact ggtaattaca cagttgccaa tgttaaaggt aataacgtaa gggacggcta ttcaactaat
41861 tcaagaatta caggtgtatt acctaataac gcaacaatca aatatgacgg cgcatattgc atcaatgggt
41931 atagatggat tacttatatt gctaatagtg gacaacgtcg ctatattgcg acaggagagg tagataaagc
42001 aggtaatagg ataagtagtt ttggtaagtt tagcacgatt tagtatttac ttagaataaa aattttgcta 42071 cattaattat agggaatctt acagttatta aataactatt tggatggatg ttaatattcc tatacacttt 42141 ttaacattac tctcaagatt taaatgtaga taacaggcag gtactacggt acttgcctat ttttttgtta 42211 taatgtaatt acattaccag taaccaatct ggcttaaaac cacatttccg gtagccaatc cggctatgca 42281 gaggacttac ttgcgtaaag tagtaagaag ctgactgcat atttaaacca cccatactag ttgctgggtg 42351 gttgtttttt atgttatatt ataaatgatc aaaccacacc acctattaat ttaggagtgt ggttattttt 42421 tatgcaaaaa aaacgaaaaa aagttcataa aaagtattgc atatcacgtt taaccgtgtt ataataaggt 42491 ataccagttg agaggaggat aaaaagtgtt agaaaatttt aaaactatag cagaaatcgc cttttataca 42561 atgtcagcaa ttgccatagc gaaaacattg aaaaaagacg ataagtaagt agacaagccc gaaagggctg 42631 tctatatata aattctaaca ctaaaatact atgaaaacaa tttacattat tttaatcatt cttatttgga 42701 taaacgtgtt tttaggcaac gatataagta aaagtgttgt tgcactgctt actactttac tgcttatcaa 42771 tttatggaag agggataaaa atgacagcaa taaaagaaat aattgaatca atagaaaagt tattcgaaaa 42841 agaaacggga tataaaattg ctaaaaattc cggattacca tatcaaactg tgcaagattt aagaaatgga 42911 aaaacatctt tatcagatgc cagatttaga acgataataa agttatacga gtatcaaaga tcgcttgaaa 42981 acgaagaaga taaataaaag gagccaaaaa tatgtttgtt acaaaagaag aatttaaaac tttgaatgta 43051 aaagaagtat ttgaatcagg taaaaacttt ataaaaatta cagatggaag acatgcaata tattgggtaa 43121 atgatagata cgtagtactt gaccataaaa aaggcgattt gtacccgcaa aaagcatacc caaaatatat 43191 caaaagaaaa ttagtaagtt aaataattag aaaaccacgt cttaattgac gtggttattt tttaggtttg 43261 cgcgtgtcaa atacgtgtca atttagttct atttctttag ttttctttct aaacttaatt gcttgtaaac 43331 cgcatagtta taggcttttc agctatatac caagataaga tttatcccgc cgtctccata aaaatatgct 43401 tggaaacctt gatttaatgg ggttttaatc tagcaagtgt caaatatgtg tcaagaaaat aattttctga 43471 cacgttgacc ttgctctttt ttatgttcat caagtaagtg agagtaggtg tctaaagtta tagatatatt 43541 ataatggcct aatcttttgc taatatattc aatagg
Table 10
Bacteriophage 96 ORFs list
Table 11
SEQUENCE INFORMATION FOR PHAGES MATCHING WITH TABLE 1
M32695 Bacteriophage PM2 nuclease cleavage site gi| 166145|gb|M32695|BM2NCS [166145] (View GenBank report.FASTA report-ASN.l report.Graphical view.l MEDLINE link, or 1 nucleotide neighbor )
M32693 Bacteriophage PM2 Hind III fragment 4 gi|166144|gb|M32693|BM24HIND3 [166144] (View GenBank report.FASTA report,ASN.l report,Graphical view.l MEDLINE link, or 1 nucleotide neighbor )
M32693 Bacteriophage PM2 Hind III fragment 4 gi|166144|gb|M32693|BM24HIND3 [166144] (View GenBank report.FASTA report,ASN.l reρort,Graphical view.l MEDLINE link, or 1 nucleotide neighbor )
M32694 Bacteriophage PM2 Hind III fragment 3 gi| 166143jgb|M32694|BM23HIND3 [166143] (View GenBank report,FASTA report,ASN.l report,Graphical view, or 1 MEDLINE link )
M26134 Bacteriophage PM2 structural protein gene containing purine/p rimidine rich regions and anti-Z-DNA-IgG binding regions, complete eds gi|289360|gb|M26134|BM2PROΗV [289360] (View GenBank report,FASTA report,ASN.l report.Graphical view.l MEDLINE link, or 1 protein link )
J02452 bacteriophage fϊ 3'-terminal region ma gi|215409|gb|J02452|PFITR3 [215409] (View GenBank report.FASTA report,ASN.l report,Graphical view, or 1 MEDLINE link )
AF020798 Bacteriophage Chpl genome DNA, complete sequence gi|217761|dbj|D00624|BCPl [217761] (View GenBank report.FASTA report^ASN.l report,Graphical view.l MEDLINE link, 12 protein links, or 1 genome link )
X72793 Clostridium botulinum C phage BONT/C1, ANTP-139, ANTP-33, ANTP-17, ANTP-70 genes and ORF-22 gi|516171|emb|X72793|CBCBONT [516171] (View GenBank report,FASTA report SN.1 report,Graphical view, 1 MEDLINE link, 6 protein links, or 4 nucleotide neighbors ;
X51464 Clostridium botulinum D Phage C3 gene for exoenzyme C3 gi| 14907|emb|X51464|CBDPE3 [14907] (View GenBank report,FASTA report^SN.l report,Graphical view.l MEDLINE link, 1 protein link, or 2 nucleotide neighbors )
D90210 Bacteriophage c-st (from C. botulinum) Cl-tox gene for botulinum Cl neurotoxin gi|217780|dbj|D90210|CSTClTOX [217780] (View GenBank report.FASTA report SN.l report,Graphical view.l MEDLINE link, or 1 protein link ) _. - -
S49407 type D neurotoxin [bacteriophage d-16 phi, host = C. botulinum, type D, CB 16, Genomic, 4087 nt] gi!260238|gb|S49407|S49407 [260238] (View GenBank report.FASTA report.ASN.l report,Graphical view.l MEDLINE link, or 1 protein link )
X53370 Bacteriophage phi29 temperature sensitive mutant TS2(98) DNA polymerase gene gi|15733|emb|X53370|POTS298 [15733] (View GenBank report,FASTA report,ASN. l report,Graphical view.l MEDLINE link, 1 protein link, or 7 nucleotide neighbors )
X53371 Bacteriophage phi29 temperature sensitive mutant TS2(24) DNA polymerase gene gi|15731|emb|X53371|POTS224 [15731] (View GenBank report.FASTA report.ASN. l report.Graphical view.l MEDLINE link, 1 protein link, or 7 nucleotide neighbors )
X05973 Bacteriophage phi29 prohead RNA gi|15680|emb|X05973|POP29PRO [15680] (View GenBank report,FASTA report-ASN.l report, Graphical view,2 MEDLINE links, or 4 nucleotide neighbors )
V01155 Left end of bacteriophage phi-29 coding for 15 potential proteins Among these are the terminal protein and the proteins encoded by the genes 1, 2 (sus), 3, and (probably) 4 gi|15659|emb|V01155|POP29B [15659] (View GenBank report,FASTA report-ASN.l report.Graphical view.l MEDLINE link, 16 protein links, or 16 nucleotide neighbors)
X73097 Bacteriophage phi-29 left origin of replication gi|312194|emb|X73097|BP29ORIL [312194] (View GenBank report,FASTA report.ASN.1 report, Graphical view.l MEDLINE link, or 5 nucleotide neighbors )
M 14430 Bacteriophage phi-29 gene- 17 gene, complete eds gi|215321|gb|M14430|P29G17A [215321] (View GenBank report,FASTA report-ASN.l report, Graphical view.l MEDLINE link, 6 protein links, or 8 nucleotide neighbors )
M14431 Bacteriophage phi-29 gene- 16 gene, complete eds gi|215319igb|M 14431 |P29G 16A [215319] (View GenBank report,FASTA report-ASN.l report, Graphical view.l MEDLINE link, 2 protein links, or 7 nucleotide neighbors )
M20693 Bacteriophage phi-29 DNA, 3' end gi|215343|gb|M20693|P29REPINB [215343] (View GenBank report,FASTA report-ASN.l report,Graphical view.l MEDLINE link, or 4 nucleotide neighbors )
M21016 Bacteriophage phi-29. DNA, 5' end gi|215342|gb|M21016|P29REPINA [215342] (View GenBank report,FASTA report-ASN.l report,Graphical view.l MEDLINE link, or 1 nucleotide neighbor )
M l 2456 Bactenophage phi 29 genes 9 10 and 11 encoding p9 tail, mcomplete, plO connector, complete and p i 1 lower collar, incomplete, respectively gι|215338|gb|M12456|P29P9 [215338] (View GenBank report.FASTA report,AS 1 report.Graphical \ ιew, l MEDLINE link, 3 protein links, or 2 nucleotide neignbors )
Ml 4782 Bacillus phage phi-29 head morphogenesis, major head protein head fiber protem, tail protein, upper collar protein, lower collar protein nre-neck- appendage protein, morphogenesιs(13), lysis, morphogenesιs( 15), encapsidation genes, complete eds gι|215323|gb|M14782|P29LATE2 [215323] (View GenBank report.FASTA report.ASN 1 reoort.Graphical view. l MEDLINE link, 1 1 protem Imks, or 1 1 nucleotide neighbors)
M26968 Bactenophage phi-29 (from Bacillus subtilis) proteins pi delta-1 genes, comϋlete eds and the susl(629) mutanon gι|341558|gb|M26968|P29P lD l A [341558] (View GenBank report,FASTA report,ASN 1 report,Graphιcal view. l MEDLINE link, 2 protem links, or 1 nucleotide neighbor )
J02448 Bactenophage fl complete genome gι| 166201 |gb|J02448|FlCCG [166201 ]
(View GenBank report.FASTA report,ASN 1 report.Graphical view, 1 MEDLINE link, 10 protein links, 205 nucleotide neighbors or 1 genome link )
M24832 Bactenophage f2 coat protein gene, partial eds gι| 166228|gb|M24832|F2CRNACA [166228] (View GenBank report,FASTA report,ASN 1 report.Graphical view, 1 MEDLINE link, 1 protem link, or 4 nucleotide neighbors )
J02451
Bacteriophage fd, strain 478, complete genome gι|215394|gb|J02451|PFDCG [215394]
(View GenBank report.FASTA report,ASN 1 report-Graphical vιe ,5 MEDLINE links, 10 protem links, 204 nucleotide neighbors, or 1 genome link )
M34834 Bactenophage fr replicase gene, 5' end gι| 166139|gb|M34834|BFRREGRA [166139] (View GenBank report,FASTA report-ASN 1 report,Graphical view.l protem link, or 9 nucleoπde neighbors )
M38325 Bactenophage fr replicase gene, 5' end gι|166137|gb|M38325|BFRREGR [166137] (View GenBank report,FASTA report-ASN 1 _eport,Graphical view, 1 protem link, or 9 nucleotide neighbors )
M35063 Bactenophage fr coat protem replicase cistron (R region) RNA gι|166134|gb|M35063|BFRRCRRA [166134] (View GenBank report.FASTA report,ASN 1 report,Graphιcal view, 1 protem link, or 3 nucleotide neighbors )
S66567 a -ha-ar l natπuretic factor/coat protειn=fusιon polypeptide [human, bactenophage rr, expression vector pFAN15, Pla≤midSyntheπcRecombinant, 510 nt] gι|435742|gb|S66567|S66567 [435742]
(View GenBank report,FASTA reportASN 1 report.Graphical view.l MEDLINE link, 1 protem lmk, or 15 nucleotide neighbors )
X15031 Bactenophage fr RNA genome gι| 15071|emb|X15031|LEBFRX [15071]
(View GenBank report.FASTA report.ASN 1 report.Graphical \ ιew, l MEDLINE link, 4 protem links, 9 nucleotide neighbors, or 1 genome link )
U51233 Mus musculus neurralizmg ann-RNA-bactenophage fr immunoglobulin vaπable region light cham (IgM) mRNA, partial eds gιfl277150|gb|U51233|MMU 1233 [1277150] (View GenBank report.FASTA report.ASN 1 report.Graphical view. l protem link, or 1669 nucleotide neighbors )
U51232
Mus musculus neurralizmg anti-RNA-bacteπophase fr immunoglobul_n vaπable region heavy chain (IgM) mRNA, partial eds gι| 1277148|gb|U51232|MMU51232 [1277148]'
(View GenBank report.FASTA report,ASN 1 report.Graphical \ lew, 1 protem link, or 1073 nucleotide neighbors )
U02303 Bactenophage Ifl, complete genome gι|3676280|gb|U02303|B2U02303 [3676280] (View GenBank report.FASTA report.ASN 1 report.Graphical view, 10 protem links, or 1 genome link )
V00604
Phage M13 genome gι|14959|emb|V00604|INM13X [14959]
(View GenBank report.FASTA report,ASN.1 repoπ,Graphιcal view.l MEDLINE link, 10 protem links, or 205 nucleotide neighbors )
A32252
Synthetic bactenophage M13 protem III probe gι|1567340|emb|A32252|A32252 [1567340]
(View GenBank report,FASTA report.ASN.1 report, or Graphical view)
A32251 Synthetic bactenophage Ml 3 protein HI probe gι| 1567339|emb|A32251|A32251 [1567339] (View GenBank report.FASTA report-ASN.1 report, or Graphical view)
Ml 2465 Bactenophage M13 mplO mutations in lac operon gι|215210|gb|M12465|M13LACMUT [215210] (View GenBank report,FASTA report-ASN. l report,Graphιcal view.l MEDLINE link, or 215 nucleotide neighbors )
M24177 Synthetic Bactenophage MI3 (clone M13.SV.B 12) SV40 early promoter region DNA gι|209416|gb|M24177|S YNS VB 12 [209416] (View GenBank report,FASTA report,ASN.1 report,Graphιcal view.l MEDLINE link, or 1 nucleotide neighbor )
M24176
Synthetic Bactenophage M13 (clone M13 SV.B 1 1) SV40 early promoter region DNA gι|209415|gb|M241761SYNS VB 11 [209415]
(View GenBank report.FASTA report-ASN. l report,Graphιcal vιew,l MEDLINE link, or 1 nucleotide neighbor )
M24175 Synthetic Bactenoohage M 13 (clone M 13 S V 8) S V40 earlv cromoter region DNA gι|208806|gb|M24175|SYNM13SV8 [208806] (View GenBank report.FASTA report.ASN 1 report.Graphical \ ιew 1 MEDLINE link, or 242 nucleotide neighbors )
M19979 Synthetic hybπds, recombinant DNA from bactenophage M13 ana plasmid ρHV33 gι|207813|gb|M19979|SYN33Ml3M [207813] (View GenBank report.FASTA report.ASN 1 report.GraDhical wew.l MEDLINE link, or 617 nucleotide neighbors )
Ml 9565
Synthetic hybπds, recombinant DNA from bactenophage M13 and plasmid pHV33 gι|207808|gb|M 19565|SYN33M 13H [207808]
(View GenBank report,FASTA report.ASN 1 report.Graphical \ ιew, l MEDLINE link, or 567 nucleotide neighbors )
M19564
Synthetic hybnds, recombinant DNA from bactenophage M13 and plasmid pHV33 gι|207807|gb|M19564|SYN33M13G [207807]
(View GenBank report,FASTA report-ASN 1 report, Graphical \ ιew,l MEDLINE link, or 12 nucleotide neighbors )
Ml 9563
Synthetic hybnds, recombinant DNA from bactenophage M13 and plasmid pHV33 gι|207806lgb|M19563|SYN33M13F [207806]
(View GenBank report.FASTA reρort,ASN 1 report.Graphical view.l MEDLINE link, or 262 nucleotide neighbors )
M19561
Synthetic hybnds, recombinant DNA from bactenophage M13 and plasmid pHV33 gι|207804|gb|M19561|SYN33M13D [207804]
(View GenBank report.FASTA report-ASN.1 report, Graphical \ lew, 1 MEDLINE link, or 27 nucleotide neighbors )
Ml 9560
Synthetic hybnds; recombinant DNA from bactenophage M13 and plasmid pHV33 gι|207803|gb|M 19560|S YN33M 13C [207803]
(View GenBank report.FASTA report.ASN 1 report.Graphical view, or 1 MEDLINE lin )
M19559
Syntheπc hybπds, recombinant DNA from bactenophage M13 and plasmid pHV33 gι|207802|gb|M 19559|SYN33M 13B [207802]
(View GenBank report,FASTA report-ASN 1 reporζGraphical view.l MEDLINE link, or 227 nucleotide neighbors )
M10568 Bactenophage M13 rephcaave form π, replicanon oπgin, specific nick location gι|215220|gb|M10568|M13OR_B [215220] (View GenBank report,FASTA rcporV-SN. l report, Graphical vαew.l MEDLINE link, or 650 nucleotide neighbors )
M10910 Bactenophage M13 gene II regulatory region and M13sjl mutant gι|215209|gb|M10910|M13IIREG [215209] (View GenBank report.FASTA report, ASN.1 report, Graphical viεw.l MEDLINE link, or 72 nucleotide neighbors )
M38295
Bactenophage Ml 3 HaeEII restriction fragment DNA gι|215208|gb|M38295[M13HAEIII [215208]
(View GenBank report,FASTA report,ASN 1 report, Graphical view, or 67 nucleotide neighbors )
E02067
DNA encoding a part of Bacteπophaee M13 tε 127 gι|217031 1 |dbj"|E02067|E02067 [217031 1 ]
(View GenBank report.FASTA repoπ ASN 1 report, or Graphical view)
J02467
Bactenophage MS2, complete genome gι|215232|gb|J02467|MS2CG [215232]
(View GenBank report.FASTA repor ASN 1 report.Graphical view, 8 MEDLINE links, 4 protem links, 20 nucleotide neighbors, or 1 genome link )
AJ004950
Bactenophage PI ban gene gι|3688226jemblAJ01 1592|BP 1011592 13688226]
(View GenBank report.FASTA report ASN 1 report.Graphical view, or 1 protem link )
U88974 Bactenophage PI structural lync transgr.cosylase (orf47), pep44b (orf44b), pep44a (orf44a), and pep43 (orf43) genes, complete eds; and pep42 (orf42) gene, partial eds gι|2661099|gb|AF035607|AF035607' [2661099] (View GenBank report,FASTA report.ASN 1 report.Graphical vιew,5 protem links, or 1 nucleotide neighbor )
AJ000741
Bactenophage P 1 darA operon gι|2462938|emb|AJ000741|BPAJ7641 [2462938]
(View GenBank report.FASTA report ASN 1 report.Graphical view. l MEDLINE link, 10 protem links, or 31 nucleotide neighbors
X01828
Bactenophage P 1 recombinase gene cm gι|15133|emb|X01828|MYPlCIN [15133]
(View GenBank report.FASTA repor ASN 1 report,Graphιcal view.l MEDLINE link, 1 protem link, or 3 nucleotide neighbors )
X98146
Bactenophage P I DNA sequence arounα the Oρ88 operator gι| 1359513|emb|X98146|BP10P880P [1359513]
(View GenBank report.FASTA report-ASN.l report, Graphical view, or 1 nucleotide neighbor )
S61175
_mml operon- ιcd=cell division represser, antl=ant_repressor (promoters
P51a, P51b} [bactenophage PI , Genomic, 728 nt] gι|385908|gb|S61 175|S611~75 [385908]
(View GenBank report.FASTA report-ASN.l report, Graphical view. l MEDLINE link, or 3 nucleotide neighbors )
X87824
Bactenophage P 1 gene 26 gι|861164|emb|X87824|XXBP 1G26 [861164]
(View GenBank report.FASTA reportASN.1 report,Graphical view, or 1 protem lin )
X15638
Phage P 1 DNA for lyric replicon contaι____g promoter P53 and two open readmg frames gι|15735|emb|X15638|PPlLREP [157351
(View GenBank report.FASTA reportASN 1 report,Graphical view. l MEDLINE link, 3 protein links, or 24 nucleotide neighbors
X17512
Bacteriophage PI DNA for immunity region imml gi| 15479|emb|X 17512|P1IMMUNIY [ 15479]
(View GenBank report.FASTA reportASN.1 report.Graphical view,2 MEDLINE links, or 4 nucleotide neighbors )
XI 6005
Bacteriophage PI el gene for Pic 1 repressor protein gi|15477|emb|X16005|PlCl [15477]
(View GenBank report.FASTA report.ASN.1 report,Graphical view.l MEDLINE link, 1 protein link, or 3 nucleotide neighbors )
X03453
Bacteriophage PI ere gene for recombinase protein gi|15135|emb|X03453|MYPlCRE [15135]
(View GenBank report.FASTA report.ASN.l report,Graphical view.l MEDLINE link, 2 protein links, or 12 nucleotide neighbors ',
X06561
Bacteriophage PI cl gene 5'-region gi|15128|emb|X06561|MYPlCl [15128]
(View GenBank reportFASTA reportASN.1 reportGraphical view, 1 MEDLINE link, 4 protein links, or 6 nucleotide neighbors )
V01534
Bacteriophage PI genome fragment (IS2 insertion spot). This regions contains four unidentified reading frames and is known as insertion hot spot for IS2 insertion sequences gi|15118|emb|V01534|MYOVPl [15118]
(View GenBank reportFASTA reportASN.1 report,Graphical view.l MEDLINE link, 4 protein links, or 3 nucleotide neighbors )
X56951
Bacteriophage PI gene 10 gi|406728|emb|X56951 |BPP1 GP 10 [406728]
(View GenBank reportFASTA report-ASN.l reportGraphical view,2 MEDLINE links, 3 protein links, or 1 nucleotide neighbor )
K02380
Bacteriophage PI replication region including repA, parA, and parB genes and incA, incB, and incC incompatibility determinants gi|215652|gb|K02380|PPlREP [215652]
(View GenBank report,FASTA report-ASN.l _eport,Graphical view,5 MEDLINE links, 4 protein links, or 8 nucleotide neighbors )
X87674
Bacteriophage PI lydA & lydB genes gi|974763|emb|X87674|BACPlLYD [974763]
(View GenBank report.FASTA report-ASN.l reportGraphical view.l MEDLINE link,- 2 protein links, or 2 nucleotide neighbors )
X87673
Bacteriophage PI gene 17 gi|974761|emb|X87673|BACPl 17 [974761]
(View GenBank reportFASTA report-ASN.l reρort,Graphical view.l MEDLINE link, 1 protein link, or 1 nucleotide neighbor )
M16618 Bacteriophage PI cl repressor binding sites gi|215600|gb|M16618|PPlCl [215600] (View GenBank reportFASTA reportASN.1 reportGraphical view.l MEDLINE link, 2 protein links, or 3 nucleotide neighbors )
SEG_PP1CIN Bactenophage P I cm gene encodmg recombmase, cixL recombination site, and 5' end of C mvertible element gι|215607|gb||SEG_PPl CIN [215607] (View GenBank report,FASTA report.ASN 1 report.Graphical view, 1 MEDLINE link, 1 protem Imk, or 4 nucleotide neighbors ) 03I73 Bactenophage PI C mvertible element πght end, and cixR recombmaπon site gι|215606igb|K03173|PPlCIN2 [215606] (View GenBank report.FASTA report-ASN 1 report, or Graphical view)
215605 Bacteriophage PI cm gene encodmg recombmase, cixL recombinanon site, and 5 end of C invertible element gi|2 l5605|lcl|X0I 828 [215605] (View GenBank report.FASTA report.ASN 1 report, or Graphical view)
M25470 Bactenophage PI tail fiber protem gene, complete eds gι|341349|gb|M25470|PPlTFPR [341349] (View GenBank report.FASTA report,ASN 1 reportGraphical view, 1 MEDLINE link, 3 protem links, or 3 nucleotide neighoors )
M34382 Bactenophage P 1 sun region proteins, complete eds gι]215661|gb|M34382|PPlSIM [215661] (View GenBank report.FASTA reportASN 1 reportGraphical view.l MEDLINE link, or 2 protem links )
M81 56
Bactenophage PI R protem (R) gene, complete eds gι|215658|gb|M81956|PPlRP [215658]
(View GenBank reportFASTA reportASN 1 reportGraphical vιew,l MEDLINE link, 2 protem links, or 4 nucleotide neighoors >
M37080 Bactenophage P 1 mini-Pi plasmid oπgin of replication gι|215657|gb|M37080|PPlREPOR [215657] (View GenBank reportFASTA report,ASN 1 report, Graphical view, 1 MEDLINE link, or 46 nucleotide neighbors )
M27041 Bactenophage PI ref gene, complete eds gι|215650|gb|M27041|PPlREF [215650] (View GenBank reportFASTA report-ASN 1 reportGraphical vιew,l MEDLINE link, 1 protem link, or 1 nucleonde neighbor )
L01408 Bactenophage PI pamπoπ protem (parB) gene, 3' end gι|215642|gb|L01408|PPlPARB [215642] (View GenBank reportFASTA report-ASN.l report,Graphιcal view.l protem link, or 41 nucleotide neighbors )
SEG_PP1PAR Bactenophage miniplasmid PI parA gene, 5' end gι|215639|gb||SEG_PPlPAR [215639] (View GenBank reportFASTA report-ASN 1 reportGraphical view. l MEDLINE Imk, 2 protem links, or 48 nucleonde neighbors )
M36425
Bactenophage mnuolasmid P I parB gene, 3' end gι|215638|gb|M36425|PPlPAR2 [215638]
(View GenBank report,FASTA reportASN 1 report, or Graphical view)
M36424 ~ ! 3
Bacteπopnage mmiplasmid PI parA gene, 5' end gι|215637igb|M36424|PPlPARl [215637] (View GenBank report.FASTA report,ASN 1 report, or Graphical view)
Ml 1129 Bactenophage P I mmiplasmid oπgm of replication region gι|215632lgb|Ml 1 129|PP10RIM [215632] (View GenBank report.FASTA report.ASN 1 reportGraphical view, 1 MEDLINE link, 1 protem link, or 43 nucleotide neighbors
M25414 Bactenophage PI c l repressor binding site, operator 88 (Op88) gi|215631|gb|M25414|PP10P88A [215631] (View GenBank reoort,FASTA report.ASN 1 report.Graphical view, 1 MEDLINE Imk, or 3 nucleotide neighbors )
M25413 Bactenophage PI cl repressor binding site, operator 68 (Op68) gι!215630|gb|M25413|PPlOP68A [215630] (View GenBank report.FASTA reportASN 1 reportGraphical view, or 1 MEDLINE link )
M25412 Bactenophage PI cl repressor binding site, operator 21 (Op21) gι|215629|gb|M25412|PP10P21A [215629] (View GenBank report.FASTA reρort,ASN 1 report.Graphical view, 1 MEDLINE Imk, or 1 nucleotide neighbor )
M 10510 Bactenophage PI recombination site loxR gι|215628|gb|M 10510|PP 1 LOXR [215628] (View GenBank reportFASTA reportASN 1 report, Graphical view.l MEDLINE link, or 1 nucleotide neighbor )
Ml 0287 Bactenophage P 1 loxP X loxP recombinanon site gι|215627|gb|Ml 0287|PP 1 LOXPX [215627] (View GenBank reportFASTA report-ASN 1 reportGraphical view. l MEDLINE link, or 13 nucleonde neighbors )
Ml 0494 Bactenophage PI recombination site loxP gι|215626|gb|M10494|PPlLOXP [215626] (View GenBank reportFASTA report-ASN.l report, Graphical view. l MEDLINE link, or 134 nucleonde neighbors )
M10511 Bactenophage PI recombinanon site loxL gι|215625|gb|M1051 1IPP1LOXL [215625] (View GenBank reportFASTA report-ASN 1 report,Graphical view.l MEDLINE link, or 1 nucleotide neighbor )
MI0512 Bactenophage P 1 recombination site loxB gι[215624|gb|Ml 0512|PP 1 LOXB [215624] (View GenBank reportFASTA report-ASN.l reportGraphical view, or 1 MEDLINE link )
M10145 Bactenophage PI eenome fragment with recombination site loxP gι|215623|gb|M10.45|PPlCREX [215623] (View GenBank reportFASTA reportASN 1 reportGraphical view. l MEDLINE link, or 21 nucleonde neighbors )
M13327 Bactenophage PI Cm recombmase acnvated cross over site, juncπon IV, clone pSHI326 gι|215622|gb|M13327|PP 1 CN26IV [215622] (View GenBank report.FASTA report.ASN 1 report.Graphical view, 1 MEDLINE link, or 7 nucleotide neighbors )
M13325 Bactenophage P 1 Cm recombmase acnvated cross over site, junction II, clone pSHI326 gι|215621|gb|M13325|PPlCN26II [215621] (View GenBank report.FASTA report.ASN 1 reportGraphical view.l MEDLINE link, or 1401 nucleonde neighbors )
M13323 Bactenophage PI Cm recombmase activated cross over site, junction TV, clone pSHI325 gι|215620|gb|M13323|PPlCN25IV [215620] (View GenBank report.FASTA report.ASN 1 report.Graphical view.l MEDLINE link, or 7 nucleonde neighbors )
M13321 Bactenophage PI Cm recombmase acnvated cross over site, juncnon II, clone pSHI325 gι|215619|gb|M 1332 HPP1CN25II [215619] (View GenBank report.FASTA reportASN 1 report.Graphical view.l MEDLINE link, or 1058 nucleotide neighbors )
MI3324 Bactenophage PI Cm recombmase acnvated cross over site, juncnon I, clone pSHI326 gι|215618|gb|M13324|PPlCIR26I [215618] (View GenBank reportFASTA report-ASN 1 report.Graphical view.l MEDLINE Imk, or 7 nucleonde neighbors )
M13319 Bactenophage PI Cm recombmase acnvated cross over site, πght juncnon, clone pSHI327 gι|215617|gb|M13319|PPlCIN27R [215617] (View GenBank report,FASTA reportASN 1 report.Graphical view, 1 MEDLINE link, or 7 nucleotide neighbors )
M13320 Bactenophage PI Cm recombmase acnvated cross over site, junction I, clone pSHI325 gι|215616|gb|M 13320|PP 1 CIN25I [215616] (View GenBank report.FASTA reportASN 1 reportGraphical view, 1 MEDLINE link, or 7 nucleonde neighbors )
M13318 Bactenophage PI Cm recombmase acnvated cross over site, left juncnon, clone pSHD24 gι|215615|gb|M13318|PPlCIN24L [215615] (View GenBank reportFASTA report-ASN 1 report, Graphical view, 1 MEDLINE link, or 1370 nucleonde neighbors )
M13317 Bactenophage PI Cm recombmase acnvated cross over site, πght juncnon, clone pSHI323 gι|215614|gb|M13317|PPlCIN23M [215614] (View GenBank reportFASTA reportASN 1 reportGraphical vιew.1 MEDLINE link, or 1055 nucleonde neighbors )
M13316 Bactenophage PI Cm recombmase activated cross over site, left junction, clone pSHI323 gι|215613|gb|M13316|PPlCIN23L [215613] (View GenBank reportFASTA report,ASN.l reportGraphical view.l MEDLINE link, or 7 nucleonde neighbors )
M13315 Bactenophage PI Cm recombmase acnvated cross over site, nght juncnon, clone pSHI322 gι|215612|gb|M13315|PP!CIN22R [215612] (View GenBank reportFASTA report,ASN 1 reportGraphical view, 1 MEDLINE link, or 7 nucleonde neighbors )
M13314 2 1 -
Bactenophage PI Cm recombmase acnvated cross over site, left juncnon, clone pSHI322 gι|21561 11gb|M13314|PPlCIN22L [215611] (View GenBank report.FASTA report.ASN 1 report.Graphical view. l MEDLINE Imk, or 1401 nucleotide neighbors )
M 13313 Bactenophage P I Cm recombmase acnvated cross over site, πght juncnon, clone pSHI321 gι|215610|gb|M13313|PPlCIN21R [215610] (View GenBank report.FASTA report-ASN. l report.Graphical view.l MEDLINE link, or 7 nucleonde neighbors )
M13312
Bactenophage PI Cm recombmase acnvated cross over site, left juncnon, clone pSHI321 gι|215609|gb|M13312|PP lCIN21L [215609]
(View GenBank report.FASTA report.ASN 1 report.Graphical view 1 MEDLINE link, or 1058 nucleotide neighbors )
Ml 6568 Bactenophage PI c4 repressor gene, complete eds gι|215603|gb|M16568|PP l C4 [215603] (View GenBank reportFASTA report-ASN 1 reportGraphical view. l MEDLINE link, 1 protem link, or 4 nucleotide neighbors )
M13326 Bactenophage PI Cm recombmase activated cross over site, juncnon III, clone pSHI326 gι|215602|gb|M13326|PP 1 C26m [215602] (View GenBank reportFASTA reportASN 1 reportGraphical view. l MEDLINE link, or 1192 nucleotide neighbors )
M 13322
Bactenophage PI Cm recombmase acnvated cross over sue, juncnon HI, clone pSHI325 gι|215601|gb|M13322|PPlC25m [215601]
(View GenBank reportFASTA report-ASN 1 report,Graphιcal view. l MEDLINE link, or 1231 nucleonde neighbors )
J05651
Bactenophage PI modulator protem (bof) gene, complete eds gι|215598|gb|J05651]PPlBOFYl [215598]
(View GenBank reportFASTA report-ASN.l report,Graphιcal view.l MEDLINE link, 1 protem link, or 3 nucleonde neighbors )
M33224 Bactenophage PI regulatory protem (bof) gene, complete eds gι|215596|gb|M33224|PPlBOFFO [215596] (View GenBank reportFASTA reportASN.1 reportGraphical view.l MEDLINE link, 1 protem Imk, or 3 nucleotide neighbors )
Ml 0288
E coli/bacteπophage PI loxR recombination site gι| 146647|gb|M10288|ECOLOXR [146647]
(View GenBank report,FASTA report SN. l report,Graphιcal view.l MEDLINE link, or 3 nucleonde neighbors )
M 10289 E.coli bacteπophage PI loxL recombination site gι| 146646|gb|M10289|ECOLOXL [146646] (View GenBank report,FASTA report-ASN.l report,Graphical view.l MEDLINE link, or 2 nucleotide neighbors )
M 10290
E coli loxB site, which can recombine with bactenophage PI loxP site gι| 146645|gb|M10290|ECOLOXB [146645]
(View GenBank report,FASTA report-ASN 1 reportGraphical view. l MEDLINE link, or 2 nucleotide neighbors )
M 10287
Bacteriophage P 1 loxP X loxP recombination site gi|215627|gb|M10287|PP l LOXPX [215627]
(View GenBank report.FASTA report.ASN.l report.Graphical view.l MEDLINE link, or 13 nucleotide neighbors )
M74046
Bacteriophage P 1 pacA and pacB genes, complete eds gi|215634|gb|M74046|PP 1 PACAB [215634]
(View GenBank report.FASTA report,ASN.l report.Graphical view.l MEDLINE link, or 2 protein links )
M95666 Bacteriophage P 1 gene 10, doc and phd genes, complete eds gi|463276|gb|M95666|PPlPHDDOC [463276] (View GenBank report.FASTA report.ASN. l report.Graphical view,2 MEDLINE links, 4 protein links, or 1 nucleotide neighbor )
M25604 Bacteriophage Q-beta mutated autonomously replicating sequence MDV1 RNA fragment gi|556359|gb|M25604|PQBARSMUT [556359] (View GenBank report.FASTA report,ASN.l reportGraphical view, l MEDLINE link, or 8 nucleotide neighbors )
V00643 fust half of the phage Q-beta gene for coat protein gi| 15088|emb|V00643|LEQBET [15088]
(View GenBank reportFASTA report-ASN.1 reportGraphical view, 1 MEDLINE link, 1 protein link, or 4 nucleotide neighbors )
M25167
Bacteriophage Q-beta RNA fragment recovered from replicase binding complex gi|556362|gb|M25167|PQBREPLICB [556362]
(View GenBank report.FASTA reportASN.1 reportGraphical view, 1 MEDLINE link, or 2 nucleotide neighbors )
M24876
Bacteriophage Q-beta replicase RNA, 5' end gi|556360|gb|M24876|PQBREPLICA [556360]
(View GenBank report.FASTA report-ASN.l reportGraphical view. l MEDLINE link, 1 protein link, or 4 nucleotide neighbors )
M25444
Synthetic bacteriophage Q-beta DNA gi|209118|gb|M25444|SYNPQBTERM [209118]
(View GenBank report.FASTA report-ASN.1 report,Graphical view, 1 MEDLINE link, or 8 nucleotide neighbors )
M25463
Bacteriophage Q-beta self-replicating micTovariant (+)'RNA gi|532489|gb|M25463|PQBMVSRRNA [532489]
(View GenBank reportFASTA report^ASN.1 reportGraphical view, or 1 MEDLINE link )
M25014 Bateriophage Q-beta RNA replicase gene, 5'end, and maturation protein gene, 3' end gi|294316|gb|M25014|PQBREPLC [294316] (View GenBank reportFASTA report-ASN. l reportGraphical view.l MEDLINE link, 2 protein links, or 2 nucleotide neighbors }
M25065
Bacteriophage Q-beta RNA sequence wi± putative stem loop gi|294315|gb|M25065|PQBLOOP [294315]
(View GenBank reportFASTA report-ASN. l reportGraphical view,l MEDLINE link, or 3 nucleotide neighbors)
Ml 0265
Bacteriophage Q-beta RNA molecule with the ability to replicate extracellularly gi|215726|gb|M 10265|PQBRNA [215726]
(View GenBank report.FASTA report.ASN.l reportGraphical view. l MEDLINE link, or 8 nucleotide neighbors )
M24815 Bacteriophage Q-beta specified replicase subunit RNA, gi|215725|gb|M24815|PQBREPL [215725] (View GenBank report.FASTA report.ASN. l report.Graphical view. l MEDLINE link, or 4 nucleotide neighbors )
M25461
Bacteriophage Q-beta plus-strand RNA, 5' terminus gi|215724|gb|M25461|PQBPS5E [215724]
(View GenBank report.FASTA report.ASN.l report, or Graphical view)
M25462 Bacteriophage Q-beta plus-strand RNA, 3' terminus gi|215723|gb|M25462|PQBPS3E [215723] (View GenBank report.FASTA report-ASN.l reportGraphical view, or 8 nucleotide neighbors )
M24871 Bacteriophage Q-beta nanovariant WSIII RNA gi|215722|gb|M24871 |PQBNVWSIC [215722] (View GenBank reportFASTA reportASN.1 reportGraphical view, 1 MEDLINE link, or 2 nucleotide neighbors )
M24870 Bacteriophage Q-beta nanovariant WSII RNA gi|2I5721|gb|M24870|PQBNVWSIB [215721] (View GenBank reportFASTA reportASN.1 reportGraphical view, 1 MEDLINE link, or 2 nucleotide neighbors )
M24869
Bacteriophage Q-beta nanovariant SI RNA gi|2 I5720|gb|M24869|PQBNVWSIA [215720]
(View GenBank report.FASTA report-ASN.l report,Graphical view.l MEDLINE link, or 2 nucleotide neighbors )
Ml 0495
Coliphage Q-beta MDV-1(+) RNA gi|215719|gb|M10495|PQBMDVlA [215719]
(View GenBank reportFASTA report-ASN.l report,Graphical view. l MEDLINE link, or 10 nucleotide neighbors )
J02484 bacteriophage qbeta coat protein cistron first half gi|215717|gb|J02484|PQBCP5 [215717]
(View GenBank reportFASTA reportASN.1 report,Graphical view,l MEDLINE link, 1 protein link, or 4 nucleotide neighbors )
M57754
Bacteriophage Q-beta minus strand RNA, 5' terminus gi|215716|gb|M57754|PQBBMS5E [215716]
(View GenBank report,FASTA reportASN.1 reportGraphical view, or 8 nucleotide neighbors )
M24297
Bacteriophage Q-beta 5'-terminal region of the minus strand gi|215715|gb|M24297|PQB5END [215715]
(View GenBank report.FASTA report-ASN.l reportGraphical view. l MEDLINE link, or 8 nucleotide neighbors )
M 10695 21 8
Bacteriophage Q-beta, MDV- 1 RNA gι|215714|gb|M 10695|PQB lIR [215714] (View GenBank report.FASTA reportASN.1 report.Graphical view,2 MEDLINE links, or 12 nucleotide neighbors )
M24827 Bacteriophage Rl 7 A protein gene, 5' end gi|216078|gb|M24827|R 17RNACIS [216078] (View GenBank report.FASTA reportASN.1 reportGraphical view.l MEDLINE link, or 5 nucleotide neighbors )
M24829 Bacteriophage R17 coat protein gene, 5' end gi|216075|gb|M24829|R17CP5 [216075] (View GenBank report.FASTA report.ASN. l report.Graphical view.l MEDLINE link, or 5 nucleotide neighbors )
J02488 bacteriophage rl7 ma synthetase initiation site gi|216080|gb[ J02488|R17RNASYN [216080]
(View GenBank report.FASTA reportASN.1 reportGraphical view,3 MEDLINE links, 2 protein links, or 6 nucleotide neighbors )
J02487 bacteriophage rl7 coat protein initiation site gi|216073|gb|J 02487|R17COATP [216073]
(View GenBank report.FASTA report-ASN.1 reportGraphical view, or 1 MEDLINE link )
J02486 bacteriophage rl7 a protein initiation site gi|216071 |gb| J02486|R17APROT [216071 ] (View GenBank report.FASTA report-ASN.1 reportGraphical view, or 1 MEDLINE link )
M24826 Bacteriophage R17 coat protein RNA fragment gi|216077|gb|M24826|Rl 7CPRAA [216077] (View GenBank report.FASTA report-ASN.1 reportGraphical view, 1 MEDLINE link, or 7 nucleotide neighbors )
M24296
Bacteriophage R17 3'-terminal fragment A RNA gi|216070|gb|M24296|R173TFA [216070]
(View GenBank reportFASTA report-ASN.l reportGraphical view, l MEDLINE link, or 9 nucleotide neighbors )
1TFN structure refinement for a 24-nucleotide ma hairpin, nmr, minimized average stπicture ribonucleic acid, hairpin, bacteriophage rl7 mol_id: 1; molecule: rl7c; chain: null; engineered: yes gi|1942336|pdb|lTFN| [1942336] (View GenBank rcport,FASTA report-ASN.l report,Graphical view, or 1 structure link )
1RPEA ma (5'-d(gpgpgpapcpupgpapcpgpapupcpapcpgp cpapgpupcpupapu-3') (24-mer ma hairpin coat protein binding site for bacteriophage rl7) (nmr, minimized average structure) gi| 1421020|pdb[ 1 RHT| [1421020] (View GenBank reportFASTA report-ASN.l report-Graphical view, or 1 stπicture lin )
M 14428
Bacteriophage S 13 circular DNA, complete genome gi|216089|gb|M14428|S13CG [216089]
(View GenBank report.FASTA report,ASN. l report.Graphical view,2 MEDLINE links, 12 protein links, 26 nucleotide neighbors, or 1 genome link )
J05393
Bacteriophage TI DNA N-6-adenine-methyltransferase (M.T1) gene, complete cd≤ gij l66163|gb|J05393|BTlNAMTA [166163]
(View GenBank report.FASTA report.ASN.l report.Graphical view. l MEDLINE link, or 2 protein links )
L46845 Bacteriophage T2 frd3, frd2 genes, comnlete eds gij951387|gb|L46845|PT2FRD32G [951387] (View GenBank report.FASTA report.ASN. l report.Graphical view,2 protein links, or 17 nucleotide neighbors )
L43611 Bacteriophage T2 fibritin (wac) gene, complete eds gi|903869|gb|L43611|PT2WAC [903869] (View GenBank report.FASTA report,ASN. l reportGraphical view.l protein link, or 4 nucleotide neighbors )
M24812 Bacteriophage T2 secondary structure RNA sequence gi|215796|gb|M24812|PT2RNA [215796] (View GenBank reportFASTA report-ASN.1 reportGraphical view,l MEDLINE link, or 4 nucleotide neighbors )
M22342
Bacteriphage T2 DNA-(adenine-N6)methyltransferase (dam) gene, complete eds gi!215792|gb|M22342|PT2DAM [215792]
(View GenBank reportFASTA reportASN.1 reportGraphical view,l MEDLINE link, 1 protein link, or 2 nucleotide neighbors )
S57515 orf61.2 {intergenic region between 41 and δl } [bacteriophage T2, Genomic, 323 nt] gi|298524|gb|S57515|S57515 [298524] (View GenBank reportFASTA reportASN.1 reportGraphical view,l MEDLINE link, or 1 protein link )
X05312 Bacteriophage T2 gene 38 for receptor recognizing protein gi| !5197jemb|X05312|MYT2G38 [15197] (View GenBank reportFASTA reportASN.1 reportGraphical view.l MEDLINE link, or 1 protein link )
X04442
Bacteriophage T2 gene 37 for receptor recognizing protein gi|15195iemb|X04442|MYT2G37 [15195]
(View GenBank report.FASTA report-ASN.l report,Graphical view. l MEDLINE link, or 1 protein link )
X12460
Bacteriophage T2 gene 32 mRNA for single-stranded DNA binding protein gill5192|emb|X12460|MYT2G32 [15192]
(View GenBank reportFASTA report-ASN.l report,Graphical view. l MEDLINE link, 2 protein links, or 14 nucleotide neighbors )
X57797 Bacteriophage T2 gene for gpl2 gi|14875|emb|X56555|BT2GP12 [14875] (View GenBank report,FASTA report,ASN.l report,Graphical view. l protem link, or 2 nucleotide neighbors )
X01755 Bacteriophage T2 tail fiber gene 36 gij 15189lemb|X01755|MYT2F36 [15189] (View GenBank report.FASTA report.ASN.l report.Graphical view.l MEDLINE link, 2 protein links, or 1 nucleotide neighbor ) 14784 Bacteriophage T3 strain amNG220B right end, tail fiber protein, lysis protein and DNA packaging proteins, complete eds gi|215810|gb|M14784|PT3RE [215810] (View GenBank reportFASTA reportASN.1 report.Graphical view.l fvIEDLINE link, 9 protein links, or 10 nucleotide neighbors )
SEG_PT3RNAPOL Bacteriophage T3 RNA polymerase III gene, 5' end gi|710559|gb||SEG_PT3RNAPOL [710559] (View GenBank report.FASTA reportASN.1 report.Graphical view, 1 MEDLINE link, 2 protein links, or 2 nucleotide neighbors )
M22610 Bacteriophage T3 RNA polymerase III gene, 3' end gi|340722|gb|M22610|PT3RNAPOL2 [340722] (View GenBank report,FASTA report,ASN.1 report, or Graphical view)
M22609 Bacteriophage T3 RNA polymerase III gene, 5' end gi|340721 |gb|M22609|PT3RNAPOL 1 [340721 ] (View GenBank report,FASTA reportASN.1 report, or Graphical view)
X05031 Bacteriophage T3 gene region 1-2.5 with primary origin of replication gi|15719|emb|X05031|POT3ORI [15719] (View GenBank reportFASTA report-ASN.l report.Graphical view.l MEDLINE link, 11 protein links, or 5 nucleotide neighbors )
X03964 Bacteriophage T3 early control region pos. 308-810 from genome left end gi|15718!emb|X03964|POT3EP [15718] (View GenBank reportFASTA report-ASN.l report, Graphical view,2 MEDLINE links, or 20 nucleotide neighbors )
XI 7255 Bacteriophage T3 gene 1 to gene 11 gi|15682|emb|X17255|POT3111G [15682]
(View GenBank reportFASTA report-ASN.l report,Graphical view,4 MEDLINE links, 36 protein links, 17 nucleotide neighbors, or 1 genome link )
X15840 Phage T3 gene 10 gi|15625|emb|X15840|PODT3G10 [15625] (View GenBank reportFASTA report-ASN.l report, Graphical view.l MEDLINE link, or 3 nucleotide neighbors )
X02981 Bacteriophage T3 gene 1 for RNA polymerase gi|15561|emb|X02981|PODOT3P [15561] (View GenBank reportFASTA report-ASN.l report, Graphical view.l MEDLINE link, 1 protein link, or 3 nucleotide neighbors )
J02503 bacteriophage t3 5' end, terminally redundant sequence (trs) gi|215816|gb|J02503|PT3TRSl [215816] (View GenBank reportFASTA report-ASN.l report, or Graphical view)
SEG_PT3TRS bacteriophage t3 5' end, terminally redundant sequence (trs) gi|215818|gb||SEG_PT3TRS [215818]
(View GenBank report.FASTA report.ASN.l report.Graphical view, or 1 MEDLINE link )
J02504 bacteriophage t3 3' end, terminally redundant sequence (trs) gi|215817|gb|J02504|PT3TRS2 [215817] (View GenBank report.FASTA report.ASN.l report, or Graphical view}
H YPERLINK htφ://www.rs.noda.sut.ac.jp/~kunisawa h t tp://www.rs.noda.sut.ac.jp/-kunisawa Bacteriophage T4 genomic database compiled by Arisaka et al.
X95646 Bacteriophage T5 DNA for region 60.5%-71% of the T5 genome gi|2791557|emb|AJ001191|BTJ001191 [2791557] (View GenBank reportFASTA report,ASN.l report.Graphical view,7 MEDLINE links, 12 protein links, or 6 nucleotide neighbors )
X56847 Bacteriophage T5 genomic region encoding early genes D10-D15 gii l5407|emb|X12930|MYT5D10 [15407] (View GenBank report.FASTA report.ASN.1 reportGraphical view,l MEDLINE link, 5 protein links, or 4 nucleonde neighbors )
AF039886 Bacteriophage T5 subclone T5.5.3r5.18r, single pass sequence, genomic survey sequence gij2811154|gb|AF039886|AF039886 [2811154] (View GenBank report,FASTA report-ASN.l report, or Graphical view)
AF039885
Bacteriophage T5 subclone T5.40f,41f, single pass sequence, genomic survey sequence gi|2811153|gb|AF039885|AF039885 [2811153]
(View GenBank report.FASTA report-ASN.l report, or Graphical view)
AF039884 Bacteriophage T5 subclone T5.26.fr, single pass sequence, genomic survey sequence gi|2811152|gb|AF039884|AF039884 [2811152] (View GenBank report.FASTA report,ASN.l report, or Graphical view)
AF039883
Bacteriophage T5 subclone 10-T5.5.7F, single pass sequence, genomic survey sequence gi|2811151|gb|AF039883|AF039883 [2811151]
(View GenBank report-FASTA report-ASN.l report, or Graphical view)
AF039882 Bacteriophage T5 subclone 41-T5.5.4BF, single pass sequence, genomic survey sequence gi|2811150|gb|AF039882|AF039882 [2811150] (View GenBank reportFASTA report,ASN.1 report, or Graphical view)
AF039881
Bacteriophage T5 subclone 39-T5.5.4aF, single pass sequence, genomic survey sequence gi|2811149|gb|AF039881|AF039881 [2811149]
(View GenBank report.FASTA report,ASN.1 report, Graphical view, or 1 nucleotide neighbor )
AF039880 Bacteriophage T5 subclone 19-T5.7.2r, single pass sequence, genomic survey sequence gi|2811148|gb|AF039880|AF039880 [28i π48] (View GenBank report.FASTA report.ASN.l report, or Graphical view)
AF039879 Bacteriophage T5 subclone 18-T5.7.2F, single pass sequence, genomic survey sequence gi|2811147|gb|AF039879|AF039879 [2811 f47] (View GenBank report.FASTA report.ASN.1 report, or Graphical view)
AF039878 Bacteriophage T5 subclone 11-T5.5.7R, sinele pass sequence, genomic survey sequence gi|2811146|gb|AF039878|AF039878 [2811146]
(View GenBank report.FASTA report.ASN.l report.Graphical view, or 2 nucleotide neighbors )
AF039877 Bacteriophage T5 subclone T5.4FR, single pass sequence, genomic survey sequence gi|2811145|gb|AF039877|AF039877 [2811145] (View GenBank reportFASTA reporζASN.1 report, or Graphical view)
AF039876
Bacteriophage T5 subclone 22-T5.16R, single pass sequence, genomic survey sequence gi|2811144|gb|AF039876|AF039876 [2811144]
(View GenBank reportFASTA reportASN.1 report or Graphical view)
AF039875 Bacteriophage T5 subclone 21-T5.16R, single pass sequence, genomic survey sequence gi|2811143|gb|AF039875|AF039875 [2811143] (View GenBank reportFASTA reportASN.1 report, or Graphical view)
AF039874
Bacteriophage T5 subclone 21-T5.16F, single pass sequence, genomic survey sequence gi|2811 !42|gb|AF039874|AF039874 [2811142]
(View GenBank reρort,FASTA report,ASN.1 report, or Graphical view)
AF039873 Bacteriophage T5 subclone 09-T5.6F, single nass sequence, genomic survey sequence gi|2811141|gb|AF039873|AF039873 [2811141] (View GenBank reportFASTA report-ASN.1 report, or Graphical view)
AF039872
Bacteriophage T5 subclone 09-T5.6R, single pass sequence, genomic survey sequence gi|2811140|gb|AF039872|AF039872 [2811140]
(View GenBank report,FASTA reportASN.1 reρort,Graphical view, or 2 nucleotide neighbors )
AF039871
Bacteriophage T5 subclone 04-T5.26.R, single pass sequence, genomic survey sequence gi|2811139|gb|AF039871|AF039871 [2811139]
(View GenBank reportFASTA report-ASN.1 report, or Graphical view)
AF039870 Bacteriophage T5 subclone 13-T5.42F, single pass sequence, genomic survey sequence gi|2811138|gb|AF039870|AF039870 [2811138] (View GenBank report,FASTA report-ASN.l report, or Graphical view)
X69460 Bacteriophage T5 lrf gene for L-shaped tail fibers gi|15415|emb|X69460|MYT5LTF [15415] (View GenBank report.FASTA report,ASN.l report.Graphical view,2 MEDLINE links, 1 protein link, or 4 nucleotide neighbors )
X03402
Bacteriophage T5 D15 gene for 5' exonuclease gi|15413|emb|X03402|MYT5EXOG [15413]
(View GenBank report.FASTA report.ASN.l report,Graphical view.l MEDLINE link, 1 protein link, or 2 nucleotide neighbors )
Z11972
Bacteriophage T5 tRNA-Tyr, tRNA-Glu, tRNA-Trp, tRNA-Phe, tRNA-Cys and tRNA-Asn genes, and ORFs 91aa, 90aa, 42aa and 172aa gi|15795|emb|Zl 1972|T56TRNAG [15795]
(View GenBank report.FASTA report,ASN.1 report.Graphical view, 1 MEDLINE link, 4 protein links, or 3 nucleotide neighbors )
X03898
Bacteriophage T5 genes for tRNA-His, -Ser and -Leu gi|15786iemb|X03898|STT5RNl [15786]
(View GenBank report.FASTA reportASN.1 report.Graphical view, or 2 MEDLINE links )
X04177 Bacteriophage T5 gene for transfer RNA-Gln gi|15421|emb|X04177|MYT5TRNQ [15421] (View GenBank report.FASTA report-ASN.1 reportGraphical view, 1 MEDLINE link, or 2 nucleotide neighbors )
X03899
Bacteriophage T5 genes for tRNA-Val, -Lys, -fMet, -Pro and -ϋe3 gi|15787|emb|X03899|STT5RN2 [15787]
(View GenBank reportFASTA reρort,ASN.1 report,Graphical view, or 1 MEDLINE link )
X03798
Bacteriophage T5 gene for tRNA-Asp (GUC) gi|15472|emb|X03798|NCT5TRDG [15472]
(View GenBank reportFASTA report,ASN.1 report,Graphical view, 1 MEDLINE link, 2 protem links, or 2 nucleotide neighbors )
Y00364
Bacteriophage T5 tRNA gene cluster (27.8%-22.4%) gi|15420|emb|Y00364|MYT5TRN [15420]
(View GenBank reportFASTA report-ASN.l reρort,Graphical view.l MEDLINE link, or 13 nucleotide neighbors )
X03140
Bacteriophage T5 DNA with rho-dependent transcription terminator (Hind HI-P fragment) gi|15417|emb|X03140|MYT5RHO [15417]
(View GenBank report,FASTA report-ASN.l _eport,Graphical view, 1 MEDLINE link, 2 protein links, or 2 nucleotide neighbors )
Z35070
Bacteriophage T6 DNA gi|535228|emb|Z35074IMYEREGBT6 [535228]
(View GenBank reportFASTA report-ASN.l reportGraphical view.l MEDLINE link, or 1 protein link )
AF060870 Colφhage T6 small subunit distal tail fiber (gene 36) gene, partial eds, and large subunit distal tail fiber (gene 37) and tail fiber adhesin (gene 38) genes, complete eds gι|3676458|gb|AF052605|AF052605 [3676458] (View GenBank report.FASTA reportASN 1 report.Graphical v lew ,3 protem links, or 2 nucleonde neighbors )
Z35072 Bactenophage T6 DNA encodmg ORF19 1 gene and gl9 gene gι|535232|emb|Z35072|MYTAILT6 [535232] (View GenBank report.FASTA reportASN 1 reportGraphical view.l MEDLINE link, or 2 protem links )
X12488 Bactenophage T6 gene 32 mRNA for single-stranded DNA binding protem gι|15843|emb|X12488|MYT6G32 [15843] (View GenBank reportFASTA report-ASN 1 report.Graphical view, 1 MEDLINE link, 1 protem link, or 14 nucleonde neighbors )
Z78095 Bactenophage T6 DNA (1506 bp) gι|1488562|emb|Z78095|BPHZ78095 [1488562] (View GenBank reportFASTA reportASN 1 report.Graphical iew.l protem link, or 4 nucleonde neighbors )
Z35079 Bactenophage T6 DNA for Ip5, Ip6 gι|535215|emb|Z35079|MY57BT6 [535215] (View GenBank report.FASTA reportASN 1 reportGraphical v ie , 1 MEDLINE link, 2 protem links, or 1 nucleonde neighbor )
X68725 E coli bactenophage T6 gene for beta-glucosyl-HMC-alpha-glucos l-transferase gι|296439|emb|X68725|ECT6 [296439] (View GenBank report.FASTA report,ASN 1 reportGraphical v lew, 1 MEDLINE link, 3 protem links, or 1 nucleonde neighbor )
X69894
Bactenophage T6 alt gene for ADP-Rϊbosyltransferase gι|15422|emb|X69894|MYT6ADP [15422]
(View GenBank reportFASTA report,ASN 1 report, Graphical view, 1 MEDLINE link, 1 protem link, or 1 nucleonde neighbor )
L46846 Bactenophage T6 frd3, frd2 genes, complete eds gι|951390|gb|L468461PT6FRD32G [951390] (View GenBank reportFASTA report-ASN 1 reportGraphical view, or 2 protem links )
M27738 Bactenophage T6 translaQonal repressor protem (regA), complete eds gι|215993|gb|M27738|PT6REGA [215993] (View GenBank reportFASTA report-ASN.l reportGraphical vιew,l MEDLINE link, 1 protem link, or 5 nucleotide neighbors )
M38465 Bactenophage T6 DNA ligase gene, complete eds gι|21599 l|gb|M38465|PT6LIG55 [215991] (View GenBank reportFASTA report-ASN 1 reportGraphical view.l MEDLINE link, 1 protem link, or 2 nucleotide neighbors )
V01146 Genome of bactenophage T7 gι|431187|emb| V01 146|T7CG [4 1187]
(View GenBank report.FASTA report.ASN 1 report.Graphical view, 13 MEDLINE links, 60 protem Imks, 105 nucleotide neighbors, or 1 genome link )
X60322 Bactenophage alpha3 genes A, B, K, C, D, E, J, F, G, H gι|14775|emb|X60322|BACALPHA [14775]
(View GenBank reportFASTA report.ASN 1 rep on, Graphical view.l MEDLINE link, 10 protem links, 22 nucleonde neighbors, or 1 genome link )
X13332 Bactenophage alpha3 DNA for ongm of replication gι|15093|emb|X13332|MIA3ORPL [15093] (View GenBank report.FASTA report.ASN 1 reportGraphical view, or 1 MEDLINE link )
X12611
Bactenophage alpha3 gene for protem A part, finger domain gι| 15092|emb|X 12611 |MIA3 AFIN [15092]
(View GenBank report.FASTA report.ASN 1 reportGraphical view.l MEDLINE link, 1 protem link, or 6 nucleonde neighbors )
X15721
Bactenophage alpha3 deletion mutation DNA for the ongm region (-on) of replication gι|14774|emb|X 15721 |B A3DMOR9 [14774]
(View GenBank report.FASTA reportASN 1 report.Graphical view.l MEDLINE link, or 11 nucleonde neighbors )
XI5720
Bactenophage alpha3 deletion mutant DNA for the ongm region (-on) of replication gι|14773|emb|X15720|BA3DMOR8 [14773]
(View GenBank reportFASTA report,ASN 1 reportGraphical vιew,l MEDLINE link, or 1 nucleonde neighbor )
X15719
Bactenophage alpha3 insertion mutant DNA for the ongm region (-on) of replication gι|14772|emb|X15719|BA3DMOR7 [14772]
(View GenBank reportFASTA reportASN 1 reportGraphical view.l MEDLINE link, or 10 nucleonde neighbors )
X15718
Bactenophage alpha3 deletion mutation DNA for ongm region (-on) of replication gι|14771|emb|X15718|BA3DMOR6 [14771]
(View GenBank report,FASTA report-ASN 1 reportGraphical vιew,l MEDLINE link, or 11 nucleonde neighbors )
X15717
Bactenophage alpha3 deletion mutatαt DNA for ongm region (-on) of replication gι|14770|emb|X15717|BA3DMOR5 [14770]
(View GenBank reportFASTA report-ASN 1 reportGraphical vιew,l MEDLINE link, or 9 nucleotide neighbors )
X15716 Bactenophage alpha3 deletion mutant DNA for ongm region (-on) of replication gι|14769|emb|X15716|BA3DMOR4 [14769] (View GenBank report.FASTA report,ASN 1 reportGraphical vιew,l MEDLINE link, or 10 nucleonde neighbors )
X15715
Bacteriophage alpha3 deletion mutant DNA for origin region (-ori) of of replication gi|14768|emb|X15715|BA3DMOR3 [14768]
(View GenBank report.FASTA report.ASN.l report,Graphical view.l MEDLINE link, or 11 nucleotide neighbors )
X15714
Bacteriophage alpha3 deletion mutant DNA for origin region (-ori) of replication gi| 14767|emb|X 15714|BA3DMOR2 [ 14767]
(View GenBank report.FASTA reportASN.1 report,Graphical view, 1 MEDLINE link, or 11 nucleotide neighbors )
X15713
Bacteriophage alpha3 deletion mutant DNA for the origin region (-ori) of replication gi|14766|emb|X15713|BA3DMORl [14766]
(View GenBank report.FASTA report.ASN.l report.Graphical view. l MEDLINE link, or 11 nucleotide neighbors )
X62059
Bacteriophage alpha3 origin of cDNA synthesis (oriGA) gi|14763|emb|X62059|AL3ORIGA [14763]
(View GenBank reportFASTA reportASN.1 reportGraphical view.l MEDLINE link, or 13 nucleotide neighbors )
X62058
Bacteriophage alpha3 origin of cDNA synthesis (oriAA) gi|14762|emb|X62058|AL3ORIAA [14762]
(View GenBank report.FASTA reportASN.1 report,Graphical view.l MEDLINE link, or 13 nucleotide neighbors )
J02444
Bacteriophage alpha3 origin of DNA replication gi|166103|gb|J02444|AL3ORI [166103]
(View GenBank reportFASTA report-ASN.l report,Graphical view.l MEDLINE link, 2 protein links, or 12 nucleotide neighbors )
M25640
Bacteriophage alpha-3 H protein gene, complete eds gi|166101|gb|M25640|AL3HP [166101]
(View GenBank report,FASTA report-ASN.l report,Graphical view.l MEDLINE link, 1 protein link, or 13 nucleotide neighbors )
M10631
Bacteriophage alpha-3 cleavage site for phage phi-X174 gene A protein gij 166099|gb|M 106311 AL3 CS A [ 166099]
(View GenBank reportFASTA repoιV.SN.1 report,Graphical view.l MEDLINE link, 1 protein link, or 3 nucleotide neighbors )
X00774
Bacteriophage alpha-3 gene J sequence gi|15431|emb|X00774|NCBA J [15431]
(View GenBank report,FASTA report-ASN.l report,Graphical view.l MEDLINE link, 3 protein links, or 2 nucleotide neighbors )
M25640
Bacteriophage alpha-3 H protein gene, complete eds gi|166101|gb|M25640|AL3HP [166101]
(View GenBank report.FASTA reportASN.1 report,Graphical view.l MEDLINE link, 1 protein link, or 13 nucleotide neighbors )
M10631
Bacteriophage alpha-3 cleavage site for phage phi-X174 gene A protein gi|l 66099|gb|M 106311 AL3 CS A [ 166099]
(View GenBank reportFASTA report-ASN.l report-Graphical view.l MEDLINE link, 1 protein link, or 3 nucleotide neighbors )
T77
J02459
Bacteriophage lambda, complete genome gi|215104|gb|J02459|LAMCG [215104]
(View GenBank report.FASTA report.ASN.1 report.Graphical view,87 MEDLINE links, 67 protein links, 190 nucleotide neighbors, or 1 genome link )
J02482
Bacteriophage phi-X174, complete genome gi|216019[gb| J02482|PX 1 CG [216019]
(View GenBank report.FASTA reportASN.1 report.Graphical view,23' EDLINE links, 11 protein links, 26 nucleotide neighbors, or 1 genome link )
J02454
Bacteriophage G4, complete genome gi|215415|gb|J02454|PG4CG [215415]
(View GenBank report.FASTA report,ASN.1 report.Graphical view,6 MEDLINE links, 1 1 protein links, 20 nucleotide neighbors. or 1 genome link )
X60323
Bacteriophage phiK complete genome gi|14781 18|emb|X60323|BPHIKCG [1478118]
(View GenBank reportFASTA report-ASN.l reportGraphical view, 10 protein links, 18 nucleotide neighbors, or 1 genome link )
L42820
Bacteriophage BF23 tail protein (hrs) gene, complete eds gi| 1048680|gb|L42820|BBFHRS [1048680]
(View GenBank report.FASTA report,ASN.1 reportGraphical view, 1 MEDLINE link, 1 protein link, or 1 nucleotide neighbor )
X54455 Bacteriophage BF23 gene 17 and gene 18 gi|14797|emb|X54455|BF231718G [14797] (View GenBank reportFASTA report-ASN.l report,Graphical view,2 protein links, or 2 nucleotide neighbors )
M37097
Bacteriophage BF23 DNA, right end of terminal repetition gi|166115|gb|M37097|BBFRIGH [166115]
(View GenBank report.FASTA report-ASN.l reportGraphical view.l MEDLINE link, or 2 nucleotide neighbors )
M37096 Bacteriophage BF23 DNA, left end of terminal repetition gi|166114|gb|M37096|BBFLEFT [166114] (View GenBank reportFASTA report-ASN.l report,Grapr_ical view.l MEDLINE link, or 1 nucleotide neighbor )
M37095
Bacteriophage BF23 A2-A3 gene, complete eds, and Al gene, 5' end gi|166110|gb|M37095|BBFA2A3 [166110]
(View GenBank reportFASTA report.ASN.1 reportGraphical view,2 MEDLINE links, 3 protein links, or 1 nucleotide neighbor )
AF056281
Bacteriophage BF23 clone bf23.mac5/6.1, genomic survey sequence gi|3090930|gb|AF056281|AF056281 [3090930]
(View GenBank report,FASTA report-ASN.l report, or Graphical view)
AF056280 Bacteriophage BF23 clone bf23.mac3, genomic survey sequence gi|3090929|gb|AF056280|AF056280 [3090929] (View GenBank reρort,FASTA report,ASN.l report, or Graphical view)
AF056279 Bacteriophage BF23 clone bf23.macl8/21.34, genomic survey sequence gi!3090928|gb|AF056279|AF056279 [3090928] (View GenBank report.FASTA reportASN.1 report, or Graphical view)*
AF056278
Bacteriophage BF23 clone bf23.macl6/19.33, genomic survey sequence gi;3090927|gb|AF056278|AF056278 [3090927]
(View GenBank report.FASTA report.ASN. l report, or Graphical view)
AF056277 Bacteriophage BF23 clone bf23.mac 16/19-33, genomic survey sequence gi|3090926|gb|AF056277|AF056277 [3090926] (View GenBank reportFASTA reportASN.1 report, or Graphical view)
AF056276 Bacteriophage BF23 clone bf23.macl2/9-9, genomic survey sequence gi|3090925|gb|AF056276|AF056276 [3090925] (View GenBank report.FASTA reportASN.1 report, or Graphical view)
AF056275 Bacteriophage BF23 clone bf23.macl 1/14-24, genomic survey sequence gi|3090924|gb|AF056275|AF056275 [3090924] (View GenBank reportFASTA report-ASN.l report, or Graphical view)
AF056274 Bacteriophage BF23 clone bf23.57r64r, genomic survey sequence gi|3090923|gb|AF056274|AF056274 [3090923] (View GenBank report.FASTA report-ASN.l reportGraphical view, or 3 nucleotide neighbors )
AF056273 Bacteriophage BF23 clone bf23.54fr, genomic survey sequence gi|3090922|gb|AF056273|AF056273 [3090922] (View GenBank report.FASTA reportASN.1 report, or Graphical view)
AF056272 Bacteriophage BF23 clone bf23.47fr.mac 10/7, genomic survey sequence gi|3090921|gb|AF056272|AF056272 [3090921] (View GenBank report,FASTA report-ASN.l report, or Graphical view)
AF056271 Bacteriophage BF23 clone bf23.23.66r, genomic survey sequence gi|3090920|gb|AF056271|AF056271 [3090920] (View GenBank reportFASTA report-ASN.l report, or Graphical view)
AF056270
Bacteriophage BF23 clone bf23.23.64f, genomic survey sequence gi|3090919|gb|AF056270|AF056270 [3090919]
(View GenBank report.FASTA report-ASN.l report, or Graphical view)
AF056269
Bacteriophage BF23 clone bf23.23.60r, genomic survey sequence gi|3090918|gb|AF056269|AF056269 [3090918]
(View GenBank report.FASTA report.ASN.l report, or Graphical view)
AF056268 Bacteriophage BF23 clone bf23.23.60f, genomic survey sequence gi|3090917|gb|AF056268|AF056268 [3090917] (View GenBank report.FASTA repσrt,ASN.1 report.Graphical view, or 1 nucleotide neighbor )
AF056267 Bacteriophage BF23 clone bf23.23.59r, genomic survey sequence gi|3090916|gb|AF056267|AF056267 [3090916] (View GenBank reportFASTA report.ASN.l report, or Graphical view)
AF056266 Bacteriophage BF23 clone bf23.23.59f, genomic survey sequence gi|3090915|gb|AF056266|AF056266 [3090915] (View GenBank reportFASTA report,ASN.1 report or Graphical view)
AF056265 Bacteriophage BF23 clone bf23.23.56r, genomic survey sequence gi|3090914|gb|AF056265|AF056265 [3090914] (View GenBank report.FASTA report-ASN.l report or Graphical view)
AF056264 Bacteriophage BF23 clone bf23.23.56f, genomic survey sequence gi|3090913|gb|AF056264|AF056264 [3090913] (View GenBank reportFASTA report-ASN.1 report, or Graphical view)
AF056263 Bacteriophage BF23 clone bf23.23.68f35r, genomic survey sequence gi|3090912|gb|AF056263|AF056263 [3090912] (View GenBank reportFASTA reportASN.1 report, or Graphical view)
AF056262 Bacteriophage BF23 clone bf23.23.43fr.66f, genomic survey sequence gi|3090911|gb|AF056262|AF056262 [3090911] (View GenBank reportFASTA reportASN.1 report, or Graphical view)
AF056261 Bacteriophage BF23 clone bf23.23.2fr, genomic survey sequence gi|3090910|gb|AF056261|AF056261 [3090910] (View GenBank report FASTA report-ASN.l report, or Graphical view)
AF056260 Bacteriophage BF23 clone bf23.23.55.f, genomic survey sequence gi|3090909|gb|AF056260|AF056260 [3090909] (View GenBank report.FASTA reportASN.1 report, or Graphical view)
AF056259
Bacteriophage BF23 clone bf23.23.53.r, genomic survey sequence gi|3090908|gb|AF056259|AF056259 [3090908]
(View GenBank report,FASTA report-ASN.l report, or Graphical view)
AF056258 Bacteriophage BF23 clone bf23.23.53.f, genomic survey sequence gi|3090907|gb|AF056258IAF056258 [3090907] (View GenBank report.FASTA report.ASN. l report, or Graphical view)
AF056257 Bacteriophage BF23 clone bf23.23.52.r, genomic survey sequence gi|3090906|gb|AF056257|AF056257 [3090906] (View GenBank report.FASTA report.ASN.1 report, or Graphical view)*
AF056256 Bacteriophage BF23 clone bf23.23.52.f, genomic survey sequence gi|3090905|gb|AF056256|AF056256 [3090905] (View GenBank reportFASTA report.ASN.1 report, or Graphical view)
AF056255
Bacteriophage BF23 clone b03.23.49.r, genomic survey sequence gi|3090904|gb|AF056255|AF056255 [3090904]
(View GenBank reportFASTA reportASN.1 report, or Graphical view)
AF056254
Bacteriophage BF23 clone bf23.23.49.f, genomic survey sequence gi|3090903|gb|AF056254|AF056254 [3090903]
(View GenBank reportFASTA reportASN.1 report, or Graphical view)
AF056253 Bacteriophage BF23 clone bf23.23.48.r, genomic survey sequence gi|3090902|gb|AF056253|AF056253 [3090902] (View GenBank reportFASTA reportASN.1 report, or Graphical view)
AF056252 Bacteriophage BF23 clone bf23.23.48.f, genomic survey sequence gi|3090901|gb|AF056252|AF056252 [3090901] (View GenBank reportFASTA reportASN.1 report, or Graphical view)
AF056251
Bacteriophage BF23 clone bf23.23.44.r, genomic survey sequence gi|3090900|gb|AF056251 IAF056251 [3090900]
(View GenBank reportFASTA report-ASN. l report, or Graphical view)
AF056250 Bacteriophage BF23 clone bf23.23.41.f, genomic survey sequence gi|3090899|gb|AF056250|AF056250 [3090899] (View GenBank reportFASTA report-ASN.l report, or Graphical view)
AF056249
Bacteriophage BF23 clone bf23.23.22.a.r, genomic survey sequence gi|3090898|gb|AF056249|AF056249 [3090898]
(View GenBank reportFASTA report-ASN.l report, or Graphical view)
AF056248
Bacteriophage BF23 clone bf23.23.22.a.f, genomic survey sequence gi|3090897|gb|AF056248IAF056248 [3090897]
(View GenBank report,FASTA reportASN.1 report, or Graphical view)
AF056247 Bacteriophage BF23 clone bf23.23.68.r, genomic survey sequence gi|3090896|gb|AF056247|AF056247 [3090896] (View GenBank report.FASTA report.ASN.1 report, or Graphical view)
Z50114
Bacteriophage BF23 DNA for putative tail protein gene gi|2464952|emb|Z50114|BF23LATE [2464952]
(View GenBank report.FASTA report, ASN.1 report.Graphical view, o protein link )
D12824 Bacteriophage BF23 genes for minor tail orotein gp24 and major tail protein gp25, complete eds gi|520578|dbj|D12824|BBF2TAIL [520578] (View GenBank report.FASTA report.ASN.1 report.Graphical view, 1 MEDLINE link, 2 protein links, or 3 nucleotide neighbors )
Z34953 Bacteriophage K3 ip9, iρ7 and ip8 genes gi|535261 |emb|Z34953 IMYK3IP978 [535261 ] (View GenBank reportFASTA report,ASN.1 report-Graphical view, 1 MEDLINE link, 3 protein links, or 1 nucleotide neighbor )
Z35075
Bacteriophage K3 DNA for Ip3 and Ip4 gi|535229|emb|Z35075|MYEORF64K [535229]
(View GenBank report.FASTA report-ASN.1 report,Graphical view, 1 MEDLINE link, or 2 protein links )
X05560
Bacteriophage K3 gene 38 for receptor recognizing protein gi|15112|emb|X05560|MYK3G38 [15112]
(View GenBank reportFASTA report-ASN.1 report, Graphical view.l MEDLINE link, or 1 protein link )
X04747
Bacteriophage 3 gene 37 for receptor recognizing protein gi|15110|emb|X04747|MYK3G37 [15110]
(View GenBank reportFASTA report-ASN.l reportGraphical view.l MEDLINE link, 1 protein link, or 2 nucleotide neighbors )
X01754 Bacteriophage K3 tail fiber gene 36 gi|15108|emb|X01754|MYK3F36 [15108] (View GenBank reportFASTA report,ASN.1 report, Graphical view, 1 MEDLINE link, or 2 protein links )
M16812 Bacteriophage K3 't lysis gene, complete eds gi|215503|gb|M16812|PK3LYST [215503] (View GenBank report,FASTA reportASN.1 report,Graphical view, 1 MEDLINE link, 1 protein link, or 4 nucleotide neighbors )
L46833
Bacteriophage K3 fτd3, frd2 genes, complete eds gi|951377|gb|L46833|PK3FRD32G [951377]
(View GenBank report.FASTA report,ASN.1 reportGraphical view,2 protein links, or 2 nucleotide neighbors )
L43613 Bacteriophage K3 fibritin (wac) gene, complete eds gi|903861|gb|L43613|PK3WAC [903861] (View GenBank report,FASTA report.ASN.1 report-Graphical view.l protein link, or 4 nucleotide neighbors )
X01753 Bactenophage 0x2 tail fiber gene 36 gι|15122|emb|X01753|MYOX2F36 [15122] (View GenBank report.FASTA report.ASN.l reportGraphical view.l MEDLINE link, 2 protein links, or 1 nucleotide neighbor )
L43612 Bactenophage 0x2 fibritin (wac) gene, complete eds gι|903848|gb|L43612|OX2WAC [903848] (View GenBank report.FASTA report.ASN 1 report.Graphical view.l protem link, or 4 nucleotide neighbors )
Z46880 Bactenophage 0X2 stp gene gι|599663|emb|Z46880|BPOX2STP [599663] (View GenBank report.FASTA reportASN.1 report.Graphical view.l MEDLINE link, 1 protem Imk, or 4 nucleotide neighbors )
X05675 Bactenophage Ox2 gene 38 for receptor-recognizing protein and flanking regions gι|15124|emb|X05675|MYOX2G38 [15124] (View GenBank reportFASTA report.ASN.1 reportGraphical view, 1 MEDLINE link, 3 protem links, or 1 nucleotide neighbor )
M33533 Bactenophage RB18 translational repressor protem (regA) and Oτf43.1, complete eds gι!216083|gb|M33533|RB18REGA [216083] (View GenBank report.FASTA reportASN.1 reportGraphical view.l MEDLINE link, 2 protein links, or 2 nucleonde neighbors )
AF033329 Bacteriophage RB 18 single-stranded binding protem (gene 32) gene, partial eds, and 5' region gι|2645788|gb|AF033329|AF033329 [2645788] (View GenBank report.FASTA report-ASN.l reportGraphical view.l protein link, or 11 nucleotide neighbors )
M86231
Bacteriophage RB69 gene 62, 3'end; RegA (regA) gene, complete eds gι|215354|gb|M86231 |P6962REGA [215354]
(View GenBank reportFASTA report.ASN.1 reportGraphical view.l MEDLINE link, 2 protem links, or 1 nucleotide neighbor )
AF033332 Bactenophage RB69 single-stranded binding protem (gene 32) gene, partial eds, and 5' region gι|2645794|gb|AF033332|AF033332 [2645794] (View GenBank reportFASTA reportASN.1 reportGraphical view.l protein link, or 12 nucleotide neighbors )
U34036 Bactenophage RB69 DNA polymerase (43) gene, complete eds gι|1237125|gb|U34036|BRU34036 [1237125] (View GenBank reportFASTA report-ASN.l reportGraphical view.l MEDLINE link, or 1 protein link )
V01145 Bactenophage HI genome fragment Each Thymine given in this sequence represents a HMU-residue (HMU = 5-hydroxvmethyluracil) gι|15557|emb|V01145|PODOHl [15557] (View GenBank reportFASTA reportASN.1 reportGraphical view, or 1 MEDLINE lin )
X05676 Bactenophage Ml gene 38 for receptor recognizing protein and flanking regions gι|15114|emb|X05676|MYMlG38 [15114] (View GenBank reportFASTA report-ASN.l report, Graphical view.l MEDLINE link, 3 protem links, or 1 nucleotide neighbor )
AF034575
Bactenophage M l putative mtegrase (int) gene, complete eds, and artP region, complete sequence gι|2662472|gb|AF034575|AF034575 [2662472]
(View GenBank report.FASTA report.ASN. l report.Graphical view. l MEDLINE link, or 1 protein link )
AF033321
Bactenophage Ml single-stranded binding protem (gene 32) gene, partial eds, and 5' region gι|2645772|gb|AF033321|AF033321 [2645772]
(View GenBank report.FASTA report.ASN.l report.Graphical view.l protein link, or 17 nucleotide neighbors )
X55190
Bacteriophage Tula 37 and 38 genes for receptor-recognizing proteins 37 and 38 (respectively), pamal eds gι|14860|emb|X55190|BPTUIA [14860]
(View GenBank report.FASTA report.ASN.1 report.Graphical view, 1 MEDLINE link, 2 protein Imks, or 2 nucleotide neighbors )
AF033334
Bactenophage Tulb single-stranded binding protem (gene 32) gene, partial eds, and 51 region gi|2645798|gb|AF033334|AF033334 [2645798]
(View GenBank reportFASTA reportASN.1 report.Graphical view, or 5 nucleotide neighbors )
X55191
Bactenophage Tulb 37 gene for receptor-recognizing protem 37 (partial eds), 38 gene for receptor-recognizing protein 38, and t gene (partial eds) gi|14863|emb|X55191|BPTUIB [14863]
(View GenBank report,FASTA report,ASN.1 reportGraphical view, 1 MEDLINE link, 3 protein links, or 3 nucleotide neighbors )
X13065
Bacteriophage phi80 early region gi|14800|emb|X13065|BP80ER [14800]
(View GenBank report.FASTA reportASN.1 report,Graphical view.l MEDLINE link, 8 protein links, or 6 nucleotide neighbors )
D00360
Bacteriophage phi80 cor gene gi|217782|dbj'|D00360|P8080COR [217782]
(View GenBank report.FASTA report.ASN.l report,Graphical view, or 1 protem link )
X01639
Bactenophage phi 80 DNA-fragment with replication origin gi|15828|emb|X01639|XXPHI80 [15828]
(View GenBank report,FASTA reportASN.1 reportGraphical view, 1 MEDLINE link, or 25 nucleotide neighbors )
X04051
Lambdoid bacteriophage phi 80 int-xis region (integrase-excisionase region) gi| 15770|emb|X04051|STPHI80X [15770]
(View GenBank reportFASTA reportASN.1 report,Graphical view.l MEDLINE link, 2 protein links, or 1 nucleotide neighbor )
X06751
Phage Phi80 DNA for major coat protein gι|15768|emb|X06751|STPHI80C [15768]
(View GenBank reportFASTA report-ASN. l report-Graphical view.l MEDLINE link, 1 protein link, or 11 nucleotide neighbors )
X75949
Bactenophage phι80 DNA for ORF xl71.S and ORF xl71.28' gi|458811|emb|X75949|ECORF171B [458811]
(View GenBank report,FASTA report.ASN.1 reportGraphical view, 1 MEDLINE link, 2 protein links, or 28 nucleotide neighbors )
L40418 Bactenophage phι-80 gene, complete eds gι| l019107|gb|L40418|P80A [1019107] (View GenBank report.FASTA report.ASN 1 report.Grapnical view. l MEDLINE link, or 1 protem link )
M24831 Bacteriophage phι-80 Tyr-tRNA gene, 3' end gι|215363|gb|M24831 IP80TGY [215363] (View GenBank report.FASTA reportASN 1 report.Graphical view, 1 MEDLINE link, or 43 nucleotide neighbors )
Ml 0670
Bactenophage phι-80 replication ongm gιj215361|gb|M10670|P80ORI [215361] (View GenBank report.FASTA report.ASN 1 report.Graphical view. l MEDLINE link, 1 protem Imk, or 1 nucleotide neιghbor )
M24825 Bactenophage phι-80 RNA fragment gι|215360|gb|M24825|P80M3A [215360] (View GenBank report.FASTA report.ASN 1 reportGraphical view.l MEDLINE link, or 1 nucleotide neighbor )
M 11919 Bacteriophage phι-80 cl immunity region encodmg the N gene gι|215358|gb|Ml 1919|P80CI [215358] (View GenBank report.FASTA reportASN 1 reportGraphical view.l MEDLINE link, 1 protem link, or 2 nucleotide neighbors )
M10891 Bactenophage phι-80 attP site DNA gι|215357|gb|M 10891 |P80ATTP [215357] (View GenBank reportFASTA reportASN 1 reportGraphical view, 1 MEDLINE link, or 1 nucleotide neighbor )
Ml 9473 Bactenophage 933J (from E coli) proviral Shiga-like toxin type 1 subunits A and B genes, complete eds gι|215072|gb|M 19473) J93SLTI [215072] (View GenBank reportFASTA report-ASN 1 reportGraphical vιew,2 MEDLINE links, 2 protem links, or 20 nucleonde neighbors )
Y10775 Bactenophage 933W lleX, stx2A and stx2B genes gι| 1938206|emb|Y10775|BP933ILEX [1938206] (View GenBank report.FASTA reportASN 1 reportGraphical vιew,2 protem links, or 36 nucleotide neighbors )
X83722 Bactenophage 933 slt-IIB gene gι|1490229|emb|X83722|B933 SLT [1490229] (View GenBank reportFASTA reportASN 1 reportGraphical vιew,2 protem links, or 20 nucleotide neighbors )
X07865 Bactenophage 933W slt-II gene for Shiga-like toxm typell subunit A and B gι|14892|emb|X07865|BWSLΗI [14892] (View GenBank reportFASTA reportASN 1 reportGraphical vιew,2 protem links, or 29 nucleotide neighbors )
Ml 6625 Bactenophage H19B (from E coli) sltLA and sltlB genes encodmg Shiga-like toxin I subunits A and B, complete eds gι|215043|gb|M16625|H19BSLT [215043] (View GenBank reportFASTA report-ASN 1 reportGraphical vιew, l MEDLINE link, 2 protem links, or 24 nucleonde neighbors )
M17358
Bactenophage H I 9B shiga-like toxin- 1 (SLT- 1 ) A and B subunit DNA, complete eds gι|215046|gb|M17358|H19BSLTA [215046]
(View GenBank report.FASTA report.ASN 1 report.Graphical view. l MEDLINE link, 2 protem links, or 20 nucleotide neighoor. )
U29728
Bactenophage N4 single-stranded DNA-bmdmg protem (N4SSB) gene, complete eds gι|939708|gb|U29728|BNU29728 [939708]
(View GenBank report.FASTA reportASN 1 reportGraphical vιew,2 MEDLINE links, or 1 protem link )
J02580 Bacteriophage PA-2 (E coli porcine strain isolate) Rz gene, 5'end, ORF2, outer membrane porm protem (lc) and ORF1 gene- complete eds gι|215366|gb|J02580|PA2LC [215366] (View GenBank report.FASTA report, ASN 1 report.Graphical view. l MEDLINE link, 4 protem Imks, or 4 nucleotide neighbor. )
U32222 Bactenophage 186, complete sequence gι|3337249|gb|U32222|B 1U32222 [3337249] (View GenBank report.FASTA reportASN 1 report.Graphical vιew,6 MEDLINE links, 46 protein links, or 5 nucleo de neignoors ;
X51522 Bactenophage P4 complete DNA genome gι|450916|emb|X51522|MYP4CG [450916]
(View GenBank reportFASTA reportASN 1 reportGraphical vιew,3 MEDLINE links, 13 protem links, 6 nucleonde neighbor. or 1 genome link )
X92588 Bactenophage 82 orf33, orfl51, orf56, orf96, rus, orf45, and Q genes gι|l 05111 l|emb|X92588|BAC82HOLL [1051111] (View GenBank report.FASTA reportASN 1 reportGraphical vιew,7 protem links, or 1 nucleotide neighbor )
J02803 Bactenophage 82 anntermmanon protem (Q) gene, complete eds gι|215364|gb|J02803|P82Q [215364] (View GenBank report.FASTA report-ASN 1 reportGraphical vιew,l MEDLINElink, or 1 protem link )
U02466 Bactenophage HK022 (cro), (ell) and (O) genes, complete eds, (P) gene, partial eds gι|407285|gb|U02466|BHU02466 [407285] (View GenBank reportFASTA report-ASN 1 reportGraphical view.l MEDLINE link, 5 protem links, or 1 nucleotide neighbor )
M26291 Bactenophage D108 regulatory DNA-bmdmg protem (ner) gene, complete eds gι|166194|gb|M26291|D18NER [166194] (View GenBank reportFASTA report-ASN 1 reportGraphical view.l MEDLINE link, 1 protem link, or 1 nucleonde neighbor )
Ml 1272
Bactenophage D108 left-end DNA gι|166193|gb|Ml 1272|D18LEDNA [166193]
(View GenBank reportFASTA reportASN 1 reportGraphical view.l MEDLINE link, or 2 nucleotide neighbors )
Ml 8902 Bactenophage D108 bl gene encodmg a replication protein, 3' end, and containing three ORFs, complete eds gι| 16619 l|gb|M18902|D18KIL [166191] (View GenBank reportFASTA reportASN 1 reportGraphical view.l MEDLINE link, 1 protem Imk, or 3 nucleotide neighbors )
M 10191
Bactenophage D 108, left end with Mu A protem binding sites LI and L2 gι| 166190|gb|M 10191 |D 18BS [ 166190]
(View GenBank report.FASTA reportASN.1 report.Graphical view. l MEDLINE link, or 5 nucleonde neighbors )
J02447 bactenophage d 108 gene a 5' end gι| 166189|gb| J02447|D 18 AAA [ 166189] (View GenBank report.FASTA report.ASN. l reportGraphical view, o'r'l MEDLINE link )
V00865 Bactenophage D 108 fragment from genes A and ner (C- terminus of ner and N-terminus of A) gι| 15437|emb|V00865|NCD 108 [15437] (View GenBank report.FASTA report.ASN.1 report.Graphical view, 1 MEDLINE link, or 2 protem Imks )
X01914 Bactenophage IKe gene for DNA binding protem gι|14957|emb|X01914|INIKEDBP [14957] (View GenBank reportFASTA reportASN.1 reportGraphical view, 1 MEDLINE link, 1 protem link, or 2 nucleonde neighbors )
AF064539 Bactenophage N15, complete genome gι|3192683|gb|AF064539|AF064539 [3192683]
(View GenBank reportFASTA reportASN.1 reportGraphical vιew,2 MEDLINE links, 60 protem links, 26 nucleotide neighbors, or I genome link )
U02303 Bactenophage Ifl, complete genome gi|3676280|gb|U02303|B2U02303 [3676280] (View GenBank report.FASTA reportASN.1 rep on, Graphical view, 10 protein Imks, or 1 genome lin )
AF007792 Bactenophage Mu late morphogenetic region gι|3551775|gb|AF007792|AF007792 [3551775] (View GenBank reportFASTA reportASN.1 reportGraphical view, or 1 nucleotide neighbor )
U24159 Bactenophage HP1 strain HPlcl, complete genome gι|1046235|gb|U24159|BHU24159 [1046235]
(View GenBank reportFASTA report-ASN. l reportGraphical vιew,6 MEDLINE links, 41 protem Imks, 8 nucleotide neighbors, or 1 genome link )
Z71579 Bactenophage S2 type A 5.6 kb DNA fragment gi|1679806|emb|Z71579|BPHSlADNA [1679806] (View GenBank reportFASTA reportASN.1 reportGraphical view,3 MEDLINE links, 9 protem links, or 9 nucleotide neighbors
X53238 Klebsiella sp. bactenophage Kl 1 gene 1 for RNA polymerase gι|14984|emb|X53238|KSKl 1RPO [14984] (View GenBank reportFASTA report-ASN.l reportGraphical view. l MEDLINE link, 1 protein link, or 1 nucleotide neighbor )
X85010 Bactenophage A51 1 ply51 1 gene gι|853748|emb|X85010|BPA511PLY [853748] (View GenBank report.FASTA report.ASN 1 report.Graphical view. l MEDLINE link, 3 protem links, or 1 nucleotide neighbor )
U29728 Bactenophage N4 single-stranded DNA-bmdmg protem (N4SSB) gene, complete eds gι|939708|gb|U29728|BNU29728 [939708] (View GenBank report.FASTA report.ASN 1 report.Graphical view 2 MEDLINE links or 1 protem link )
J02445 bactenophage bol 3 '-terminal region ma gι|166152|gb|J02445|BOlTR3 [166152] (View GenBank report.FASTA reportASN 1 report.Graphical view 1 MEDLINE link or 5 nucleonde neighbors )
L06183 Bactenophage L5 (from Leucoπostoc oenos) genome gι|289353|gb|L06183|BL5GENM [289353] (View GenBank reportFASTA reportASN 1 reportGraphical view, or 1 genome lin )
AF074945 Mycoplasma arthnndis bactenophage MAV1, complete genome gι|351 1243|gb|AF074945|AF074945 [3511243] (View GenBank report.FASTA reportASN 1 report, Graphical view, 15 protem links, 3 nucleonde neighbors, or 1 genome link )
LI 3696 Bactenophage L2 (from Mycoplasma), complete genome gι|289338|gb|L13696|BL2CG [289338] (View GenBank reportFASTA report-ASN 1 reportGraphical vιew,3 MEDLINE links, 14 protem links, or 1 genome link )
X80191
Bactenophage PP7 mRNA for maturation, coat lysis and replicase proteins gι|517237|emb|X80191 |BPP7PR [51 237]
(View GenBank reportFASTA reportASN.1 reportGraphical view, 1 MEDLINE link 4 protem Imks, or 1 genome link )
M19377 Bactenophage PD from Pseudomonas aeniginosa (New York strain), complete genome gι|2I5380|gb|M19377|PF3COMNY [215380] (View GenBank reportFASTA report-ASN 1 reportGraphical view, 1 MEDLINE link 9 orotem Imks, or 5 nucleonde neighbors )
M11912
Bactenophage PD from Pseudomonas aeniginosa (Nymegen strain), complete genome gι|215371|gb|M11912|PF3COMN [215371]
(View GenBank reportFASTA report-ASN.l reportGraphical view.l MEDLINE link, 9 protem links, 5 nucleonde neighbors or 1 genome lin )
V00605 Bactenophage Pfl gene encodmg DNA binding protem gι|14970|emb|V00605|INOPFl [14970] (View GenBank report.FASTA reportASN.1 report, Graphical view.l proteine link, or 1 nucleotide neighbor )
L05626 Bactenophage PR4 capsid protem (P6) gene, complete eds gι|215735|gb|L05626|PR4P6MAJA [215735] (View GenBank reportFASTA report-ASN 1 reportGraphical vιew,l MEDLINE link, 1 υrotein link, or 1 nucleotide neighbor )
D 13409
Bactenophage phiCTX (isolated from Pseudomonas aeniginosa) cosR, attP, int genes gι|217776|dbj|D 13409|BPHCOSR [217776]
(View GenBank report.FASTA report.ASN 1 report.Graphical view.l MEDLINE link, 3 protem links, or 3 nucleotide neighbors )
D 13408 Bactenophage phiCTX (isolated from Pseudomonas aeniginosa) cosL, ctx genes gι|217775|dbj|D 13408|BPHCOSLCTX [217775] (View GenBank report.FASTA report.ASN 1 report.Graphical view, 2 iNΪEDLINE links, or 3 nucleonde neighbors )
M24832 Bactenophage f2 coat protem gene, partial eds gι|166228|gb|M24832|F2CRNACA [166228] (View GenBank report.FASTA report, ASN 1 report.Graphical view, 1 MEDLINE link, 1 protem Imk, or 4 nucleotide neighbors )
S72011 Bactenophage 21 lsocitrate dehydrogenase (led) and mtegrase (int) genes.partial eds gι|2618967|gb|AF017629|AF017629 [2618967] (View GenBank reportFASTA reportASN 1 reportGraphical view, 1 MEDLINEhnk, 2 protem links, or 44 nucleotide neighbors )
AFO 17628
Bactenophage 21 lsocitrate dehydrogenase (led) and mtegrase (int) genes, partial eds gι|2618964|gb|AF017628|AF017628 [2618964]
(View GenBank reportFASTA reportASN 1 reportGraphical view.l MEDLINElink, 2 protem links, or 44 nucleonde neighbors )
AFO 17627 Bactenophage 21 lsocitrate dehydrogenase (led) and mtegrase (int) genes, partial eds gι|2618961 |gb|AF017627|AF017627 [2618961] (View GenBank report.FASTA reportASN 1 reportGraphical view.l MEDLINElink, 2 protem links, or 44 nucleotide neighbors )
AF0I7626
Bacteriophage 21 lsocitrate dehydrogenase (led) gene, partial eds, and mtegrase (mt) gene, partial eds gι|2618958|gb|AF017626|AF017626 [2618958]
(View GenBank reportFASTA report-ASN.l reportGraphical view.l MEDLINE link, 2 protem links, or 49 nucleotide neighbors )
AFO 17625 Bactenophage 21 lsocitrate dehydrogenase (led) and mtegrase (mt) genes, partial eds gι|2618955|gb|AFO 17625) AFO 17625 [2618955] (View GenBank reportFASTA report-ASN 1 reportGraphical view.l MEDLINElink, 2 protem links, or 44 nucleotide neighbors )
AFO 17624
Bactenophage 21 lsocitrate dehydrogenase (led) and mtegrase (mt)genes, partial eds gι|2618952|gb|AF017624|AF017624 [2618952]
(View GenBank reportFASTA report-ASN 1 reportGraphical view.l MEDLINElink, 2 protem links, or 44 nucleonde neighbors )
AFO 17623 Bactenophage 21 lsocitrate dehydrogenase (led) and mtegrase (mt) genes, partial eds gι|2618949|gb|AF017623|AF017623 [2618949] (View GenBank report.FASTA reportASN.1 reportGraphical view.l MEDLINE link, 2 protem Imks, or 44 nucleonde neighbors )
AFO 17622
Bactenophage 21 lsocitrate dehydrogenase (ted) and mtegrase (mt) genes, partial eds gι|2618946|gb|AF017622| AFO 17622 [2 18946]
(View GenBank reportFASTA reportASN 1 report, GraDhical view.l MEDLINE link, 2 protem Imks, or 44 nucleotide neighbors )
AFO 17621
Bactenophage 21 lsocitrate dehydrogenase (led) and mtegrase (mt) genes, partial eds gι|2618943|gb|AF017621|AF017621 [2618943]
(View GenBank reportFASTA reportASN.1 report.Graphical view.l MEDLINE link, 2 protem links, or 44 nucleotide neighbors )
D26449
Bactenophage PS 17 FI gene for tail sheath protem (gpFI) and FII gene for tail tube protem (gpFII), complete eds gι|452162|dbj|D26449|BPSFIFπ [452162]
(View GenBank report.FASTA report-ASN.1 reportGraphical view, or 2 protem links )
X87627 Bactenophage D3112 A and B genes gι|974768|emb|X87627|BPD3112AB [974768] (View GenBank report.FASTA report.ASN.l report.Graphical view.l MEDLINElink, 2 protein links, or 1 nucleonde neighbor )
U32623 Bactenophage D3 transcnptional activator C_I (ell) gene, complete eds gι|984852|gb|U32623|BDU32623 [984852] (View GenBank reportFASTA reportASN.1 reportGraphical view.l protein link, or 1 nucleotide neighbor )
L34781 Bactenophage phi 11 holm homologue (ORF3) gene, complete eds and pepndoglycan hydrolase (lytA) gene, partial eds gι|511838|gb|L34781 |BPHHOLIN [511838] (View GenBank reportFASTA report-ASN.1 reportGraphical view.l MEDLINE link, 4 protein links, or 2 nucleotide neighbors )
L14810 Bactenophage P22 (gplO) gene, complete eds, and (gp26) gene, complete eds gι|294053|gb|L 14810|P22GP 1026X [294053] (View GenBank report.FASTA reportASN.1 reportGraphical view.l MEDLINE link, 2 protein links, or 2 nucleotide neighbors )
X87420 Bacteriophage ESI 8 genes 24, c2, cro, cl, 18, and oL and oR operators gi|1143407|emb|X87420|BPES18GEN [1143407] (View GenBank reportFASTA report.ASN.1 reportGraphical vιew,5 protein links, or 9 nucleotide neighbors )
L42820 Bacteriophage BF23 tail protein (hrs) gene, complete eds gι| 1048680|gb|L42820|BBFHRS [ 1048680] (View GenBank reportFASTA report-ASN.l reportGraphical view.l MEDLINElink, 1 protein link, or 1 nucleotide neighbor )
X14980 Bactenophage PRD1 XV gene for protein P 15 (lytic enzyme) gι|15802|emb|X14980|TEPRDlXV [15802] (View GenBank reportFASTA report-ASN.l reportGraphical view.l MEDLINElink, 1 protein link, or 4 nucleotide neighbors )
X06321 Bactenophage PR 1 gene 8 for DNA terminal protem gι|15800|emb|X06321|TEPRD18 [15800] (View GenBank reportFASTA reportASN.1 reportGraphical view.l MEDLINE link, 2 protem links, or 10 nucleotide neighbors )
X14336 Filamentous Bactenophage 12-2 genome gι|14920|emb|X14336|INBI22 [14920]
(View GenBank reportFASTA reportASN.1 reportGraphical view.l MEDLINE link, 9 protein links, 1 nucleotide neighbor, or 1 genome link )
L05001 240
Bacteriophage X glucosyl transferase gene, complete eds gi|216044|gb|L05001 |PXFCLUS YLT [216044] (View GenBank report.FASTA report.ASN. l report, Graphical view. l MEDLINE link, or 1 protein link )
M29479
Bacteriophage p4 sid and psu genes partial eds, and delta gene, complete eds gi|215701| gb|M29479|PP4SDP [215701]
(View GenBank report.FASTA report.ASN. l reportGraphical view,3 protein links, or 4 nucleotide neighbors )
SEG_PP4PSUSID
Bacteriophage P4 capsid size determination protein (sid) gene, 5' end gi|215698|gb||SEG_PP4PSUSID [215698]
(View GenBank report.FASTA report.ASN. l report.Graphical view. l MEDLINE link, 2 protein links, or 1 nucleotide neighbor )
M29650 Bacteriophage P4 polarity suppression protein (psu) gene, complete eds gi|215697|gb|M29650|PP4PSUSID2 [215697] (View GenBank report.FASTA reportASN.1 report, or Graphical view)
M29651 Bacteriophage P4 capsid size determination protein (sid) gene, 5' end gi|215696|gb|M29651 |PP4PSUSID 1 [215696] (View GenBank reportFASTA report-ASN.1 report, or Graphical view)
M27748 Bacteriophage P4 gop, beta, and ell genes, complete eds and int gene, 3' end gi|215691 |gb|M27748|PP4GOPBC [215691 ] (View GenBank reportFASTA reportASN.1 reportGraphical view, 1 MEDLINE link, 4 protein links, or 1 nucleotide neighbor )
K02750
Bacteriophage IKe, complete genome gi|215061|gb|K02750|IKECG [215061]
(View GenBank report.FASTA report-ASN. l reportGraphical view. l MEDLINElink, 10 protein links, 4 nucleotide neighbors, or 1 genome link )
L40418 Bacteriophage phi-80 gene, complete eds gi|1019107|gb|L40418|P80A [1019107] (View GenBank reportFASTA reportASN.1 reportGraphical view. l MEDLINE link, or 1 protein link )
AF032122 Bacteriophage Sfil integrase (int) gene, partial eds; and bactoprenol glucosyl transferase (bgt), and glucosyl tranferase II (gtrll) genes, complete eds gi|2465412|gb|AF021347| AF021347 [2465412] (View GenBank reportFASTA reportASN.1 reportGraphical view.l MEDLINElink, 4 protein links, or 2 nucleotide neighbors )
M35825 Bacteriophage SF6 fragment D lysozyme gene, complete eds gi|216105|gb|M35825|SF6LYZ [216105] (View GenBank reportFASTA reportASN.1 reportGraphical view, or 1 protein link )
Z35479 Bacteriophage C16 ipl gene gi|534936|emb|Z35479|BC16IPl [534936] (View GenBank reportFASTA reportASN.1 reportGraphical view.l MEDLINE link, 1 protein link, or 2 nucleotide neighbors )
X12638
Bacteriophage 21 DNA for gene 2 gi|296141|emb|X12638|B21GENE2 [296141]
(View GenBank report.FASTA report.ASN. l report.Graphical view. l MEDLINE link, 1 protein link, or 1 nucleotide neighbor )
X02501
Bacteriophage 21 DNA for left end sequence with genes 1 and 2 gi|15825|emb|X02501|XXPHA21 [15825]
(View GenBank report.FASTA reportASN.1 reportGraphical view.l MEDLINE link, 2 protein links, or 3 nucleotide neighbors )
M65239 Bacteriophage 21 lysis genes S, R, and Rz, complete eds gi|215466|gb|M65239|PH2LYSGEN [215466] (View GenBank report.FASTA report.ASN.1 report.Graphical view, 1 MEDLINE link, 3 protein links, or 1 nucleotide neighbor )
M58702 Bacteriophage 21 late gene regulatory region gi|215465|gb|M58702|PH2LATEGE [215465] (View GenBank report.FASTA reportASN.1 reportGraphical view, or 1 MEDLINE link )
M81255 Bacteriophage 21 head gene operon gi|215454|gb|M81255|PH2HEADTL [215454] (View GenBank reportFASTA reportASN.1 reportGraphical view,2 MEDLINE links, 10 protein links, or 4 nucleotide neighbors )
M23775 Bacteriophage 21 glycoprotein 1 gene, complete eds, and glycoprotein gene, 5' end gi|215451|gb|M23775|PH2GPA [215451] (View GenBank reportFASTA reportASN.1 reportGraphical view, 1 MEDLINE link, 2 protein links, or 3 nucleotide neighbors )
M61865 Bacteriophage 21 excisionase (xis), integrase (int) and isocitrate dehydrogenase (icd), complete eds gi|215448|gb|M61865|PH22XISAA [215448] (View GenBank reportFASTA report-ASN.l reportGraphical view,2 protein links, or 9 nucleotide neighbors )
S72011
Bacteriophage 21 isocitrate dehydrogenase (icd) and integrase (int) genes, partial eds gi|2618967|gb|AF017629|AF017629 [2618967]
(View GenBank reportFASTA report-ASN.l reportGraphical view.l MEDLINE link, 2 protein links, or 44 nucleotide neighbors )
AF017628
Bacteriophage 21 isocitrate dehydrogenase (icd) and integrase (int) genes, partial eds gi|2618964|gb|AF017628|AF017628 [2618964]
(View GenBank reportFASTA reportASN.1 reportGraphical view, 1 MEDLINE link, 2 protein links, or 44 nucleotide neighbors )
AF017627
Bacteriophage 21 isocitrate dehydrogenase (icd) and integrase (int) genes, partial eds gi|2618961 |gb|AF017627|AF017627 [2618961]
(View GenBank reportFASTA reportASN.1 reportGraphical view.l MEDLINE link, 2 protein links, or 44 nucleotide neighbors )
AF017626
Bacteriophage 21 isocitrate dehydrogenase (icd) gene, partial eds; and integrase (int) gene, partial eds gi|2618958|gb|AF017626|AF017626 [2618958]
(View GenBank reportFASTA report-ASN.l reportGraphical view.l MEDLINE link, 2 protein links, or 49 nucleotide neighbors )
WO 00/32825 r<" i ay 'υ
AFO 17625 242
Bactenophage 21 isocitrate dehydrogenase (icd) and mtegrase (mt) genes, partial eds gι|2618955|gb|AFO 17625|AF017625 [2618955] (View GenBank report.FASTA report.ASN.1 report.Graphical view, 1 MEDLINE link, 2 protem links, or 44 nucleotide neighbors
AFO 17624 Bactenophage 21 isocitrate dehydrogenase (icd) and mtegrase (mt) genes, partial eds gι|2618952|gb|AF017624|AF017624 [2618952] (View GenBank report.FASTA reportASN.1 report.Graphical view, 1 MEDLINE link, 2 protem links, or 44 nucleonde neighbors )
AFO 17623 Bacteriophage 21 isocitrate dehydrogenase (icd) and mtegrase (mt) genes, partial eds gi|2618949|gb|AF017623|AF017623 [2618949] (View GenBank report.FASTA report.ASN.1 report.Graphical view, 1 MEDLINE link, 2 protem links, or 44 nucleotide neighbors )
AFO 17622 Bacteriophage 21 isocitrate dehydrogenase (icd) and mtegrase (mt) genes, partial eds gi|2618946|gb|AF017622|AF017622 [2618946] (View GenBank report.FASTA report-ASN.l report.Graphical view.l MEDLINE link, 2 protem links, or 44 nucleotide neighbors )
AF017621 Bactenophage 21 isocitrate dehydrogenase (icd) and integrase (mt) genes, partial eds gι|2618943|gb) AFO 176211 AFO 17621 [2618943] (View GenBank report.FASTA reportASN.1 reportGraphical view.l MEDLINE link, 2 protem links, or 44 nucleotide neighbors )
M57455 Bactenophage 42D (clone pDB17) (from Staphylococcus aureus) staphylokinase gene, complete eds gι|215344|gb|M57455|P42STK [215344] (View GenBank reportFASTA reportASN.1 report.Graphical view.l protem link, or 9 nucleonde neighbors )
Y12633
Bacteriophage 85 DNA, promoter sequence of unknown gene gi|2058285|emb|Y12633|B85PROM [2058285]
(View GenBank report.FASTA report-ASN.l report, or Graphical view)
X98146
Bacteriophage PI DNA sequence around the Op88 operator gi|1359513|emb|X98146|BP10P880P [1359513]
(View GenBank reportFASTA reportASN.1 reportGraphical view, or 1 nucleonde neighbor )
Y07739 Staphylococcus phage Twort holTW, plyT genes gi|2764979|emb|Y07739|BPTWGHOLG [2764979] (View GenBank reportFASTA reportASN.1 reportGraphical view, or 2 protein links )
L07580
Bacteriophage phi-11 rinA and rin B genes, required for the activation of Staphylococcal phage phi-11 int expression gi| 166160|gb|L07580|BPHRINAB [ 166160] (View GenBank reportFASTA report-ASN.l reportGraphical view,l MEDLINE link, or 2 protein links)
M34832 Bacteriophage phi-11 integrase (int) and excisionase (xis) genes, complete eds gi|166157|gb|M34832|BPHINTXIS [166157] (View GenBank reportFASTA report-ASN.l report.Graphical view.l MEDLINE link, 2 protein links, or 2 nucleonde neighbors )
M20394
Bacteriophage phi-1 1 S.aureus attachment site (attP) gi| 166156|gb|M20394|BPHATTP [166156]
(View GenBank reportFASTA reportASN.1 report.Graphical view. l MEDLINE link, or 4 nucleotide neighbors )
X23128 Bacteriophage phi- 13 integrase gene gi|758228|emb|X82312|PHI13INT [758228] (View GenBank reportFASTA reportASN.1 report.Graphical view.l protein link, or 3 nucleotide neighbors )
X61719 S.aureus phi- 13 lysogen right chromosome/bacteriophage DNA junction gi|46625|emb|X61719|SAP13RJNC [46625] (View GenBank report.FASTA reportASN.1 report.Graphical view, or 1 MEDLINE link )
X61718 S.aureus phi- 13 lysogen left chromosomal bacteriophage DNA junction gi|46624|emb|X61718|S AP 13LJNC [46624] (View GenBank reportFASTA reportASN.1 reportGraphical view, or 1 MEDLINE link )
X61717
Bacteriophage phi- 13 core sequence for attachment gi| 14799|emb|X61717|BP13ATTP [14799]
(View GenBank reportFASTA reportASN.1 reportGraphical view,2 MEDLINE links, or 3 nucleotide neighbors )
U01875 Bacteriophage phi- 13 putative regulatatory region and integrase (int) gene, partial eds gi|437118|gb|U01875|U01875 [437118] (View GenBank report.FASTA reportASN.1 reportGraphical view,3 MEDLINE links, or 4 nucleotide neighbors )
X67739
S.aureus Bacteriophage phi-42 attP gene gi|14809|emb|X67739|BPATTPA [14809]
(View GenBank reportFASTA report-ASN.l reportGraphical view.l MEDLINE link, or 3 nucleotide neighbors )
U01872 Bacteriophage phi-42 integrase (int) gene, complete eds gi|437115|gb|U01872|U01872 [437115] (View GenBank reportFASTA report-ASN.l reportGraphical view,3 MEDLINE links, 2 protein links, or 3 nucleotide neighbors )
X94423 Staphylococcus aureus bacteriophage phi-42 DNA with ORFs (restriction modification system) gi| 1771597|emb|X94423|SARMS [1771597] (View GenBank reportFASTA reportASN.1 reportGraphical view,2 protein links, or 1 nucleotide neighbor )
M27965 Bacteriophage L54a (from S.aureus) int and xis genes, complete eds gi|215096|gb|M27965|L54INTXIS [215096] (View GenBank reportFASTA reportASN.1 report.Graphical view, MEDLINE 1 link, 2 protein links, or 3 nucleotide neighbors )
U72397 Bacteriophage 80 alpha holin and amidase genes, complete eds gi| 1763241 |gb|U72397|B 8U72397 [ 1763241 ] (View GenBank report.FASTA report-ASN.1 reportGraphical view,2 protein links, or 2 nucleotide neighbors )
AB009866
Bacteriophage phi PVL proviral DNA, complete sequence gi|3341907|dbj|AB009866|AB009866 [3341907]
(View GenBank report.FASTA report.ASN.l report.Graphical view,63 protein links, or 1 nucleotide neighbor )
Z47794
Bacteriophage Cp-1 DNA, complete genome gi|2288892|emb|Z47794|BPCPlXX [2288892]
(View GenBank report.FASTA report.ASN.l reportGraphical view,3 MEDLINE links, 28 protein links, 1 nucleotide neighbor, or
1 genome link )
SEG_CP7RSIT
Bacteriophage Cp-7 (S.pneumoniae) 5' inverted terminal repeat gi| 166186|gb||SEG_CP7RSIT [ 166186]
(View GenBank report.FASTA report, ASN.1 report, Graphical view, or 1 MEDLINE link )
Ml 1635
Bacteriophage Cp-7 (S.pneumoniae) DNA, 3' inverted terminal repeat gi|166185|gb|M11635|CP7RSIT2 [166185]
(View GenBank report.FASTA reportASN.1 report, or Graphical view)
Ml 1636
Bacteriophage Cp-7 (S.pneumoniae) 5' inverted terminal repeat gi|166184|gb|Ml 1636|CP7RSIT1 [166184]
(View GenBank report.FASTA report-ASN.l report, or Graphical view)
SEG_CP5RSIT
Bacteriophage Cp-5 (S.pneumoniae), 5' inverted terminal repeat gi| 166181|gb||SEG_CP5RSIT [166181]
(View GenBank report.FASTA reportASN.1 reportGraphical view, or 1 MEDLINE link )
Ml 1633
Bacteriophage Cp-5 (S.pneumoniae) 3' inverted terminal repeat gi|166180|gb|M11633|CP5RSIT2 [166180]
(View GenBank report.FASTA reportASN.1 report, or Graphical view)
Ml 1634
Bacteriophage Cp-5 (S.pneumoniae), 5' inverted terminal repeat gi|166179|gb|M11634|CP5RSITl [166179]
(View GenBank reportFASTA report-ASN.l report, or Graphical view)
M34780
Bacteriophage Cp-9 mura idase (cpl9) gene gi|166187|gb|M34780|CP9CPL [166187]
(View GenBank reportFASTA reportASN.1 reportGraphical view.l MEDLINE link, 1 protein link, or 1 nucleotide neighbor )
M34652
Bacteriophage HB-3 amidase (hbl) gene, complete eds gi|215055|gb|M34652|HB3HBLA [215055]
(View GenBank reportFASTA reportASN.1 reportGraphical view.l MEDLINE link, or 1 protein link )
U64984
Streptococcus pyogenes phage T12 repressor, excisionase (xis), integrase(int) and erythrogenic toxin A precursor (speA) genes, complete eds gi|1877426|gb|U40453|SPU40453 [1877426]
(View GenBank reportFASTA reportASN.1 reportGraphical view,2 MEDLINE links, 4 protein links, or 22 nucleotide neighbors )
X12375
Phage CP-T1 (Vibrio cholerae) DNA for packaging signal (pac site) gi|15435|emb|X12375|NCCPPAC [15435]
(View GenBank report.FASTA report.ASN. l report.Graphical view. l MEDLINE link, or 1 protein link )
AF087814
Vibrio cholerae filamentous bacteriophage fs-2 DNA, complete genome sequence gi|3702207|dbj|AB002632|AB002632 [3702207]
(View GenBank report.FASTA reportASN.1 reportGraphical view, 1 MEDLINE link, 9 protein links, or 1 genome link )
D83518
Bacteriophage KVP40 gene for major capsid protein precursor, complete eds gi|3046858|dbj|D83518|D83518 [3046858]
(View GenBank report.FASTA report.ASN. l report.Graphical view.l MEDLINE link, or 1 protein link )
AF033322 Bacteriophage PST single-stranded binding protein (gene 32) gene, partial eds, and 5' region gi|2645774|gb|AF033322|AF033322 [2645774] (View GenBank reportFASTA reportASN.1 reportGraphical view.l protein link, or 17 nucleotide neighbors )
X94331
Bacteriophage L cro, 24, c2, and cl genes gi|1469213|emb|X94331|BLCR024C [1469213]
(View GenBank report.FASTA reportASN.1 report.Graphical view.l MEDLINE link, or 4 protein links )
U82619 Shigella flexneri bacteriophage V glucosyl transferase (gtr), integrase (int) and exeisionase (xis) genes, complete eds gi|2465470|gb|U82619|SFU82619 [2465470] (View GenBank reportFASTA reportASN.1 reportGraphical view.l MEDLINE link, 8 protein links, or 1 nucleotide neighbor )
lable 12
NCBI Entrez Nucleotide QUERY Key words: bacteriophage and lysis 56 citations found (all selected)
AJ011581
Bacteriophage PS119 lysis genes 13, 19, 15, and packaging gene 3, complete eds gil3676084JemblAJ011581IBPS011581 [3676084]
(View GenBank report.FASTA reportASN.1 report.Graphical vie\v,4 protein links, or 1 nucleotide neighbor )
AJ011580
Bacteriophage PS34 lysis genes 13, 19, 15, antitermiπator gene 23, and packaging gene 3, complete eds gil3676078lemblAJ011580IBPS011580 [3676078]
(View GenBank report.FASTA reportASN.1 reportGraphical view,5 protein links, or 2 nucleotide neighbors )
AJ011579
Bacteriophage PS3 lysis genes 13, 19, 15, and packaging gene 3 gil3676O73lemblAJ011579IBPS011579 [3676073]
(View GenBank reportFASTA reportΛSN.l reportGraphical view,4 protein links, or 1 nucleotide neighbor )
AF034975
Bacteriophage H-19B essential recombination function protein (erf), _dl protein (kil), regulatory protein clll (cIII), protein gpl7 (17), N protein (N), cl protein (cl), cro protein (cro), ell protein (ell), O protein (O), P protein (P), ren protein (ren), Roi (roi), Q protein (Q),
Shiga-like to in A (slt-lA) and B (slt-IB) subunits, and putative holin protein (S) genes, complete eds gil2668751lgblAF0349751 [2668751]
(View GenBank reportFASTA reportASN, 1 reportGraphical view.l MEDLINE link, 20 protein links, or 30 nucleotide neighbors )
U37314
Bacateriophage lambda Rzl protein precursor (Rzl) gene, complete eds gill017780lgblU37314IBLU37314 [1017780]
(View GenBank reportFASTA reportASN.1 reportGraphical view,2 MEDLINE links, I protein link, or 9 nucleotide neighbors )
U00005
E. coli hflA locus encoding the hilX, hflK and hflC genes, hfq gene, complete eds; miaA gene, partial eds gil436153lgblU00005IECOHFLA [436153]
(View GenBank reportFASTA reportΛSN.l reportGraphical view,4 MEDLINE
links, 5 protein links, or 8 nucleotide neighbors
U32222
Bacteriophage 186, complete sequence gi!3337249lgblU32222IBlU32222 [3337249]
(View GenBank reportFASTA reportΛSN.l report.Graphical vie\v,6 MEDLINE links, 46 protein links, or 5 nucleotide neighbors )
AF064539
Bacteriophage N15, complete genome gil3192683lgblAF0645391AF064539 [3192683]
(View GenBank report.FASTA reportΛSN.l report.Graphical view,2 MEDLINE links, 60 protein links, 26 nucleotide neighbors, or 1 genome link )
AF063097
Bacteriophage P2, complete genome gil3139086lgblAF063097IAF063097 [3139086]
(View GenBank reportFASTA reportΛSN.l reportGraphical view,21 MEDLINE links, 42 protein links, 3 nucleotide neighbors, or 1 genome link )
Z97974
Bacteriophage phiadh lys, hoi, intG, rad.and tec genes gil2707950lemblZ97974IBPHIADH [2707950]
(View GenBank reportFASTA reportΛSN.l reportGraphical view,2 MEDLINE links, 9 protein links, or 1 nucleotide neighbor )
AF059243
Bacteriophage NL95, complete genome gil30885451gbIAF059243IAF059243 [3088545]
(View GenBank report.FASTA reportΛSN.l reportGraphical view,2 MEDLINE links, 4 protein links, 3 nucleotide neighbors, or 1 genome link )
AF052431
Bacteriophage Ml 1 A-protein, coat protein, A 1 -protein, and replicase genes, complete eds gil2981208lgWAF052431l [2981208]
(View GenBank reportFASTA reportΛSN.l reportGraphical view,2 MEDLINE links, 4 protein links, or 8 nucleotide neighbors )
Y07739
Staphylococcus phage Twort holTW, plyTW genes gil2764979iemWY07739IBPTWGHOLG [2764979] (View GenBank reportFASTA reportΛSN.l reportGraphical view, or 2 protein links )
X94331
Bacteriophage L cro, 24, c2, and cl genes gil 1469213lemblX94331IBLCR024C [ 1469213]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, or 4 protein links )
X78410
Bacteriophage phiadh holin and lysin genes gil793848lemblX7841CHLGHOLLYS [793848]
(View GenBank report.FASTA reportΛSN.l report.GraphicaLyiew.l MEDLINE link, 2 protein links, or 1 nucleotide neighbor )
X99260
Bacteriophage B103 genomic sequence gill429229lemblX99260IBB103G [1429229]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 17 protein links, or 12 nucleotide neighbors )
AJ000741
Bacteriophage PI darA operon gil2462938lemWAJ000741IBPAJ7641 [2462938]
(View GenBank reportFASTA reportΛSN.l report.Graphical view.l MEDLINE link, 10 protein links, or 31 nucleotide neighbors )
X87420
Bacteriophage ES18 genes 24, c2, cro, cl, 18, and oL and oR operators gil 1143407lemblX87420IBPES 18GEN [1143407]
(View GenBank reportFASTA reportΛSN.l report.Graphical view,5 protein links, or 9 nucleotide neighbors )
L35561
Bacteriophage phi-105 ORFs 1-3 gil532218lgblL35561IPH50RFHTR [532218]
(View GenBank reportFASTA reportΛSN.l reportGraphical view.l MEDLINE link, or 3 protein links )
D10027
Group II RNA coliphage GA genome gil217784ldbjlDI00271PGAXX [217784]
(View GenBank reportFASTA reportΛSN.l reportGraphical view.l MEDLINE link, 3 protein links, 5 nucleotide neighbors, or 1 genome link )
V01128
Bacteriophage phi-X174 (cs70 mutation) complete genome gill5535lemb!V01128IPHIX174 [15535]
(View GenBank reportFASTA reportΛSN.l reportGraphical view,4 MEDLINE links, 11 protein links, or 26 nucleotide neighbors )
S81763 coat gene...replicase gene [bacteriophage KU1, host=Escherichia coli, group II RNA phage, Genomic RΝA, 3 genes, 120 πt] gill438766lgblS81763IS81763 [1438766]
(View GenBank reportFASTA reportASN.1 report.Graphical view, or 1 MEDLINE link )
U38906
Bacteriophage rlt integrase, repressor protein (rro), dUTPase, holin and lysin genes, complete eds gil 1353517lgblU38906IBRU38906 [1353517]
(View GenBank report.FASTA reportΛSN.l report.Graphical view,2 MEDLINE links, 50 protein links, or 3 nucleotide neighbors )
X91149
Bacteriophage phi-C31 DNA cos region gil 1107473lemblX91149IAPHIC3 IC [1107473]
(View GenBank reportFASTA reportΛSN.l reportGraphical view.l MEDLINE link, 6 protein links, or 1 nucleotide neighbor )
V00642 phage MS2 genome gill5081lemblV00642ILEMS2X [15081]
(View GenBank reportFASTA reportΛSN.l reportGraphical view,8 MEDLINE links, 4 protein links, or 20 nucleotide neighbors )
V01146
Genome of bacteriophage T7 gil431187lemb!V01146IT7CG [431187]
(View GenBank report .FAST A reportΛSN.l reportGraphical view,13 MEDLINE links, 60 protein links, 105 nucleotide neighbors, or 1 genome link )
X78401
Bacteriophage P22 right operon, orf 48, replication genes 18 and 12, nin region genes, ninG phosphatase, late control gene 23, orf 60, complete- eds, late control region, start of lysis gene 13 gil512343lemblX78401IPOP22NIN [512343]
(View GenBank reportFASTA reportΛSN.l reportGraphical view,2 MEDLINE links, 13 protein links, or 4 nucleotide neighbors )
Y00408
Bacteriophage T4 gene t for lysis protein gill5368lemWY00408IMYT4T [15368]
(View GenBank reportFASTA reportΛSN.l reportGraphical view.l MEDLINE link, 1 protein link, or 3 nucleotide neighbors )
Z26590
Bacteriophage mv4 lysA and lysB genes gil410500lemblZ26590IMV4LYSAB [410500]
(View GenBank report.FASTA reportΛSN.l report.Graphical view, or 4 protein links )
X07809
Phage phiX174 Ivsis (E) gene upstream region gil l5094lemblXCf7809IMIPHlXE [15094]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 2 protein links, or 4 nucleotide neighbors )
Z34528
Lactococcal bacteriophage c2 lysin gene gi!506455lemblZ34528ILBC2LYSIN [506455]
(View GenBank reportFASTA reportΛSN.l report.Graphical view.l MEDLINE link, 1 protein link, or 4 nucleotide neighbors )
X15031
Bacteriophage fr RNA genome gill5071lemblX15031ILEBFRX [15071]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 4 protein links, 9 nucleotide neighbors, or 1 genome link )
X80191
Bacteriophage PP7 mRNA for maturation, coat, lysis and replicase proteins gil517237lemblX80191IBPP7PR [517237]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 4 protein links, or 1 genome link )
X85010
Bacteriophage A511 ply511 gene gil853748lemblX8501CHBPA511PLY [853748]
(View GenBank report-FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 3 protein links, or 1 nucleotide neighbor )
X85009
Bacteriophage A500 hol500 and ply500 genes gil853744lemblX85009IBPA500PLY [853744]
(View GenBank reportFASTA reportΛSN.l reportGraphical view.l MEDLINE link, 3 protein links, or 4 nucleotide neighbors )
X85008
Bacteriophage A 118 holl 18 and plyl 18 genes gil853740lemblX85008IBPA118PLY [853740]
(View GenBank reportFASTA reportΛSN.l reportGraphical view.l MEDLINE link, 3 protein links, or 1 nucleotide neighbor )
Z35638
Bacteriophage phi-X174 genes for lysis protein and beta-lactamase gil520996lemblZ35638IBPLYSPR [520996]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 2 protein links, or 516 nucleotide neighbors )
J02459
Bacteriophage lambda, complete genome gil215104lgblJ02459ILAMCG [215104]
(View GenBank report.FASTA report.ASN.l report.Graphical view,87 MEDLINE links, 67 protein links, 190 nucleotide neighbors, or 1 genome link )
X87674
Bacteriophage PI lydA & lydB genes gil974763lemblX87674IB ACPI LYD [974763]
(View GenBank report.FASTA reportΛSN.l reportGraphical view.l MEDLINE link, 2 protein links, or 2 nucleotide neighbors )
X87673
Bacteriophage PI gene 17 gil974761lemblX87673IBACP117 [974761]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 1 protein link, or 1 nucleotide neighbor )
M14784
Bacteriophage T3 strain amNG220B right end, tail fiber protein, lysis protein and DNA packaging proteins, complete eds gil215810)gblM14784IFT3RE [215810]
(View GenBank reportFASTA reportΛSN.l reportGraphical view.l MEDLINE link, 9 protein links, or 10 nucleotide neighbors )
M11813
Bacteriophage PZA (from B.subtilis), complete genome gil216046lgblMl 1813IPZACG [216046]
(View GenBank reportFASTA reportΛSN.l reportGraphical view,3 MEDLINE links, 27 protein links, 17 nucleotide neighbors, or 1 genome link )
M16812
Bacteriophage K3 't' lysis gene, complete eds gil215503lgblM16812IPK3LYST [215503]
(View GenBank report.FASTA reportΛSN.l reportGraphical view.l MEDLINE link, 1 protein link, or 4 nucleotide neighbors )
J04356
Bacteriophage P22 proteins 15 (complete eds), and 19 (31 end) genes gil215265lgblJ04356IP2215P [215265]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 3 protein links, or 2 nucleotide neighbors )
J04343
Bacteriophage JP34 coat and lysis protein genes, complete eds, and replicase protein gene, 51 end gil215076lgblJ04343IJP3COLY [215076]
(View GenBank report.FASTA reportΛSN. l reportGraphical view.l MEDLINE link, 3 protein links, or 2 nucleotide neighbors )
J02482
Bacteriophage phi-X174, complete genome gil216019lgblJ02482IPXlCG [216019]
(View GenBank report.FASTA reportΛSN.l report.Graphical view,23 MEDLINE links, 11 protein links, 26 nucleotide neighbors, or 1 genome link )
M99441
Bacteriophage T4 anti-sigma 70 protein (asiA) gene, complete eds and lysis protein, 3' end gil215820lgblM99441IPr4ASIA [215820]
(View GenBank report.FASTA reportΛSN.l reportGraphical view3 MEDLINE links, 2 protein links, or 2 nucleotide neighbors )
M65239
Bacteriophage 21 lysis genes S, R, and Rz, complete eds gil215466lgblM65239IPH2LYSGEN [215466]
(View GenBank report.FASTA reportΛSN.l report,Graphical view.l MEDLINE link.3 protein links, or 1 nucleotide neighbor )
M10637
Phage G4 D/E overlapping gene system, encoding D (morphogenetic) and E
(lysis) proteins gil215427lgblM10637IPG4DE [215427]
(View GenBank reportFASTA reportΛSN.l reportGraphical view.l MEDLINE link, 2 protein links, or 12 nucleotide neighbors )
J02454
Bacteriophage G4, complete genome gil215415lgblJ02454IPG4CG [215415]
(View GenBank reportFASTA reportΛSN.l reportGraphical view,6 MEDLINE links, 11 protein links, 20 nucleotide neighbors, or 1 genome link )
J02580
Bacteriophage PA-2 (Rcoli porcine strain isolate) Rz gene, 5'end; ORF2, outer membrane porin protein (lc) and ORF1 genes, complete eds gil215366lgblJ02580IPA2LC [215366]
(View GenBank reportFASTA reportΛSN.l reportGraphical view.l MEDLINE link, 4 protein links, or 4 nucleotide neighbors )
M 14782
Bacillus phage phi-29 head morphogenesis, major head protein, head fiber protein, tail protein, upper collar protein, lower collar protein, pre-neck appendage protein, morphogenesis(13), lysis, morphogenesis(15), encapsidation genes, complete eds gil215323lgblM14782IP29LATE2 [215323]
(View GenBank report.FASTA reportASN.1 report.Graphical view.l MEDLINE link, 11 protein links, or 11 nucleotide neighbors )
M10997
Bacteriophage P22 lysis genes 13 and 19, complete eds gil215262lgbiM 10997IP221319 [215262]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 2 protein links, or 3 nucleotide neighbors )
J02467
Bacteriophage MS2, complete genome gil215232lgblJ024671MS2CG [215232]
(View GenBank report.FASTA reportΛSN.l report.Graphical view,8 MEDLINE links, 4 protein links, 20 nucleotide neighbors, or 1 genome link )
M14035
Bacteriophage lambda lysis S gene with mutations leading to nontethality of S in the plasmid pRGl gil215180lgblM14O351LAMLYS [215180]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 1 protein link, or 14 nucleotide neighbors )
U04309
Bacteriophage phi-LC3 putative holin (lysA) gene and putative murein hydrolase (lysB) gene, complete eds gil530796lgblU04309IBPU04309 [530796]
(View GenBank report.FASTA report-ASN.l reportGraphical view.l MEDLINE link, 2 protein links, or 1 nucleotide neighbor )
Table 13
NCBI Entrez Nucleotide QUERY
Key word: holin
51 citations found (all selected)
AF034975
Bacteriophage H-19B essential recombination function protein (erf), kil protein (kil), regulatory protein cIII (cIII), protein gpl7 (17), N protein (N), cl protein (cl), cro protein (cro), ell protein (ell), O protein (O), P protein (P), ren protein (ren), Roi (roi), Q protein (Q),
Shiga-like toxin A (slt-IA) and B (slt-IB) subunits, and putative holin protein (S) genes, complete eds gil2668751lgblAF034975l [2668751]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 20 protein links, or 30 nucleotide neighbors )
U52961
Staphylococcus aureus holin-like protein LrgA (lrgA) and LrgB (lrgB) genes, complete eds gill841516lgblU52961ISAU52961 [1841516]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 2 protein links, or 1 nucleotide neighbor )
U28154
Haemophilus somnus cryptic prophage genes, capsid scaffolding protein gene, partial eds, major capsid protein precursor, endonuclease, capsid completion protein, tail synthesis proteins, holin, and lysozyme genes, complete eds gill765928lgblU28154IHSU28154 [1765928]
(View GenBank report,FASTA reportΛSN.l report.Graphical view,l MEDLINE link, or 13 protein links )
AF032122
Streptococcus thermophilus bacteriophage Sfil9 central region of genome gil2935682lgblAF032122l [2935682]
(View GenBank report-FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 14 protein links, or 2 nucleotide neighbors )
AF032121
Streptococcus thermophilus bacteriophage Sfι21 central region of genome gil2935667lgblAF032121IAF032121 [2935667]
(View GenBank report,FASTA reportΛSN.l report.Graphical view,l MEDLINE- *' link, 14 protein links, or 2 nucleotide neighbors )
AF021803
Bacillus subtilis 168 prophage SPbeta N-acetylmuramoyl-L-alanine amidase
(blyA), holin-like protein (bhlA), holin-like protein (bhJB), and yolK genes, complete eds; and yolJ gene, partial eds gil2997594lgblAF021803IAF021803 [2997594]
(View GenBank report.FASTA reportΛSN. l report.Graphical view.l MEDLINE link, 5 protein links, or 1 nucleotide neighbor )
AF057033
Streptococcus thermophilus bacteriophage sfi l l gp502 (orf502), gρ284 (orf284), gP129 (orfl29), gp!93 (orf 193), gpl l9 (orf 119), gp348 (or048), gP53 (orf53), gpl 13 (orf 113), gpl04 (orf 104), gpl 14 (orf 114), gpl28 (orfl28), gpl68 (orf 168), gpl 17 (orfl l7), gpl05 (orfl05), putative minor tail protein (orf 1510), putative minor structural protein (orf512), putative minor structural protein (orflOOO), gp373 (orf373), gp57 (orf57), putative anti-receptor (orf695), putative minor structural protein (orf669), gpl49 (orfl49), putative holin (orf 141), putative holin (orf87), and lysin (orf288) genes, complete eds gil3320432lgblAF057033IAF057033 [3320432]
(View GenBank report.FASTA reportΛSN.l report.Graphical view,25 protein links, or 1 nucleotide neighbor )
U32222
Bacteriophage 186, complete sequence gil3337249lgblU32222IB 1U32222 [3337249]
(View GenBank report.FASTA reportΛSN.l report.Graphical view,6 MEDLINE links, 46 protein links, or 5 nucleotide neighbors )
AB009866
Bacteriophage phi PVL proviral DNA, complete sequence gil3341907ldbj I AB009866IAB 009866 [3341907]
(View GenBank reportFASTA reportΛSN.l report.Graphical view,63 protein links, or 1 nucleotide neighbor )
AF009630
Bacteriophage HL170, complete genome gil3282260lgblAF009630IAF009630 [3282260]
(View GenBank report,FASTA reportΛSN.l report.Graphical view,63 protein links, 3 nucleotide neighbors, or 1 genome link )
AF064539
Bacteriophage N15, complete genome
gil3192683lgblAF064539IAF064539 [3192683]
(View GenBank report,FASTA reportΛSN.l report,Graphical view,2 MEDLINE links, 60 protein links, 26 nucleotide neighbors, or 1 genome link )
AF063097
Bacteriophage P2, complete genome gil3139086lgblAF063097IAF063097 [3139086]
(View GenBank report,FASTA reportΛSN.l report.Graphical view,21 MEDLINE links, 42 protein links, 3 nucleotide neighbors, or 1 genome lin )
Z97974
Bacteriophage phiadh lys, hoi, intG, rad,and tec genes gil2707950lemblZ97974IBPHIADH [2707950]
(View GenBank report.FASTA reportΛSN.l report.Graphical view,2 MEDLINE links, 9 protein links, or 1 nucleotide neighbor )
X95646
Streptococcus thermophilus bacteriophage Sfi21 DNA; lysogeny module,
8141 bp gil22927471emblX95646IBSFI21LYS [2292747]
(View GenBank report,FASTA reportΛSN.l report.Graphical view,2 MEDLINE links, 19 protein links, or 3 nucleotide neighbors )
SEG LLHLYSINO
Bacteriophage LL-H structural protein gene, partial eds; minor structural protein gp61 (g57), unknown protein, unknown protein, structural protein (g20), unknown protein, unknown protein, major capsid protein (g34), main tail protein gpl9 (gl7), holin (hoi), muramidase
(mur), unknown protein, unknown protein, unknown protein, unknown protein, unknown protein, and unknown protein genes, complete eds; unknown protein gene, partial eds; and unknown protein, unknown protein, unknown protein, unknown protein, unknown protein, minor structural protein gp75 (g70), minor structural protein gp89 (g88), minor structural protein gp58 (g71), unknown protein, unknown protein, unknown protein, and unknown protein genes, complete eds gil lOO4337lgbllSEG_LLHLYSIN0 [1004337]
(View GenBank report,FASTA reportΛSN.l report.Graphical view,4 MEDLINE links, 31 protein links, or 1 nucleotide neighbor )
M96254
Bacteriophage LL-H holin (hoi), muramidase (mur), and unknown protein genes, complete eds gill004336lgblM962541LLHLYSIN03 [1004336]
(View GenBank report-FASTA report-ASN.l report, or Graphical view)
Y07740
Staphylococcus phage 187 ply 187 and hoi 187 genes gil2764982lemblY07740IBP187PLYH [2764982] (View GenBank report.FASTA reportΛSN.l report.Graphical view, or 2 protein links )
U88974
Streptococcus thermophilus bacteriophage 01205 DNA sequence gil2444080lgblU88974l [2444080]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 57 protein links, or 6 nucleotide neighbors )
Z99117
Bacillus subtilis complete genome (section 14 of 21): from 2599451 to
2812870 gil2634966lemblZ99117IBSUB0014 [2634966]
(View GenBank report.FASTA reportΛSN.l report.Graphical view,233 protein links, 51 nucleotide neighbors, or 1 genome link )
Z99115
Bacillus subtilis complete genome (section 12 of 21): from 2195541 to
2409220 gil2634478lemblZ99115IBSUB0012 [2634478]
(View GenBank report-FASTA reportΛSN.l report,Graphical view,244 protein links, 64 nucleotide neighbors, or 1 genome link )
Z99110
Bacillus subtilis complete genome (section 7 of 21): from 1194391 to
1411140 gil2633472lemblZ99110IBSUB0007 [2633472]
(View GenBank report,FASTA reportΛSN.l report.Graphical view,226 protein links, 31 nucleotide neighbors, or 1 genome link )
X78410
Bacteriophage phiadh holin and lysin genes gil793848lemblX78410ILGHOLLYS [793848]
(View GenBank report-FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 2 protein links, or 1 nucleotide neighbor )
Z93946
Bacteriophage Dp-1 dph and pal genes and 5 open reading frames gill934760lemblZ93946IBPDPlORFS [1934760] (View GenBank report.FASTA reportΛSN.l report.Graphical view, or 6 protein links )
AFO 11378
Bacteriophage ski complete genome gil2392824lgblAF011378IAF011378 [2392824]
(View GenBank report.FASTA reportΛSN.l report.Graphical view ,54 protein links, 2 nucleotide neighbors, or 1 genome link )
Z47794
Bacteriophage Cp-1 DNA, complete genome gil2288892lemblZ47794IBPCPlXX [2288892]
(View GenBank report.FASTA reportΛSN.l report.Graphical view3 MEDLINE links, 28 protein links, 1 nucleotide neighbor, or 1 genome link )
L35561
Bacteriophage phi-105 ORFs 1-3 gil532218lgblL35561IPH50RFHTR [532218]
(View GenBank report.FASTA reportΛSN.l report,Graphical view.l MEDLINE link, or 3 protein links )
D49712
Bacillus licheniformis DNA for ORFs, xpaL2 homologous protein and xpaLl homologous protein, complete and partial eds gill514423ldbjlD49712ID49712 [1514423]
(View GenBank report,FASTA reportΛSN.l report,Graphical view,2 MEDLINE links, or 4 protein links )
X90511
Lactobacillus bacteriophage phigle DNA for Rorf 162, Holin, Lysin, and
Rorfl75 genes gill926386lemblX90511ILBPHIHOL [1926386]
(View GenBank report-FASTA reportΛSN.l report-Graphical view ,4 protein links, or 1 nucleotide neighbor )
X98106
Lactobacillus bacteriophage phigle complete genomic DNA gill926320lemblX98106ILBPHIGlE [1926320]
(View GenBank report,FASTA reportΛSN.l report.Graphical view.l MEDLINE
link, 50 protein links, or 4 nucleotide neighbors )
U72397
Bacteriophage 80 alpha holin and amidase genes, complete eds gill763241lgblU723971B8U72397 [1763241]
(View GenBank report,FASTA reportΛSN.l report.Graphical view,2 protein links, or 2 nucleotide neighbors )
U38906
Bacteriophage rlt integrase, repressor protein (rro), dUTPase, holin and lysin genes, complete eds gil 1353517lgblU38906IBRU38906 [1353517]
(View GenBank reportJFASTA reportΛSN.l report.Graphical view,2 MEDLINE links, 50 protein links, or 3 nucleotide neighbors )
X91149
Bacteriophage phi-C31 DNA cos region gill l07473lemblX91149IAPHIC3lC [1107473]
(View GenBank report,FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 6 protein links, or 1 nucleotide neighbor )
U24159
Bacteriophage HP1 strain HPlcl, complete genome gill046235lgblU24159IBHU24159 [1046235]
(View GenBank report,FASTA reportΛSN.l report.Graphical view,6 MEDLINE links, 41 protein links, 8 nucleotide neighbors, or 1 genome link )
Z26590
Bacteriophage mv4 lysA and lysB genes gil410500lemblZ26590IMV4LYSAB [410500]
(View GenBank report,FASTA reportΛSN.l report,Graphical view, or 4 protein links )
Z70177
B.subtilis DNA (28 kb PBSX/skin element region) gill225934lemblZ70177IBSPBSXSE [1225934]
(View GenBank report-FASTA reportΛSN.l report.Graphical view32 protein links, or 4 nucleotide neighbors )
Z36941
B.subtilis defective prophage PBSX xhlA, xhlB, and xylA genes gil535793lemblZ36941IBSPBSXXHL [535793]
(View GenBank report.FASTA reportΛSN. l report.Graphical view ,4 protein links, or 5 nucleotide neighbors )
X89234
L.innocua DNA for phagelysin and holin gene gil 1134844lemblX89234ILICPLYHOL [ 1134844]
(View GenBank report.FASTA report-ASN.l report.Graphical view.l MEDLINE link, 2 protein links, or 4 nucleotide neighbors )
X85010
Bacteriophage A511 ply511 gene gil853748lemblX85010IBPA511PLY [853748]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 3 protein links, or 1 nucleotide neighbor )
X85009
Bacteriophage A500 hol500 and ply500 genes gil853744lemblX85009IBPA500PLY [853744]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 3 protein links, or 4 nucleotide neighbors )
X85008
Bacteriophage A118 holll8 and plyllδ genes gil853740lemblX85008IBPA118PLY [853740]
(View GenBank report,FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 3 protein links, or 1 nucleotide neighbor )
L34781
Bacteriophage phi 11 holin homologue (ORF3) gene, complete eds and peptidoglycan hydrolase (lytA) gene, partial eds gil511838lgblL347811BPHHOUN [511838]
(View GenBank report,FASTA reportΛSN.l report,Graρhical view.l MEDLINE link, 4 protein links, or 2 nucleotide neighbors )
U11698
Serratia marcescens SM6 extracellular secretory protein (nucE), putative phage lysozyme (nucD), and transcriptional activator (nucC) genes, complete eds gil509550lgblU11698ISMU11698 [509550]
(View GenBank report,FASTA reportΛSN.l report.Graphical view.l MEDLINE
link, 3 protein links, or 1 nucleotide neighbor )
U31763
Serratia marcescens phage-holin analog protein (regA), putative phage lysozyme (regB), and transcriptional activator (regQ genes, complete eds gil965068lgblU31763ISMU31763 [965068]
(View GenBank report,FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 3 protein links, or 1 nucleotide neighbor )
X87674
Bacteriophage PI lydA & lydB genes gil974763lemblX87674IBACPlLYD [974763]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 2 protein links, or 2 nucleotide neighbors )
L48605
Bacteriophage c2 complete genome gill l46276lgblL48605IC2PVCG [1146276]
(View GenBank report,FASTA reportΛSN.l report.Graphical view3 MEDLINE links, 39 protein links, 3 nucleotide neighbors, or 1 genome link )
L33769
Bacteriophage WL67 DNA polymerase subunit (ORF3-5), essential recombination protein (ORF13), lysin (ORF24), minor tail protein
(ORF31), -erminase subunit (ORF32), holin (ORF37), unknown protein (ORF
1-2,6-12,14-23,25-3033-36), complete genome gil522252lgblL33769IL67CG [522252]
(View GenBank report,FASTA reportΛSN.l report,Graphical view.l MEDLINE link, 37 protein links, 2 nucleotide neighbors, or 1 genome link )
131348
Bacteriophage Tuc2009 integrase (int) gene, complete eds; lysin (lys) gene, 3' end gil508612lgblL31348ITU2INT [508612]
(View GenBank reportFASTA reportΛSN.l report,Grap__ical view.2 MEDLINE links, 3 protein links, or 3 nucleotide neighbors )
131364
Bacteriophage Tuc2009 holin (S) gene, complete eds; lysin (lys) gene, _ _ complete eds gil496281lgblL31364ITU2SLYS [496281]
(View GenBank report,FASTA reportΛSN.l report,Graphical view.l MEDLINE link, 2 protein links, or 1 nucleotide neighbor )
1 1366
Bacteriophage Tuc2009 structural protein (mp2) gene, complete eds gil496278lgb!I31366ITU2MP2A [496278]
(View GenBank report-FASTA reportΛSN.l report-Graphical view.l MEDLINE link, 2 protein links, or 1 nucleotide neighbor )
131365
Bacteriophage Tuc2009 structural protein (mpl) gene, complete eds gil496276lgblL31365ITU2MPlA [496276]
(View GenBank report,FASTA reportΛSN.l report.Graphical view.l MEDLINE link, or 1 protein link )
U04309
Bacteriophage phi-LC3 putative holin (lysA) gene and putative murein hydrolase (lysB) gene, complete eds gil530796lgblU04309IBPU04309 [530796]
(View GenBank report.FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 2 protein links, or 1 nucleotide neighbor )
Table 14
NCBI Entrez Nucleotide QUERY Key word: bacteriophage and kil 5 citations found (all selected)
AF034975
Bacteriophage H-19B essential recombination function protein (erf), kil protein (kil), regulatory protein cIII (cIII), protein gpl7 (17), N protein (N), cl protein (cl), cro protein (cro), ell protein (ell), O protein (O), P protein (P), ren protein (ren), Roi (roi), Q protein (Q),
Shiga-like toxin A (slt-IA) and B (slt-IB) subunits, and putative holin protein (S) genes, complete eds gil2668751lgblAF034975l [2668751]
(View GenBank report,FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 20 protein links, or 30 nucleotide neighbors )
X15637
Bacteriophage P22 P(L) operon encompassing ral, 17, kil and arf genes gill5646lemblX15637IPOP22PL [15646]
(View GenBank report,FASTA reportΛSN.l report.Graphical view.l MEDLINE link, 7 protein links, or 2 nucleotide neighbors )
J02459
Bacteriophage lambda, complete genome gil215104lgblJ02459ILAMCG [215104]
(View GenBank reportFASTA reportΛSN.l reportGraphical view,87 MEDLINE links, 67 protein links, 190 nucleotide neighbors, or 1 genome link )
M64097
Bacteriophage Mu left end gil215543lgblM64097IPMULEFTEN [215543]
(View GenBank report,FASTA reportΛSN.l reportGraphical view.2 MEDLINE links, 39 protein links, or 15 nucleotide neighbors )
M18902
Bacteriophage D108 kil gene encoding a replication protein, 31 end; and containing three ORFs, complete eds gil 166191 lgblM18902ID18KIL [166191]
(View GenBank reportFASTA reportΛSN.l report.Graphical view.l MEDLINE link, 1 protein link, or 3 nucleotide neighbors ) - -
Table 15
Table 16
Phage 44AHJD complete genome sequence. 16668 nucleotides.
1 tccatttctt tactaaactt aaaaatgctg tgcaacaact taaccaactt atctaaccta ttacatattc
71 atcaaataca aaatttatgt atctattgac ttttattcaa aattatgatt tcaacatata ataaaattaa
141 tttacttatt taaatattct atgatataat tagttataaa atatttggag gtgtataaat gacagaattt
211 gatgaaatcg taaaaccaga cgacaaagaa gaaacttcag aatcaactga agaaaattta gaatcaactg
281 aagaaacttc agaatcaact gaagaatcaa ctgaagaatc aactgaagaa tcaactgaag ataaaacagt
351 agaaacaatc gaagaagaaa atgaaaacaa attagaacct actacaacag atgaagatag ttcgaaattt
421 gaccctgttg tattagaaca acgtattgct tcattagaac aacaagtgac tactttttta tcttcaoaaa
491 tgcaacaacc acaacaagta caacaaacac aatcagatgt aacagaatca aacaaagaag ataacgacta
561 ttcagatgaa gaactagttg ataagttaga tttagattag gaggaattta aacatgtatg agggaaacaa
631 oatgcgttct atgatgggta catcatatga agattcaaga ttaaataaac gaacagaatt aaatgaaaac
701 atgtcaattg atacaaataa aagtgaagat agttatggtg tacaaattca ttcactttca aaacaatcat
771 ttacaggtga cgttgaggag gaataataaa ttatggcaca acaatctaca aaaaatgaaa ctgcactttt
841 agtagcaaag tcagctaaat cagcgttaca agattttaat catgattatt caaaatcttg gacatttggc
911 gacaaatggg ataattcaaa tacaatgttc gaaacatttg taaataaata tttattccct aagattaatg
981 agactttatt aatcgatatt gcattaggta atcgttttaa ttggttagct aaagagcaag attttattgg
1051 acaatatagt gaagaatacg tgattatgga cacagtacca attaacatgg acttatctaa aaatgaggaa
1121 ttaatgttga aacgtaatta tccacgtatg gcaactaagt tatatggtaa cggaattgtg aagaaacaaa
1191 aattcacatt aaacaacaat gatacacgtt tcaatttcca aacattagca gacgcaacta attacgcttt
1261 aggtgtatac aaaaagaaaa tttctgatat taatgtatta gaagaaaaag aaatgcgtgc aatgttagtt
1331 gattactcat tgaatcaatt atccgaaaca aatgtacgta aagcaacatc aaaagaagat ttagcaagca
1401 aagtttttga agcaatccta aacttacaaa acaacagtgc taaatataat gaagtacatc gtgcatcagg
1471 tggtgcaatt ggacaatata caactgtatc aaaattaaaa gatattgtga ttttaacaac agattcatta
1541 aaatcttatc ttttagatac taagattgca aacacattcc agattgcagg cattgatttc acagatcacg
1611 ttattagttt tgacgactta ggtggcgtgt ttaaagtaac aaaagaattt aagttacaaa accaagattc
1681 aattgacttt ttacgtgcgt atggagatta tcaatcacaa ttaggagata caattccagt tggtgctgta
1751 tttacttatg atgtatctaa acttaaagag tttactggca acgttgaaga aattaaacca aaatcagatt
1821 tatatgcgtt tattttggat attaattcaa ttaaatataa acgttacaca aaaggtatgt taaaaccacc
1891 attccataac cctgaatttg atgaagttac acaotggatt cattactatt catttaaagc cattagtcca
1961 ttctttaata aaattttaat tactgaccaa gat taaatc caaaaccaga ggaagaatta caagaataaa
2031 aggagcgtaa aatatgaaca acgataaaag aggtttaaac gttgagttat caaaggaaat cagcaaaaga
2101 gttgttgaac atcgcaacag atttaaacgt cttatgttta atcgttattt ggaattttta ccgctactaa
2171 tcaactatac caatcgtgat acggttggta tagattttat tcagttagaa tcagctttaa gacaaaacat
2241 taatgtagtt gttggtgaag ctagaaataa gcaaattatg attcttggtt atgtaaataa cacttaottt
2311 aatcaagcac caaatttttc atcaaacttt aatttccaat ttcaaaaacg attaactaaa gaagatatat
2381 attttattgt aoctgactat ttaatacctg atgattgtct acaaattcat aagctatatg ataactgtat
2451 gagtggtaac tttgttgtca tgcaaaataa accaattcaa tataatagtg atatagaaat tatagaacat
2521 tatactgatg aattagcaga agttgcttta tctcgctttt ctttaatcat gcaagcaaaa tttagcaaga
2591 tatttaaatc agaaattaat gacgagtcaa tcaatcaact tgtgtccgaa atatataacg gtgcaccatt
2661 tgttaaaatg tcacctatgt ttaatgcaga tgacgatatc attgatttaa caagtaatag cgtaatccca
2731 gcattaactg aaatgaaacg ggaatatcaa aacaaaatta gtgaattaag taactattta ggcattaatt
2801 cattagccgt tgataaagaa agcggtgttt cagacgaaga ggcaaaaagt aatcgtggat ttaccacatc
2871 aaacagtaat atctatttaa aaggtcgtga accaattacg tttttatcaa agcgttatgg tttagatatt
2941 aaaccgtatt acgatgatga aacaacgtct aaaatatcaa tggtagacac actttttaaa gatgaaagca
3011 gtgatataaa tggctagata cacaatgact ttatacgatt tcattaaatc agaattgatt aaaaaaggtt
3081 tcaatgaatt tgtaaatgat aataaattaa cgttttatga tgatgaattt caattcatgc aaaaaatgct
3151 gaagttcgac aaagacgttt tagctatcgt taatgaaaaa gtatttaaag gtttttcatt gaaagatgaa
3221 ttatcagatt tactttttaa aaaatcattt acgattcatt ttttagatag agaaatcaac agacaaacag
3291 ttgaagcatt tggcatgcaa gtgattactg tatgtattac acatgaggat tatttaaatg tggtttattc
3361 atcaagtgaa gttgaaaaat acttacaatc acaaggcttc acagaacaca atgaagatac aacaagtaac
3431 actgatgaaa catcgaatca aaatgctaca tctttagaca attcaactgg catgactgca aacagaaacg
3501 cttatgtgtc attaccacaa agtgaggtta acattgatgt tgataataca acgttacgat tcgctgataa
3571 taatacgatt gataacggta aaactgtgaa taaatcgagt aacgaaagta atcaaaacgc aaaacgtaat
3641 caaaatcaaa aaggtaatgc aaaaggtaca caattcacta agcagtattt aattgataat attgataaag
3711 cgtacgattt aagaaagaaa attttaaatg aatttgataa aaaatgtttt ttacaaattt ggtagaggtg
3781 gttaaataat ggcatataat gaaaacgatt ttaaatattt tgatgacatt cgtccatttt tagacgaaat
3851 ttataaaacg agagaacgtt atacaccgtt ttacgatgat agagcagatt ataatactaa ttcaaaatca
3921 tattatgatt atatttcaag attatcaaaa ctaattgaag tattagcacg tcgtatttgg gactatgaca
3991 atgaattaaa aaaacgtttc aaaaattggg acgacttaat gaaagcattt ccagagcaag cgaaagactt
4061 atttagaggt tggttaaacg acggtacgat tgacagtatt attcatgacg agtttaaaaa atatagcgca
4131 ggattaacat cggcatttgc tttatttaaa gttactgaaa tgaaacaaat gaatgacttt aaatcagaag
4201 ttaaagactt aattaaagat attgaccgtt tcgttaatgg gtttgaatta aatgagcttg aaccaaagtt—
4271 tgtgatgggc tttggtggta ttcgcaacgc agttaaccaa tctattaata ttgataaaga aacaaatcac
4341 atgtactcta cacaatccga ttctcaaaaa cctgaaggtt tttggataaa taaattaaca cctagtggtg
4411 acttaatttc aagcatgcgt attgtacagg gtggtcatgg tacaacaatc ggattagaac gtcaatccaa
4481 tggtgaaatg aaaatctggt tacatcacga tggtgttgca aaactgttac aagtcgcata taaagataat
4551 tatgtattag atttagaaga ggctaaaggt ttaacagatt atacaccaca gtcactttta aacaaacaca
4621 catttacacc gttaattgat gaagcaaatg acaaactcat tttaagattc ggtgacggaa caatacaggt
4691 tcgttcaaga gcagacgtaa aaaatcacat tgataatgta gaaaaagaaa tgacaattga taattcagaa
4761 aacaatgata a cgttggat gcaaggcatt gctgttgatg gtgatgattt atactggtta agtggtaaca
4831 gttcagttaa ttcacatgtt caaatcggta aatattcatt aacaacaggt caaaagattt atgattatcc
4901 atttaagtta tcatatcaag acggtattaa tttcccacgt gataacttta aagagcctga gggtatttgc
4971 atttatacaa atccaaaaac aaaacgtaaa tcgttattac ttgctatgac aaacggcggt ggtggaaaac
5041 gtttccataa tttatatggt ttcttccaac ttggtgagta tgaacacttt gaagcattac gcgcaagagg
5111 ttcacaaaac tataaattaa caaaagacga cggtcgtgca ttatctattc cagaccatat cgacgattta
5181 aatgacttaa cgcaagctgg tttttattat attgacgggg gtactgcaga aaaacttaag aatatgccaa
5251 tgaatggtag caagcgtata attgacgctg gttgtttcat taatgtatac cctacaacac aaacattagg
5321 tacggttcaa gaattaacac gtttctcaac aggtcgtaaa atggttaaaa tggtgcgtgg tatgacttta
5391 gacgtattta cgttaaaatg ggattatgga ttatggacaa caatcaaaac tgacgcacca tatcaagaat
5461 atttggaagc aagtcaatac aataactgga ttgcttatgt aacaacagct ggtgagtatt acattacagg
5531 taaccaaatg gaattattta gagacgcgcc agaagaaatt aaaaaagtgg gtgcatggtt acgtgtgtca
5601 agtggtaacg cagtcggtga agtaagacaa acattagagg ctaatatatc ggaatataaa gaattcttca
5671 gtaatgttaa tgcggaaaca aaacatcgtg aatatggttg ggtagcaaaa catcaaaaat aggagtgata
5741 taaatgaaat cacaacaaca agcaaaagaa tggatatata agcatgaggg ggcaggtgtt gactttgatg
5811 gtgcatatgg atttcaatgt atggacttat cagttgctta tgtgtattac attactgacg gtaaagttcg
5881 catgtggggt aatgσtaaag acgcgataaa taatgacttt aaaggtttag cgacggtgta taaaaataca
5951 ccgagcttta aacctcaatt aggggacgtt gctgtatata caaatggaca atatggacat attcaatgtg
6021 tgttaagtgg aaatcttgat tattatacat gcttagaaca aaactggtta ggcggcggtt ttgacggttg
6091 ggaaaaagca accattagaa cacattatta tgacggtgta actcacttta ttagacctaa attttcaggt
6161 agtaatagca aagcattaga aacatcaaaa gtaaatacat ttggaaaatg gaaacgaaac caatacggca
6231 catattatag aaatgaaaat ggtacattta catgtggttt tttaccaata tttgcacgtg tcggtagtcc
6301 aaaattatca gaacctaatg gctattggtt ccaaccaaac ggttatacac catataacga agtttgttta
6371 tcagatggtt acgtatggat tggttataac tggcaaggca cacgttatta tttaccagtg cgccaatgga
6441 atggaaaaac aggtaatagt tacagtgttg gtattccttg 99999tgttc tcataatggg tattttagcc
6511 tttttctttg aatttagttg gaaaagatac aaataagagg tgtaaacaat ggctgataga atcgtaagaa
6581 gtttaagaca agttgaaaca attgaacgtt tattggagga aaaaaatgag aaagttaacg aattttaagt
6651 ttttctataa cacaccgttt acagactatc aaaacacgat tcattttaat agtaataaag aacgtgatga
6721 ttatttttta aatggtcgtc attttaaatc gttagactat tcaaaacaac cgtataattt tatacgtgat
6791 agaatggaaa tcaatgttga tatgcagtgg catgacgcac aaggtattaa ctacatgacg tttttatcag
6861 attttgagga tagaagatat tacgcttttg taaaccaaat cgaatacgtg aatgacgttg tggttaaaat
6931 atattttgtc a tgatacca ttatgacgta tacacaaggg aatgtattag agcaactctc aaacgtcaat
7001 at gaacgtc aacatttatc aaaacgcacg tataactata tgttaccaat gttacgtaat aatgatgatg
7071 tgttaaaagt atcaaataaa aactatgttt ataaccaaat gcaacaatat ttggaaaatt tagtattatt
7141 ccagtcaagc gctgatttat caaagaaatt tggtactaaa aaagagccaa acttagatac gtcaaaaggt
7211 acgatttatg acaatatcac atcaccagtc aacttatacg ttatggaata tggtgacttt attaacttta
7281 tggataaaat gagtgcctat ccatggatta cgcaaaactt tcaaaaggtt caaatgttac ctaaagactt
7351 tattaataca aaagacttag aggacgttaa aaccagtgaa aaaattacag gattaaaaac attaaaacag
7421 gg ggtaaat caaaagaatg gagtctaaaa gatttatcat taagtttctc aaatcttcaa gagatgatgt
7491 tatctaaaaa agatgaattt aaacatatga tacgtaatga gtatatgaca attgaatttt atgactggaa
7561 tggaaatacg atgttactcg acgctggtaa gatttcacaa aaaactggtg ttaagttacg tacaaaatca
7631 attattggtt atcataatga agttcgagta tatccagtag attataacag tgctgaaaac gacagaccaa
7701 tactcgctaa aaataaagaa atattgattg atacgggttc attcttaaat acaaatataa catttaatag
7771 ttttgcacaa gtaccaatat taatcaataa tggtatctta ggacaatcac aacaagccaa ccgacaaaaa
7841 aatgcagaaa gtcaattaat tacaaatcgt attgataatg tattaaatgg tagcgacccg aaatcacgct
7911 tttatgacgc tgtgagtgta gcaagtaatt taagtccaac tgctttattt ggtaagttta atgaagaata
7981 taatttctac aaacaacaac aagctgaata taaagattta gccttacaac caccttctgt aactgaatca
8051 gaaatgggca acgcattcca aattgcgaat agcattaacg gtttaacgat gaaaattagt gtaccgtcac
8121 ctaaagaaat tacattttta caaaaatatt atatgttgtt tggttttgaa gtgaatgact ataattcatt
8191 tattgaacca attaacagta tgactgtttg caattattta aaatgtacag gtacgtatac t tacgtgac
8261 atcgacccca tgttaatgga acaattaaaa gcaattttag aatctggtgt aagattttgg cataatgacg
8331 gttcaggtaa tccaatgtta caaaatccat taaataacaa atttagagag ggggtataat atgaacgaag
8401 taaaattcag atttacagac tcagaagcgt ttcacatgtt tatatacgct ggggatttaa aattactcta
8471 ctttttattt gtattaatgt tcgttgatat tattacaggt atttcaaaag caattaaaaa taataactta
8541 tggtcaaaaa aatcaatgag aggattttct aaaaaat at tgatattctg tattatcatt ttagcaaaca
8611 tcattgacca gattttacaa ttaaaaggtg gtctactcat gattacaata ttttattata ttgcaaatga
8681 gggactttct attgtagaaa attgtgcaga aatggacgta ttagtaccag aacaaattaa agataaatta
8751 agagtcatta aaaatgatac tgaaaagagt gataacaatg aacgatcaag agaagataga taaatttacg
8821 cattcctata ttaatgatga ttttggttta acgatagacc agttagtccc taaagtaaaa ggatatgggc
8891 gctttaatgt atggcttggt ggtaatgaaa gtaaaatcag acaagtatta aaagcagtaa aagagatagg
8961 tgtttcacct actctttttg ccgtatatga aaaaaatgag ggttttagtt ctggacttgg ttggttaaac
9031 catacgtctg cacgtggtga ttatttaaca gatgctaaat tcatagcaag aaagttagta tcacaatcaa
9101 aacaagctgg acaaccgtct tggtatgacg caggtaacat cgtccacttt gtaccacaag acgtacaaag
9171 aaaaggtaat gcagattttg caaaaaatat gaaagcaggt acaattggac gtgcatatat tccattaaca
9241 gcagctgcta cttgggcggc atattatcct ttaggtttga aagcatcata taacaaagta caaaactatg
9311 gtaatccatt tttagacggt gcgaatacta ttctagcttg gggtggtaaa ttagacggta aaggtggatc
9381 acctagtgat tcgtctgaca gtggtagtag tggtgacagt ggtagttcac tactcgcttt agcaaaacaa
9451 gccatgcaag aattattaaa aaaaatacaa gacgcattac aatgggacgt tcatagtatt ggtagtgata
9521 aattttttag taatgattat tttacattag aaaaaacatt taacaacaca tatcatatta aaatgacgat..
9591 tggtttactt gattcattaa aaaaactgat tgatagcgtt caagtagata gtgggagtag tagttctraat
9661 cctactgatg atgacggaga ccataaacca attagtggta aatcagtcaa gccaaatgga aaaagtggtc
9731 gtgtgattgg tggtaactgg acatatgcac agttaccaga aaaatataaa aaagcaattg gtgtaccttt
9801 attcaaaaaa gaatacttat acaaaccagg taacatattt cctcaaacgg gtaatgcagg acaatgtaca
9871 gaattaacat gggcgtatat gtcacaacta catggtaaaa gacaacctac cgacgacggt caaataacaa
9941 acggtcagcg tgtatggtac gtctataaaa agttaggtgc aaaaacaaca cataatccaa cagtaggtta
10011 tggtttctct agtaaaccac catacttaca agcaactgca tatggtattg gtcacacagg tgttgttgta
10081 gcagtttttg aagatggttc gtttttagtt gcaaactata atgtaccacc atatgttgca ccatcacgtg
10151 tggtattgta tacactcatt aatggcgtac caaataatgc tggtgataat attgtattct ttagtggtat
10221 tgcttaatta actatgctat aatgaacaca tgctagtaat gctagtaaat aaaatacaaa acataatcaa
10291 ttttcgtaca catttttcat gttatctcaa aaagaaaagg agactgttat tttaacagtt gccttttttt
10361 atttcatcat gttcacgttt taatatatgc aaatcagatt tgttatgtac tgaacgttca actggaaata
10431 agtcgttaag tgaaaatgaa ccgatgtcac tttcaatata aagaatatca tcaaattgac tatggtcgaa
10501 attttctcta gcgtctttta atataaattc acgtttcata ttaagttcat cagtaaaata ttcatcatat
10571 acattaccac atacaatttc agttttagac ggatatatcg atattgtacc ttgctcatta tagatacttt
10641 tattgttttc aataatggca ccgtcaaaga attgttcacg tacaaaggtt tcaaaatcga cgcttgtatc
10711 aaaggcgttt ttcggtatac cagcagaagc aattttaatc tttccattca cttcatatgc atatttctta
10781 tgattcagta caaacatctt atctatctgt tcgttttcaa tatcccattt acctaaggct atcgggtcga
10851 ataaactggg gttcaataag ggtttaacaa cggatttcat atacaaacta tcagtatcgc aataaataaa
10921 attgtcgtca atttcacttt ccgttaagta ttggaaagga accaataagt tatacaatga acgtgatgtg
10991 acaaatgtag agaataatat attacgttca gtgtttttgt aaccgttaat gatattgtat agttcattgt
11061 tatcatctaa acggaataag ttaaaatgtg aacgtaatgc aggtatgcca tataatccat ttaaaacgac
11131 tttagataac ataacctcct catttgagta tgggtgttcg ttgatatcat cagtaatgtg atagtcgtaa
11201 ggtgatgtca tattgatttt gttttttaac ttaccttgtg ttttaataaa atagttttga aaaataatat
11271 cacgtgcatg aaagtattca cattcatata taacaaacga a taacacgt atatgcatgc aatcaatacc
11341 cgtaatgtct tgaatcattc ttaatgtatt tgtattgata ttaacgtaat cattatcatt attatagtat
11411 tttacaatca tttgacgtaa tacacgtgat ttaattttaa ttaataaatc atcgttaaat acatctttat
11481 caatcttata taatgaaaaa taattgtcat catctaaaaa agtagggatt aacgttggtt ctgaatagtg
11551 ttcgtaaaag tataaccatg ttggaatttt ttcatgatac atcacataag gataactcga attgatgtca
11621 atagaaaaac aaggctcatc aattagtttg tttatgtatt tggtgttata catatttaaa ccaccacgat
11691 agaatgattt aatatagtca taaaaattca tatcatggaa atgataatgt gtataagata ttttaatatc
11761 ttgatattgg ttgagtaact gaaaacgtgt catttcatta ttcaagtaag attccataat attcaatgaa
11831 aatgttaatt tgttatagtc aaaatttgga aatatatcac tataatgaat atggcacata cctaatataa
11901 tcacgtcatt atgaatgtat gtaagttgtt caggtgtgag ttttgcaaaa catttcacag catagtcata
11971 ggcttcacta tcattcatat cattatcttt atcaaaaatc gtataattaa aatctgtttt aagttgtgat
12041 tctgttaaat aaccaccatc aagtaatttc ttacctaatg ttgcaattga tgtattggtt ttcataaagt
12111 tatcaataat attaaattta aaaccattta aaaacattgt taaatctaaa ttgattgaag atttaacacg
12181 tttttctaaa attacatttt gatttttggc taaaatagta gcctctttca tttttaatgt gtgttcattt
12251 tcttctgcag attttaaata tatattttcg cgtgtaatat tatcaaaata acgcatggtg tctttaagta
12321 aaaaatgatt atcgtattta ttacagttat gtgcaatcat gataatatct gtttttgatt ttgtgattgt
12391 atcacgtctt ttcacatacg tataaaatgc gtcataaaaa gattcgaaac tcggaaatac ttcaacatca
12461 atttcataac cattaaacca accaattgct acagaataag taacgttttt atatttggtt ggtttttttc
12531 gtccgttaac tttattgtac gctaatgttt ctatatccca gtataaaatc attcgacgtt catgtttatg
12601 atattgcatg cattctagta atcccataat cttacacacc ttttataagc catattgttt cattagatac
12671 tttttcgtat tctctatata gttatcttcg tatatttttt cttttctttc aaactcactc atatttttct
12741 tcatttcatt ttttatatga aattttataa ttttattcat atctaaatat aaatatctat cattatcaac
12811 cacgtaattt ttagagtaag cattgtcaaa atgtaaattg cttggattgt agtaataacg ttccatgttt
12881 tctttataaa acatatcatc acgtaaatag gtaacatgat tgtctatatc cctaatttta gtacaaaatt
12951 catattgttt tgtatatggt acaacgataa tatttgtcat aaaagtagtt acattataca tgactttaat
13021 atatttatca tcagttttga tatagaagaa atcaccgttt tgattgatgt gatttcttaa attatcatcc
13091 gccaaattat attcgttaaa ttcaaattct ccagttgtca tagcgtcgtc atttgaatta aacgcacgtg
13161 tgttacgttt ttcattcacg taatcgtttc gtcgcatttc taaaaaaatg tttttgtaaa gtcttgatgt
13231 attcatttta tgcttttgta ataaattgta tatatttaaa ttggataata taggacttga aaagttgact
13301 gcattaccta gtaaaaacat tttagggaat ccaatataat caacgttacc atggttacgg tcgattgatt
13371 catatattgt ttttaactta tcccactcat caattaaata atcatcttca agtgctaaaa actcatcata
13441 tataataata ggatagtgtt ttaaaaagtt agaatgatat tttaaatcag tggcactatt caaatctgta
13511 atcacaccaa tttctttatc ttgatagata atagctaaat agtccctagc acttctgaac gtgacacgtt
13581 ttgatttaaa tagtggattt tcatctatga tttcttcaat aaaatcacgg taagcgtcac gtaatgtata
13651 atgacgtgat aataaagtaa attttatatc aagtttaata gctaaataaa taaaaaatga aacatagttg
13721 aacgattttc catcagaacg gtttgaaata gatatataat aatctatatc atcattcata agtt atcaa
13791 ctaattctat ttgattatac ttatctggga ttttttttct gacatgattg acagcatttt gataatctct
13861 taccatgtct aaacgatttt gttttaccat gtttttgctc cttgtaatag tttatgatgt cgtttacagt
13931 gttaaattta ttcgtcaaat gttgcataat ataaaaagtt atacctcaca tcttcatcat caatatttgt
14001 cactggtcta tctgatttac caatttcttt atataaagta tcgatttctt taatatattt atacattgaa
14071 gaattattat ttttagcttg taaattatat aaagcgtatt tatgcttttt agcgttttta ttattagaat
14141 catcattacg gttatatatt tcaagaatat aatttaattt tttatgtctt gaacctctta ccaatgatac
14211 agcatttaca tatgatacgt ttctttcttt aggaaaatag ggcagatgtg caaaatgttt ccatgtgtca
14281 atgtacgcct cttgtaaatc tttatcatca aatttaaaat taacattact aaaatcattt aaaaataaat
14351 ctttttcttg ctcttttcta gcttctcttt cttttttcca tctatccatt tcagacgtat gtctaaccaa
14421 tgttatcaac ctccatataa agcataaata accattaaaa agataatata gaatataatc aatgtagtga
14491 ataaaacacc aaatgacacg cgtatatgca gtgtcataag tatgataagt gtaattaaaa atgctaaaag
14561 gaaaacaatg gctatgttta ataggttatt catggtcaat cactttccca ttatcgtata tgactttgtt
14631 ttgataaata atcattaatt cgctttcaag aggtttatca aaatttgata atacgtcgtc aattgtaacg
14701 tttaataaaa tttctcttat taattcatta cttaaataat ttctataata aaatacaagt atattaaaaa
14771 catgtttttt aatatcaatg tcgatatcta acgtaaataa ctctttttca atttcaaaat catcatattg
14841 tttgtcaaac tcaatataca catcacccat atttattttt actatacatt ttttattaga tgaagtaaat
14911 ttttcaaatt tatcattata ataatctcta tttgttaaaa ggtaataaat taaattattt aatctaaaa
14981 tagttttaat tttcattttt atatctcctt aatgtattct atgatatacg cgtatttttt agtgaacagg
15051 ttatattcat aatatgaata tacaacttta gcgtcatata aatcttcaaa cattgagatt tgatgtggaa
15121 aatgtccttt aatctcatcg caatataata ataccgtttt gtatttacgt tccatttaaa cacctcataa
15191 aaaatagggg ataagtatcc cctatgaaat tgtattaaaa tgatacttga ccaaaattga ttgagtaacc
15261 tttttgacct tttttgtttt catattcata aattgtgaat tgaacttctc cagcattgat aatgtcaaca
15331 acgtcctcat ctgctctcat ttctttaatt aattctgtta agtggttcgg taagtttacg ttatagtcat
15401 cagtgacgat aacaccttgt tcaccgaatt ttgattcttt gtttgtgaat aatgctctaa cgatatactc
15471 ttttttcata ccgtattttt ctactaattc tgatagtttg ataaattctc tttctttttc ctcaaattca
15541 aatctcgcta atgtgttttg gtgtcttgat aaaatatctt ttacgtttgt cattttattt ctcctcttat
15611 ttaaattatt tgctttctgc aattgcgatt tgtagtaaat cattgtaata aacttgaatt gttttcgttg
15681 tgcgtgtagt ggacaatagt ttacatgtgt ctggtaataa ttcttttgct tgtgttttgg ttaaatgata
15751 ctcgtgaagt ggtaaaaatt cctcaatgta ttcattatca tcatctaagt aatgaagtat ataacctttg
15821 acacgtaagg taacaatgtc gtcaactttc attattatat cactcctttc taaaaaacgt aaacgttata
15891 cgtttcataa aatcctttat gcatattcca ttgttctatt gggtcatcac cagcaatata agacaatatt
15961 gattctggtt tagtttcgtt gtttagttca tcatttaaga attgaacaac agaactatta tagtttaata
16031 atagttgttg gcaagccgat aataagttaa ttgcattgtc aaatgtataa gctggattcc attgaatcag
16101 tttattgaat agttgcaaca tttcagtata ggcttgtcct ttttcttctg gtgcattatc aacattaacc
16171 attattatca cttcctaata aagttgaaat tacgcgtaaa acagaattat gatttaaatc ttcaatttca
16241 tcaatgtcaa catcataaaa tgaaatttca ttttctgttc tatcaaataa cgctatacat aaacttccat
16311 tcttaaaacg aaaaacatgc ttcaactcaa tgttttttgt ttcattttcc atttttgtta ctccttgttt
16381 tgattacata cttagtatag caaacgttta aaagttttgt caatagtttt tcttaaaaaa gtttaaataa
16451 ttttaaaact actatttaat agaagaaata agattttaag ttcaaatcat aattttgaat aaaagtcaat
16521 agatacataa attttgtatt tgatgaatat gtaataggtt agataagttg gttaagttgt tgcacagtat
16591 ttttaagttt agtaaagaaa tgataagtaa atttataagt tttgatttgt ataatcgttt attttaaacc
16661 ggtggggt
Table 17
Table 18 Predicted amino acid sequences
44AH DORF001
12627 atgggattactagaatgcatgcaatatcataaacatgaacgtcgaatgattttatactgggatatagaaacattagcgtacaat
1 M G L L E C M Q Y H K H E R R M I Y W D I E T L A Y N
12543 aaagttaacggacgaaaaaaaccaaccaaatataaaaacgttacttattctgtagcaattggttggtttaatggttatgaaatt
29 K V N G R K K P T K Y K N V T Y S V A I G F N G Y E I
12459 gatgttgaagtatttccgagtttcgaatctttttatgacgcattttatacgtatgtgaaaagacgtgatacaatcacaaaatca
57 D V E V F P S F E S F Y D A F Y T Y V K R R D T I T K S
12375 aaaacagatattatcatgattgcacataactgtaataaatacgataatcattttttacttaaagacaccatgcgttattttgat
85 K T D I I M I A H N C N K Y D N H F L K D T M R Y F D
12291 aatattacacgcgaaaatatatatttaaaatctgcagaagaaaatgaacacacattaaaaatgaaagaggctactattttagcc
113 N I T R E N I Y L K S A E E N E H T K M K E A T I L A
12207 aaaaatcaaaatgtaattttagaaaaacgtgttaaatcttcaatcaatttagatttaacaatgtttttaaatggttttaaattt
141 K N Q N V I L E K R V K S S I N D L T M F L N G F K F
12123 aatattattgataactttatgaaaaccaatacatcaattgcaacattaggtaagaaattacttgatggtggttatttaacagaa
169 N I I D N F M K T N T S I A T L G K K L L D G G Y T E
12039 tcacaacttaaaacagattttaattatacgatttttgataaagataatgatatgaatgatagtgaagcctatgactatgctgtg
197 S Q K T D F N Y T I F D K D N D M N D S E A Y D Y A V
11955 aaatgttttgcaaaactcacacctgaacaacttacatacattcataatgacgtgattatattaggtatgtgccatattcattat
225 K C F A K L T P E Q T Y I H N D V I I L G M C H I H Y
11871 agtgatatatttccaaattttgactataacaaattaacattttcattgaatattatggaatcttacttgaataatgaaatgaca
253 S D I F P N F D Y N K T F Ξ L N I M E Ξ Y L N N E M T
11787 cgttttcagttactcaaccaatatcaagatattaaaatatcttatacacattatcatttccatgatatgaatttttatgactat
281 R F Q L N Q Y Q D I K I S Y T H Y H F H D M N F Y D Y
11703 attaaatcattctatcgtggtggtttaaatatgtataacaccaaatacataaacaaactaattgatgagccttgtttttctatt
309 I K S F Y R G G L N M Y N T K Y I N K L I D E P C F S I
11619 gacatcaattcgagtt tccttatgtgatgtatcatgaaaaaattccaacatggttatacttttacgaacactattcagaacca
337 D I N S S Y P Y V M Y H E K I P T W L Y F Y E H Y S E P
11535 acgttaatccctacttttttagatgatgacaattatttttcattatataagattgataaagatgtatttaacgatgatttatta
365 T L I P T F D D D N Y F S L Y K I D K D V F N D D L
11451 attaaaattaaatcacgtgtattacgtcaaatgattgtaaaatactataataatgataatgattacgttaatatcaatacaaat
393 I K I K S R V L R Q M I V K Y Y N N D N D Y V N I N T N
11367 acattaagaatgattcaagacattacgggtattgattgcatgcatatacgtgttaattcgtttgttatatatgaatgtgaatac
421 T R M I Q D I T G I D C M H I R V N S F V I Y E C E Y
11283 tttcatgcacgtgatattatttttcaaaactattttattaaaacacaaggtaagttaaaaaacaaaatσaatatgacatcacct
449 F H A R D I I F Q N Y F I K T Q G K L K N K I N M T S P
11199 tacgactatcacattactgatgatatcaacgaacaccca actcaaatgaggaggttatgttatctaaagtcgttttaaatgga
477 Y D Y H I T D D I N E H P Y S N E E V M L S K V V N G
11115 ttatatggcatacctgcattacgttcacattttaacttattccgtttagatgataacaatgaactatacaatatcattaacggt
505 L Y G I P A IJ R S H F N F R L D D N N E L Y N I I N G
11031 tacaaaaacactgaacgtaatatattat ctctacatttgtcacatcacgttcattgtataacttattggttcctttccaatac
533 Y K N T E R N I L F Ξ T F V T S R S L Y N L L V P F Q Y
10947 ttaacggaaagtgaaattgacgacaattttatttattgcgatactgatagtttgtatatgaaatccgttgttaaacccttattg
561 L T E S E I D D N F I Y C D T D Ξ Y M K S V V K P L L
10863 aaccccagtttattcgacccgatagccttaggtaaatgggatattgaaaacgaacagatagataagatgtttgtactgaatcat
589 N P S F D P I A G K D I E N E Q I D K M F V L N H
10779 aagaaatatgcatatgaagtgaatggaaagattaaaattgcttctgctggtataccgaaaaacgcctttgatacaagcgtcgat
617 K K Y A Y E V N G K I K I A S A G I P K N A F D T S V D
10695 tttgaaacctttgtacgtgaacaattctttgacggtgccattattgaaaacaataaaagtatctataatgagcaaggtacaata
645 F E T F V R E Q F F D G A I I E N N K S I Y N E Q G T I
10611 tcgatatatccgtctaaaactgaaattgtatgtggtaatgtatatgatgaatattttactgatgaacttaatatgaaacgtgaa
673 S I Y P S K T E I V C G N V Y D E Y F T D E N M K R E
10527 tttatattaaaagacgctagagaaaatttcgaccatagtcaatttgatgatat ctttatattgaaagtgacatcggttcattt
701 F I K D A R E N F D H S Q F D D I L Y I E S D I G S F
10443 tcacttaacgacttatttccagttgaacgttcagtacataacaaatctgatttgcatatattaaaacgtgaacatgatgaaata
729 S L N D L F P V E R S V H N K S D L H I K R E H D E I
10359 aaaaaaggcaactgttaa 10342
757 K K G N C * 44AHJDORF002
3789 atggcatataatgaaaacgattttaaatattttgatgacattcgtccatttttagacgaaatttataaaacgagagaacgttat
1 M A Y N E N D F K Y F D D I R P F D E I Y K T R E R Y
3873 acaccgttttacgatgatagagcagattataatactaattcaaaatcatattatgattatatttcaagattatcaaaactaatt
29 T P F Y D D R A D Y N T N S K S Y Y D Y I S R I- S __ E" I
3957 gaagtattagcacgtcgtatttgggactatgacaatgaattaaaaaaacgtttcaaaaattgggacgacttaatgaaagcattt
57 E V A R R I W D Y D N E L K K R F K N W D D L M K A F
4041 ccagagcaagcgaaagacttatttagaggttggttaaacgacggtacgattgacagtattattcatgacgagtttaaaaaatat
85 P E Q A K D L F R G L N D G T I D S I I H D E F K K Y
4125 agcgcaggattaacatcggcatttgctttatttaaagttactgaaatgaaacaaatgaatgactttaaatcagaagttaaagac
113 S A G L T S A F A F K V T E M K Q M N D F K S E V K D
4209 ttaattaaagatattgaccgtttcgttaatgggtttgaattaaatgagcttgaaccaaagtttgtgatgggctttggtggtatt
141 L I K D I D R F V N G F E L N E L E P K F V M G F G G I
4293 cgcaacgcagttaaccaatctattaatattgataaagaaacaaatcacatgtactctacacaatccgattctcaaaaacctgaa
169 R N A V N Q S I N I D K E T N H M Y S T Q S D S Q K P E
4377 ggtttttggataaataaattaacacctagtggtgacttaatttcaagcatgcgtattgtacagggtggtcatggtacaacaatc
197 G F W I N K L T P S G D Ii I S S M R I V Q G G H G T T I
4461 ggattagaacgtcaatccaatggtgaaatgaaaatctggttacatcacgatggtgttgcaaaactgttacaagtcgcatataaa
225 G E R Q S N G E M K I W H H D G V A K L Q V A Y K
4545 gataattatgtattagatttagaagaggctaaaggtttaacagattatacaccacagtcacttttaaacaaacacacatttaca
253 D N Y V L D L E E A K G T D Y T P Q S L N K H T F T
4629 ccgttaattgatgaagcaaatgacaaactcattttaagattcggtgacggaacaatacaggttcgttcaagagcagacgtaaaa
281 P L I D E A N D K L I L R F G D G T I Q V R S R A D V K
4713 aatcacattgataatgtagaaaaagaaatgacaattgataattcagaaaacaatgataatcgttggatgcaaggcattgctgtt
309 N H I D N V E K E M T I D N S E N N D N R W M Q G I A V
4797 gatggtgatgatttatactggttaagtggtaacagttcagttaattcacatgttcaaatcggtaaatatt attaacaacaggt
337 D G D D Y W L S G N S S V N S H V Q I G K Y S L T T G
4881 caaaagatttatgattatccatttaagttatcatatcaagacggtattaatttcccacgtgataactttaaagagcctgagggt
365 Q K I Y D Y P F K L S Y Q D G I N F P R D N F K E P E G
4965 atttgcatttatacaaatccaaaaacaaaacgtaaatcgttattacttgctatgacaaacggcggtggtggaaaacgtttccat
393 I C I Y T N P K T K R K S L A M T N G G G G K R F H
5049 aatttatatggtttcttccaacttggtgagtatgaacactttgaagcattacgcgcaagaggttcacaaaactataaattaaca 421 N Y G F F Q G E Y E H F E A L R A R G S Q N Y K T 5133 aaagacgacggtcgtgcattatctattccagaccatatcgacgatttaaatgacttaacgcaagctggtttttattatattgac 449 K D D G R A L S I P D H I D D L N D T Q A G F Y Y I D 5217 gggggtactgcagaaaaacttaagaatatgccaatgaatggtagcaagcgtataattgacgctggttgtttcattaatgtatac 477 G G T A E K L K N M P M N G S K R I I D A G C F I N V Y 5301 cctacaacacaaacattaggtacggttcaagaattaacacgtttctcaacaggtcgtaaaatggttaaaatggtgcgtggtatg
505 P T T Q T G T V Q E L T R F S T G R K M V K M V R G M 5385 ac tagacgtatttacgttaaaatgggattatggattatggacaacaatcaaaactgacgcaccatatcaagaatatttggaa 533 T D V F T K W D Y G L W T T I K T D A P Y Q E Y L E 5469 gcaagtcaatacaataactggattgcttatgtaacaacagctggtgagtattacattacaggtaaccaaatggaattatttaga 561 A S Q Y N N W I A Y V T T A G E Y Y I T G N Q M E L F R 5553 gacgcgccagaagaaattaaaaaagtgggtgcatggttacgtgtgtcaagtggtaacgcagtcggtgaagtaagacaaacatta 589 D A P E E I K K V G A W L R V S S G N A V G E V R Q T 5637 gaggctaatatatcggaatataaagaattcttcagtaatgttaatgcggaaacaaaacatcgtgaatatggttgggtagcaaaa 617 E A N I S E Y K E F F S N V N A E T K H R E Y G W V A K 5721 catcaaaaatag 5732
645 H Q K * 44AHJDORF003
6626 atgagaaagttaacgaattttaagtttttctataacacaccgtttacagactatcaaaacacgattcattttaatagtaataaa
1 M R K T N F K F F Y N T P F T D Y Q N T I H F N S N K
6710 gaacgtgatgattattttttaaatggtcgtcattttaaatcgttagactattcaaaacaaccgtataattttatacgtgataga
29 E R D D Y F L N G R H F K S L D Y S K Q P Y N F I R D R
6794 atggaaatcaatgttgatatgcagtggcatgacgcacaaggtattaactacatgacgtttttatcagattttgaggatagaaga
57 M E I N V D M Q W H D A Q G I N Y M T F L S D F E D R R
6878 tattacgcttttgtaaaccaaatcgaatacgtgaatgacgttgtggttaaaatatattttgtcattgataccattatgacgtat
85 Y Y A F V N Q I E Y V N D V V V K I Y F V I D T I M T Y
6962 acacaagggaatgtattagagcaactctcaaacgtcaatattgaacgtcaacatttatcaaaacgcacgtataactatatgtta
113 T Q G N V E Q L S N V N I E R Q H L S K R T Y N Y M L
7046 ccaatgttacgtaataatgatgatgtgttaaaagtatcaaataaaaactatgtttataaccaaatgcaacaatatttggaaaat
141 P M R N N D D V K V S N K N Y V Y N Q M Q Q Y E N
7130 ttagtattattccagtcaagcgctgatttatcaaagaaatttggtactaaaaaagagccaaacttagatacgtcaaaaggtacg
169 L V F Q S S A D L S K K F G T K K E P N L D T S K G T
7214 atttatgacaatatcacatcaccagtcaacttatacgttatggaatatggtgactttattaactttatggataaaatgagtgcc
197 I Y D N I T S P V N L Y V M E Y G D F I N F M D K M S A
7298 tatccatggattacgcaaaactttcaaaaggttcaaatgttacctaaagactttattaatacaaaagacttagaggacgttaaa
225 Y P W I T Q N F Q K V Q M L P K D F I N T K D L E D V K
7382 accagtgaaaaaattacaggattaaaaacattaaaacagggtggtaaatcaaaagaatggagtctaaaagatttatcattaagt
253 T S E K I T G K T L K Q G G K S K E S K D S S
7466 ttctcaaatcttcaagagatgatgttatctaaaaaagatgaatttaaacatatgatacgtaatgagtatatgacaattgaattt
281 F S N L Q E M M L S K K D E F K H M I R N E Y M T I E F
7550 tatgactggaatggaaatacgatgttactcgacgctggtaagatttcacaaaaaactggtgttaagttacgtacaaaatcaatt
309 Y D W N G N T M L D A G K I S Q K T G V K R T K S I
7634 attggttatcataatgaagttcgagtatatccagtagattataacagtgctgaaaacgacagaccaatactcgctaaaaataaa
337 I G Y H N E V R V Y P V D Y N S A E N D R P I L A K N K
7718 gaaatattgattgatacgggttcattcttaaatacaaatataacatttaatagttttgcacaagtaccaatattaatcaataat
365 E I L I D T G S F N T N I T F N S F A Q V P I L I N N
7802 ggtatcttaggacaatcacaacaagccaaccgacaaaaaaatgcagaaagtcaattaattacaaatcgtattgataatgtatta
393 G I L G Q S Q Q A N R Q K N A E S Q I T N R I D -J V L- -
7886 aatggtagcgacccgaaatcacgcttttatgacgctgtgagtgtagcaagtaatttaagtccaactgctttatttggtaagttt
421 N G S D P K S R F Y D A V S V A S N S P T A F- G K F
7970 aatgaagaatataatttctacaaacaacaacaagctgaatataaagatttagccttacaaccaccttctgtaactgaatcagaa
449 N E E Y N F Y K Q Q Q A E Y K D A Q P P S V T E S E
8054 atgggcaacgcattccaaattgcgaatagcattaacggtttaacgatgaaaattagtgtaccgtcacctaaagaaattacattt
477 M G N A F Q I A N S I N G L T M K I S V P S P K E I T F
8138 ttacaaaaatattatatgttgtttggttttgaagtgaatgactataattcatttattgaaccaattaacagtatgactgtttgc
505 L Q K Y Y M L F G F E V N D Y N S F I E P I N Ξ M T V C
8222 aattatttaaaatgtacaggtacgtatactatacgtgacatcgaccccatgttaatggaacaattaaaagcaattttagaatct
533 N Y Ii K C T G T Y T - R D I D P L M E Q K A I L E S
8306 ggtgtaagattttggcataatgacggttcaggtaatccaatgttacaaaatccattaaataacaaatttagagagggggtataa 8389
561 G V R F W H N D G S G N P M Q N P N N K F R E G V * 44AHJDORF004
8764 atgatactgaaaagagtgataacaatgaacgatcaagagaagatagataaatttacgcattcctatattaatgatgattttggt
1 M I L K R V I T M N D Q E K I D K F T H S Y I N D D F G
8848 ttaacgatagaccagttagtccctaaagtaaaaggatatgggcgctttaatgtatggcttggtggtaatgaaagtaaaatcaga
29 L T I D Q V P K V K G Y G R F N V W G G N E S K I R
8932 caagtattaaaagcagtaaaagagataggtgtttcacctactctttttgccgtatatgaaaaaaatgagggttttagttctgga
57 Q V K A V K E I G V S P T L F A V Y E K N E G F S S G
9016 cttggttggttaaaccatacgtctgcacgtggtgattatttaacagatgctaaattcatagcaagaaagttagtatcacaatca
85 G W N H T S A R G D Y T D A K F I A R L V S Q S
9100 aaacaagctggacaaccgtcttggtatgacgcaggtaacatcgtccactttgtaccacaagacgtacaaagaaaaggtaatgca
113 K Q A G Q P S Y D A G N I V H F V P Q D V Q R K G N A
9184 gattttgcaaaaaatatgaaagcaggtacaattggacgtgcatatattccattaacagcagctgctacttgggcggcatattat
141 D F A K N M K A G T I G R A Y I P T A A A T W A A Y Y
9268 cctttaggtttgaaagcatcatataacaaagtacaaaactatggtaatccatttttagacggtgcgaatactattctagcttgg
169 P L G L K A S Y N K V Q N Y G N P F L D G A N T I L A W
9352 ggtggtaaattagacggtaaaggtggatcacctagtgattcgtctgacagtggtagtagtggtgacagtggtagttcactactc
197 G G K D G K G G S P S D S S D S G S S G D S G S S
9436 gctttagcaaaacaagccatgcaagaattattaaaaaaaatacaagacgcattacaatgggacgttcatagtattggtagtgat
225 A A K Q A M Q E L L K K I Q D A Q W D V H S I G S D
9520 aaattttttagtaatgattattttacattagaaaaaacatttaacaacacatatcatattaaaatgacgattggtttacttgat
253 K F F S N D Y F T L E K T F N N T Y H I K M T I G L Ii D
9604 tcattaaaaaaactgattgatagcgttcaagtagatagtgggagtagtagttctaatcctactgatgatgacggagaccataaa
281 S L K K I D S V Q V D S G S S S S N P T D D D G D H K
9688 ccaattagtggtaaatcagtcaagccaaatggaaaaagtggtcgtgtgattggtggtaactggacatatgcacagttaccagaa
309 P I S G K S V K P N G K S G R V I G G N T Y A Q L P E
9772 aaatataaaaaagcaattggtgtacctttattcaaaaaagaatacttatacaaaccaggtaacatatttcctcaaacgggtaat
337 K Y K K A I G V P F K K E Y L Y K P G N I F P Q T G N
9856 gcaggacaatgtacagaattaacatgggcgtatatgtcacaactacatggtaaaagacaacctaccgacgacggtcaaataaca
365 A G Q C T E L T W A Y M S Q L H G K R Q P T D D G Q I T
9940 aacggtcagcgtgtatggtacgtctataaaaagttaggtgcaaaaacaacacataatccaacagtaggttatggtttctctagt
393 N G Q R V W Y V Y K K G A K T T H N P T V G Y G F S S
10024 aaaccaccatacttacaagcaactgcatatggtattggtcacacaggtgttgttgtagcagtttttgaagatggttcgttttta
421 K P P Y Q A T A Y G I G H T G V V V A V F E D G S F L
10108 gttgcaaactataatgtaccaccatatgttgcaccatcacgtgtggtattgtatacactcattaatggcgtaccaaataatgct
449 V A N Y N V P P Y V A P S R V V L Y T I N G V P N N A
10192 ggtgataatattgtattctttagtggtattgcttaa 10227
477 G D N I V F F S G I A * 44AHJDORF005
13890 atggtaaaacaaaatcgtttagacatggtaagagattatcaaaatgctgtcaatcatgtcagaaaaaaaatcccagataagtat
1 M V K Q N R L D M V R D Y Q N A V N H V R K K I P D K Y
13806 aatcaaatagaattagttgatgaacttatgaatgatgatatagattattatatatctatttcaaaccgttctgatggaaaatcg
29 N Q I E L V D E M N D D I D Y Y I S I S N R S D G K S
13722 ttcaactatgtttcattttttatttatttagctattaaacttgatataaaatttactttattatcacgtcattatacattacgt
57 F N Y V S F F I Y L A I K L D I K F T L L S R H Y T R
13638 gacgcttaccgtgattttattgaagaaatcatagatgaaaatccactatttaaatcaaaacgtgtcacgttcagaagtgctagg
85 D A Y R D F I E E I I D E N P F K S K R V T F R S A R
13554 gactatttagctattatctatcaagataaagaaattggtgtgattacagatttgaatagtgccactgatttaaaatatcattct
113 D Y A I I Y Q D K E I G V I T D L N S A T D K Y H S
13470 aactttttaaaacactatcctattattatatatgatgagtttttagcacttgaagatgattatttaattgatgagtgggataag
141 N F L K H Y P I I I Y D E F A L E D D Y I D E D K
13386 ttaaaaacaatatatgaatcaatcgaccgtaaccatggtaacgttgattatattggattccctaaaatgtttttactaggtaat
169 K T I Y E S I D R N H G N V D Y I G F P K M F Ii li G N
13302 gcagtcaacttttcaagtcctatattatccaatttaaatatatacaatttattacaaaagcataaaatgaatacatcaagactt
197 A V N F S S P I L S N L N I Y N L L Q K H K M N T S R L
13218 tacaaaaacatttttttagaaatgcgacgaaacgattacgtgaatgaaaaacgtaacacacgtgcgtttaattcaaatgacgac
225 Y K N I F E M R R N D Y V N E K R N T R A F N S N D D
13134 gctatgacaactggagaatttgaatttaacgaatataatttggcggatgataatttaagaaatcacatcaatcaaaacggtgat
253 A M T T G E F E F N E Y N A D D N L R N H I N Q N G D
13050 ttcttctatatcaaaactgatgataaatatattaaagtcatgtataatgtaactacttttatgacaaatattatcgttgtacca
281 F F Y I K T D D K Y I K V M Y N V T T F M T N I I V V P
12966 tatacaaaacaatatgaattttgtactaaaattagggatatagacaatcatgttacctatttacgtgatgatatgttttataaa
309 Y T K Q Y E F C T K I R D I D N H V T Y L R D D M F Y K
12882 gaaaacatggaacgttattactacaatccaagcaatttacattttgacaatgcttactctaaaaattacgtggttgataatgat
337 E N M E R Y Y Y N P S N H F D N A Y S K N Y V V D N D
12798 agatatttatatttagatatgaataaaattataaaatttcatataaaaaatgaaatgaagaaaaatatgagtgagtttgaaaga
365 R Y L Y L D M N K I I K F H I K N E M K K N M S E F E R
12714 aaagaaaaaatatacgaagataactatatagagaatacgaaaaagtatctaatgaaacaatatggcttataa 12643
393 K E K I Y E D N Y I E N T K K M K Q Y G * 44AHJDORF006
803 atggcacaacaatctacaaaaaatgaaactgcacttttagtagcaaagtcagctaaatcagcgttacaagattttaatcatgat
1 M A Q Q S T K N E T A L V A K S A K S A Q D F N H D
887 tattcaaaatcttggacatttggcgacaaatgggataattcaaatacaatgttcgaaacatttgtaaataaatatttattccct
29 Y S K S T F G D K D N S N T M F E T F V N K Y L F P
971 aagattaatgagactttattaatcgatattgcattaggtaatcgttttaattggttagctaaagagcaagattttattggacaa
57 K I N E T L L I D I A L G N R F N W L A K E Q D F I G Q
1055 tatagtgaagaatacgtgattatggacacagtaccaattaacatggacttatctaaaaatgaggaattaatgttgaaacgtaat
85 Y S E E Y V I M D T V P I N M D L S K N E E L M K R N
1139 tatccacgtatggcaactaagttatatggtaacggaattgtgaagaaacaaaaattcacattaaacaacaatgatacacgtttc
113 Y P R M A T K L Y G N G I V K K Q K F T N N N D T R F
1223 aatttccaaacattagcagacgcaactaattacgctttaggtgtatacaaaaagaaaatttctgatattaatgtattagaagaa
141 N F Q T L A D A T N Y A L G V Y K K K I S D I N V L E E
1307 aaagaaatgcgtgcaatgttagttgattactcattgaatcaattatccgaaacaaatgtacgtaaagcaacatcaaaagaagat
169 K E M R A M L V D Y S N Q L S E T N V R K A T S K E D
1391 ttagcaagcaaagtttttgaagcaatcctaaacttacaaaacaacagtgctaaatataatgaagtacatcgtgcatcaggtggt
197 L A S K V F E A I N Q N N S A K Y N E V H R A S G G
1475 gcaattggacaatatacaactgtatcaaaattaaaagatattgtgattttaacaacagattcattaaaatcttatcttttagat
225 A I G Q Y T T V S K K D I V I L T T D S L K S Y L D
1559 actaagattgcaaacacattccagattgcaggcattgatttcacagatcacgttattagttttgacgacttaggtggcgtgttt
253 T K I A N T F Q I A G I D F T D H V I S F D D G G V F 1643 aaagtaacaaaagaatttaagttacaaaaccaagattcaattgactttttacgtgcgtatggagattatcaatcacaattagga 281 K V T K E F K L Q N Q D S I D F L R A Y G D Y Q S Q L G 1727 gatacaattccagttggtgctgtatttacttatgatgtatctaaacttaaagagtttactggcaacgttgaagaaattaaacca 309 D T I P V G A V F T Y D V S K L K E F T G N V E E I K P 1811 aaatcagatttatatgcgtttattttggatattaattcaattaaatataaacgttacacaaaaggtatgttaaaaccaccattc 337 K S D L Y A F I L D I N S I K Y K R Y T K G M L K P P F 1895 cataaccctgaatttgatgaagttacacactggattcattactattcatttaaagccattagtccattctttaataaaatttta 365 H N P E F D E V T H I H Y Y S F K A I S P F F N K I 1979 attactgaccaagatgtaaatccaaaaccagaggaagaattacaagaataa 2029
393 I T D Q D V N P K P E E E L Q E * 4AHJDORF007
2044 atgaacaacgataaaagaggtttaaacgttgagttatcaaaggaaatcagcaaaagagttgttgaacatcgcaacagatttaaa
1 M N N D K R G L N V E S K E I S K R V V E H R N R F K
2128 cgtcttatgtttaatcgttatttggaatttttaccgctactaatcaactataccaatcgtgatacggttggtatagattttatt
29 R L M F N R Y E F L P L I N Y T N R D T V G I D F I
2212 cagttagaatcagctttaagacaaaacattaatgtagttgttggtgaagctagaaataagcaaattatgattcttggttatgta
57 Q L E S A L R Q N I N V V V G E A R N K Q I M I G Y V
2296 aataacacttactttaatcaagcaccaaatttttcatcaaactttaatttccaatttcaaaaacgattaactaaagaagatata
85 N N T Y F N Q A P N F S S N F N F Q F Q K R L T K E D I
2380 tattttattgtacctgactatttaatacctgatgattgtctacaaattcataagctatatgataactgtatgagtggtaacttt
113 Y F I V P D Y I P D D C Q I H K Y D N C M S G N F
2464 gttgtcatgcaaaataaaccaattcaatataatagtgatatagaaattatagaacattatactgatgaattagcagaagttgct
141 V V M Q N K P I Q Y N S D I E I I E H Y T D E L A E V A
2548 ttatctcgcttttctttaatcatgcaagcaaaatttagcaagatatttaaatcagaaattaatgacgagtcaatcaatcaactt 169 L S R F S L I M Q A K F S K I F K S E I N D E S I N Q L 2632 gtgtccgaaatatataacggtgcaccatttgttaaaatgtcacctatgtttaatgcagatgacgatatcattgatttaacaagt 197 V S E I Y N G A P F V K M Ξ P M F N A D D D I I D L T S 2716 aatagcgtaatcccagcattaactgaaatgaaacgggaatatcaaaacaaaattagtgaattaagtaactatttaggcattaat 225 N S V I P A T E M K R E Y Q N K I S E L S N Y L G I N 2800 tca agccgttgataaagaaagcggtgtttcagacgaagaggcaaaaagtaatcgtggatttaccacatcaaacagtaatatc 253 S A V D K E S G V S D E E A K S N R G F T T S N S N I 2884 tatttaaaaggtcgtgaaccaattacgtttttatcaaagcgttatggtttagatattaaaccgtattacgatgatgaaacaacg 281 Y L K G R E P I T F L S K R Y G L D I K P Y Y D D E T T 2968 tctaaaatatcaatggtagacacactttttaaagatgaaagcagtgatataaatggctag 3027
309 S K I S M V D T F K D E S S D I N G * 44AHJDORF008
3020 atggctagatacacaatgactttatacgatttcattaaatcagaattgattaaaaaaggtttcaatgaatttgtaaatgataat
1 M A R Y T M T Y D F I K S E L I K K G F N E F V N D N
3104 aaattaacgttttatgatgatgaatttcaattcatgcaaaaaatgctgaagttcgacaaagacgttttagctatcgttaatgaa
29 K T F Y D D E F Q F M Q K M Ii K F D K D V L A I V N E
3188 aaagtatttaaaggtttttcattgaaagatgaattatcagatttactttttaaaaaatcatttacgattcattttttagataga
57 K V F K G F S K D E L S D L L F K K S F T I H F L D R
3272 gaaatcaacagacaaacagttgaagcatttggcatgcaagtgattactgtatgtattacacatgaggattatttaaatgtggtt
85 E I N R Q T V E A F G M Q V I T V C I T H E D Y L N V V
3356 tat catcaagtgaagttgaaaaatacttacaatcacaaggcttcacagaacacaatgaagatacaacaagtaacactgatgaa
113 Y S S S E V E K Y L Q S Q G F T E H N E D T T S N T D E
3440 acatcgaatcaaaatgctacatctttagacaattcaactggcatgactgcaaacagaaacgcttatgtgtcattaccacaaagt
141 T S N Q N A T S L D N S T G M T A N R N A Y V S L P Q S
3524 gaggttaacattgatgttgataatacaacgttacgattcgctgataataatacgattgataacggtaaaactgtgaataaatcg-
169 E V N I D V D N T T R F A D N N T I D N G K T V _I—K*"S
3608 agtaacgaaagtaatcaaaacgcaaaacgtaatcaaaatcaaaaaggtaatgcaaaaggtacacaattcactaagcagtattta
197 S N E S N Q N A K R N Q N Q K G N A K G T Q F T K Q Y L
3692 attgataatattgataaagcgtacgatttaagaaagaaaattttaaatgaatttgataaaaaatgttttttacaaatttggtag 3775
225 I D N I D K A Y D R K K I L N E F D K K C F L Q I W * 44AHJDORF009
5744 atgaaatcacaacaacaagcaaaagaatggatatataagcatgagggggcaggtgttgactttgatggtgcatatggatttcaa
1 M K S Q Q Q A K E W I Y K H E G A G V D F D G A Y G F Q
5828 tgtatggacttatcagttgcttatgtgtattacattactgacggtaaagttcgcatgtggggtaatgctaaagacgcgataaat
29 C M D L S V A Y V Y Y I T D G K V R M W G N A K D A I N
5912 aatgactttaaaggtttagcgacggtgtataaaaatacaccgagctttaaacctcaattaggggacgttgctgtatatacaaat
57 N D F K G L A T V Y K N T P S F K P Q L G D V A V Y T N
5996 ggacaatatggacatattcaatgtgtgttaagtggaaatcttgattattatacatgcttagaacaaaactggttaggcggcggt
85 G Q Y G H I Q C V S G N L D Y Y T C E Q N W G G G
6080 tttgacggttgggaaaaagcaaccattagaacacattattatgacggtgtaactcactttattagacctaaattttcaggtagt
113 F D G W E K A T I R T H Y Y D G V T H F I R P K F S G S
6164 aatagcaaagcattagaaacatcaaaagtaaatacatttggaaaatggaaacgaaaccaatacggcacatattatagaaatgaa
141 N S K A L E T S K V N T F G K W K R N Q Y G T Y Y R N E
6248 aatggtacatttacatgtggttttttaccaatatttgcacgtgtcggtagtccaaaattatcagaacctaatggctattggttc
169 N G T F T C G F L P I F A R V G S P K Ξ E P N G Y W F
6332 caaccaaacggttatacaccatataacgaagtttgtttatcagatggttacgtatggattggttataactggcaaggcacacgt
197 Q P N G Y T P Y N E V C L S D G Y V W I G Y N W Q G T R
6416 tattatttaccagtgcgccaatggaatggaaaaacaggtaatagttacagtgttggtattccttggggggtgttctcataa 6496
225 Y Y L P V R Q W N G K T G N S Y S V G I P W G V F S * 44AHJDORF010
14420 ttggttagacatacgtctgaaatggatagatggaaaaaagaaagagaagctagaaaagagcaagaaaaagatttatttttaaat
1 V R H T S E M D R W K K E R E A R K E Q E K D L F N
14336 gattttagtaatgttaattttaaatttgatgataaagatttacaagaggcgtacattgacacatggaaacattttgcacatctg
29 D F S N V N F K F D D K D Q E A Y I D T K H F A H
14252 ccctattttcctaaagaaagaaacgtatcatatgtaaatgctgtatcattggtaagaggttcaagacataaaaaattaaattat
57 P Y F P K E R N V S Y V N A V S L. V R G S R H K K N Y
14168 attcttgaaatatataaccgtaatgatgattctaataataaaaacgctaaaaagcataaatacgctttatataatttacaagct
85 I L E I Y N R N D D S N N K N A K K H K Y A Y N Q A
14084 aaaaataataattcttcaatgtataaatatattaaagaaatcgatactttatataaagaaattggtaaatcagatagaccagtg
113 K N N N S S M Y K Y I K E I D T L Y K E I G K S D R P V
14000 acaaatattgatgatgaagatgtgaggtataactttttatattatgcaacatttgacgaataa 13938
141 T N I D D E D V R Y N F L Y Y A T F D E * 44AHJDORF011
15593 atgacaaacgtaaaagatattttatcaagacaccaaaacacattagcgagatttgaatttgaggaaaaagaaagagaatttatc
1 M T N V K D I L S R H Q N T L A R F E F E E K E R E F I
15509 aaactatcagaattagtagaaaaatacggtatgaaaaaagagtatatcgttagagcattattcacaaacaaagaatcaaaattc
29 K L S E L V E K Y G M K K E Y I V R A F T N K E S K F
15425 ggtgaacaaggtgttatcgtcactgatgactataacgtaaacttaccgaaccacttaacagaattaattaaagaaatgagagca
57 G E Q G V I V T D D Y N V N L P N H T E I K E M R A
15341 gatgaggacgttgttgacattatcaatgctggagaagttcaattcacaatttatgaatatgaaaacaaaaaaggtcaaaaaggt
85 D E D V V D I I N A G E V Q F T I Y E Y E N K K G Q K G
15257 tactcaatcaattttggtcaagtatcattttaa 15225
113 Y S I N F G Q V S F * 44AHJDORF012
8391 atgaacgaagtaaaattcagatttacagactcagaagcgtttcacatgtttatatacgctggggatttaaaattactctacttt
1 M N E V K F R F T D S E A F H M F I Y A G D L K L Y F
8475 ttatttgtattaatgttcgttgatattattacaggtatttcaaaagcaattaaaaataataacttatggtcaaaaaaatcaatg
29 L F V M F V D I I T G I S K A I K N N N L S K K S M
8559 agaggattttctaaaaaattattgatattctgtattatcattttagcaaacatcattgaccagattttacaattaaaaggtggt
57 R G F S K K L I F C I I I L A N I I D Q I L Q L K G G
8643 ctactcatgattacaatattttattatattgcaaatgagggactttctattgtagaaaattgtgcagaaatggacgtattagta
85 L L M I T I F Y Y I A N E G S I V E N C A E M D V L V
8727 ccagaacaaattaaagataaattaagagtcattaaaaatgatactgaaaagagtgataacaatgaacgatcaagagaagataga
113 P E Q I K D K L R V I K N D T E K S D N N E R S R E D R
8811 taa 8813
141 * 44AHJDORF013
14996 atgaaaattaaaactacttttagattaaataatttaatttattaccttttaacaaatagagattattataatgataaatttgaa
1 M K I K T T F R L N N L I Y Y Ii L T N R D Y Y N D K F E
14912 aaatttacttcatctaataaaaaatgtatagtaaaaataaatatgggtgatgtgtatattgagtttgacaaacaatatgatgat
29 K F T S S N K K C I V K I N M G D V Y I E F D K Q Y D D
14828 tttgaaattgaaaaagagttatttacgttagatatcgacattgatattaaaaaacatgtttttaatatacttgtattttattat
57 F E I E K E L F T L D I D I D I K K H V F N I L V F Y Y
14744 agaaattatttaagtaatgaattaataagagaaattttattaaacgttacaattgacgacgtattatcaaattttgataaacct
85 R N Y S N E I R E I L N V T I D D V L S N F D K P
14660 cttgaaagcgaattaatgattatttatcaaaacaaagtcatatacgataatgggaaagtgattgaccatgaataa 14586
113 E S E L M I I Y Q N K V I Y D N G K V I D H E * 44AHJDORF113
199 atgacagaatttgatgaaatcgtaaaaccagacgacaaagaagaaacttcagaatcaactgaagaaaatttagaatcaactgaa
1 M T E F D E I V K P D D K E E T S E S T E E 'N L E S T E- .
283 gaaacttcagaatcaactgaagaatcaactgaagaatcaactgaagaatcaactgaagataaaacagtagaaacaatcgSagaa
29 E T S E S T E E S T E E S T E E S T E D K T V E T- I E E
367 gaaaatgaaaacaaattagaacctactacaacagatgaagatagttcgaaatttgaccctgttgtattagaacaacgtattgct
57 E N E N K L E P T T T D E D S S K F D P V V L E Q R I A
451 tcat agaacaacaagtgactacttttttatcttcacaaatgcaacaaccacaacaagtacaacaaacacaatcagatgtaaca
85 S L E Q Q V T T F S S Q M Q Q P Q Q V Q Q T Q S D V T
535 gaatcaaacaaagaagataacgactattcagatgaagaactagttgataagttagatttagattag 600
113 E S N K E D N D Y S D E E V D K L D L D *
44AHJDORF114
16172 atggttaatgttgataatgcaccagaagaaaaaggacaagcctatactgaaatgttgcaactattcaataaactgattcaatgg
1 M V N V D N A P E E K G Q A Y T E M Q L F N K L I Q W
16088 aatccagcttatacatttgacaatgcaattaacttattatcggcttgccaacaactattattaaactataatagttctgttgtt
29 N P A Y T F D N A I N L S A C Q Q L N Y N S S V V
16004 caattcttaaatgatgaactaaacaacgaaactaaaccagaatcaatattgtcttatattgctggtgatgacccaatagaacaa
57 Q F L N D E L N N E T K P E S I L S Y I A G D D P I E Q
15920 tggaatatgcataaaggattttatgaaacgtataacgtttacgttttttag 15870
85 W N H K G F Y E T Y N V Y V F *
44AHJDORF014
6243 atgaaaatggtacatttacatgtggttttttaccaatatttgcacgtgtcggtagtccaaaatta agaacctaatggctatt
1 M K M V H I. H V V F Y Q Y -. H V S V V Q N Y Q N M A I
6327 ggttccaaccaaacggttatacaccatataacgaagtttgtttatcagatggttacgtatggattggttataactggcaaggca
29 G S N Q T V I H H I T K F V Y Q M V T Y G V I T G K A
6411 cacgttattatttaccagtgcgccaatggaatggaaaaacaggtaatagttacagtgttggtattccttggggggtgttctcat
57 H V I I Y Q C A N G M E K Q V I V T V L V F L G G C S H
6495 aatgggtattttagcctttttctttga 6521
85 N G Y F S F L *
44AHJDORF015
15403 gtgacgataacaccttgttcaccgaattttgattctttgtttgtgaataatgctctaacgatatactcttttttcataccgtat
1 V T I T P C S P N F D S F V N N A T I Y Ξ F F I P Y
15487 ttttctactaattctgatagtttgataaattctctttctttttcctcaaattcaaatctcgctaatgtgttttggtgtcttgat
29 F S T N S D S I N S L S F S S N S N L A N V F W C L D
15571 aaaatatcttttacgtttgtcattttatttctcctcttatttaaattatttgctttctgcaattgcgatttgtag 15645
57 K I S F T F V I F L L L F K L F A F C N C D L *
44AHJDORF016
15852 atgaaagttgacgacattgttaccttacgtgtcaaaggttatatacttcattacttagatgatgataatgaatacattgaggaa
1 M K V D D I V T R V K G Y I H Y L D D D N E Y I E E
15768 tttttaccacttcacgagtatcatttaaccaaaacacaagcaaaagaattattaccagacacatgtaaactattgtccactaca
29 F L P H E Y H T K T Q A K E L L P D T C K L S T T
15684 cgcacaacgaaaacaattcaagtttattacaatgatttactacaaatcgcaattgcagaaagcaaataa 15616
57 R T T K T I Q V Y Y N D L Q I A I A E S K *
44AHJDORF017
10757 atggaaagattaaaattgcttctgctggtataccgaaaaacgcctttgatacaagcgtcgattttgaaacctttgtacgtgaac
1 M E R L K L L L V Y R K T P I Q A S I L K P Y V N
10673 aattctttgacggtgccattattgaaaacaataaaagtatctataatgagcaaggtacaatatcgatatatccgtctaaaactg
29 N S L T V P L K T I K V S I M S K V Q Y R Y I R L K L
10589 aaattgtatgtggtaatgtatatgatgaatattttactgatgaacttaatatga 10536
57 K L Y V V M Y M N I L M N L I *
44AHJDORF018
1098 atgttaattggtactgtgtccataatcacgtattcttcactatattgtccaataaaatcttgctctttagctaaccaattaaaa
1 M L I G T V S I I T Y S S Y C P I K S C S L A N Q L K
1014 cgattacctaatgcaatatcgattaataaagtctcattaatcttagggaataaatatttatttacaaatgtttcgaacattgta
29 R P N A I S I N K V S L I G N K Y L F T N V S N I V
930 tttgaattatcccatttgtcgccaaatgtccaagattttgaataa 886
57 F E L S H L S P N V Q D F E *
44AHJDORF019
9836 atgttacctggtttgtataagtattcttttttgaataaaggtacaccaattgcttttttatatttttctggtaactgtgcatat
1 M P G L Y K Y Ξ F L N K G T P I A F L Y F S G N C A Y
9752 gtccagttaccaccaatcacacgaccactttttccatttggcttgactgatttaccactaattggtttatggtctccgtcatca
29 V Q P P I T R P F P F G L T D P L I G W S P S S
9668 tcagtaggattagaactactactcccactatctacttga 9630
57 S V G L E L L L P L S T *
44AHJDORF121
16362 atggaaaatgaaacaaaaaacattgagttgaagcatgtttttcgttttaagaatggaagtttatgtatagcgttatttgataga
1 M E N E T K N I E L K H V F R F K N G S C I A L F D R
16278 acagaaaatgaaatttcattttatgatgttgacattgatgaaattgaagatttaaatcataattctgttttacgcgtaatttca
29 T E N E I S F Y D V D I D E I E D L N H N S V L R V I S
16194 actttattaggaagtgataataatggttaa 16165
57 T L L G S D N N G *
44AH DORF020
13865 atgtctaaacgattttgttttaccatgtttttgctccttgtaatagtttatgatgtcgtttacagtgttaaatttattcgtcaa
1 M S K R F C F T M F L J- L V I V Y D V V Y S V K F I R Q
13949 atgttgcataatataaaaagttatacctcacatcttca catcaatatttgtcactggtctatctgatttaccaatttctttat
29 M L H N I K Ξ Y T S H L H H Q Y L S V Y L I Y Q F Y
14033 ataaagtatcgatttctttaa 14053
57 I K Y R F *
44AHJDORF123 ~ __ *"
614 atgtatgagggaaacaacatgcgttctatgatgggtacatcatatgaagattcaagattaaataaacgaacagaβttaaatgaa
1 M Y E G N N M R S M M G T S Y E D S R N K R T E N E
698 aacatgtcaattgatacaaataaaagtgaagatagttatggtgtacaaattcattcactttcaaaacaatcatttacaggtgac
29 N M S I D T N K S E D S Y G V Q I H S S K Q S F T G D
782 gttgaggaggaataa 796
57 V E E E *
44AHJDORF021
5816 atgcaccatcaaagtcaacacctgccccctcatgcttatatatccattcttttgcttgttgttgtgatttcatttatatcactc
1 M H H Q S Q H L P P H A Y I S I L L V V V I S F I S
5732 ctatttttgatgttttgctacccaaccatattcacgatgttttgtttccgcattaacattactgaagaattctttatattccga
29 L F L M F C Y P T I F T M F C F R I N I T E E F F I F R
5648 tatattagcctctaa 5634
57 Y I S L *
44AHJDORF022
8611 atgtttgctaaaatgataatacagaatatcaataattttttagaaaatcctctcattgatttttttgaccataagttattattt
1 M F A K M I I Q N I N N F L E N P I D F F D H K F
8527 ttaattgcttttgaaatacctgtaataatatcaacgaacattaatacaaataaaaagtag 8468
29 L I A F E I P V I I S T N I N T N K K *
44AHJDORF023
6494 atgagaacaccccccaaggaataccaacactgtaactattacctgtttttccattccattggcgcactggtaaataataacgtg
1 M R T P P K E Y Q H C N Y Y L F F H S I G A L V N N N V
6410 tgccttgccagttataaccaatccatacgtaaccatctgataaacaaacttcgttatatggtgtataaccgtttggttggaacc
29 C L A S Y N Q S I R N H L I N K R Y M V Y N R V G T
6326 aatagccattag 6315
57 N S H *
44AHJDORF024
14275 gtgtcaatgtacgcctcttgtaaatctttatcatcaaatttaaaattaacattactaaaatcatttaaaaataaatctttttct
1 V S M Y A S C K S S S N K T L L K S F K N K S F S
14359 tgctcttttctagcttctctttcttttttccatctatccatttcagacgtatgtctaaccaatgttatcaacctccatataaag
29 C S F A S S F F H S I S D V C T N V I N L H I K
14443 cataaataa 14451
57 H K *
44AHJDORF025
15175 atggaacgtaaatacaaaacggtattattatattgcgatgagattaaaggacattttccacatcaaatctcaatgtttgaagat
1 M E R K Y K T V L L Y C D E I K G H F P H Q I S M F E D
15091 ttatatgacgctaaagttgtatattcatattatgaatataacctgttcactaaaaaatacgcgtatatcatagaatacattaag
29 Y D A K V V Y S Y Y E Y N L F T K K Y A Y I I E Y I K
15007 gagatataa 14999
57 E l *
44AHJDORF026
14593 atgaataacctattaaacatagccattgttttccttttagcatttttaattacacttatcatacttatgacactgcatatacgc
1 M N N L N I A I V F L A F L I T L I I M T L H I R
14509 gtgtcatttggtgttttattcactacattgattatattctatattatctttttaatggttatttatgctttatatggaggttga
14426
29 V S F G V F T T L I I F Y I I F M V I Y A L Y G G *
44AHJDORF027
12916 atgattgtctatatccctaattttagtacaaaattcatattgttttgtatatggtacaacgataatatttgtcataaaagtagt
1 M I V Y I P N F S T K F I F C I Y N D N I C H K S S
13000 tacattatacatgactttaatatatttatcatcagttttgatatagaagaaatcaccgttttgattgatgtgatttcttaa
13080
29 Y I I H D F N I F I I S F D I E E I T V I D V I S *
44AHJDORF029
15183 gtgtttaaatggaacgtaaatacaaaacggtattattatattgcgatgagattaaaggacattttccacatcaaatctcaatgt
1 V F K W N V N T K R Y Y Y I A M R L K D I F H I K S Q C
15099 ttgaagatttatatgacgctaaagttgtatattcatattatgaatataacctgttcactaaaaaatacgcgtatatcatag
15019
29 L K I Y M T K L Y I H I M N I T C S L K N T R I S *
44AHJDORF028
9235 atggaatatatgcacgtccaattgtacctgctttcatattttttgcaaaatctgcattaccttttctttgtacgtcttgtggta
1 M E Y M H V Q L Y L S Y F Q N L H Y L F F V R L V V
9151 caaagtggacgatgttacctgcgtcataccaagacggttgtccagcttgttttgattgtgatactaactttcttgctatga 9071
29 Q S G R C Y R H T K T V V Q L V L I V I L T F L *
44AHJDORF030
14487 gtgaataaaacaccaaatgacacgcgtatatgcagtgtcataagtatgataagtgtaattaaaaatgctaaaaggaaaacaatg
1 V N K T P N D T R I C S V I S M I S V I K N A K R K T M
14571 gctatgtttaataggttattcatggtcaatcactttcccat at gtatatgactttgttttgataaataatcattaa 14648
29 A M F N R L F M V N H F P I I V Y D F V I N N H *
44AHJDORF031
11039 atgatattgtatagttcattgttatcatctaaacggaataagttaaaatgtgaacgtaatgcaggtatgccatataatccattt
1 M I L Y S S L S S K R N K L K C E R N A G M P Y N P F
11123 aaaacgactttagataacataacctcct atttgagtatgggtgttcgttgatatcatcagtaatgtga 11191
29 K T T L D N I T S S F E Y G C S L I S S V M *
44AHJDORF135
693 atgaaaacatgtcaattgatacaaataaaagtgaagatagttatggtgtacaaat cattcactttcaaaacaatc__tteacag
1 M K T C Q L I Q I K V K I V M V Y K F I H F Q N^H L Q
777 gtgacgttgaggaggaataataaattatggcacaacaatctacaaaaaatgaaactgcacttttag 842
29 V T R R N N K L W H N N Q K M K L H F *
44AHJDORF033
3795 atgccatta ttaaccacctctaccaaatttgtaaaaaacattttt atcaaattcatttaaaattttctttcttaaatcgtac
1 M P L F N H L Y Q I C K K H F L S N S F K I F F L K S Y
3711 gctttatcaatattat aattaaatactgcttagtgaattgtgtaccttttgcattacctttttga 3646
29 A L S I S I K Y C L V N C V P F A P F *
44AHJDORF032
9455 atggcttgttttgctaaagcgagtagtgaactaccactgtcaccactactaccactgtcagacgaatcactaggtgatccacct
1 M A C F A K A S S E L P Ξ P L P L S D E S L G D P P
9371 ttaccgtctaatttaccaccccaagctagaatagtattcgcaccgtctaaaaatggattaccatag 9306
29 L P S N P P Q A R I V F A P S K N G P *
44AHJDORF034
14146 atgatgattctaataataaaaacgctaaaaagcataaatacgctttatataatttacaagctaaaaataataattcttcaatgt
1 M M I L I I K T L K S I N T L Y I I Y K L K I I I L Q C
14062 ataaatatattaaagaaatcgatactttatataaagaaattggtaaatcagatagaccagtga 14000
29 I N I L K K S I L Y I K K L V N Q I D Q *
44AHJDORF035
13957 atgcaacatttgacgaataaatttaacactgtaaacgacatcataaactattacaaggagcaaaaacatggtaaaacaaaatcg
1 M Q H L T N K F N T V N D I I N Y Y K E Q K H G K T K S
13873 tttagacatggtaagagattatcaaaatgctgtcaatcatgtcagaaaaaaaatcccagataa 13811
29 F R H G K R L S K C C Q S C Q K K N P R *
44AHJDORF036
10165 gtgtatacaataccacacgtgatggtgcaacatatggtggtacattatagtttgcaactaaaaacgaaccatcttcaaaaactg
1 V Y T I P H V M V Q H M V V H Y S Q K T N H L Q K L
10081 ctacaacaacacctgtgtgaccaataccatatgcagttgcttgtaagtatggtggtttactag 10019
29 Q Q H C D Q Y H M Q V S M V V Y *
44AHJDORF037
14788 atgtcgatatctaacgtaaataactctttttcaatttcaaaatcatcatattgtttgtcaaactcaatatacacatcacccata
1 M S I S N V N N S F S I S K S S Y C L S N S I Y T S P I
14872 tttatttttactatacattttttattagatgaagtaaatttttcaaatttatcattataa 14931
29 F I F T I H F L L D E V N F S N L S L *
44AH DORF038
3671 gtgtaccttttgcattacctttttgattttgattacgttttgcgttttgattactttcgttact gatttattcacagttttac
1 V Y L H Y L F D F D Y V R F D Y F R Y S I Y S Q F Y
3587 cgttatcaatcgtattattatcagcgaatcgtaacgttgtattatcaacatcaatgttaa 3528
29 R Y Q S Y Y Y Q R I V T L Y Y Q H Q C *
44AHJDORF039
1743 gtgctgtatttacttatgatgtatctaaacttaaagagtttactggcaacgttgaagaaattaaaccaaaatcagatttatatg
1 V Y L L M M Y L N L K S L L A T L K K L N Q N Q I Y M
1827 cgtttattttggatattaattcaattaaatataaacgttacacaaaaggtatgttaa 1883
29 R F W I L I Q L N I N V T Q K V C *
44AHDORF040
9740 gtggtaactggacatatgcacagttaccagaaaaatataaaaaagcaattggtgtacctttattcaaaaaagaatacttataca
1 V V T G H M H S Y Q K N I K K Q L V Y Y S K K N T Y T
9824 aaccaggtaacatatttcctcaaacgggtaatgcaggacaatgtacagaattaa 9877
29 N Q V T Y F L K R V M Q D N V Q N *
44AHJDORF041
15836 atgtcgtcaactttcattattatatcactcctttctaaaaaacgtaaacgttatacgtttcataaaatcctttatgcatattcc
1 M S S T F I I I S L L S K K R K R Y T F H K I L Y A Y S
15920 attgttctattgggtcatcaccagcaatataagacaatattgattctggtttag 15973
29 I V L G H H Q Q Y K T I L I L V *
44AHJDORF0 2
5151 atgcacgaccgtcgtcttttgttaatttatagttttgtgaacctcttgcgcgtaatgcttcaaagtgttcatactcaccaagtt
1 M H D R R L I Y S F V N L R V M L Q S V H T H Q V
5067 ggaagaaaccatataaattatggaaacgttttccaccaccgccgtttgtcatag 5014
29 G R N H I N Y G N V F H H R R S *
44AHJDORF043
4539 atgcgacttgtaacagttttgcaacaccatcgtgatgtaaccagattttcatttcaccattggattgacgttctaatccgattg
1 M R L V T V Q H H R D V T R F S F H H I D V L I R L
4455 ttgtaccatgaccaccctgtacaatacgcatgcttgaaattaagtcaccactag 4402
29 L Y H D H P V Q Y A C L K L S H H *
44AHJDORF044
12917 atgttacctatttacgtgatgatatgttttataaagaaaacatggaacgttattactacaatccaagcaatttacattttgaca
1 M L P I Y V M I C F I K K T W N V I T T I Q A I Y I T
12833 atgcttactctaaaaattacgtggttgataatgatagatatttatatttag 12783
29 M T K I T W L I M I D I Y I *
44AHJDORF149
770 atgattgttttgaaagtgaatgaatttgtacaccataactatcttcacttttatttgtatcaattgacatgttttcatttaatt
1 M I V L K V N E F V H H N Y L H F Y Y Q T C F H L I
686 ctgttcgtttatttaatcttgaatcttcatatgatgtacccatcatag 639
29 F V Y L I N H M Y P S *
44AHJDORF046
4891 atgattatccatttaagttatcatatcaagacggtattaatttcccacgtgataactttaaagagcctgagggtatt-tgcSttt
1 M I I H S Y H I K T V L I S H V I T L K S L R V^F A F
4975 atacaaatccaaaaacaaaacgtaaatcgttattacttgctatga 5019
29 I Q I Q K Q N V N R Y Y L L *
44AHJDORF047
11911 atgaatgtatgtaagttgttcaggtgtgagttttgcaaaacatttcacagcatagtcataggcttcactatcattcatatcatt
1 M N V C K L F R C E F C K T F H S I V I G F T I I H I I
11995 atctttatcaaaaatcgtataattaaaatctgttttaagttgtga 12039
29 I F I K N R I I K I C F K L *
44AHJDORF045
10655 atggcaccgtcaaagaattgttcacgtacaaaggtttcaaaatcgacgcttgtatcaaaggcgtttttcggtataccagcagaa
1 M A P S K N C S R T K V S K S T L V S K A F F G I P A E
10739 gcaattttaatctttccattcacttcatatgcatatttcttatga 10783
29 A I I F P F T S Y A Y F *
44AHJDORF048
15340 atgaggacgttgttgacattatcaatgctggagaagttcaattcacaatttatgaatatgaaaacaaaaaaggtcaaaaaggtt
1 M R T L L T L S M L E K F N S Q F M N M K T K K V K K V
15256 actcaatcaattttggtcaagtatcattttaatacaatttcatag 15212
29 T Q S I L V K Y H F N T I S *
44AHJDORF049
5784 atgagggggcaggtgttgactttgatggtgcatatggatttcaatgtatggacttatcagttgcttatgtgtattacattactg
1 M R G Q V L T L M V H M D F N V W T Y Q L L M C I T L
5868 acggtaaagttcgcatgtggggtaatgctaaagacgcgataa 5909
29 T V K F A C G V M L K T R *
44AHJDORF050
13158 gtgtgttacgtttttcattcacgtaatcgtttcgtcgcatttctaaaaaaatgtttttgtaaagtcttgatgtattcattttat
1 V C Y V F H S R N R F V A F L K K C F C K V L M Y S F Y
13242 gcttttgtaataaattgtatatatttaaattggataatatag 13283
29 A F V I N C I Y L N W I I *
44AHJDORF051
11066 atgataacaatgaactatacaatatcattaacggttacaaaaacactgaacgtaatatattattctctacatttgtcacatcac
1 M I T M N Y T I S L T V T K T L N V I Y Y S L H S H H
10982 gttcattgtataacttattggttcctttccaatacttaa 10944
29 V H C I T Y W F L S N T *
44AHJDORF052
14338 atgattttagtaatgttaattttaaatttgatgataaagatttacaagaggcgtacattgacacatggaaacattttgcacatc
1 M I L V M L I L N L M I K I Y K R R T T H G N I L H I
14254 tgccctattttcctaaagaaagaaacgtatcatatgtaa 14216
29 C P I F L K K E T Y H M *
44AHJDORF053
3348 atgtggtttattcatcaagtgaagttgaaaaatacttacaatcacaaggcttcacagaacacaatgaagatacaacaagtaaca
1 M W F I H Q V K L K N T Y N H K A S Q N T M K I Q Q V T
3432 ctgatgaaacatcgaatcaaaatgctacatctttag 3467
29 L M K H R I K M L H L *
4 AHJDORF054
7551 atgactggaatggaaatacgatgttactcgacgctggtaagatttcacaaaaaactggtgttaagttacgtacaaaatcaatta
1 M T G M E I R C Y S T V R F H K K L V L S Y V Q N Q L
7635 ttggttatcataatgaagttcgagtatatccagtag 7670
29 V I I M K F E Y I Q *
44AHJDORF055
15705 atgtgtctggtaataattcttttgcttgtgttttggttaaatgatactcgtgaagtggtaaaaattcctcaatgtattcattat
1 M C L V I I L L V F W L N D T R E V V K I P Q C I H Y
15789 catcatctaagtaatgaagtatataacctttga 15821
29 H H L S N E V Y N L *
44AHJDORF056
5512 gtgagtattacattacaggtaaccaaatggaattatttagagacgcgccagaagaaattaaaaaagtgggtgcatggttacgtg
1 V S I T Ii Q V T K W N Y L E T R Q K K K K W V H G Y V
5596 tgtcaagtggtaacgcagtcggtgaagtaa 5625
29 C Q V V T Q S V K *
44AHJDORF057
10121 atgtaccaccatatgttgcaccatcacgtgtggtattgtatacactcattaatggcgtaccaaataatgctggtgataatattg
1 M Y H H M L H H H V W Y C I H S L M A Y Q I M L V I I L
10205 tattctttagtggtattgcttaattaa 10231
29 Y S L V V L L N *
44AHJDORF058
10767 atgcatatttcttatgattcagtacaaacatc tatctatctgttcgttttcaatatcccatttacctaaggctatcgggtcga
1 M H I S Y D S V Q T S Y L S V R F Q Y P I Y L R L S G R
10851 ataaactggggttcaataagggtttaa 10877
29 I N W G S I R V *
44AHJDORF164
702 atgttttcatttaattctgttcgtttatttaatcttgaatcttcatatgatgtacccatcatagaacgcatgttgtttccctca
1 M F S F N S V R F N E S S Y D V P I I E R M F P S
618 tacatgtttaaattcctcctaatctaa 592
29 Y M F K F L I *
44AHJDORF059
8360 atggattttgtaacattggattacctgaaccg attatgccaaaatcttacaccagattctaaaattgcttttaatt*ttcca
1 M D F V T D Y N R H Y A K I H Q I K L L i l V P
8276 ttaacatggggtcgatgtcacgtatag 8250
29 T W G R C H V *
44AHJDORF060
6257 atgtaccattttcatttctataatatgtgccgtattggtttcgtttccattttccaaatgtatttacttttgatgtttctaatg
1 M Y H F H F Y N M C R I G F V S I F Q M Y L L L M F M
6173 ctttgctattactacctgaaaatttag 6147
29 C Y Y Y K I *
44AHJDORF061
15551 atgtgttttggtgtcttgataaaatatcttttacgtttgtcattttatttctcctcttatttaaattatttgctttctgcaatt
1 M C F G V L I K Y L L R Ξ F Y F S S Y L N Y L S A I
15635 gcgatttgtagtaaatcattgtaa 15658
29 A I C S K S *
44AHJDORF062
4285 gtggtattcgcaacgcagttaaccaatctattaatattgataaagaaacaaatcacatgtactctacacaatccgattctcaaa
1 V V F A T Q L T N L L I L I K K Q I T C T L H N P I L K
4369 aacctgaaggtttttggataa 4389
29 N K V F G *
44AHJDORF063
9487 atgcgtcttgtattttttttaataattcttgcatggcttgttttgctaaagcgagtagtgaactaccactgtcaccactactac
1 M R L V F F L I I L A W V L K R V V N Y H C H H Y Y
9403 cactgtcagacgaatcactag 9383
29 H C Q T N H *
44AHJDORF065
5029 gtggtggaaaacgtttccataatttatatggtttcttccaacttggtgagtatgaacactttgaagcattacgcgcaagaggtt
1 V V E N V S I I Y M V S S N L V S M N T L K H Y A Q E V
5113 cacaaaactataaattaa 5130
29 H K T I N *
44AHJDORF064
2609 atgacgagtcaatcaatcaacttgtgtccgaaatatataacggtgcaccatttgttaaaatgtcacctatgtttaatgcagatg
1 M T S Q S I N C P K Y I T V H H L K C H L C M Q M
2693 acgatatcattgatttaa 2710
29 T I S L I *
44AHJDORF066
10481 atgatattctttatattgaaagtgacatcggttcattttcacttaacgacttatttccagttgaacgttcagtacataacaaat
1 M I F F I L K V T S V H F H L T T Y F Q L N V Q Y I T N
10397 ctgatttgcatatattaa 10380
29 I C I Y *
Table 19
Sequence similarities between ORFs 44AHJD and public databases
Phage : 44AHJD
Database : nr
Query= sid| 110871 | lan|44AHJDORF001 Phage 44AHJD ORF| 10342-12627 | -1 (761 letters) gi 118848|sp|P19894|DPO _BPM2 DNA POLYMERASE >gi | 76896 |pir| | JQO . 55 le-06 gi 1072656|pir| |S51275 DNA polymerase - phage CP-1 >gij 836593 |e. 53 6e-06 gi 1429230 j emb|CAA67649| (X99260) DNA polymerase [Bacteriophage. 49 le-04 gi 1572479 jemb|CAA65712J (X96987) DNA polymerase [Bacteriophage. 46 0.001 gi 118851 |sp|P06950|DPOL_BPPZA DNA POLYMERASE (EARLY PROTEIN GP. 45 0.002 gi 2435429 (AF012250) unassigned reading frame (possible DNA po. 45 0.002 gi 1084487|pir| |S41618 DNA polymerase - slime mold (Physarum po. 45 0.002 gi 4877819|gb|AAD31446.l| (AF133505) DNA polymerase [Neurospora. 44 0.004 g 461962|sp|P33537|DPOM_NEUCR PROBABLE DNA POLYMERASE >gi|2833. 44 0.004 gi 2499511 |sp|Q1247l|6P22_YEAST 6-PHOSPHOFRUCTO-2-KINASE 2 (PHO. 41 0.041 gi 2258375 jgbj AAD11909.1 | (AF007261) transcription initiation f. 40 0.070 gi 15734|emb|CAA37450| (X53370) DNA polymerase (AA 1-575) [Bact. 39 0.092
Query= sid | 110872 | Ian | 44AHJDORF002 Phage 44AHJD ORF| 3789-5732 | 3 (647 letters) gi|135273|sp|P27622|TAGC_BACSU TEICHOIC ACID BIOSYNTHESIS PROTE .. 112 7e-24 gi j 142847 (M64050) DNase inhibitor (Bacillus subtilis] 52 le-05 gi|4038407 (AF103943) factor C protein precursor [Streptomyces .. 39 0.10
Query= sid| 110873 | lan|44AHJDORF003 Phage 44AHJD ORF| 6626-8389 | 2 (587 letters) gi|l38123|sp|P0433l|VG9_BPPH2 TAIL PROTEIN (LATE PROTEIN GP9) >.. 92 8e-18 gi| 138124 jsp|P07534|VG9_BPPZA TAIL PROTEIN (LATE PROTEIN GP9) >.. 82 le-14 gi|1429238|emb|CAA67657| (X99260) tail protein [Bacteriophage B.. 78 2e-13 gi|215339 (M12456) p9 tail protein [Bacteriophage phi-29] >gi|2.. 71 2e-ll gi|1181968|emb|CAA87738.1| (Z47794) tail protein [Bacteriophage.. 54 3e-06 gij 1181970 jemb|CAA87740.1| (Z47794) tail protein [Bacteriophage.. 42 0.010
Query= sid| 110875 | lan|44AHJDORF005 Phage 44AHJD ORF| 12643-13890 | -1 (415 letters) gi 13845203 (AE001399) GAF domain protein (cyclic nt signal tran... 52 6e-06 gij 3758843 |emb|CAB11128.1| (Z98551) predicted using hexExon; MA... 49 5e-05 gi 13845297 (AE001421) hypothetical protein [Plasmodium falciparum] 48 le-04 gij 4493936 |emb|CAB38972.l| (AL034556) predicted using hexExon; ... 47 2e-04 gi 13845165 (AE001390) hypothetical protein [Plasmodium falciparum] 46 6e-04
Query= sid| 110877 | lan| 44AHJDORF007 Phage 44AHJD ORF| 2044-3027 | 1 (327 letters) gi|ll81960|emb|CAA87731.1| (Z47794) connector protein [Bacterio... 46 Se-04 gij 1429239 jem jcAA67658| (X99260) upper collar protein [Bacteri... 45 8e-04 gijl37915|sp|P07535|VG10_BPPZA UPPER COLLAR PROTEIN (CONNECTOR ... 44 0.002 gi|137914|sp|P04332|VG10_BPPH2 UPPER COLLAR PROTEIN (CONNECTOR ... 41 0.009
Query= sid| 110878 | lan| 44AHJDORF008 Phage 44AHJD ORF| 3020-3775 | 2 (251 letters) g 498246 8|gb|AAD30963.2| (AF118151) SNFl/AMP-activated kinase ... 52 3e-06 gi 1730077|sp|P18160|KYKl_DICDI NON-RECEPTOR TYROSINE KINASE SP ... 46 2e-04 gi 3758855 jemb|CAB11140.1| (Z98551) predicted using hexExon; MA... 46 2e-04 gi 585795 |sp|P21538|REBl_YEAST DNA-BINDING PROTEIN REB1 (QBP) >... 46 3e-04 gi 172372 (M58728) DNA-binding protein [Saccharomyces cerevisiae] 46 3e-04 gi 2952545 (AF051898) coronin binding protein [Dictyostelium di... 45 6e-04 gi 535260 |emb|CAA82996| (Z30339) STARP antigen [Plasmodium reic ... 45 7e-04 gi 1429240|emb|CAA67659| (X99260) lower collar protein [Bacteri... 44 0.001
28.
Query= sid 1110879 | Ian | 44AHJDORF009 Phage 44AHJD ORF| 5744-6496 | 2 (250 letters) gi 2764981 |emb|CAA69021.l| (Y07739) N-acetylmuramoyl-L-alanine ... 180 le-44 gi 1136751 sp|P24556|ALYS_STAAU AOTOLYSIN (N-ACETYLMURAMOYL-L-AL... 118 6e-26 gi 1763243 (U72397) amidase [bacteriophage 80 alpha] 118 6e-26 gi 4574237 |gb|AAD23962.l|AF106851_l (AF106851) LytN [Staphyloco... 84 9e-16 gi 3767593 jdb |BAA33856.1| (AB015195) LytN [Staphylococcus aureus] 84 9e-16 gi 2764983 jemb|CAA69022.l| (Y07740) cell wall hydrolase Plyl87 ... 77 2e-13 gi 3287732 jsp|O05156|ALEl_STACP GLYCYL-GLYCINE ENDOPEPTIDASE AL... 73 2e-12 gi 79926|pir| |A25881 lysostaphin precursor - Staphylococcus aim... 69 3e-ll gi 126496 | spjpi0548|LSTP_STAST LYSOSTAPHIN PRECURSOR (GLYCYL-GL... 69 3e-ll gi 3287967 |sp|P10547|LSTP_STASI LYSOSTAPHIN PRECURSOR (GLYCYL-G... 69 3e-ll gi 3341932 jdb |BAA31898.1| (AB009866) amidase (peptidoglycan hy... 68 6e-ll
Query= sid | 110882 | Ian | 44AHJDORF012 Phage 44AHJD ORF| 8391-8813 | 3 (140 letters) gi|l40528|sp|P2481l|YQXH_BACSU HYPOTHETICAL 15.7 KD PROTEIN IN 80 6e-15 gij 41266311 bj |BAA36651.l| (AB016282) ORF45 [bacteriophage phi- 76 le-13 gij 141088 |sp|P26835|YNGD_CLOPE HYPOTHETICAL 14.9 KD PROTEIN IN 61 4e-09 gij 2293160 (AF008220) YtkC [Bacillus subtilis] >gi | 2635548 | emb| 36 0.099 gijll81973|emb|CAA87743.l| (Z47794) holin protein [Bacteriophag 31 3.3
Table 20 Homolgies between phage 44 AHJD ORFs and proteins in public databases
Query= pt| 110871 44AHJDORF001 Phage 44AHJD ORF ] 10342-12627 | -1 1 (761 letters)
>gi|H8848|sp|P19894|DPOL_BPM2 DNA POLYMERASE >gi | 76896 |pir| | JQ0161 DNA-directed DNA polymerase (EC 2.7.7.7) - phage M2 >gi 1215509 (M33144) DNA polymerase [Bacteriophage M2] Length = 572
Score = 55.4 bits (131), Expect = le-06
Identities = 96/426 (22%), Positives = 159/426 (36%), Gaps = 88/426 (20%)
Query: 229 KLTPEQLTYIHNDVIILGMCHIHYSDIFPNFDYNKLTFSLNIMESYLNNEMTR FQ 283
++TPE+ YI ND+ 1+ DI +++T + ++ + + T+ F Sbjct: 154 EITPEEYEYIKNDIEIIARA LDIQFKQGLDRMTAGSDSLKGFKDILSTKKFNKVFP 209
Query: 284 LLNQYQDIKISYTHYHFHDMNFYDYIKSFYRGGLNMYNTKYINKLIDEPCFΞIDINSSYP 343
L+ D +1 + YRGG N KY K I E D+NS YP
Sbjct: 210 KLSLPMDKEI-- RKAYRGGFT LNDKYKEKEIGEGMV-FDVNSLYP 252
Query: 344 YVMYHEKIPT LYFYEHYSEPTLIPTFLDDDNYFSLYKIDKDVFNDDLLIKIKSRVLRQM 403
MY +P Y P + + D + LY I + F +L K + +
Sbj ct : 253 SQMYSRPLP YGAPIVFQGKYEKDEQYPLY- IQRIRFEFEL KEGYIPTI 299
Query: 404 XXXXXXXXXXXXXXXXXXLRMIQ-DITGIDCMHIRVNSFVIYECEYFHARDIIFQNYFIK 462
+ ++ +T +D 1+ + + +Y EY F +
Sbjct: 300 QIKKNPFFKGNEYLKNSGVEPVELYLTNVDLELIQEH-YELYNVEYIDGFK FRE 352
Query: 463 TQGKLKNKINMTSPYDYHITDDINEHPYSNEEVMLSKWLNGLYG IPAL 511
G K+ 1+ + H + L+K++LN LYG +P L
Sbjct: 353 KTGLFKDFIDKWTYVKTH EEGAKKQLAKLMLNSLYGKFASNPDVTGKVPYL 403
Query: 512 RSHFNL-FRLDDNNELYNIINGYKNTERNILFSTFVTSRSLYNLLVPFQYLTESEIDDNF 570
+ +L FR+ D YK+ + F+T+ + + + Q D Sbjct: 404 KDDGSLGFRVGDEE YKDPVYTPM-GVFITAARFTTITAAQACY DRI 449
Query: 571 IYCDTDSLYMKSWKPLLNPSLFDPIALGKDIENEQIDKMFVLNHKK YAYEVNG 625
IYCDTDS+++ P + + DP LG E+ + L K Y EV+G Sbjct: 450 IYCDTDSIHLTGTEVPEIIKDIVDPKKLGYWAHES-TFKRAKYLRQKTYIQDIYVKEVDG 508
Query: 626 KIKIAS 631
K+K S Sbjct: 509 KLKECS 514
>gi|l072656 |pir I |S51275 DNA polymerase - phage CP-1
>gi|836593|emb|CAA87725.l| (Z47794) DNA polymerase [Bacteriophage CP-1] Length = 568
Score = 53.5 bits (126), Expect = 6e-06
Identities = 104/464 (22%) , Positives = 169/464 (36%) , Gaps = 66/464 (14%)
Query: 230 LTPEQLTYIHNDVIIL--GMCHIHYSDIFPNFDYNKLTFSLNIMESYLNNEMTRFQLLNQ 287
+ PE + YIH DV IL G+ ++Y + F Y + +L + +F+ Sbjct: 152 IKPE IDYIHVDVAILARGIFAMYYEENFTK--YTSASEALTEFKRIFRKSKRKFRDFFP 209
Query: 288 YQDIKISYTHYHFHDMNFYDYIKSFYRGGLNMYNTKYINKLIDEPCFSIDINSSYPYVMY 347
D K+ D+ + G + K+ + +++ DINS YP M
Sbjct: 210 ILDEKVD DFCRKHIVGAGRLPTLKHRGRTLNQLIDIYDINSMYPATML 257
Query: 348 HEKIPT LYFYEHYSEPTLIPTFLDDDNYFSLY-KIDKDVFNDDL-LIKIKSRVLRQMXX 405 - _ —
+P + + Y P + +D+Y+ + K D D+ L I+IK ++ __ ~
Sbjct: 258 QNALPIGIP--KRYKGK---PKEIKEDHYYIYHIKADFDLKRGYLPTIQIKKKLDALRIG 312 "
Query: 406 XXXXXXXXXXXXXXXXLRMIQDITGIDCMHIRVNSFVIYECEYFHARDIIFQNYFIKTQG 465
L + + H + E F +F +Y Sbjct: 313 VRTSDYVTTSKNEVIDLYLTNFDLDLFLKHYDATIMYVETLE-FQTEΞDLFDDYI 366
Query: 466 KLKNKINMTSPYDYHITDDINEHPYΞNEEVMLΞKWLNGLYGIPALR--SHFNLFRLDDN 523
+ Y Y E+ S E +K++LN LYG + S L LDD Sbjct: 367 TTYRYK KENAQSPAEKQKAKIMLNSLYGKFGAKIISVKKLAYLDDK 412
Query: 524 NELYNIINGYKNTERNIL FSTFVTSRSLYNLLVPFQYLTESEIDDNFIYCDTDS 577
L +KN + + + FVTS + + ++ Q E DNF+Y DTDS Sbjct: 413 GILR FKNDDEEEVQPVYAPVALFVTSIARHFIISNAQ ENYDNFLYADTDS 462
Query: 578 LYMKSWKPLLNPSLFDPIALGK DIENEQIDKMFVLNHKKYAYEVNGKIKIASAGIPKN 637
L++ +L+ DP GKW E + K L K Y E+ + + K Sbjct: 463 LHLFHSDSLVLD IDPSEFGKWAHEGRAV-KAKYLRSKLYIEELIQEDGTTHLDV-KG 517
Query: 638 AFDTSVDFETFVREQFFDGAIIENNKSIYNEQGTIΞIYPSKTEI 681
A T E E F GA E ++ +G IY + +1 Sbjct: 518 AGMTPEIKEKITFENFVIGATFEGKRASKQIKGGTLIYETTFKI 561
>gi|l429230|emb|CAA67649| (X99260) DNA polymerase [Bacteriophage B103] Length = 572
Score = 49.2 bits (115), Expect = le-04
Identities = 93/422 (22%), Positives = 155/422 (36%), Gaps = 88/422 (20%)
Query: 229 KLTPEQLTYIHNDVIILGMCHIHYΞDIFPNFDYNKLTFSLNIMESYLNNEMTR FQ 283
++TPE+ YI ND+ 1+ DI +++T + ++ + + T+ F Sbjct: 154 EITPEEYEYIKNDIEIIARA LDIQFKQGLDRMTAGSDSLKGFKDILSTKKFNKVFP 209
Query: 284 LLNQYQDIKISYTHYHFHDMNFYDYIKSFYRGGLNMYNTKYINKLIDEPCFSIDINSSYP 343
L+ D +1 + YRGG N KY K I E D+NS YP
Sbjct: 210 KLSLPMDKEI RRAYRGGFT LNDKYKEKEIGEGMV-FDVNSLYP 252
Query: 344 YVMYHEKIPTWLYFYEHYSEPTLIPTFLDDDNYFSLYKIDKDVFNDDLLIKIKSRVLRQM 403
MY +P Y P + + D + LY I + F +L K + + Sbjct: 253 SQMYSRPLP -YGAPIVFQGKYEKDEQYPLY-IQRIRFEFEL KEGYIPTI 299
Query: 404 XXXXXXXXXXXXXXXXXXLRMIQ-DITGIDCMHIRVNSFVIYECEYFHARDIIFQNYFIK 462
++ +T +D 1+ + + +Y EY F +
Sbjct: 300 QIKKNPFFKGNEYLKNSGAEPVELYLTNVDLELIQEH-YEMYNVEYIDGFK FRE 352
Query: 463 TQGKLKNKINMTSPYDYHITDDINEHPYSNEEVMLSKWLNGLYG IPAL 511
G K 1+ + H + L+K++ + LYG +P L
Sbjct: 353 KTGLFKEFIDK TYVKTH EKGAKKQLAKLMFDSLYGKFASNPDVTGKVPYL 403
Query: 512 RSHFNL-FRLDDNNELYNIINGYKNTERNILFSTFVTSRSLYNLLVPFQYLTESEIDDNF 570
+ +L FR+ D YK+ + F+T+ + + + Q D Sbjct: 404 KEDGSLGFRVGDEE YKDPVYTPM-GVFITAWARFTTITAAQAC DRI 449
Query: 571 IYCDTDSLYMKSWKPLLNPSLFDPIALGKWDIENEQIDKMFVLNHKK YAYEVNG 625
IYCDTDS+++ P + + DP LG E+ + L K YA EV+G Sbjct: 450 IYCDTDSIHLTGTEVPEIIKDIVDPKKLGYAHES-TFKRAKYLRQKTYIQDIYAKEVDG 508
Query: 626 KI 627
K+ Sbjct: 509 KL 510
>gi|1572479|emb|CAA65712| (X96987) DNA polymerase [Bacteriophage GA-1] Length = 578
Score = 46.1 bits (107), Expect = 0.001
Identities = 80/376 (21%) , Positives = 146/376 (38%) , Gaps = 54/376 (14%)
Query: 234 QLTYIHNDVIILGMCHIHYSDIFPNFDYNKLTFSLNIMESYLNNEMTRFQLLNQYQDIKI 293
++ Y+ +D++I+ + +F N D+ +T + + +Y EM + +Y + Sbjct: 162 EIEYLKHDLLIVALA---LRSMFDN-DFTSMTVGSDALNTY--KEMLGVKQ EKYFPVL- 214
Query: 294 SYTHYHFHDMNFYDYIKSFYRGGLNMYNTKYINKLIDEPCFSIDINSSYPYVMYHEKIPT 353
+ 1+ Y+GG N KY + + D+NS YP +M ++ +P Sbjct: 215 --SLKVNSEIRKAYKGGFTWVNPKYQGETVYGGMV-FDVNSMYPAMMKNKLLP- 264
Query: 354 WLYFYEHYSEPTLIPTFLDDDNYFSLYKIDKDVFNDDLLIKIKSRVLRQMXXXXXXXXXX 413 Y EP + + + LY F + KI ++
Sbjct: 265 YGEPVMFKGEYKKNVEYPLYIQQVRCFFELKKDKIPCIQIKGNARFGQNEYLS 317
Query: 414 XXXXXXXXLRMIQDITGIDCMHIRVNSFVIYECEYFHARDIIFQNYFIKTQGKLKNKINM 473
L +T +D 1+ + + I+E E+ +F+ + I
Sbjct: 318 TSGDEYVDLY VTNVD ELIKKH-YDIFEEEFIGG--FMFKGF IGF 359
Query: 474 TSPYDYHITDDINEHPYSNEEVMLSKWLNGLYGIPALRSHFN--LFRLDDNNELYNIIN 531
Y + N S E+ + +K++LN LYG A + LD+N L Sbjct: 360 FDEYIDRFMEIKNSPDSSAEQΞLQAKLMLNSLYGKFATNPDITGKVPYLDENGVLKFRKG 419
Query: 532 GYKNTERNILFST---FVTSRSLYNLLVPFQYLTESEIDDNFIYCDTDSLYMKΞWKPLL 588
K ER+ +++ F+T+ + N+L Q L FIY DTDS++++ + + Sbjct: 420 ELK--ERDPVYTPMGCFITAYARENILSNAQKLYP RFIYADTDSIHVEGLGEVDA 472
Query: 589 NPSLFDPIALGKWDIE 604
+ DP LG WD E Sbjct: 473 IKDVIDPKKLGYWDHE 488
>gi| 118851 |sp|P06950|DPOL_BPPZA DNA POLYMERASE (EARLY PROTEIN GP2)
>gi| 75812 jpir| |ERBP2Z DNA-directed DNA polymerase (EC 2.7.7.7) - phage PZA >gi|216051 (M11813) gene 2 product
[Bacteriophage PZA] >gi | 224741 |prf | | 1112171E ORF 2
[Bacteriophage PZA] Length = 572
Score = 45.3 bits (105), Expect = 0.002
Identities = 98/461 (21%), Positives = 166/461 (35%), Gaps = 110/461 (23%)
Query: 198 QLKTDFNYTIFDKDNDMNDSEAYDYAVKCFAKLTPEQLTYIHNDVIILGMCHIHYSDIFP 257
++ DF T+ D D + Y ++TP++ YI ND+ 1+ + I Sbjct: 129 KIAKDFKLTVLKGDIDYHKERPVGY EITPDEYAYIKNDIQIIAEALL IQF 178
Query: 258 NFDYNKLTFSLNIMESYLNNEMTR FQLLNQYQDIKISYTHYHFHDMNFYDYIKSF 312
+++T + ++ + + T+ F L+ D ++ Y Sbjct: 179 KQGLDRMTAGSDDLKGFKDIITTKKFKKVFPTLSLGLDKEVRYA 222
Query: 313 YRGGLNMYNTKYINKLIDEPCFSIDINSSYPYVMYHEKIPTWLYFYEHYSEPTLIPT--F 370
YRGG N ++ K I E D+NS YP MY +P Y EP + Sbjct: 223 YRGGFTWLNDRFKEKEIGEGMV-FDVNSLYPAQMYSRLLP YGEPIVFEGKYV 273
Query: 371 LDDDNYFSLYKID KDVF-TODLLIKIKSRVLRQ-1XXXXX_XXXX__XXXXXXXLRMI 425
D+D + 1 K+ + + IK +SR + Sbjct: 274 WDEDYPLHIQHIRCEFELKEGYIPTIQIK-RSRFYKGNEYLKSSGGEIADLW 324
Query: 426 QDITGIDCMHIRVNSFVIYECEYFHARDIIFQNYFIKTQGKLKNKINMTSPYDYHITDDI 485
++ +D + + + +Y EY F T G K+ 1+ + I
Sbjct: 325 --VSNVD-LELMKEHYDLYNVEYISGLK FKATTGLFKDFIDKWTHIKTTΞEGAI 375
Query: 486 NEHPYSNEEVMLSKWLNGLYG IPALRSHFNL-FRLDDNNELYNIINGY 533
+ L+K++LN LYG +P L+ + L FRL G
Sbjct: 376 KQ LAKLMLNSLYGKFASNPDVTGKVPYLKENGALGFRL GE 415
Query: 534 KNTERNIL--FSTFVTSRSLYNLLVPFQYLTESEIDDNFIYCDTDSLYMKSWKPLLNPS 591
+ T+ + F+T+ + Y + Q D IYCDTDS+++ P + Sbjct: 416 EETKDPVYTPMGVFITAWARYTTITAAQACF DRIIYCDTDSIHLTGTEIPDVIKD 470
Query: 592 LFDPIALGKWDIENEQIDKMFVLNHKKYAY EVNGKI 627
+ DP LG E+ + L K Y EV+GK+ Sbjct: 471 IVDPKKLGYWAHES-TFKRAKYLRQKTYIQDIYMKEVDGKL 510
>gi|2435429 (AF012250) unassigned reading frame (possible DNA polymerase) [Physarum polycephalum] Length = 544
Score = 44.9 bits (104), Expect = 0.002
Identities = 118/545 (21%) , Positives = 206/545 (37%) , Gaps = 104/545 (19%)
Query: 179 TSIATLGKKLLDGGYLTESQLKTDFNYTIFDKDNDMNDSEAYDYAVKCFAKLTPEQLTYI 238 ~ ~-
T + L K L D + T Q F N M Y + CF L P++ I - Sbjct: 62 TQLFNLLKSLQDSSFYTFKQ FTYQNIM YSLEISCF--LYPKKKILI 105
Query: 239 HNDVII GMCHIHYSDIFPNFD YNKL--TFSLNIMESY-LNNEMTRFQLLNQYQD 290
D+ +1 Y+D+ ++ YN++ +++NI Y L+ ++ + Sbjct: 106 -KDLYNFFSENIIYNDWKDYKLLAILYNEIQTAYNININRKYILSTASLSLRIFKKSFP 164
Query: 291 IKISYTHYHFHD.lNFYDYIKSFYRGG-_WrYNT_7riNKLIDEPCFSIDINSSYPYVMYHEK 350
K + D + +YI+ Y GG N I + + + + D+NS YPY+M EK Sbjct: 165 EKYRLIPHLTRDED--NYIRKSYIGGRNE IFEHVAQRNYFYDVNSLYPYIMKKEK 217
Query: 351 IPTWLYFYEHYSEPTLIPTFLDD-DNYFS LYKIDKDVFNDDLL IKIKSRVLRQ 402
+P + Y + + F + +N+F L I+K N +L + IK+ V Sbjct: 218 MPIGI PEYRDKEYMKKFEKNIENFFGFIDVLITIEKTNNNIPVLPYRMGIKNNV-EV 273
Query: 403 MXXXXXXXXXXXXXXXXXXLRMIQDITGIDCMHIRVNSFVIYECEYFHARDIIFQNYFIK 462
L + Q 1+ IY + ++++F+ Y +
Sbjct: 274 GIIYAKGTLRGIYFSEEIKLALKQGYKIIE IYSAYEYKEKEWFEEYVEQ 323
Query: 463 TQGK-LKNKINMTSPYDYHITDDINEHPYSNEEVMLSKWLNGLYG -IPALRS 513
+ LK K D + D L K +LN LYG I +
Sbjct: 324 MYNRRLKAK DPALKD LYKKLLNTLYGRFGLVYEQIDIISP 363
Query: 514 HFNLFRLDDNNELYNIINGYKNTERNILFΞTFVTSRSLYNLLVPFQYLTESEIDDNFIYC 573
L + DN + + + + N ++ + ++ + F Y T + + IY Sbjct: 364 EKEL--ITDNTYISHDTTEFIDITANTCYNNIAITSAITSYARIFMYNTILNYNLHVIYI 421
Query: 574 DTDSLYMKSWKPLLNPSLFDPIALGKWDIENEQIDKMFVLNHKKYAY-EVNGKIKIASA 632
DTD L++K+ P+ + +L +GK+ +E+ + F+ N K Y Y +N I Sbjct: 422 DTDGLFLKN PIPDIALTTSKEMGKFRLESINAEAHFIAN-KFYIYAPINSPIIYKFK 477
Query: 633 GIPK NAFDTSVDFETFVR EQFFDGAIIENNKSIYNEQGT ISIYPSK 678
GIP N D + + +F +I Y+ Q + I Y + Sbjct: 478 GIPLQKPIFNIHDIITQHKKILNITLGHHYFTFSIRLNNNQTYSFQASRKRKLIPNYKTT 537
Query: 679 TEIVC 683
I+C Sbjct: 538 PWIIC 542
>gi| 1084487 |pir I I S41618 DNA polymerase - slime mold (Physarum polycephalu ) >gi| 509721 | dbj |BAA06121.1 | (D29637) DNA polymerase [Physarum polycephalum] Length = 547
Score = 44.9 bits (104), Expect = 0.002
Identities = 118/545 (21%) , Positives = 206/545 (37%) , Gaps = 104/545 (19%)
Query: 179 TSIATLGKKLLDGGYLTESQLKTDFNYTIFDKDNDMNDSEAYDYAVKCFAKLTPEQLTYI 238
T + L K L D + T Q F N M Y + CF L P++ I Sbjct: 65 TQLFNLLKSLQDSSFYTFKQ FTYQNIM YSLEISCF--LYPKKKILI 108
Query: 239 HNDVIILGMCHIHYSDIFPNFD YNKL--TFSLNIMESY-LNNEMTRFQLLNQYQD 290
D+ +1 Y+D+ ++ YN++ +++NI Y L+ ++ + Sbjct: 109 -KDLYNFFSENIIYNDWKDYKLLAILYNEIQTAYNININRKYILSTASLSLRIFKKSFP 167
Query: 291 IKISYTHYHFHDrøFYDYIKSFYRGGLNHϊTCTKYINKLIDEPCFSIDINSSYPYVMYHEK 350
K + D + +YI+ Y GG N I + + + + D+NS YPY+M EK Sbjct: 168 EKYRLIPHLTRDED--NYIRKSYIGGRNE IFEHVAQRNYFYDVNSLYPYIMKKEK 220
Query: 351 IPTWLYFYEHYSEPTLIPTFLDD-DNYFS LYKIDKDVFNDDLL IKIKSRVLRQ 402
+P + Y + + F + +N+F L I+K N +L + IK+ V Sbjct: 221 MPIGI---PEYRDKEYMKKFEKNIENFFGFIDVLITIEKTNNNIPVLPYRMGIKNNV-EV 276
Query: 403 MXXXXXXXXXXXXXXXXXXLRMIQDITGIDCMHIRVNSFVIYECEYFHARDIIFQNYFIK 462
L + Q 1+ IY + ++++F+ Y +
Sbjct: 277 GIIYAKGTLRGIYFSEEIKLALKQGYKIIE IYSAYEYKEKEWFEEYVEQ 326
Query: 463 TQGK-LKNKINMTSPYDYHITDDINEHPYSNEEVMLSKWLNGLYG IPALRS 513
+ LK K D + D L K +LN LYG I +
Sbjct: 327 MYNRRLKAK DPALKD LYKKLLNTLYGRFGLVYEQIDIISP 366
Query: 514 HFNLFRLDDNNELYNIINGYKNTERNILFSTFVTSRSLYNLLVPFQYLTESEIDDNFIYC 573
L + DN + + + + N ++ + ++ + F Y T + + IY Sbjct: 367 EKEL--ITDNTYISHDTTEFIDITANTCYNNIAITSAITSYARIFMYNTILNYNLHVIYI 424
Query: 574 DTDSLYMKSVVKPLLNPSLFDPIALGKWDIENEQIDKMFVLNHKKYAY-EVNGKIKIASA 632
DTD L++K+ P+ + +L +GK+ +E+ + F+ N K Y Y +N I Sbjct: 425 DTDGLFLKN PIPDIALTTSKEMGKFRLESINAEAHFIAN-KFYIYAPINSPIIYKFK 480
Query: 633 GIPK NAFDTSVDFETFVR EQFFDGAIIENNKSIYNEQGT ISIYPSK 678
GIP N D + + +F +I N + Q + I Y +
Sbj ct : 481 GIPLQKPIFNIHDIITQHKKILNITLGHHYFTFSIRLNNNQTYSFQASRKRKLIPNYKTT 540
Query: 679 TEIVC 683
I+C Sbjct: 541 P IIC 545
>gi|4877819|gb|AAD31446.l| (AF133505) DNA polymerase [Neurospora crassa] Length = 1035
Score = 44.1 bits (102), Expect = 0.004
Identities = 36/172 (20%), Positives = 82/172 (46%), Gaps = 14/172 (8%)
Query: 521 DDNNELYNIINGYKNTERNILFSTFVTSRSLYNLLVPFQYLTESEIDDNFIYCDTDSLYM 580
+ N EL + ++G K+ I ++ + + ++ ++ ++++ S Y DTDS+++ Sbjct: 817 EKNYELLSYLDGEKDDGFIINSTSIAAATASWSRILMYKHIINSA YTDTDSIFV 870
Query: 581 KSWKPLLNPSLFDPIALGKWDIENEQIDKMFVLNHKKYAYEVNGKIKIASAGIPKNAFD 640
+ KPL + + + K + + 1 + ++ KY + GK++I GI KN + Sbjct: 871 E KPLDSAFIGEGCGKFKAEYNGQLIKRAIFISGKLYLLDFGGKLEIKCKGITKNKDN 927
Query: 641 TSVDFETFVREQFFDG---AIIENNKSIYNEQGTISIYPSKTEIVCGNVYDE 689
T+ + + E ++G + + E GT+++ K ++ G YD+ Sbjct: 928 TTHNLDINDFEALYNGESRVLFQERWGRSLELGTVTVKYQKYNLISG--YDK 977
>gi|461962|sp|P33537|DPOM_NEUCR PROBABLE DNA POLYMERASE
>gi|283351|pir| |S26985 probable DNA-directed DNA polymerase (EC 2.7.7.7) - Neurospora crassa mitochondrion plasmid maranhar (SGC3) >gi|578156|emb|CAA39046| (X55361) putative DNA polymerase [Neurospora crassa] Length = 1021
Score = 44.1 bits (102), Expect = 0.004
Identities = 36/172 (20%), Positives = 82/172 (46%), Gaps = 14/172 (8%)
Query: 521 DDNNELYNIINGYKNTERNILFSTFVTSRSLYNLLVPFQYLTESEIDDNFIYCDTDSLYM 580
+ N EL + ++G K+ I ++ + + ++ ++ ++++ S Y DTDS+++ Sbjct: 815 EKNYELLSYLDGEKDDGFIINSTSIAAATASWSRILMYKHIINSA YTDTDSIFV 868
Query: 581 KSWKPLLNPSLFDPIALGKWDIENEQIDKMFVLNHKKYAYEVNGKIKIASAGIPKNAFD 640
+ KPL + + + K + + 1 + ++ K Y + GK++I GI KN + Sbjct: 869 E KPLDSAFIGEGCGKFKAEYNGQLIKRAIFISGKLYLLDFGGKLEIKCKGITKNKDN 925
Query: 641 TSVDFETFVREQFFDG---AIIENNKSIYNEQGTISIYPSKTEIVCGNVYDE 689
T+ + + E ++G + + E GT+++ K ++ G YD+ Sbjct: 926 TTHNLDINDFEALYNGESRVLFQERWGRSLELGTVTVKYQKYNLISG--YDK 975
>gi|249951l|sp|Q12471|6P22_YEAST 6-PHOSPHOFRUCTO-2-KINASE 2 (PHOSPHOFRUCTOKINASE 2 II) (6PF-2-K 2) >gi| 2131162 |pir| |S61066 6-phosphofructo-2-kinase (EC 2.7.1.105) - yeast (Saccharomyces cerevisiae) >gi|2131163 |pir| |S71026 6-phosphofructo-2-kinase (EC 2.7.1.105) - yeast (Saccharomyces cerevisiae) >gi|l085116|emb|CAA6237l| (X90861)
6-phospho ucto-2-kinase [Saccharomyces cerevisiae] >gi|l420028|emb|CAA99157| (Z74878) ORF YOL136C [Saccharomyces cerevisiae] >gi | 1628439 | emb|CAA64733 | (X95465) 6-phosphofructo-2-kinase [Saccharomyces cerevisiae] Length = 397
Score = 40.6 bits (93), Expect = 0.041
Identities = 48/208 (23%) , Positives = 92/208 (44%) , Gaps = 29/208 (13%)
Query: 175 MKTNTSIATLGKKLLDGGYLTESQLKTDFNYTIFDKDNDMNDSEAYDYAVKCFAKLTPEQ 234
++ S AT+ K LL L+ + + FN K+ND ++ +A++T ++
Sbjct: 139 IRRQISCATISKPLL LSNTSSEDLFN PKNNDKKET YARITLQK 181
Query: 235 LTY-IHNDVIILGMCHIHYSDIFPNFDYNKLTFSLNIMESYLNNEMTRFQLLN---QYQD 290
L + I+ND +G+ S I + F + S+ +E++ F L+ Q Sbjct: 182 LFHEINNDECDVGIFDATNSTI ERRRFIFEEVCSFNTDELSSFNLVPIILQVSC 235
Query: 291 IKISYTHYHFHD.røFY-DYIKSFYRGG__JMYNTKYINKLIDEPCFSID-INSSYPYVMYH 348
S+ Y+ H+ +F DY+ Y + + + + FS+D N + Y+ H Sbjct: 236 FNRSFIKYNIHNKSFNEDYLDKPYELAIKDFAKRLKHYYSQFTPFSLDEFNQIHRYISQH 295
Query: 349 EKIPTWLYFYEHYSEPTLIPTFLDDDNY 376
E+I T L+F+ + + P L+ +Y Sbjct: 296 EEIDTSLFFFNVINAGWEPHSLNQSHY 323
>gi|2258375|gb|AAD11909.l| (AF007261) transcription initiation factor sigma [Reclinomonas americana] Length = 532
Score = 39.9 bits (91), Expect = 0.070
Identities = 49/205 (23%) , Positives = 84/205 (40%) , Gaps = 14/205 (6%)
Query: 100 NHFLLKDTMRYFDNITRENIYLKSAEENEHTLKMKEATILAKNQNVIL EKRVKSΞIN 156
N+ + + F + ++IY+ + +KE L K NVI+ K +K N Sbjct: 177 NYLVKNSYI__LFKTVPHDSIYMNYSYIQTPLNILKEYLQLIKIINVIILQINKNIKKKNN 236
Query: 157 LDLTMFLNGFKFNIIDNFM---KTNTSIATLGKKLLDGGYLTESQLKTDFNYTIFDKDND 213
L++++FL F + N++ K + + + K L Y+T L T Y K Sbjct: 237 LNISLFLYKFYQELKWNYIFINKISRNTQKINIKTLKNSYITFYNLITFIQYYTTKKQRL 296
Query: 214 MNDSEAYDYAVKCFAK--LTPEQLTYIHNDVIILGMCHIHYΞDIFPNFDYN-KLTFSLNI 270
D +K F K P+ +N +1 G+ HI+ + N K+T I Sbjct: 297 KKDIFYKQIFIKTFLKQHKIPKINKIKNNSLIKYGLTHIYDMILISILRENIKVTLKNRI 356
Query: 271 MESYLNNEMTRFQLLNQYQDIKISY 295
+ +Y+ T + QY +KI Y Sbjct: 357 IFNYMPYITT---ISKQY--VKIGY 376
>gi|l5734|emb|CAA37450| (X53370) DNA polymerase (AA 1-575) [Bacteriophage phi-29] Length = 575
Score = 39.5 bits (90), Expect = 0.092
Identities = 41/150 (27%) , Positives = 64/150 (42%) , Gaps = 36/150 (24%)
Query: 497 LSKWLNGLYG IPALRSHFNL-FRLDDNNELYNIINGYKNTERNIL--F 542
L+K++LN LYG +P L+ + L FRL G + T+ +
Sbjct: 381 LAKLMLNSLYGKFASNPDVTGKVPYLKENGALGFR GEEETKDPVYTPM 429
Query: 543 STFVTSRSLYNLLVPFQYLTESEIDDNFIYCDTDSLYMKSWKPLLNPSLFDPIALGKWD 602
F+T+ + Y + Q D IYCDTDS+++ P + + DP LG W Sbjct: 430 GVFITAWARYTTITAAQACY DRIIYCDTDSIHLTGTEIPDVIKDIVDPKKLGYWA 484
Query: 603 IENEQIDKMFVLNHKKYAY EVNGKI 627
E+ ++ L K Y EV+GK+ Sbjct: 485 HES-TFKRVKYLRQKTYIQDIYMKEVDGKL 513
Query= pt| 110872 44AHJDORF002 Phage 44AHJD ORF | 3789-5732 | 3 1 (647 letters)
>gi|l35273|sp|P27622|TAGC_BACSU TEICHOIC ACID BIOSYNTHESIS PROTEIN C >gi|478126 |pir| |D49757 techoic acid biosynthesis protein tagC - Bacillus subtilis (strain 168) >gi| 143727 (M57497) putative [Bacillus subtilis] >gi|2636103|emb|CAB15594.1| (Z99122) alternate gene name: dinC [Bacillus subtilis] Length = 442
Score = 112 bits (278), Expect = 7e-24
Identities = 91/314 (28%) , Positives = 147/314 (45%) , Gaps = 58/314 (18%)
Query: 152 FELNELEPKFVMGFGGIRNAVNQSINIDKETNHMYSTQSDS QKPEGFWINKLTPSG 207
F+ + PK V QS N D++ + +Y+TQ S + + I +L+ G
Sbjct: 7 FDFTNITPKLFTELRVADKTVLQSFNFDEKNHQIYTTQVASGLGKDNTQSYRITRLSLEG 66
Query: 208 DLISSMRIVQGGHGTTIGLERQSNGEMKIWLHHD GVAKLLQVAYKDNYVLDLEEA 262
+ SM + GGHGT IG+E + NG + IW +D ++L+ YK LD E + Sbjct: 67 LQLDSMLLKHGGHGTNIGIENR-NGTIYIWSLYDKPNETDKΞELVCFPYKAGATLD-ENS 124
Query: 263 KGLTDYTPQSLLNKHTFTPLIDEANDKLILRFGDGTIQVRSRADVKNHIDNVEKEMTIDN 322
K L ++ H TP +D N +L +R + D KN+ N ++ +TI N
Sbjct: 125 KELQRFSNMPF--DHRVTPALDMKNRQLAIR QYDTKNN- -NNKQWVTIFN 170
Query: 323 SE NNDN RWMQGIAVDGDDLYWLSGNSSVNSHVQIGKYSLTTGQKI 367
+ N +N ++QG +D LYW +G+++ S+ + +
Sbjct: 171 LDDAIANKNNPLYTINIPDELHYLQGFFLDDGYLYWYTGDTNSKSYPNL ITV 222
Query: 368 YDYPFKLSYQDGINFPRD NFKEPEGICIYTNPKTKRKSLLLAMTNGGGGKRFH 420
+D K+ Q I +D NF+EPEGIC+YTNP+T KSL++ +T+G G R Sbjct: 223 FDSDNKIVLQKEITVGKDLSTRYENNFREPEGICMYTNPETGAKSLMVGITSGKEGNRIS 282
Query: 421 NLYGFFQLGEYEHF 434
+Y + YE+F Sbjct: 283 RIYAYH SYENF 293
>gi 1142847 (M64050) DNase inhibitor [Bacillus subtilis] Length = 125
Score = 51.9 bits (122), Expect = le-05
Identities = 35/116 (30%) , Positives = 55/116 (47%) , Gaps = 10/116 (8%)
Query: 152 FELNELEPKFVMGFGGIRNAVNQSINIDKETNHMYSTQSDS QKPEGFWINKLTPSG 207
F+ + PK V QS N D++ + +Y+TQ S + + I +L+ G
Sbjct: 7 FDFTNITPKLFTELRVADKTVLQSFNFDEKNHQIYTTQVASGLGKDNTQSYRITRLSLEG 66
Query: 208 DLISSMRIVQGGHGTTIGLERQSNGEMKIWLHHD GVAKLLQVAYKDNYVLD 258
+ SM + GGHGT IG+E + NG + IW +D ++L+ YK LD Sbjct: 67 LQLDSMLLKHGGHGTNIGMENR-NGTIYIWSLYDKPNETDKSELVCFPYKAGATLD 121
>gi 14038407 (AF103943) factor C protein precursor [Streptomyces griseus] Length = 324
Score = 39.1 bits (89), Expect = 0.10
Identities = 61/269 (22%) , Positives = 102/269 (37%) , Gaps = 33/269 (12%)
Query: 172 VNQSINIDKETNHMYSTQSDSQKPEG FWINKLTPSGDLISSMRIVQGGHGTTIGLER 228
V QS D ++ Q S P+ I +L SG+ + M ++ GHG +IG + Sbjct: 66 VQQSFTFDIVNRRLFVAQLKΞGSPDDSGDLCITQLDFSGNKLGHMYLLGFGHGVSIGAQ- 124
Query: 229 QSNGEMKIWLHHDGVAKLLQVAYKDNYVLDLEEAKGLTDYTPQSLLNKHTFTP 281
+ + D + + + + G T S L KH P Sbjct: 125 PVGADTYLWTEVD VNSNARGTRLARFKWNNGATLSRTSSALAKHQPVPGATEMTC 179
Query: 282 LIDEANDKLILRFGDGTIQVRSRADVKNHIDNVEKEMTIDNSENNDNRWMQGIAVDGDDL 341
ID N+++ +R+ + + +V + V + D QG A+ G +
Sbjct: 180 AIDPVNNRMAIRYLTASGRRYGIYNVADIAAGVYDKPLSDVPHPTGLGTFQGYALYGSYV 239
Query: 342 YWLSGN SSVNSHVQIGKYSLTTGQKIYDYPFKLSYQDGINFPRDNFKEPEGIC 394
Y L+GN + NS+V + TG + + + G F+EPEG+ Sbjct: 240 YQLTGNPYGPDNPNPGNSYVS--SVDVNTGALVQ RAFTRAGSTL---TFREPEGMG 290
Query: 395 IYTNPKTKRKSLLLAMTNGGGGKRFHNLY 423
IY + + L L +G G R NL+ Sbjct: 291 IYRTAAGEVR-LFLGFASGVAGDRRSNLF 318
Query= pt| 110873 44AHJDORF003 Phage 44AHJD ORF | 6626-8389 | 2 1 (587 letters)
>gi| 138123 |sp|P0433l|VG9_BPPH2 TAIL PROTEIN (LATE PROTEIN GP9) >gi| 75850 jpir| |WMBPT9 gene 9 protein - phage phi-29 >gij 215327 (M14782) tail protein [Bacteriophage phi-29] >giJ225364 |prf I |1301270D gene 9 [Bacillus sp.] Length = 599
Score = 92.4 bits (226), Expect = 8e-18
Identities = 126/618 (20%) , Positives = 251/618 (40%) , Gaps = 71/618 (11%)
Query: 5 TNFKFFYNTPFT-DYQNTIHFNSNKERDDYFLNGRHFKSLDYSKQPY-NFIRDRMEINVD 62
TN + + PF+ DY+NT F S+ + ++F R + + SK + F ++ ++V Sbjct: 9 TNVRILADVPFSNDYKNTRWFTSSSNQYNWF--NRKSRVYEMSKVTFMGFRENKPYVSVS 66
Query: 63 MQWHDAQGINYMTFLS-DFEDRRYYAFVNQIEYVNDVWKIYFVIDTIMTYTQGNVLEQL 121
+ +Y+ F + D+ ++ +YAFV ++E+ N V ++F ID + T+ ++ Sbjct: 67 LPIDKLYSASYIMFQNADYGNKWFYAFVTELEFKNSAVTYVHFEIDVLQTWMFDMKFQES 126
Query: 122 SNVNIERQHLSKRTYNYMLPMLRNNDDVLKVSNKNYVYNQMQQYLENLVLFQSSADLSKK 181
I R+H+ K + P + D+ L ++ + + + ++F S Sbjct: 127 F---IVREHV-KLWNDDGTPTINTIDEGLSYGSEYDIVSVENHKPYDDMMFLVIISKSIM 182
Query: 182 FGT--KKEPNLDTSKGTIYDNITSPVNLYVMEYGDFINFMDKMSAYPWITQNFQK V 235
GT ++E L+ ++ + + P+ Y+ + + D +1 N V Sbjct: 183 HGTPGEEESRLNDINASL-NGMPQPLCYYIHPF YKDGKVPKTYIGDNNANLSPIV 236
Query: 236 QMLPKDFINTKDLEDVKTSEKITGLKTLKQGGKSKEWSLK-DLSL SFSNLQ 285
ML F + D+ + +T LK K+ + LK D + N+
Sbjct: 237 NMLTNIFSQKSAVNDI-VNMYVTDYIGLKLDYKNGDKELKLDKDMFEQAGIADDKHGNVD 295
Query: 286 EMMLSK ---KDEFKHMIRNEYMTIEFYDWNGNTMLLDAGKISQK 326
+ + K KD+ ++ Y E D+ GN M L 1+
Sbjct: 296 TIFVKKIPDYEALEIDTGDKWGGFTKDQESKLMMYPYCVTEITDFKGNHMNLKTEYINNS 355
Query: 327 TGVKLRTKSIIGYHNEVRVYPVDYNSAENDRPILAKNKEILIDTGSFLNTNITFNSFAQV 386
+K++ + +G N+V DYN+ D + N+ S +N N Sbjct: 356 K-LKIQVRGSLGVSNKVAYSVQDYNA---DSALSGGNRLTASLDSSLINNNPN 404
Query: 387 PILINNGILGQSQQANRQ--KNAESQLITNRIDNVLNG---SDPKSRFYDAVSVASNLSP 441
I I N L Q N+ +N +S ++ N I ++ G + + A+ +AS++ Sbjct: 405 DIAILNDYLSAYLQGNKNSLENQKSSILFNGIMGMIGGGISAGASAAGGSALGMASSV-- 462
Query: 442 TALFGKFNEEYNFYKQQQAEYKDLALQPPSVTESEMGNAFQIANSINGLTMKISVPSPKE 501
T + + QA+ D+A PP +T+ AF N G+ + +
Sbjct: 463 TGMTSTAGNAVLQMQAMQAKQADIANIPPQLTKMGGNTAFDYGNGYRGVYVIKKQLKAEY 522
Query: 502 ITFLQKYYMLFGFEVNDYNSFIEPINSMTVCNYLKCTGTYTIRDIDPMLMEQLKAILESG 561
L ++ +G+++N + + NY++ + DI+ +++++ I ++G Sbjct: 523 RRSLSSFFHKYGYKINRVKK--PNLRTRKAFNYVQTKDCFISGDINNNDLQEIRTIFDNG 580
Query: 562 VRFWHNDGSGNPMLQNPL 579
+ WH D GN ++N L Sbjct: 581 ITLWHTDNIGNYSVENEL 598
>gi|l38124|sp|P07534|VG9_BPPZA TAIL PROTEIN (LATE PROTEIN GP9) >gi|75849|pir I |WMBP9Z gene 9 protein - phage PZA >gij 216058 (M11813) tail protein [Bacteriophage PZA] Length = 599
Score = 81.9 bits (199), Expect = le-14
Identities = 127/618 (20%) , Positives = 248/618 (39%) , Gaps 71/618 (11%)
Query: 5 TNFKFFYNTPFT-DYQNTIHFNΞNKERDDYFLNGRHFKSLDYSKQPYNFIRDRME-INVD 62
TN + + PF+ DY+NT F S+ + ++F + + SK + R+ I+V Sbjct: 9 TNVRILADVPFSNDYKNTRWFTSSSNQYNWF--NSKTRVYEMSKVTFQGFRENKSYISVS 66
Query: 63 MQWHDAQGINYMTFLS-DFEDRRYYAFVNQIEYVNDVWKIYFVIDTIMTYTQGNVLEQL 121
++ +Y+ F + D+ ++ +YAFV ++EY N ++F ID + T+ N+ Q Sbjct: 67 LRLDLLYNASYIMFQNADYGNKWFYAFVTELEYKNVGTTYVHFEIDVLQTW-MFNIKFQE 125
Query: 122 SNVNIERQHLSKRTYNYMLPMLRNNDDVLKVSNKNYVYN--QMQQYLENLVLFQSSADLS 179
S I R+H+ K + P + D+ L ++ + + + Y + + L S + Sbjct: 126 SF--IVREHV-KLWNDDGTPTINTIDEGLNYGSEYDIVSVENHRPYDDMMFLWISKSIM 182
Query: 180 KKFGTKKEPNLDTSKGTIYDNITSPVNLYVMEY GD FINFMDK 221
+ E L+ ++ + + P+ Y+ + GD +N +
Sbjct: 183 HGTAGEAESRLNDINASL-NGMPQPLCYYIHPFYKDGKVPKTFIGDNNANLSPIVNMLTN 241
Query: 222 MSAYPWITQNFQKVQMLPKDFINTK- DLEDVKTSEKITGLKTLKQGGKSKEWS 273
+ + N V M D+I K +L+ K + G+ K G + Sbjct: 242 IFSQKSAVNNI--VNMYVTDYIGLKLDYKNGDKELKLDKDMFEQAGIADDKHGNVDTIFV 299
Query: 274 LKDL---SLSFSNLQEMMLSKKDEFKHMIRNEYMTIEFYDWNGNTMLLDAGKISQKTGVK 330
K +L + KD+ ++ Y E D+ GN M L I +K Sbjct: 300 KKIPDYETLEIDTGDKWGGFTKDQESKLMMYPYCVTEVTDFKGNHMNLKTEYIDNNK-LK 358
Query: 331 LRTKSIIGYHNEVRVYPVDYNSAENDRPILAKNKEILIDTGSFLNTNITFNSFAQVPILI 390 ++ + +G N+V DYN+ + L+ + L+T++ N+ + 1+
Sbjct: 359 IQVRGSLGVSNKVAYSIQDYNAGGS LSGGDRLTAS LDTSLINNNPNDIAII- 409
Query: 391 NNGILGQSQQANRQ--KNAESQLITNRIDNVLNGSDPKSRFYDAVSVASNLSP -- 441
N L Q N+ +N +S ++ N I +L G A + A SP
Sbjct: 410 -NDYLSAYLQGNKNSLENQKSSILFNGIVGMLGGG VSAGASAVGRSPFGLAΞSV 462
Query: 442 TALFGKFNEEYNFYKQQQAEYKDLALQPPSVTESEMGNAFQIANSINGLTMKISVPSPKE 501
T + + QA+ D+A PP +T+ AF N G+ + +
Sbjct: 463 TGMTSTAGNAVLDMQALQAKQADIANIPPQLTKMGGNTAFDYGNGYRGVYVIKKQLKAEY 522
Query: 502 ITFLQKYYMLFGFEVNDYNSFIEPINSMTVCNYLKCTGTYTIRDIDPMLMEQLKAILESG 561
L ++ +G+++N + + NY++ + DI+ +++++ I ++G Sbjct: 523 RRSLSSFFHKYGYKINRVKK--PNLRTRKAYNYIQTKDCFISGDINNNDLQEIRTIFDNG 580
Query: 562 VRFWHNDGSGNPMLQNPL 579
+ WH D GN ++N L Sbjct: 581 ITLWHTDDIGNYSVENEL 598
>gi|l429238|emb|CAA67657| (X99260) tail protein [Bacteriophage B103] Length = 598
Score = 77.6 bits (188), Expect = 2e-13
Identities = 130/623 (20%), Positives = 240/623 (37%), Gaps = 86/623 (13%)
Query: 5 TNFKFFYNTPFT-DYQNTIHFNSNKERDDYFLNGRHFKSLDYSKQPYNFI RDRMEIN 60
T+ + F N PF+ DY++T F + + YF + K + NF+ I Sbjct: 9 TDVRIFSNVPFSNDYKSTRWFTNADAQYSYF NAKPRVHVINECNFVGLKEGTPHIR 64
Query: 61 VDMQWHDAQGINYMTFLS-DFEDRRYYAFVNQIEYVNDVWKIYFVIDTIMTYTQGNVLE 119
V+ + D YM F + + ++ +Y FV ++EYVN V +YF ID I Tt + Sbjct: 65 VNKRIDDLYNACYMIFRNTQYSNKWFYCFVTRLEYVNSGVTNLYFEIDVIQTW-MFDFKF 123
Query: 120 QLSNVNIERQHLSKRTYNYMLPMLRNNDDVLKVSNKNYVYNQMQQYLENLVLFQSSADLS 179
Q S + E Q + P+ D+ L + V Q ++F S
Sbjct: 124 QPSYIVREHQEMWDANNE PLTNTIDEGLNYGTEYDWAVEQYKPYGDLMFMVCISKS 180
Query: 180 KKFGTKKEPNLDTSKGTIYDNITS PVNLYVMEYGDFINFMDKMSAYPWITQNFQKVQ 236
K T E G I NI P++ YV + + D S P +T +VQ Sbjct: 181 KMHATAGET---FKAGEIAANINGAPQPLSYYVHPF YEDGSS- -PKVTIGSNEVQ 230
Query: 237 ML-PKDFINTKDLEDVKTSEKITGLKT LKQGGKSKEWSLKDLSLSFSNL 284
+ P DF+ ++ + ++ T + +K SL+D + + Sbjct: 231 VSKPTDFLKNMFTQEHAVNNIVSLYVTDYIGLNIHYDESAKTMSLRDTMFEHAQIADDKH 290
Query: 285 QEMMLSKKDEFKHMIRNEYMTIEFY DWNGNTMLLDAGK 322
+E + +F NE + Y D+ GN + + Sbjct: 291 PNVNTIYLKEVKEYEEKTIDTGYKFASFANNEQSKLLMYPYCVTTITDFKGNQIDIKNEY 350
Query: 323 ISQKTGVKLRTKSIIGYHNEVRVYPVDYNS AENDRPILAKNKEILIDTGSFLNTNIT 379
++ + +K++ + +G N+V DYN+ D+ + A NT++
Sbjct: 351 VNG-SNLKIQVRGSLGVSNKVTYSVQDYNADTTLSGDQNLTAS CNTSLI 398
Query: 380 FNSFAQVPILINNGILGQSQQANRQ--KNAESQLITNRIDNVLN GSDPKSRFYDAVS 434
N+ V 1+ N L Q N+ +N + ++ N + ++L G+ + AV Sbjct: 399 NNNPNDVAII--NDYLSAYLQGNKNSLENQKDSILFNGVMSMLGNGIGAVGSAATGSAVG 456
Query: 435 VASNLSPTALFGKFNEEYNFYKQQQAEYKDLALQPPSVTESEMGNAFQIANSINGLTMKI 494
VAS S T + + QA+ D+A PP + + A+ N G+ +
Sbjct: 457 VAS--SATGMVSSAGNAVLQIQGMQAKQADIANTPPQLVKMGGNTAYDYGNGYRGVYVIK 514
Query: 495 SVPSPKEITFLQKYYMLFGFEVNDYNSFIEPINSMTVCNYLKCTGTYTIRDIDPMLMEQL 554
+ L + +G++ N + + + NY++ I +++ ++++ Sbjct: 515 KQIKEEYRNILSDFSRKYGYKTNLVK--MPNLRTRESYNYVQTKDCNIIGNLNNEDLQKI 572
Query: 555 KAILESGVRFWHNDGSGNPMLQN 577
+ I +SG+ WH D G+ L N Sbjct: 573 RTIFDSGITLWHADPVGDYTLNN 595
>gi 1215339 (M12456) p9 tail protein [Bacteriophage phi-29]
>gi | 224163 |prf| | 1011232C protein p9,tail [Bacteriophage phi-29]
Length = 335
Score = 71.0 bits (171), Expect = 2e-ll
Identities = 64/293 (21%) , Positives = 123/293 (41%) , Gaps = 20/293 (6%)
Query: 292 KDEFKHMIRNEYMTIEFYDWNGNTMLLDAGKISQKTGVKLRTKSIIGYHNEVRVYPVDYN 351
KD+ ++ Y E D+ GN M L 1+ +K++ + +G N+V DYN Sbjct: 57 KDQESKLMMYPYCVTEITDFKGNHMNLKTEYINNSK-LKIQVRGSLGVSNKVAYSVQDYN 115
Query: 352 SAENDRPILAKNKEILIDTGSFLNTNITFNSFAQVPILINNGILGQSQQANRQ--KNAES 409
+ D + N+ S +N N I I N L Q N+ +N +S
Sbjct: 116 A DSALSGGNRLTASLDSSLINNNPN DIAILNDYLSAYLQGNKNSLENQKS 165
Query: 410 QLITNRIDNVLNG---SDPKSRFYDAVSVASNLSPTALFGKFNEEYNFYKQQQAEYKDLA 466
++ N I ++ G + + A+ +AS++ T + + QA+ D+A
Sbjct: 166 SILFNGIMGMIGGGISAGASAAGGSALGMASSV--TGMTSTAGNAVLQMQAMQAKQADIA 223
Query: 467 LQPPSVTESEMGNAFQIANSINGLTMKISVPSPKEITFLQKYYMLFGFEVNDYNSFIEPI 526
PP +T+ AF N G+ + + L ++ +G+++N + Sbjct: 224 NIPPQLTKMGGNTAFDYGNGYRGVYVIKKQLKAEYRRSLSSFFHKYGYKINRVKK--PNL 281
Query: 527 NSMTVCNYLKCTGTYTIRDIDPMLMEQLKAILESGVRFWHNDGSGNPMLQNPL 579
+ NY++ + DI+ +++++ I ++G+ WH D GN ++N L Sbjct: 282 RTRKAFNYVQTKDCFISGDINNNDLQEIRTIFDNGITLWHTDNIGNYSVENEL 334
>gi|ll81968|emb|CAA87738.l| (Z47794) tail protein [Bacteriophage CP-1] Length = 230
Score = 53.9 bits (127), Expect = 3e-06
Identities = 29/113 (25%) , Positives = 54/113 (47%) , Gaps = 3/113 (2%)
Query: 1 MRKLTNFKFFYNTPF-TDYQNTIHFNSNKERDDYFLNGRHFKSLDYSKQPYNFIRDRMEI 59
M++ T + +PF DY N I+F + + +D+F + Y + + + I Sbjct: 1 MQESTKIWLYAKSPFKNDYANVINFETRESMEDFFTKKNPHIEIVYEYDKFQYTQRNGSI 60
Query: 60 NVDMQWHDAQGINYMTFLSDFEDRRYYAFVNQIEYVNDVWKIYFVIDTIMTY 112
V + + + YM F+++ R YYAFV + Y+N+ +1 + +D TY Sbjct: 61 WSGRVEKYENVTYMRFINN--GRTYYAFVFDVLYINEDATRIIYEVDVWNTY 111
>gi|ll81970|emb|CAA87740.l| (Z47794) tail protein [Bacteriophage CP-1] Length = 586
Score = 42.2 bits (97), Expect = 0.010
Identities = 79/381 (20%) , Positives = 139/381 (35%) , Gaps = 92/381 (24%)
Query: 277 LSLSFSNLQEMMLSK--KDEFK---HMIR_ΪEYMTIEFYDWNGNTMLLDAG KISQKT 327
L +++ +QE + S KD+ + ++ +E+ IE YD GN+ + I +
Sbjct: 187 LKIAYDQIQEGLRSYMGKDDLEIEVQLLNSEFTEIELYDIYGNSYVYQPQYLPRTIDEAH 246
Query: 328 GVKLRTKSIIGYHNEVRVYPVDYNSAEN DRPIL 360
K+ +G N+V + ++YN+A N D+ IL Sbjct: 247 KYKVIVSGSLGDSNQVHINFLEYNNANNVSYADKNILDSLESGDWAEHNPEHFKYGLNDV 306
Query: 361 -AKNKEILIDT-GSFLNTNITFNSFAQVPILINNGILGQSQQANRQKNAESQLITNRIDN 418
K+ IL D S++ ++ Q+ N +L QS + ++ A + + Sbjct: 307 TGKSVAILNDAEASYIQSHKNQMEHTQLTFKENRDMLKQSVDLSNKQVATANSQASYNAQ 366
Query: 419 VLNGSDPKSRFYDAVSVASNLSPTALFGKF NEEYNFYKQQQ-- 459
S +++ + S N++ L G F N +YN QQ
Sbjct: 367 FAVDSANINQWTEGASGILNVAGNLLTGNFGGALGGLASGGMKVFNANRDYNDKWQQGF 426
Query: 460 AEYKDLALQPPSVTESEMGNAFQIANSIN 488
A DL QP SV + AFQ N + Sbjct: 427 TSENNALKSQSNALANMKSKIALDQSIRAYNATMADLQNQPISVQQIGNDLAFQSGNRLT 486
Query: 489 GLTMKISVPSPKEITFLQKYYMLFGFEVNDY-NSFIEPINSMTVCNYLKCTGTY--TIRD 545
+ K+S+ + + +Y +G VN + N + + S NY+K T+R Sbjct: 487 DVYWKVSLAQKEIMGRANEYIKCYGVLVNWFTNDALSVMRSRKRFNYIKMINVNLGTLR- 545
Query: 546 IDPMLMEQLKAILESGVRFWH 566
+ M ++AI +SGVR W+ Sbjct: 546 ANQSHMNAIQAIFQSGVRIWN 566
Query= pt| 110875 44AHJDORF005 Phage 44AHJD ORF | 12643-13890 | -1 1 (415 letters)
>gi 13845203 (AE001399) GAF domain protein (cyclic nt signal transduct.) [Plasmodium falciparum] Length = 1245
Score = 52.3 bits (123), Expect = 6e-06
Identities = 59/246 (23%) , Positives = 105/246 (41%) , Gaps = 27/246 (10%)
Query: 174 ESIDRNHGNVDYIGFPKMFLLGNAVNFSSPILSNLNIYNLLQKHKMNTSRLYKNIFLEMR 233
+S D N+ N + + N+V FS+ N IY++L N +YK + E+ Sbjct: 854 DSSDNNOTTONNN-INNNNNYNNNNSVIFS NEKIYDML NRDNIYKKVKKEIF 904
Query: 234 RNDYVNEKRNTRAFNSNDDAMTTGEFEFNEYNLADDNLRNHINQNGDFFYIKTDDKYI-- 291
D + + + +N + M + N N ++N+ N+ N NGD Y KY Sbjct: 905 EGDSIIKT ENKPNLTNKNY^INNDNIDNNNNN N NID NNNNGDNIYNDDLKKYY N 964
Query: 292 KVMYNVTTFMTNIIVVPYTKQYEFCTKIR-DIDNHVTYLRDDMFYKENMERYYYNPSNLH 350
++N ++ + + + K E K+ I + L +F+K NM + + L+ Sbjct: 965 TSIFNKDLYVKHFVDIIMNKSLEEIIKMNVYISERINSL LFHKGNM LNDVTKLY 1018
Query: 351 FDNAYSKNYWDNDRYLYLDMNKIIKFHIKNEMKKNMSEFERKEKIYEDN YIENTK 406
NAY + N K I F + E K + F+ +KIY+ N + N K Sbjct: 1019 MSNAYGEKCFFFN FPQIKEIIFVNEYEKKMDMKYFKMLKKIYKYNLNKIFSNNYK 1073
Query: 407 KYLMKQ 412
+++K+ Sbjct: 1074 FFIIKK 1079
>gi|3758843|emb|CAB11128.l| (Z98551) predicted using hexExon;
MAL3P6.23 (PFC0820w) , Hypothetical protein, len: 4982 aa [Plasmodium falciparum] Length = 4981
Score = 49.2 bits (115), Expect = 5e-05
Identities = 67/287 (23%) , Positives = 110/287 (37%) , Gaps = 60/287 (20%)
Query: 127 ITDLNSATDLKYHSNFLKHYPIIIYDEFLALEDDYLIDEWDKLKT IYESIDRNHGN 182
I D+N + D+ + +++ I YD +++DK++ IY +ID++ N
Sbjct: 3619 IMDINKSKDISKNMEIVQN---IEYD NKYDKIRNDMDAIYMAIDKDMDN 3664
Query: 183 VDYIGFPKMFLLGNAVNFSSPILSNLNIYNL LQKHKMNTSRLYKNIFLEMRRNDYV 238
+ 1 + F L N S +N YNL ++ K N R Y N F +D Sbjct: 3665 IGIINCMRYFNLYKNYNNLSNECNNRE-YNLNELYMEDIKRNMKR-YDNNFNINHYDDNN 3722
Query: 239 NEKRNTRAFNS-JDDAMTTGEFEFNEYNI_M.DNLR_mNQNGDFFYIKTDDKYIKVMY-IVT 298
N N N+N++ N N ++N N+ N NG F+ D Sbjct: 3723 NNNNNNNNNNNNNNNNNNNNNNNNNNNNNNNNNNNNNNNNGGCFFFHVD 3771
Query: 299 TFMTNIIWPYTKQYEFCTKIRDIDNHVTYLRDDMFYKENMERYYYNPSNLHFDNAYSKN 358
K FCTK ++F +N+E N N N Y+ N
Sbjct: 3772 -- -KDLFFCTK KNIFPCKNIETVCKNEYNKKIYNNYTCN 3807
Query: 359 YWDNDRYLYLDMNKIIKFHIKNEMKKNMSEFERKEK-IYEDNYIEN 404
V+N + ++IK + + N E+ + EK +Y + EN Sbjct: 3808 ISVNNTLNCLNIIKELIKLNNNKKKILNYYEYHKVEKLLYYRHSFEN 3854
Score = 35.6 bits (80), Expect = 0.70
Identities = 62/290 (21%), Positives = 121/290 (41%), Gaps = 65/290 (22%)
Query: 2 VKQNRLDMVRDYQNAVN--HVRKKIPDKYNQIELVDELMNDDIDYYISISNRSDGKΞFNY 59
+K+N ++ +N +N +V++ DK N I D++I+ SN + +SF
Sbjct: 4445 IKRNNINKSNIKRNNINKSNVKRSNTDKSNVIS DFHIT-SNNNITRSFT- 4492
Query: 60 VSFFIYLAIKLDIKFTLLSRHYTLRDAYRDFIEEIIDENPLFKSKRVTFRSARDYLAIIY 119
A D F LS TL +Y +F + + I
Sbjct: 4493 ATLTDSIFNTLΞE--TLNYSYDNFFSNMDN IKI 4523
Query: 120 QDKEIGVITDLNSATDLKYHSNFLKHYPIIIYDEFL ALEDDYLIDEWDKLKTIYE 174
+ El ITD++ +YH N+LK + +E++ + +D + DE ++T+ E Sbjct: 4524 KKNEINNITDVDYGNKKEYHENYLKVKQNKVNEEYIEETFKΞDKDCSIKDEACTIRTLSE 4583
Query: 175 S--IDRNHGNVDYIGFPKMFLLGNAVNFSSPILSNLNIYNLLQKHKMN--TSRLYKNIFL 230
S I N N+D + + + S P N++ N ++K+ +N R+ KN
Sbjct: 4584 SCNISENISNID MDDEDHISFPNGRNVHDNNYMKKNHVNYDKMRVGKNKIP 4634
Query: 231 EMRRNDYVNEKRNTRAFNSNDDAMTTGEFEFNEYNLADDNLRNHINQNGD 280
D + +++ + +D M++ ++ E ++ + L + NG+ Sbjct: 4635 SFTHFDKILDEKKKK SDKDMSSSKWLEREEHIKEIKLEKNEYMNGN 4680
Score = 34.0 bits (76), Expect = 2.0
Identities = 47/211 (22%), Positives = 84/211 (39%), Gaps = 32/211 (15%)
Query: 210 IYNLLQKHKMNTSRLYKNIFLEMRRNDYVNEKRNTRAFNSNDDAMTTGEFEFNEYNLADD 269
I++LLQK LY+N+ + R + N+ T E ++ + ++
Sbjct: 918 IFSLLQKDSSPLLVLYENVHI REGEKYGRNE--ATDNEVDYKKGDIIKH 964
Query: 270 NLRNHINQNGDFFYIKTD---DKYIKVMYNVTTFMTNIIWPYTKQYEFCTKIRDIDNHV 326
N+ N + D + D+ K MY + V E K D+ N+ Sbjct: 965 NVTNEHGNHSDSYPYGNSLNLDRKPKNMYE-DIYKEKGFVKSDCSNIEI--KKNDMINND 1021
Query: 327 TYLRDDMFYKENMERYYYNPSNLHFDNAYSKNYWDNDRYLYLDMNKII KFHIKNE 382
Y +++ FY+++ Y+ + YV++ +YL +N ++ F +KN+ Sbjct: 1022 VYKKNE-FYEDSRINMIYDEDEIKTWFLIPHKYVIN IIYLFLNILLTDESNFKLKNK 1077
Query: 383 MKKNMSEFERKEKIYEDN YIENTKKY 408
E K IYEDN ++N KKY Sbjct: 1078 KYGYFVNEETKGTIYEDNNGLQEILKNGKKY 1108
Score = 33.6 bits (75), Expect = 2.7
Identities = 42/198 (21%) , Positives = 77/198 (38%) , Gaps = 42/198 (21%)
Query: 222 SRLYKNIFLEMR RNDYVNEKRNTRAF- NSNDDAMTTGEFEFNEYNLA 267
S LY I++ + +N + K+NT + N+++D TT E + +
Sbjct: 411 SVLYSIIYMNKKYKKKNFIITNKKNTNVYFENDVIQLSVENTSEDTFTTNTRESSLNSGM 470
Query: 268 DDNLR_raiNQNGDFFYIKTDDKYIKVMYNVTTFMTNIIVVPYTKQYEFCTKIRDIDNHVT 327
+++R +N D +DDK ++Y N YTK E Sbjct: 471 MNDMRYSVNNYADEKVYHSDDKSDHLIYKHVHDEKNKYDEMYTKTKE 517
Query: 328 YLRDDMFYKE.MERYYYNPSN___FDNAYSKNYVVDNDRYLYLDMNKIIKFHIKNEMKKNM 387
+++ YK N+ + N K LD+ K I H+KN+ + N
Sbjct: 518 --NENIIYKSNIVDKKTCDISSEMVNGKDK LDVEKYIGSHVKND-ENNK 563
Query: 388 SEFERK-EKIYEDNYIEN 404
+ ++K + + + YI+N Sbjct: 564 EKLKKKIDNVNKKEYIDN 581
>gi 13845297 (AE001421) hypothetical protein [Plasmodium falciparum] Length = 2380
Score = 48.0 bits (112), Expect = le-04
Identities = 87/390 (22%) , Positives = 160/390 (40%) , Gaps = 65/390 (16%)
Query: 20 VRKKIPDKYNQIELVDELMNDDIDYYISISNRSDGKSFNYVSFF IYLAIKLDIKF 74
+++K +K ++ + +N D + ++ R K+ NY++ +YL I DI Sbjct: 1049 LQRiαmNKCSKNRNRNRYINKDSNIHL.1NLIRIKFKNL_r_MNMNSFEIELYLKINNDIFL 1108
Query: 75 TLLSRHYTLRDAYR DFIEEIIDEN-PLFKSKRVTFRSARDYLAIIYQDKEIGVI 127
+Y +++ Y + + + EN + +++ ++ + Y +K+ Sbjct: 1109 QFNKHNYNVQNFYNFSITLINIMSKYYSENFYAYNLEKIVYKFLLNNKNFEYIEKQYSSK 1168
Query: 128 TDLNSATDLKYHSNFLKHYPIIIYDEFLA LEDDYLIDEWDKLKTIYESIDRNHGNV 183
D+N D+ ++ +K+ II EFL L+ D I + KLKT ++
Sbjct: 1169 EDMNEL-DILVNTYDMKYDKII---EFLKNNGYLKIDRYIYFYPKLKT DI 1214
Query: 184 DYIGFPKMFLLGNAVNFSSPILSNLNIYNLLQKHKMNTSRLY KNIF--LEMRRN 235
F ++FL N + L NI +++ K + Y K IF + M+ + Sbjct: 1215 ILFFFKEIFLNDNILKIDRKFLKK-NITIMIEVLKEIFFKEYVKRCITKVIFFPVHMKEH 1273
Query: 236 DYVNEKR --NTRAFNSNDDAMTTGEFEFNEYNLADDNLRNHINQNGDFFYIKTD 287
D+V K N+ FN+ D + N YN D+ N+ N N +Y K
Sbjct: 1274 DHVMNK-T-YNNQYVNNSNMFNTRGDHNNNNQTNDNHYNHHYDDTHNNNNNNNSKYY-KNK 1332
Query: 288 DKYIKVMYNVTTFMTNIIV VPYTKQYEFCTKIRDIDNHVTYLRDDMFYKEN ME 340
+K K+MY +++ + V K + K I + Y+ ++ N + Sbjct: 1333 NKN-KIMYEKERKSSSLFISNNVQDVKPIKHYLKYSSIYKNFIYIISEIKNFNNKITKIN 1391
Query: 341 RY-YYNPSNLHFDNAYSKNYWDNDRYLYL 369
RY YYN NL+ D+ ND YL+L
Sbjct: 1392 RYNYYNYMNLNIDDL NDAYLFL 1413
Score = 32.5 bits (72), Expect = 6.0
Identities = 46/183 (25%) , Positives = 73/183 (39%) , Gaps = 26/183 (14%)
Query: 225 YKNIFLEMRRNDYVNEKRNTRAFNSNDDAMTTGEFEFNEYNLADDNLRNHINQNGDFFYI 284
+KNI ++ ++N + NSN + + N N+ +N N IN + I Sbjct: 27 HKNINKNIKNKKFINIDNSNNCNNSNSNNSNSNNNNNNNNNIVRNN-NNFINADKKKNVI 85
Query: 285 KTDDKYIKVMYNVTTFMTNIIVVPYTKQYEFCTKIRDIDNHVTYLRDDMFYKENMERYYY 344
+D IK V NI Y ++ + D+ N+ + + KE ER Sbjct: 86 I_.EDDDIKNKELVDESFVNIFF--YENYFKNLFNLNDVSNNKVI--NIIEQKEGDER 138
Query: 345 NPSNLHFDNAYΞKNYVVDNDRYLYLDMNKIIKFHIKNEMKKNMSEFERKEKIYEDNYIEN 404
N N N +KN V DN +NK IKN +N++E Y N++ + Sbjct: 139 NADN NLKNKNIVRDN INK IKN--TRNVNEILIYNNKYIINFLND 180
Query: 405 TKK 407
T K Sbjct: 181 TTK 183
>gi|4493936|emb|CAB38972.l| (AL034556) predicted using hexExon;
MAL3P5.6 (PFC0600W) , Hypothetical protein, len: 250 aa [Plasmodium falciparum] Length = 249
Score = 47.3 bits (110), Expect = 2e-04
Identities = 53/215 (24%) , Positives = 87/215 (39%) , Gaps = 30/215 (13%)
Query: 209 NIYNLLQKHKMNTSRLYKNIFLEMRRNDYVNEKRNTRAFNSNDDAMTTGEFEF--NEYNL 266
NIYN L++ YKN N ++ +N N+N EFE N YN Sbjct: 13 NIYNKLEEK YKNFLKLKNMNSHMGASQNMNV-NNNYTMNELEEFEKINNNYNN 64
Query: 267 ADDNLRNHINQNGDFFYIKTD DKYIKVMYNVTTFMTNIIWPYTKQYEFCTKIRD 321
++N+ N+IN D+ IK +K ++ YN + 1 T +++ Sbjct: 65 NNNNINNNINNYYDYMNIKVSQSVQHNKRLQDFYNNKNSFQHYIKKLKTCRFDADDIRNL 124
Query: 322 IDNHVTYLRDDMFYK ENMERYYYNPSNLHFDNAYSKNYWDNDRYLYLDMNKIIK 376
++ + Y RD+ K EN + N + N+ S NY DN+ LY +N++ K Sbjct: 125 LEKRLAYERDNTLIKNIQEEENKKGIGINGNFGSESNSSSSNY--DNNYLLYRKINRLNK 182
Query: 377 FHIKNEMKKNMSEFERKEKIYEDNYIENTKKYLMK 411
+ ++ KI KKY++K
Sbjct: 183 TNTNKSKNRSRKRKRINSKI DKKYIIK 209
>gi 13845165 (AE001390) hypothetical protein [Plasmodium falciparum] Length = 1247
Score = 45.7 bits (106), Expect = 6e-04
Identities = 52/239 (21%) , Positives = 94/239 (38%) , Gaps = 38/239 (15%)
Query: 206 SNLNIYNLLQKHKMNTSRLYKNIFLEMRRNDYVNEKRNTRAFNSNDDAMTTGEFEFNEYN 265
+N N +N ++K K R I +N + +N ++N+D E N N Sbjct: 474 NNTNKVMEIKKRKKKFKREKNKIINNSFQNQEAEDDKNNNNNDNNNDNHNDNNNENNNEN 533
Query: 266 LADDNLRNHINQNGDFFYI-KTDDKYIK VMYNVTTFMTNIIWPYTKQYEFCTKIR 320
D+N N+ + N D I D+ Y +YN T ++ YTK + + + Sbjct: 534 NNDNNNENNNDINNDINNIHNNDNNYYNNDNINLYNEMTKKKCMLDNSYTKYFFYIFTL- 592
Query: 321 DIDNHVTYLRDDMFYKENME RYYYN-- PSNLHFDNAYS 356 __
+ + ++ + FY++N + ++YYN + N
Sbjct: 593 ---DMLPSIKFETFYEKNTDHKNFNENYKFYYNTDDDTDIINAIKKKNVKNKKKNGNIVI 649
Query: 357 KNYWDNDRYLYLDMNKIIKFHIKNEMKKNMSEFER KEKIYEDNYIENTKKYLMK 411
KNY+ N+ Y YL+ N+ + I + K +E K+ 1+ ++Y E K K
Sbjct: 650 KNYINHNE-YSYLEYNENKNYEINKKEKLLTENYEYDMYIKDNIHYNDYSEGDGKQTKK 707
Score = 41.0 bits (94), Expect = 0.016
Identities = 58/245 (23%) , Positives = 96/245 (38%) , Gaps = 43/245 (17%)
Query: 207 NLNIYNLLQKHKMNTSRLYKNIFLEMRRNDYVNEKRNTRAFNSNDDAMTTGEFEFNEYNL 266
N+N+YN + K K Y F + D + + + N D E YN Sbjct: 564 NINLYNEMTKKKCMLDNSYTKYFFYIFTLDMLPSIKFETFYEKNTDHKNFNENYKFYYNT 623
Query: 267 ADD NLRNHINQNGDFF---YIKTDDKYIKVMYNVT-TFMTNIIWPYTKQ 312
DD N++N +NG+ YI ++ Y + YN + N T+
Sbjct: 624 DDDTDIINAIKKKNVKNK-KKNGNIVIKNYINHNE-YΞYLEYNENKNYEINKKEKLLTEN 681
Query: 313 YEFCTKIRDIDNHVTYLRDDMFYKENMERYYYNPSNLHFDNAYSK NYV--VD 362
YE+ I+D ++ Y D + + YN +N +N Y K +Y+ VD Sbjct: 682 YEYDMYIKDNIHYNDYSEGDGKQTKKASSFLYNNNN---NNKYKKEDNKTQIISYMDHVD 738
Query: 363 NDR YLYLDMNKIIKFHIK-NEM KKNMSEFERKEKIYEDNYIENTKKY 408
N+ Y + +++ F +K N+M K+ F +E I + +EN K+
Sbjct: 739 NENGVKGLKKRNLFYNNSDQLYNFDVKDNDMIKYEKRQSKNFVEEEFINGNRKMENEDKH 798
Query: 409 LMKQY 413
L K Y Sbjct: 799 LKKHY 803
Query= pt| 110877 44AHJDORF007 Phage 44AHJD ORF | 2044-3027 |l 1 (327 letters)
>gi|1181960|emb|CAA87731.1| (Z47794) connector protein [Bacteriophage CP-1] Length = 337
Score = 45.7 bits (106), Expect = 5e-04
Identities = 44/184 (23%) , Positives = 84/184 (44%) , Gaps = 13/184 (7%)
Query: 127 QIHKLYDNCMSGNFWMQNKPIQYNSDIEIIEHYTDELAEVALSRFSLIMQAKFSK--IF 184
++HK + + +V+ N Y I +E + ++LA++ L+ L A+ + IF Sbjct: 125 ELHKDNPDKIKRPCIVIPNNNF-YEPYIGYLELFCEKLADIELT-IQLNRNAQITPYFIF 182
Query: 185 KSEINDESINQLVSEIYNGAPFVKMSPMFNAD DDIIDLTSNSVIPALTEMKR 236
N S+ + ++I N P V ++ + D D I + L ++
Sbjct: 183 ADNTNVLSMKNIFNKIANFEPWYLNKQKDQDGQDSFKQLSDYIQVFRTDAPFLLDKLHD 242
Query: 237 EYQNKISELSNYLGINSLAVDKESGVSDEEAKSNRGFTTSNSNIYLKGREP-ITFLSKRY 295
E +++L ++GIN+ DK+ + EA SN G ++N + K R + ++K Y Sbjct: 243 EKLRVMNQLLTFIGINNNPΞDKKERLWSEAISNNGVISANIEVGWKSRRKFVELINKCY 302
Query: 296 GLDI 299
GL+I Sbjct: 303 GLEI 306
>gi|l429239|emb|CAA67658| (X99260) upper collar protein [Bacteriophage B103] Length = 308
Score = 44.9 bits (104), Expect = 8e-04
Identities » 40/159 (25%), Positives = 73/159 (45%), Gaps = 11/159 (6%)
Query: 150 YNSDIEI IEHYTDELAEVA-LΞRFΞLIMQAKFSKIFKSEINDESINQLVSEIYNG 203
YN+D++ +E + +LAE+ + + Q I ++ N S+ + ++ Sbjct: 121 YNNDLKCSTLPALEMFAQDLAELKEIIAVNQNAQKTPVLIAANDNNQLSLKNIYNQYEGN 180
Query: 204 APFVKMSPMFNADD-DIIDLTSNSVIPALTEMKREYQNKISELSNYLGINSLAVDKESGV 262
AP + + + D+ + + V+ L K N E+ YLGI + ++K+ + Sbjct: 181 APVIFVHESLDLDNLKVFKTDAPYWDKLNAQKNAVWN EVMTYLGIKNANLEKKERM 237
Query: 263 ΞDEEAKSNRGFTTSNSNIYLKGR-EPITFLSKRYGLDIK 300
E SN S+ NIYLK R E +S+ YGL++K Sbjct: 238 VTSEVDSNDEQIESSGNIYLKARQEACNKISELYGLNLK 276
>gi|137915|sp|P07535|VG10_BPPZA UPPER COLLAR PROTEIN (CONNECTOR
PROTEIN) (LATE PROTEIN GP10) >gi | 75851 |pir | |WMBP10 gene
10 protein - phage PZA >gi| 216059 (M11813) upper collar protein [Bacteriophage PZA] Length = 309
Score = 43.8 bits (101), Expect = 0.002
Identities = 38/160 (23%) , Positives = 75/160 (46%) , Gaps = 13/160 (8%)
Query: 150 YNSDIEI IEHYTDELAEVALSRFSLIMQAKFSKIF--KSEINDESINQLVSEIYN 202
YN+D+ +E + ELAE+ S+ A+ + + ++ N S+ Q+ ++ Sbjct: 122 YNNDMSFPTTPTLELFAAELAELK-EIISVNQNAQKTPVLIRANDNNQLSLKQVYNQYEG 180
Query: 203 GAPFVKMSPMFNADD-DIIDLTSNSVIPALTEMKREYQNKISELSNYLGINSLAVDKESG 261
AP + ++D ++ + V+ L K N E+ +LGI + ++K+ Sbjct: 181 NAPVIFAHEALDSDSIEVFKTDAPYWDKLNAQKNAVWN EMMTFLGIKNANLEKKER 237
Query: 262 VSDEEAKSNRGFTTSNSNIYLKGR-EPITFLSKRYGLDIK 300
+ +E SN S+ ++LK R E +++ YGLD+K Sbjct: 238 MVTDEVSSNDEQIESΞGTVFLKΞREEACEKINELYGLDVK 277
>gi|l37914|sp|P04332|VG10_BPPH2 UPPER COLLAR PROTEIN (CONNECTOR
PROTEIN) (LATE PROTEIN GP10) >gi | 75852 | pir] | WMBPC9 gene 10 protein - phage phi-29 >gi| 215328 (M14782) upper collar protein [Bacteriophage phi-29] >gi| 215340 (M124S6) plO connector protein [Bacteriophage phi-29] >gi| 2241611 rf | |1011232A protein plO, connector [Bacteriophage phi-29] >gi | 225365 | rf | | 1301270E gene 10 [Bacteriophage phi-29] Length = 309
Score = 41.4 bits (95), Expect = 0.009
Identities = 37/160 (23%), Positives = 75/160 (46%), Gaps = 13/160 (8%)
Query: 150 YNSDIEI IEHYTDELAEVALSRFSLIMQAKFSKIF--KSEINDESINQLVSEIYN 202
YN+D+ +E + ELAE+ S+ A+ + + ++ N S+ Q+ ++ Sbjct: 122 YNNDMAFPTTPTLELFAAELAELK-EIISVNQNAQKTPVLIRANDNNQLSLKQVYNQYEG 180
Query: 203 GAPFVKMSPMFNADD-DIIDLTSNSVIPALTEMKREYQNKISELSNYLGINSLAVDKESG 261
AP + ++D ++ + V+ L K N E+ +LGI + ++K+ Sbjct: 181 NAPVIFAHEALDSDSIEVFKTDAPYWDKLNAQKNAVWN EMMTFLGIKNANLEKKER 237
Query: 262 VSDEEAKSNRGFTTSNSNIYLKGR-EPITFLSKRYGLDIK 300
+ +E SN S+ ++LK R E +++ YGL++K Sbjct: 238 MVTDEVSSNDEQIESSGTVFLKSREEACEKINELYGLNVK 277
Query= pt| 110878 44AHJDORF008 Phage 44AHJD ORF | 3020-3775 | 2 1 (251 letters)
>gi|4982468|gb|AAD30963.2| (AF118151) SNFl/AMP-activated kinase [Dictyostelium discoideum] Length = 718
Score = 52.3 bits (123), Expect = 3e-06
Identities = 28/118 (23%) , Positives = 56/118 (46%) , Gaps = 5/118 (4%)
Query: 121 YLQSQGFTEHNEDTTSNTDETSNQNATSLDNSTGMTANRNAYV SLPQSEVNIDVDN 176
+ + GF N ++ SN + +N N + N+ T N N + ++ + +N + +N Sbjct: 382 FTTTTGFNPTNSNSISNNNNNNNNNNNNTTNNNNNTTNNNNSIINNNNINNNNINNNNNN 441
Query: 177 TTLRFADNNTIDNGKTVNKSSNESNQNAKRNQNQKGNAKGTQFTKQYLID-NIDKAYD 233
+NN I+N N ++N +N N N N N+ + T+ + I N++ +Y+ Sbjct: 442 -lNNNINNNNII NNNNNNNNNNπ_NNNNNNNNNNNSSISGGTEVFSISP -_ΛNS 499
Score = 37.5 bits (85), Expect = 0.094
Identities = 17/111 (15%) , Positives = 45/111 (40%)
Query: 130 HNEDTTΞNTDETSNQNATSLDNSTGMTANRNAYVΞLPQSEVNIDVDNTTLRFADNNTIDN 189
+N + +N + +N N + +N++ ++ + P + + +++ N+ ++ Sbjct: 456 NNNNNNNNNNNNNNNNNNNNNNNΞSISGGTEVFSISPNLNNSYNSNSSGNSNGSNSNNNS 515
Query: 190 GKTVNKSSNESNQNAKRNQNQKGNAKGTQFTKQYLIDNIDKAYDLRKKILN 240
N +N +N N N N N ID+++ + + + N
Sbjct: 516 NNNTNNDNNNNNNNNNNNNNNNNNNNNNNNNNNNCIDSVNNSLNNENDVNN 566
Score = 32.8 bits (73), Expect = 2.4
Identities = 31/140 (22%) , Positives = 57/140 (40%) , Gaps = 14/140 (10%)
Query: 109 LNWYSSSEVEKYLQSQGFTEHNEDTTS---NTDETSNQNATSLDNSTGMTANRNAYVSL 165
LN Y+S+ S N +T + N + +N N + +N+ N N + Sbjct: 494 LNNSYNSNSSGNSNGSNSNNNSNNNTNNDNNNNNNNNNNNNNNNNNNNNNNNNNNNCIDS 553
Query: 166 PQSEVN--IDVDNTTLRFADNNTIDNGKTVNKSS- -NESNQNAKRNQNQKGNAK 215
+ +N DV+N+ + +NN D+G N ++ N N + N GN
Sbjct: 554 VNNSLNNENDVNNSNINNNNNNNSDDGSNNNSYEGGGDVLLLSDLNGNNQLGGNDNGNW 613
Query: 216 GTQFTKQYLIDNIDKAYDLR 235
Q L++++D D++ Sbjct: 614 NLNNNFQ-LLNSLDLNSDIQ 632
Score = 31.7 bits (70), Expect = 5.4
Identities = 25/115 (21%) , Positives = 48/115 (41%) , Gaps = 10/115 (8%)
Query: 130 HNEDTTSNTDETSNQNATSLDNST GMTAN-RNAYVSLPQSEVNIDVDNTTLRFADNN 185
+N + +N + +N N +S+ T ++ N N+Y S S N + N+ +N Sbjct: 462 NNNNNNNNNNNNNNNNNSSISGGTEVFSISPNLNNSYNS--NSSGNSNGSNΞNNNSNNNT 519
Query: 186 TIDNGKTVNKSSNESNQNAKRNQNQKGNAKGTQFTKQYLIDNIDKAYDLRKKILN 240
DN N ++N +N N N N N + ++++ D+ +N
Sbjct: 520 NNDN NNNNNNNNNNNNNNNNNNNNNNNNNNCIDSVNNSLNNENDVNNSNIN 570
Score = 31.7 bits (70), Expect = 5.4
Identities = 15/104 (14%) , Positives = 43/104 (40%)
Query: 110 NWYSSSEVEKYLQSQGFTEHNEDTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSE 169
N+ +++ + + +N + +N + +N N + +N+ + + V Sbjct: 434 NINNNNNNNNNNINNNNIINNNNNNNNNNNNNNNNNNNNNNNNNNSSISGGTEVFSISPN 493
Query: 170 VNIDVDNTTLRFADNNTIDNGKTVNKSSNESNQNAKRNQNQKGN 213
+N ++ + ++ + +N N +++ +N N N N N Sbjct: 494 UINSYNSNSSGNSNGSNSNNNSNNNTNNDNN_mNNNNNNNNNNN 537
Score = 30.9 bits (68), Expect = 9.2
Identities = 16/84 (19%) , Positives = 34/84 (40%)
Query: 130 HNEDTTSNTDETSNQNATΞLDNSTGMTANRNAYVSLPQSEVNIDVDNTTLRFADNNTIDN 189
+N + +N + +N N + +N+ + S+ + N N++ +N+ +N Sbjct: 455 NNNNNNNNNNNNNNNNNNNNNNNNSSISGGTEVFSISPNLNNSYNSNSSGNSNGSNSNNN 514
Query: 190 GKTVNKSSNESNQNAKRNQNQKGN 213
+ N +N N N N N Sbjct: 515 SNNNTNNDNNNNNNNNNNNNNNNN 538
>gi|l730077|sp|P18160|KYKl_DICDI NON-RECEPTOR TYROSINE KINASE SPORE LYSIS A (TYROSINE-PROTEIN KINASE 1) >gi | 974334 (U32174) non-receptor tyrosine kinase [Dictyostelium discoideum] Length = 1584
Score = 46.5 bits (108), Expect = 2e-04
Identities = 29/106 (27%), Positives = 48/106 (44%), Gaps = 4/106 (3%)
Query: 130 HNEDTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSEVNID VDNTTLRFADN-N 185
+NED +SN + +N N + +N+ N N + + N + ++NTT N N Sbjct: 442 N EDISSNNNNNN^I NNNNNNN N NNNNNNNN^IN SNSSNT-INN INNTTNNNNS SN 501
Query: 186 TIDNGKTVNKSSNESNQNAKRNQNQKGNAKGTQFTKQYLIDNIDKA 231
+N N +SN +N N N N N TK+ I + D++ Sbjct: 502 NNNNNNNSNSNSNSNNNNINNNNNNNNNNNNIYLTKKPSIGSTDES 547
Score = 34.0 bits (76), Expect = 1.1
Identities = 20/117 (17%) , Positives = 46/117 (39%)
Query: 87 NRQTVEAFGMQVITVCITHEDYLNWYSSSEVEKYLQSQGFTEHNEDTTΞNTDETSNQNA 146 N G IT T + + ++ ++ + +N + +N + +N N
Sbjct: 415 NNNNNNIIGNGKITTTTTTSTSPSSINNNEDISSNNNNNNNNNNNNNNNNNNNNNNNNNN 474
Query: 147 TSLDNSTGMTANRNAYVSLPQSEVNIDVDNTTLRFADNNTIDNGKTVNKSSNESNQN 203
+ ++++ T N N + + N + +N N+ +N N ++N +N N Sbjct: 475 NNNNSNSSNTNNNNINNTTNNNNSNSNNNNNNNNSNSNSNSNNNNINNNNNNNNNNN 531
Score = 33.2 bits (74), Expect = 1.8
Identities = 18/88 (20%) , Positives = 35/88 (39%)
Query: 130 HNEDTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSEVNIDVDNTTLRFADNNTIDN 189
+N + ++N + +N N T T + S+ +E +N +NN +N Sbjct: 405 NNNNNSNNNNNNNNNNIIGNGKITTTTTTSTSPSSINNNEDISSNNNNNNNNNNNNNNNN 464
Query: 190 GKTVNKSSNESNQNAKRNQNQKGNAKGT 217
N ++N +N N+ + T Sbjct: 465 NNNNNNNNNNNNNNSNSSNTNNNNINNT 492
Score = 32.5 bits (72), Expect = 3.1
Identities = 18/94 (19%) , Positives = 37/94 (39%)
Query: 120 KYLQSQGFTEHNEDTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSEVNIDVDNTTL 179
K + S N + +N++ +N N ++ + +T S N D+ + Sbjct: 392 KNVNSTSILVPNGNNNNNSNNNNNNNNNNIIGNGKITTTTTTSTSPSSINNNEDISSNNN 451
Query: 180 RFADNNTIDNGKTVNKSSNESNQNAKRNQNQKGN 213
+NN +N N ++N +N N + + N Sbjct: 452 NN^] NNNNNNI_raN_I_JN _MN NNNNSNSSNTN 485
Score = 32.5 bits (72), Expect = 3.1
Identities = 24/110 (21%) , Positives = 44/110 (39%) , Gaps = 10/110 (9%)
Query: 138 TDETSNQNATSLDNSTGMTANRNAYVSLPQSEVNIDVDNTTLRFADNNTIDNGK 191
T T++ + +S++N+ +++N N + + N + +N +NN N Sbjct: 429 TTTTTSTSPSSINNNEDISSNNNNNNNNNNNNNNNNNNNNNNNNNNNNNNSNSSNTNNNN 488
Query: 192 TVNKSSNESNQNAKRNQNQKGNAKGTQFTKQYLIDNIDKAYDLRKK 237
T N +SN +N N N N N+ +N + L KK
Sbjct: 489 I_TOTTNNNNSNSNNNNNNNNSNSNSNSNNNNINNNNNN_J_raNNNIYLTKK 538
>gi|3758855|emb|CAB11140.l| (Z98551) predicted using hexExon;
MAL3P6.11 (PFC0760c), Hypothetical protein, len: 3395 aa [Plasmodium falciparum] Length = 3394
Score = 46.5 bits (108), Expect = 2e-04
Identities = 52/202 (25%) , Positives = 96/202 (46%) , Gaps = 32/202 (15%)
Query: 21 FNEFVNDNKLTFYDDEFQFMQKMLKFD-KDVLAIVNEKVFKGFSLKDELSDL--LFKKSF 77
F ++ ++ K T D+ M+K K D DV + NEK++ L ++L+ + + KK Sbjct: 665 FEKYCSNIKNTLIRDD---MKKFRKPDISDVHILHNEKIYLEKLLNEKLNYIKDIEKKLD 721
Query: 78 TIHFLDREINRQTVEAFGMQV ITVCITHEDYLNWYSSSEVEKYLQSQGFTEHNE 132
+H + IN+ + + +QV I V + DY + S + + K + +N Sbjct: 722 ELHGV---INKNKEDIYILQVEKQTLIKVISSVYDYTKME-SENHIFKMNTTWNKMLNNV 777
Query: 133 DTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSEVNIDVDNTTLRFADNNTIDNGKT 192
+SN D +NQN +++N+ + N+N N +++N + N +N Sbjct: 778 HMSSNKDY-NNQNNQNIENNQNIENNQN NQNIEN NQNIENNQNN 820
Query: 193 VNKSSNESNQNAKRNQNQKGNA 214
N +N++NQN + NQN + NA Sbjct: 821 QNNQNNQNNQNNQNNQNNQNNA 842
Score = 33.6 bits (75), Expect = 1.4
Identities = 46/221 (20%) , Positives = 89/221 (39%) , Gaps = 37/221 (16%) - _ —
Query: 10 DFIKSELIKKGFNEFVNDNKLTFYDDEFQFMQKMLKFDKDVLAIVNEKVFKGFSLKDELS 69 ~~
D +K E K N + +L Y + + M+K K + V K SL
Sbjct: 367 DSLKIEYNKSKTNIQQLNEQLVNYKNFIKEMEKKYK QLWKNNSLFSITH 416
Query: 70 DLLFKKSFTIHFLDREINRQTVEAFGMQVITVCITH---EDYLNWYSSSEVEKYLQSQG 126
D + K+ I + R + + + ++ + I H +D+L+V+Y + + L + Sbjct: 417 DFINLKNSNIIIIRRTSDMKQI FKMYNLDIEHFNEQDHLSVIY IYEILYNTN 468
Query: 127 FTEHNEDTTSNTDETSNQNATΞLDNSTGMTANRNAYVSLPQSEVNIDVDNTTLRFADNNT 186
+N D +N D +N N + +N+ N N N + +N + Sbjct: 469 -D^MNNDNDNNND ^JNNNNNNDNNNINNDNNNN NNNYNNIMM M 512
Query: 187 IDNGKTVNKSSNESNQNAKRNQNQKGNAKGTQFTKQYLIDN 227
I+N + N ++ + N + N N + N + + +Y I+N Sbjct: 513 IENMNSGNHPNSNNLHNYRHNTNDENNLΞSLKTSFRYKINN 553
Score = 32.8 bits (73), Expect = 2.4
Identities = 28/122 (22%) , Positives = 53/122 (42%) , Gaps = 2/122 (1%)
Query: 119 EKYLQSQGFTEHNEDTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSEVNID-VDNT 177
E Y S + +++ N + +N + + DN+ N N ++ +N D ++N Sbjct: 2838 ENYPVSTHYDNNDDINKDNINNDNNNDNINDDNNNDNINNDNNNDNINNDNINNDNINND 2897
Query: 178 TLRFADNNTIDNGKTVNKSSNESNQNAKRNQNQKGNAKGTQFTKQYLIDNIDKAYDLRKK 237
+N+ +NG SSN ++ N NN KN +G + + + + YD K Sbjct: 2898 NNNDNNNDNSNNGFVCELSSNINDFNNILNVN-KDNFQGINKSNNFSTNLSEYNYDAYVK 2956
Query: 238 IL 239
1+ Sbjct: 2957 IV 2958
Score = 32.5 bits (72), Expect = 3.1
Identities = 46/249 (18%) , Positives = 101/249 (40%) , Gaps = 31/249 (12%)
Query: 9 YDFIKSELIKKGFNEFVNDNKLTFYDDEFQFMQKMLKFDKDVLAIVNEKVFKGFSLKDEL 68
Y+++K ++ N N NK E Q++ K+ + + + +E K L++ Sbjct: 2150 YNYVK---VQNATNREDNKNK ERNLSQEIYKYINENIDLTSELEKKNDMLENYK 2200
Query: 69 SDL LFKKSFTIHFLDREINRQTVEAFGMQVITVCITHEDYLNWYSSSEVEKYL 122
++L ++K + I L + M+ + + N + E+ + L Sbjct: 2201 NELKEKNEEIYKLNNDIDMLSNNCKKLKESIMMMEKYKIIMN NNIQEKDEIIENL 2255
Query: 123 QSQGFTEHNEDTTSNTDETSNQNATSLDNSTGMTAN RNAYVSLPQSE VNIDV 174
+++ + +D +N + ++S M+ + N + +L +S N+D+
Sbjct: 2256 KNK-YNNKLDDLINNYSWDKSIVSCFEDSNIMSPSCNDILNVFNNLSKSNKKVCTNMDI 2314
Query: 175 DNTTLRFADNNTIDNGKTVNKSSNESNQNAKRNQNQKGNAKGTQFTKQYLIDNIDKAYDL 234
N + ++I+N +N +N +N N N N N K YL++N+ D Sbjct: 2315 CMENMDSI--SSINNVNNINNVNNINNVNNINNVNNINNVKNIVDINNYLVNNLQLNKDN 2372
Query: 235 RKKILNEFD 243
1+ +F+ Sbjct: 2373 DNIIIIKFN 2381
Score = 32.1 bits (71), Expect = 4.1
Identities = 20/103 (19%) , Positives = 48/103 (46%) , Gaps = 2/103 (1%)
Query: 115 SSEVEKYLQSQGFTEHNEDTTSNTDETSNQN--ATSLDNSTGMTANRNAYVSLPQSEVNI 172
+++ EKY EH + N D +N+N L ++ ++ + N S ++E+ Sbjct: 3264 NNDEEKYSCHDDKNEHTNNDLLNIDHDNNKNNITDELYSTYNVSVSHNKDPSNKENEIQN 3323
Query: 173 DVDNTTLRFADNNTIDNGKTVNKSSNESNQNAKRNQNQKGNAK 215
+ + D N ++ N ++E+++N + ++N + + K Sbjct: 3324 LISIDSSNENDENDENDENDENDENDENDENDENDENDENDEK 3366
Score = 30.9 bits (68), Expect = 9.2
Identities = 27/118 (22%) , Positives = 53/118 (44%) , Gaps = 15/118 (12%)
Query: 104 THEDYLNWYSSSEV EKYLQSQGFTEHNEDTTSNTDETSNQNATSLDNSTGMTANR 159
T+ D LN+ + +++ E Y HN+D ++ +E QN S+D+S N Sbjct: 3280 TNNDLLNIDHDNNKNNITDELYSTYNVSVSHNKDPSNKENEI--QNLISIDSSNENDEND 3337 '
Query: 160 NAYVSLPQSEVNIDVDNTTLRFADNNTIDNGKTVNKSSNESNQNAKRNQNQKGNAKGT 217
+++ N + D D N ++ N +E+++N + ++N N +GT Sbjct: 3338 EN DENDENDENDEN DENDENDENDENDEKDENDENDENDENFDNNNEGT 3386
>gι|585795|sp|P21538|REBl_YEAST DNA-BINDING PROTEIN REB1 (QBP)
>gι|626139 |pιr I |S45907 DNA-bmding protem REB1 - yeast (Saccharomyces cerevisiae) >gι | 536280 | emb| CAA84992 | (Z35918) ORF YBR049C [Saccharomyces cerevisiae]
>gι|559944 |emb|CAA86391| (Z46260) REB1 DNA-bindmg protem [Saccharomyces cerevisiae]
Length = 810
Score = 45 7 bits (106), Expect = 3e-04
Identities = 34/158 (21%), Positives = 72/158 (45%), Gaps = 14/158 (8%)
Query 83 DREINRQTVEAFGMQVITVCITHEDYLNWYSSSEVEKYLQSQGFTEHNEDTTSNTDETS 142
D+ N+++VE ++ + V + H+++ +++ K+ + Q E + D N ++ S Sbjct 7 DKNANQESVEEAVLKYVGVGLDHQNHDPQLHTKDLENKHSKKQNIVESSSDVDVNNNDDS 66
Query 143 NQNATSLDNSTGMTANRNAYVSLPQSEVNIDVDNTTLRFADNNTID NGKTVNKSSNE 199
N+N + D+S ++A L +E + +VD+ N +D N+ +E Sbjct 67 NRNEDNNDDSENISA LNANESSSNVDHANSNEQHNAVMDWYLRQTAHNQQDDE 119
Query 200 SNQNAKRNQNQKGNAKGTQFTKQYLIDNIDKAYDLRKK 237
++N N GN F++ ++ +D D KK Sbjct 120 DDEN--NNNTDNGNDSNNHFSQSDIV--VDDDDDKNKK 153
>gι 1172372 (M58728) DNA-bmding protem [Saccharomyces cerevisiae] Length = 809
Score = 45 7 bits (106) , Expect = 3e-04
Identities = 34/158 (21%), Positives = 72/158 (45%), Gaps = 14/158 (8%)
Query 83 DREINRQTVEAFGMQVITVCITHEDYLNWYSSSEVEKYLQSQGFTEHNEDTTSNTDETS 142
D+ N+++VE ++ + V + H+++ +++ K+ + Q E + D N ++ S Sbjct 7 DKNANQESVEEAVLKYVGVGLDHQNHDPQLHTKDLENKHSKKQNIVESΞNDVDVNNNDDS 66
Query 143 NQNATSLDNSTGMTANRNAYVSLPQSEVNIDVDNTTLRFADNNTID NGKTVNKSSNE 199
N+N + D+S ++A L +E + +VD+ N +D N+ +E Sbjct 67 NRNEDNNDDSENISA LNANESSSNVDHANSNEQHNAVMDWYLRQTAHNQQDDE 119
Query 200 SNQNAKRNQNQKGNAKGTQFTKQYLIDNIDKAYDLRKK 237
++N N GN F++ ++ +D D KK Sbjct 120 DDEN--NNNTDNGNDSNNHFSQSDIV--VDDDDDKNKK 153
>gι 12952545 (AF051898) coron binding protein [Dictyostelium discoideum] Length = 560
Score = 44 9 bits (104) , Expect = 6e-04
Identities = 26/83 (31%) , Positives = 39/83 (46%) , Gaps = 5/83 (6%)
Query 131 NEDTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSEVNIDVDNTTLRFADNNTIDNG 190
N + +N +N N+ S +NS +N N+ + P N D DN T +NNT +N Sbjct 404 NNNNNNNIINNNNSNSNSNNNSNN-NSNNNSNRNSPNHNNNGDNDNNT NNNTNNNN 458
Query 191 KTVNKSSNESNQNAKRNQNQKGN 213
N ++N +N N N N N Sbjct 459 NNNNNNNNNNNNNNNNNNNNNNN 481
Score = 41 4 bits (95), Expect = 0 006
Identities = 22/88 (25%) , Positives = 43/88 (48%) , Gaps = 6/88 (6%)
Query 130 HNEDTTSNTDETSNQNATSLDN STGMTANRNAYVSLPQSEVNIDVDNTTLRFADNNT 186
+ ++ +N++ SN N+ + +N + G AN++ + P + +N + DN +NN Sbjct 337 NRNNSNNNSNNNSNNNSNNSNNRNITNGSNANKS NSPNNNLNTNNDNKNNNSNNNNN 393
Query 187 IDNGKTVNKSSNESNQNAKRNQNQKGNA 214
+N S+N +N N N N N+ Sbjct 394 SNNNSNNGNSNNNNNNNIINNNNSNSNS 421
Score = 40 6 bits (93), Expect = 0 011
Identities = 24/101 (23%) , Positives = 41/101 (39%) , Gaps = 2/101 (1%)
Query 115 SSEVEKYLQSQGFTEHNEDTTSNTDETSNQNATΞLDNSTGMTANRNAYVSLPQSEVNIDV 174 S+ L + ++N +N ++ N S +N+ N N S + N +
Sbjct: 370 SNSPNNN-JTNITONKNNNSNNNNNSNNNSNNGNSNNNNNNNIINNNNSNSNSNNNSNNNS 429
Query: 175 DNTTLRFADN--NTIDNGKTVNKSSNESNQNAKRNQNQKGN 213
+N + R + N N DN N ++N +N N N N N Sbjct: 430 NNNSN NSP HNNNGDNDNNTNn.T nM^NNNNNNNNNNN 470
Score = 40.2 bits (92), Expect = 0.014
Identities = 21/80 (26%) , Positives = 39/80 (48%) , Gaps = 9/80 (11%)
Query: 130 HNEDTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSEVNIDVDNTTLRFADNNTIDN 189
+N D +NT+ +N N + +N+ N N N + +N +ADN+ ++
Sbjct: 442 NNGDNDNNTNNNTNNNNNNNNNNNNNNNNNNN ---NNNNNNNNNNYADNSNNNS 492
Query: 190 GKTVNKSSNESNQNAKRNQN 209
+ N +SN +N N +N+N Sbjct: 493 SNSNNNNSNSNNNNDNKNEN 512
Score = 39.5 bits (90), Expect = 0.024
Identities = 26/111 (23%) , Positives = 44/111 (39%) , Gaps = 20/111 (18%)
Query: 112 VYSSSEVEKYLQSQ--GFTEHNEDTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSE 169
VY + K+ ++ G +N ++ +N++ SN N ++N N N Sbjct: 296 VYCTHHHTKFYETHRNGLLNNNNNSNNNSNSNSNNNNNGINNRNNSNNNSN 346
Query: 170 VNIDVDNTTLRFADNNTIDNGKTVNKSS NESNQNAKRNQNQKGNA 214
+ N ++N I NG NKS+ N +N N N N N+ Sbjct: 347 ---NNSNNNSNNSNNRNITNGSNANKSNSPNNNLNTNNDNKNNNSNNNNNS 394
Score = 37.5 bits (85), Expect = 0.094
Identities = 24/96 (25%) , Positives = 41/96 (42%) , Gaps = 1/96 (1%)
Query: 124 SQGFTEHNEDTTSNTDETSNQNATSLDNSTGM-TANRNAYVSLPQSEVNIDVDNTTLRFA 182
S + +N + SN + ++ N DN+T T N N + + N + +N Sbjct: 421 SNNNSNNNSNNNSNRNSPNHNNNGDNDNNTNNNTNNNNNNNNNNNNNNNNNNN^ 480
Query: 183 DNNTIDNGKTVNKSSNESNQNAKRNQNQKGNAKGTQ 218
+NN DN + +SN +N N+ N + K Q Sbjct: 481 NNNYADNSNNNSSNSNNNNSNSNNNNDNKNENSDNQ 516
Score = 35.6 bits (80), Expect = 0.36
Identities = 25/99 (25%) , Positives = 42/99 (42%) , Gaps = 18/99 (18%)
Query: 130 HNEDTTSNTDETSNQNATSLDNST-GMTANRNAYVSLPQSEVNIDVDNTTLRFADNNTID 188
+N + SN + +N N ++ N T G AN++ + P + +N + DN +NN + Sbjct: 339 NNSNNNSNNNSNNNSNNSNNRNITNGSNANKS NSPNNNLNTNNDNKNNNSNNNNNSN 395
Query: 189 NGKTV NKSSNESNQNAKRNQNQKGN 213
N N S++ SN N+ N N N
Sbjct: 396 NNSNNGNSNNNNNNNIINNNNSNSNSNNNSNNNSNNNSN 434
Score = 35.2 bits (79), Expect = 0.47
Identities = 21/94 (22%) , Positives = 42/94 (44%) , Gaps = 5/94 (5%)
Query: 124 SQGFTEHNEDTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSEVNIDVDNTTLRFAD 183
+ G + ++ +N T+N N + N+ N N+ + N + +N + + Sbjct: 362 TNGSNANKSNSPNNNLNTNNDNKNNNSNN NNNSNNNSNNGNSNNNNNNNIINNNN 416
Query: 184 NNTIDNGKTVNKSSNESNQNAKRNQNQKGNAKGT 217
+N+ N + N S+N SN+N+ + N N T Sbjct: 417 SNSNSNNNSNNNSNNNSNRNSPNHNNNGDNDNNT 450
Score = 35.2 bits (79), Expect = 0.47
Identities = 29/118 (24%) , Positives = 53/118 (44%) , Gaps = 12/118 (10%)
Query: 115 SSEVEKYLQS-QGFTEHNEDTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSEVNID 173 __
SS+ E ++ +GF + + T+N ++N D S+G + + + V+ P+S +N Sbjct: 114 SSDSEADIEDDKGFQD--KPITTNNSGSNNPLKNLKDYSSGSSGSSRΞGVNQPRSNINNS 171
Query: 174 VDNTTLRFADNNT IDNGKTVNKSSNESNQNAKRNQNQKGNAKGTQFTKQ 222
D + + +N+ I + T + NQN +NQNQ N Q +Q
Sbjct: 172 NDKYKSKSSSSNSNSSSSGGSLISSLLTGGNTYQNQNQNQNQNQNQNNNQSQLQQQQQ 229
Score = 34.4 bits (77), Expect = 0.81
Identities = 24/94 (25%), Positives = 38/94 (39%), Gaps = 12/94 (12%)
Query: 131 NEDTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSEVNIDVDNTTLRFADNNTIDNG 190
N +T +N + +N N + +N+ N N S N N +NN+ N Sbjct: 451 NNNTNNNNNNNNNNNNNNNNNNNNNNNNNNNNNYADNSNNNSSNSN NNNSNSNN 504
Query: 191 KTVNKSSNESNQNAKR NQNQKGNAKGTQ 218
NK+ N NQ+ R ++NQK + Q Sbjct: 505 NNDNKNENSDNQSVLRSNEKFTDENQKNGSDDQQ 538
Score = 33.6 bits (75), Expect = 1.4
Identities = 22/90 (24%) , Positives = 35/90 (38%)
Query: 124 SQGFTEHNEDTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSEVNIDVDNTTLRFAD 183
S N SN +++++ N N+ N N + + N + +N Sbjct: 353 SNNSNNRNITNGSNANKSNSPNNNIjπϊINDNKNNNSNNNNNSNNNSNNGNSNNNNNNNII 412
Query: 184 NNTIDNGKTVNKSSNESNQNAKRNQNQKGN 213
NN N + N S+N SN N+ RN N Sbjct: 413 NNNNSNSNSNNNSNNNSNNNSNRNSPNHNN 442
>gi|535260|emb|CAA82996| (Z30339) ΞTARP antigen [Plasmodium reichenowi] Length = 655
Score = 44.5 bits (103), Expect = 7e-04
Identities = 31/114 (27%) , Positives = 47/114 (41%) , Gaps = 14/114 (12%)
Query: 128 TEHNEDTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSEVN IDVDNTTLRF 181
T++N T TD + + +N+T A N + ++ N D +NT + Sbjct: 433 TDNNNTNTKATDSNNTNTKATDNNNTNTKATDNNNTNTKATDNNNTNTKATDNNNTNTKA 492
Query: 182 ADNNTI DNGKTVNKSSNESNQNAKRNQNQKGNAKGT---QFTKQYLIDN 227
DNN DN T K+++ +N N K N N K T T QY+ N Sbjct: 493 TDNNNTNTKATDNNNTNTKATDNNNTNTKATDNNNTNTKATDNNNNTNQYVFAN 546
Score = 44.5 bits (103), Expect = 7e-04
Identities = 30/103 (29%) , Positives = 44/103 (42%) , Gaps = 13/103 (12%)
Query: 128 TEHNEDTTSNTDETSNQNATSLDNS TGMTANRNAYVSLPQSEVN IDVDNTTL 179
T++N T TD+++N + + DN+ T T N N S D +NT Sbjct: 401 TDNNNTDTKATDKSNNTDTKATDNNNNTDTKATDNNNTNTKATDSNNTNTKATDNNNTNT 460
Query: 180 RFADNNTI DNGKTVNKSSNESNQNAKRNQNQKGNAKGT 217
+ DNN DN T K+++ +N N K N N K T Sbjct: 461 KATDNNNTNTKATDNNNTNTKATDNNNTNTKATDNNNTNTKAT 503
Score = 42.6 bits (98), Expect = 0.003
Identities = 27/96 (28%) , Positives = 43/96 (44%) , Gaps = 10/96 (10%)
Query: 128 TEHNEDTTSNTDETSNQNATSLD-NSTGMTANRNAYVΞLPQSEVNIDVDNTTLRFADNNT 186
T++N +T + + +N N + D N+T A N + ++ N NT + DNN Sbjct: 422 TDNNNNTDTKATDNNNTNTICATDSNNTNTKATDNNNTNTKATDNN NTNTKATDNNN 477
Query: 187 I DNGKTVNKSSNESNQNAKRNQNQKGNAKGT 217
DN T K+++ +N N K N N K Sbjct: 478 TNTKATDNNNTNTKATDNNNTNTKATDNNNTNTKAT 513
Score = 41.8 bits (96), Expect = 0.005
Identities = 35/150 (23%) , Positives = 59/150 (39%) , Gaps = 9/150 (6%)
Query: 85 EINRQTVEAFGMQVITVCITHEDYLNWYSSSEVEKYLQSQGFTEHNEDTTSNTDETSNQ 144 __
E N+ ++ G T+ + N + E + +Q T +N TT+ + N Sbjct: 118 ETNKTNIKLTGNNSTTINTNLTENTNA--TKKLTENVITNQILTGNNNTTTNTSΞTEHNN 175
Query: 145 NATSLDNSTGMTANRNAYVSLPQSEVNIDVDNTTLRFADNNTIDNGKTVNKSSNESNQNA 204 N + NSTG T+ NI + N L +N T + T + ++ +N N+
Sbjct: 176 NINTNTNSTGNTSTTKKLTE NI-ITNQILTGNNNTTTNTSSTEHNNNINTNTNS 228
Query: 205 KRNQNQKGNAKGTQFTKQYLIDNIDKAYDL 234
N N N T + DNI+ +L Sbjct: 229 TDNSNTNTNLTDITTTTKKWTDNINTTQNL 258
Score = 41.8 bits (96), Expect = 0.005
Identities = 30/101 (29%), Positives = 43/101 (41%), Gaps = 13/101 (12%)
Query: 130 HNEDTTSNTDETSNQNATSLDNS-TGMTANRNAYVSLPQSEVNIDV DNTTLRFA 182
+N DT S ++ ++ AT DN+ T T N N + N D +NT + Sbjct: 363 NNTDTISTDNDNTDTKATDNDNTDTKATDNNNNTDTKATDNNNTDTKATDKSNNTDTKAT 422
Query: 183 DNN -TIDNGKTVNKSSNESNQNAKRNQNQKGNAKGT 217
DNN DN T K+++ +N N K N N K T Sbjct: 423 DNNNNTDTKATDNNNTNTKATDSNNTNTKATDNNNTNTKAT 463
Score = 40.6 bits (93), Expect = 0.011
Identities = 31/121 (25%), Positives = 47/121 (38%), Gaps = 31/121 (25%)
Query: 128 TEHNEDTTSNTDETSNQNAT SLDNSTGMTANRNAYVSLPQSEVN 171
TEHN + +NT+ T N + T ++ + +T N N + +E N Sbjct: 171 TEHNNNINTNTNSTGNTSTTKKLTENIITNQILTGNNNTTTNTSSTEHNNNINTNTNSTD 230
Query: 172 IDVDNTTLRFADN NTIDNGKTVNKSSNESNQNAKRNQNQKGNAKG 216
D+ TT ++ DN T N TV+ +N +N N K N N K Sbjct: 231 NSNTNTNLTDITTTTKKWTDNINTTQNLTTSTNTTTVSTDNNNNNINTKPTDNNNTNIKS 290
Query: 217 T 217
T Sbjct: 291 T 291
Score = 38.3 bits (87), Expect = 0.055
Identities = 28/98 (28%) , Positives = 41/98 (41%) , Gaps = 10/98 (10%)
Query: 128 TEHNEDTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSEVNIDVD-NTTLRFADNNT 186
TEHN + +NT+ S N+ + N T +T + + N+ NTT DNN Sbjct: 216 TEHNNNINTNTN--STDNSNTNTNLTDITTTTKKWTDNINTTQNLTTSTNTTTVSTDNNN 273
Query: 187 IDNGKTVNKSSNESNQNAKRNQNQKGNAKGT 217
DN T KS++ N K N+ + K T Sbjct: 274 NNINTKPTDNNNTNIKSTDNYNTGTKETDNKNTDIKAT 311
Score = 37.5 bits (85), Expect = 0.094
Identities = 31/106 (29%) , Positives = 45/106 (42%) , Gaps = 18/106 (16%)
Query: 128 TEHNEDTTSNTDETSNQN ATSLDNSTGMTANRNAYVSLPQSEVN IDVDN 176
T++N +T +T T N N AT N+T A N + ++ N D +N Sbjct: 390 TDNNNNT--DTKATDNNNTDTKATDKSNNTDTKATDNNNNTDTKATDNNNTNTKATDSNN 447
Query: 177 TTLRFADNN TIDNGKTVNKSSNESNQNAKRNQNQKGNAKGT 217
T + DNN DN T K+++ +N N K N N K T Sbjct: 448 TNTKATDNNNTNTKATDNNNTNTKATDNNNTNTKATDNNNTNTKAT 493
Score = 35.2 bits (79), Expect = 0.47
Identities = 24/109 (22%) , Positives = 46/109 (42%) , Gaps = 6/109 (5%)
Query: 128 TEHNEDTTSNTDETSNQNATSLDNSTGMTANRNAYVSLPQSEVN IDVDNTTLRF 181
T++N T TD + + +N+T A N + ++ N D +NT + Sbjct: 473 TDNNNTNTKATDNNNTNTKATDNNNTNTKATDNNNTNTKATDNNNTNTKATDNNNTNTKA 532
Query: 182 ADNNTIDNGKTVNKSSNESNQNAKRNQNQKGNAKGTQFTKQYLIDNIDK 230
DNN N + +E+ + K N++ N++ + K + +DK Sbjct: 533 TDNNNNTNQYVFANNYDETTSDDKLNKDΞCDNSEEKENIKSMINAYLDK 581
Score = 34.4 bits (77), Expect = 0.81
Identities = 26/126 (20%) , Positives = 46/126 (35%) , Gaps = 7/126 (5%)
Query: 99 ITVCITHEDYLNWYSSSEVEKYLQSQGFTEHNEDTTSNTDETSNQNATSLDNSTGMTAN 158
IT T+ + ++ S + V S T +++ +N T N N ++ T Sbjct: 318 ITTDNTNTNVISTDNSKTNVISKDNSNTHTISTDNSKTNVISTDNNNTDTISTDNDNTDT 377
Query: 159 RNAYVSLPQSEVNIDVDNTTLRFADNNTID NGKTVNKSSNESNQNAKRNQNQK 211
+ ++ + +NT + DNN D N + N +N + K N Sbjct: 378 iATDNDNTDTPATDNNNNTDTKATDNNNTDTKATDKSNNTDTKATDNNNNTDTKATDNNN 437
Query: 212 GNAKGT 217
N K T Sbjct: 438 TNTKAT 443
Score = 34.4 bits (77), Expect = 0.81
Identities = 30/100 (30%) , Positives = 44/100 (44%) , Gaps = 14/100 (14%)
Query: 131 NEDTTSNTDETSNQNATSLDNS-TGMTANRNAY VΞLPQSEVNI DVDNTTLRFAD 183
N + T TD T N N S DNS T + + N+ +S S+ N+ D +NT D Sbjct: 313 NNNITITTDNT-NTNVISTDNSKTNVISKDNSNTHTISTDNSKTNVISTDNNNTDTISTD 371
Query: 184 NNTIDNGKTVNKSS NESNQNAKRNQNQKGNAKGT 217
N+ D T N ++ N +N + K N + K T Sbjct: 372 NDNTDTKATDNDNTDTKATDNNNNTDTKATDNNNTDTKAT 411
Score = 34.4 bits (77), Expect = 0.81
Identities = 28/101 (27%), Positives = 41/101 (39%), Gaps = 15/101 (14%)
Query: 131 NEDTTSNTDETSNQNATSLDNSTGMTA--NRNAYVSLPQSEVNIDV DNTTLRFA 182
N DT + ++ ++ AT +N+T A N N N D +NT +
Sbjct: 374 NTDTKATDNDNTDTKATDNNNNTDTKATDNNNTDTKATDKSNNTDTKATDNNNNTDTKAT 433
Query: 183 DNNTIDNGK TVNKSSNESNQNAKRNQNQKGNAKGT 217
DNN N K T K+++ +N N K N N K T Sbjct: 434 DNNN-TNTKATDSNNTNTKATDNNNTNTKATDNNNTNTKAT 473
Score = 32.5 bits (72), Expect = 3.1
Identities = 30/110 (27%) , Positives = 40/110 (36%) , Gaps = 23/110 (20%)
Query: 131 NEDTTSNTDETSNQNATSLDNS TGMTANRNAYVSLPQS EVNIDVDNTTLRF 181
N +TT N ++N S DN+ T T N N + + D NT ++ Sbjct: 251 NINTTQNLTTSTNTTTVSTDNNNNNINTKPTDNNNTNIKSTDNYNTGTKETDNKNTDIKA 310
Query: 182 ADNNTI DNGKTVNKSSNESNQNAKRNQNQKGNAKGT 217
DNN I DN KT S + SN + N K N T
Sbjct: 311 TDNNNITITTDNTNTNVISTDNSKTNVIΞKDNSNTHTISTDNSKTNVIST 360
>gi|l429240|emb|CAA67659| (X99260) lower collar protein [Bacteriophage B103] Length = 293
Score = 43.8 bits (101), Expect = 0.001
Identities = 53/204 (25%) , Positives = 79/204 (37%) , Gaps = 42/204 (20%)
Query: 56 EKVFKG FSLKDELSDLLFKKSFTIHFLD REINRQTVEAFGMQVITVCITHED 107
EK+ KG F + + D ++K F HF+ REI +T F + T I + Sbjct: 26 EKIEKGRPKLFDFQYPIFDESYRKVFETHFIRNFYMREIGFETEGLFKFNLETWLIINMP 85
Query: 108 YLNWYSSSEVEKY LQSQGFTEH NEDTT SNTDETSNQNA 146
Y N ++ S E+ KY L + G ++ N DTT SNT + NA Sbjct: 86 YFNKLFES-ELIKYDPLENTRLNTTGNKKNDTERNDNRDTTGSMKADGKSNTKTSDKTNA 144
Query: 147 TSLDNSTGMTA NRNAYVSLPQSEVNIDVDN--TTLRFADNNTIDNGKTVNKS 196
T G T NR P S +N+ ++ TL +A + 1+ T NK
Sbjct: 145 TGSSKEDGKTTGSVTDDNFNRKIDSDQPDSRLNLTTNDGQGTLEYA--SAIEENNTNNKR 202
Query: 197 SNESNQNAKRNQNQKGNAKGTQFT 220
+ N + + GT T Sbjct: 203 NTTGTNNVTSSAESESTGSGTSDT 226
Query= pt| 110879 44AHJDORF009 Phage 44AHJD ORF | 5744-6496 | 2 1 (250 letters)
>gi|276498l|emb|CAA69021.l| (Y07739) N-acetylmuramoyl-L-alanine amidase [Staphylococcus phage Twort] Length = 467
Score = 180 bits (452) , Expect = le-44
Identities = 89/157 (56%) , Positives = 109/157 (68%) , Gaps = 8/157 (5%)
Query: 1 MKSQQQAKEWIYKHEGAGVDFDGAYGFQCMDLΞVAYVYYITDGKVRMWGNAKDAINNDFK 60
MK+ +QA+ +1 G DFDG YG+QCMDL+V Y+Y++TDGK+RMWGNAKDAINN F Sbjct: 1 MKTLKQAESYIKSKVNTGTDFDGLYGYQCMDLAVDYIYHVTDGKIRMWGNAKDAINNSFG 60
Query: 61 GLATVYKNTPSFKPQLGDVAVYTNGQ---YGHIQCVLS GNLDYYTCLEQNWLGGGF 113
G ATVYKN P+F+P+ GDV V+T G YGHI V + G+L Y T LEQNW G G Sbjct: 61 GTATVYKNYPAFRPKYGDVVVWTTGNFATYGHIAIVTNPDPYGDLQYVTVLEQNWNGNGI 120
Query: 114 DGWEKATIRTHYYDGVTHFIRPKFSGSNS-KALETSK 149
E ATIRTH Y G+THFIRP F+ +S K +T K Sbjct: 121 YKTELATIRTHDYTGITHFIRPNFATESSVKKKDTKK 157
Score = 61.7 bits (147), Expect = 6e-09
Identities = 41/125 (32%) , Positives = 57/125 (44%) , Gaps = 8/125 (6%)
Query: 125 YYDGVTHFIRPKFSGSNSKALETSKVNTFGKWKRNQYGTYYRNENGTFTC-GFLPIFARV 183
YY+G T P +K + +T G W N YGTYY++E+ TF C I R Sbjct: 346 YYEGKTPV--PTWNQKAKTKPVKQSSTSG-WNVNNYGTYYKSESATFKCTARQGIVTRY 402
Query: 184 GSPKLSEPNGYWFQPNGYTPYNEVCLSDGYVWIGYNWQGTR-YYLPVRQWNGKTGNSYSV 242
P + P Y+ VC DGYVWI + G + ++PVR W+ N+ + Sbjct: 403 TGPFTTCPQAGVLYYGQSVTYDTVCKQDGYVWISWTTNGGQDVWMPVRTWD KNTDIM 459
Query: 243 GIPWG 247
G WG Sbjct: 460 GQLWG 464
>gi 1113675 I sp I P24556 I ALYS_STAAU AUTOLYSIN
(N-ACETYLMURAMOYL-L-ALANINE AMIDASE)
>gi| 79887|pir I I Q1147 N-acetylmuramoyl-L-alanine amidase (EC 3.5.1.28) - Staphylococcus aureus >gi| 153067 (M76714) peptidoglycan hydrolase [Staphylococcus aureus] Length = 481
Score = 118 bits (292) , Expect = 6e-26
Identities = 56/117 (47%) , Positives = 68/117 (57%) , Gaps = 1/117 (0%)
Query: 135 PKFSGSNSKALETSKVNTFGK-WKRNQYGTYYRNENGTFTCGFLPIFARVGSPKLSEPNG 193
P + SN + ++ V WKRN+YGTYY E+ FT G PI R P LS P G Sbjct: 365 PVATVSNESSASSNTVKPVASAWKRNKYGTYYMEESARFTNGNQPITVRICVGPFLSCPVG 424
Query: 194 YWFQPNGYTPYNEVCLSDGYVWIGYNWQGTRYYLPVRQWNGKTGNSYSVGIPWGVFS 250
Y FQP GY Y EV L DG+VW+GY W+G RYYLP+R WNG + +G WG S Sbjct: 425 YQFQPGGYCDYTEVMLQDGHVWVGYTWEGQRYYLPIRTWNGSAPPNQILGDLWGEIS 481
Score = 78.0 bits (189), Expect = 7e-14
Identities = 48/109 (44%) , Positives = 62/109 (56%) , Gaps = 6/109 (5%)
Query: 15 EGAGVDFDGAYGFQCMDLSVAYVYYITDGKVRMWGNAKDA-INNDFKGLATVYKNTPSFK 73
EG + D YGFQC D + A + + G + AKD N+F GLATVY+NTP F Sbjct: 18 EGKQFNVDLWYGFQCFDYANAG-WKVLFGLLLKGLGAKDIPFANNFDGLATVYQNTPDFL 76
Query: 74 PQLGDVAVYTNGQ YGHIQCVLSGNLDYYTCLEQNWLGGGF-DGWEK 118
Q GD+ V+ + YGH+ V+ LDY EQNWLGGG+ DG E+ Sbjct: 77 AQPGDMWFGSNYGAGYGHVAWVIEATLDYIIVYEQNWLGGGWTDGIEQ 125
>gi|l763243 (U72397) amidase [bacteriophage 80 alpha] Length = 481
Score = 118 bits (292), Expect = 6e-26
Identities = 56/117 (47%), Positives = 68/117 (57%), Gaps = 1/117 (0%)
Query: 135 PKFSGSNSKALETSKVNTFGK-WKRNQYGTYYRNENGTFTCGFLPIFARVGSPKLSEPNG 193
P + SN + ++ V WKRN+YGTYY E+ FT G PI R P LS P G Sbjct: 365 PVATVSNESSASSNTVKPVASAWKRNKYGTYYMEESARFTNGNQPITVRKVGPFLSCPVG 424
Query: 194 YWFQPNGYTPYNEVCLSDGYVWIGYNWQGTRYYLPVRQWNGKTGNSYSVGIPWGVFS 250
Y FQP GY Y EV L DG+VW+GY W+G RYYLP+R WNG + +G WG S Sbjct: 425 YQFQPGGYCDYTEVMLQDGHVWVGYTWEGQRYYLPIRTWNGSAPPNQILGDLWGEIS 481
Score = 83.5 bits (203), Expect = 2e-15
Identities = 50/115 (43%) , Positives = 65/115 (56%) , Gaps = 6/115 (5%)
Query: 9 EWIYKHEGAGVDFDGAYGFQCMDLSVAYVYYITDGKVRMWGNAKDA-INNDFKGLATVYK 67
EW+ EG + D YGFQC D + A + + G + AKD N+F GLATVY+ Sbjct: 12 EWLKTSEGKQFNVDLWYGFQCFDYANAG-WKVLFGLLLKGLGAKDIPFANNFDGLATVYQ 70
Query: 68 NTPSFKPQLGDVAVYTNGQ---YGHIQCVLSGNLDYYTCLEQNWLGGGF-DGWEK 118
NTP F Q GD+ V+ + YGH+ V+ LDY EQNWLGGG+ DG E+ Sbjct: 71 NTPDFLAQPGDMWFGSNYGAGYGHVAWVIEATLDYIIVYEQNWLGGGWTDGIEQ 125
>gi|4S74237|gb|AAD23962.l|AF106851_l (AF106851) LytN [Staphylococcus aureus] Length = 383
Score = 84.3 bits (205), Expect = 9e-16
Identities = 48/128 (37%) , Positives = 68/128 (52%) , Gaps = 7/128 (5%)
Query: 15 EGAGVDFDGAYGFQCMDLSVAYVYYITDGKVRMWGNAKDAINNDFKGLATVYKNTPSFKP 74
E G DFDG+YG+QC DL Y ++ ++ +G N+F A +Y NTP+FK Sbjct: 252 ENRGWDFDGSYGWQCFDLVNVYWNHLYGHGLKGYGAKDIPYANNFNSEAKIYHNTPTFKA 311
Query: 75 QLGDVAVYT---NGQYGHIQCVLSGNLD YYTCLEQNWLGGGFDGWEKATIRTHYYD 127
+ GD+ V++ G YGH VL+G+ D + L+QNW GG+ E A H Y+ Sbjct: 312 EPGDLVVFSGRFGGGYGHTAIVLNGDYDGKLMKFQSLDQNWNNGGWRKAEVAHKVVHNYE 371
Query: 128 GVTHFIRP 135
FIRP Sbjct: 372 NDMIFIRP 379
>gi|3767593|dbj|BAA33856.l| (AB015195) LytN [Staphylococcus aureus] Length = 383
Score = 84.3 bits (205), Expect = 9e-16
Identities = 48/128 (37%) , Positives = 68/128 (52%) , Gaps = 7/128 (5%)
Query: 15 EGAGTOFDGAYGFQCMDLSVAYVYYITDGKVRMWGNAKDAINNDFKGLATVYKNTPSFKP 74
E G DFDG+YG+QC DL Y ++ ++ +G N+F A +Y NTP+FK Sbjct: 252 ENRGWDFDGSYGWQCFDLVNVYWNHLYGHGLKGYGAKDIPYANNFNSEAKIYHNTPTFKA 311
Query: 75 QLGDVAVYT---NGQYGHIQCVLSGNLD YYTCLEQNWLGGGFDGWEKATIRTHYYD 127
+ GD+ V++ G YGH VL+G+ D + L+QNW GG+ E A H Y+ Sbjct: 312 EPGDLWFSGRFGGGYGHTAIVLNGDYDGKLMKFQSLDQNWNNGGWRKAEVAHKWHNYE 371
Query: 128 GVTHFIRP 135
FIRP Sbjct: 372 NDMIFIRP 379
>gi| 2764983 |emb|CAA69022.l| (Y07740) cell wall hydrolase Plyl87 [Staphylococcus phage 187] Length = 628
Score = 76.9 bits (186), Expect = 2e-13
Identities = 50/144 (34%), Positives = 68/144 (46%), Gaps = 18/144 (12%)
Query: 5 QQAKEWIYKHEGAGVDFDGAYGFQCMDLSVAYVYYITDGKVRMW GNAKDAINNDF 59
+Q +W G+GVD DG YG QC DL Y++ R W GNA+D + Sbjct: 12 KQWDWAINLIGSGVDVDGYYGRQCWDLP-NYIFN RYWNFKTPGNARDMAWYRY 64
Query: 60 KGLATVYKNTPSFKPQLGDVAVYTNGQY GHIQCVLS-GNLDYYTCLEQNWLGGGF 113
V++NT F P+ GD+AV+T G Y GH V+ Y+ ++QNW Sbjct: 65 PEGFKVFRNTSDFVPKPGDIAVWTGGNYNWNTWGHTGIWGPSTKSYFYSVDQNWNNSNS 124
Query: 114 DGWEKATIRTHYYDGVTHFIRPKF 137
A H Y GVTHF+RP + Sbjct: 125 YVGSPAAKIKHSYFGVTHFVRPAY 148
>gi|3287732|sp|O05156|ALEl_STACP GLYCYL-GLYCINE ENDOPEPTIDASE ALE-1 PRECURSOR >gi 11890068 I dbj |BAA13069| (D86328) ALE-1 [Staphylococcus capitis] Length = 362
Score = 73.4 bits (177), Expect = 2e-12
Identities = 47/117 (40%), Positives = 61/117 (51%), Gaps = 10/117 (8%)
Query: 132 FIRPKFSGSNSKALETSKVNTFGKWKRNQYGTYYRNENGTFTCGFLPIFARVGSPKLSEP 191
F++ GSNS TS N G +K N+YGT Y++E+ +FT I R+ P S P Sbjct: 252 FLKSAGYGSNS TSSSNNNG-YKTNKYGTLYKSESASFTAN-TDIITRLTGPFRSMP 305
Query: 192 NGYWFQPNGYTPYNEVCLΞDGYVWIGYNW-QGTRYYLPVRQWNGKTGNSYSVGIPWG 247
+ Y+EV DG+VW+GYN G R YLPVR WN TG +G WG Sbjct: 306 QSGVLRKGLTIKYDEVMKQDGHVWVGYNTNSGKRVYLPVRTWNESTG ELGPLWG 359
>gi I 79926 |pir I I A25881 lysostaphin precursor - Staphylococcus simulans >gi| 153047 (M15686) lysostaphin (ttg start codon) [Staphylococcus simulans] Length = 389
Score = 69.5 bits (167), Expect = 3e-ll
Identities = 48/133 (36%), Positives = 62/133 (46%), Gaps = 20/133 (15%)
Query: 131 HFIRPKFSGSNSKALETS- - -KVNTFGK WKRNQYGTYYRNENGTFTCG 175
HF R S SNS A + K +GK WK N+YGT Y++E+ +FT
Sbjct: 258 HFQRMVNSFSNSTAQDPMPFLKSAGYGKAGGTVTPTPNTGWKTNKYGTLYKSESASFTPN 317
Query: 176 FLPIFARVGSPKLSEPNGYWFQPNGYTPYNEVCLSDGYVWIGYNW-QGTRYYLPVRQWNG 234
I R P S P + Y+EV DG+VW+GY G R YLPVR WN Sbjct: 318 -TDIITRTTGPFRSMPQSGVLKAGQTIHYDEVMKQDGHVWVGYTGNSGQRIYLPVRTWNK 376
Query: 235 KTGNSYSVGIPWG 247
T ++G+ WG Sbjct: 377 STN---TLGVLWG 386
>gi|l26496|sp|P10548|LSTP_STAST LYSOSTAPHIN PRECURSOR
(GLYCYL-GLYCINE ENDOPEPTIDASE) >gi | 79927 |pir| | S01079 lysostaphin precursor - Staphylococcus simulans bv. staphylolyticus >gi | 581744 | emb| CAA29494 | (X06121) lysostaphin (AA 1-480) [Staphylococcus simulans bv. staphylolyticus] Length = 480
Score = 69.5 bits (167), Expect = 3e-ll
Identities = 48/133 (36%), Positives = 62/133 (46%), Gaps = 20/133 (15%)
Query: 131 HFIRPKFSGSNSKALETS- --KVNTFGK WKRNQYGTYYRNENGTFTCG 175
HF R S SNS A + K +GK WK N+YGT Y++E+ +FT
Sbjct: 349 HFQRMVNSFSNSTAQDPMPFLKSAGYGKAGGTVTPTPNTGWKTNKYGTLYKSESASFTPN 408
Query: 176 FLPIFARVGSPKLSEPNGYWFQPNGYTPYNEVCLSDGYVWIGYNW-QGTRYYLPVRQWNG 234
I R P S P + Y+EV DG+VW+GY G R YLPVR WN Sbjct: 409 -TDIITRTTGPFRSMPQSGVLKAGQTIHYDEVMKQDGHVWVGYTGNSGQRIYLPVRTWNK 467
Query: 235 KTGNSYSVGIPWG 247
T ++G+ WG Sbjct: 468 STN---TLGVLWG 477
>gi|3287967|sp|P10547|LSTP_STASI LYSOSTAPHIN PRECURSOR
(GLYCYL-GLYCINE ENDOPEPTIDASE) >gi| 2072411 (U66883) lysostaphin [Staphylococcus simulans] Length = 493
Score = 69.5 bits (167), Expect = 3e-ll
Identities = 48/133 (36%) , Positives = 62/133 (46%) , Gaps = 20/133 (15%)
Query: 131 HFIRPKFSGSNSKALETS---KVNTFGK WKRNQYGTYYRNENGTFTCG 175
HF R S SNS A + K +GK WK N+YGT Y++E+ +FT
Sbjct: 362 HFQRMVNSFSNSTAQDPMPFLKSAGYGKAGGTVTPTPNTGWKTNKYGTLYKSESASFTPN 421
Query: 176 FLPIFARVGSPKLSEPNGYWFQPNGYTPYNEVCLΞDGYVWIGYNW-QGTRYYLPVRQWNG 234
I R P S P + Y+EV DG+VW+GY G R YLPVR WN Sbjct: 422 -TDIITRTTGPFRSMPQSGVLKAGQTIHYDEVMKQDGHVWVGYTGNSGQRIYLPVRTWNK 480
Query: 235 KTGNSYSVGIPWG 247
T ++G+ WG Sbjct: 481 STN---TLGVLWG 490
>gi I 3341932 I dbj |BAA31898.l| (AB009866) amidase (peptidoglycan hydrolase) [bacteriophage phi PVL] Length = 484
Score = 68.3 bits (164), Expect = 6e-ll
Identities = 52/150 (34%), Positives = 71/150 (46%), Gaps = 17/150 (11%)
Query: 3 SQQQAKEWIYKHEGAGVDFDGAYGFQCMDLSVAYVYYITDGKVRMWGNAKDAINNDFKGL 62
++ QA++W G + D YGFQC D + + + I G+ R+ G I D K Sbjct: 4 TKNQAEKWFDNΞLGKQFNPDLFYGFQCYDYASMF-FMIATGE-RLQGLYAYNIPFDNKAR 61
Query: 63 ATVY KNTPSFKPQLGDVAVYTN GQYGHIQCVLSGNLDYYTCLEQNWLGGGF- - 113
Y KN SF PQ D+ V+ + G GH++ V S NL+ +T QNW G G+ Sbjct: 62 IEKYGQIIKNYDSFLPQKLDIWFPSKYGGGAGHVEIVESANLNTFTSFGQNWNGKGWTN 121
Query: 114 DGW--EKATIRTHYYDGVTHFIRPKF 137
GW E T HYYD +FIR F Sbjct: 122 GVAQPGWGPETVTRHVHYYDDPMYFIRLNF 151
Query= pt| 110882 44AH DORF012 Phage 44AHJD ORF | 8391-8813 | 3 1 (140 letters)
>gi|140528|sp|P2481l|YQXH_BACSU HYPOTHETICAL 15.7 KD PROTEIN IN SPOIIIC-CWLA INTERGENIC REGION (ORF2) >gi|322189|pir| |B44816 orf2 5'of autolytic amidase - Bacillus subtilis >gi| 142801 (M59232) open reading frame 2 [Bacillus subtilis] >gi | 1217874 | dbj |BAA06959 | (D32216) ORF121 [Bacillus subtilis] >gi| 1303767 | dbj |BAA12423 | (D84432) YqdD [Bacillus subtilis]
>gi|2635036|emb|CAB14532 I (Z99117) alternate gene name: yqdD; similar to holin [Bacillus subtilis] Length = 140
Score = 80.4 bits (195), Expect = 6e-15
Identities = 45/130 (34%), Positives = 67/130 (50%), Gaps = 3/130 (2%)
Query: 4 VKFRFTDSEAFHMFIYAGDLKLLYFLFVLMFVDIITGISKAIKNNNLWSKKSMRGFSKKX 63
+ F D ++F G +K L L VL +D++TG+ KA K L S+ + G+ +K Sbjct: 8 INFETLDLARVYLF---GGVKYLDLLLVLSIIDVLTGVIKAWKFKKLRSRSAWFGYVRKL 64
Query: 64 XXXXXXXXXXXXXXXXXXKGGLLMITIFYYIANEGLSIVENCAEMDVLVPEQIKDKLRVI 123
G L T+ +YIANEGLSI EN A++ V +P I D+L+ I Sbjct: 65 LNFFAVILANVIDTVLNLNGVLTFGTVLFYIANEGLΞITENLAQIGVKIPSSITDRLQTI 124
Query: 124 KNDTEKSDNN 133
+N+ E+S NN Sbjct: 125 ENEKEQSKNN 134
>gi I 41266311 dbj |BAA36651.l| (AB016282) ORF45 [bacteriophage phi-105] Length = 135
Score = 76.1 bits (184), Expect = le-13
Identities = 44/115 (38%), Positives = 61/115 (52%), Gaps = 4/115 (3%)
Query: 21 GDLKLLYFLFVLMFVDIITGISKAIKNNNLWSKKSMRGFSKKXXXXXXXXXXXXXXXXXX 80
G++K L + VL +DIITG+ KA K L S+ + G+ +K Sbjct: 17 GEVKYLDLMLVLNIIDIITGVIKAWKFKELRSRSAWFGYVRKMLSFLWIVANAIDTIMD 76
Query: 81 XKGGLLMITIFYYIANEGLSIVENCAEMDVLVPEQIKDKLRVIKND TEKSD 131
G L T+ +YIANEGLSI EN A++ V +P I D+L VI++D TEK D Sbjct: 77 LNGVLTFATVLFYIANEGLSITENLAQIGVKIPAVITDRLHVIESDNDQKTEKDD 131
>gi|l41088|sp|P26835|YNGD_CLOPE HYPOTHETICAL 14.9 KD PROTEIN IN NAGH 3'REGION (ORFD) >gi | 1075967 | pir | | S43905 hypothetical protein D - clostridium perfringens >gi| 455154 (M81878)
ORF D [Clostridium perfringens] Length = 132
Score = 60.9 bits (145), Expect = 4e-09
Identities = 38/127 (29%) , Positives = 63/127 (48%) , Gaps = 3/127 (2%)
Query: 1 MNEVKFRFTDSEAFHMFIY-AGDLKLLYFLFVLMFVDIITGISKAIKNNNLWSKKSMRGF 59
+N +K+ +1+ A D+ L+ L V +F+D +TG+ K K+ L S +RG
Sbjct: 5 INYIKWGIVSLGTLFTWIFGAWDIPLITLL-VFIFLDYLTGVIKGCKSKELCSNIGLRGI 63
Query: 60 SKKXXXXXXXXXXXXXXXXXXXKGGLLMITI-FYYIANEGLSIVENCAEMDVLVPEQIKD 118
+KK + I ++YI NEG+SI+ENCA + V +PE++K
Sbjct: 64 TKKGLILWLLVAVMLDRLLDNGTWMFRTLIAYFYIMNEGISILENCAALGVPIPEKLKQ 123
Query: 119 KLRVIKN 125
L+ + N Sbjct: 124 ALKQLNN 130
>gi 12293160 (AF008220) YtkC [Bacillus subtilis]
>gi|2635548|emb|CAB15042| (Z99119) similar to autolytic amidase [Bacillus subtilis] Length = 134
Score = 36.4 bits (82), Expect = 0.099
Identities = 25/109 (22%) , Positives = 41/109 (36%)
Query: 17 FIYAGDLKLLYFLFVLMFVDIITGISKAIKNNNLWSKKSMRGFSKKXXXXXXXXXXXXXX 76
F + G L LM ++ I+ K + L KK KK Sbjct: 20 FFFGGFQYSFLILLSLMAIEFIΞTTLKETIIHKLSFKKVFARLVKKLVTLALISVCHFFD 79
Query: 77 XXXXXKGGLLMITIFYYIANEGLSIVENCAEMDVLVPEQIKDKLRVIKN 125
+G + + 1 +YI E + IV + + + VP+ + D L +KN Sbjct: 80 QLLNTQGSIRDLAIMFYILYESVQIWTASSLGIPVPQMLVDLLETLKN 128
>gi| 1181973 |emb|CAA87743.l| (Z47794) holin protein [Bacteriophage CP-1] Length = 134
Score = 31.3 bits (69), Expect = 3.3
Identities = 27/88 (30%), Positives = 36/88 (40%), Gaps = 5/88 (5%)
Query: 29 LFVLMFVDIITGISKAIKNNNLWSKKSMRGFSKKXXXXXXXXXXXXXXXXXXXK--GGLL 86
LF L+ D ITG KA K S ++G K G +L
Sbjct: 18 LFALILFDFITGFLKAWKWKVTDSWTGLKGVIKHTLTFIFYYFVAVFLTYIHAMAVGQIL 77
Query: 87 MITIFYYIANEGLSIVENCAEMDVLVPE 114
++ I Y A LSI+EN A M V +P+ Sbjct: 78 LVIINLYYA---LSIMENLAVMGVFIPK 102
Table 21
Phage 182 complete genome sequence. 17833 nucleotides.
1 tagaatattg tcataaaaca caaacataat aatgcatatt attgtttaca aatatgtaat ttcgtgatat 71 aatatatttg taagttaaag gaggtgacaa aagaacaaat cataaatgct ttagaaattg caaaaactat 141 tggaggaaaa ataatgaaat attcactaca acaaatagat gaaattaaat caacaatttt cagaattaga 211 ttaaaaaggc atgaactaga ggaattggtg gacgaagtaa acgatattgc taaagatccg gaggaaagat
281 atcttttatc gttttattac acagaagaag aacgtttgtt tgaaattccc tctgcaagat taatagatta
351 ttacaacgaa aagatcacaa atctgaaatc ggaaatcata tcactcgaaa aaagattaca aaaactagta 421 aaataattac acaaaaagct ttacaaatat aacacatcat gttatactaa aagagtagta agggaacgga 491 aaatacctta cttcacacct caatcattct tatcaaaata caaaaggagg gaaaataatg ggtcgaaaac 561 taatgcaacg aaacgtaaca tcaactaaag tagaattctc agaagttatc gtacaagatg gagcgccaac 631 aattgtacca tgcgaaccag ttgtcttaac aggaaaactt tcagaagaaa aagctttatc agcgatcaaa 701 cgtaaaaacc ctgataaaaa cgtagttgta acaaatgttt cacatgaaac agcgctttac acaatgccag 771 tcgataaatt tatcgagtta gcagacaaat caacacaagc ctaataaaaa caaaactaaa acaaaacaga 841 ggagattata atcatggaaa tcgtaaaaag cacatttgac acacaaacac cagaaggaat gttacaagta 911 ttcaatgcca caaacggggc ttcaattccg ttacgtaacg caattggcga agtactagaa ttgaaagata 981 ttctagttta ctcagacgaa gtttctggtt ttggtggagc cgaaccatca caagcagaac tagtcgcttt 1051 cttcacagaa gatggtaaaa cttatgcggg tgtatcagca gtagcaacaa aatcagctaa aaacctaatt 1121 gatatgatga ctgctaaccc tgacatcaaa ccaaaaattt cttttgtcga aggaaaatca aacggtggac 1191 aaaaatttgt aaatctacaa gtggtttcac tgtagcataa aaatacagga atctagtaag ccacttagcg 1261 aatctcgcta ggtggttttt attatgtttc tacattgagg tgtgtagaat tgaccgtaag aatatcaaag 1331 aatgatagag ccaagttaga gaaaatctac ggtaaatcta acaaagctcg taaaaaatac aatcgtttaa 1401 gacaaaaagg agttgaggaa aggcaacttc caactgttcc aacatcaaag aaaagactta ttgactacgt 1471 aaaatcaaca aatatgagtc gtagtgattt taacaagatg ttagacgagt tggtagattt tgcacaacct 1541 tacaacgaga attacatttt tgagatcaac aagcgaaatg ttgcaatctc aagagcgcaa atcaaagaag 1611 cgcaaattaa aacagagcaa gctcaaaaag cgaaagaaga acactacaaa gagcttaaca aagttgaagt 1681 taagaagccc acagaaaaca caattgtcac accaactatt ttaacagagt taggtgctga cttacctttt 1751 caagcaatac cagattttaa tattgacgct ttcacttctc cagaaggagt tcagtcttat ttagaaaata 1821 taggaaaaca agacgaacaa tattttgacg aaagagacca actttattac gacaatttca gacaagcgat 1891 gtttactatt ttcaattcag acgctgacga tattgttcgt ttacttgact caatggggct tgatctattt 1961 atgaaaacat atgttagtaa cttcttagac atgaaccttg actacattta tgacgaagca gaagtacaac 2031 agaaaaaaga acaagtttac agtaagattg caaaagtgat cgagtctgaa acaggtggag aagtcccctc 2101 atataacccc acgaagaaca tcacaattaa ttcagaaaca ggagaagaat tatgattaag aaatatactg 2171 gcgactttga aacaacaact gatctcaacg attgtcgtgt atggtcgtgg ggcgtatgcg atatagacaa 2241 cgttgacaat atgacgttcg gtttagaaat cgattctttt tttgagtggt gtaaaatgca aggcagcaca 2311 gacatttatt tccacaacga aaaatttgac ggagagttta tgctttcatg gttattcaaa aatggtttca 2381 aatggtgtaa agaagcaaaa gaagatcgaa cattctccac actcatatca aatatgggtc aatggtatgc 2451 tttggaaatt tgttgggaag ttaattacac aacaacaaaa tcaggtaaaa cgaaaaaaga gaaatctcga 2521 acaataattt atgatagcct taaaaaatat ccttttccag tgaaacaaat tgcagaagct tttaattttc 2591 ctataaaaaa aggcgaaata gattatacaa aagaaagacc tattggttac aaaccaacaa aagatgaatg 2661 ggagtattta aagaacgaca ttcagattat ggcgatggca ttaaaaattc aattcgatca aggactaact 2731 cgaatgacta gaggaagcga cgctttaggc gattacaaag attggctaaa agctacacat ggaaaatcaa 2801 ctttcaaaca atggtttcct attttgtctt tagggtttga taaagactta cgtaaagcat acaaaggcgg 2871 cttcacttgg gtaaacaaag tttttcaagg gaaagaaata ggtgacggca ttgtctttga tgtcaactct 2941 ttgtatccct ctcaaatgta cgtaagacct ttaccatatg gaacacctct attctacgaa ggagaataca 3011 aaccgaacaa cgactatccg ctgtacattc aaaatatcaa agtaagattc cgtttaaagg agggttatat 3081 tccaaccatt caagttaagc aaagttcatt attcattcaa aacgaatatc ttgaatcaag tgtaaacaag 3151 ttaggagttg acgaattaat cgatcttact cttacaaatg ttgacctaga attatttttt gaacactacg 3221 atattttaga gatacattac acttacggat atatgttcaa agcttcttgt gatatgttca aaggctggat 3291 cgataaatgg atcgaagtaa agaacaccac cgaaggggct agaaaagcta acgccaaagg tatgttaaat 3361 agcttgtatg gaaagttcgg aacaaaccct gacattacag gaaaagtgcc ttacatgggc gaggacggca 3431 ttgttcgatt gacactagga gaagaagaat taagagatcc tgtttatgtt ccgcttgcta gttttgtgac 3501 ggcttggggt agatatacta ccattacaac cgctcaaaaa tgttttgatc gcattattta ttgtgataca 3571 gatagcattc atctagtagg aacagaagtt ccagaagcaa tcgatcactt ggttgatcct aaaaaacttg 3641 gttattgggg gcatgaaagc acatttcaac gagcaaaatt cattcggcag aaaacatacg tagaagaaat 3711 tgatggcgaa ttaaatgtaa agtgtgctgg tatgccagat cgaataaaag agattgtaac ttttgacaat 3781 tttgaagttg gtttttcaag ctatggaaag ttgctaccta aaagaacaca aggtggcgtg gtattagtag 3851 acacaatgtt tacaatcaaa taaggaggac taataatgga actatataaa gcaatgttta tcgtacgtga 3921 tgaaggtact attgacggtt acgatactga acactatgta gatatttctt tacatgactt tgaagaaata 3991 tatggaaaag aaacacgtga aattgaagca gtaacattag taaaaacagg aaatttaaaa aaataaatta 4061 tttacatcct ttgcaaagta tggtaaaata ttcttgtgat agttgacaag agtcaaattt ggcgagattg 4131 ggcgaatgta cacgtgaaat atcgtgcgct cccgttaagt tatggacaca taaacgtttt gaccgtcaac
4201 caatcgcaaa aaccttttag gagtagccct taaatgtggc tactcttttt tgtgtttcac agaattatgt _. - "
4271 ttcacgtgaa acagttttta tggtataata gaatcaaaag gaggtggaga ttatggaaat taaagaacatr
4341 gaatcaattt taaatggtat tcttgaaagt gtcacagacg gtgaagcaag atcaaagatt gtagaacatc
4411 ttgaagcatt gcgagaagac tacggagcaa caactgaagc tttgacatca gcaaatagca cacttgaaaa
4481 gttaaagaaa gataacgaag cgttggttat ttcaaactca aaattgttcc gagaacgagc gatcgtagaa
4551 ccagcagaaa ataacgaacc agaaacagac cagaatatta cactagacga tttaggaatt taaggaggaa
4621 aaaacatggc tgacaaaatc acagaacaag atgttcttcg tgccacaaat gtagaaacac cagtacaatt
4691 aatgactgct atttataata gttcatcatc tctttttcag gcgaacgtac ctatgccaaa tgcagataac
4761 atcgaagcgg ttggtgcagg gatcacacgt ttagacgtag taaaaaacga atttatttca actttagttg
4831 accgtattgg taaagtagtt atccgataca aatcttggcg taaccctttg aaaatgttta aaaaaggaaa
4901 catgccttta ggtcgaacga ttgaagaaat ttttgttgac attgcacagg aacataagtt caaccctgac
4971 gagtctgtta caggggtatt taaacaggaa gttcccgatg taaaaacatt gttccacgaa attaatcgtg
5041 aaggttacta caaacaaacg atccaagaag catggttaga aaaagcattt acttcatggg ataatttcaa
5111 tagtttcgtt gctggtgtaa tgaacgcttt atacacaggt gacgaagtaa gcgaatttga atacacgaaa
5181 ttattaatag caaactacca agaaaaagag ctattcaaag agatcgaaat tggcgaaatt actgaatcaa
5251 atgcaaaaga atttatccgt aagatcaaat caacctctaa caaattagaa tttatgagtt ccgcttacaa
5321 cgctcaagga gttaaaacat ctacctcaaa atctgatcaa tacgttatta ttgacgccga cacagacgca
5391 accattgacg ttgacgtttt agcagcggca ttcaatatga gtaaaactga ctttgtagga cacaaaatcg
5461 ttattgatga gtttcctaaa aaagaaggcg aagaatcgtc aaatattgtg gcagttattg tagatagtga
5531 atggtttatg atctacgaca aattgtacaa aacaacaagt ctatacaacc ctgaagggtt atattggaat
5601 tattggttgc accaccacca actatattct acttctcaat tcgggaacgc tgttgctttt gttaaatcag
5671 caacaaaacc tgtcacaaaa gttgcttttg caagtgcaac aactagtgtt gttaaaggat catctaaaga
5741 tatcgcattg acatttacac cagtagaagc aacaaaccaa caaggagaag ttgtttcatc agcaccagca
5811 ttggttaagg caaccgtaaa acaaacagca ggtaaagcga ctgccgtaac cgtagaaggc ttagaagtcg
5881 gtcaatcatt agtaacattc acagctatcg gaggtcaaca agcaacggtt cttgttacgg ttacttctga
5951 ctaaggagga caattatggc aagaaggtat acaaatgtaa aattgttggc taacgtgcct tttgataaca
6021 cctatacaca cacaagatgg tttaaaactc aacaggaaca ggaatcgtac tttaattcgt ttcctgttct
6091 taacgagaat agagattgtt cttatcaaag ggatacacaa ctcgggggag tttttagagt agataaacac
6161 aaagacgcct tatatgcttg taactatctc atctttaaaa acgaagaaac ttatcctagt aaatggcagt
6231 atgcctttgt tactgatatt gaatataaga atgacaacac aagtttcgtt acctttgaaa ttgatgtttt
6301 acaaacttat cgtttcgata ttggtatacg agaaagtttc attgcaaaag aacaccctca actttattat
6371 tcgaatggaa tacctttcat taatacaatt gaagagtcgc ttgattacgg tagagaatac acaacaacaa
6441 atgtaacaac ttttcatcct aacgatggag tcaattttct tgttattcta acaagtgaag caatgccagt
6511 tggagataag gaagataaat caggaggatc aatagtaggt ggcccatctc ctttttccta ttatttactt
6581 cctatcaatt caagtgggga ggtatacaaa ccaaatgggg caggcaatgc taattttgga gagtacatgg
6651 cgtttcttac aacgaaagaa ccttttttaa ataagatagt cgggatgtat gtaacgtcgt atacaggtat
6721 accattcatt gtggatcacg cgaacaaaac ggtaaggtat aatgcaggag gttcttataa gatcatgctt
6791 ccaacctacg ctagtgatcc aacaggaaca atgaaaacat tcgctttctt ttgtgtaaaa gaagcaagaa
6861 cattcgtacc taaaagaatt gatcttgtag ggaacgtgta taactacttt agagaagctt ttccgtttaa
6931 tgttaaggaa tcaaaactat ttatgtatcc ctattgttta atagaaatta cagatacaaa aggacatgta
7001 atgactttaa gacctgaata tcttacaggt ggtaaattga gtgtatatgt aaaaggttcg ttaggaattt
7071 ctaataaagt gatgatcgag ccgattgatt atgatgtaag taactcaacc attattacca atttaagtga
7141 caagatgtta atcgataatg atcctaacga tgtaggagtt aaatctgact atgcttctgc attcatgcaa
7211 ggaaacaaaa actccttgat tgctcaagag caaaacattc gcaatacttt cagacatggt atgggaaaca
7281 gtgcaatgag tacaggagga gcgatctttt cagccttagc aagtaacaac ccttttgttg gtttgactaa
7351 catcatggga gcaggacaac aagtaaacaa ctatgtttct gaaaaagaaa acggtttgaa cctcttggca
7421 ggtaaagtgg cagatatcga aaatattcca gataatgtaa cacagcttgg atcaaactta tctttcacaa
7491 caggaaactt tcaaaactat tatcaattgc gcttcaaaca aattaaatat gagtatgcaa caagacttga
7561 tcgttacttc tcaatgtatg gcacaaagag caatcgagta gctacaccaa acttacaaac aagaaaagca
7631 tggaatttca ttaaattaaa agaaccaaat attgtaggca caatgagtaa cgatgtatta acacgtgtga
7701 aacaaatttt tagtgcaggc gttacgcttt ggcatacgaa tgatgttttg aattataacc aagacaacgg
7771 agatgtatag gaaggaggaa taagatgagt agacgaaaag gtgcaggact tgctagaaat aaccgttata
7841 cagcaaaaag cagaccttat ccaaatgaac cctattcaag tgatgtagaa gaaatcagct actatgaaca
7911 ttatcgtaga caactcacgc tccttacgtt tcagttgttt gaatgggaaa atttgccaaa atcaattgac
7981 cctcgttatt tagaaattgc tttacacact aatggttatc ttggtttctt taaagaccct acacttgggt
8051 tcatggtttg cgcaggggca gaagatggtc aaatcgatca ttatcacaac cctattttct ttacagcaaa
8121 cgaagcaatg tatcacaaga gatatcctgt tttaagatat gatgatgatg atgataaatc aaaatgtatc
8191 atgttgtata ataatgactt gaaagttcct acgttaccaa gtttacatcg ttttgcttta gatatggcgg
8261 acataaacca gatatcacga gtgaatcgaa gagcgcaaaa aacacctgta attattcaaa ctgatgaaaa
8331 gaaatacttc tcattgctac aagcttataa ccaaattgac gaaaataatc aggctgtttt tgtggataaa
8401 gatatggagt ttgacgaatc ttttaatgta tggcaaacaa atgctccata tgtagtagat aaactacgat
8471 cagaattgaa cgaagtatgg aatgaagtgt taacttttct aggtatcaac aatgctaacg tagataagac
8541 tgcacgtgta caaacatcag aagtcttatc taacaatgaa cagattgaaa gttcaggtaa catcttgtta
8611 aaatcaagaa aagagttttg cgatcgtgta aatcgtgtct ttggcgatga acttgacgga aagattgacg
8681 tgaagtttag aacagacgcc gttcgacaat tacaactggc ggcaggtcaa tcaaaaaaag accagatgag
8751 tggagggttg ccaagtgcta cttaaacgtt atattgaaag tttcacttat taccaacctg aattatctcg
8821 aaaagaacgt attgaagttg gccgaaaaca attgtttgat tttgattatc cgttttatga cgaaacaaaa
8891 cgagcagaat ttgaaacaaa atttatcaat cacttttact tgagagagat aggctcagaa acgatgggat
8961 catttaagtt taatcttgac gaatatttaa atctaaacat gccctattgg aataaaatgt tcctatcaaa
9031 tcttgaagag tttccgattt ttgatgacat ggactacacc attgatgaga aacagaaatt gttaaatgag
9101 attgatacaa acatcaaagc gaatcgtgat gaatcgaaga accaaacgaa gcaagtagat caaacagaca
9171 acagaaacaa aaatacacgt gacacaggaa caaccgattc tttctcaagg aacacttata cagacacccc
9241 tcaaaaagat ttgagaattg ccagcaatgg agatggaaca ggtgtaatca attatgcaac aaatatcaca
9311 gaagatttga gtaaagaaac aacaagctcc acaggcgttg aaacaaacaa cgacaaaaca aatcaaaata
9381 cacgaagcaa tgcttctgaa aaagaaacaa agaacacaga cattaataaa gatcaaaatc aaaccaaaga
9451 tacgattaca cgatataaag gtaaaaaggg aaacactgat tatgctgact tactcgaaaa atatcgtaga
9521 agtgttttga gaattgagaa aatgatcttt agagaaatga acaaggaagg cttatttctc cttgtttatg
9591 gagggaggta gcaacaatgg tagattttaa ccccgacaag cggtttgacg gtttacccgc tgtattcaaa -
9661 gaacgcttta gcaaatatcc tcatactgaa tacagatatg aattactatt agatgaagaa gtatcggctt
9731 taattgccta tctgaatgaa gttggtgctt tagttaatga tatgagtggt tatttaaatt actttatcga
9801 acattttgtt gagaagttag aagagatcac aaatgacaca ctcaaaaaat ggttgtctga tggtacgtta
9871 gaaaatttaa tcaatgatac tgtttttgca aattatatca aagaaatcaa aagattacaa atcttggttg
9941 ctgaaacacg tgctaacagt gtgaatattc ttttgacaaa aaataaaccg gatgttgctg atgatcgaac
10011 attttggtat aagattcaac gcgacaatac tgattatgga gccgatccta ttgacacgtt acgtattgtt
10081 gcaatcaata aagttagtgg ctggaatacc gctacaggag atatttatct taacattaaa ggaacggagg
10151 gtgtataatg gcagacatta gaacacaact aacaagtgaa gatggatcag acaatttatt tccaatttca
10221 aaagccgtta atattatgac taatagcggt acgaatgtag aaggagaatt gggtacactc aaacaaaatg
10291 acgaaacaat gaatacctca gttcaaaatg ctgtagttac tgccaatcaa gcaaaagatt ctgtagctga
10361 attaaatgta aatgttggta aactaaccaa tcgaataaca acattagaga gtacagtggc taatcttgat
10431 ggtattcgtt atgtagaggt gtaatatggc agataaaaat attcaaatgc aggataaaga tcataatcgt
10501 ttaatgcctg ttacaattgc taaaaatgtt ctaacaggcg actctaatct tgaattagtt aatgctgaaa
10571 taagaggtaa cgctagtgaa gctaaaacac ttgcacaaca agctaaagaa actgctgctg gtttgtcaac
10641 agaaattgac acagtaacat caaccgcaaa tcaagcgttg acgaaggctg gtacagcaca acaaaccgca
10711 gaacaagcga aaacaacagc aaacagtatc agcgcagttg caacggcagc taaaaacaca gctgattcag
10781 cacaaaaaag tgcaactgat ctagctgttc gagtaagcag tttagaggac acagcaatac aatatactgt
10851 attaccatag gaggaaaaat aatggcaaat aaaaatattc aaatgaagga tagcaatgac aataatttat
10921 atccaagtgt tcgagcagaa aacttgttag atttgaccag tcgtgctgaa ttaacaatga caaattgtca
10991 attatatgca gctggtgata aaacaaatgc aatctcttat ctcggtgcag taggtatgct cgaaggtatg
11061 ataaagttta ctgaaagttt gacaaaccct gtgatcacaa cgctaccaga aggttttaga ccaataagaa
11131 caaaacgtat tggttgtttc gcaaaatatt acacaccaaa tccaacagat acaaaagaaa tggtttatgt
11201 atcaatcaca cctgatggca aagtaactgt aaatgacaat gtaggtaaaa tcgaatatct atccctagat
11271 aattgcgttt tccctctaaa ataaggaggt tcatatggaa gaacgaattg atattcaaat gaacaagatg
11341 aaagaagaaa atcaaaagaa ttacctattg caccctgaaa cgaacccgaa acaagttgtt tttgatgaaa
11411 cattgcatgg aaatgaaaat caggagagtt tcaacaattt tgttgacaca agaaaaatga caactacaat
11481 tgatgtaagt gcttatgggg ttatcgctga cggtgtaaca gattgtacac caatattaaa taaattactt
11551 gaagaaaaaa gcgaaatggg tatcactttt tattttcctc cttgtgaacg tgattcatat tatcgctttg
11621 ctaacaccat tgaattgaaa cgtgatgtac ctgtagttac tttcttagga tcgggagaaa cgacattaaa
11691 gtttgaaaca atgacggcat ttaatgtaaa catcgaaagt ttcaatattg atggttttgc attatggttg
11761 ccacaaggcg ctcaaagtgg taaaggaatt ttctttaatg atac cgcaa ttacaatcgt tttgactttg
11831 atttgtttgt tcgtaactgt actttaaatg aaggaacgta tgttgttgtt gctagaggta gaggggttac
11901 atttgaaaat tgtctattct ctaatatctc tcaagcaatt atcaaaacag cttttcccga tgtaaatggt
11971 atgtggcaag ggaacgatat caatactagg ggtacaggtt ttagaggttt ctttgtgaaa aacaaccgta
12041 ttcatttttg tacagcgatc attatcgaca atgacgatga ttatcagaat gtaattaatt tctgtgaaat
12111 ttctggtaac acaatcgaag gtggcgtaag ttattatcga ggatatgcgc ataacttgca tgtccaaaac
12181 aacaaccatt ttctagcata cggaaataga aacgctttgt ttgagtttca agatgtggat caagcttata
12251 ttgatgtaga tgtttattgt cgtaactcac aagtcgaggg aatgaatagt acagctattt cacgtttaat
12321 tgttgtttac ggacattacc gaaacttaaa gattacaggt aaattatatc gttgtcaagg acatgttatc
12391 acgttgtatg gcggtggcgt taatttctat tgtgacttga tggcacaaga agcacctttg acggacggtt
12461 accggtttat tcaaacggct gacaatcgag ttaactatga tgggtttgtt gttcgtggtt tgtctaattc
12531 aacaaaagta aatacaccaa tgatctataa agcacctcag actgttttct ataatcgtag aatcgatcat
12601 gtgctaacag gtccaaatgc aagtaatgta tataactagg aggatatgag atggcaactc ttacaaatga
12671 acaaatagct agaggacaaa caatcgctaa aatactttca aaatatggct ataataaaaa ttcacaagta
12741 ggagttgtcg ccaatctcca ttgggaatcg gctggtttga acccgaacag caatgaatat ggtggaggcg
12811 gatatgggtt aggtcaatgg acgcctaaaa gcaatcttta tcgccaagca caaatttgtg ggttgtctaa
12881 tgctaaagct gaaacgttgg aaggtcaagc agagatcatc gctcaagggg ataaaacagg tcaatggatg
12951 gataatacac ctgtttcttc tgcaggttat actaaccctc agaccctttc agcatttaaa caatctgcaa
13021 atattgatgt tgctacaatt aattttatgt gtcactggga acgccctggt aaacttcata tcgaagaaag
13091 acttgatctt gcacaagctt atagtaagca tattgacggt agcggtggcg gtggcgtaaa acgttgctat
13161 ggaaccccaa tcaagaatac aaatcttgat cctaaaagtt tcatgagtgg acaacttttt ggcacgcatg
13231 caggaaacgg cagaccaaat aatttccatg atggtttgga ctttggttca attgatcacc ctggcaatga
13301 aatgattgca tgttgcgatg gaacagtaac acatgttgga acaatgggag cattaagagc gtattttgtg
13371 ataaatgatg gtacttacaa tatcgtttat caagaattta gttataacca gtcaaata a aaggtaaaag
13441 ttggcgacaa agttaagaac ggacaagttt gcgcaatacg tgacgcggat catttacatt taggttttac
13511 taaaaaagat tttatgactg cgttaggatc ttctttcata gatgatggaa catgggaaga ccctttgaag
13581 tttttagggc aatgttttgg agatggagat actggcggag ataatgacga taacaataag gataaaaatg
13651 atcttattta tctattgcta tccgatgcct tgaatggttg gaaattttaa taaggagaaa aaggtatgat
13721 agaatatatc acacaatggt tggcagatga taatcatctt gtttatggtt tgattatatg gttaatggtt
13791 gcaatgatta tcgattttgt gttaggtttt acaattgcca aatttaacaa ggaaatcgac tttagtagtt
13861 ttaaagctaa agcaggtatc attgttaagg tggcagaaat ggttttagtg gtttacttta ttcctgtagc
13931 agtaaaattc ggtgcagtag gtattacaat gtatataaca atgttggttg gtttgatttt atcagaaatt
14001 tatagtatac taggacatat ttcagatatc gatgatgata ataattggac tgattatgtt aagaagtttt
14071 tagacggaac actcaacaga aaggacgata ttaaatgatg aatggtattg atatctctag ttatcaaaca
14141 ggaattgatc tttcaaaagt tccatgcgat tttgtaaata ttaaagcaac aggcggaaca ggttatgtaa
14211 accctgattg tgaccgagca tttcaacaag ctttgtcttt aggtaaaaag attggtgtgt atcattttgc
14281 gcatgagagg ggtttagaag gtacacctca acaagaagcg caattctttt tagataatat taagggttac
14351 attggtaaag ctgttcttat tcttgacttt gaagggtcaa atcagaaaga tgtaaattgg gcgaaagcat
14421 ttcttgatta tgtttataat aaaacaggcg ttaaagcatg gttttatacg tatacagcaa acctcaatac
14491 aactgatttt tctagtattg caaaaggcga ttatggttta tgggttgctg aatatggatc aaatcaacca
14561 caaggctact ctcaaccagc gccacctaaa acaaataatt ttccaattgt tgcctgtttt cagtttacaa
14631 gtaaaggacg tttaccagga tacaacggca atcttgattt gaatgttttc tatggcgatg gtaatacatg
14701 ggatctgtat gtaggtaaaa aacaggatca aattgttcct cctgaaaata aaatatttga cgccacaagt
14771 gatgagttta ttttcactct tacaacaggt agcacaagcg tgttttattt tgacggagaa acgatctttg
14841 aattgtctga tccaacacaa ctcgatcata ttagaggaac atacaatcat gttcatggaa aagaaatccc
14911 atcaatggtg tggacacctg aacaatttga tatttactta aaaatgtatg aaaagaaacc agtatataaa
14981 taggagtgta tagtatgaca aatagcttag gcgttaaact tgaagagaaa aacttatact ataaccctaa
15051 caatgcttta ggttttaatt gcctaatgtt gtttgtaata ggcgcacgtg gtataggtaa aacttatggt
15121 tataaaaaat ttgttgttaa tcgctttatt aaacacggcg aacaatttat ttatttaaga agattcaaaa
15191 cagaacttaa aaagattcct caatttttca aaacaatggc gaaagaattt cctgatcata aacttgaagt
15261 aaaaggaaaa gaattctatt gtgatgataa attaatgggt tgggctgttc cacttagtac gtggggaatt
15331 gaaaaatcta atgaatatcc cgaagttcgt acaattttgt ttgatgagtt tttaattgag aaatcaaaaa
15401 tcacttattt accaaacgaa gctgaagcct tattgaacat gatggaaacg gttttccgaa gacgtacaaa
15471 tacaagatgt gttatgttga gtaatgcaac tagtgtagtg aacccttatt tcttgtattt caatctgcag
15541 ccagatttga ataagcgttt taatctatat caagatcgag gtatattgat tgaattgtgt gattcaaaag
15611 actttgcaga agtgaagaga gaaacacctt ttggtagatt gattcgtgga acagaatacg aagattttag
15681 tatcaacaat gagtttgtca atgatagtga tacgtttatt gaaaagagaa gtaaaaatag tagtttctta
15751 tgcgccattg cttttgaagg gaaaatcttt gggtattgga tagacgctga aacaggttgt gtctatgtga
15821 gttatgatta tcaaccaaat acaaatcatt tttatgcaat gactacgaaa gaccatgaag aaaatagatt
15891 gctgatgaaa aattggcgaa ataattatta tctttcaaca gtggcgaaag cattcaagaa tagttatctg
15961 cggtttgata acattgttat taagaattta cattatgatt tgtttaataa gatgaaaatc tggtaaccct
16031 attttagtag agctaccacg attagttcta ttacaatgat gaatagtaga taacatagta attgtagtct
16101 gcgatagttt tgttttggtt ctttggcgtt agtgattttt gctaacgcct ttttgtttgc ttttggatcg
16171 ggtgtgttaa tgtagacgaa atcttttctc atagttcttt ctccttatac agttttaata attccctgta
16241 aaatgtagct ataggacgtc catttctttc tattctaacg caattcacta tatccatttc taggtatata
16311 cggctatatt ttaatgcttt tgttaaggtg agaggttcgg ttttgtgtat caaaacctcc caaccatcta
16381 tataaaatac tgtgatatcg tatattggtt ccttgtagaa tgtagccatt attccacctc ctttaaatag
16451 ccttttggta tttgtaacgc taactgatag cgagaaccaa cttttacgta tgaagttact aatttcattg
16521 cctgacaata cttttcaaga atgttaaatt gactcgattc gggtaatagc gttgaatgag ttaacaaaag
16591 ttcggtgata tttatttccg gaacgtcgaa atcttgtaaa gtcccctcta tgatctctat tttttcattg
16661 tctgaaaggt tacgtttaca gtagaaacgt aaccattcaa ttagttcgcg gtgttctttg aatgttcgtg
16731 caatcatttt aattcctcct atttgtccgt aatttgttta tatccgtcat gtttcaattg ttccgcatag
16801 tgttcaacgc ttttcattga tttcgttatt gcgatattaa tgcaatggct atcaagataa acatagttat
16871 atttatcatg tgttaacacg aactcttttg taacgtaatc aatgtataaa attaattgtt ttcctccttg
16941 tgttatttct gacttgatag acgctaaact atcgttgtca tctttagtta gttgatttaa accctctaaa
17011 attaatgata aattgttaat catgtaaaac actcctttta tattaatttg atattgatac caccaatcga
17081 ataagattgg tagcattgta tcgaattaat atgttatttc tgtagttttc catgaatact cggaaataag
17151 atccatatct aattccttta gttcttcaaa agataacaaa caatattcct catcgcctac ctcatcaata
17221 tcaataagat aatgtttatt gttttcggta tctatgatat gataattcat atcccactca ttaaaggggt
17291 gaagtagaga tacctctcct ttttcagcta ttaatgattt attgttcata tgaaacactc cttttatatt
17361 aatttgatat tgataccacc aatcaaatgt gattggtagc attgtattaa attaatattc tggataattt
17431 attgagaaag tccagttatc atcaaatgaa attgttttat tttcaagtaa ctttttagcc tcatccacct
17501 caaattctaa atagaggaat ttactaagtt tatcctcatc tctaaaaatt ttcatacata ccacgttatt
17571 tgaataaatt tctgtgtata cgatcggttc attcatgttt atcatccttt ctttattaca tatatagtat
17641 atcatgtatt tacatatatg tcaatcattt aattcattta ttttaatgat ttatttgatt gtttttttat
17711 gatcctttct ttattacatc tatattatat catgtatgat tgtatttgtc aacaattaaa ttcatataaa
17781 tgtagtttgg ggtcagttac atttgtgtta tcaaaaaaag ataatattct att
Table 22
Table 23 Predicted amino acid sequences of ORFs from phage 182
182ORF001
5966 atggcaagaaggtatacaaatgtaaaattgttggctaacgtgccttttgataacacctatacacacacaagatggtttaaaact
1 M A R R Y T N V K L L A N V P F D N T Y T H T R W F K T
6050 caacaggaacaggaatcgtactttaattcgtttcctgttcttaacgagaatagagattgttcttatcaaagggatacacaactc
29 Q Q E Q E S Y F N S F P V L N E N R D C S Y Q R D T Q L
6134 gggggagtttttagagtagataaacacaaagacgccttatatgcttgtaactatctcatctttaaaaacgaagaaacttatcct
57 G G V F R V D K H K D A L Y A C N Y L I F K N E E T Y P
6218 agtaaatggcagtatgcctttgttactgatattgaatataagaatgacaacacaagtttcgttacctttgaaattgatgtttta
85 S K W Q Y A F V T D I E Y K N D N T S F V T F E I D V L
6302 caaacttatcgtttcgatattggtatacgagaaagtttcattgcaaaagaacaccctcaactttattattcgaatggaatacct
113 Q T Y R F D I G I R E S F I A K E H P Q L Y Y S N G I P
6386 ttcattaatacaattgaagagtcgcttgattacggtagagaatacacaacaacaaatgtaacaacttttcatcctaacgatgga
141 F I N T I E E S L D Y G R E Y T T T N V T T F H P N D G
6470 gtcaattttcttgttattctaacaagtgaagcaatgccagttggagataaggaagataaatcaggaggatcaatagtaggtggc
169 V N F L V I L T S E A M P V G D K E D K S G G S I V G G
6554 ccatctcctttttcctattatttacttcctatcaattcaagtggggaggtatacaaaccaaatggggcaggcaatgctaatttt
197 P S P F S Y Y L L P I N S S G E V Y K P N G A G N A N F
6638 ggagagtacatggcgtttcttacaacgaaagaaccttttttaaataagatagtcgggatgtatgtaacgtcgtatacaggtata
225 G E Y M A F L T T K E P F L N K I V G M Y V T S Y T G I
6722 ccattca tgtggatcacgcgaacaaaacggtaaggtataatgcaggaggttcttataagatcatgcttccaacctacgctagt
253 P F I V D H A N K. T V R Y N A G G S Y K I M L P T Y A S
6806 gatccaacaggaacaatgaaaacattcgctttcttttgtgtaaaagaagcaagaaσattcgtacctaaaagaattgatcttgta
281 D P T G T M K T F A F F C V K E A R T F V P K R I D L V
6890 gggaacgtgtataactactttagagaagcttttccgtttaatgttaaggaatcaaaactatttatgtatccctattgtttaata
309 G N V Y N Y F R E A F P F N V K E S K L F M Y P Y C L I
6974 gaaattacagatacaaaaggacatgtaatgactttaagacctgaatatcttacaggtggtaaattgagtgtatatgtaaaaggt
337 E I T D T K G H V M T L R P E Y L T G G K L S V Y V K G
7058 tcgttaggaatttctaataaagtgatgatcgagccgattgattatgatgtaagtaactcaaccattattaccaatttaagtgac
365 S L G I S N K V M I E P I D Y D V S N S T I I T N L S D
7142 aagatgttaatcgataatgatcctaacgatgtaggagttaaatctgactatgcttctgcattcatgcaaggaaacaaaaactcc
393 K M L I D N D P N D V G V K S D Y A S A F M Q G N K N S
7226 ttgattgctcaagagcaaaacattcgcaatactttcagacatggtatgggaaacagtgcaatgagtacaggaggagcgatcttt
421 L I A Q E Q N I R N T F R H G M G N S A M S T G G A I F
7310 tcagccttagcaagtaacaacccttttgttggtttgactaacatcatgggagcaggacaacaagtaaacaactatgtttctgaa
449 S A L A S N N P F V G L T N I M G A G Q Q V N N Y V S E
7394 aaagaaaacggtttgaacctcttggcaggtaaagtggcagatatcgaaaatattccagataatgtaacacagcttggatcaaac
477 K E N G L N L L A G K V A D I E N I P D N V T Q L G S N
7478 ttatctttcacaacaggaaactttcaaaactattatcaattgcgcttcaaacaaattaaatatgagtatgcaacaagacttgat
505 L S F T T G N F Q N Y Y Q L R F K Q I K Y E Y A T R L D
7562 cgttacttctcaatgtatggcacaaagagcaatcgagtagctacaccaaacttacaaacaagaaaagcatggaatttcattaaa
533 R Y F S M Y G T K S N R V A T P N L Q T R K A W N F I K
7646 ttaaaagaaccaaatattgtaggcacaatgagtaacgatgtattaacacgtgtgaaacaaatttttagtgcaggcgttacgctt
561 L K E P N I V G T M S N D V L T R V K Q I F S A G V T L
7730 tggcatacgaatgatgttttgaattataaccaagacaacggagatgtatag 7780
589 W H T N D V L N Y N Q D N G D V * 182ORF002
2152 atgattaagaaatatactggcgactttgaaacaacaactgatctcaacgattgtcgtgtatggtcgtggggcgtatgcgatata
1 M I K K Y T G D F E T T T D L N D C R V W S W G V C D I
2236 gacaacgttgacaatatgacgttcggtttagaaatcgattctttttttgagtggtgtaaaatgcaaggcagcacagacatttat
29 D N V D N M T F G L E I D S F F E W C K M Q G S T D I Y
2320 ttccacaacgaaaaatttgacggagagtttatgctttcatggttattcaaaaatggtttcaaatggtgtaaagaagcaaaagaa
57 F H N E K F D G E F M L S W L F K N G F K W C K E A K E
2404 ga cgaacattctccacactcatatcaaatatgggtcaatggtatgctttggaaatttgttgggaagttaattacacaacaaca
85 D R T F S T L I S N M G Q W Y A L E I C W E V N Y T T T
2488 aaatcaggtaaaacgaaaaaagagaaatctcgaacaataatttatgatagccttaaaaaatatccttttccagtgaaacaaatt
113 K S G K T K K E K S R T I I Y D S L K K Y P F P V K Q I
2572 gcagaagcttttaattttcctataaaaaaaggcgaaatagattatacaaaagaaagacctattggttacaaaccaacaaaagat
141 A E A F N F P I K K G E I D Y T K E R P I G Y K P T K D
2656 gaatgggagtatttaaagaacgacattcagattatggcgatggcattaaaaattcaattcgatcaaggactaactcgaatgact
169 E W E Y L K N D I Q I M A M A L K I Q F D Q G L T R M T
2740 agaggaagcgacgctttaggcgattacaaagattggctaaaagctacacatggaaaatcaactttcaaacaatggtttcctatt
197 R G S D A L G D Y K D W L K A T H G K S T F K -Q W F .P. T "
2824 ttgtctttagggtttgataaagacttacgtaaagcatacaaaggcggcttcacttgggtaaacaaagtttttcaagggaaagaa
225 L S L G F D K D L R K A Y K G G F T W V N K V F ~Q G K E
2908 ataggtgacggcattgtctttgatgtcaactctttgta ccctctcaaatgtacgtaagacctttaccatatggaacacctcta
253 I G D G I V F D V N S L Y P S Q M Y V R P L P Y G T P L
2992 ttctacgaaggagaatacaaaccgaacaacgactatccgctgtacattcaaaatatcaaagtaagattccgtttaaaggagggt
281 F Y E G E Y K P N N D Y P L Y I Q N I K V R F R L K E G
3076 tatattccaaccattcaagttaagcaaagttcattattcattcaaaacgaatatcttgaatcaagtgtaaacaagttaggagtt
309 Y I P T I Q V K Q S S L F I Q N E Y L E S S V N K L G V
3160 gacgaattaatcgatcttactcttacaaatgttgacctagaattattttttgaacactacgatattttagagatacattacact
337 D E L I D L T L T N V D L E L F F E H Y D I L E I H Y T
3244 tacggatatatgttcaaagcttcttgtgatatgttcaaaggctggatcgataaatggatcgaagtaaagaacaccaccgaaggg
365 Y G Y M F K A S C D M F K G W I D K W I E V K N T T E G
3328 gctagaaaagctaacgccaaaggtatgttaaatagcttgtatggaaagttcggaacaaaccctgacattacaggaaaagtgcct
393 A R K A N A K G M L N S L Y G K F G T N P D I T G K V P
3412 tacatgggcgaggacggcattgttcgattgacactaggagaagaagaattaagagatcctgtttatgttccgcttgctagtttt
421 Y M G E D G I V R L T L G E E E L R D P V Y V P L A S F
3496 gtgacggcttggggtagatatactaccattacaaccgctcaaaaatgttttgatcgcattatttattgtgatacagatagcatt
449 V T A W G R Y T T I T T A Q K C F D R I I Y C D T D S I
3580 catctagtaggaacagaagttccagaagcaatcgatcacttggttgatcctaaaaaacttggttattgggggcatgaaagcaca
477 H L V G T E V P E A I D H L V D P K K L G Y W G H E S T
3664 tttcaacgagcaaaattcattcggcagaaaacatacgtagaagaaattgatggcgaattaaatgtaaagtgtgctggtatgcca
505 F Q R A K F I R Q K T Y V E E I D G E L N V K C A G M P
3748 gatcgaataaaagagattgtaacttttgacaattttgaagttggtttttcaagctatggaaagttgctacctaaaagaacacaa
533 D R I K E I V T F D N F E V G F S S Y G K L L P K R T Q
3832 ggtggcgtggtattagtagacacaatgtttacaatcaaataa 3873
561 G G V V L V D T M F T I K * 182ORF003
11305 atggaagaacgaattgatattcaaatgaacaagatgaaagaagaaaatcaaaagaattacctattgcaccctgaaacgaacccg
1 M E E R I D I Q M N K M K E E N Q K N Y L L H P E T N P
11389 aaacaagttgtttttgatgaaacattgcatggaaatgaaaatcaggagagtttcaacaattttgttgacacaagaaaaatgaca
29 K Q V V F D E T L H G N E N Q E S F N N F V D T R K M T
11473 actacaattgatgtaagtgcttatggggttatcgctgacggtgtaacagattgtacaccaatattaaataaattacttgaagaa
57 T T I D V S A Y G V I A D G V T D C T P I L N K L L E E
11557 aaaagcgaaatgggtatcactttttattttcctccttgtgaacgtgattcatattatcgctttgctaacaccattgaattgaaa
85 K S E M G I T F Y F P P C E R D S Y Y R F A N T I E L K
11641 cgtgatgtacctgtagttactttcttaggatcgggagaaacgacattaaagtttgaaacaatgacggcatttaatgtaaacatc
113 R D V P V V T F L G S G E T T L K F E T M T A F N V N I
11725 gaaagtttcaatattgatggttttgcattatggttgccacaaggcgctcaaagtggtaaaggaattttctttaatgatactcgc
141 E S F N I D G F A L W L P Q G A Q S G K G I F F N D T R
11809 aattacaatcgttttgactttgatttgtttgttcgtaactgtactttaaatgaaggaacgtatgttgttgttgctagaggtaga
169 N Y N R F D F D L F V R N C T L N E G T Y V V V A R G R
11893 ggggttacatttgaaaattgtctattctctaatatctctcaagcaattatcaaaacagcttttcccgatgtaaatggtatgtgg
197 G V T F E N C L F S N I S Q A I I K T A F P D V N G M W
11977 caagggaacgatatcaatactaggggtacaggttttagaggtttctttgtgaaaaacaaccgtattcatttttgtacagcgatc
225 Q G N D I N T R G T G F R G F F V K N N R I H F C T A I
12061 attatcgacaatgacgatgattatcagaatgtaattaatttctgtgaaatttctggtaacacaatcgaaggtggcgtaagttat
253 I I D N D D D Y Q N V I N F C E I S G N T I E G G V S Y
12145 tatcgaggatatgcgcataacttgcatgtccaaaacaacaaccattttctagcatacggaaatagaaacgctttgtttgagttt
281 Y R G Y A H N L H V Q N N N H F L A Y G N R N A L F E F
12229 caagatgtggatcaagcttatattgatgtagatgtttattgtcgtaactcacaagtcgagggaatgaatagtacagctatttca
309 Q D V D Q A Y I D V D V Y C R N S Q V E G M N S T A I S
12313 cgtttaattgttgtttacggacattaccgaaacttaaagattacaggtaaattatatcgttgtcaaggacatgttatcacgttg
337 R L I V V Y G H Y R N L K I T G K L Y R C Q G H V I T L
12397 tatggcggtggcgttaatttctattgtgacttgatggcacaagaagcacctttgacggacggttaccggtttattcaaacggct
365 Y G G G V N F Y C D L M A Q E A P L T D G Y R F I Q T A
12481 gacaatcgagttaactatgatgggtttgttgttcgtggtttgtctaattcaacaaaagtaaatacaccaatgatctataaagca
393 D N R V N Y D G F V V R G L S N S T K V N T P M I Y K A
12565 cctcagactgttttctataatcgtagaatcgatcatgtgctaacaggtccaaatgcaagtaatgtatataactag 12639
421 P Q T V F Y N R R I D H V L T G P N A S N V Y N * 182ORF004
4626 atggctgacaaaatcacagaacaagatgttcttcgtgccacaaatgtagaaacaccagtacaattaatgactgctatttataat
1 M A D K I T E Q D V L R A T N V E T P V Q L M T A I Y N
4710 agttcatcatctctttttcaggcgaacgtacctatgccaaatgcagataacatcgaagcggttggtgcagggatcacacgttta
29 S S S S L F Q A N V P M P N A D N I E A V G A G I T R L
4794 gacgtagtaaaaaacgaatttatttcaactttagttgaccgtattggtaaagtagttatccgatacaaatcttggcgtaaccct
57 D V V K N E F I S T L V D R I G K V V I R Y K S W R N P
4878 ttgaaaatgtttaaaaaaggaaacatgcctttaggtcgaacgattgaagaaatttttgttgacattgcacaggaacataagttc
85 L K M F K K G N M P L G R T I E E I F V D I A Q E H K F
4962 aaccctgacgagtctgttacaggggtatttaaacaggaagttcccgatgtaaaaacattgttccacgaaattaatcgtgaaggt
113 N P D E S V T G V F K Q E V P D V K T L F H E I N R E G
5046 tactacaaacaaacgatccaagaagcatggttagaaaaagcatttacttcatgggataatttcaatagtttcgttgctggtgta
141 Y Y K Q T I Q E A W L E K A F T S W D N F N S F V A G V
5130 atgaacgctttatacacaggtgacgaagtaagcgaatttgaatacacgaaattattaatagcaaactaccaagaaaaagagcta
169 M N A L Y T G D E V S E F E Y T K L L I A N Y Q E K E L
5214 ttcaaagagatcgaaattggcgaaattactgaatcaaatgcaaaagaatttatccgtaagatcaaatcaacctctaacaaatta
197 F K E I E I G E I T E S N A K E F I R K I K S ~T S_a T L
5298 gaatttatgagttccgcttacaacgctcaaggagttaaaacatctacctcaaaatctgatcaatacgttatfeattgacgccgac
225 E F M S S A Y N A Q G V K T S T S K S D Q Y V I I D A D
5382 acagacgcaaccattgacgttgacgttttagcagcggcattcaatatgagtaaaactgactttgtaggacacaaaatcgttatt
253 T D A T I D V D V L A A A F N M S K T D F V G H K I V I
5466 gatgagtttcctaaaaaagaaggcgaagaatcgtcaaatattgtggcagttattgtagatagtgaatggtttatgatctacgac
281 D E F P K K E G E E S S N I V A V I V D S E W F M I Y D
5550 aaattgtacaaaacaacaagtctatacaaccctgaagggttatattggaattattggttgcaccaccaccaactatattctact
309 K L Y K T T S L Y N P E G L Y W N Y W L H H H Q L Y S T
5634 tctcaattcgggaacgctgttgcttttgttaaatcagcaacaaaacctgtcacaaaagttgcttttgcaagtgcaacaactagt
337 S Q F G N A V A F V K S A T K P V T K V A F A S A T T S
5718 gttgttaaaggatcatctaaagatatcgcattgacatttacaccagtagaagcaacaaaccaacaaggagaagttgtttcatca
365 V V K G S S K D I A L T F T P V E A T N Q Q G E V V S S
5802 gcaccagcattggttaaggcaaccgtaaaacaaacagcaggtaaagcgactgccgtaaccgtagaaggcttagaagtcggtcaa
393 A P A L V K A T V K Q T A G K A T A V T V E G L E V G Q
5886 tcattagtaacattcacagctatcggaggtcaacaagcaacggttcttgttacggttacttctgactaa 5954
421 S L V T F T A I G G Q Q A T V L V T V T S D * 182ORF005
12651 atggcaactcttacaaatgaacaaatagctagaggacaaacaatcgctaaaatactttcaaaatatggctataataaaaattca
1 M A T L T N E Q I A R G Q T I A K I L S K Y G Y N K N S
12735 caagtaggagttgtcgccaatctccattgggaatcggctggtttgaacccgaacagcaatgaatatggtggaggcggatatggg
29 Q V G V V A N L H W E S A G L N P N S N E Y G G G G Y G
12819 ttaggtcaatggacgcctaaaagcaatctttatcgccaagcacaaatttgtgggttgtctaatgctaaagctgaaacgttggaa
57 L G Q W T P K S N L Y R Q A Q I C G L S N A K A E T L E
12903 ggtcaagcagagatcatcgctcaaggggataaaacaggtcaatggatggataatacacctgtttcttctgcaggttatactaac
85 G Q A E I I A Q G D K T G Q W M D N T P V S S A G Y T N
12987 cctcagaccctttcagcatttaaacaatctgcaaatattgatgttgctacaattaattttatgtgtcactgggaacgccctggt
113 P Q T L S A F K Q S A N I D V A T I N F M C H W E R P G
13071 aaacttcatatcgaagaaagacttgatcttgcacaagcttatagtaagcatattgacggtagcggtggcggtggcgtaaaacgt
141 K L H I E E R L D L A Q A Y S K H I D G S G G G G V K R
13155 tgctatggaaccccaatcaagaatacaaatcttgatcctaaaagtttcatgagtggacaactttttggcacgcatgcaggaaac
169 C Y G T P I K N T N L D P K S F M S G Q L F G T H A G N
13239 ggcagaccaaataatttccatgatggtttggactttggttcaattgatcaccctggcaatgaaatgattgcatgttgcgatgga
197 G R P N N F H D G L D F G S I D H P G N E M I A C C D G
13323 acagtaacacatgttggaacaatgggagcattaagagcgtattttgtgataaatgatggtacttacaatatcgtttatcaagaa
225 T V T H V G T M G A L R A Y F V I N D G T Y N I V Y Q E
13407 tttagttataaccagtcaaatataaaggtaaaagttggcgacaaagttaagaacggacaagtttgcgcaatacgtgacgcggat
253 F S Y N Q S N I K V K V G D K V K N G Q V C A I R D A D
13491 catttacatttaggttttactaaaaaagattttatgactgcgttaggatcttctttcatagatgatggaacatgggaagaccct
281 H L H L G F T K K D F M T A L G S S F I D D G T W E D P
13575 ttgaagtttttagggcaatgttttggagatggagatactggcggagataatgacgataacaataaggataaaaatgatcttatt
309 L K F L G Q C F G D G D T G G D N D D N N K D K N D L I
13659 tatctattgctatccgatgccttgaatggttggaaattttaa 13700
337 Y L L L S D A L N G W K F * 182ORF006
14995 atgacaaatagcttaggcgttaaacttgaagagaaaaacttatactataaccctaacaatgctttaggttttaattgcctaatg
1 M T N S L G V K L E E K N L Y Y N P N N A L G F N C L M
15079 ttgtttgtaataggcgcacgtggtataggtaaaacttatggttataaaaaatttgttgttaatcgctttattaaacacggcgaa
29 L F V I G A R G I G K T Y G Y K K F V V N R F I K H G E
15163 caatttatttatttaagaagattcaaaacagaacttaaaaagattcctcaatttttcaaaacaatggcgaaagaatttcctgat
57 Q F I Y L R R F K T E L K K I P Q F F K T M A K E F P D
15247 cataaacttgaagtaaaaggaaaagaattctattgtgatgataaattaatgggttgggctgttccacttagtacgtggggaatt
85 H K L E V K G K E F Y C D D K L M G W A V P L S T W G I
15331 gaaaaatctaatgaatatcccgaagttcgtacaattttgtttgatgagtttttaattgagaaatcaaaaatcacttatttacca
113 E K S N E Y P E V R T I L F D E F L I E K S K I T Y L P
15415 aacgaagctgaagccttattgaacatgatggaaacggttttccgaagacgtacaaatacaagatgtgttatgttgagtaatgca
141 N E A E A L L N M M E T V F R R R T N T R C V M L S N A
15499 actagtgtagtgaacccttatttcttgtatttcaatctgcagccagatttgaataagcgttttaatctatatcaagatcgaggt
169 T S V V N P Y F L Y F N L Q P D L N K R F N L Y Q D R G
15583 atattgattgaattgtgtgattcaaaagactttgcagaagtgaagagagaaacaccttttggtagattgattcgtggaacagaa
197 I L I E L C D S K D F A E V K R E T P F G R L I R G T E
15667 tacgaagattttagtatcaacaatgagtttgtcaatgatagtgatacgtttattgaaaagagaagtaaaaatagtagtttctta
225 Y E D F S I N N E F V N D S D T F I E K R S K N S S F L
15751 tgcgccattgcttttgaagggaaaatctttgggtattggatagacgctgaaacaggttgtgtctatgtgagttatgattatcaa
253 C A I A F E G K I F G Y W I D A E T G C V Y V S Y D Y Q
15835 ccaaatacaaatcatttttatgcaatgactacgaaagaccatgaagaaaatagattgctgatgaaaaattggcgaaataattat
281 P N T N H F Y A M T T K D H E E N R L L M K N W R N N Y
15919 tatctttcaacagtggcgaaagcattcaagaatagttatctgcggtttgataacattgttattaagaatttacattatgatttg
309 Y L S T V A K A F K N S Y L R F D N I V I K N L H Y D L
16003 tttaataagatgaaaatctggtaa 16026
337 F N K M K I W *
182ORF007
7795 atgagtagacgaaaaggtgcaggacttgctagaaataaccgttatacagcaaaaagcagaccttatccaaatgaaccctattca
1 M S R R K G A G L A R N N R Y T A K S R P Y P N j) P Y S
7879 agtgatgtagaagaaatcagctactatgaacattatcgtagacaactcacgctccttacgtttcagttgtttgaatgggaaaat
29 S D V E E I S Y Y E H Y R R Q L T L L T F Q L F E W E N
7963 ttgccaaaatcaattgaccctcgttatttagaaattgctttacacactaatggttatcttggtttctttaaagaccctacactt
57 L P K S I D P R Y L E I A L H T N G Y L G F F K D P T L
8047 gggttcatggtttgcgcaggggcagaagatggtcaaatcgatcattatcacaaccctattttctttacagcaaacgaagcaatg
85 G F M V C A G A E D G Q I D H Y H N P I F F T A N E A M
8131 tatcacaagagatatcctgttttaagatatgatgatgatgatgataaatcaaaatgtatcatgttgtataataatgacttgaaa
113 Y H K R Y P V L R Y D D D D D K S K C I M L Y N N D L K
8215 gt cctacgttaccaagtttacatcgttttgctttagatatggcggacataaaccagatatcacgagtgaatcgaagagcgcaa
141 V P T L P S L H R F A L D M A D I N Q I S R V N R R A Q
8299 aaaacacctgtaattattcaaactgatgaaaagaaatacttctcattgctacaagcttataaccaaattgacgaaaataatcag
169 K T P V I I Q T D E K K Y F S L L Q A Y N Q I D E N N Q
8383 gctgtttttgtggataaagatatggagtttgacgaatcttttaatgtatggcaaacaaatgctccatatgtagtagataaacta
197 A V F V D K D M E F D E S F N V W Q T N A P Y V V D K L
8467 cgatcagaattgaacgaagtatggaatgaagtgttaacttttctaggtatcaacaatgctaacgtagataagactgcacgtgta
225 R S E L N E V W N E V L T F L G I N N A N V D K T A R V
8551 caaacatcagaagtcttatctaacaatgaacagattgaaagttcaggtaacatcttgttaaaatcaagaaaagagttttgcgat
253 Q T S E V L S N N E Q I E S S G N I L L K S R K E F C D
8635 cgtgtaaatcgtgtctttggcgatgaacttgacggaaagattgacgtgaagtttagaacagacgccgttcgacaattacaactg
281 R V N R V F G D E L D G K I D V K F R T D A V R Q L Q L
8719 gcggcaggtcaatcaaaaaaagaccagatgagtggagggttgccaagtgctacttaa 8775
309 A A G Q S K K D Q M S G G L P S A T * 182ORF008
14105 atgatgaatggtattgatatctctagttatcaaacaggaattgatctttcaaaagttccatgcgattttgtaaatattaaagca
1 M M N G I D I S S Y Q T G I D L S K V P C D F V N I K A
14189 acaggcggaacaggttatgtaaaccctgattgtgaccgagcatttcaacaagctttgtctttaggtaaaaagattggtgtgtat
29 T G G T G Y V N P D C D R A F Q Q A L S L G K K I G V Y
14273 cattttgcgcatgagaggggtttagaaggtacacctcaacaagaagcgcaattctttttagataatattaagggttacattggt
57 H F A H E R G L E G T P Q Q E A Q F F L D N I K G Y I G
14357 aaagctgttcttattcttgactttgaagggtcaaatcagaaagatgtaaattgggcgaaagcatttcttgattatgtttataat
85 K A V L I L D F E G S N Q K D V N W A K A F L D Y V Y N
14441 aaaacaggcgttaaagcatggttttatacgtatacagcaaacctcaatacaactgatttttctagtattgcaaaaggcgattat
113 K T G V K A W F Y T Y T A N L N T T D F S Ξ I A K G D Y
14525 ggtttatgggttgctgaatatggatcaaatcaaccacaaggctactctcaaccagcgccacctaaaacaaataattttccaatt
141 G L W V A E Y G S N Q P Q G Y S Q P A P P K T N N F P I
14609 gttgcctgttttcagtttacaagtaaaggacgtttaccaggatacaacggcaatcttgatttgaatgttttctatggcgatggt
169 V A C F Q F T S K G R L P G Y N G N L D L N V F Y G D G
14693 aatacatgggatctgtatgtaggtaaaaaacaggatcaaattgttcctcctgaaaataaaatatttgacgccacaagtgatgag
197 N T W D L Y V G K K Q D Q I V P P E N K I F D A T S D E
14777 tttattttcactcttacaacaggtagcacaagcgtgttttattttgacggagaaacgatctttgaattgtctgatccaacacaa
225 F I F T L T T G S T S V F Y F D G E T I F E L S D P T Q
14861 ctcgatcatattagaggaacatacaatcatgttcatggaaaagaaatcccatcaatggtgtggacacctgaacaatttgatatt
253 L D H I R G T Y N H V H G K E I P S M V W T P E Q F D I
14945 tacttaaaaatgtatgaaaagaaaccagtatataaatag 14983
281 Y L K M Y E K K P V Y K * 182ORF009
8765 gtgctacttaaacgttatattgaaagtttcacttattaccaacctgaattatctcgaaaagaacgtattgaagttggccgaaaa
1 V L L K R Y I E S F T Y Y Q P E L S R K E R I E V G R K
8849 caattgtttgattttgattatccgttttatgacgaaacaaaacgagcagaatttgaaacaaaatttatcaatcacttttacttg
29 Q L F D F D Y P F Y D E T K R A E F E T K F I N H F Y L
8933 agagagataggctcagaaacgatgggatcatttaagtttaatcttgacgaatatttaaatctaaacatgccctattggaataaa
57 R E I G S E T M G S F K F N L D E Y L N L N M P Y W N K
9017 atgttcctatcaaatcttgaagagtttccgatttttgatgacatggactacaccattgatgagaaacagaaattgttaaatgag
85 M F L S N L E E F P I F D D M D Y T I D E K Q K L L N E
9101 attgatacaaacatcaaagcgaatcgtgatgaatcgaagaaccaaacgaagcaagtagatcaaacagacaacagaaacaaaaat
113 I D T N I K A N R D E S K N Q T K Q V D Q T D F. N K N
9185 acacgtgacacaggaacaaccgattctttctcaaggaacacttatacagacacccctcaaaaagatttgagaattgccagcaat
141 T R D T G T T D S F S R N T Y T D T P Q K D L R I A S N
9269 ggagatggaacaggtgtaatcaattatgcaacaaatatcacagaagatttgagtaaagaaacaacaagctccacaggcgttgaa
169 G D G T G V I N Y A T N I T E D L S K E T T S S T G V E
9353 acaaacaacgacaaaacaaatcaaaatacacgaagcaatgcttctgaaaaagaaacaaagaacacagacattaataaagatcaa
197 T N N D K T N Q N T R S N A S E K E T K N T D I N K D Q
9437 aatcaaaccaaagatacgattacacgatataaaggtaaaaagggaaacactgattatgctgacttactcgaaaaatatcgtaga
225 N Q T K D T I T R Y K G K K G N T D Y A D L L E K Y R R
9521 agtgttttgagaattgagaaaatgatctttagagaaatgaacaaggaaggcttatttctccttgtttatggagggaggtag 9601
253 S V L R I E K M I F R E M N K E G L F L L V Y G G R *
182ORF010
1310 ttgaccgtaagaatatcaaagaatgatagagccaagttagagaaaatctacggtaaatctaacaaagctcgtaaaaaatacaat
1 L T V R I S K N D R A K L E K I Y G K S N K A R K K Y N
1394 cgtttaagacaaaaaggagttgaggaaaggcaacttccaactgttccaacatcaaagaaaagacttattgactacgtaaaatca
29 R L R Q K G V E E R Q L P T V P T S K K R L I D Y V K S
1478 acaaatatgagtcgtagtgattttaacaagatgttagacgagttggtagattttgcacaaccttacaacgagaattaσattttt
57 T N M S R S D F N K M L D E L V D F A Q P Y N E JJ~Y I F
1562 gagatcaacaagcgaaatgttgcaatctcaagagcgcaaatcaaagaagcgcaaattaaaacagagcaagctcaaaaagcgaaa
85 E I N K R N V A I S R A Q I K E A Q I K T E Q A Q K A K
1646 gaagaacactacaaagagcttaacaaagttgaagttaagaagcccacagaaaacacaattgtcacaccaactattttaacagag
113 E E H Y K E L N K V E V K K P T E N T I V T P T I L T E
1730 ttaggtgctgacttaccttttcaagcaataccagattttaatattgacgctttcacttctccagaaggagttcagtcttattta
141 L G A D L P F Q A I P D F N I D A F T S P E G V Q S Y L
1814 gaaaatataggaaaacaagacgaacaatat ttgacgaaagagaccaactttattacgacaatttcagacaagcgatgtttact
169 E N I G K Q D E Q Y F D E R D Q L Y Y D N F R Q A M F T
1898 attttcaattcagacgctgacgatattgttcgtttacttgactcaatggggcttgatctatttatgaaaacatatgttagtaac
197 I F N S D A D D I V R L L D S M G L D L F M K T Y V S N
1982 ttcttagacatgaaccttgactacatttatgacgaagcagaagtacaacagaaaaaagaacaagtttacagtaagattgcaaaa
225 F L D M N L D Y I Y D E A E V Q Q K K E Q V Y S K I A K
2066 gtgatcgagtctgaaacaggtggagaagtcccctcatataaccccacgaagaacatcacaattaattcagaaacaggagaagaa
253 V I E S E T G G E V P S Y N P T K N I T I N S E T G E E
2150 ttatga 2155
281 L * 182ORF011
9607 atggtagattttaaccccgacaagcggtttgacggtttacccgctgtattcaaagaacgctttagcaaatatcctcatactgaa
1 M V D F N P D K R F D G L P A V F K E R F S K Y P H T E
9691 tacagatatgaattactattagatgaagaagtatcggctttaattgcctatctgaatgaagttggtgctttagttaatgatatg
29 Y R Y E L L L D E E V S A L I A Y L N E V G A L V N D M
9775 agtggttatttaaattactttatcgaacattttgttgagaagttagaagagatcacaaatgacacactcaaaaaatggttgtct
57 S G Y L N Y F I E H F V E K L E E I T N D T L K K W L S
9859 gatggtacgttagaaaatttaatcaatgatactgtttttgcaaattatatcaaagaaatcaaaagattacaaatcttggttgct
85 D G T L E N L I N D T V F A N Y I K E I K R L Q I L V A
9943 gaaacacgtgctaacagtgtgaatattcttttgacaaaaaataaaccggatgttgctgatgatcgaacattttggtataagatt
113 E T R A N S V N I L L T K N K P D V A D D R T F W Y K I
10027 caacgcgacaatactgattatggagccgatcctattgacacgttacgtattgttgcaatcaataaagttagtggctggaatacc
141 Q R D N T D Y G A D P I D T L R I V A I N K V S G W N T
10111 gctacaggagatatttatcttaacattaaaggaacggagggtgtataa 10158
169 A T G D I Y L N I K G T E G V * 182ORF012
10872 atggcaaataaaaatattcaaatgaaggatagcaatgacaataatttatatccaagtgttcgagcagaaaacttgttagatttg
1 M A N K N I Q M K D S N D N N L Y P S V R A E N L L D L
10956 accagtcgtgctgaattaacaatgacaaattgtcaattatatgcagctggtgataaaacaaatgcaatctcttatctcggtgca
29 T S R A E L T M T N C Q L Y A A G D K T N A I S Y L G A
11040 gtaggtatgctcgaaggtatgataaagtttactgaaagtttgacaaaccctgtgatcacaacgctaccagaaggttttagacca
57 V G M L E G M I K F T E S L T N P V I T T L P E G F R P
11124 ataagaacaaaacgtattggttgtttcgcaaaatattacacaccaaatccaacagatacaaaagaaatggtttatgtatcaatc
85 I R T K R I G C F A K Y Y T P N P T D T K E M V Y V S I
11208 acacctgatggcaaagtaactgtaaatgacaatgtaggtaaaatcgaatatctatccctagataattgcgttttccctctaaaa
113 T P D G K V T V N D N V G K I E Y L S L D N C V F P L K
11292 taa 11294
141 * 182ORF013
10456 atggcagataaaaatattcaaatgcaggataaagatcataatcgtttaatgcctgttacaattgctaaaaatgttctaacaggc
1 M A D K N I Q M Q D K D H N R L M P V T I A K N V L T G
10540 gactctaatcttgaattagttaatgctgaaataagaggtaacgctagtgaagctaaaacacttgcacaacaagctaaagaaact
29 D S N L E L V N A E I R G N A S E A K T L A Q Q A K E T
10624 gctgctggtttgtcaacagaaattgacacagtaacatcaaccgcaaatcaagcgttgacgaaggctggtacagcacaacaaacc
57 A A G L S T E I D T V T S T A N Q A L T K A G T A Q Q T
10708 gcagaacaagcgaaaacaacagcaaacagtatcagcgcagttgcaacggcagctaaaaacacagctgattcagcacaaaaaagt
85 A E Q A K T T A N S I S A V A T A A K N T A D S A Q K S
10792 gcaactgatctagctgttcgagtaagcagtttagaggacacagcaatacaatatactgtattaccatag 10860
113 A T D L A V R V S S L E D T A I Q Y T V L P * 182ORF014
13716 atgatagaatatatcacacaatggttggcagatgataatcatcttgtttatggtttgattatatggttaatggttgcaatgatt
1 M I E Y I T Q W L A D D N H L V Y G L I I W L M V A M I
13800 atcgattttgtgttaggttttacaattgccaaatttaacaaggaaatcgactttagtagttttaaagctaaagcaggtatcatt
29 I D F V L G F T I A K F N K E I D F S S F K A K A G I I
13884 gttaaggtggcagaaatggttttagtggtttactttattcctgtagcagtaaaattcggtgcagtaggtattacaatgtatata
57 V K V A E M V L V V Y F I P V A V K F G A V G I T M Y I
13968 acaatgttggttggtttgattttatcagaaatttatagtatactaggacatatttcagatatcgatgatgataataattggact
85 T M L V G L I L S E I Y S I L G H I S D I D D D N N W T
14052 gattatgttaagaagtttttagacggaacactcaacagaaaggacgatattaaatga 14108
113 D Y V K K F L D G T L N R K D D I K * 182ORF015
854 atggaaatcgtaaaaagcacatttgacacacaaacaccagaaggaatgttacaagtat caatgccacaaacggggcttcaatt
1 M E I V K S T F D T Q T P E G M L Q V F N A T N G A S I
938 ccgttacgtaacgcaattggcgaagtactagaattgaaagatattctagtttactcagacgaagtttctggttttggtggagcc
29 P L R N A I G E V L E L K D I L V Y S D E V S G F G G A
1022 gaaccatcacaagcagaactagtcgctttcttcacagaagatggtaaaacttatgcgggtgtatcagcagtagcaacaaaatca
57 E P S Q A E L V A F F T E D G K T Y A G V S A V A T K S
1106 gctaaaaacctaattgatatgatgactgctaaccctgacatcaaaccaaaaatttcttttgtcgaaggaaaatcaaacggtgga
85 A K N L I D M M T A N P D I K P K I S F V E G -K S_N -Θ G~
1190 caaaaatttgtaaatctacaagtggtttcactgtag 1225 _,
113 Q K F V N L Q V V S L * 182ORF016
17033 atgattaacaatttatcattaattttagagggtttaaatcaactaactaaagatgacaacgatagtttagcgtctatcaagtca
1 M I N N L S L I L E G L N Q L T K D D N D S L A S I K S
16949 gaaataacacaaggaggaaaacaattaattttatacattgattacgttacaaaagagttcgtgttaacacatgataaatataac
29 E I T Q G G K Q L I L Y I D Y V T K E F V L T H D K Y N
16865 tatgtttatcttgatagccattgcattaatatcgcaataacgaaatcaatgaaaagcgttgaacactatgcggaacaattgaaa
57 Y V Y L D S H C I N I A I T K S M K S V E H Y A E Q L K
16781 catgacggatataaacaaattacggacaaatag 16749
85 H D G Y K Q I T D K *
182ORF017
154 atgaaatattcactacaacaaatagatgaaattaaatcaacaattttcagaattagattaaaaaggcatgaactagaggaattg
1 M K Y S L Q Q I D E I K S T I F R I R L K R H E L E E L
238 gtggacgaagtaaacgatattgctaaagatccggaggaaagatatcttttatcgttttattacacagaagaagaacgtttgttt
29 V D E V N D I A K D P E E R Y L L S F Y Y T E E E R L F
322 gaaattccctctgcaagattaatagattattacaacgaaaagatcacaaatctgaaatcggaaatcatatcactcgaaaaaaga
57 E I P S A R L I D Y Y N E K I T N L K S E I I S L E K R
406 ttacaaaaactagtaaaataa 426
85 L Q K L V K *
182ORF018
16737 atgattgcacgaacattcaaagaacaccgcgaactaattgaatggttacgtttctactgtaaacgtaacctttcagacaatgaa
1 M I A R T F K E H R E L I E W L R F Y C K R N L S D N E
16653 aaaatagagatcatagaggggactttacaagatttcgacgttccggaaataaatatcaccgaacttttgttaactcattcaacg
29 K I E I I E G T L Q D F D V P E I N I T E L L L T H S T
16569 ctattacccgaatcgagtcaatttaacattcttgaaaagtattgtcaggcaatgaaattagtaacttcatacgtaaaagttggt
57 L L P E S S Q F N I L E K Y C Q A M K L V T S Y V K V G
16485 tctcgctatcagttagcgttacaaataccaaaaggctatttaaaggaggtggaataa 16429
85 S R Y Q L A L Q I P K G Y L K E V E *
182ORF019
4323 atggaaattaaagaacatgaatcaattttaaatggtattcttgaaagtgtcacagacggtgaagcaagatcaaagattgtagaa
1 M E I K E H E S I L N G I L E S V T D G E A R S K I V E
4407 catcttgaagcattgcgagaagactacggagcaacaactgaagctttgacatcagcaaatagcacacttgaaaagttaaagaaa
29 H L E A L R E D Y G A T T E A L T S A N S T L E K L K K
4491 gataacgaagcgttggttatttcaaactcaaaattgttccgagaacgagcgatcgtagaaccagcagaaaataacgaaccagaa
57 D N E A L V I S N S K L F R E R A I V E P A E N N E P E
4575 acagaccagaatattacactagacgatttaggaatttaa 4613
85 T D Q N I T L D D L G I *
182ORF020
10158 atggcagacattagaacacaactaacaagtgaagatggatcagacaatttatttccaatttcaaaagccgttaatattatgact
1 M A D I R T Q L T S E D G S D N L F P I S K A V N I M T
10242 aatagcggtacgaatgtagaaggagaattgggtacactcaaacaaaatgacgaaacaatgaatacctcagttcaaaatgctgta
29 N S G T N V E G E L G T L K Q N D E T M N T S V Q N A V
10326 gttactgccaatcaagcaaaagattctgtagctgaattaaatgtaaatgttggtaaactaaccaatcgaataacaacattagag
57 V T A N Q A K D S V A E L N V N V G K L T N R I T T L E
10410 agtacagtggctaatcttgatggtattcgttatgtagaggtgtaa 10454
85 S T V A N L D G I R Y V E V *
182ORF021
17339 atgaacaataaatcattaatagctgaaaaaggagaggtatctctacttcacccctttaatgagtgggatatgaattatcatatc
1 M N N K S L I A E K G E V S L L H P F N E W D M N Y H I
17255 atagataccgaaaacaataaacattatcttattgatattgatgaggtaggcgatgaggaatattgtttgttatcttttgaagaa
29 I D T E N N K H Y L I D I D E V G D E E Y C L L S F E E
17171 ctaaaggaattagatatggatcttatttccgagtattcatggaaaactacagaaataacatattaa 17106
57 L K E L D M D L I S E Y S W K T T E I T Y *
182ORF022
12868 gtgggttgtctaatgctaaagctgaaacgttggaaggtcaagcagaga catcgctcaaggggataaaacaggtcaatggatgg
1 V G C L M L K L K R W K V K Q R S S L K G I K Q V N G W
12952 ataatacacctgtttcttctgcaggttatactaaccctcagaccctttcagcatttaaacaatctgcaaatattgatgttgcta
29 I I H L F L L Q V I L T L R P F Q H L N N L Q I L M L L
13036 caattaattttatgtgtcactgggaacgccctggtaaacttcatatcgaagaaagacttgatcttgcacaagcttatagtaagc
57 Q L I L C V T G N A L V N F I S K K D L I L H K L I V S
13120 atattgacggtagcggtggcggtggcgtaa 13149
85 I L T V A V A V A *
182ORF023
12189 atggttgttgttttggacatgcaagttatgcgcatatcctcgataataacttacgccaccttcgattgtgttaccagaaatttc
1 M V V V L D M Q V M R I S S I I T Y A T F D C V T R N F
12105 acagaaattaattacattctgataatcatcgtcattgtcgataatgatcgctgtacaaaaatgaatacggttgtttttcacaaa
29 T E I N Y I L I I I V I V D N D R C T K M N T V V F H K
12021 gaaacctctaaaacctgtacccctagtattgatatcgttcccttgccacataccatttacatcgggaaaagctgttttgataat
57 E T S K T C T P S I D I V P L P H T I Y I G K S C F D N
11937 tgcttgagagatattagagaatag 11914
85 C L R D I R E *
182ORF024
6174 atgcttgtaactatctcatctttaaaaacgaagaaacttatcctagtaaatggcagtatgcctttgttactgatattgaata a
1 M L V T I S S L K T K K L I L V N G S M P L L- L I L ~N V
6258 agaatgacaacacaagtttcgttacctttgaaattgatgttttacaaacttatcgtttcgatattggtatacgagaaagtttca
29 R M T T Q V S L P L K L M F Y K L I V S I L V Y E K V S
6342 ttgcaaaagaacaccctcaactttattattcgaatggaatacctttcattaatacaattgaagagtcgcttgattacggtagag
57 L Q K N T L N F I I R M E Y L S L I Q L K S R L I T V E
6426 aatacacaacaacaaatgtaa 6446
85 N T Q Q Q M *
182ORF025
548 atgggtcgaaaactaatgcaacgaaacgtaacatcaactaaagtagaattctcagaagttatcgtacaagatggagcgccaaca
1 M G R K L M Q R N V T S T K V E F S E V I V Q D G A P T
632 at gtaccatgcgaaccagttgtcttaacaggaaaactttcagaagaaaaagctttatcagcgatcaaacgtaaaaaccctgat
29 I V P C E P V V L T G K L S E E K A L S A I K R K N P D
716 aaaaacgtagttgtaacaaatgtttcacatgaaacagcgctttacacaatgccagtcgataaatttatcgagttagcagacaaa
57 K N V V V T N V S H E T A L Y T M P V D K F I E L A D K
800 tcaacacaagcctaa 814
85 S T Q A *
182ORF026
13259 atggaaattatttggtctgccgtttcctgcatgcgtgccaaaaagttgtccactcatgaaacttttaggatcaagatttgtatt
1 M E I I W S A V S C M R A K K L S T H E T F R I K I C I
13175 cttgattggggttccatagcaacgttttacgccaccgccaccgctaccgtcaatatgcttactataagcttgtgcaagatcaag
29 L D W G S I A T F Y A T A T A T V N M L T I S L C K I K
13091 tctttcttcgatatgaagtttaccagggcgttcccagtgacacataaaattaattgtagcaacatcaatatttgcagattgttt
57 S F F D M K F T R A F P V T H K I N C S N I N I C R L F
13007 aaatgctga 12999
85 K C *
182ORF027
14896 atgaacatgattgtatgttcctctaatatgatcgagttgtgttggatcagacaattcaaagatcgtttctccgtcaaaataaaa
1 M N M I V C S S N M I E L C W I R Q F K D R F S V K I K
14812 cacgcttgtgctacctgttgtaagagtgaaaataaactcatcacttgtggcgtcaaatattttattttcaggaggaacaatttg
29 H A C A T C C K S E N K L I T C G V K Y F I F R R N N L
14728 atcctgttttttacctacatacagatcccatgtattaccatcgccatagaaaacattcaaatcaagattgccgttgtatcctgg
57 I L F F T Y I Q I P C I T I A I E N I Q I K I A V V S W
14644 taa 14642
85 *
182ORF028
14430 atgtttataataaaacaggcgttaaagcatggttttatacgtatacagcaaacctcaatacaactgatttttctagtattgcaa
1 M F I I K Q A L K H G F I R I Q Q T S I Q L I F L V L Q
14514 aaggcgattatggtttatgggttgctgaatatggatcaaatcaaccacaaggctactctcaaccagcgccacctaaaacaaata
29 K A I M V Y G L L N M D Q I N H K A T L N Q R H L K Q I
14598 attttccaattgttgcctgttttcagtttacaagtaaaggacgtttaccaggatacaacggcaatcttgatttga 14672
57 I F Q L L P V F S L Q V K D V Y Q D T T A I L I *
182ORF029
17606 atgaatgaaccgatcgtatacacagaaatttattcaaataacgtggtatgtatgaaaatttttagagatgaggataaacttagt
1 M N E P I V Y T E I Y S N N V V C M K I F R D E D K L S
17522 aaattcctctatttagaatttgaggtggatgaggctaaaaagttacttgaaaataaaacaatttcatttgatgataactggact
29 K F L Y L E F E V D E A K K L L E N K T I S F D D N W T
17438 ttctcaataaattatccagaatattaa 17412
57 F S I N Y P E Y *
182ORF030
16429 atggctacattctacaaggaaccaatatacgatatcacagtattttatatagatggttgggaggttttgatacacaaaaccgaa
1 M A T F Y K E P I Y D I T V F Y I D G W E V L I H K T E
16345 cctctcaccttaacaaaagcattaaaatatagccgtatatacctagaaatggatatagtgaattgcgttagaatagaaagaaat
29 P L T L T K A L K Y S R I Y L E M D I V N C V R I E R N
16261 ggacgtcc atagctacattttacagggaattattaaaactgtataaggagaaagaactatga 16199
57 G R P I A T F Y R E L L K L Y K E K E L *
182ORF031
8603 atgttacctgaactttcaatctgttcattgttagataagacttctgatgtttgtacacgtgcagtcttatctacgttagcattg
1 M L P E L S I C Ξ L L D K T S D V C T R A V L S T L A L
8519 ttgatacctagaaaagttaacacttcattccatacttcgttcaattctgatcgtagtttatctactacatatggagcatttgtt
29 L I P R K V N T S F H T S F N S D R S L S T T Y G A F V
8435 tgccatacattaaaagattcgtcaaactccatatctttatccacaaaaacagcctga 8379
57 C H T L K D S S N S I S L S T K T A *
182ORF032
11413 atgtttcatcaaaaacaacttgtttcgggttcgtttcagggtgcaataggtaattcttttgattttcttctttcatcttgttca
1 M F H Q K Q L V S G S F Q G A I G N S F D F L L S S C S
11329 tttgaatatcaattcgttcttccatatgaacctccttattttagagggaaaacgcaattatctagggatagatattcgatttta
29 F E Y Q F V L P Y E P P Y F R G K T Q L Ξ R D R Y S I L
11245 cctacattgtcatttacagttactttgccatcaggtgtgattgatacataa 11195
57 P T L S F T V T L P S G V I D T *
182ORF033
4942 atgtcaacaaaaatttcttcaatcgttcgacctaaaggcatgtttccttttttaaacattttcaaagggttacgccaagatttg
1 M S T K I S S I V R P K G M F P F L N I F K G L R Q D L
4858 tatcggataactactttaccaatacggtcaactaaagttgaaataaattcgttttttactacgtctaaacgtgtgatccctgca
29 Y R I T T L P I R S T K V E I N S F F T T S K R V I P A
4774 ccaaccgcttcgatgttatctgcatttggcataggtacgttcgcctga 4727 - —
57 P T A S M L S A F G I G T F A * __
182ORF034
6160 gtgtttatctactctaaaaactcccccgagttgtgtatccctttgataagaacaatctctattctcgttaagaacaggaaacga
1 V F I Y S K N S P E L C I P L I R T I S I L V K N R K R
6076 attaaagtacgattcctgttcctgttgagttttaaaccatcttgtgtgtgtataggtgttatcaaaaggcacgttagccaacaa
29 I K V R F L F L L S F K P S C V C I G V I K R H V S Q Q
5992 ttttacatttgtataccttcttgccataattgtcctccttag 5951
57 F Y I C I P S C H N C P P *
182ORF035
15758 atggcgcataagaaac ac atttttacttctcttttcaataaacgtatcactatcattgacaaactcattgttgatactaaaa
1 M A H K K L L F L L L F S I N V S L S L T N S L L I L K
15674 tcttcgtattctgttccacgaatcaatctaccaaaaggtgtttctctcttcacttctgcaaagtcttttgaatcacacaattca
29 S S Y S V P R I N L P K G V S L F T S A K S F E S H N S
15590 atcaatatacctcgatcttga 15570
57 I N I P R S *
182ORF036
2315 atgtctgtgctgccttgcattttacaccactcaaaaaaagaatcgatttctaaaccgaacgtcatattgtcaacgttgtcta a
1 M S V L P C I L H H S K K E S I S K P N V I L S T L S I
2231 tcgcatacgccccacgaccatacacgacaatcgttgagatcagttgttgtttcaaagtcgccagtatatttcttaatcataatt
29 S H T P H D H T R Q S L R S V V V S K S P V Y F L I I I
2147 cttctcctgtttctgaattaa 2127
57 L L L F L N *
182ORF037
12280 gtgagttacgacaataaacatctacatcaatataagcttgatccacatcttgaaactcaaacaaagcgtttctatttccgtatg
1 V S Y D N K H L H Q Y K L D P H L E T Q T K R F Y F R M
12196 ctagaaaatggttgttgttttggacatgcaagttatgcgcatatcctcgataataacttacgccaccttcgattgtgttaccag
29 L E N G C C F G H A S Y A H I L D N N L R H L R L C Y Q
12112 aaatttcacagaaattaa 12095
57 K F H R N *
182ORF038
14769 gtgatgagtttattttcactcttacaacaggtagcacaagcgtgttttattttgacggagaaacgatctttgaattgtctgatc
1 V M S L F S L L Q Q V A Q A C F I L T E K R S L N C L I
14853 caacacaactcgatcatattagaggaacatacaatcatgttcatggaaaagaaatcccatcaatggtgtggacacctgaacaat
29 Q H N S I I L E E H T I M F M E K K S H Q W C G H L N N
14937 ttgatatttacttaa 14951
57 L I F T *
182ORF039
9992 atgttgctgatgatcgaacattttggtataagattcaacgcgacaatactgattatggagccgatcctattgacacgttacgta
1 M L L M I E H F G I R F N A T I L I M E P I L L T R Y V
10076 ttgttgcaatcaataaagttagtggctggaataccgctacaggagatatttatcttaacattaaaggaacggagggtgtataat
29 L L Q S I K L V A G I P L Q E I F I L T L K E R R V Y N
10160 ggcagacattag 10171
57 G R H *
182ORF040
16202 atgagaaaagatttcgtctacattaacacacccgatccaaaagcaaacaaaaaggcgttagcaaaaatcactaacgccaaagaa
1 M R K D F V Y I N T P D P K A N K K A L A K I T N A K E
16118 ccaaaacaaaactatcgcagactacaattactatgttatctactattcatcattgtaatagaactaatcgtggtagctctacta
29 P K Q N Y R R L Q L L C Y L L F I I V I E L I V V A L L
16034 aaatag 16029
57 K *
182ORF041
3886 atggaactatataaagcaatgtttatcgtacgtgatgaaggtacta tgacggttacgatactgaacactatgtagatatttct
1 M E L Y K A M F I V R D E G T I D G Y D T E H Y V D I S
3970 ttacatgactttgaagaaatatatggaaaagaaacacgtgaaattgaagcagtaacattagtaaaaacaggaaatttaaaaaaa
29 L H D F E E I Y G K E T R E I E A V T L V K T G N L K K
4054 taa 4056
57 *
182ORF042
10832 gtgtcctctaaactgcttactcgaacagctagatcagttgcacttttttgtgctgaatcagctgtgtttttagctgccgttgca
1 V S S K L L T R T A R S V A L F C A E S A V F L A A V A
10748 actgcgctgatactgtttgctgttgttttcgcttgttctgcggtttgttgtgctgtaccagccttcgtcaacgcttga 10671
29 T A L I L F A V V F A C S A V C C A V P A F V N A *
182ORF043
10652 gtgtcaatttctgttgacaaaccagcagcagtttctttagcttgttgtgcaagtgttttagcttcactagcgttacctcttatt
1 V S I S V D K P A A V S L A C C A S V L A S L A L P L I
10568 tcagcattaactaattcaagattagagtcgcctgttagaacatttttagcaattgtaacaggcattaaacgattatga 10491
29 S A L T N S R L E S P V R T F L A I V T G I K R L *
182ORF044
6457 atgaaaagttgttacatttgttgttgtgtattctctaccgtaatcaagcgactcttcaattgtattaatgaaaggtattccatt
1 M K S C Y I C C C V F S T V I K R L F N C I N E R Y S I
6373 cgaataataaagttgagggtgttcttttgcaatgaaactttctcgtataccaatatcgaaacgataagtttgtaa 6299
29 R I I K L R V F F C N E T F S Y T N I E T I S L *
182ORF045
6729 atgaatggtatacctgtatacgacgttacatacatcccgactatcttatttaaaaaaggttctttcgttgtaagaaacgccatg
1 M N G I P V Y D V T Y I P T I L F K K G S F V -V R_N 7. M
6645 tactctccaaaattagcattgcctgccccatttggtttgtatacctccccacttgaattgataggaagtaaataa 6571
29 Y S P K L A L P A P F G L Y T S P L E L I G S K *
182ORF046
2372 atggtttcaaatggtgtaaagaagcaaaagaagatcgaacattctccacactcatatcaaatatgggtcaatggtatgctttgg
1 M V S N G V K K Q K K I E H S P H S Y Q I W V N G M L W
2456 aaatttgttgggaagttaattacacaacaacaaaatcaggtaaaacgaaaaaagagaaatctcgaacaataa 2527
29 K F V G K L I T Q Q Q N Q V K R K K R N L E Q *
182ORF047
13353 atgctcccattgttccaacatgtgttactgttccatcgcaacatgcaatcatttcattgccagggtgatcaattgaaccaaagt
1 M L P L F Q H V L L F H R N M Q S F H C Q G D Q L N Q S
13269 ccaaaccatcatggaaattatttggtctgccgtttcctgcatgcgtgccaaaaagttgtccactcatga 13201
29 P N H H G N Y L V C R F L H A C Q K V V H S *
182ORF048
3395 atgtcagggtttgttccgaactttccatacaagctatttaacatacctttggcgttagcttttctagccccttcggtggtgttc
1 M S G F V P N F P Y K L F N I P L A L A F L A P S V V F
3311 tttacttcgatccatttatcgatccagcctttgaacatatcacaagaagctttgaacatatatccgtaa 3243
29 F T S I H L S I Q P L N I S Q E A L N I Y P *
182ORF049
1578 atgttgcaatctcaagagcgcaaatcaaagaagcgcaaattaaaacagagcaagctcaaaaagcgaaagaagaacactacaaag
1 M L Q S Q E R K S K K R K L K Q S K L K K R K K N T T K
1662 agcttaacaaagttgaagttaagaagcccacagaaaacacaattgtcacaccaactattttaa 1724
29 S L T K L K L R S P Q K T Q L S H Q L F *
182ORF050
8012 atggttatcttggtttctttaaagaccctacacttgggttcatggtttgcgcaggggcagaagatggtcaaatcgatcattatc
1 M V I L V S L K T L H L G S W F A Q G Q K M V K S I I I
8096 acaaccctattttctttacagcaaacgaagcaatgtatcacaagagatatcctgttttaa 8155
29 T T L F S L Q Q T K Q C I T R D I L F *
182ORF051
9390 atgcttctgaaaaagaaacaaagaacacagacattaataaagatcaaaatcaaaccaaagatacgattacacgatataaaggta
1 M L L K K K Q R T Q T L I K I K I K P K I R L H D I K V
9474 aaaagggaaacactgattatgctgacttactcgaaaaatatcgtagaagtgttttga 9530
29 K R E T L I M L T Y S K N I V E V F *
182ORF052
4096 gtgatagttgacaagagtcaaatttggcgagattgggcgaatgtacacgtgaaatatcgtgcgctcccgttaagttatggacac
1 V I V D K S Q I W R D W A N V H V K Y R A L P L S Y G H
4180 ataaacgttttgaccgtcaaccaatcgcaaaaaccttttaggagtagcccttaa 4233
29 I N V L T V N Q S Q K P F R S S P *
182ORF053
15656 gtggaacagaatacgaagattttagtatcaacaatgagtttgtcaatgatagtgatacgtttattgaaaagagaagtaaaaata
1 V E Q N T K I L V S T M S L S M I V I R L L K R E V K I
15740 gtagtttcttatgcgccattgcttttgaagggaaaatctttgggtattggatag 15793
29 V V S Y A P L L L K G K S L G I G *
182ORF054
8136 gtgatacattgcttcgtttgctgtaaagaaaatagggttgtgataatgatcgatttgaccatcttctgcccctgcgcaaaccat
1 V I H C F V C C K E N R V V I M I D L T I F C P C A N H
8052 gaacccaagtgtagggtctttaaagaaaccaagataaccattagtgtgtaa 8002
29 E P K C R V F K E T K I T I S V *
182ORF055
8324 atgaaaagaaatacttctcattgctacaagcttataaccaaattgacgaaaataatcaggctgtttttgtggataaagatatgg
1 M K R N T S H C Y K L I T K L T K I I R L F L W I K I W
8408 agtttgacgaatcttttaatgtatggcaaacaaatgctccatatgtag 8455
29 S L T N L L M Y G K Q M L H M »
182ORF056
6549 gtggcccatctcctttttcctattatttacttcctatcaattcaagtggggaggtatacaaaccaaatggggcaggcaatgcta
1 V A H L L F P I I Y F L S I Q V G R Y T N Q M G Q A M L
6633 attttggagagtacatggcgtttcttacaacgaaagaaccttttttaa 6680
29 I L E S T W R F L Q R K N L F *
182ORF057
8264 atgtccgccatatctaaagcaaaacgatgtaaacttggtaacgtaggaactttcaagtcattattatacaacatgatacatttt
1 M S A I S K A K R C K L G N V G T F K S L L Y N M I H F
8180 gatttatcatcatcatcatcatatcttaaaacaggatatctcttgtga 8133
29 D L S S S S S Y L K T G Y L L *
182ORF058
5176 gtgtattcaaattcgcttacttcgtcacctgtgtataaagcgttcattacaccagcaacgaaactattgaaattatcccatgaa
1 V Y S N S L T S S P V Y K A F I T P A T K L L K L S H E
5092 gtaaatgctttttctaaccatgcttcttggatcgtttgtttgtag 5048
29 V N A F S N H A S W I V C L *
182ORF059
15876 atggtctttcgtagtcattgcataaaaatgatttgtatttggttgataatcataactcacatagacacaacctgtttcagcgtc
1 M V F R S H C I K M I C I W L I I I T H I D T T C F S V
15792 tatccaatacccaaagattttcccttcaaaagcaatggcgcataa 15748
29 Y P I P K D F P F K S N G A * - --
182ORF060 _
15404 gtgatttttgatttctcaattaaaaactcatcaaacaaaattgtacgaacttcgggatattcattagatttttcaattccccac
1 V I F D F S I K N S S N K I V R T S G Y Ξ L D F S I P H
15320 gtactaagtggaacagcccaacccattaatttatcatcacaatag 15276
29 V L S G T A Q P I N L S S Q *
182ORF061
2102 atgaggggacttctccacctgtttcagactcgatcacttttgcaatcttactgtaaacttgttcttttttctgttgtacttctg
1 M R G L L H L F Q T R S L L Q S Y C K L V L F S V V L L
2018 cttcgtcataaatgtagtcaaggttcatgtctaagaagttactaa 1974
29 L R H K C S Q G S C L R S Y *
182ORF062
1992 atgtctaagaagttactaacatatgttttcataaatagatcaagccccattgagtcaagtaaacgaacaatatcgtcagcgtct
1 M Ξ K K L L T Y V F I N R S S P I E S S K R T I S S A S
1908 gaattgaaaatagtaaacatcgcttgtctgaaattgtcgtaa 1867
29 E L K I V N I A C L K L S *
182ORF063
14306 gtgtaccttctaaacccctctcatgcgcaaaatgatacacaccaatctttttacctaaagacaaagcttgttgaaatgctcggt
1 V Y L L N P S H A Q N D T H Q S F Y L K T K L V E M L G
14222 cacaatcagggtttacataacctgttccgcctgttgctttaa 14181
29 H N Q G L H N L F R L L L *
182ORF064
7356 atgatgttagtcaaaccaacaaaagggttgttacttgctaaggctgaaaagatcgctcctcctgtactcattgcactgtttccc
1 M M L V K P T K G L L L A K A E K I A P P V L I A L F P
7272 ataccatgtctgaaagtattgcgaatgttttgctcttga 7234
29 I P C L K V L R M F C S *
182ORF065
3582 atgaatgctatctgtatcacaataaataatgcgatcaaaacatttttgagcggttgtaatggtagtatatctaccccaagccgt
1 M N A I C I T I N N A I K T F L S G C N G S I S T P S R
3498 cacaaaactagcaagcggaacataaacaggatctcttaa 3460
29 H K T S K R N I N R I S *
182ORF066
4234 atgtggctactcttttttgtgtttcacagaattatgtttcacgtgaaacagtttttatggtataatagaatcaaaaggaggtgg
1 M W L L F F V F H R I M F H V K Q F L W Y N R I K R R W
4318 agattatggaaattaaagaacatgaatcaattttaa 4353
29 R L W K L K N M N Q F *
182ORF067
13882 atgatacctgctttagctttaaaactactaaagtcgatttccttgttaaatttggcaattgtaaaacctaacacaaaatcgata
1 M I P A L A L K L L K S I S L L N L A I V K P N T K S I
13798 atcattgcaaccattaaccatataatcaaaccataa 13763
29 I I A T I N H I I K P *
182ORF068
7267 atgtctgaaagtattgcgaatgttttgctcttgagcaatcaaggagtttttgtttccttgcatgaatgcagaagcatagtcaga
1 M S E S I A N V L L L S N Q G V F V S L H E C R S I V R
7183 tttaactcctacatcgttaggatcattatcgattaa 7148
29 F N S Y I V R I I I D *
182ORF069
5027 gtggaacaatgtttttacatcgggaacttcctgtttaaatacccctgtaacagactcgtcagggttgaacttatgttcctgtgc
1 V E Q C F Y I G N F L F K Y P C N R L V R V E L M F L C
4943 aatgtcaacaaaaatttcttcaatcgttcgacctaa 4908
29 N V N K N F F N R S T *
182ORF070
1031 gtgatggttcggctccaccaaaaccagaaacttcgtctgagtaaactagaatatctttcaattctagtacttcgccaattgcgt
1 V M V R L H Q N Q K L R L Ξ K L E Y L S I L V L R Q L R
947 tacgtaacggaattgaagccccgtttgtggcattga 912
29 Y V T E L K P R L W H *
182ORF071
11741 atggttttgcattatggttgccacaaggcgctcaaagtggtaaaggaattttctttaatgatactcgcaattacaatcgttttg
1 M V L H Y G C H K A L K V V K E F S L M I L A I T I V L
11825 actttgatttgtttgttcgtaactgtactttaa 11857
29 T L I C L F V T V L *
182ORF072
11723 atgtttacattaaatgccgtcattgtttcaaactttaatgtcgtttctcccgatcctaagaaagtaactacaggtacatcacgt
1 M F T L N A V I V S N F N V V S P D P K K V T T G T S R
11639 ttcaattcaatggtgttagcaaagcgataa 11610
29 F N S M V L A K R *
182ORF073
2876 gtgaagccgcctttgtatgctttacgtaagtctttatcaaaccctaaagacaaaataggaaaccattgtttgaaagttgatttt
1 V K P P L Y A L R K S L S N P K D K I G N H C L K V D F
2792 ccatgtgtagcttttagccaatctttgtaa 2763
29 P C V A F S Q S L *
182ORF074
8923 gtgattgataaattttgtttcaaattctgctcgttttgtttcgtcataaaacggataatcaaaatcaaacaattgttttcggcc
1 V I D K F C F K F C S F C F V I K R I I K I K Q L F S A
8839 aacttcaatacgttcttttcgagataa 8813
29 N F N T F F S R * - —
182ORF075 _
7463 gtgttacattatctggaatattttcgatatctgccactttacctgccaagaggttcaaaccgttttctttttcagaaacatagt
1 V L H Y L E Y F R Y L P L Y L P R G S N R F L F Q K H S
7379 tgtttacttgttgtcctgctcccatga 7353
29 C L L V V L L P *
182ORF076
2426 atgagtgtggagaatgttcgatcttcttttgcttctttacaccatttgaaaccatttttgaataaccatgaaagcataaactct
1 M S V E N V R S S F A Ξ L H H L K P F L N N H E S I N S
2342 ccgtcaaatttttcgttgtggaaataa 2316
29 P S N F Ξ L W K *
182ORF077
11858 atgaaggaacgtatgttgttgttgctagaggtagaggggttacatttgaaaattgtctattctctaatatctctcaagcaatta
1 M K E R M L L L L E V E G L H L K I V Y S L I Ξ L K Q L
11942 tcaaaacagcttttcccgatgtaa 11965
29 S K Q L F P M *
182ORF078
7671 gtgcctacaatatttggttcttttaatttaatgaaattccatgcttttcttgtttgtaagtttggtgtagctactcgattgctc
1 V P T I F G S F N L M K F H A F L V C K F G V A T R L L
7587 tttgtgccatacattgagaagtaa 7564
29 F V P Y I E K *
182ORF079
7488 gtgaaagataagtttgatccaagctgtgttacattatctggaatattttcgatatctgccactttacctgccaagaggttcaaa
1 V K D K F D P S C V T L S G I F S I S A T L P A K R F K
7404 ccgttttctttttcagaaacatag 7381
29 P F S F S E T *
182ORF080
4473 gtgtgctatttgctgatgtcaaagcttcagttgttgctccgtagtcttctcgcaatgcttcaagatgttctacaatctttgatc
1 V C Y L L M S K L Q L L L R S L L A M L Q D V L Q S L I
4389 ttgcttcaccgtctgtga 4372
29 L L H R L *
529
Table 24
Sequence similarities phage 182 and public databases
Phage: 182
Database: nr
Query= sid| 110156 | lan| 182ORF001 Phage 182 ORF| 5966-7780 | 2 (604 letters) gi|138124|sp|P07534|VG9_BPPZA TAIL PROTEIN (LATE PROTEIN GP9) >. 384 e-105 gi|l38123|sp|P04331|VG9_BPPH2 TAIL PROTEIN (LATE PROTEIN GP9) >. 374 e-103 gi|l429238|gnl|PID|ell73412 (X99260) tail protein [Bacteriophag . 346 3e-94 gi|215339 (M12456) p9 tail protein [Bacteriophage phi-29] >gi|2. 208 8e-53 gijll81970|gnl|PID|e221269 (Z47794) tail protein [Bacteriophage. 62 8e-09 gi|ll81968|gnl|PID|e221267 (Z47794) tail protein [Bacteriophage. 56 6e-07 gij 2500030 jsp|Q59968|CARA_SULSO CARBAMOYL-PHOSPHATE SYNTHASE SM. 49 8e-05
Query= sid| 110157 | lan| 182ORF002 Phage 182 ORF| 2152-3873 | 1 (573 letters) gi|118848|sp|P19894|DPOL_BPM2 DNA POLYMERASE >gi | 76896 | pir| | JQ0.. 665 0.0 gijl429230|gnl|PID|ell73404 (X99260) DNA polymerase [Bacterioph.. 657 0.0 gijll8849|sp|P03680|DPOL_BPPH2 DNA POLYMERASE (EARLY PROTEIN GP .. 654 0.0 gi|H8851|sp|P06950|DPOL_BPPZA DNA POLYMERASE (EARLY PROTEIN GP .. 654 0.0 gij 15732 (X53371) DNA polymerase (AA 1-575) [Bacteriophage phi-29 651 0.0 gij 15734 (X53370) DNA polymerase (AA 1-575) [Bacteriophage phi-29 651 0.0 gijl572479|gnl|PID|e242301 (X96987) DNA polymerase [Bacteriopha. 565 e-160 gijl072656|pir| |S51275 DNA polymerase - phage CP-1 >gi | 836593 |g. 301 le-80 gi|H8847|sp|P22374|DPOM_ASCIM PROBABLE DNA POLYMERASE >gi|8385. 71 3e-ll giJ461962JspjP33537JDPOM_NEUCR PROBABLE DNA POLYMERASE >giJ2833. 65 le-09 giJ461963 j sp j P33538 |DPOM_NEUIN PROBABLE DNA POLYMERASE >gi j 1018. 62 le-08 gi 11084487 |pir I I S41618 DNA polymerase - slime mold (Physarum po. 61 3e-08 gi|2435429 (AF012250) unassigned reading frame (possible DNA po. 61 3e-08 gi|578157|gnl|PID|e246743 (X52106) DNA polymerase [Neurospora i. 59 le-07 giJ2147969|pir I |S72369 probable DNA-polymerase - Gelasinospora . 58 2e-07 gi I 2147968 jpi j j S62752 probable DNA-polymerase - Gelasinospora . 58 2e-07 gi 13511140 (AF061244) B type DNA polymerase [Agrocybe aegerita] 57 3e-07 gi|118850|sp|P10479|DPOL_BPPRD DNA POLYMERASE (PROTEIN PI) >gi | . 56 6e-07 gi 1578144 (X63909) putative DNA-polymerase, B-type [Morchella c. 47 3e-04 giJ232013|sp|P30322|DPOM_AGABT PROBABLE DNA POLYMERASE >gi|3208. 46 6e-04
Query= sid| 110159 | lan| 182ORF004 Phage 182 ORF| 4626-5954 | 3 (442 letters) gi|l38117|sp|P13849|VG8_BPPH2 MAJOR HEAD PROTEIN (LATE PROTEIN . 309 2e-83 gi j 138118 j sp j P07531 j VG8_BPPZA MAJOR HEAD PROTEIN (LATE PROTEIN . 305 3e-82 gijl429236|gnl|PID|ell73410 (X99260) major head protein [Bacter. 300 le-80 gijll81958|gnl|PID|e221257 (Z47794) major head protein [Bacteri. 152 6e-36
Query= sid| 110160 | lan| 182ORF005 Phage 182 ORF| 12651-13700 | 3 (349 letters) gi|l37932|sp|P15132 |VG13_BPPH2 MORPHOGENESIS PROTEIN 1 (LATE PR. 52 8e-06 gijl429242 |gnl| PID|ell73416 (X99260) morphogenesis protein [Bac. 48 7e-05 gi|l37933|sp|P07538|VG13_BPPZA MORPHOGENESIS PROTEIN 1 (LATE PR. 47 2e-04
Query= sid| 110161 | lan| 182ORF006 Phage 182 ORF| 14995-16026 | 1 (343 letters) gi|l37944|sp|P11014 |VG16_BPPH2 ENCAPSIDATION PROTEIN (LATE PROT. 402 e-111 gijl37945|spjP0754l|VG16_BPPZA ENCAPSIDATION PROTEIN (LATE PROT. 402 e-111 gijl429245|gnl|PID|ell73419 (X99260) encapsidation protein [Bac. 381 e-105 gi 11181972 jgnlj PID je221271 (Z47794) encapsidation protein [Bact. 159 2e-38
Query= sid| 110162 | lan| 182ORF007 Phage 182 ORF| 7795-877511 (326 letters)
gι|l429239|gnll PID|ell73413 (X99260) upper collar protein [Bact 271 5e-72 gι|137915|sp|P07535|VG10_BPPZA UPPER COLLAR PROTEIN (CONNECTOR 256 le-67 gij 137914 jspj P04332JVG10_BPPH2 UPPER COLLAR PROTEIN (CONNECTOR 256 2e-67 gij 1181960 |gnl| PID| e221259 (Z47794) connector protem [Bacter o 148 6e-35
Query= sιd| 110163 | lan| 182ORF008 Phage 182 ORF| 14105-14983 | 2 (292 letters) gi 4210750|gnl|PID|el374037 (AJ132604) LysL protein [Lactococcu 139 2e -32 462559|sp|P34020|LYC_CLOAB AUTOLYTIC LYSOZYME (1, 4-BETA-N-AC 75 8e -13 gi 2327014 (U82823) putative lysozyme [Saccharopolyspora erythr 64 2e -09 gi 126652|sp|P25310|LYCM_STRGL LYSOZYME Ml PRECURSOR (1,4-BETA- 60 2e -08 gi 127789 I sp|P19386JLYCA_BPCP9 LYSOZYME (ENDOLYSIN) (MURAMIDASE 60 2e -08 gi 67761 |pιr I I MUBPCP N-acetylmuramoyl-L-alanme amidase (EC 3 5 59 3e -08 gi 4105636 (AF049087) lys [Leuconostoc oenos bacteriophage 10MC] 59 3e -08 gi 623084 (L02496) muramidase, muramidase [Bacteriophage LL-H] 57 le -07 gi 127787|sp|P15057|LYCA_BPCPl LYSOZYME (ENDOLYSIN) (MURAMIDASE 57 2e -07 gi 126597 |sp|P0072lJLYCH_CHASP N, O-DIACETYLMURAMIDASE (LYSOZYME 57 2e -07 i 127788]sp|P19385|LYCA_BPCP7 LYSOZYME (ENDOLYSIN) (MURAMIDASE 57 2e- -07 gi 67762 |pιr j |MUBPC7 N-acetylmuramoyl-L-alamne amidase (EC 3 5 56 3e- 07 gi 3025168|sp|P7642l|YEGX_ECOLI HYPOTHETICAL 32 0 KD PROTEIN IN 53 2e- 06 gi 4204413 (AF047001) Lys44 [Oenococcus oeni temperate bacterio 53 3e- 06 gi 2116978|gnl|PID|dl020940 (D88151) cortical fragment-lytic en 52 5e- 06 gi 2392844 (AF011378) lysin [Bacteriophage ski] 48 8e- 05
Query= sιd| 110164 | lan| 182ORF009 Phage 182 ORF| 8765-9601 | 2 (278 letters)
.|l429240|gnl|PID|ell73414 (X99260) lower collar protein [Bact 180 le -44 .|l3792l|sp|P04333|VGll_BPPH2 LOWER COLLAR PROTEIN (LATE PROTE 171 5e -42 . j 215341 (M12456) pll lower collar protein [Bacteriophage phi-29] 98 9e -20 . J224162 |prf I 11011232B protein pll, lower collar [Bacteriophage 97 le -19 . j 535260 (Z30339) STARP antigen [Plasmodium reichenowi] 50 le -05 L 14049753 (AF063866) ORF MSV230 hypothetical protein [Melanopl 49 4e -05 L J2131557 |pιr I |S70306 hypothetical protein YEL077c - yeast (Sa 48 5e -05 LJ 131782 |sp|P12753|RA50_YEAST DNA REPAIR PROTEIN RAD50 (153 KD 48 7e- -05 . |2131309|pιr I |S70305 hypothetical protein YBL113c - yeast (Sa 47 2e- -04 L 1499325 (Z26314) STARP antigen [Plasmodium falciparum] 46 3e 04 | 3845171 (AE001391) πbosome releasing factor (OO, TP) [Plasm 46 3e -04 -J731903|sp|P40434|YIR7_YEAST HYPOTHETICAL 197 5 KD PROTEIN IN 45 5e- -04 -|l632829|gnl|PID|e276379 (Y08924) AARP2 protein [Plasmodium f 45 5e- 04 .jll7649θjsp|P40889|YJW5_YEAST HYPOTHETICAL 197 6 KD PROTEIN I 45 5e- 04 -jl077300|pιr| |S51848 hypothetical protein HRD1054 - yeast (Sa 45 Se- 04 L 12425143 (AF020407) WimA [Dictyostelium discoideum] 45 6e- 04 LJll8196l|gnl|PID|e221260 (Z47794) collar protein [Bacteriopha 45 6e- 04 j 2132657 jpir | | S64819 probable membrane protein YLL067C - yeas 45 8e- 04 -I 213304l|pιr j |S65341 probable membrane protein YPR204W - yeas 45 8e- 04 -I 730275 I sp| P39793 | PBPA_BACSU PENICILLIN-BINDING PROTEINS 1A/1 45 8e- 04
Query= sιd| 110165 | lan| 182ORF010 Phage 182 ORF| 1310-2155 ] 2 (281 letters) gι|l35604|sp|P06812|TERM_BPNF DNA TERMINAL PROTEIN >gι| 75815 | pi 69 3e-ll gιjl572478|gnl|PID|e242334 (X96987) terminal protein [Bacteπop 65 3e-10 gι|142923l|gnl|PIDJell73405 (X99260) terminal protein [Bacterio 64 le-09
Query= sιd| 110166 | lan| 182ORF011 Phage 182 ORF| 9607-10158 | 1 (183 letters) gi 1137928 I sp I P0753 IVG12_BPPZA PRE-NECK APPENDAGE PROTEIN (LATE 51 6e-06 gij 14292411 gnl I PID|ell73415 (X99260) pre-neck appendage protein 51 6e-06 gι|l37927|sp|P20345|VG12_BPPH2 PRE-NECK APPENDAGE PROTEIN (LATE 50 le-05
Query= sid] 110169 | lan| 182ORF014 Phage 182 ORF| 13716-14108 | 3 (130 letters) gι|l37936|sp|P11188|VG14_BPPH2 LYSIS PROTEIN (LATE PROTEIN GP14 97 6e-20 gιjl37938|spjP07539|VG14_BPPZA LYSIS PROTEIN (LATE PROTEIN GP14 96 8e-20 gι|1429243|gnl|PID|ell73417 (X99260) lysis protem [Bacteriopha 96 8e-20 gij 215332 (M14782) lysis protein [Bacteriophage phi-29] 94 5e-19
Query= sιd| 110170 | lan| 182ORF015 Phage 182 ORF| 854-1225 | 2 (123 letters)
gi|l5670 (V01155) reading frame 10 (may be gene 4) [Bacteriopha... 70 5e-12 gi| 138072 |sp|P069S3|VG5A_BPPZA EARLY PROTEIN GP5A >gi | 75836 | ir ... 69 7e-12
Query= sid| 110174 | lan| 182ORF019 Phage 182 ORF| 4323-4613 | 3 (96 letters) gi|l429235|gnl|PID|ell73409 (X99260) head morphogenesis protein... 61 2e-09 gi|l38111|sp| P13848 | VG7_BPPH2 HEAD MORPHOGENESIS PROTEIN (LATE ... 57 3e-08 gi 1138112 jsp I P07533 JVG7_BPPZA HEAD MORPHOGENESIS PROTEIN (LATE ... 54 le-07
Query= sid| 110180 | lan| 182ORF025 Phage 182 ORF| 548-814 | 2 (88 letters) gi|138099|sp|P06955|VG6_BPPZA EARLY PROTEIN GP6 >gi | 75841 |pir| | ... 55 7e-08 gi| 138098 jsp|P03685|VG6_BPPH2 EARLY PROTEIN GP6 >gi j 75840 | pir j j ... 54 2e-07 gi j 1429234 |gnl| PID|ell73408 (X99260) gene 6 product [Bacterioph... 54 2e-07
Table 25
Homologies between 182 ORFs and proteins in public databases
Phage: 182
Database: Swissprot
Query= sid| 110156 | lan| 182ORF001 Phage 182 ORF| 5966-7780 | 2 (604 letters) gi|l38124|spiP07534|VG9_BPPZA TAIL PROTEIN (LATE PROTEIN GP9) 384 e-106 gijl38123 j sp j P04331 j VG9_BPPH2 TAIL PROTEIN (LATE PROTEIN GP9) 374 e-103 giJ2500030|sp|Q59968|CARA_SULSO CARBAMOYL-PHOSPHATE SYNTHASE SM. 49 2e-05
Query= sid| 110157 | lan| 182ORF002 Phage 182 ORF| 2152-3873 | 1 (573 letters) gi|H8848|sp|P19894|DPOL_BPM2 DNA POLYMERASE 665 0.0 gijll8849|sp|P0368θJDPOL_BPPH2 DNA POLYMERASE (EARLY PROTEIN GP2) 654 0.0 gij 118851 jsp|P06950 I DPOL_BPPZA DNA POLYMERASE (EARLY PROTEIN GP2) 654 0.0 gi 1118847 j sp j P22374 j DPOM_AΞCIM PROBABLE DNA POLYMERASE 71 7e-12 gi j 46196 j sp j P33537 j DPOM_NEUCR PROBABLE DNA POLYMERASE 65 3e-10 gij 461963 j sp j P33538 j DPOM_NEUIN PROBABLE DNA POLYMERASE 62 3e-09 gijll885θjspjpi0479JDPOL_BPPRD DNA POLYMERASE (PROTEIN PI) 56 2e-07 giJ232013|spjP30322|DPOM_AGABT PROBABLE DNA POLYMERASE 46 2e-04 gijll8887|sp|P10582|DPOM_MAIZE DNA POLYMERASE (S-l DNA ORF 3) 46 2e-04
Query= sid| 110159 | lan| 182ORF004 Phage 182 ORF|4626-5954 | 3 (442 letters) gi|l38117|sp|P13849|VG8_BPPH2 MAJOR HEAD PROTEIN (LATE PROTEIN .., 309 6e-84 gi j 138118 j s | P07531 j VG8_BPPZA MAJOR HEAD PROTEIN (LATE PROTEIN ... 305 7e-83
Query= sid| 110160 | lan| 182ORF005 Phage 182 ORF| 12651-13700 | 3 (349 letters) gi| 137932 | sp | P15132 | VG13_BPPH2 MORPHOGENESIS PROTEIN 1 (LATE PR... 52 2e-06 gijl37933 j sp j P07538 j VG13_BPPZA MORPHOGENESIS PROTEIN 1 (LATE PR... 47 6e-05
Query= sid| 110161 | lan| 182ORF006 Phage 182 ORF| 14995-16026 | 1 (343 letters) gi|l37945|sp|P07541|VG16_BPPZA ENCAPSIDATION PROTEIN (LATE PROT... 402 e-112 gijl37944|sp|P11014 jvG16_BPPH2 ENCAPSIDATION PROTEIN (LATE PROT... 402 e-112
Query= sid| 110162 | lan| 182ORF007 Phage 182 ORF| 7795-8775 | 1 (326 letters) gi|l37915|sp|P07535|VG10_BPPZA UPPER COLLAR PROTEIN (CONNECTOR ... 256 3e-68 gijl37914 j sp j P04332 j VG10_BPPH2 UPPER COLLAR PROTEIN (CONNECTOR ... 256 5e-68
Query= sid| 110163 | lan| 182ORF008 Phage 182 ORF| 14105-14983 | 2 (292 letters) gi|462559|sp|P34020|LYC_CLOAB AUTOLYTIC LYSOZYME (1, 4-BETA-N-AC. 75 2e -13 gi|l26652 j sp j P25310 | YCM_STRGL LYSOZYME Ml PRECURSOR (1,4-BETA-. 60 5e -09 gi j 127789 | sp j P19386 j LYCA_BPCP9 LYSOZYME (ENDOLYSIN) (MURAMIDASE. 60 5e -09 gij 127787 | s j P15057 j LYCA_BPCP1 LYSOZYME (ENDOLYSIN) (MURAMIDASE. 57 4e -08 gij 126597 j sp | P00721 JLYCH_CHASP N, 0-DIACETYLMURAMIDASE (LYSOZYME. 57 4e- -08 gij 127788 jspj i9385 j YCA_BPCP7 LYSOZYME (ENDOLYSIN) (MURAMIDASE. 57 5e- 08 gi|3025168|sp|P7642l|YEGX_ECOLI HYPOTHETICAL 32.0 KD PROTEIN IN. 53 5e- 07
Query= sid| 110164 | lan| 182ORF009 Phage 182 ORF| 8765-9601 | 2 (278 letters) gi|l37921|sp|P04333|VGll_BPPH2 LOWER COLLAR PROTEIN (LATE PROTE . 171 le-42 gij 131782 | spj P12753 JRA50_YEAST DNA REPAIR PROTEIN RAD50 (153 KD. 48 2e-05 gijll76490|sp|P40889|YJW5_YEAST HYPOTHETICAL 197.6 KD PROTEIN I. 45 le-04 giJ731903|sp|P40434|YIR7_YEAST HYPOTHETICAL 197.5 KD PROTEIN IN. 45 le-04 gi| 730275 j spj P39793 j PBPA_BACSU PENICILLIN-BINDING PROTEINS 1A/1. 45 2e-04 gijll68610|sp|P41696|AZFl_YEAST ASPARAGINE-RICH ZINC FINGER PRO. 44 3e-04
gi|731587|sp| 38900| H19_YEAST HYPOTHETICAL 70.1 KD PROTEIN IN ... 44 3e-04
Query= sid| 110165 | lan| 182ORF010 Phage 182 ORF| 1310-2155 | 2 (281 letters) gi|l35604|sp|P06812|TERM_BPNF DNA TERMINAL PROTEIN 69 8e-12
Query* sid| 110166 | lan| 182ORF011 Phage 182 ORF| 9607-10158 | 1 (183 letters) gi 1137928 I sp|P07537 I VG12_BPPZA PRE-NECK APPENDAGE PROTEIN (LATE... 51 2e-06 gi|137927Jsp|P20345JvG12_BPPH2 PRE-NECK APPENDAGE PROTEIN (LATE... 50 3e-06
Query= sid| 110169 | lan| 182ORF014 Phage 182 ORF| 13716-14108 | 3 (130 letters) gi|l37936|sp|P11188|VG14_BPPH2 LYSIS PROTEIN (LATE PROTEIN GP14) 97 2e-20 gij 137938 I spjP07539|VG14_BPPZA LYSIS PROTEIN (LATE PROTEIN GP14) 96 2e-20
Query= sid| 110170 | lan| 182ORF015 Phage 182 ORF| 854-1225 | 2 (123 letters) gi|l38072|sp|P06953|VG5A_BPPZA EARLY PROTEIN GP5A 69 2e-12
Query= sid| 110174 | lan| 182ORF019 Phage 182 ORF| 4323-4613 | 3 (96 letters) gi|l3811l|sp|P13848|VG7_BPPH2 HEAD MORPHOGENESIS PROTEIN (LATE ... 57 9e-09 gi j 138112 jsp I P07533JVG7_BPPZA HEAD MORPHOGENESIS PROTEIN (LATE ... 54 4e-08
Query= sid| 110180 | lan| 182ORF025 Phage 182 ORF| 548-814 | 2 (88 letters) gi|138099|sp|P06955|VG6_BPPZA EARLY PROTEIN GP6 55 2e-08 gi] 138098 | spj P03685 j VG6_BPPH2 EARLY PROTEIN GP6 54 5e-08
BLASTP 2.0 8 [Jan-05-1999]
Query= sιd| 110156 | Ian | 182ORF001 Phage 182 ORF| 5966-7780 | 2 (604 letters)
>gι|138124 |sp|P07534 |VG9_BPPZA TAIL PROTEIN (LATE PROTEIN GP9) >gι| 75849 |pιr| |WMBP9Z gene 9 protein - phage PZA >gι j 216058 (M11813) tail protein [Bacteriophage PZA] Length = 599
Score = 384 bits (975) , Expect = e-105
Identities = 231/610 (37%), Positives = 344/610 (55%), Gaps = 36/610 (5%)
Query. 6 TNVKLLANVPFDNTYTHTRWFKTQQEQESYFNSFPVLNENRDCSYQRDTQLGGVFRVDKH 65
TNV++LA+VPF N Y +TRWF + Q ++FNS + E ++Q + V Sbjct. 9 TNVRILADVPFSNDYKNTRWFTSSSNQYNWFNSKTRVYEMSKVTFQGFRENKSYISVSLR 68
Query. 66 KDALYACNYLIFKNEETYPSKWQYAFVTDIEYKNDNTSFVTFEIDVLQTYRFDIGIRESF 125
D LY +Y++F+N + Y +KW YAFVT++EYKN T++V FEIDVLQT+ F+I +ESF Sbjct: 69 LDLLYNASYIMFQNAD-YGNKWFYAFVTELEYKNVGTTYVHFEIDVLQTWMFNIKFQESF 127
Query: 126 IAKEHPQLYYSNGIPFINTIEESLDYGREYTTTNVTTFHPNDGVNFLVILTSEAM- -PVG 183
I +EH +L+ +G P INTI+E L+YG EY +V P D + FLV+++ M G Sbjct. 128 IVREHVKLWNDDGTPTINTIDEGLNYGSEYDIVSVENHRPYDDMMFLWISKSIMHGTAG 187
Query. 184 DKEDKSG---GSIVGGPSPFSYYLLPINSSGEVYKPN-GAGNANFGEYMAFLT TKEP 236
+ E + S+ G P P YY+ P G+V K G NAN + LT +++ Sbjct: 188 EAESRLNDINASLNGMPQPLCYYIHPFYKDGKVPKTFIGDNNANLSPIVNMLTNIFΞQKS 247
Query: 237 FLNKIVGMYVTSYTGIPFIVDHANKTVRYNAGGSYKIMLPTYASDPTGTMKTFAFFCVKE 296
+N IV MYVT Y G+ + +K ++ + + + A D G + T VK+ Sbjct. 248 AVNNIVNMYVTDYIGLKLDYKNGDKELKLDKDMFEQAGI ADDKHGNVDTIF VKK 301
Query: 297 ARTFVPKRIDLVGNVYNYFREAFPFNVKEΞKLFMYPYCLIEITDTKGHVMTLRPEYLTGG 356
+ ID G+ + F + +ESKL MYPYC+ E+TD KG+ M L+ EY+ Sbjct: 302 IPDYETLEID-TGDKWGGFTKD QESKLMMYPYCVTEVTDFKGNHMNLKTEYIDNN 355
Query: 357 KLSVYVKGSLGISNKVMIEPIDYDVSNSTI ITNLSDKMLIDNDPNDVGVKSDYASA 412
KL + V+GSLG+SNKV DY+ S +T D LI+N+PND+ + +DY SA Sbjct: 356 KLKIQVRGSLGVSNKVAYSIQDYNAGGSLSGGDRLTASLDTSLINNNPNDIAIINDYLSA 415
Query: 413 FMQGNKNSLIAQEQNIRNTFRHGMGNSAMSTGGAIFSALASNNPFVGLTNIMGAGQQVNN 472
++QGNKNSL Q+ +1 GM +S G ++ +PF +++ G N Sbjct: 416 YLQGNKNSLENQKSSILFNGIVGMLGGGVSAG ASAVGRSPFGLASSVTGMTSTAGN 471
Query. 473 YVSEKENGIiNLLAGKVADIENIPDNVTQLGSNLSFTTGN-FQNYYQLRFKQIKYEYATRL 531
V + + L K ADI NIP +T++G N +F GN ++ Y ++ KQ+K EY L Sbjct. 472 AVLD MQALQAKQADIANIPPQLTKMGGNTAFDYGNGYRGVYVIK-KQLKAEYRRSL 526
Query: 532 DRYFSMYGTKSNRVATPNLQTRKAWNFIKLKEPNIVGTMSNDVLTRVKQIFSAGVTLWHT 591
+F YG K NRV PNL+TRKA+N+I+ K+ I G ++N+ L ++ IF G+TLWHT Sbjct: 527 SSFFHKYGYKINRVKKPNLRTRKAYNYIQTKDCFISGDINNNDLQEIRTIFDNGITLWHT 586
Query: 592 NDVLNYNQDN 601
+D+ NY+ +N Sbjct: 587 DDIGNYSVEN 596
Query= sid | 110157 | lan| 182ORF002 Phage 182 0RF| 2152-3873 | 1 (573 letters)
>gι|118848|sp|P19894|DPOL_BPM2 DNA POLYMERASE >gι | 76896 | pir | | JQ0161 DNA-directed DNA polymerase (EC 2.7.7.7) - phage M2 >gι 1215509 (M33144) DNA polymerase [Bacteriophage M2] Length = 572
Score = 665 bits (1697), Expect = 0.0
Identities = 327/589 (55%), Positives = 420/589 (70%), Gaps = 38/589 (6%)
Query 3 KKYTGDFETTTDLNDCRVWSWGVCDIDNVDNMTFGLEIDΞFFEWCKMQGSTDIYFHNEKF 62 K ++ DFETTT L+DCRVW++G +1 N+DN G +D F +W M+ D+YFHN KF
Sbjct: 4 KMFΞCDFETTTKLDDCRVWAYGYMEIGNLDNYKIGNSLDEFMQWV-MEIQADLYFHNLKF 62
Query: 63 DGEFMLSWLFKNGFKWCKEAKEDRTFSTLISNMGQWYALEICWEVNYXXXXXXXXXXXXX 122
DG F+++WL ++GFKW E + T++T+IS MGQWY ++IC+ Sbjct: 63 DGAFIVNWLEQHGFKWSNEGLPN-TYNTIIΞKMGQWYMIDICFGYK GKRKL 112
Query: 123 XXIIYDSLKKYPFPVKQIAEAFNFPIKKGEIDYTKERPIGYKPTKDEWEYLKNDIQIMAM 182
+IYDSLKK PFPVK+IA+ F P+ KG+IDY ERP+G++ T +E+EY+KNDI+I+A Sbjct: 113 HTVIYDSLKKLPFPVKKIAKDFQLPLLKGDIDYHTERPVGHEITPEEYEYIKNDIEIIAR 172
Query: 183 ALKIQFDQGLTRMTRGSDALGDYKDWLKATHGKSTFKQWFPILSLGFDKDLRKAYKGGFT 242
AL IQF QGL RMT GSD+L +KD L F + FP LSL DK++RKAY+GGFT Sbjct: 173 ALDIQFKQGLDRMTAGSDSLKGFKDILST KKFNKVFPKLSLPMDKEIRKAYRGGFT 228
Query: 243 WVNKVFQGKEIGDGIVFDVNSLYPSQMYVRPLPYGTPLFYEGEYKPNNDYPLYIQNIKVR 302
W+N ++ KEIG+G+VFDVNSLYPSQMY RPLPYG P+ ++G+Y+ + YPLYIQ 1+ Sbjct: 229 WLNDKYKEKEIGEGMVFDVNSLYPSQMYSRPLPYGAPIVFQGKYEKDEQYPLYIQRIRFE 288
Query: 303 FRLKEGYIPTIQVKQSSLFIQNEYLESSVNKLGVDELIDLTLTNVDLELFFEHYDILEIH 362
F LKEGYIPTIQ+K++ F NEYL++S GV E ++L LTNVDLEL EHY++ + Sbjct: 289 FELKEGYIPTIQIKKNPFFKGNEYLKNS GV-EPVELYLTNVDLELIQEHYELYNVE 343
Query: 363 YTYGYMFKASCDMFKGWIDKWIEVKNTTEGARKANAKGMLNSLYGKFGTNPDITGKVPYM 422
Y G+ F+ +FK +IDKW VK EGA+K AK MLNSLYGKF +NPD+TGKVPY+ Sbjct: 344 YIDGFKFREKTGLFKDFIDKWTYVKTHEEGAKKQLAKLMLNSLYGKFASNPDVTGKVPYL 403
Query: 423 GEDGIVRLTLGEEELRDPVYVPLASFVTAWGRYTTITTAQKCFDRIIYCDTDSIHLVGTE 482
+DG + +G+EE +DPVY P+ F+TAW R+TTIT AQ C+DRIIYCDTDSIHL GTE Sbjct: 404 KDDGSLGFRVGDEEYKDPVYTPMGVFITAWARFTTITAAQACYDRIIYCDTDSIHLTGTE 463
Query: 483 VPEAIDHLVDPKKLGYWGHESTFQRAKFIRQKT YVEEIDGEL 524
VPE I +VDPKKLGYW HESTF+RAK++RQKT YV+E+DG+L Sbjct: 464 VPEIIKDIVDPKKLGYWAHESTFKRAKYLRQKTYIQDIYVKEVDGKLKECSPDEATTTKF 523
Query: 525 NVKCAGMPDRIKEIVTFDNFEVGFSΞYGKLLPKRTQGGWLVDTMFTIK 573
+VKCAGM D IK+ VTFDNF VGFSS GK P + GGWLVD++FTIK Sbjct: 524 SVKCAGMTDTIKKKVTFDNFAVGFSSMGKPKPVQVNGGWLVDSVFTIK 572
Query= sid| 110159 | lan| 182ORF004 Phage 182 ORF| 4626-5954 | 3 (442 letters)
>gi|l38117|sp|P13849|VG8_BPPH2 MAJOR HEAD PROTEIN (LATE PROTEIN GP8) >gi| 75845 jpir| |WMBP89 gene 8 protein - phage phi-29 >gi|215325 (M14782) major head protein [Bacteriophage phi-29] >gi I 225362 I rf | |1301270B gene 8 [Bacillus sp.] Length = 448
Score = 309 bits (783), Expect = 2e-83
Identities = 176/440 (40%) , Positives = 250/440 (56%) , Gaps = 27/440 (6%)
Query: 4 KITEQDVLRATNVETPVQLMTAIYNSSSSLFQANVPMPNADNIEAVGAGITRLDWKNEF 63
+IT DV + + ++ Al NS F++ VP+ A+N+ VGAGI V+N+F Sbjct: 2 RITFNDVKTSLGITESYDIVNAIRNSQGDNFKSYVPLATANNVAEVGAGILINQTVQNDF 61
Query: 64 ISTLVDRIGKWIRYKSWRNPLKMFKKGNMPLGRTIEEIFVDIAQEHKFNPDESVTGVFK 123
I++LVDRIG WIR S NPLK FKKG +PLGRTIEEI+ DI +E +++ +E+ VF+ Sbjct: 62 ITSLVDRIGLWIRQVSLNNPLKKFKKGQIPLGRTIEEIYTDITKEKQYDAEEAEQKVFE 121
Query: 124 QEVPDVKTLFHEINREGYYKQTIQEAWLEKAFTSWDNFNSFVAGVMNALYTGDEVSEFEY 183
+E+P+VKTLFHE NR+G+Y QTIQ+ L+ AF SW NF SFV+ ++NA+Y EV E+EY Sbjct: 122 REMPNVKTLFHERNRQGFYHQTIQDDSLKTAFVSWGNFESFVSSIINAIYNSAEVDEYEY 181
Query: 184 TKLLIANYQEKELFKEIEIGEITESNA--KEFIRKIKSTSNKLEFM--SSAYNAQGVKTS 239
KLL+ NY K LF ++I E T S EF++K+++T+ KL S +N+ V+T Sbjct: 182 MKLLVDNYYSKGLFTTVKIDEPTSSTGALTEFVKKMRATARKLTLPQGSRDWNSMAVRTR 241
Query: 240 TSKSDQYXXXXXXXXXXXXXXXXXXXFNMSKTDFVGHKIVIDEFPKKEGEESSNIVAVIV 299
+ D + FNM++TDF+G+ VID F S+ + AV+V Sbjct: 242 SYMEDLHLIIDADLEAELDVDVLAKAFNMNRTDFLGNVTVIDGF ASTGLEAVLV 295
Query: 300 DSEWFMIYDKLYKTTSLYNPEGLYWNYWLHHHQLYSTSQFGNAVAFVKSATKPVTKVAFA 359
D +WFM+YD L+K ++ NP GLYWNY+ H Q S S+F NAVAFV VT+V + Sbjct: 296 DIΦWF.1vYDNLHKMETVRNPRGLYϊnr-YYHWQTLSVSRFANAVAFVSGDVPAVTQVIVS 355
Query: 360 SATTSWKGSSKDIALTFTPVEATNQQGEWSSAPALVKATVKQTAGKATAVTVEGLEVG 419
+V +G + V ATN + V V G +T + G Sbjct: 356 PNIAAVKQGGQQQFT AYVRATNAKDHKV VWSVEGGSTGTAI TG 398
Query: 420 QΞLVTFTAIGGQQATVLVTV 439
L++ + Q TV TV Sbjct: 399 DGLLSVSGNEDNQLTVKATV 418
Query= sid| 110160 | lan| 182ORF005 Phage 182 ORF| 12651-13700 | 3 (349 letters)
>gi|l37932|sp|P15132|VG13_BPPH2 MORPHOGENESIS PROTEIN 1 (LATE
PROTEIN GP13) >gi | 75858 |pir | | MBP23 gene 13 protein - phage phi-29 >gi| 215331 (M14782) morphogenesis protein
[Bacteriophage phi-29] >gi | 225368 |prf ] | 1301270H gene 13
[Bacteriophage phi-29] Length = 365
Score = 51.5 bits (121), Expect = 8e-06
Identities = 44/166 (26%), Positives = 70/166 (41%), Gaps = 14/166 (8%)
Query: 6 NEQIARGQTIAKILSKYGYNKNSQVGWANLHWESA---GLNPNSNEXXXXXXXXX-QWT 61
+E Q I LS G+ K + G++ N+ ES GL N +E QWT
Sbjct: 12 SEMKVNAQYILNYLSSNGWTKQAICGMLGNMQSESTINPGLWQNLDEGNTSLGFGLVQWT 71
Query: 62 PKSNLYRQAQICGLSNAKAETLEGQAEIIAQGDKTGQWMDNTPVSSAGYTNPQTLSAFKQ 121
P SN A GL ++ II + + QW++ ++ Y K
Sbjct: 72 PASNYINWANSQGLPYKDMDS--ELKRIIWEVNNNAQWINLRDMTFKEY IKS 121
Query: 122 SANIDVATINFMCHWERPGKLHIEERLDLAQAYSKHIDGSGGGGVK 167
+ + F+ +ERP + ER D A+ + K++ G GGGG++ Sbjct: 122 TKTPRELAMIFLASYERPANPNQPERGDQAEYWYKNLSGGGGGGLQ 167
Query= sid| 110161 | lan| 182ORF006 Phage 182 ORF| 14995-16026 | 1 (343 letters)
>gi|l37945|sp|P0754l|VG16_BPPZA ENCAPSIDATION PROTEIN (LATE PROTEIN GP16) >gi|75861|pir| |WMBP16 gene 16 protein - phage PZA >gi I 216065 (M11813) morphogenesis protein C [Bacteriophage PZA] Length = 332
Score = 402 bits (1023), Expect = e-111
Identities = 186/332 (56%) , Positives = 244/332 (73%) , Gaps = 2/332 (0%)
Query: 11 EKNLYYNPNNALGFNCLMLFVIGARGIGKTYGYKKFWNRFIKHGEQFIYLRRFKTELKK 70
+K+L+YNP L ++ ++ FVIGARGIGK+Y K + +NRFIK+GEQFIY+RR+K EL K Sbjct: 2 DKSLFYNPQKMLSYDRILNFVIGARGIGKSYAMKVYPINRFIKYGEQFIYVRRYKPELAK 61
Query: 71 IPQFFKTMAKEFPDHKLEVKGKEFYCDDKLMGWAVPLSTWGIEKSNEYPEVRTILFDEFL 130
+ +F +A+EFPDH+L VKG+ FY D KL GWA+PLS W EKSN YP V TI+FDEF+ Sbjct: 62 VSNYFNDVAQEFPDHELWKGRRFYIDGKLAGWAIPLSVWQSEKSNAYPNVSTIVFDEFI 121
Query: 131 IEKSKITYLPNEAEALLNMMETVFRRRTNTRCVMLSNATSWNPYFLYFNLQPDLNKRFN 190
EK Y+PNE ALLN+M+TVFR R RC+ LSNA SWNPYFL+FNL PD+NKRFN Sbjct: 122 REKDNSNYIPNEVSALLNLMDTVFRNRERVRCICLSNAVSWNPYFLFFNLVPDVNKRFN 181
Query: 191 LYQDRGILIELCDSKDFAEVKRETPFGRLIRGTEYEDFSINNEFVNDSDTFIEKRSKNSS 250
+Y D LIE+ DS DF+ +R+T FGRLI GTEY + S++N+F+ DS FIEKRSK+S Sbjct: 182 VYDD--ALIEIPDSLDFSSERRKTRFGRLIDGTEYGEMSLDNQFIGDSHVFIEKRSKDSK 239
Query: 251 FLCΛIAFEGKIFGYWIDAETGCVYVSYDYQPNTNHFYAMTTKDHEENRLLMKNWRNNYYL 310
F+ +1 + G G W+D G +YV + P+T + Y +TT D EN +L+ N++NNY+L Sbjct: 240 FVFSIVYNGFTLGVW\7DVNQGI_MYVDTAHDPSTKNVYTLTTDDLNENMMLITNYKNNYHL 299
Query: 311 STVAKAFKNSYLRFDNIVIKNLHYDLFNKMKI 342
+A AF N YLRFDN VI+N+ Y+LF KM+I Sbjct: 300 RKLASAFMNGYLRFDNQVIRNIAYELFRKMRI 331
Query= sid| 110162 | lan| 182ORF007 Phage 182 ORF| 7795-877511 (326 letters)
>gi|1429239|emb|CAA67658| (X99260) upper collar protein [Bacteriophage B103]
Length = 308
Score = 271 bits (685) , Expect = 6e-72
Identities = 131/275 (47%), Positives = 187/275 (67%), Gaps = 5/275 (1%)
Query: 36 YYEHYRRQLTLLTFQLFEWENLPKSIDPRYLEIALHTNGYLGFFKDPTLGFMVCAGAEDG 95
+Y HY + L L +QLFEWE LP S+DP YLE ++H GY+GF+KDP +G++ C GA G Sbjct: 22 WYYHYYQYLCSLAYQLFEWERLPPSVDPΞYLEKΞIHQFGYVGFYKDPRIGYIACQGALSG 81
Query: 96 QIDHYHNPIFFTANEAMYHKRYPVLRYDDDDDKSKCIMLYNNDLKVPTLPSLHRFALDMA 155
+DHY+ P F A+ Y + + Y D +K+ + +YNNDLK TLP+L FA D+A Sbjct: 82 TVDHYNLPDRFHASSVGYQNTFKLYNYSDMKEKNMGVAIYNNDLKCSTLPALEMFAQDLA 141
Query: 156 DINQISRVNRRAQKTPVIIQTDEKKYFSLLQAYNQIDENNQAVFVDKDMEFDESFNVWQT 215
++ +1 VN+ AQKTPV+I ++ SL YNQ + N +FV + ++ D + V++T Sbjct: 142 ELKEIIAVNQNAQKTPVLIAANDNNQLSLKNIYNQYEGNAPVIFVHESLDLD-NLKVFKT 200
Query: 216 NAPYWDKLRSELNEVWNEVLTFLGINNANVDKTARVQTSEVLSNNEQIESSGNILLKSR 275
+APYWDKL ++ N VWNEV+T+LGI NAN++K R+ TSEV SN+EQIESSGNI LK+R Sbjct: 201 DAPYWDKLNAQKNAVWNEVMTYLGIKNANLEKKERMVTSEVDSNDEQIESSGNIYLKAR 260
Query: 276 KEFCDRVNRVFGDELDGKIDVKFRTDAVRQLQLAA 310
+E C++++ ++G L VKFR D V Q++L A Sbjct: 261 QEACNKISELYGLNL KVKFRYDIVEQMRLNA 291
Query= sid| 110163 | lan| 182ORF008 Phage 182 ORF] 14105-14983 | 2 (292 letters)
>gi|4210750|emb|CAA10710 I (AJ132604) LysL protein [Lactococcus lactis] Length = 235
Score = 139 bits (347) , Expect = 2e-32
Identities = 85/210 (40%), Positives = 114/210 (53%), Gaps = 14/210 (6%)
Query: 2 MNGIDISSYQTGIDLSKVPCDFVNIKATGGTGYVNPDCDRAFQQALSLGKKIGVYHFAHE 61
MNGIDIΞSYQ ++ VP DFV IKAT GT Y+NP + Q + K +G YHFA Sbjct: 1 MNGIDISSYQAELNAGIVPSDFVIIKATEGTNYINPTWEEQAGQVIQTNKLLGFYHFAS- 59
Query: 62 RGLEGTPQQEAQFFLDNIKGYIGKAVLILDFEGS--NQKDVNWAKAFLDYVYNKTGVKAW 119
G P EA FF+ +K YIGKAVL+LDFE N A+ FL+ V KTG+ Sbjct: 60 ---VGNPIAEADFFISWKNYIGKAVLVLDFEAGAINAWGNVGARQFLNRVKEKTGINPM 116
Query: 120 FYTYTANLNTTDFSSIAKGDYGLWVAEYGSNQPQGYSQPAPPKTNN FPIVACFQF 174
Y + ++S+I+ + LWVA+Y S P GY + P T+ + A Q+ Sbjct: 117 IYMSSDVTRQFNWSTISSTN-PLWVAQYASMNPTGYQ--SEPWTDGKGYGAWSSAAIHQY 173
Query: 175 TSKGRLPGYNGNLDLNVFYGDGNTWDLYVG 204
+S G L ++GNLD+N+ Y + N W G Sbjct: 174 SSAGSLSNWSGNLDINLAYINANQWKSLAG 203
Query= sid| 110164 | lan| 182ORF009 Phage 182 ORF] 8765-9601 | 2 (278 letters)
>gi| 1429240 |emb|CAA67659| (X99260) lower collar protein [Bacteriophage B103] Length = 293
Score = 180 bits (451) , Expect = le-44
Identities = 115/296 (38%), Positives = 161/296 (53%), Gaps = 33/296 (11%)
Query: 3 LKRYIESFTYYQPELSRKERIEVGRKQLFDFDYPFYDETKRAEFETKFINHFYLREIGSE 62
L YIE ++ Y+ LS E+IE GR +LFDF YP +DE+ R FET FI +FY+REIG E Sbjct: 8 LSTYIEMWSQYETGLSMAEKIEKGRPKLFDFQYPIFDESYRKVFETHFIRNFYMREIGFE 67
Query: 63 TMGSFKFNLDEYLNLNMPYWNKMFLSNLEEF-PIFDDMDYTIDEKQKLLNEIDTNIKANR 121
T G FKFNL+ +L +NMPY+NK+F S L ++ P+ + T K+ DT NR Sbjct: 68 TEGLFKFNLETWLIINMPYFNKLFESELIKYDPLENTRLNTTGNKKN DTERNDNR 122
Query: 122 D ESKNQTKQVDQTDNRNKNTRDTGTT DSFSRNTYTDTPQKDLRIASNG 169
D + K+ TK D+T+ + D TT D+F+R +D P L + +N
Sbjct: 123 DTTGSMKADGKSNTKTSDKTNATGSSKEDGKTTGSVTDDNFNRKIDSDQPDSRLNLTTN- 181
Query: 170 DGTGVINYATNITEDLSKETTSSTGVETNNDKTNQNTRSNAS EKETKNTD 219
DG G + YA+ I E+ + ++TG TNN ++ + S S T N
Sbjct: 182 DGQGTLEYASAIEENNTNNKRNTTG--TNNVTΞSAESESTGSGTSDTVTTDNANTTTNDK 239
Query: 220 INKDQNQTKDTITRYKGKKGNTDYADLLEKYRRSVLRIEKMIFREMNKEGLFLLVY 275
+N N +D I GK G YA L++ YR ++LRIEK IF EM + LF+LVY Sbjct: 240 LNSQINNVEDYIESKIGKSGTQSYASLVQDYRAALLRIEKRIFDEMQE--LFMLVY 293
Query= sid| 110165 | lan| 182ORF010 Phage 182 ORF| 1310-2155 | 2 (281 letters)
>gi|l35604|sp|P06812|TERM_BPNF DNA TERMINAL PROTEIN
>gi| 75815 jpir | ] ERBPNP terminal protein - phage NF >giJ579177|emb|CAA68440| (Y00363) gene E product (AA 1-267) [Bacteriophage NF] Length = 266
Score = 74.9 bits (181), Expect = 6e-13
Identities = 73/275 (26%), Positives = 129/275 (46%), Gaps = 37/275 (13%)
Query: 3 VRISKNDRAKLEKIYGKSNKARKKYNRLRQK-GVE---ERQLPTVPTSKKRLIDYVKSTN 58
+RI+ ND+A K+ K+ KA K +R ++K G++ E +LP + + + Sbjct: 7 IRITNNDKALYAKLV-KNTKA--KISRTKKKYGIDLSNEIELPPLESFQ 52
Query: 59 MSRSDFNKMLDELVDFAQPYNENYIFEINKRNVAISRAQIKEAQIKTEQAQKAKEEHYKE 118
+R +FNK + F N+NY F NK + S+A+I E T++AQ+ +E +E Sbjct: 53 -TREEFNKWKQKQESFTNRANQNYQFVKNKYGIVASKAKINEIAKNTKEAQRIVDEQREE 111
Query: 119 L- NKVEVKKPTENTIVTPTILTELGADLPFQAIPDFNIDAFTSPEGVQSYLEN 170
+ K + I++P+ +T G P DFN D S +++ E Sbjct: 112 IEDKPFISGGKQQGTVGQRMQILSPSQVT--GISRP SDFNFDDVRSYARLRTLEEG 165
Query: 171 IG-KQDEQYFDERDQLYYDNFRQAMFTIFNSD--ADDIVRLLDSMGLDLFMKTYVSNFLD 227
+ K Y+D R + NF + + FNSD +D++V L + D F + Y+ F + Sbjct: 166 MAEKASPDYYDRRMTQMHQNFIEIVEKSFNSDWLSDELVERLKKIPPDDFFELYLM-FDE 224
Query: 228 MNLDYIYDEAEVQQKKEQVYSKIAKVIESETGGEV 262
++ +Y E E + E + +KI ++ G+V Sbjct: 225 ISFEYFDSEGEDVEASEAMLNKIHSYLDRYERGDV 259
Query= sid| 110166 | lan| 182ORF011 Phage 182 ORF| 9607-10158 | 1 (183 letters)
>gi|l42924l|emb|CAA67660 I (X99260) pre-neck appendage protein [Bacteriophage B103] Length = 860
Score = 50.8 bits (119), Expect = 6e-06
Identities = 29/105 (27%) , Positives = 56/105 (52%) , Gaps = 6/105 (5%)
Query: 8 KRFDGLPAVFKERFSKYPHTEYRYELLLDEEVSALIAYLNEVGALVNDMSGYLNYFIEHF 67
+RF+ L + + + +Y T + + L E+++ +1 YLN++G L ND+ N +E Sbjct: 7 RRFEKLGEMMVQVYERYLPTAFDESMTLLEKMNKIIEYLNQIGRLTNDWEEWNKVMEWI 66
Query: 68 V-EKLEEITNDTLKKWLSDGTLENLINDTVFANYIKEIKRLQILV 111
+ + LE+ +TL+KW +G +L+ I E+K+ + V Sbjct: 67 LNDGLEDYVKETLEKWYEEGKFADLV IQVIDELKQFGVSV 106
Query-. sid| 110169 | lan| 182ORF014 Phage 182 ORF| 13716-14108 | 3 (130 letters)
>gi|l37936|sp|P11188|VG14_BPPH2 LYSIS PROTEIN (LATE PROTEIN GP14) >gi|75860 jpir I |WMBP29 gene 14 protein - phage phi-29 >gijl5678Jemb|CAA28631| (X04962) gene 14 product (AA
1-393) [Bacteriophage phi-29] >gι | 225369 |prf | | 1301270J gene 14 [Bacteriophage phi-29] Length = 131
Score = 96.7 bits (237), Expect = 6e-20
Identities = 53/131 (40%), Positives = 81/131 (61%), Gaps = 3/131 (2%)
Query. 1 MIEYITQWL-ADDNHLVYGLIIWLMVAMIIDFVLGFTIAKFNKEIDFSSFKAKAGIIVKV 59
MI ++ +L D+ L+Y L +LMV M++D VLG AK N I FSSFK K G+++KV Sbjct. 3 MIAWMQHFLETDETKLIYWLT-FLMVCMWDTVLGVLFAKLNPNIKFSSFKIKTGVLIKV 61
Query. 60 AEMVLWYFIPVAVKFGAVGITMYITMLVGLILSEIYSILGHISDIDDDNNWTDYVKKFL 119
+EM+L + IP AV F A G+ + T+ L +SEIYSI GH+ +DD +++ + ++ F Sbjct: 62 SEMILALLAIPFAVPFPA-GLPLLYTVYTALCVSEIYSIFGHLRLVDDKSDFLEILENFF 120
Query 120 DGTLNRKDDIK 130
T + + K Sbjct. 121 KRTSGKNKEEK 131
Query= sιd| 110170 | lan| 182ORF015 Phage 182 ORF| 854-1225 | 2 (123 letters)
>gι| 15670|emb|CAA24483 I (V01155) reading frame 10 (may be gene 4) [Bacteriophage phi-29] Length = 124
Score = 69.9 bits (168), Expect = 6e-12
Identities = 39/119 (32%) , Positives = 64/119 (53%) , Gaps = 3/119 (2%)
Query. 3 IVKSTFDTQTPEGMLQVFNATNGASIPLRNAI-GEVLELKDILVYSDEVSGFGGAEPSQA 61
IVK+TFDT+T EG +++FNA G +N G ++E I Y +G A+ + Sbjct: 6 IVKATFDTETLEGQIKIFNAQTGGGQSFKNLPDGTIIEANAIAQYKQVSDTYGDAK--EE 63
Query: 62 ELVAFFTEDGKTYAGVSAVATKSAKNLIDMMTANPDIKPKISFVEGKSNGGQKFVNLQV 120
+ F DG Y+ +S ++A +LID++T + K+ V+G S+ G F +LQ+ Sbjct: 64 TVTTIFAADGSLYSAISKTVAEAASDLIDLVTRHKLETFKVKWQGTSSKGNVFFSLQL 122
Query= sιd| 110174 | lan| 182ORF019 Phage 182 0RF| 4323-4613 | 3 (96 letters)
>gι| 1429235 | emb | CAA67654 | (X99260) head morphogenesis protein [Bacteriophage B103] Length = 101
Score = 60.9 bits (145), Expect = le-09
Identities = 34/96 (35%), Positives = 53/96 (54%), Gaps = 5/96 (5%)
Query. 1 MEIKEHESILNGILESVTDGEARSKIVEHLEALREDYGATTEALTSANSTLEKLKKDNEA 60
ME HE ILN + + + R+++ L+ LR DYG+ + S EKL+ +N Sbjct: 3 MERDSHEEILNKLNDPELEHSERTEL---LQQLRADYGSVLSEFSELTSATEKLRAENSD 59
Query: 61 LVISNSKLFRERAIVEPAEN--NEPETDQNITLDDL 94
L++SNSKLFR+ I + E + E + IT++DL Sbjct: 60 LIVSNSKLFRQVGITKEKEEEIKQEELSETITIEDL 95
Query= sιd| 110180 | lan| 182ORF025 Phage 182 ORF | 548-814 | 2 (88 letters)
>gι|138099|sp|P06955|VG6_BPPZA EARLY PROTEIN GP6
>gι j 758411 pir | |ERBP6Z gene 6 protem - phage PZA >gι 1216047 (M11813) gene 6 product [Bacteriophage PZA] >gι|224746|prf I |1112171K ORF 6 [Bacteriophage PZA] Length = 96
Score = 55.0 bits (130), Expect = 8e-08 Identities = 28/79 (35%) , Positives = 45/79 (56%)
.40
Query: 4 KLMQRNVTSTKVEFSEVIVQDGAPTIVPCEPWLTGKLSEEKALSAIKRKNPDKNVWTN 63
K+MQR +T T V +++++ DG + G LS E+A +KRK + V V +
Sbjct: 3 KMMQREITKTTVNVAKMVMVDGEVQVEQLPΞETFVGNLSMEQAQWRMKRKYKGEPVQWS 62
Query: 64 VSHETALYTMPVDKFIELA 82
V T +Y +PV+KF+E+A Sbjct: 63 VEPNTEVYELPVEKFLEVA 81
Table 26 Secondary structure prediction for ORF 182ORF008
1 MMNGIDISSY QTGIDLSKVP CDFVNI ATG GTGYVNPDCD RAFQQALSLG KKIGVYHFAH
CCCCCCCCCC CCCCCCCCCC CCEEEEEECC CCCCCCCCCC HHHHHHHHHC CCCCEEEEEE 61 ERGLEGTPQQ EAQFF DNIK GYIGKAVLIL DFEGSNQKDV N AKAFLDYV YNKTGVKAWF
CCCCCCCCHH HHHHHHHHHC CCCCEEEEEE CCCCCCCHHH HHHHHHHHHH HCCCCCEEEE 121 YTYTANLNTT DFSSIAKGDY GLWVAEYGSN QPQGYSQPAP PKTNNFPIVA CFQFTSKGRL
EEECCCCCCC CCCEECCCCC CEEEEECCCC CCCCCCCCCC CCCCCCCEEE EEEECCCCCC 181 PGYNGNLDLN VFYGDGNTWD LYVGKKQDQI VPPENKIFDA TSDEFIFTLT TGSTSVFYFD
CCCCCCCCEE EEECCCCCCE EEECCCCCCC CCCCCCCCCC CCCEEEEEEC CCCCEEEECC 241 GETIFELSDP TQLDHIRGTY NHVHGKEIPS MVWTPEQFDI YLKMYEKKPV YK
CCEEEECCCC CCHHHHCCEE CCCCCCEECC CCCCCCCHHH HHHHHCCCCE EC
Secondary structure prediction for ORF 182ORF014
1 IEYITQWLA DDNHLVYGLI IWLMVAMIID FVLGFTIAKF NKEIDFSSFK AKAGIIVKVA
CCCCEECCCC CCCCHHHHHH HHHHHHHHHH HHHHHHHHHC CCCCCHHHHH HHHCEEEEEE
61 EMVLWYFIP VAVKFGAVGI TMYITMLVGL ILSEIYSILG HISDIDDDNN WTDYVKKFLD
EEEEEEEECC CEEECCCEEE EEEEEEEEEE EEEEEEEECC CCCCCCCCCC CEEEEEEECC
121 GTLNRKDDIK
CCCCCCCEEC
Table 27 Enterococcus accession numbers 242/242 gi|2895751 |gb| AF044978.11 AF044978 [2895751 ] gij4098267|gb|U76614.1|BLU76614 [4098267] gi|4803755|dbj|AB026843.1|AB026843 [4803755] gi|47019|emb| Y00116.1 |SFAMB 1 [47019] gi|4769001 |gb| AF 140549.1 |AF 140549 [4769001 ] g_|4158179|emb|AL035206.1|SC9B5 [4158179] gi|4760901 |gb|AF099088.1 |AF099088 [4760901 ] gi|4165458|emb|X79343.1 JEF 16SSPA [4165458] gi|4704705 |gb| AF 121254.11 AF 121254 [4704705] gi|4165457|emb|X79342.1 IEFTR ALA [4165457] gi|3342117|gb|AF076604.1 |AF076604 [3342117] gi|4165456|emb|X79341.1IEF23SRNA [4165456] gi|4688824|emb|AJ132470.1|ESP132470 gi|4150978|emb|Y14027.1jEFY14027 [4150978] [4688824] gi|4127803|emb|AJ223161.1|EFAJ3161 [4127803] gi|4732085|gb|AF125553.1|AF125553 [4732085] gi|2956685|emb|Y16413.1|EFENTIJO [2956685] gi|4732082|gb| AF 125552.1 |AF 125552 [4732082] gi|2665346|emb|Y13922.1|EHY13922 [2665346] gi|4732079|gb|AF125551.1|AF125551 [4732079] gi|4324675|gb|AF109375.1 |AF109375 [4324675] gi|4732076|gb|AF125550.1|AF125550 [4732076] gi|4234627|gb| AF061013.11 AF061013 [4234627] gi|4732073|gb| AF 125548.11 AF 125548 [4732073] gi|4234626|gb|AF061012.11 AF061012 [4234626] gi|4732070|gb| AF 125547.11 AF 125547 [4732070] gi|4234625|gb| AF061011.11 AF061011 [4234625] gi|4732067|gb|AF125546.1|AF125546 [4732067] gi|4234624|gb|AF061010.1|AF061010 [4234624] gi|4732064|gb|AF 125545.1 [AF 125545 [4732064] gi|4234623|gb|AF061009.1|AF061009 [4234623] gi|4732061 |gb| AF 125544.1 j AF 125544 [4732061] gi|4234622|gb|AF061008.1|AF061008 [4234622] gi|4704653|gb|AF114715.1|AF114715 [4704653] gi|4234621|gb|AF061007.1|AF061007 [4234621] gi|4704564|gb|AFl 02550.1 |AF 102550 [4704564] gi|4234620|gb|AF061006.11 AF061006 [4234620] gi|4688827|emb[AJ238249.1|EFA238249 [4688827] gi|4234619|gb| AF061005.11 AF061005 [4234619] gi|4680606|gb|AF125198.1|AF125198 [4680606] gi|4234618|gb|AF061004.1 |AF061004 [4234618] gi|4633279|gb|AF117609.1|AFl 17609 [4633279] gi|4234617|gb| AF061003.11 AF061003 [4234617] gi|4633124|gb|AF110130.1|AFl 10130 [4633124] gi|4234616|gb[AF061002.11 AF061002 [4234616] gi|4590399|gb|AF124258.1|AF124258 [4590399] gi|4234615|gb|AF061001.1|AF061001 [4234615] gi|4590336|gb|AF108380.1|AF108380 [4590336] gi|4234614 |gb| AF061000.11 AF061000 [4234614] gi|4590335|gb|AF108379.1[AF108379 [4590335] gi|3138990|gb|AF060241.11 AF060241 [3138990] gi|4019167|gb|U21300.1|CXU21300 [4019167] gi|3138986|gb|AF060240.1|AF060240 [3138986] gi|4545122|gb| AF077816.11 AF077816 [4545122] gi|4204535|gb|AF094803.1|AF094803 [4204535] gi|4433610|gb|AF106614.1|AF106614 [4433610] gi|4204534|gb|AF094802.1|AF094802 [4204534] gi|4468838|emb|AJ132039.1|EFA132039 gi|4204533 |gb| AF094801.11 AF094801 [4204533] [4468838] gi|4204532|gb|AF094800.1 |AF094800 [4204532] gi|4468121|emb[AJ132958.1|BPH132958 gi|4204531 |gb|AF094799.1 |AF0_94799 [4204534} [4468121] gi|4204530|gb| AF094798.11 AF094798 [4204530] g_|4456104|emb|Y17302.1|EHI17302 [4456104] gi|4204529|gb|AF094797.11 AF094797 [4204529] gi|4433611|gb|AF106615.1|AF106615 [4433611] gi|4204528|gb|AF094796.1|AF094796 [4204528] gi|4433607|gb|AF106611.1|AF106611 [4433607] gi|4204527|gb| AF094795.11 AF094795 [4204527]
_>4J gi|4204526|gb|AF094794.1 |AF094794 [4204526] gi|2149899|gb|U94707.1 |EFU94707 [2149899] gi|4204525|gb|AF094793.1|AF094793 [4204525] gi|2149149|gb|U82366.1|LSU82366 [2149149] gi|4204524|gb|AF094792.1 |AF094792 [4204524] gi| 1469463 |gb|U49512.1|EFU49512 [1469463] gi|4204523 |gb| AF094791.1 ] AF094791 [4204523] gi|1244503|gb|U35366.1|EFU35366 [1244503] gi|4204522|gb|AF094790.1 |AF094790 [4204522] gi|833854|gb|U26268.1|EFU26268 [833854] gi|4204521 |gb| AF094789.1 IAF094789 [4204521 ] gi|841200|gb|U18931.1|CPU18931 [841200] gi|4204520|gb|AF094788.1|AF094788 [4204520] gi|460079|gb|U00457.1 |U00457 [460079] gi|4204519|gb| AF094787.11 AF094787 [4204519] gi|460077|gb|U00456.1 |U00456 [460077] gi|4204518|gb|AF094786.1 |AF094786 [4204518] gi|535661|gb|L34675.1|INSTRANSPO [535661] gi|4204517|gb| AF094785.11 AF094785 [4204517] gi|3023041 |gb|AF007787.1 |AF007787 [3023041 ] gi|4204516|gb| AF094784.1 j AF094784 [4204516] gi|431124|gb|L15633.1|TRN916ENT [431124] gi|4204515 |gb| AF094783.11 AF094783 [4204515] gi|388106|gb|L23802.1|ENEEBSA [388106] gi|4204514|gb|AF094782.1|AF094782 [4204514] gi|3608387|gb|AF071085.1 |AF071085 [3608387] gi|4204513 |gb| AF094781.11 AF094781 [4204513] gi|3551851|gb|AF076027.1|AF076027 [3551851] gi|4204512|gb|AF094780.1 |AF094780 [4204512] gi|3551773|gb|U94770.1 |SPU94770 [3551773] gi|3873186|gb|AF034779.11 AF034779 [3873186] gi|3551743 |gb|U57498.1 |ECU57498 [3551743] gi|4151367|gb|AF093508.1|AF093508 [4151367] gi|3243178|gb|AF063010.1 |AF063010 [3243178] gi|2828136|gb| AF039903.1 |AF039903 [2828136] gi|3136316|gb| AF063900.11 AF063900 [3136316] gi|2828135|gb|AF039902.1|AF039902 [2828135] gi|3540256[gb|AF052459.1|AF052459 [3540256] gi|2828134jgb| AF039901.11 AF039901 [2828134] g.|755215|gb|U17696.1|LLU17696 [755215] gi|2828133|gb|AF039900.1|AF039900 [2828133] gi|3421437|gb|AF082295.11 AF082295 [3421437] gi|2828132|gb|AF039899.1|AF039899 [2828132] gi|3421436|gb| AF082294.11 AF082294 [3421436] gi|2828131 |gb|AF039898.1 |AF039898 [2828131] gi|3421435 |gb|AF082293.1 |AF082293 [3421435] gi|4103866|gb|AF028812.11 AF028812 [4103866] gi|3421434|gb|AF082292.1 |AF082292 [3421434] gi|4103864|gb|AF028811.1 |AF028811 [4103864] gi|3341430|emb|Y17797.1|EFY17797 [3341430] gi|2605925|gb|AF029727.1 |AF029727 [2605925] gi|3319647|emb|X69092.1 |EHPBP3RA [3319647] gi|1402750|gb|U60038.1|EFU60038 [1402750] gi|3292886|emb|AJ007584.1|EFA7584 [3292886] gi|1835780|gb|U86375.1|EFU86375 [1835780] gi|3261536|emb|AL021958.1|MTV041 [3261536] gi|3831555|gb|AF047608.1|AF047608 [3831555] gi|3250708|emb|Z95150.1 |MTCY164 [3250708] gi|3790617[gb|AF097414.1 |AF097414 [3790617] gi|3249688|gb|AF070678.1 |AF070678 [3249688] gi|3767587|dbj|AB005036.1|AB005036 [3767587] gi|3249687|gb|AF070677.1 |AF070677 [3249687] gi|3757810|gb|AF042288.1|AF042288 [3757810] gi|3249686|gb| AF070676.11 AF070676 [3249686] gi|3747039|gb|AF093509.1|AF093509 [3747039] gi|3219158|dbj|AB015233.1|AB015233 [3219158] gi[3660559jdbj|AB017811.1JAB017811 [3660559] gi|2765275|emb|Y12924.1|SPY12924 [2765275] gi|l 147743|gb|U42211.1|EHU42211 [1147743] gi|3183687|emb|Yl 1621.1|EA16SRRN [3183687] gi|3676412|gb|AF051917.1|AF051917 [3676412] gi|2765274|e_nb|Y12923.1|EFYl_2923 [2765 74 - gi|3676164|emb|AJ011113.1|EFA011113 gi|2765273|emb|Y12922.1|ESY12922 [2765273] [3676164] gi|2765272|emb|Y12921.1|ESY12921 [2765272] gi|2612869|gb|AF005726.1|AF005726 [2612869] gi|2765271 |emb| Y 12920.1 |EDY12920 [2765271 ] gi|2353762|gb|AF016233.1|AF016233 [2353762] gi|2765270|emb|Y12919.1|ESY12919 [2765270]
gi|2765269|emb| Yl 2918.1 |ECY12918 [2765269] gi|2058762|gb|B07882.1 IB07882 [2058762] gi|2765268|emb|Y12917.1|ECY12917 [2765268] gi|2058761|gb|B07881.1 IB07881 [2058761] gi|2765267|emb|Y12916.1|EPY12916 [2765267] gi|2058760[gb|B07880.1 IB07880 [2058760] gi|2765266|emb|Y12915.1]ESY12915 [2765266] gi|2058759|gb|B07879.1 IB07879 [2058759] gi|2765265|emb|Y12914.1|ERY12914 [2765265] gi|2058758|gb|B07878.1 IB07878 [2058758] gii2765264|emb|Y12913.1|EMY12913 [2765264] gi|2058757|gb|B07877.1 IB07877 [2058757] gi|2765263|emb|Y12912.1|EHY12912 [2765263] gi|2058756|gb|B07876.1 IB07876 [2058756] gi|2765262|emb|Y12911.1|E Y12911 [2765262] gi|2058755|gb|B07875.1 IB07875 [2058755] gi|2765261|emb|Y12910.1|EGY12910 [2765261] gi|2058754|gb|B07874.1 IB07874 [2058754] gi|2765260|emb|Y12909.1|EDY12909 [2765260] gi|2058753|gb|B07863.1 IB07863 [2058753] gi|2765259|emb|Y12908.1|ECY12908 [2765259] gi|2058752|gb|B07862.1 IB07862 [2058752] gi|2765258|emb|Y12907.1|EAY12907 [2765258] gi|2058751|gb|B07861.1 IB07861 [2058751] gi|2765257!emb|Y12906.1|EFY12906 [2765257] gi|2058750|gb|B07860.1 IB07860 [2058750] gi|2765256|emb|Y12905.1|EFY12905 [2765256] gi|2058749|gb|B07859.1 IB07859 [2058749] gii2894541|emb|AJ223332.1|EFAJ3332 [2894541] gi|2058748|gb|B07858.1 |B07858 [2058748] gi|2894539|emb|AJ223331.1|EFAJ3331 [2894539] gi|2058747|gb|B07857.1 IB07857 [2058747] gi|3108058|gb|AF060881.1|AF060881 [3108058] gi|2058746|gb|B07856.1 IB07856 [2058746] gi|3087776|emb|AJ223633.1|EFAJ3633 [3087776] gi|2058745|gb|B07855.1 IB07855 [2058745] gi|3080754|gb|AF016483.1|AF016483 [3080754] gi|2058744|gb|B07854.1 IB07854 [2058744] gi|2197119|gb| AF003921.11 AF003921 [2197119] gi|2058743|gb|B07853.1 IB07853 [2058743] gi|2982722|dbj|AB012213.1|AB012213 [2982722] gi|2058742|gb|B07852.1 |B07852 [2058742] gi|2982721|dbj|AB012212.1|AB012212 [2982721] gi|2058741|gb|B07851.1 IB07851 [2058741] gi|2058780|gb|B07890.1 IB07890 [2058780] gi|2058740|gb|B07850.1 IB07850 [2058740] gi|2058779|gb|B07889.1 |B07889 [2058779] gi|2947527|gb|T25933.1 |T25933 [2947527] gi|2058778|gb|B07888.1 IB07888 [2058778] gi|2924302|emb|X81655.1|EHERMAM [2924302] gi|2058777|gb|B07887.1 IB07887 [2058777] gi|2664256|emb|Y12234.1|EFAS48C [2664256] gi|2058776|gb|B07886.1 IB07886 [2058776] gi|2879906|dbj|D85752.1|D85752 [2879906] gi|2058775|gb|B07885.1 IB07885 [2058775] gi|2746216|gb|AF028836.11 AF028836 [2746216] gi|2058774|gb|B07884.1 IB07884 [2058774] gi|2745825|gb|AF039139.1 |AF039139 [2745825] gi|2058773|gb|B07873.1 IB07873 [2058773] gi|2696019|dbj|AB007844.1|AB007844 [2696019] gi|2058772|gb|B07872.1 IB07872 [2058772] gi|48999|emb|X62280.1 |EHPBP5G [48999] gi|2058771|gb|B07871.1 IB07871 [2058771] gi|2654477|gb|U89914.1|BFU89914 [2654477] gi|2058770|gb|B07870.1 IB07870 [2058770] gi|43347|emb|X68646.1|EHPSRAA [43347] gi|2058769|gb|B07869.1 IB07869 [2058769] gi|2613034|gb|AH005624.1|SEG_EDDH4RR [2613034] gi|2058768|gb|B07868.1 IB07868 [2058768] gi|2613033 |gb| AF029775.1 |EDDH4RR2 [2613Q33] gi|2058767|gb|B07867.1 IB07867 [2058767] gi|2613032|gb|AF029774.1|EDDH4RRl [2613032] gi|2058766|gb|B07866.1 IB07866 [2058766] gi|2058765|gb|B07865.1 IB07865 [2058765] gi|2613031|gb|AH005623.1|SEG_EDDHIRR [2613031] gi|2058764|gb|B07864.1 IB07864 [2058764] gi]2613030|gb| AF029773.1 |EDDHIRR2 [2613030] gi|2058763|gb|B07883.1 IB07883 [2058763]
gi|2613029|gb|AF029772.1|EDDHIRRl [2613029] gi|2231992|gb|U94530.1 |EFU94530 [2231992] gi|2613028|gb|AH005622.1|SEG_EDH19RR gi|2231990|gb|U94529.1 |EFU94529 [2231990] [2613028] gi|2231988|gb|U94528.1 |EFU94528 [2231988] gi|2613027|gb|AF029771.1|EDH19RR2 [2613027] gi|2231986|gb|U94527.1 JEFU94527 [2231986] gi|2613026|gb|AF029770.1|EDH19RRl [2613026] gi|2231984|gb|U94526.1 [EFU94526 [2231984] gi|2613025 |gb| AH005621.1 |SEG_EDISRR gi|2231982|gb|U94525.1|ECU94525 [2231982] [2613025] gi|2231980|gb|U94524.1 |ECU94524 [2231980] gi|2613024|gb|AF029769.1 |EDISRR2 [2613024] gi|2613023|gb|AF029768.1|EDISRRl [2613023] gi|2231978|gb|U94523.1|ECU94523 [2231978] gi|2231976|gb|U94522.1 |ECU94522 [2231976] gi|1881226|dbj|AB001488.1|AB001488 [1881226] gi|2231974|gb|U94521.1 ]ECU94521 [2231974] gi|2547160|gb| AF023104.1 |AF023104 [2547160] gi|2196685|gb|U25090.1 |EFU25090 [2196685] gi|2547159|gb| AF023103.11 AF023103 [2547159] gi|2547158|gb|AF023102.1|AF023102 [2547158] gi|2197120|gb|AF003922.1|AF003922 [2197120] gi|2547157|gb|AF023101.1|AF023101 [2547157] gi|2196683|gb|U25095.1 |EFU25095 [2196683] gi|2415383|gb|AF015775.1|AF015775 [2415383] gi|2196681 |gb|U25094.1 |EFU25094 [2196681] gi|2388636|gb|U94356.1|EFU94356 [2388636] gi|2196679|gb|U25093.1 |EFU25093 [2196679] gi|2388634|gb|U94355.1|ECU94355 [2388634] gi|2196677|gb|U25092.1|EFU25092 [2196677] gi|2196675|gb|U25091.1 |EFU25091 [2196675] gi|2340825|dbj|D26045.1|D26045 [2340825] gi|2226147|emb[Y14080.1|BSY14080 [2226147] gi|2196673|gb|U24682.1 |EFU24682 [2196673] gi|2327026|gb|U87997.1 |EFU87997 [2327026] gi|532533|gb|U09422.1|EFU09422 [532533] gi|2318058|gb|AF012532.1 |AF012532 [2318058] gi|48727 l|dbj|Dl 7462.1|ENENTP [487271] gi|468459|dbj|D28859.1 |ENEPPD 1 [468459] gi|1848175|emb|X87189.1|EM23S5SSP [1848175] gi|1848174|emb|X87187.1|EM16S23SS [1848174] gi|440135|dbj|D16334.1|ENEATPK [440135] gi|1848173|emb|X87188.1|EM16S23SP [1848173] gi|391680|dbj|D 13816.1|ENENAABS [391680] gi|1402524|dbj|D78257.1ID78257 [1402524] gij 1848172|emb|X87185.1 |EH23S5SSP [ 1848172] gi|1848171|emb|X87184.1|EH16S23SS [1848171] gi|709995|dbj|D30808.1|BACYCB20 [709995] gi|1848170|emb|X87181.1|EF23S5SSP [1848170] gi|2109265|gb|U91527.1|EFU91527 [2109265] gi|1848169|emb|X87183.1|EF23S5SPA [1848169] gi|1041112|dbj|D78016.1|ENEPPDlA [1041112] gi|1848168|emb|X87191.1|EF23S5SAC [1848168] gi|1339880|dbj|D85392.1|ENERPA [1339880] gi|1848167|emb|X87180.1|EF16S23SS [1848167] gi|1339878|dbj|D85393.1|ENEGElE [1339878] gi| 1848166|emb|X87182.1 |EF16S23 SP [ 1848166] gi|662918|emb|Z46807.1|EHCOPAYZ [662918] gi|769796|emb|X86176.1 |EFRPODDNE [769796] gi|1848165|emb|X87190.1|EF16S23SC [1848165] gi|1848164|emb|X87186.1|EF16S23SA [1848164] gi|1854638|gb[U51479.1|EGU51479 [1854638] gij 1857221 |gb|U72706.1 |EFU72706 [ 1857221 ] gi| 1848156|emb|X87179.1 |ED23S5SSP [ 1848156] gi| 1857219|gb|U72704.1|EFU72704 [ 1857219] gi|1848155|emb|X87178.1|ED16S23SS [1848155] gi| 1848154|emb|X87177.1 |ED 16S23SA [ 1848154] gij 1857217|gb|U72705.1 |ECU72705 [ 1857217] gi|2274942|emb|AJ000346.1|EHNAPBC [2274942] gi|1272655|emb|X96978.1 |EFPPDlGNS I12-726'55] gij 1272652|emb|X96976.1 |EFPLSEP1 G [ 1272652] gi|2274939|emb| AJ000042.1 |EFGLS24B [2274939] gi| 1279406|emb|X96977.1 |EFPAD 1 ORF [1279406] gi|414575|gb|L12710.1|ENEAAC [414575] gi|2245603|gb|AF006008.1|AF006008 [2245603] gi|1070149|emb|X93211.1|EFTNFOl [1070149]
gi|1065723|emb|X92947.1|EFTETMGN [1065723] gij 1469341 |gb|U30931.1 |ESU30931 [1469341] gi|1019639|gb|L38972.1|PH4COINJN [1019639] gi|488331 |gb|M77276.1 |S YNGIP2122 [488331 ] gi|l 151151|gb|U43087.1|EFU43087 [1151151] gi|1046177|gb|U39733.1| [1046177] gi|1098507|gb|U17283.1|BMU17283 [1098507] gi| 1236613|gb|U49939.1 [CVU49939 [1236613] gij 1498072|gb|U64887.1 |EFU64887 [1498072] gi|47491 |emb|X55766.1 |SS 16SR5G [47491 ] gi| 1498071 |gb|U64886.1 [EFU64886 [1498071] gi|47490|emb|X55767.1 |SS 16SR3G [47490] gi|1469783|gb|U58049.1|EHU58049 [1469783] gi|47061|emb|X56353.1|SFTET916 [47061] gij 1763666|gb|U81452.1 |EFU81452 [1763666] gi|49022|emb]X62755.1|SFNPRG [49022] gi|624694jgb|L38973.1 |PH4SEQ [624694] gi|47047|emb|X17214.1jSFPASAl [47047] gi|1730458|embjZ83305.1|EFVANRES [1730458] gi|47044|emb|X68847.1 ISFNOXAA [47044] gi| 1419498|emb|X84796.1 |ECPFW4 [ 1419498] gi|47033|emb|V01547.1|SFKANR [47033] gi| 1419497|emb|X84795.1 |ECPFW3 [1419497] gi|47018|embjX02027.1|SF5SRNA [47018] gij 1419496|emb|X84794.1 |ECPFW1 [ 1419496] gi|511044jemb|X75752.1 |MP 16SRNA0 [511044] gi|254400|gb|S43266.1 |S43266 [254400] gi|511043|emb|X75751.1|MP16SR243 [511043] gi|239025|gb|S66277.1 |S66277 [239025] gi|886481 |emb|X82819.1 |ESPLPAM [886481 ] gi| 1054931 |gb|U38590.1 [EFU38590 [1054931] gij517387|emb|X76177.1|ES16SRR [517387] gi| 1244573 |gb|U39788.1 [EH 39788 [1244573] gi|472916|emb|X76913.1 |EHNTPOP [472916] gi|1244571|gb|U39789.1|EGU39789 [1244571] gi|43351|emb|X55133.1|ES16SRRN [43351] gi| 1244569|gbjU39790.1 |EFU39790 [ 1244569] gij 1143442jemb|X92687.1 |EFPBP5G [ 1143442] gi|1255020|gb|U39777.1|ESU39777 [1255020] gi|963032|embjZ50854.1|EHARPQTOU [963032] gijl255018|gb|U39775.1|EPU39775 [1255018] gij886479|emb|X84818.1 |EHDNAPSR [886479] gi| 1255016[gb|U39778.1 |EDU39778 [1255016] gi|551437|emb|X81654.1|EHIS1216 [551437] gi|1255014jgb|U39776.1|ECU39776 [1255014] gij467805|emb|X78425.1|EFPBP5 [467805] gi|1255012|gb|U39774.1jEAU39774 [1255012] gi|296721 |emb|X55961.1 |EFPD78 [296721 ] gi|1619922|gb|U69267.1|IVU69267 [1619922] gi|287946|emb|Z19137.1|EFPTSHGN [287946] gi|790436|emb|X84861.1 |EFEFMPBP5 [790436] gi|49042|emb|X63285.1|EHNAKA [49042] gi|790434|emb|X84858.1jEFD63RPSR [790434] gi|49019|emb|X62658.1|EFSEAl [49019] gi|790432|emb|X84862.1|EF721PBP5 [790432] gi|43337|emb|Z12296.1|EFSPREG [43337] gi|790430|emb|X84860.1 |EF63RPBP5 [790430] gi|43335|emb|X56895.1|EFPVANAG [43335] gi|790428|emb|X84859.1|EF366PBP5 [790428] gi|43333 |emb|X 16421.1 |EFPF54 [43333] gij 1572800|gb|U70854.1 |CELF38A5 [ 1572800] gi|43331 |emb|X62657.1 |EFORF3 [43331 ] gi|1041816|gb|U17153.1|EFU17153 [1041816] gij 1065721 |emb|X92945.1|EFCAT501 [1065721] gi| 1086523|gb|U39859.1 |EFU39859 [ 1086523] gi|806551 |emb|Z49243.1 |EF411 OSOD [806551] gi|403564|gb|U01917.1 |EFUO 1917 [403564] gi|806549|emb|Z49244.1 |EF4105SOD [806549] gi| 1515474|gb|U66286.1 |EFU66286 [1515474] gi|505530|emb|X79542.1 |EFAS48 [505530] gi|1513068|gb|U15554.1|LMU15554 [1513068] gi|43323|emb|X62656.1|EFASPl-[43323_L - gi| 1296520|emb|X94181.1 |EFENTAORF gi|40840|emb|X56422.1|EC16SRNAG [40840] [1296520] gi|48189|emb|X04388.1 |TN1545TR [48189] gi|1488069|gb|U63997.1|EFU63997 [1488069] gi|928814|gb|L40841.1 |ENETRANSPO [928814] gi|1209525|gb|U35369.1|EFU35369 [1209525] gi|141856|gb|L01794.1|ADlREPABC [141856]
gijl49125|gb|M90647.1|IP8VANY [149125] gi| 153852|gb|AH000939.1 |SEG_STRTN916 [153852] gi|141862|gb|M87836.1|ADlTRAEl [141862] gi|153851|gb|M22645.1|STRTN9162 [153851] gi|141860|gb|M84374.1|ADlTRAA [141860] gijl53850|gb|M20864.1|STRTN9161 [153850] gi|141853|gb|M62888.1jADlPADl [141853] gi|153660[gb|M36878.1|STRIF2BA [153660] gi|1101637|dbj|D31674.1|EVM16RNA7 [1101637] gi|153585|gb|M13771.1|STRBRP [153585] gi|1101636|dbj|D31675.1jENE16RNA8 [1101636] gij 153575|gb|M64265.1 |STRATPEFHA [153575] gi|497792|dbj|D31676.1 |ENC 16RNA9 [497792] gi|153565|gb|M90060.1|STRATPASEA [153565] gi|1022729|gb|U36195.1|EFU36195 [1022729] gi| 152969|gb|M92376.1 |STABLAIA [ 152969] gi|488338|gb|M77279.1|SYNGIP3124 [488338] gi|309660|gbjL14285.1|PCFPRGWZY [309660] gi|488335|gb|M77278.1 |SYNGIP2563 [488335] gi|433714|gb|L12033.1|ENESATA [433714] gi|488333|gb|M77277.1|SYNGIP2124 [488333] gi|488329|gb|M77275.1|SYNGIP2121 [488329] gi|290645|gb|L15304.1|ENEVANB2A [290645] gi|388267|gb|L19532.1|ADlTRAC [388267] gi|148331|gb|M84146.1|ENEVANR [148331] gi|148329|gb|M64304.1|ENEVANH [148329] gi|493016|gb|U03756.1 |EFU03756 [493016] gi|148326|gb|M68910.1|ENEVANCRES [148326] gi|453536|gb|L28754.1|INSTRAN [453536] gi|148324|gb|M75132.1|ENEVANC [148324] gi|153658|gb|M58002.1|STRHYDROLA [153658] gi|148323|gb|L06138.1|ENEVANB [148323] gi|475427|gb|U00681.1 |EFU00681 [475427] gi| 14832 l|gb|M85225.1|ENETETM [148321] gi|818704|gb|U24692.1 |EFU24692 [818704] gi|148320|gb|L00925.1|ENERTRNA [148320] gi|155036|gb|M97297.1|TRNVAN [155036] gij 148319|gbjL00924.1 |ENERRNA [ 148319] gi|150552|gb|M64978.1|PCFPRGAB [150552] gi|148317|gb|M81466.1|ENERECA [148317] gi|786274|gb|U22541.1 |EHU22541 [786274] gi|148315|gb|M81961.1|ENENAPA [148315] gi|786273|gb|U22540.1 |EHU22540 [786273] gi|148312|gbjM38386.1|ENEMSPDPS [148312] gij559858|gb|L37110.1|ADlCLYL [559858] gi|148310|gb|M37185.1|ENEGELE [148310] gi|643614|gb|U16659.1|ECU16659 [643614] gi|148307|gb|L07892.1|ENEBLACREG [148307] gi|643612|gb|U16658.1|ECU16658 [643612] gi|148305|gb|M60253.1|ENEBELAA [148305] gi|290641|gb|L13292.1|ENECOPPUMP [290641] gi|148303|gbjM77639.1|ENEB14NAM [148303] gi|624701 |gb|L29639.1 |ENEVANCRF [624701 ] gi|290644|gb|L 16515.1 |ENERGTG [290644] gi|624699|gbjL29638.1|ENEVANCR [624699] gi| 154954|gb|M37184.1 |TRN916 [154954] gi|624692|gb|L29641.1 |ENEDDLA [624692] gi|148301|gbjM69221.1|ENEAAD9A [148301] gi|624690|gb|L29640.1 |ENEDDL [624690] gij 148308|gb|M38052.1|ENECYLB [148308] gij493094|gbjL32813.1jENERRD [493094]
Table 28
Phage Dpi complete genome sequence. 56506 nucleotides.
1 ataataaaaa tatgaagcag atattgggtt aattattgct taacaaaatg caccgaattt gtgtataata
71 taagtgaagc agttttgtaa acctgacatc ctgctaaata aaaataaagg aggctcgaac atgagtcaaa
141 acactacacg cactgacgct gaattgacag gcgttactct tttaggaaac caagacacca aatacgatta
211 tgactataat ccagacgtcc ttgaaacttt ccctaacaaa catcctgaaa ataattacct agtaacattt
281 gacggatatg aattcacttc cctttgccct aaaacaggac agcctgactt cgcgaatgtt ttcattagtt
351 acattccaaa cgaaaagatg gttgaatcta aatcattgaa attgtactta ttcagtttcc gtaaccacgg
421 tgacttccac gaagattgca tgaacattat tttgaatgac ttgtatgaat tgatggaacc taagtacatt
491 gaagtcatgg gcctattcac tcctcgtggt ggaatttcaa tttacccatt cgtcaacaaa gtgaatcctc
561 aatttgcaac tcctgaactt gaacagcttc aacttcaacg caaattgaac ttccttggaa atgttcaagg
631 tcttggacga gctattcgat aggaggctgg aatgaaatca gtagttttat tatccggcgg agtcgactca
701 gccacttgtt tagcaattga agttgacaag tggggttcta aaaatgttca tgctatagca ttcaattacg
771 gacaaaagca tgaagcagaa cttgaaaatg ctgctaatgt tgcaatgttc tacggagtca agttcaccat
841 tcttgaaatt gactcgaaaa tctactcaag ctctagctct tccttattac aaggaaaagg cgaaatttca
911 catggaaaat cttacgctga aatcctagca gagaaggaag tagttgacac ctatgttcca tttagaaatg
981 gactaatgct ttcacaggct gcggcttatg cttattcggt tggagcttct tacgtcgtat atggtgctca
1051 cgcagacgat gcggctggag gtgcttaccc tgattgcact cctgagttct ataattcaat gtcaaatgca
1121 atggaatatg gaactggagg caaggtaacc cttgtcgctc ctctacttac tctaaccaag gcgcaagtcg
1191 ttaaatgggg aattgattta gatgttcctt atttcttgac tcgttcatgt tatgaaagtg acgctgaaag
1261 ttgtggaact tgcgcaactt gtatcgaccg caaaaaggca ttcgaagaaa atggaatgac tgaccctatt
1331 cattataagg agaattgata tgagagtttc taaaacctta acattcgacg cagctcatca actagttgga
1401 cattttggaa aatgcgcaaa tttgcacggg catacttaca aagtcgaaat ttcattagca ggcggaactt
1471 atgaccacgg ttcgagtcaa gggatggttg ttgactttta tcacgtcaag aaaatcgcag gtacattcat
1541 tgacagactt gaccacgctg ttcttcttca agggaatgaa ccaatcgctt tagcaaatgc agttgacacc
1611 aagcgagttc tatttggatt tagaactacg gctgagaata tgtcaagatt ccttacctgg actctcacgg
1681 agcttatgtg gaagcatgct cgtatcgact ctatcaaact atgggaaact cctacaggtt gcgcagaatg
1751 tacttactac gagattttca cagaagacga gattgaaatg ttcaagaacg taacctttat cgacaaagac
1821 gaaaagatta ctgtccgcga aattttagag caggagcagg ataatggtta atcaatacaa tcagcctgaa
1891 agaggcaaga ttcgaatcaa tgttcgcgac cctgagaaaa tgcctatcat ggaaattttc ggtcctacaa
1961 ttcaaggtga aggaatggtt ataggtcaaa agactatttt cattcgaact ggtggatgcg actatcattg
2031 caactggtgt gactcagcct ttacctggaa cggtactact gagccggaat atatcacagg caaagaagct
2101 gctagtcgaa tcttgaaact agctttcaat gataaaggtg aacagatttg taaccacgtg acattgactg
2171 gaggaaatcc tgccttaatc aacgagccta tggctaagat gatttcgatt ctaaaagaac atggattcaa
2241 gtttggtctc gaaactcaag gaactcgatt ccaagaatgg ttcaaagaag taagcgatat cactattagt
2311 cctaaaccgc cttcaagtgg aatgagaact aatatgaaaa ttcttgaagc tattgtagat agaatgaatg
2381 atgaaaacct tgactggtca tttaaaatcg ttatctttga cgaaaatgac ctagcttatg cgcgtgatat
2451 gtttaaaact ttcgaaggca agttacgtcc agtgaactac ctttcagttg ggaatgcaaa cgcatacgaa
2521 gaaggaaaaa tcagtgatag gcttcttgaa aagttgggat ggctttggga taaagtgtat gaagacccag
2591 ctttcaacaa tgttcgacct ttaccgcaac ttcatacact tgtttatgat aataaaagag gagtataaaa
2661 tgaaaattga gcatctagat aaaatcggta acgtattagg gagagagaac ggatgggctt cccttaagcc
2731 ggatgaaatt gtaaccttgg acaatactga ggcagccgtt caaagacttt ttggtctatt aggcgaggac
2801 gcagaacgtg acgggttgca agatactcca ttccgttttg ttaaagcact cgctgaacat accgtagggt
2871 atcgagaaga ccctaaactt catctcgaaa aaacattcga cgtcgaccat gaagaccttg ttcttgtgaa
2941 agacattcca ttcaattctt tatgtgagca tcatttagct ccgttcgtag ggaaggtgca tattgcatac
3011 attcctaagg ataagattac aggtctttca aaattcggtc gagtggttga aggatacgct aaacgacttc
3081 aagtacaaga gcgcttgact caacaaatcg ctgacgctat tcaggaagtt ctaaatcctc aagcagttgc
3151 ggtcatcgta gaggctgagc atacttgcat gagcggacgc ggtattaaga agcacggggc aacgacagtg
3221 acttcaacta tgcgaggtct tttccaagat gacgcatctg ctcgagcaga attgcttcag ttgattaaaa
3291 agtaggaggc ggaaaatgaa taaaagtgca accttttggc ttgttcgaac agctcttatt gcggctctat
3361 atgtgacatt gaccgttgca ttttctgcta ttagttatgg acctattcaa tttagagtca gtgaagcctt
3431 gattcttcta cctttatgga accatagatg gactccgggg attgtattag gaacaattat tgcaaacttc
3501 ttttcacctc ttggactgat tgacgtttta ttcggttcac ttgctacctt ccttggagta gtggcaatgg
3571 tgaaagttgc taagatggca agtcctctat attcacttat ctgtccagtt cttgctaatg cttaccttat
3641 tgcgctggaa cttcgaatag tttactcttt acctttttgg gaatctgtca tctatgtagg aattagtgaa
3711 gcgattatcg ttttaatttc atacttcctt atttccacgc tggcgaagaa caatcatttt agaacactga
3781 taggagcgaa aaatgggatt taatctatac ttcgcaggag gtcacgctat tagcactgac gattatttga
3851 aggaaagagg agccaatcgc ctattcaatc aactgtacga aagaaacggg attggcaaaa ggtggattga
3921 gcataagaaa accaatccaa gcactacttc aaaactattc gtcgactcta gtgcatattc tgctcatacc
3991 aaaggggctg aagttgacat tgacgcctat atcgaatacg tgaatgataa cgtgggaatg tttgactgta
4061 tcgccgaact cgataaaatt cctggtgtat ttagacagcc taagacacgt gaacagcttt tggaagcacc
4131 acaaatttct tgggataatt atctatacat gcgcgagcga atggttgaga aagacaagct cttacctatt - -
4201 ttccatatgg gagaagactt taaatggctc aacttgatgc tcgaaactac attcgaaggc gg~aaagcata ""
4271 ttccttacat tggaatttca ccagccaatg actcgactac gaagcataaa gacaagtgga tggaaagagt
4341 attcgaagtt attcgaaaca gttctaatcc agacgttaag actcacgcat ttgggatgac agttactagc
4411 caattagagc gtcacccatt ctatagcgcc gactctactt ctgtactgct cacaggagcg atgggaaaca
4481 ttatgacgtc aaaaggatta gttgacttgt cacagaagaa tggaggaatt gatgctgtcc gtaggctgcc
4551 aaaaccggtt caagttgaaa ttgaatccat tatcgaagaa actggagcgc attttagcct agagcaatta
4621 gttgaggact ataaacttcg agcattgttc aatgttcaat acatgctgaa ttgggcagag aactatgaat
4691 tcaagggaat taaaaatcgt caacgtcgac tattttagat aagagctttt cgctcttatt ttttttaaaa
4761 aaaaatgaac tttttataca aaaacgcttg actttattca ctcattatcg tataatcata atataaataa
4831 aacgaataag aggtaaataa aatgacagca gttcaacaag ttaagttcta cttagaagaa gccggcgctc
4901 actttctaaa agatgttgag tacagtgaca acttagagca agcaattatg aaagatattc ttaaatggaa
4971 tggcgctcat agagatgagc acgatatgaa aataacttca tacgaagtat tatagagagg ggtaaggcta
5041 tgaaaaaagt tcaaacttat caagaatatc taaaactagt tgagttcaaa cgtcaacttt ctttaaatct
5111 tcgagaagga aaaataggag tcgatgaagc ggttattcaa ttattcacct tctatagttt caacaatatc
5181 gaggaacctc ctttcattgt actcaaaatg caagaggctg ccgtgaacgg gacttatgaa gcaaaactca
5251 atatgcttaa aagatttaaa attatttaga aacggcttta caaactcgcg ataattcgtg tatattatat
5321 atatcaaaaa aaggaggctc atattatgag tattaagttc aaaaccgaag aactttcaaa aattgtttct
5391 cagctcaata agttgaagcc tagcaagttg ctagaaatca caaactattg gcatattttt ggtgacggcg
5461 aatgcgtcat gtttacagcg tatgatggct caaacttcct tcgatgcatt atcgacagcg atgttgaaat
5531 tgacgtgatt gtgaaagcag agcagtttgg aaaacttgta gaaaagacca cggccgcaac cgtcacatta
5601 gttcctgaag aatcttcgct aaaagttatt gggaatggtg agtacaatat tgatattgtt acagaagatg
5671 aagagtaccc tacattcgac cacttgctcg aagacgtgag tgaagaaaat gctctcactt tgaaaagctc
5741 gctgttctac ggaatcgcca atatcaacga ttctgcggta tctaaatcag gagcagatgg aatttatacc
5811 ggcttcctgt taaaaggcgg aaaagcaatt actacagaca tcattcgcgt atgtatcaac cctatcaagg
5881 aaaagggact agaaatgctc attccttaca acctaatgag tattttagca agtattcctg atgagaagat
5951 gtacttctgg caaattgacg atactactgt ctatatttca tcggcttcag tcgaaattta tggaaaattg
6021 atggaaggta tggaagatta tgaagacgtt tcacagcttg actcaattga gtttgaagat gatgcggcta
6091 tccctacagc agaaatcctg agcgtattag accgccttgt actattcact tcagcctttg acaaaggaac
6161 cgtcgaattc ttattcttga aagaccgact tcgaattaaa acttctacta gcagttatga agacatcatg
6231 tacgcatctg ctggcaagaa agtttcgaag aaagaattca cttgccacct taacagctta ctcttgaagg
6301 aaattgtatc aaccgtcacc gaagaaaact tcactgtctc ttatggaagc gaaaccgcaa ttaagatttc
6371 atcgaatggt gtcgtttact tcctagcact tcaagagccg gaagaataat ggccaagtcc aatttaacta
6441 gaattgcaaa gatggttaga gcaggaaaca gtgaaggtcc tgcttcatct tttgtcaatt cgctgacccg
6511 ggttattgaa cgaactcagc ctgaatataa tccttcgaca tattataagc ccagcggggt tggtggatgt
6581 attcgaaaaa tgtatttcga aagaatcggt gagtctatta tagataacgc agattctaac ctaattgcaa
6651 tgggcgaagc tggaacattt aggcacgaag ttctccaaga gtacatggtt aaaatggctg aaatcgatga
6721 ggactttgaa tggttgaatg tagcagagtt cttgaaagaa aatccagttg aaggaactat cgtcgacgag
6791 cgtttcaaga aaaacgatta tgaaacgaag tgtaagaacg aacttcttca actttcattc ttgtgtgacg
6861 gactagttcg atataaaggc aagctctaca ttttagagat taagactgaa accatgttca agttcactaa
6931 acatactgag ccctatgaag aacacaagat gcaagcaact tgctacggaa tgtgtctagg agtcgatgat
7001 gtcattttcc tttatgaaaa tcgagataac ttcgaaaaga aagcctacac gtttcacatc acagacgaga
7071 tgaaaaatca agtccttgga aaaattatga cctgcgaaga gtatgtagag aaaggcgaaa gtcctaaaat
7141 ctattgctct tcagcctatt gcccatattg tagaaaggaa ggtcgaaatc tgtgagctat actggaaaaa
7211 tgttcgagga agactttttc gaaggtgcaa aagactttga gaaagatgct ttcacggtcc gtctatatga
7281 taccactaat ggatttcgag gagttgcaaa tccctgcgat tatatagccg caactaactt tgggaccttg
7351 tttattgaac tgaaaactac taaagaagct tctttgagct ttaataacat cactgataat caatggttcc
7421 agctatcacg cgcagatgga tgcaaattta ttctcgccgg aattttagtg tatttccaaa agcatgaaaa
7491 gattatatgg tatccaattt caagccttga aaaaattaaa cggtctggag ttaaaagcgt caacccaaac
7561 ttcatcgatg cagggtatga agtttcttac aagaagcgtc gaactagatt gaccattcct ttccaaaatg
7631 ttctagatgc agttgagctt cattacaagg agaaaagcaa tggcaagacc taagttacct caaattgata
7701 ttcgagaaga agaaatacga gatgctcaag acgtagcaga ctcgtatggt gcgattatca ataaagtagt
7771 cgacgaaatt gttgaagcag cttgcggttc acttgaccag gcaatggaag aaattcaaat agttgtaagc
7841 caaaatcctg tcattatgga agaccttaac tactacattg gctatcttcc cactcttctt tatttcgccg
7911 cagatagggc ggaaatggtg ggaatacaaa tggattcaag ttctgctatc aggaaagaaa aatacgataa
7981 tctatacatt ttagccgccg ggaaaactat tcctgacaag caagcagaaa ctcgaaaact tgtcatgaat
8051 gaagaagtca tcgaaaatgc ttacaagcga gcctacaaga aagttcaatt aaagctagaa caggccgata
8121 aggtattagc atctttaaaa cgaattcaaa cctggcaact agcagagtta gaaactcagt caaataattc
8191 aaaaggagta ttattaaatg caaaaagacg tagacgtgaa aatgattgac cctaaacttg accgattaaa
8261 atacacaggt gattgggttg atgtacgaat tagttctatc actaaaattg acgccgacag cgccgatgtc
8331 tcaagatgtc gaaaagtgct tcaaaaggct caagtatatt cagtggcggc aggtgaatgc attaaaattg
8401 cacacggatt tgctcttgaa cttcctaagg gatatgaagc aatcttgcat cctcgttcca gtctttttaa
8471 gaaaactggt ctaatcttcg tttctagcgg agtgattgac gaaggttaca aaggtgacac tgatgaatgg
8541 ttctcagttt ggtatgctac tcgtgacgca gatatcttct acgaccaaag aattgcccaa tttagaattc
8611 aggaaaagca acctgctatc aagttcaatt tcgtagaatc tttaggaaat gcggctcgtg gaggccatgg
8681 aagtacaggt gatttctaat gaaattggaa cagttgatga aggactggaa taaggattcg aaagctcttg
8751 tagcagttca aggacttgaa cgtgaagcgc ttccaagaat ccctttttct gcgccttcta tgaattatca
8821 aacctacggc gggctccctc gaaaaagggt agttgaattc ttcggtcctg agtcaagtgg gaaaactact
8891 tcagctctcg acattgtcaa gaatgcgcaa atggtatttg agcaggaatg ggaacagaag actgaagaac
8961 tcaaggaaaa gctggaaaat gcgcgtgcat ccaaagctag caagactgct gtcaaggaac ttgaaatgca
9031 actcgatagt cttcaagagc ctcttaagat tgtatatctt gaccttgaga atacattaga cactgagtgg
9101 gctaaaaaga ttggagtcga tgttgacaat atttggatag ttcgccctga aatgaacagc gctgaagaaa
9171 tacttcaata tgttttagac attttcgaaa caggtgaagt tggcctagta gttctagatt ccttgcctta
9241 catggtcagt caaaacctta ttgatgaaga gttgactaaa aaggcctatg caggaatctc agcgcctttg
9311 actgaattta gtcgaaaggt tactcctctt cttactcgct acaatgcaat attcctaggc atcaatcaaa
9381 ttcgagaaga tatgaatagt cagtacaatg cctattcaac tccaggcgga aagatgtgga agcatgcttg
9451 tgcagttcga cttaaattta gaaaaggtga ctaccttgac gaaaacggtg catcattgac ccgtactgct
9521 cgaaaccctg cagggaatgt agtagagtca ttcgtcgaga agaccaaagc atttaagccg gacagaaaat^
9591 tagtttccta tacgctttcc tatcatgatg gaattcaaat tgaaaatgac cttgtagatg tcgctgfcga
9661 atttggagtc attcaaaagg caggggcatg gttcagtatc gtcgaccttg aaactggaga aattatgaca
9731 gatgaagacg aagaaccatt gaagttccaa ggcaaggcaa atctagttcg acgcttcaag gaggatgact
9801 acttattcga catggtgatg actgcggttc acgaaattat cactcgagaa gaaggctaat gcaaaaatct
9871 ctatttggac ctaagctagt gcctgctagt tcaaggcgca agaaaagaac ggttccaaaa cctaaaccta
9941 aaatcgatga gcaagtggtt gagcttatga accgcagaga gcgtcaagtg cttgttcata gttgcatcta
10011 ttattatttt aatgactcaa ttatagcaga cgggcagtat gacaaatgga gccacgaact atattctctt
10081 atagtttcgc accctgatga gtttcgacag actgttctct ataacgagtt taaacagttt gacggaaata
10151 ctggaatggg tcttccatac gactgtcagt ttgctgtaag ggtcgcagaa aggcttttaa gaaaatgaat
10221 ttagcttcta aataccgtcc tcaaactttc gaggaagtgg tagctcaaga atatgtcaaa gaaattcttt
10291 tgaatcaatt acaaaatggc gctatcaaac acggctatct attctgtggt ggcgctggaa ctggtaaaac
10361 cactactgct cgaattttcg cgaaggatgt gaacaaagga cttggctctc ctattgaaat tgatgctgct
10431 tctaataatg gggtagaaaa tgttcgaaac attattgaag attctagata caagtctatg gacagcgagt
10501 tcaaagttta catcattgac gaggttcata tgctttcaac cggagcattt aatgcgctgt tgaaaacatt
10571 agaagagccc tcatcgggaa ccgtgttcat tctatgtact actgaccctc aaaagattcc tgacactatt
10641 ctcagtcgag ttcaacggtt tgactttact cgaattgata atgacgacat cgttaatcaa cttcaattta
10711 ttatcgaaag tgaaaatgaa gaaggagctg gttatagtta tgagcgtgac gccctttcgt ttattgggaa
10781 acttgcaaat ggaggaatgc gtgacagtat cacaaggctc gaaaaagtcc ttgattatag tcatcacgtt
10851 gacatggaag ccgtttctaa tgcactagga gttccggact acgaaacatt cgcttcactt gttgaagcta
10921 ttgccaacta tgacggctca aagtgtttag aaattgtaaa tgacttccac tactcaggaa aagacttgaa
10991 attagtgact cgaaacttta cagacttcct tttagaggtt tgtaagtatt ggctagttcg agatatttca
11061 atcactcaac ttcctgctca ttttgaaagt aagctagagc aattctgtga ggcttttcaa tatcctactc
11131 tattgtggat gctagaagaa atgaatgaac ttgctggagt tgttaaatgg gagcctaatg ctaaaccgat
11201 aattgaaacc aaacttcttt tgatgagcaa ggaggagtga catgattgga cagggacttg ttaaatctac
11271 catttcgaaa tggaaacaac ttccaaaata tataatcgtc gaaggtgaag taggttcagg acggaagacc
11341 ttaatccgtt atattgcttc gaaatttgac gctgattcta ttgtagtagg aacgagtgta gatgacattc
11411 gaaacatcat tcaggatgca cagactattt tcaaggcgag aatctacgtg atagacggaa atagcctgtc
11481 aatgtcagct cttaactcgc ttttgaagat agcggaagag ccacctttaa actgtcatat agccatgact
11551 gttgatagca tcaataatgc tttacctacg cttgcaagta gagcaaaagt tctaaccatg ctaccttata
11621 ctaatgaaga gaaaatgcag tttgtcaagt cctacaagaa ggtagatact tcaggaattg acgaccgagc
11691 gattgtagac tattgcaatc ttgccagcaa tcttcaaatg cttgaagaca tattagaata tggcgcagaa
11761 gagctatttg aaaaggttac aacattttat gacttaatat gggaggcaag tgctagcaat tcgctaaagg
11831 ttactaattg gctcaaattt aaggaaactg atgaaggaaa aattgagcct aaacttttcc tcaactgtct
11901 tttaaattgg tcgacagttg tcatcaggaa gcactatgta gaaatgtctt tcgaagaact tgaggcccat
11971 gaccttttag tgagggaagc atctaggtgt ttgcgaaagg tatctaaaaa gggctcaaat gcgcgtgtct
12041 gcgtgaacga atttatcagg agggtcaaac aagttgagtg atttagtatc atttcaaaaa gacattcgaa
12111 ccaataatct aaagccgttc tatatcttgt acggcgaaga aattggtctt atgaatgttt atctcaatca
12181 aatgggaaat gtagttcgag aaacttcggt ttcaacagtc tggaaaaccc tcactcaaaa agggctcgtt
12251 tctaatcatc gaatattcgc tgttcgagat gataaggagt ttctgtctaa tgagtcgagg tggaaaaggc
12321 ttccggatgt tagatatggg acacttgttt tgatggttac taaaattgac aagcgaagca agttgctaaa
12391 ggcctttcct gataattgtg ttgagtttga gaaaatgact gacgcgcagt tgaaaaggca ttttgtgtct
12461 aaatactcga ctattgatag cgacatgatt gacatggtta tccagttctg tctaaacgat tactctagaa
12531 ttgacaatga attggacaag ctgtcgcgat tgaaaaaggt tgacgcatca gtagttgaat ccattgtcaa
12601 gcacaagacc gaaattgaca ttttcagcct agttgatgat gtattggaat ataggccgga gcaggcaatt
12671 atgaaagtga ctgaactttt agccaaagga gaaagtccta ttggattgct taccttgctt tatcaaaatt
12741 ttaataacgc ttgtcttgtg ctaggagccg atgagcctaa agaagccaat ctaggcatta agcagttctt
12811 aatcaataag attgtctata actttcaata cgagctggac tcagcctttg aaggcatggc tattttaggt
12881 caagctatcg agggcataaa gaatggtcgc tatacagaaa gttcagtggt ctatatttct ttgtataaaa
12951 ttttttcact tacttaacaa ataagctgaa atctgtgtat attacagtat aagcaaagga ggacagccta
13021 tgacagaagt tgcggtaaat agcccgcaaa aggtgagagt agttatggtc gggaatattg aatttctcga
13091 atatttaaaa aggaagtacg gaacagaaac ttccatcagt tatattatag aaaatgaaag gggtctaata
13161 tgacagactt taaaaaacgc ttcaagaaag cagtaacaga aacaatcaat cgtgacggta tcgagaacct
13231 tatggattgg ctcgaaaatg ataccaattt cttctcaagt ccagcaagca ctcgatacca tggaagctat
13301 gaaggtggac ttgtcgagca ctcattaaac gtgttcaatc aactactttt cgaaatggat accatggtag
13371 gcaaaggctg ggaagacatt tacccaatgg aaacagttgc aatcgtagca ctatttcacg acctttgcaa
13441 agttggtcag tatcgtgaaa ctgaaaaatg gcgcaagaac agcgacggtg aatgggaaag ctatttagca
13511 tatgaatacg accctgagca acttacaatg ggacatggtg caaaatctaa tttccttctt caacgtttca
13581 ttcaactcac gccagttgaa gctcaagcaa ttttctggca tatgggagcc tatgatatta gtccttatgc
13651 aaatttgaat ggatgtggag cagccttcga aactaatcca cttgcattct taatccatcg cgcagatatg
13721 gccgcaactt atgtagtcga aaatgaaaac ttcgaatact ctcaaggtcc agttgaacaa gaggctgagg
13791 ttgaagaagt agttgaagaa aaacctaaga gttcaactcg taagaaacct gcgcctaagg aagaaaaagt
13861 tgaagaggct gaagaaaaac caaaagctgg aatcactcga cgtcgcaaac ctgcgccaaa agaggaagag
13931 gtagaagagc ctaaagaaga gcctaagaaa gcatcttcta aaattcgaat gcctaaaaag actgaaaagg
14001 tcgaagaggt agaaagcgca gacgagccga aagttgaaga agcagaggac gacaatgtgg tggtacctgc
14071 tggatatgtt cgagatgtct actacttcta cagtgaagtc gctgacgttt actacaagaa agatgtcgac
14141 gagcctgacg atgacagcga cattcttgta gacgaagaag agtacatgga cgcaatgtgt cctgtattag
14211 aagaagactt cttctacgaa cttgacggca aggttcacaa attagcaaaa ggtgaacgct tgccggaaga
14281 atacgacgaa gaaacttggg aacctatcac tgaagcagaa tacatcaagc gaacagaaaa acctaaagca
14351 gttgcaaaac ctactcgaaa aactccagcg ccttctcgtc gccctcgccc ttaaaagaaa ggttgaaata
14421 aaatgtgtga aaattgtcaa aacgaaacat tcaatactag aattttcaat gaagatgaaa gtggctatgt
14491 cgacgcctca ttcacttaca aggagattcg cgacaccgca gcagctatta gcaatcgagc ggtagaaaag
14561 aaagaccgtg acagcctttt agtcgctaca gttatggctc ttcccgtttc tcacgcagaa gatttaggca
14631 agagactttg tattgcaaat tctcgattgg aagcatttcg tgaagctgtt caagaggctc tcgagaatga
14701 aaaggctgaa gatttaaagg acgttatctt aggtcttatc gacgttgaca aaaaaattgg caaccttgca
14771 ttgcaattag ttgaatcagg agcattataa tggaacgaat aaagacgcta tttcacgtga tttatgctaa
14841 cggcactcat ttagaagtag cagctttgtt cgataccgtt gatgattatg atgacgttat agaggacatc
14911 caggggtata ttgatacccc tgacctttat aatcaaagga gcattagaat ggcgccttac aatcctgaca
14981 tcaatggtga cgctattgct actgacattt tactacgact agatgatatt atctacgtcg acgcaacttg
15051 tgaaactatt aaatacgagg agcctattgc atgaacaatc agcgaaagca aatgaacaaa cgaatcgtcg
15121 aacttcgcga agactatcaa cgtgcaagag gtcgaataaa cttccttctt gctgtaaagg accacggcga
15191 agaactcgaa aaccttgaag cctttgtggg atacattgac aatctagtcg aatgttttcc tgaaagccaa
15261 cgaaatgtct tgaggctatg tgtattagat gaccttccag tcactaatgc ggccgctgaa attggatacc
15331 actatacatg ggttcaccaa cttcgagaca aagcagttga aacacttgaa gaaattttag atggggataa
15401 cattattcgc tctaaacacg gaatcgaaat taaggagaaa cttgatgaat tatatggtaa aagtcattct
15471 agttagtgtc tttgtactgt cagccttttg catgacttgc tcaatggttt atttggttac aggtaagcaa
15541 gaggaccacc gtagtaccgt cgcccttgta tttggcgctc tcgtaagctc tgcggcgttc tattcgacac
15611 tctttatcct cgcctatctg ccatgacatc acgcgcatac aaaccaattc ccacgcgcag agctagtgct
15681 aaacaagaga aggcagttgc taagcagttg ggaggaaaag tacagcctaa ttcaggagcc actgactact
15751 acaaaggtga cgtcgtaaca gactcaatgc ttatagaatg caagacagtt atgaagccac aaagttcagt
15821 cagcttgaaa aaggaatggt tcctaaaaaa tgaacaggaa aggttcgctc aaaaactcga ctattctgct
15891 atcgctttcg actttggtga cggaggcgaa cagtatatag caatgtctat aagtcagttc aagcgaatat
15961 tagaggatag aaatgataac cttatttaaa ataaacagtg aaggaacagt tactccaatt aaagggtcag
16031 ccatgcaact gtacgcagac cttattccta tacaagagga cgatatacag ttcgttgata taactggact
16101 tgaccctatt gttcgagaaa acgtacttga gctcatttca cggagccgtg taggagtttc aaaatatggt
16171 acaaacctcg accagaatga tgtcgacgat ttcctacagc acgccaaaga agaagcgctc gactttgcta
16241 actacctaac caagctacaa agtcaacaaa agcaaaataa atagacctat ttctaggtct atttttatta
16311 ttgataaatt ccagcaattt gacgagcgca atcttctagc gcagatacta ggtggcggct ttcttgttta
16381 ccttgttcat ttcttgcttt aattctttcg ttaaggcgtt cgattcttgt agttaatttc ttgatgattt
16451 caattctagc atcaacttcc atgtcgcgag taagtgtgac tccagtttca gcgacaggac atgctttgaa
16521 tactgcaatg tcaagttcgc tctttctaat aactgagcct aggtctaagt acaagttagg attgattcca
16591 gtgaccttat attgtttctc agtttctttt acaggaatgc tttcatagtg gaaagtgtag ttcttgtgac
16661 cgtctttcca atctgctgta agataaccga aataaagtgt tgtttccata attgacctct ttctgcgtcc
16731 ttgacgcttg ttttatttat attatgatta tacgataata aaggaataaa gtcaagcact ttttacaaaa
16801 aagttgaact tttttaaata tttttttttg aaaataaaaa gccctaataa tagagctttt agtttagcag
16871 aaaattaagt tcatcttcat aagcaagaat ctgtccgtac tggtaagaaa tagctgattc aatatccggc
16941 atttcgtgga ctcctttttt aagttcgtcg atagtacagt tacaatgacc tattcttgac tgaagttcct
17011 caatcctttc gagtcgcttt tcattttgtg tatcaattgt tttcgagtct aggtgagtga aggaacttgc
17081 aatagtttga atggcttcaa aaaagtccgt tattgaaact cctttataag aaagctcatt ccgtgtatag
17151 caggaaagca aagcgttcca gctagtgatt tgaatttgag ggttaggaga gtttcgataa gctacaaaat
17221 ttagaatatc tttgtagtca atatcagctt cagtatgatt gttgataaat accttcattt tataaccctt
17291 ccaaatcttc gtcctcgtca tcgttttcat agcaggcgat aacttcaacc cactcgtcgt cctcaccttc
17361 gtttcgaact cgaatgctaa ggacttccat gtcctcaaca tcttcgaatc cttcattagg tgcatatcct
17431 tcccactcta aatcgtcgta gtcgaagata gttacaagac gtccgtcaaa ttttactgtt tcctttactg
17501 ttgccatttt agtttcctcc ttatgcgata tatagtttga taatttgaga ttcgatgtca ccatagttga
17571 tgaacttaac ttggtcgacc gtttcttcca tgtattcgcc catgtcttcg attcttccgt cttgaatcat
17641 ttggccgttt tcgttgataa tttcgtacca ccattcatca ccgaattgtt tgattgcttc tttaactgtt
17711 ttcattttac tacctccact ttttcgtcca ttagtgattc gttatcatag aaccgaatac gtccatcact
17781 aagacgttct aggcttaccc atttacgacc ttgacggtca gttactttaa attcagtacc ttttgcattt
17851 acaactttca ttcctacttg caaatcttta acttttacca ttttatatga ctcctttatt tgtttttctt
17921 tatagtatta ttatacgata atgagtgaat aaagtcaagt gtttttgtaa acttttttaa attttttaat
17991 tttttttttc aaaaaaataa cgagccgaag ctacgttatt tatttatctg ctcaagggct tgttgaattg
18061 cctcatagcc tttacgacgt gctacctttc cagctttaga gccgggtgaa aagtcccaaa cagtttcgtc
18131 tactttaaag tcatccgcct tggcatagtc gagcaggagc tggatagctt tttgccattt ccgccaattc
18201 ttggaaaact cacctatatt agcacaacgc aaaacaagtg ctctagtatg ctggctagac ataatgaact
18271 ctaaaaagtt gtccaaggtt ataggaaggt cctttggaaa ctcataaggc tctttgacat cgtatttgaa
18341 aaggctgaca atttcactgt ccttaaatag ttcaccgtct ttatacataa taccttgaac aatttcagta
18411 ggctctgctc cgctatctag tacatcgcca accgtgtgac aataggcttt aagaactgca aaaaaacctg
18481 gggcgtctgc acgcgcaacc tggagctcct taacagtcat ccaaggctga ggtttcttac aaacaatcct
18551 aattccttca aaatagctct tgtccgggtc aatagtgcct aacattgtca gcctgttttt atttatataa
18621 aggtcgaaat atacttgaat ttcatctgta ttaggcagcc acttaacagt gacttttcta taagcgattg
18691 cttttacatt tacttttttc gagagatttg tagggataag cattttcctt ttgacattta ctttttttcg
18761 ctttttgttc tttgccatgc tagtatctcc atttctgttg gtcttgcttt ttagctctgt tcagttcagc
18831 tgcttctcgc gatgcaatag tttcgagaat atgcctgttc ataggctcac aatattccgc caaagatttg
18901 ccagttatgg tggcgtcaat taagtaacca tctattgact ccttaccata aaatacaaaa tcgtcttggc
18971 atactagcct tttataatag ccatttcctg cgcgtgtttc aattttaact aagctcattt tcacccaaac
19041 ttgtagacga taaggagttc ctggaacttc gaacaggagc ctcctttttt catcgtctac ttgtttaata
19111 catgagtttt gaaaatggat aactttccat ttattttcca tagtttcacc ttattccatg tacccgtcaa
19181 caatccataa ttgaaaaggc ttatcttctc tataaggccg tgataatttt agtccagttc ccactacatt
19251 tgaaagcgcg attaggtcat ctaggctgtc tagctcgagt tcgattacaa ggttgccagt atcaatttca
19321 caaaagtaag cgacatttcc aactttctct agtgcttcac gatacctatc atatgtcgcc tcttcgtcaa
19391 atagtcgcgc agaataaact tcgaatttca ttttagttac cgccttccaa aatttcatcg ggcataatct
19461 ttgcattctc gccatgaaac cgcccttcaa tatacgcttc aagattgaag tcatgttgag gtctgtcaat
19531 tccttccttc tttaaatttc gaaatgtgtc ctgaagcgca ttttttgttt gctcgctagg taggaccata
19601 agtgaatatt cttccacctg ctttttaaat cgaatggcta aggctgacaa aaagcctttg aggtatgaat
19671 tcttgtagga aggttcgcga gtaggaagtc ggtcaatacg gtaacgaaga taaagcaaag cagcctcata
19741 tattttagac actaattcag cgtcttgttt ttcgccgaag aaaattattc gacttttatt caagcgcata
19811 tcacgctgat taatacaaaa gcacctaaaa ttagtcgcga gaatatgacc aagttcacgt tcccaccaaa
19881 atattcgacc tgcttctttc ccaacagctt gagaagtctc gaactgttta ggttcatcaa attgttcaac
19951 ttgagcaagt gcgatattat tctttagcat caacttttga gccataagaa gggcagtttg cccctcttcg
20021 tcactcgggt tgtcatttgc taattgaata agatttttaa ttttttcaat aattttttcg ttattcatat
20091 tagtcacttt ctatcatatt ttcgagcttt cgaaaagtca atgtcgtcta cttcaattgt cttgtcataa
20161 gtccaagcgc gacaagtgtc gaaatgaaat aggctacaaa acatcttttc attatggtcg aaactttcag_
20231 tacatttttc aatatctact tcaagttcga gaacgacaat agtatcaaca tttcgaagcg ataaaaaggc
20301 tagagccttt tcataacttt ctgctaggta aataactcca gctgaaggct tcaatccttc agctagaatt
20371 ttaccaagat tatcaaaatc agtggcgtga taaagtttca ttagttactt ccttacatat ctagagtcac
20441 tacataaata gaagcagttt tatcttccaa gtcctactca atagcttcct cttcgctgag tttttcgagt
20511 tttaaaactg tcgcttcagc tacaacatta gcaaagttcg aaccgttgag aatgttttcg atatttcctg
20581 cgcctaagac ttcagcttgg tcattgttca ctaccattag gtattcatta gtaagtgctt tagcaaagtt
20651 tgaaaatttc attttatttt ccctttattt gtttttcttt atactattat tatacaataa tgattgaata
20721 aagtaaagca ttttttataa aaaagttgaa ctttttttac aattttttga actatttaaa aattataaaa
20791 tgggtggaaa atttaggcga caatttatac ccattttcaa cctcatttat aaacaatcta atatagaaaa
20861 ggacttaata agtaaataaa aaagcgccct gaaaatacct acaaatccca tagtccgtaa gtaaaaacaa
20931 aaattagggg cgacataaaa gtcgagcact atcttaatct attaccagtc tcatatacaa tcgacacaga
21001 tttagcaggc ttttagcaaa ctttcgaaca gcatgaaaaa gcatacaatt agaggaacag attatagaaa
21071 aagcacttcc acaaacaagt tctcaaaatg ctctcaaaaa ccgtaaaatt agtaagtttg aacttttcga
21141 acttctaaac ttttcgaata atcgagccta atttagaggt cgaaaaactc aatttctcga aaagtcgaac
21211 ctgctcgaaa acctcaaaac actcgaaaag tcgagcatag aaaggggtcg aaaagtcgag aatgctcgaa
21281 aaactcaacc ggttcgaaaa cctcaatcct tcgaaaagtc gaaccattcg aaaagttcaa aagttcgaaa
21351 aactcaacca ttcgagagta ggaattaagg acataccagt tcaacctttt tagcttcaaa atcactcttt
21421 ttctcattat aggactataa attcagtcaa ttgtaagtca cgcgcaaatt tgttacaatg taaacgataa
21491 aatataaagg agggtcaata aatggcgaaa gctactggac caaaagttcg aagaggaaaa actcctccac
21561 ggccaaaaga caaaaaagga atcaaagcaa atgcgcgtgt caataaagac cagttcgtag agtatgacta
21631 taaaggcatc aagatgacaa ttaaggaacg tgatgctaga atgaaattgg aatttattag aggcatgact
21701 attcaggaaa ttgcagcccg ctatggatta aatgaaaagc gtgttggcga aatacgggct cgcgataaat
21771 gggtgaaggc taagaaagag ttcgagaatg aaaaggctct tgttactaat gatacattga ctcaaatgta
21841 tgcagggttt aaagtctcag tcaatattaa atatcacgcc gcctgggaga aactaatgaa catcgtcgaa
21911 atgtgtttag ataatcctga cagatattta tttactaaag aaggaaatat tagatggggc gcattagatg
21981 tcctttcgaa ccttatagat agagctcaaa aaggacaaga aagagcgaat ggaatgcttc cggaagaggt
22051 tcgatataga ctacaaattg agcgcgagaa aattacattg ctccgggcca aaatgggcga ccaggaaatt
22121 gaaggcgagg ttaaagataa cttcgtagaa gcactagata aagcagctca agccgtttgg caagaattta
22191 gtgacgcaac aggttcctac attaaaggag tgactgataa tgacaataag cctgagaaat aaactaccta
22261 agttcaactt cgtccctttt agtaagaaac aactccagct cctaacatgg tggacaaagg gctcaccttt
22331 tcgaactttc gatatcgtca tagcagacgg ttccattcgt tcaggaaaaa cagtatcgat ggctctttca
22401 ttttcccttt gggccatgac ggaattcaac ggacaaaact ttgccatctg tggtaagaca attcactcag
22471 ctcgacgaaa tgttattcag cctctaaagc aaatgctcac aagtcgcggg tatgaaattc gagatgttcg
22541 aaatgaaaat ctacttatta ttagacactt tagaaatggc gaagaaattg tcaactactt ctatatattt
22611 ggaggaaaag atgagtcgag tcaagacctt atacaggggg taacattagc aggtatcttc tgtgatgagg
22681 tggcactgat gcctgaatcg tttgtcaacc aagcgacagg gcgctgttcc gtaacaggtt cgaaaatgtg
22751 gttctcttgt aacccggcca atcctaatca ctacttcaag aagaactgga ttgacaaaca ggtcgaaaag
22821 cgtatcttat atcttcactt tacaatggac gacaacccta gcttgacgga tagcattaaa aggcgctatg
22891 agaaaatgta tgctggagtc ttcaggaaaa gatttattct cggcctttgg gtaacagcag atggtctagt
22961 ttattcaatg ttcaatgaag agcagcatgt caaaaagctc aatatagaat tcgaccgttt attcgtagca
23031 ggcgactttg gtatctataa tgcaacaacc ttcggccttt atggattctc gaaacgtcat aagcgctacc
23101 atctaattga gtcatactac cactcagggc gcgaggcgga agagcaacta actgaggcgg atgttaattc
23171 gaatattcaa tttagttcag ttctacaaaa gactactaaa gagtacgcaa atgatttagt cgatatgata
23241 cgaggaaagc aaatcgaata tataattctc gacccgtctg cttctgctat gattgttgaa cttcaaaagc
23311 atccttatat agctagaaag aatatcccta tcattcctgc tcgaaatgac gtgacgcttg gcatttcatt
23381 tcacgctgaa ctcttggctg agaatagatt tacactcgac cctagcaaca cgcacgacat tgatgaatac
23451 tatgcttaca gctgggacag taaagcgagc caaacgggag aagatagagt cattaaagag catgaccact
23521 gcatggatag gaacagatat gcctgtctca ctgacgctct aatcaacgat gacttcggtt tcgaaataca
23591 aatattatcc ggaaaaggcg ctagaaacta actaaacact tttatagaaa ttagtgtata atataagtag
23661 gaggatttta aacatggcta aaaaatcaaa agctatctca cacacagacg aactgattag tcagtcgttt
23731 gacagcccct tggcaaagaa tcaaaagttc aagaaagagc ttcaggaagt tgaaaagtat tatcaatact
23801 tcgacggatt tgatgtcacg gacttgaata ctgactatgg gcaaacatgg aagattgacg aagactcagt
23871 cgactataaa cctactcgag aaattcgaaa ctatattcga caacttatca aaaagcaatc acgctttatg
23941 atgggtaaag agccagagct tatctttagt ccagttcaag acaatcaaga tgaacaggct gagaacaagc
24011 gtattctatt cgactctatt ttaaggaatt gtaaattctg gagcaaaagt acaaatgcat tagtcgacgc
24081 cacagtaggt aagcgggtat tgatgacagt agtagcaaat gccgctcaac aaattgacgt ccagttttat
24151 tcaatgcctc agttcaccta tacagttgac cctagaaacc cttccagctt gctttctgtt gacattgttt
24221 atcaggacga gcgtacaaaa ggaatgagca ctgaaaaaca actttggcat cattatagat atgaaatgaa
24291 agctggaaca agtcaatcag gaattgcaac agctttagaa gacattgaag aacaatgttg gctcacttat
24361 gccttaacgg atggagagtc gaaccaaatc tatatgacag aaagtggcca aactactatc aaggagacag
24431 aggctaaact tgtagaaatt gaagacaacc taggaaacaa gattgaagtt cctttaaaag ttcaagaatc
24501 cgccccaacc ggcttgaagc aaattccttg tcgagttatt cttaatgaac cattgactaa tgacatatac
24571 gggacaagcg atgtcaaaga ccttatcaca gtagcagata acttgaacaa aactattagt gacttacgag
24641 attcacttcg atttaaaatg ttcgagcagc ctgttatcat tgatggctct tctaagtcaa ttcaaggaat
24711 gaagattgcg ccaaacgctt tggtcgacct taagagtgac cctacttcct caatcggcgg tactggaggc
24781 aagcaagctc aagtcacttc catttcagga aacttcaact tccttccagc ggctgaatat tatttagagg
24851 gcgctaagaa agccatgtat gaactaatgg accagccaat gcctgaaaag gtacaggagg cgccatcagg
24921 aattgcaatg cagttcttat tctacgacct aatttctcga tgtgacggaa aatggattga gtgggatgat
24991 gctattcaat ggctcattca aatgctggaa gaaattttag caacagtgaa tgttgacttg ggaaatattc
25061 ctcaagatat tcaatcaagt tatcaaacac ttacgacaat gactatcgaa caccactatc caattcctag
25131 cgatgaactt tctgctaagc aacttgcgct cactgaagtt caaactaatg tacgcagcca ccaatcttac
25201 attgaagaat tcagtaagaa ggaaaaggcg gacaaggaat gggaacgcat tttggaagaa cttgctcagc
25271 ttgacgaaat ctcagctgga gcattgcctg tattagcaaa cgaattaaac gaacaagagg agcctcaaga
25341 tgaaacgagt gaagaagacg aagttgatga caaagaaaaa gaacaaactg aacaaccaac cgaagaagga
25411 gtcgacccag acgttcaagg ttaattgtga ccattgtgag cataagttcg accttacatc taaacagatt
25481 atttcgaaac atatcgaaaa gggcgtagag tggagattct tcgaatgtcc taagtgccat tatcggtfeca
25551 ccacttatgt aggaaacaag gaaattgaaa accttattcg atttagaaat acttgtcgag ctaaaatgaa
25621 gcaggaactt caaaaaggag ctgctgctaa tcaaaacact taccattcat atcgaattca ggatgagcaa
25691 gctgggcata aaatctcagg gcttatggcg aagctaaaga aggagataaa cattgaaaaa cgagaaaaag
25761 aatgggtatc tatatagctg ggaaaaggct attcatgaaa ataatattcg tctaaccctt gaacaggaac
25831 aagctgtact gaaagccttc agcgatgcag gaactgattt aattgcaaag attaaaaagt ctcgaaatgg
25901 atacttgcct aaaagaatct ataaagacta cgcttacgac ctgcacgctg ttcttgttca actaatgact
25971 gaatactctc ataaggcggc aatgaacgca gtagatggcc aggtagttca tattctacaa gtattagcag
J. j
26041 aagatggaaa tgctacggct gaaaagttcg aaaaggaagt cagggctgca tctttagtat tttcacgaag
26111 agcagccgag gcagttgtca aaggtgaaat ctataaggac ggcaaaaacc tctcgaaacg tgtttggtct
26181 tcagccgcac gcgcaggaaa tgatgttcaa caaatagtca cacaaggcct agcaagtgga atgtctgcta
26251 cagatatggc taaaatgctc gagaaatata tcgaccctaa ggttcgaaaa gattgggact ttgataagat
26321 agctgagaag ctagggaaac ctgctgctca taaatatcaa aatctcgaat acaatgccct tcgacttgct
26391 cgaactacca ttagccattc cgccacagct ggagtgagac aatggggcaa ggttaatcct tatgctcgaa
26461 aagttcaatg gcattctgtt cacgctccag gtcgaacgtg tcaagcgtgt atcgatttag atggtgaagt
26531 atttcctatc gaagaatgtc ctttcgacca tcctaatgga atgtgctacc aaactgtatg gtacgaaaac
26601 tcactcgaag aaatcgctga tgagttgaga ggctgggtag acggagaacc taatgatgta ttagacgaat
26671 ggtacgacga tttaagttca ggaaaagttg agaaatacag cgacctcgac tttgttaaaa gttattaggc
26741 tcggttcaat accgagtctt tttgtctata aattgtctaa tttcgagaac cttcgaaaag tagtaaaatg
26811 atattcagtt atgttataat ataagttgaa aaggaacctt gtcgccttaa tgactcgaaa ttggtttcac
26881 tgttccaatt aaataaaaac agcagattca gccggagggc ggaaaactca ggaggaaaat aaatggctta
26951 tcaattagaa gacttgttaa aaggtctaga tgaaccaact atcaaacagg tgaaggaaat tatttcgaaa
27021 acttcgaaag aactcgatgc taaaattttc attgacggcg acggtcaaca ttttgtacct cacgcacgtt
27091 tcgatgaagt tgttcaacag cgcgatgcag ctaacggctc aattaattct ataaagaac aagtcgcgac
27161 gctttctaaa caggtcaaag ataacggtga tgcgcagacc actatccaaa accttcaaga gcaactcgac
27231 aagcagtctc aacttgcaaa aggcgctgtg attacttcag ctcttcatcc gttgattagt gactccattg
27301 ctccagcagc agacattctt ggatttatga accttgacaa cattacggtc gaaagtgacg gtaaagttaa
27371 aggtcttgat gaagagttga aagctgttcg tgagtctcgt aaatacttat tcaaagaagt cgaagttccc
27441 gcagaacaag aggctcaagc taagtcgcca gccgggactg gaaatttagg aaatccaggt cgtgtcggtg
27511 gtggtgttcc cgaacctcgt gaaatcggct cttttggtaa gcaacttgct gctgctcaac aaacggcagg
27581 agcacaagaa caatcatcat tctttaaata ataggaggaa ctaactatgc ctaatgtgcg agttaagaaa
27651 actgatttta atcaaaccac tcgaagcatt gtcgcaattc ctgaccacta cgttgctttg gctgctcaaa
27721 ttccagctac cgcagcaact caagtaggga acaagaaata cattcttgcc ggaacttgcg tgaaaaatgc
27791 tactacattt gaaggacgca aaactggact cgaagtagta tctaccggtg aacaattcga cggagttatc
27861 ttcgctgacc aagaagtgtt tgaaggtgaa gaaaaagtaa ccgtgacagt attagttcac ggattcgtca
27931 aatatgcagc ccttcgaaaa gttggcgatg ctgtgcctga atctaaaaac gcaatgattc ttgtcgttaa
28001 ataggaggaa ttatagatga atatttatga ttatatcaac gcaggggaga ttgctagcta cattcaagca
28071 cttccttcaa acgctcttca ataccttgga ccaactcttt tccctaatgc tcaacaaaca gggacagaca
28141 tttcatggct caagggtgca aataatttgc cagtaactat ccagccatct aactacgacg cgaaagcaag
28211 tcttcgtgaa cgtgctggat ttagcaaaca agctactgag atggcattct tccgtgagtc tatgcgactt
28281 ggtgaaaaag accgtcaaaa cttgcaaatg ctattgaacc aaagttcagc tcttgcccaa ccacttatca
28351 ctcaactcta taatgatact aagaaccttg tagacggtgt tgaagcgcaa gcagaataca tgcgtatgca
28421 attgcttcaa tacggtaaat tcactgtcaa atcaactaac agcgaggctc aatacactta cgactacaac
28491 atggatgcta agcaacaata tgcagtcact aagaaatgga ctaacccagc tgaaagtgac cctatcgctg
28561 acattttagc agcaatggat gacatcgaaa atcgtacagg tgttcgccct actcgaatgg tcttgaaccg
28631 aaacacttat aaccaaatga ctaagagtga ctctatcaag aaagctcttg caattggtgt tcaaggttct
28701 tgggaaaact tcttgcttct tgcaagtgac gctgagaaat tcatcgctga aaaaacaggt cttcaaatcg
28771 ctgtctactc taagaaaatt gctcagttcg ctgacgctga caaacttcct gacgttggta acattcgtca
28841 gttcaacttg attgacgacg gtaaagtggt attgcttcca cctgacgcag ttggtcacac ttggtacggt
28911 actactccag aagcattcga cttggcttca ggcggaacag acgctcaagt tcaagttctt tcaggcggac
28981 ctaccgttac aacttatctt gaaaaacatc ctgtcaacat tgcaacagtt gtatcagctg ttatgattcc
29051 atcattcgaa ggaattgact atgtaggagt tctcacaact aattaggagg tcgctatatg gctacattga
29121 aagctcttag caccttaatc gtttccggag cagtagtgca ttcagggtcg gtattttctt gccctgaagc
29191 gcttgcttcg tctttaattg aacgcaattt tgcgttcgag attaaggcgg ctgaagatgg agaaacggta
29261 gaaactgttc ctcaaacaat tgaatcagtt gaagaaattg acgaagttga acaaatgcgc gaagagtatg
29331 cggctaaaac cgttcctgag ctcgttgaat tagcaagagc taatggaatt gacatttctt caatttctcg
29401 aaaaagcgaa tatatcgacg ctttaattaa gtacgaacta ggagagtaaa atggcagctc aaacggacat
29471 tgaattagtc aaaatcaata tcgataacga taattctccg tcaccaatga ctgaccaaag tatctcagct
29541 cttttagaca agcataaatc tgtcgcctat gttagttata tgatttgctt aatgaagacc cggaatgacg
29611 tggtaaccct tggacctatc agtctaaaag gtgacgcaga ctactggaaa caaatggcgc aattctatta
29681 tgaccaatat aagcaagaac agcttgaaac tgatgaaaag tcgaacgctg gttcgacaat cttaatgaaa
29751 agggctgatg ggacatgagt tatgacgtga attatgttaa gaatcaagtt cgtagagcca ttgaaaccgc
29821 tcctactaaa atcaaggtac ttcgaaactc ttgggtcagt gatggatatg gaggaaagaa aaaggataaa
29891 gcgaatgaag tcgtagcaga cgaccttgtt tgtttagttg ataattcaac tgttcctgac cttttagcca
29961 attctactga cgcgggaaaa atttttgccc aaaatggagt gaaaattttc attctatatg atgaaggcaa
30031 aatcattcaa cgagccgata ctatcgaaat taaaaactca ggaagacggt acagggtagt agaaacccac
30101 aatcttctcg agcaagacat tttgatagaa cttaaattgg aggtgaacga ctaatgtctc agcctgaatt
30171 agtatggaag cctgaagaat ttgttagtaa ctgtgaacgg tatcgaaaca agtttcaagt cgctgtcata
30241 acagtctgcg aagtcgctgc tactaagatg gaagaatacg caaagacgca tgctatttgg acagaccgta
30311 cagggaatgc tcgacagaaa ctcaaaggag aagctgcttg ggtaagcgca gaccaaatca tgatagctgt
30381 atcacatcac atggactacg ggttttggct agaactagct catggtcgaa aatacaaaat tctcgaacag
30451 gctgtagaag acaatgtcga agaacttttt agagcgttga gaaggttatt agactaggag tgaacatgac
30521 taaacgaacg acaatgatgg acagattgaa ggaaattctt cctacatttc agctctcgcc tgctcctatg
30591 cttccaggag ttgaatttga cgagcaagat acagataggc cggatgacta cattgttctt cgatatagtc
30661 atagaatgcc cagcgcaaca aatagcctag gaagttttgc ttattggaaa gttcaaatct acgtccattc
30731 aaactcaatt attggtatcg acgaatatag cagaaaggtt cgaaacatta tcaaggacat gggctacgaa
30801 gtaacctatg cagaaactgg tgactacttc gacacaatgc tttctagata ccgactagaa atcgaatata
30871 gaattccaca aggaggaaac taataatgag taaagacatt ctttacggaa tcaagctcgt gcaaaTcgag
30941 gagcttgacc cattgactca gttgccaaaa gtcggcggag ctaactttgt cgtagatacg gcagaaacag
31011 cagaactcga agccgtgacc tcggagggaa ctgaagatgt gaaacgcaat gacacgcgca ttcttgctat
31081 cgtgcgtact ccagaccttt tatacggtta tgacttaaca ttcaaggaca acacgtttga ccctgaaatc
31151 atggccctaa ttgaaggtgg tacagtacgt caacaaggcg gaactattgc tggatacgac accccaatgc
31221 ttgcacaagg tgcttctaat atgaaaccat ttagaatgaa catctatgtg ccaaactatg taggtgactc
31291 aattgtcaac tacgtgaaaa tcactttgaa taactgtacc ggtaaagctc cagggctttc aatcgggaaa
31361 gagttctacg ctcctgagtt caacatcaag gcacgtgaag caaccaaagc aggtttgcca gttaagtcaa
31431 tggactatgt ggcacaactt ccagcggttc ttcgtcgcgt gacattcgat ttgaacggtg gaacaggaac
31501 cgccgacgca gt cgagttg aagcaggtaa gaagatttct ccaaaaccag ttgaccctac cttaaσaggt
31571 aaggctttca aaggctggaa agttgaagga gaatcaacta tttgggactt cgacaaccac atgatgcctg
31641 accgagacgt caaactcgta gcacaatttg catagaaatt tagaaagaag ggtctgttat gactaatatt
31711 atcacagctg agcagtttaa gcaacttgca tttcaaatca tcgcacttcc aggattttca aaaggtagtg
31781 aacctatcca tgttaaaatt cgagcagcag gtgtcatgaa cctaatcgct aacgggaaaa tccctaatac
31851 gcttttaggt aaagtgacag aactgtttgg agaaacttcg acagtcacta aagacaatgc tagtctagca
31921 tcaattactg accaacagaa gaaagaagcg ctcgaccgat tgaacaaaac cgataccggt attcaagaca
31991 tggctgaact tcttcgagta ttcgcagaag cttcaatggt agagcctact tacgctgaag tcggcgagta
32061 tatgacagat gagcaactta tgacaatctt cagtgcaatg tacggtgaag tgactcaagc tgaaaccttt
32131 cgtacagacg aaggaaatgt ctaatgtcat agcagtcgct actgaatttc atattagacc tagcgaggtg
32201 gtcgggatgc aaactgattt aggcaaatac tgcttcgacg cagcagccgt tgcttatatt agatatttgc
32271 aggaagacaa gactcctagg tatcctggtg acgaaaagaa aaatccagga ttgcaaatgc ttatggagtg
32341 actattttca gtcgctcctc tttttgtata tagaaaggaa attacatgga ttttgggtca attgcagcaa
32411 aaatgacttt ggatatctca aacttcacaa gtcaattaaa tcttgctcaa agtcaagcgc aacggctcgc
32481 actagagtct tcgaagtcct ttcaaattgg ttctgcttta acaggattag ggaaaggact tacgactgcg
32551 gttacccttc ctcttatggg atttgcagcc gcctctatta aagtagggaa tgaattccaa gctcaaatgt
32621 cccgtgttca agctattgca ggagcgacag cggaagagct tggtagaatg aagactcaag caatcgacct
32691 tggtgctaaa actgctttta gtgcaaaaga ggcggctcaa ggtatggaaa atctagcttc agccggtttc
32761 caggtaaatg aaatcatgga cgctatgcca ggggtacttg acctggctgc cgtatctgga ggagatgtgg
32831 ccgcgagctc cgaggccatg gctagttcac ttcgagcctt tggattagag gcaaaccagg cgggtcacgt
32901 ggctgacgta tttgctcgag cagcagctga tacgaacgca gaaactagcg acatggcaga ggcgatgaaa
32971 tacgtcgcac ccgttgctca ctctatgggc ttgagccttg aagaaacggc tgcgtctatt gggattatgg
33041 ccgacgccgg tattaagggc tcgcaagccg gaaccacgct tagaggcgct ctctcgcgta ttgccaaacc
33111 tacgaaagcg atggtcaaat caatgcagga attaggagtt tcgttctacg acgcgaacgg aaacatgatt
33181 ccactaagag aacaaatcgc tcaactgaaa acagctactg caggactaac acaagaggaa cgaaatcgtc
33251 accttgttac cttgtatggc caaaactcgt tgtcaggtat gcttgcacta ttagacgcag gtcctgagaa
33321 attggataag atgaccaatg ctctcgtgaa ctcggacgga gctgctaagg aaatggcaga aactatgcag
33391 gacaaccttg ctagtaaaat cgagcaaatg ggaggagctt tcgagtctgt tgctattatt gttcaacaaa
33461 tccttgagcc tgcacttgct aaaatcgtgg gagcaatcac aaaagttctc gaagcattcg taaatatgtc
33531 acctatcggt caaaagatgg ttgtcatatt cgcaggaatg gttgcagccc ttggaccact gcttctaatt
33601 gcaggaatgg tgatgacaac tattgtcaag ttaagaattg ctattcagtt tttaggtcca gcatttatgg
33671 gaacgatggg aaccattgca ggagttatag caatattcta tgctctggtc gccgtgttca tgatagccta
33741 cacaaaatcg gagagattta gaaactttat caacagtctt gcgcctgcta ttaaagctgg gtttggagga
33811 gcgttggaat ggctacttcc acgactgaaa gagttaggag aatggttaca gaaggcaggc gagaaggcga
33881 aagagttcgg tcagtctgta gggtctaaag tgtcaaaact gctcgaacag tttggaataa gtatcggtca
33951 ggcaggaggc tcgattggtc agttcattgg aaatgttctc gaaaggctag gaggcgcatt tggaaaagta
34021 ggaggagtca tttcaattgc tgtttcactt gtaacaaaat tcggtctcgc atttctaggg attacaggac
34091 cactcgggat tgctattagt ctgttagttt catttttgac agcttgggct agaacaggtg agttcaacgc
34161 agacggaatt actcaagtat tcgaaaactt gacaaacaca attcagtcga cggctgattt catctctcaa
34231 taccttccag tctttgtcga aaaaggaact caaattttag ttaagattat tgaaggaatt gcatctgctg
34301 ttcctcaagt agttgaagtg atttcacaag tcattgaaaa tattgtgatg acaatttcga cagttatgcc
34371 tcaattagtc gaagcaggaa ttaagatact cgaagcgctt ataaatggtc ttgttcaatc tcttcctact
34441 atcattcaag cagctgttca aattatcact gctttattca atggtcttgt tcaggcactt cctacgctta
34511 ttcaagcagg tcttcaaatt ttgtcagctc tcataaacgg actagttcaa gcgcttccgg caattattca
34581 agcagctgtt caaattatca tgtcgcttgt tcaagcacta attgaaaact tgcctatgat aatcgaagca
34651 gcgatgcaga ttataatggg tctagtcaac gcactgattg aaaatatagg acctatctta gaagcaggga
34721 ttcaaattct aatggcttta atcgagggac ttattcaagt gcttcctgaa ctaattacag cagcgattca
34791 aatcattact tcactattag aagcaatctt gtcgaacctt cctcaacttc tagaagccgg agttaaattg
34861 cttttatcac ttcttcaagg gttgctaaat atgcttcctc aactaattgc aggggctttg caaatcatga
34931 tggcacttct taaagcagtt atcgacttcg tccctaaact tcttcaagca ggt ttcaac ttcttaaggc
35001 attgattcaa ggtattgctt cacttctcgg ctcactttta tcgacagctg gaaacatgct ttcatcatta
35071 gttagcaaga ttgctagctt tgtgggacag atggtttcag gaggtgcgaa cctgattcga aacttcatta
35141 gtggtattgg gtcaatgatt ggttcagctg tctctaaaat tggcagcatg ggaacttcaa ttgtttctaa
35211 ggttactgga ttcgctggac aaatggtaag cgcaggggtc aaccttgttc gaggatttat caatggtatc
35281 agttccatgg taagttctgc ggtaagtgcg gcggctaata tggctagcag tgcattaaat gccgttaagg
35351 gattcttagg tattcactct ccttcacgtg tcatggagca gatgggtatc tatacgggtc aagggttcgt
35421 aaatggtatt ggtaacatga ttcgaactac acgtgacaag gctaaagaaa tggctgaaac tgttactgaa
35491 gctctcagcg acgtgaagat ggatattcaa gaaaatggag ttatagaaaa ggttaaatca gtttacgaaa
35561 agatggctga ccaacttcct gaaactcttc cagctcctga tttcgaagat gttcgtaaag cagccggttc
35631 gcctcgagtg gacttgttca atacaggaag tgacaaccct aaccaacctc agtcacaatc taaaaacaat
35701 caaggcgagc aaaccgttgt caacattgga acaatcgtag ttcgaaacaa tgacgacgtt gacaaactgt
35771 cgagaggatt gtataataga agtaaagaaa ctctatcagg gtttggtaac attgtaacac cgtaaaggag
35841 aaatagatgg ctagcagaca gacgctattg gtcgacggaa ttgaccttgt cgacaaaggt gcaaccgtgc
35911 tagaatatgt aggactcact ttcgcaggat ttaaggactc aggatttaaa aaccctgaag gcatagacgg
35981 agtattagat tctccgtcta atgctatgtc cgctcttact ggaagcgtga ccttaatgtt ccacggagaa
36051 accgaaaagc aagttaatca aaaatacagg cagttcaaac aatttattcg ctcgaagtca ttttggagaa
36121 tttcgacact tgaagaccct ggatactatc gaacgggaaa atttttagga gaaaccgagc aaggaaaacf
36191 tgtagacgtt caagccttta aagatacttc ccttgtagtt aaattaggga ttcagttcaa agatgcttac
36261 gagtacagcg actcaactgt tcgaaaggtt tataagtttc aacccgcttt gggaggcgat agcttaccta
36331 acccaggaag acctactcga caatttagag tagaaataag aactacttct caaatcaaag gatattttcg
36401 aattggcgaa aaaagttcag gacagtttgt tgagttcggt actaattcag tattgatgga aagtggctcg
36471 attattattc taaatcttgg aacttttgaa cttattaaaa ttagcagtgc aaatcaagcg actaacttat
36541 ttagatacat taaacgaggc gcattcttca agattcctaa tggaaattca acaattacca ttgaataccg
36611 agccgatgac gcagcagctt ggacctctac tcttcccgct caagttgaac tgtttctaaa tccgtcttac
_>_o
36681 tattagaaag ggaatatatg attgacaata atttacctat gagtccaatt cctggcgaaa ttgttcaagt
36751 atatgaccaa aacttcaatc taattggagc aagtgatgaa atctttagca agcattacga agacgaaatt
36821 gtgactcgag ctcgaggaaa agaaactttc acttttgaaa gtattgaaac ctcatctatc tatcaacact
36891 taaaggttga aaacattatc cagtatggag gaagatggtt tcgaattaaa tatgctcagg acgtagaaga
36961 tgtcaaaggg cttaccaagt ttacctgcta cgcattatgg tatgaactag cagaaggctt gcctaggaag
37031 ttgaaacacg ttgcttcttc tgtaggcgct gtcgcgctag atattatcaa agacgcaggt gaatgggttc
37101 gactagtttg tcctcctgac ggtgctaaca aacaagttcg aagcataaca gccgcagaaa attcaatgct
37171 ttggcatctt cgatatcttg caaagcaata caatttagaa ttgacatttg gttatgaaga aattatcaag
37241 caagaggtta gaattgttca aaccgttgta tttcttcagc cttatgtcga gtctaaagta gactttcctc
37311 ttgtagttga agagaatttg aaatatgtca ctaggcagga agattctcga aacctgtgta cggcttacaa
37381 gttgacaggt aaaaaggaag aaggcagtca agagccttta acgtttgctt ctatcaacaa tggaagtgaa
37451 tatctcattg atgtttcgtg gtttactaca cgccacatga agcctcgata tattgctaaa tctaaaagcg
37521 acgaacattt tagaattaaa gaaaatttga tgagtgctgc gcgtgcttat cttgacatct acagtcgccc
37591 actaattgga tatgaggctt cagcggtcct ttataacaag gttcctgact tgcatcatac tcaactaatt
37661 gtcgacgacc attatgatgt tatcgagtgg cgaaagatat ctgctcgaaa aattgactac gacgaccttt
37731 caaactctac tatcattttc caagaccctc gaaaagactt gatggacttg ctaaatgagg acggcgaagg
37801 agtcctttca ggggaaactg taaatgagtc ccaagttgtt attagatacg cagatgacat tttagggact
37871 aattttaatg cagaatctgg gaaatacatt ggtgtcctta atactaataa gaaaccgagc gaattagttc
37941 ctgacgactt tacatggatt cgactagaag gtcctaaagg tgacgcaggt ttaccgggag ctcctgggcg
38011 tgatggagtc gacggtgtac ctggaaagag cggagtaggg atagcagata cagctatcac ttatgctgta
38081 tccgtttccg gaacgcaaga gcctgaaaat ggatggagcg aacaagttcc tgaactcata aaaggtcgat
38151 tcttgtggac taaaacattt tggagatata ctgacggctc acatgaaact ggatactccg ttgcctatat
38221 agggcaagac ggaaattccg gaaaagacgg aatcgcaggt aaggacggag taggtatagc cgcaactgaa
38291 gtcatgtatg caagttcgcc atctgctact gaagctccag ctggtggatg gtctacgcaa gttcctaccg
38361 tcccaggtgg tcagtattta tggactcgaa caagatggcg ctacactgac caaactgatg aaattggata
38431 ttcagtttca agaatgggcg agcagggtcc taaaggtgac gcaggtcgtg acggtattgc aggaaagaac
38501 ggaatagggt tgaagtcaac ttcagtttct tatggaatta gtcccactga ttctgcgatt cctggagtat
38571 gggcttcaca agttccttct ttaatcaaag gtcaatatct ttggactcga actatttgga cctataccga
38641 ttcaactacc gaaacgggct atcaaaaaac ctacattcca aaagacggga atgacggtaa aaatggaatt
38711 gctggtaagg atggggtagg aattaagtct acgaccatta cctacgcagg ctcaacctca ggaacagttg
38781 cgcctacttc aaattggact tctgctattc caaatgttca accgggattc ttcttgtgga cgaaaactgt
38851 ttggaactat actgatgaca ctagcgaaac aggttactca gtttccaaga taggtgaaac aggtcctaga
38921 ggagttcaag gtcttcaagg tcctcaaggg cttcaaggaa ttcctggacc tgcaggagct gacggacgtt
38991 cgcaatatac tcacctcgct ttctctaata gtccaaacgg tgagggattt agtcatactg acagcggacg
39061 agcatacgtc ggtcagtatc aagatttcaa tcccgtccat tcaaaagacc ctgcagccta tacatggacg
39131 aaatggaagg ggaatgacgg agctcaaggg atacccggga agccaggcgc agacggtaag actaattatt
39201 tccatatagc ttacgcttca agtgcagacg gatcacgtga gttcagtttg gaagataata atcaacaata
39271 tatgggttat tactccgatt atgagcaagc agatagcagg gatcgaacta agtat gatg gtttgaccgc
39341 cttgccaatg ttcaagtggg aggtcgaaac gagttcctta attctttatt tgaatttggt ttaaaacctc
39411 gctattctag ttacaatcta atggacggac aagatcaaac gcaaggacag atatctgcta ctattgacga
39481 acgtcaacgg ttcaaaggtg ctaactcttt acgacttgac tcaacatgga acggtaaacc gcagaaccaa
39551 aaactgacct tttctttagg aggagatacg cgattaggta ctccaaccga gtggtctaat ttagaaggtc
39621 gtatcagttt ctgggctaag gcctctagga acggagtgag cttagctgca cggccgggtt atcgtagtaa
39691 cgtatttacc gcaaccttaa ccgatcaatg gaagttctac gattttaaat tctttgacaa agttaattca
39761 aattgtaccg ctgaagcaat tttccatgta ttcactcaaa gttgttcagt gtggctcaat catattaaaa
39831 tcgaacttgg taatatctct actcctttta gtgaagcaga ggaagacctt aaatatcgaa ttgactcaaa
39901 agccgatcaa aagctaacta accaacagtt gacggcactc acggaaaagg ctcaactaca tgacgcagaa
39971 ctgaaagcta aggctacaat ggagcagtta agtaacttag aaaaggctta tgaaggtaga atgaaagcta
40041 atgaagaagc tatcaaaaaa tcggaagccg acctaatctt agcggcaagt cgaattgaag ctactatcca
40111 agaacttggc gggctacggg aactgaagaa gttcgtcgac agttacatga gctcttctaa tgaaggtcta
40181 attatcggta agaacgacgg tagctctacc attaaggtat caagtgaccg aatttctatg ttctccgcag
40251 ggaatgaagt tatgtacctt acgcaagggt tcattcacat cgataacggg atctttaccc aatccattca
40321 agtcggccga tttagaacgg aacaatactc gtttaatcca gacatgaacg tgattcggta tgtaggataa
40391 ggagaataac atgacaaaat ttatcaactc atacggccct cttcacttga acctttacgt cgaacaagtt
40461 agtcaggacg taacgaacaa ctcctcgcga gttagttggc gagctactgt cgaccgcgat ggagcttatc
40531 gaacgtggac ttatggaaat attagtaacc tttccgtatg gttaaatggt tcaagtgttc atagcagtca
40601 cccagactac gacacgtccg gcgaagaggt aacgctcgca agtggagaag tgactgttcc tcacaatagt
40671 gacgggacaa agacaatgtc cgtttgggct tcgtttgacc ctaataacgg cgttcacgga aatatcacta
40741 tctctactaa ttacacttta gacagtattc caaggtctac acagatttct agttttgagg gaaatcgaaa
40811 tctaggatct ttacatacgg ttatctttaa ccgaaaagtg aactctttta cgcatcaagt ttggtaccga
40881 gttttcggta gcgactggat agatttaggt aagaaccata ctactagcgt atcctttacg ccgtcactgg
40951 acttagcaag gtacttacct aaatcaagtt ccggaacaat ggacatctgt attcgaacct ataacggaac
41021 tacgcaaatt ggtagtgacg tctattcaaa cggatggagg ttcaacatcc ccgattcagt acgtcctact
41091 ttttcgggca tttctttagt agacacgact tcagcggttc gacagatttt aacagggaac aacttcctcc
41161 aaatcatgtc gaacattcaa gtcaacttca acaatgcttc cggcgcttac ggatccacta tccaagcatt
41231 tcacgctgag ctcgtaggta aaaaccaagc tatcaacgaa aacggcggca aattgggtat gatgaacttt
41301 aatggctccg ctaccgtaag agcatgggtt acagacacgc gaggaaaaca atcgaacgtc caagacgtat
41371 ctatcaatgt tatagaatac tatggaccgt ctatcaattt ctccgttcaa cgtactcgtc aaaatcctgc
41441 aattatccaa gctcttcgaa atgctaaggt cgcacctata acggtaggag gtcaacagaa aaacatcatg
41511 caaattacct tctccgtggc gccgttgaac actactaatt tcacagaaga tagaggttcg gcgteaggga
41581 cgttcactac tatttcccta atgactaact cgtccgcgaa cttagctggt aactacgggc cggacaagtc
41651 ttacatagtt aaggctaaaa tccaagacag gttcacttcg actgaattta gtgctacggt agctaccgaa
41721 tcagtagttc ttaactatga caaggacggt cgacttggag ttggtaaggt tgtagaacaa gggaaggcag
41791 ggtcaattga tgcagcaggt gatatatatg ctggaggtcg acaagttcaa cagtttcagc tcactgataa
41861 taatggagca ttgaacaggg gtcaatataa cgatgtttgg aataagcgtg aaacagagtt tacatggcga
41931 agtaacaaat acgaggacaa ccctacggga actcgaggtg aatggggact atttcaaaat ttctggttag
42001 atagctggaa aatggttcaa tccttcatta caatgtcagg aagaatgttc atcaggacag cgaacgatgg
42071 aaacagctgg agacctaaca agtggaaaga ggttctattt aagcaagact tcgaacagaa taattggcag
42141 aaacttgttc ttcaaagtgg gtggaaccat cactcaacct atggcgacgc attctattcg aaaactcttg
42211 acggcatagt atatttgaga ggaaatgtgc ataaaggact tatcgacaaa gaggctacta ttgcagtact
42281 tcctgaagga tttagaccga aagtttcaat gtatcttcag gctctcaata actcatatgg aaatgccatt
42351 ctatgtatat acactgacgg aagacttgtg gtgaaatcga atgtagataa ttcttggtta aatttagaca
42421 atgtctcatt tcgtatttaa tttgagctga aatcatgtta taatattttt tagaaaggag gtgagaacta
42491 tgttgaacct tacaaaatcg cgccaaattg tggcagagtt cactattgga caaggagctg aaaagaaact
42561 tgtcaaaaca acgattgtga acattgatgc aaacgcagta tcaaccgtct ctgaaactct tcatgaccca
42631 gacttgtatg ctgcgaaccg tcgagaactt cgagctgacg agcaaaaact tcgcgaaact cgttacgcaa
42701 tcgaagatga aattctagct gaacagtcaa agactgaaac agctctaaca gctgaataag gaggcgtcaa
42771 tctatgccaa tgtggctaaa cgacacagca gtcttgacga cgattattac agcgtgcagc ggagtgctta
42841 ctgtcctact aaataagtta ttcgaatgga aatcgaataa agccaagagc gttttagagg atatctctac
42911 aactcttagc actcttaaac agcaggtcga cgggattgac caaacgacag tagcaatcaa tcaccaaaat
42981 gacgtcattc aagacggaac tagaaaaatt caacgttacc gtctttatca cgacttaaaa agggaagtga
43051 taacaggcta tacaactctc gaccatttta gagagctctc tattttattc gaaagttata agaaccttgg
43121 cggaaatggt gaagttgaag ccttgtatga aaaatacaag aaattaccaa ttagggagga agatttagat
43191 gaaactatct aacgaacaat atgacgtagc aaagaacgtg gtaaccgtag tcgttccagc agcgattgca
43261 ctaattacag gtcttggagc gttgtatcaa tttgacacta ctgctatcac aggaaccatt gcacttcttg
43331 caacttttgc aggtactgtt ctaggagttt ctagccgaaa ctaccaaaag gaacaagaag ctcaaaacaa
43401 tgaggtggaa taatgggagt cgatattgaa aaaggcgttg cgtggatgca ggcccgaaag ggtcgagtat
43471 cttatagcat ggactttcga gacggtcctg atagctatga ctgctcaagt tctatgtact atgctctccg
43541 ctcagccgga gcttcaagtg ctggatgggc agtcaatact gagtacatgc acgcatggct tattgaaaac
43611 ggttatgaac taattagtga aaatgctccg tgggatgcta aacgaggcga catcttcatc tggggacgca
43681 aaggtgctag cgcaggcgct ggaggtcata cagggatgtt cattgacagt gataacatca ttcactgcaa
43751 ctacgcctac gacggaattt ccgtcaacga ccacgatgag cgttggtact atgcaggtca accttactac
43821 tacgtctatc gcttgactaa cgcaaatgct caaccggctg agaagaaact tggctggcag aaagatgcta
43891 ctggtttctg gtacgctcga gcaaacggaa cttatccaaa agatgagttc gagtatatcg aagaaaacaa
43961 gtcttggttc tactttgacg accaaggcta catgctcgct gagaaatggt tgaaacatac tgatggaaat
44031 tggtattggt tcgaccgtga cggatacatg gctacgtcat ggaaacggat tggcgagtca tggtactact
44101 tcaatcgcga tggttcaatg gtaaccggtt ggattaagta ttacgataat tggtattatt gtgatgctac
44171 caacggcgac atgaaatcga atgcgtttat ccg tataac gacggctggt atctactatt accggacgga
44241 cgtctggcag ataaacctca attcaccgta gagccggacg ggctcattac tgctaaagtt taaaatatag
44311 agaggaggaa gctcttttct taatattgtt tctcttaatc ccgcaaggtt tcgaccctgc ggggttttgt
44381 gtcgtatatt actctattta cttattcgaa gatttcaatt ataattaaat agtcaacatg attcatgatt
44451 gttgatatga ccctttccgc cctacataat ttgtggggcg tttatttttt ataaaaattt tttacaaaat
44521 gcttgacaac attcactcat tatcgtataa tacaattata aaaataaata aagccgaaag gcgaggagga
44591 cattatgtca aaaattaaat tcgaaaacct taaaaaaggc gatgttgtgc tacgagctaa atctcaaacg
44661 aagtttaaaa tcgtttcaat tttagcagac gaaaagaaag cagaccttga atcattagaa gacggaggtg
44731 aacttcacct ttcagcttca actctcgaac gttggtacac aatggaagat gaaactgaac ctaaaaaaga
44801 agaagctgct aaacctgcta aaaaggctgc tcctgcagtt gctcgacctg ctcgaaaagg tagagtcgtt
44871 cccaaaccta aaaaagaagt ccttgaggaa gaaattcctg aagttaagga acagccggaa gaagttggtt
44941 cagttagtga gaaatctact gttcgaaaac ctgctcctaa aaaagaaagc gtgatggcga ttactaaggc
45011 tcttgaaagt cgaattgttg aagcctttcc tgcgtctact cgaatcgtca ctcagtctta catcgcctat
45081 cgctctaaga agaacttcgt tactatcgaa gaaactcgaa aaggtgtttc tattggagtt cgcgcaaaag
45151 ggttgacaga agaccaaaag aaacttcttg catctattgc tcctgcatct tacgaatggg cgattgacgg
45221 aatttttaaa ctcgtcaagg aagaagatat tgacaccgca atggaattga ttgaagcttc tcacctttct
45291 tcgctatgat tgaaatcgtt atagcacgtt cgaaagctag gcgaggtcga accctattta ttgaaacatg
45361 ggcaagcact gatgaagatg cagttaaaat ggcagaaaag atttccagct tgcccaatgt agtcgagacg
45431 tcttctaata acttcgaact accttataag tatttcaata atgttataga cgctctagat gaatgggagc
45501 ttcacatctt cggcgaactt gataaagatg ttcaagacta cattgactct cgaaaccgaa tagcttcttc
45571 aagcaatgag cagttttcgt tcaagactac tccattcgcg caccaggttg aatgtttcga atacgcacaa
45641 gagcatccat gtttcctttt aggcgatgag caaggtttag ggaaaactaa acaggcaatt gatattgcag
45711 ttagcaggaa ggcaagtttc aaacattgtt taatcgtatg ttgcatatca gggctcaaat ggaattgggc
45781 aaaagaagta ggtattcatt caaatgagtc agctcatatt ttaggaagtc gagtcactaa agatgggaaa
45851 ttagtgattg acggagtttc taaacgggca gaagacttgc ttggtggcca cgacgaattc ttccttatca
45921 ctaacattga aactcttcgc gatgctgtgt tcattaaata cttaaatgaa ctgacaaaaa gcggagaaat
45991 tggaatggtt attattgacg agattcacaa gtgtaagaac ccttcaagta agcaaggggc ttcaattcaa
46061 aagctccaaa gttattacaa gatgggactt acaggaactc ctctaatgaa taacccaatc gatgtattca
46131 atgttatgaa gtggctaggg gcggaacatc atacactgac tcagttcaaa gagcgatact gtatcgtcga
46201 ccagttcaat caaatcactg gatatcgaaa tctagctgaa cttcgcgagc ttgtcaacga ctacatgctt
46271 agaagaacga aggaagaagt tttagacctg cctgaaaaga ttcgagtcac agagtatgtc gacatgaact
46341 cgaaacagtc aaaaatctat aaggaagttt tgactaaact tgttcaagaa atagataaag tcaagctcat
46411 gcctaaccct ctagccgaaa cgattcgact tcgacaagcg actggaaatc cttcgatttt aactactcaa
46481 gatgtcaagt cttgcaagtt cgaaagatgt atcgaaattg tcgaggaatg tatccagcaa ggaaagtcct
46551 gcgtgatatt tagcaattgg gaaaaggtta ttgaacctct tgctaagata ctttcgaaga cagtcaaatg
46621 caacctggta acaggagaaa ccgcagataa gttcaacgaa attgaagaat ttatgaatca cagaaaggct
46691 tctgttattt taggaactat aggtgcgcta ggaacaggat ttactttgac gaaagcggat acggttattt
46761 tcttagatag tccgtggaca cgcgcagaaa aggaccaagc cgaagatagg tgtcatagaa tεggcgcaaa
46831 aagttctgtc actatctaca cgcttgtcgc caaaggtact gttgacgaac gtatagaaga ccttattgaa
46901 cggaaaggag aattagcaga ttatatcgta gatggtaagc ctatgaaatc taaaattggt aaccttttcg
46971 atatcctgct taaatagaat gaaaactatc tccatattaa ggaaagacac taaaaggaag ccggacagga
47041 acggaagaaa aactgcactc gaactagctc aagagattga tatgtcacct agtgagttag cagagctcct
47111 tcaaattcct gaaaggacgg caaccagaat tttaaaactc gacaaactgc tcaacaaaga gcaatgctca
47181 ataatagaaa ggtatataaa tgaaattcac tgaaggaaaa aattggtata aagttggaga gatatgtcaa
47251 atgttgaacc gctctctatc tacgattaat gtttggtatg aagcaaaaga cttcgctgaa gaaaataaca
47321 ttcacttccc gtttgttctt cctgaaccta gaacagacct tgaccatcgt ggttctcgat tctgggatga
47391 cgaaggcgtg aacaaactca aacgatttag ggacaaccta atgcgcggtg acttggcatt ctacactcga
47461 actcttgtag ggaaaactga aagggaagca attcaagaag atgctaaagc atttaaacgt gaacatggat
47531 tggagaatta aatgaaattt gaagatgaaa aacagttcat cgctgcaatt gaagaagccg gtgaattaaa
47601 tgctaccaaa ggcgacatgg agaaacaagt caaaagtctt cgtgatgctc taaaagagta catgaaagaa
47671 aatgacattg aatctgctca aggtaagcac ttttctgcta ccttctacac gacagagcgc tcaactatgg
47741 acgaagaacg cttgaaagaa attatcgaaa aattagttga cgaagccgag acggaagaaa tgtgtgaaaa
47811 actttcaggg cttatcgaat acaagcctgt catcaatacg aaacttctcg aggatatgat ttatcacggc
47881 gagattgacc aagaagcaat tcttccagca gttgtcattt ctgttacaga aggcattcgt tttggaaagg
47951 ctaaaattta gcgatatttt tggttctgcg acgtttttag ggttagcaga atccaatcac accacttgcg
48021 caggcaaccg ctgtctgcgt taattttaga aggttaatat tataccataa ggaggagata agtggcaagg
48091 caaagaatag gcaattcagg aaagcctaaa aatgaaattg aactaacatt caaagacaag cctaaaactc
48161 gttctacctt attcaagaag gacgtggcaa caggtctttc aaaagtcgag catgattatt ttcaaatagt
48231 tgaagcactt aacggaaaac aattcgaacc taatatgaag caggtgtcat ctttctttat agttcagtat
48301 gaatttattt tcaatattaa gtgcatcgat tataactggt tcaacttttc gagcactatg aaaaatgttc
48371 gaacttattt aaacattgag tcgaacattg aactttgtcg atttttagct gaaagttttg ttaaatatga
48441 aaatgttcga aaaagattga acctaagcga aaggttcata acggtctcga ctttcaaaag agcctggatt
48511 ttggacgaac tcgaaggaaa aacgggttca aaattcgaag gattttatta gtttagtaga ctatttttag
48581 attttttaaa atgtggttta caaaatgacc tcaataggcg tataatttat caatcttgat tctttcgggc
48651 cggtatatat acaccaataa tcgagaaata ataaattata gtatcgaaaa tataaaaagg agaaaagttg
48721 gaaaatttag ctgatagaat atggaagaaa aagttaaatg accttttcga gagaagtggg ctacctcaaa
48791 agtatttcga acctcaagtg ttagtcgaac gaaaagccga caaggaatgt tgggaatggc tagaagctgt
48861 tcgagcaaat atagtcgaag aagttcgaaa cggtcttagc attgttattg cttcgaatac tgtcgggaat
48931 gggaaaacta gctgggcggt tcgacttttg caacgctatt tagcagaaac tgcacttgac ggaagaattg
49001 ttgagaaagg aatgtttgta gtgtcagctc aactattgac tgagttcggc gactataatt attttcaaac
49071 catgcaagaa tttctcgaac gtttcgagcg ccttaagact tgtgagctat tagtcataga cgaaataggt
49141 ggaggttcct taaccaaggc ctcttatcct tatctgtatg acttggttaa ttatagggtt gacaataact
49211 tgtcgactat ttatacgact aattatactg acgatgaaat tattgacctt ttaggccaaa ggctttatag
49281 tcgtatatat gatacttcag tggttctaga ttttcaggca agcaatgtaa gaggattgga ggtaagcgaa
49351 attgaatcat agatatagta acatcacaac tatttttctt tggcagattg tctttctttg tatttgctgc
49421 gcggtgtcct attgtgcagg agtgcataat gagcgagagt ctcaagataa ggtgattcaa agttataagc
49491 agaaagaaaa gtcagccgtc tacttgacag tcgatagttc aggagcttgg ctaggaagtg ctccgggagc
49561 caaggaaagt cctctctaca atgaaaaggg acagcatgta ggaaaattga aagaggtggg agagtgatac
49631 agcttcaagt cttaaataaa gttctcgaag aaaagagctt atccatttta gaaaataatg gaattgacca
49701 agaatacttc acggattatt tagacgagta tcaatttatt caagaacact tttcgagata tggaagagtt
49771 ccggacgacg aaactattct cgaccatttt cctggattcg aatttttcga aattggcgaa actgatgaat
49841 accttatcga caagctaaaa gaggagcatc tatataattc acttgttcca attttaacgg aagcggctga
49911 ggacattcaa gtagatagta acattgcgat tgcgaatata attccaaaac tagaagaact tttcaatcgc
49981 tctaaattcg taggcggact agacattgct cgaaatgcta aacttcgact agactgggcg aatactatta
50051 gaaaccatga cggtgaaaga cttggaatat cgacagggtt tgaactattg gacgacgtgc ttggaggctt
50121 acttcctggt gaggatttga ttgtcataat ggctcgacct ggacaaggta agtcgtggac tattgataaa
50191 atgcttgcaa ctgcttggaa gaacgggcat gatgtccttc tatatagcgg ggaaatgagt gaaatgcaag
50261 ttggtgctcg tatagatact attctttcga atgttagcat caattcaatt accaaaggga tttggaacga
50331 ccatcagttc gaaaaatatg aggaccatat tcaagcaatg actgaggctg aaaattccct tgtggtagtc
50401 acgcccttta tgattggagg aaagaacctt acccctgcaa ttttagatag catgatatct aaatatagac
50471 catctgtggt ggggattgac cagctttcac tcatgagcga gtcttatcca agcagggagc agaagcgaat
50541 ccagtacgcc aacatcacca tggacctata taagatttct gctaaatatg gaattcctat tgtgcttaat
50611 gtccaagcag ggcgttcggc taaaactgaa ggcgctgaaa gtatggaact agaacatata gcagaaagtg
50681 atggagtagg tcaaaatgct agcagagtta tcgctatgaa gcgtgacgaa aaatccggca tacttgaact
50751 atctgtcgtt aaaaaccgat atggcgaaga ccgaaaaatc atcgaatata tgtgggacgt tgaaactgga
50821 acctatactc ttataggatt caaagaggaa ggcgaagaag gaactgaaaa aggcgaaagc tctccattga
50891 aagcaaaagc ctctaggtcg actgctcgtc ttcgaagtaa ggttacaagg gaaggagttg aagcattttg
50961 atgaaagtaa atggtcttca aattgaagcg actcctgaac aaataattga aaaactttcg agacaacttg
51031 aagacgaagg aacattcatt tttagacgaa ctaagtcgct tggaagcaac tatcaattct catgcccgtt
51101 tcatgcagga gggactgaaa agcatccctc ttgtggcatg agtagaaatc cttcttattc aggaagtaag
51171 gtgacggaag ctggaacggt tcactgtttc acttgcggct acacttcagg actaactgaa ttcgtctcga
51241 atgtattagg tcgaaacgat ggagggttct atggaaacca gtggctgaaa aggaattttg gaacatctag
51311 cgaagtagtt aggcaaggcg tcagccctga agcgtttcga agaaatggga gaactgaaaa agtcgagcat
51381 aaaatcattc ctgaagagga acttgataaa taccggttta ttcatcctta tatgtatgaa cggaaattga
51451 cggacgagct catcgagatg tttgatgtag gttatgacaa actgcatgat tgcatcacct ttccagtacg
51521 gaacctcaag ggcgaaacag tattcttcaa ccgtcgaagt gttcgttcta agtttcacca gtacggtgaa
51591 gatgacccta aaacggaatt tctttatggc caatatgagc ttgtagcatt tcgagactat tttgaaaaac
51661 ctattagtca agtattcgtg actgagtctg ttatcaactg cttgactctt tggtcaatga agattccagc
51731 agtcgctctt atgggagtag gtggaggaaa tcaaatcaat ttactaaaac gacttcctta tagaaatatt
51801 gttctagcac ttgaccctga taacgctggg cagacagcgc aggaaaaact ctaccgacag ttaaagcgaa
51871 gcaaggtcgt tagatttttg aactacccta aagagttcta tgataataag tgggatataa acgaccatcc
51941 ggaattatta aattttaatg atttagtctt gtagaaattc atttattatc gtataataaa gttagaaaat
52011 tttaaaaaga ggtcatatca atatgaaaga agcgaataga ctagtttcta gctatgtagg attcgaatgc
52081 tggactgacg aagaatgtat caggaacttt gaactagacc ctgatatgtc aattgcgtct gcttatcafcs
52151 gttattttgg gatgctttat tcctatgcaa aaaggtttaa atgcttatct cgacatgaca ttgaaagcat
52221 tgcattcgag actatttcaa aatgtttggc aacgttcaaa tcaaaccaag gggccaagtt ttcaacttac
52291 cttacaagac tcttcaagaa tagaatagtc ttagaatata ggtacctaaa tgcaccttcc atgaatcgaa
52361 attggtatgt agaagtgacg ttcgatagcg tttcgacaaa tgaagaaggc gacgatttta gtatcctatc
52431 gacagttggc tattgtgaag actacggaaa aattgaaatt gaagcaagtc ttgacttcat gacgctttct
52501 aatacagagt atgcttatat ctcgtctgtc attcaaaacg gtccttcagt aagcgacgca gaaattgcgc
52571 gtgaaattgg agtaagcagg tctgctatta gtcagtctaa gaagtcacta aaaaataaat taaaagattt
?58
52641 tatataactg gtttacaaat cacgtgaatt tcgtgtatat tatatatgaa aggacaaact ttgaaacctt
52711 aaaaacttca aaaatctttc aaccattaaa aacttataaa ggagaatcga tatgggaaaa gtatcaattc
52781 aaaaatcagg aacatttagc tcagggtcta ataacgagtt tttcacactc gctgaccacg gtgacagcgc
52851 aattgtcact ctattgtatg atgacccgga aggcgaagac atggattatt tcgtagtcca cgaagcagac
52921 gttgacggtc gtcgacgcta tatcaattgc aatgctattg gcgaagacgg ggaaacagtc catcctgata
52991 attgtccatt atgccaaaac ggattccctc gtattgaaaa actatttctt caactttaca accatgatac
53061 gggaaaagtt gaaacatggg accgaggccg ttcttatgtt caaaagattg ttacatttat caataaatat
53131 ggaagccttg tgactcagcc ttttgaaatt attcgttcag gagctaaagg tgaccaacga actacttatg
53201 aattccttcc agagcgtccg gaagacagtg ctactcttga agattttcca gaaaagagcg aacttcttgg
53271 aactctaatt ttagacctcg acgaagacca aatgtttgac gtggttgacg gcaagttcac tcttcaagaa
53341 gagcgttctt caagtcgttc aaattcacgt agaggagcat ctcctgcgcc tagacgaggt tccggtcgag
53411 aatcttcaca aggtcgaaca gctgaaagaa ctccttcagt tagtcgaaga actcctccaa cacgaggtcg
53481 aggattctaa catgagggcg cgagccctct ttattattga ttaagaaagg gaaaataatg gcacaaaaag
53551 gactctttgg tgcaaagcct cgttctagca agaagaacga tgctcagtta cttgctcaac ggaaaaacag
53621 gaagcctgca gttgaggtta cttacatttc aggaaacgct ctaaaggacg cagttgctag agctcgtact
53691 ctttcaacta ggattcttgg acacgttctt gatagacttg agttaatcac tgaggaagca aaactcgagc
53761 agtatgtaga caaaatgatt gaagacggaa taggttctat tgacgtagaa actgatggac tcgatactat
53831 tcacgatgag ctggcaggag tctgcttgta ctcacctagt caaaaaggaa tctatgctcc tgtcaatcat
53901 gttagcaata tgacgaagat gcgaattaag aatcaaattt ctcctgagtt catgaagaaa atgcttcaac
53971 ggattgtaga ttcaggaatt cctgtcatct atcataattc gaaatttgac atgaaatcga tttattggcg
54041 actcggcgtc aaaatgaatg agccagcgtg ggatacatat ttagccgcaa tgcttttaaa tgaaaacgag
54111 tctcacagct tgaaaagtct tcactctaaa tatgttagga acgaagaaaa cgcagaggtt gcaaaattta
54181 atgacttatt taaaggaatt ccttttagtt taattcctcc tgatgttgcc tatatgtatg cggcctatga
54251 ccctttgcaa actttcgaac tctatgaatt tcaagaacaa tacttgactc caggaactga acaatgtgaa
54321 gaatataacc tggaaaaagt ctcatgggtt cttcataata ttgagatgcc tctaattaaa gttctcttcg
54391 acatggaagt ctacggtgtc gacttagacc aagataagct ggcagaaatt agagaacagt ttactgccaa
54461 tatgaacgag gctgagcaag agtttcaaca gcttgtcagc gaatggcagc ctgaaattga agaacttcga
54531 caaactaatt tccagagcta tcaaaaactc gaaatggatg caagaggtcg agtgacgg a agcatttcca
54601 gtcctactca attagcaatt ctgttttatg atatcatggg attgaaaagt cctgaaaggg ataaacctag
54671 aggaacaggc gaaagtattg tcgagcattt tgataacgat atctcaaaag cacttttgaa atatagaaaa
54741 tatgcaaaat tagtttcgac ctatacaaca cttgaccaac accttgcaaa gcctgacaat cgaattcaca
54811 ctacattcaa acagtacgga gctaagacag ggcgtatgtc aagtgagaat cctaacttac agaatattcc
54881 ttctcgcggt gagggtgcag tagttcgaca aatctttgca gccagtgaag ggcattacat tattggtagt
54951 gactactctc aacaagaacc tcgttcattg gcggaattaa gtggcgacga aagtatgcga catgcttacg
55021 aacaaaacct ggacctatat tcagttatcg gttcgaaact ttatggtgtt ccctatgaag agtgtttaga
55091 gttctatccc gacggaacga ctaacaagga aggaaaactt cgaagaaatt ctgtcaagtc cgttctttta
55161 ggtcttatgt acggccgcgg ggctaactca atcgctgagc agatgaatgt atctgtcaaa gaagcgaata
55231 aggttattga agatttcttc accgagttcc ctaaagtggc agactatatc atattcgttc aacagcaggc
55301 gcaggacttg ggatatgttc aaacagctac cggtcgaaga agaaggcttc ctgatatgag tcttcctgaa
55371 tacgagttcg agtatatcga cgctagcaag aacgaagatt tcgacccctt taactttgac gcagaccaac
55441 agatggacga tactgttcct gaacatatta tcgaaaaata ttgggcccag ctagatagag cctggggatt
55511 taagaagaag caagaaatta aagaccaggc aaaagccgaa ggaattctta ttaaggataa cggaggcaag
55581 atagctgatg ctcagcgcca atgtttgaac tcagttattc aaggaacggc agccgacatg actaagtacg
55651 caatgattaa ggtacacaat gacgctgaat tgaaagaatt aggattccat ttaatgattc cagttcacga
55721 tgagttacta ggtgaggttc ctatcaagaa cgcaaaacgg ggagcagaaa ggttgacaga agttatgatt
55791 gaagcagcca aggacattat tagtcttcca atgaaatgtg accccagtat agtagaaaga tggtatggtg
55861 aagaaattga aatctaaaat ctattcagtt gcatatataa ttctagtagt tattgcgaac cttgtgacaa
55931 tttatttcga acctttaaat gtgaaaggaa ttttaattcc tccaagcagt tggtttatgg gattcacttt
56001 cctgcttata aatctaataa gcaagtacga gaagccaaaa tttgcaggtt ctttgatatg ggtagggtta
56071 ttccttacct cgttgatttg ctttatgcaa aacctaccac aatcgcttgt cgtggcttca ggagttgcat
56141 tttggataag tcaaaaagca agtgtcttta tattcgacaa gctctcgaat aaattagact cgaagattgc
56211 aaatgctttg tctagcaaca tcggttctat tatagacgca accatatgga tttcattagg actgagtcct
56281 cttggaattg gaacggttgc atatatagat attccgtcag ccgtactagg ccaagttcta gttcagttta
56351 tcttgcagtc aattgcttcg agatatttga aaaagtagtc aggaaaattc ctgattatct tgcagtcaat
56421 tgcttcgaga tatttgaaaa agtagt agg aaaattcctg attatttttt ttacaaaaac gcttgacttt
56491 attcattcat tattat
Table 29
Phage dpi ORFs list
Predicted Dp-1 amino acid sequences dplORFOOl
36698 atgattgacaataatttacctatgagtccaattcctggcgaaattgttcaagtatatgaccaaaacttcaatctaattggagca
1 M I D N N L P M S P I P G E I V Q V Y D Q N F N L I G A
36782 agtgatgaaatctttagcaagcattacgaagacgaaattgtgactcgagctcgaggaaaagaaactttcacttttgaaagtatt
29 S D E I F S K H Y E D E I V T R A R G K E T F T F E S I
36866 gaaacctcatctatctatcaacacttaaaggttgaaaacattatccagtatggaggaagatggtttcgaattaaatatgctcag
57 E T S S I Y Q H L K V E N I I Q Y G G R W F R I K Y A Q
36950 gacgtagaagatgtcaaagggcttaccaagtttacctgctacgcattatggtatgaactagcagaaggcttgcctaggaagttg
85 D V E D V K G L T K F T C Y A L W Y E L A E G P R K L
37034 aaacacgttgcttcttctgtaggcgctgtcgcgctagatattatcaaagacgcaggtgaatgggttcgactagtttgtcctcct
113 K H V A S S V G A V A L D I I K D A G E W V R V C P P
37118 gacggtgctaacaaacaagttcgaagcataacagccgcagaaaattcaatgctttggcatcttcgatatcttgcaaagcaatac
141 D G A N K Q V R S I T A A E N S M L W H L R Y A K Q Y
37202 aatttagaattgacatttggttatgaagaaattatcaagcaagaggttagaattgttcaaaccgttgtatttcttcagccttat
169 N L E L T F G Y E E I I K Q E V R I V Q T V V F L Q P Y
37286 gtcgagtctaaagtagactttcctcttgtagttgaagagaatttgaaatatgtcactaggcaggaagattctcgaaacctgtgt
197 V E S K V D F P V V E E N L K Y V T R Q E D S R N C
37370 acggcttacaagttgacaggtaaaaaggaagaaggcagtcaagagcctttaacgtttgcttctat aacaatggaagtgaatat
225 T A Y K L T G K K E E G S Q E P L T F A S I N N G S E Y
37454 ctcattgatgtttcgtggtttactacacgccacatgaagcctcgatatattgctaaatctaaaagcgacgaacattttagaatt
253 L I D V S W F T T R H M K P R Y I A K S K S D E H F R I
37538 aaagaaaatttgatgagtgctgcgcgtgcttatcttgacatctacagtcgcccactaattggatatgaggcttcagcggtcctt
281 K E N M S A A R A Y D I Y S R P L I G Y E A S A V L
37622 tataacaaggttcctgacttgcatcatactcaactaattgtcgacgaccattatgatgttatcgagtggcgaaagatatctgct
309 Y N K V' P D L H H T Q L I V D D H Y D V I E R K I S A
37706 cgaaaaattgactacgacgacctttcaaactctactatcattttccaagaccctcgaaaagacttgatggacttgctaaatgag
337 R K I D Y D D L S N S T I I F Q D P R K D M D L L N E
37790 gacggcgaaggagtcctttcaggggaaactgtaaatgagtcccaagttgttattagatacgcagatgacattttagggactaat
365 D G E G V S G E T V N E S Q V V I R Y A D D I L G T N
37874 tttaatgcagaatctgggaaatacattggtgtccttaatactaataagaaaccgagcgaattagttcctgacgactttacatgg
393 F N A E S G K Y I G V N T N K K P S E L V P D D F T
37958 attcgactagaaggtcctaaaggtgacgcaggtttaccgggagctcctgggcgtgatggagtcgacggtgtacctggaaagagc
421 I R E G P K G D A G P G A P G R D G V D G V P G K S
38042 ggagtagggatagcagatacagctatcacttatgctgtatccgtttccggaacgcaagagcctgaaaatggatggagcgaacaa
449 G V G I A D T A I T Y A V S V S G T Q E P E N G W S E Q
38126 gttcctgaactcataaaaggtcgattcttgtggactaaaacattttggagatatactgacggctcacatgaaactggatactcc
477 V P E L I K G R F L W T K T F W R Y T D G S H E T G Y S
38210 gttgcctatatagggcaagacggaaattccggaaaagacggaatcgcaggtaaggacggagtaggtatagccgcaactgaagtc
505 V A Y I G Q D G N S G K D G I A G K D G V G I A A T E V
38294 atgtatgcaagttcgccatctgctactgaagctccagctggtggatggtctacgcaagttcctaccgtcccaggtggtcagtat
533 M Y A S S P S A T E A P A G G W S T Q V P T V P G G Q Y
38378 ttatggactcgaacaagatggcgctacactgaccaaactgatgaaattggatattcagtttcaagaatgggcgagcagggtcct
561 L W T R T R W R Y T D Q T D E I G Y S V S R M G E Q G P
38462 aaaggtgacgcaggtcgtgacggtattgcaggaaagaacggaatagggttgaagtcaacttcagtttcttatggaattagtccc
589 K G D A G R D G I A G K N G I G L K S T S V S Y G I S P
38546 actgattctgcgattcctggagtatgggcttcacaagttccttctttaatcaaaggtcaatatctttggactcgaactatttgg
617 T D S A I P G V A S Q V P S L I K G Q Y W T R T I
38630 acctataccgattcaactaccgaaacgggctatcaaaaaacctacattccaaaagacgggaatgacggtaaaaatggaattgct
645 T Y T D S T T E T G Y Q K T Y I P K D G N D G K N G I A
38714 ggtaaggatggggtaggaattaagtctacgaccattacctacgcaggctcaacctcaggaacagttgcgcctacttcaaattgg
673 G K D G V G I K S T T I T Y A G S T S G T V A P T S N W
38798 acttctgctattccaaatgttcaaccgggattcttcttgtggacgaaaactgtttggaactatactgatgacactagcgaaaca
701 T S A I P N V Q P G F F L W T K T V W N Y T D D T S E T
38882 ggttactcagtttccaagataggtgaaacaggtcctagaggagttcaaggtcttcaaggtcctcaagggcttcaaggaattcct
729 G Y S V S K I G E T G P R G V Q G L Q G P Q G L Q G I P
38966 ggacctgcaggagctgacggacgttcgcaatatactcacctcgctttctctaatagtccaaacggtgagggatttagtcatact
757 G P A G A D G R S Q Y T H L A F S N S P N G E G F S H T
39050 gacagcggacgagcatacgtcggtcagtatcaagatttcaatcccgtccattcaaaagaccctgcagcctatacatggacgaaa
785 D S G R A Y V G Q Y Q D F N P V H S K D P A A Y T W T K
39134 tggaaggggaatgacggagctcaagggatacccgggaagccaggcgcagacggtaagactaattatttccatatagcttacgct
813 W K G N D G A Q G I P G K P G A D G K T N Y F H I A Y A
39218 tcaagtgcagacggatcacgtgagttcagtttggaagataataatcaacaatatatgggttattactccgattatgagcaatjca
841 S S A D G S R E F S L E D N N Q Q Y M G Y Y S D Y TT Q A
39302 gatagcagggatcgaactaagtatcgatggtttgaccgccttgccaatgttcaagtgggaggtcgaaacgagttccttaattct
869 D S R D R T K Y R W F D R L A N V Q V G G R N E F L N S
39386 ttatttgaatttggtttaaaacctcgctattctagttacaatctaatggacggacaagatcaaacgcaaggacagatatctgct
897 L F E F G L K P R Y S S Y N M D G Q D Q T Q G Q I S A
39470 actattgacgaacgtcaacggttcaaaggtgctaactctttacgacttgactcaacatggaacggtaaaccgcagaaccaaaaa
925 T I D E R Q R F K G A N S L R L D S T W N G K P Q N Q K
39554 ctgaccttttctttaggaggagatacgcgattaggtactccaaccgagtggtctaatttagaaggtcgtatcagtttctgggct
953 T F S XJ G G D T R L G T P T E S N L E G R I Ξ F A
39638 aaggcctctaggaacggagtgagcttagctgcacggccgggttatcgtagtaacgtatttaccgcaaccttaaccgatcaatgg
981 K A S R N G V S A A R P G Y R S N V F T A T L T D Q W
39722 aagttctacgattttaaattctttgacaaagttaattcaaattgtaccgctgaagcaattttccatgtattcactcaaagttgt
1009 K F Y D F K F F D K V N S N C T A E A I F H V F T Q S C
39806 tcagtgtggctcaatcatattaaaatcgaacttggtaatatctctactccttttagtgaagcagaggaagaccttaaatatcga
1037 S V W L N H I K I E L G N I S T P F S E A E E D K Y R
39890 attgactcaaaagccgatcaaaagctaactaaccaacagttgacggcactcacggaaaaggctcaactacatgacgcagaactg
1065 I D S K A D Q K L T N Q Q L T A T E K A Q L H D A E L
39974 aaagctaaggctacaatggagcagttaagtaacttagaaaaggcttatgaaggtagaatgaaagctaatgaagaagctatcaaa 1093 K A K A T M E Q S N L E K A Y E G R M K A N E E A I K
40058 aaatcggaagccgacctaatcttagcggcaagtcgaattgaagctactatccaagaacttggcgggctacgggaactgaagaag 1121 K S E A D L I L A A S R I E A T I Q E L G G L R E L K K 40142 ttcgtcgacagttacatgagctcttctaatgaaggtctaattatcggtaagaacgacggtagctctaccattaaggtatcaagt 1149 F V D S Y M S S S N E G L I I G K N D G S S T I K V S S 40226 gaccgaatttctatgttctccgcagggaatgaagttatgtaccttacgcaagggttcattcacatcgataacgggatctttacc 1177 D R I S M F S A G N E V M Y T Q G F I H I D N G I F T 40310 caatccattcaagtcggccgatttagaacggaacaatactcgtttaatccagacatgaacgtgattcggtatgtaggataa 40390
1205 Q S I Q V G R F R T E Q Y S F N P D M N V I R Y V G * dplORF002
32386 atggattttgggtcaattgcagcaaaaatgactttggatatctcaaacttcacaagtcaattaaatcttgctcaaagtcaagcg
1 D F G S I A A K M T L D I S N F T S Q L N L A Q S Q A
32470 caacggctcgcactagagtcttcgaagtcctttcaaattggttctgctttaacaggattagggaaaggacttacgactgcggtt
29 Q R A E S S K S F Q I G S A L T G G K G T T A V
32554 acccttcctcttatgggatttgcagccgcctctattaaagtagggaatgaattccaagctcaaatgtcccgtgttcaagctatt
57 T L P M G F A A A S I K V G N E F Q A Q M S R V Q A I
32638 gcaggagcgacagcggaagagcttggtagaatgaagactcaagcaatcgaccttggtgctaaaactgcttttagtgcaaaagag
85 A G A T A E E L G R K T Q A I D L G A K T A F S A K E
32722 gcggctcaaggtatggaaaatctagcttcagccggtttccaggtaaatgaaatcatggacgctatgccaggggtacttgacctg
113 A A Q G M E N A S A G F Q V N E I M D A M P G V D L
32806 gctgccgtatctggaggagatgtggccgcgagctccgaggccatggctagttcacttcgagcctttggattagaggcaaaccag
141 A A V S G G D V A A S S E A A S S L R A F G E A N Q
32890 gcgggtcacgtggctgacgtatttgctcgagcagcagctgatacgaacgcagaaactagcgacatggcagaggcgatgaaatac
169 A G H V A D V F A R A A A D T N A E T S D M A E A M K Y
32974 gtcgcacccgttgctcactctatgggcttgagccttgaagaaacggctgcgtctattgggattatggccgacgccggtattaag
197 V A P V A H S M G L S L E E T A A S I G I M A D A G I K
33058 ggctcgcaagccggaaccacgcttagaggcgctctctcgcgtattgccaaacctacgaaagcgatggtcaaatcaatgcaggaa
225 G S Q A G T T L R G A S R I A K P T K A M V K S M Q E
33142 ttaggagtttcgttctacgacgcgaacggaaacatgattccactaagagaacaaatcgctcaactgaaaacagctactgcagga
253 L G V S F Y D A N G N M I P L R E Q I A Q L K T A T A G
33226 ctaacacaagaggaacgaaatcgtcaccttgttaccttgtatggccaaaactcgttgtcaggtatgcttgcactattagacgca
281 T Q E E R N R H L V T L Y G Q N S L S G M L A L D A
33310 ggtcctgagaaattggataagatgaccaatgctctcgtgaactcggacggagctgctaaggaaatggcagaaactatgcaggac
309 G P E K L D K M T N A L V N S D G A A K E M A E T M Q D
33394 aaccttgctagtaaaatcgagcaaatgggaggagctttcgagtctgttgctattattgttcaacaaatccttgagcctgcactt
337 N L A S K I E Q M G G A F E S V A I I V Q Q I L E P A L
33478 gctaaaatcgtgggagcaatcacaaaagttctcgaagcattcgtaaatatgtcacc atcggtcaaaagatggttgtcatattc
365 A K I V G A I T K V -i E A F V N M S P I G Q K M V V I F
33562 gcaggaatggttgcagcccttggaccactgcttctaattgcaggaatggtgatgacaactattgtcaagttaagaattgctatt
393 A G M V A A L G P L L L I A G M V M T T I V K R I A I
33646 cagtttttaggtccagcatttatgggaacgatgggaaccattgcaggagttatagcaatattctatgctctggtcgccgtgttc
421 Q F L G P A F M G T M G T I A G V I A I F Y A V A V F
33730 atgatagcctacacaaaatcggagagatttagaaactttatcaacagtcttgcgcctgctattaaagctgggtttggaggagcg
449 M I A Y T K S E R F R N F I N S L A P A I K A G F G G A
33814 ttggaatggctacttccacgactgaaagagttaggagaatggttacagaaggcaggcgagaaggcgaaagagttcggtcagtct
477 E W L P R K E L G E W Q K A G E K A K E F G Q S
33898 gtagggtctaaagtgtcaaaactgctcgaacagtttggaataagtatcggtcaggcaggaggctcgattggtcagttcattgga
505 V G S K V Ξ K E Q F G I S I G Q A G G Ξ I G Q F I G
33982 aatgttctcgaaaggctaggaggcgcatttggaaaagtaggaggagtcatttcaattgctgtttcacttgtaacaaaattcggt
533 N V E R G G A F G K V G G V I S I A V S V T K F G
34066 ctcgcatttctagggattacaggaccactcgggattgctattagtctgttagtttcatttttgacagcttgggctagaacaggt
561 L A F G I T G P G I A I S -J L V S F T A W A R T G
34150 gagttcaacgcagacggaattactcaagtattcgaaaacttgacaaacacaattcagtcgacggctgatttcatctctcaatac
589 E F N A D G I T Q V F E N L T N T I Q S T A D F I S Q Y
34234 cttccagtctttgtcgaaaaaggaactcaaattttagttaagattattgaaggaattgcatctgctgttcctcaagtagttgaa
617 P V F V E K G T Q I L V K I I E G I A S A V P Q V V E
34318 gtgatttcacaagtcattgaaaatattgtgatgacaatttcgacagttatgcctcaattagtcgaagcaggaattaagataett
645 V I S Q V I E N I V M T I S T V M P Q L V E A G I ~K" I L
34402 gaagcgcttataaatggtcttgttcaatctcttcctactatcattcaagcagctgttcaaattatcactgctttattcaatggt
673 E A L I N G V Q S L P T I I Q A A V Q I I T A L F N G
34486 cttgttcaggcacttcctacgcttattcaagcaggtcttcaaattttgtcagctctcataaacggactagttcaagcgcttccg
701 L V Q A L P T I Q A G L Q I S A L I N G V Q A L P
34570 gcaattattcaagcagctgttcaaattatcatgtcgcttgttcaagcactaattgaaaacttgcctatgataatcgaagcagcg
729 A I I Q A A V Q I I S L V Q A L I E N L P M I I E A A
34654 atgcagattataatgggtctagtcaacgcactgattgaaaatataggacctatcttagaagcagggattcaaattctaatggct
757 M Q I I M G L V N A I E N I G P I L E A G I Q I L M A
34738 ttaatcgagggacttattcaagtgcttcctgaactaattacagcagcgattcaaatcattacttcactattagaagcaatcttg
785 I E G L I Q V P E L I T A A I Q I I T S L L E A I
34822 tcgaaccttcctcaacttctagaagccggagttaaattgcttttatcacttcttcaagggttgctaaatatgcttcctcaacta
813 S N L P Q L E A G V K L S L L Q G L L N P Q L
34906 attgcaggggctttgcaaatcatgatggcacttcttaaagcagttatcgacttcgtccctaaacttcttcaagcaggtgttcaa
841 I A G A Q I M M A L K A V I D F V P K L L Q A G V Q
34990 cttcttaaggcattgattcaaggtattgcttcacttctcggctcacttttatcgacagctggaaacatgctttcatcattagtt
869 L K A Ii l Q G I A S -i G S L L S T A G N M L S S L V
35074 agcaagattgctagctttgtgggacagatggtttcaggaggtgcgaacctgattcgaaacttcattagtggtattgggtcaatg
897 S K I A S F V G Q M V S G G A N I R N F I S G I G S M
35158 attggttcagctgtctctaaaattggcagcatgggaacttcaattgtttctaaggttactggattcgctggacaaatggtaagc
925 I G S A V S K I G S M G T S I V S K V T G F A G Q M V S
35242 gcaggggtcaaccttgttcgaggatttatcaatggtatcagttccatggtaagttctgcggtaagtgcggcggctaatatggct
953 A G V N L V R G F I N G I S S M V S S A V S A A A N M A
35326 agcagtgcattaaatgccgttaagggattcttaggtattcactctccttcacgtgtcatggagcagatgggtatctatacgggt
981 S S A L N A V K G F G I H S P S R V M E Q M G I Y T G
35410 caagggttcgtaaatggtattggtaacatgattcgaactacacgtgacaaggctaaagaaatggctgaaactgttactgaagct
1009 Q G F V N G I G N M I R T T R D K A K E M A E T V T E A
35494 ctcagcgacgtgaagatggatattcaagaaaatggagttatagaaaaggttaaatcagtttacgaaaagatggctgaccaactt
1037 L S D V K M D I Q E N G V I E K V K S V Y E K M A D Q L
35578 cctgaaactcttccagctcctgatttcgaagatgttcgtaaagcagccggttcgcctcgagtggacttgttcaatacaggaagt
1065 P E T Ii P A P D F E D V R K A A G S P R V D Ii F N T G S
35662 gacaaccctaaccaacctcagtcacaatctaaaaacaatcaaggcgagcaaaccgttgtcaacattggaacaatcgtagttcga
1093 D N P N Q P Q S Q S K N N Q G E Q T V V N I G T I V V R
35746 aacaatgacgacgttgacaaactgtcgagaggattgtataatagaagtaaagaaactctatcagggtttggtaacattgtaaca
1121 N N D D V D K S R G Y N R S K E T L S G F G N I V T
35830 ccgtaa 35835
1149 P * dplORF003
53538 atggcacaaaaaggactctttggtgcaaagcctcgttctagcaagaagaacgatgctcagttacttgctcaacggaaaaacagg
1 M A Q K G F G A K P R S S K K N D A Q L A Q R K N R
53622 aagcctgcagttgaggttacttacatttcaggaaacgctctaaaggacgcagttgctagagctcgtactctttcaactaggatt
29 K P A V E V T Y I S G N A K D A V A R A R T S T R I
53706 cttggacacgttcttgatagacttgagttaatcactgaggaagcaaaactcgagcagtatgtagacaaaatgattgaagacgga
57 G H V D R L E L I T E E A K L E Q Y V D K M I E D G
53790 ataggttctattgacgtagaaactgatggactcgatactattcacgatgagctggcaggagtctgcttgtactcacctagtcaa
85 I G S I D V E T D G L D T I H D E A G V C L Y S P S Q
53874 aaaggaatctatgctcctgtcaatcatgttagcaatatgacgaagatgcgaattaagaatcaaatttctcctgagttcatgaag
113 K G I Y A P V N H V S N M T K M R I K N Q I S P E F M K
53958 aaaatgcttcaacggattgtagattcaggaattcctgtcatctatcataattcgaaatttgacatgaaatcgatttattggcga
141 K Q R I V D S G I P V I Y H N S K F D K S I Y W R
54042 ctcggcgtcaaaatgaatgagccagcgtgggatacatatttagccgcaatgcttttaaatgaaaacgagtctcacagcttgaaa
169 L G V K M N E P A W D T Y L A A M L L N E N E S H S L K
54126 agtcttcactctaaatatgttaggaacgaagaaaacgcagaggttgcaaaatttaatgacttatttaaaggaattccttttagt
197 S H S K Y V R N E E N A E V A K F N D F K G I P F S
54210 ttaattcctcctgatgttgcctatatgtatgcggcctatgaccctttgcaaactttcgaactctatgaatttcaagaacaatac
225 L I P P D V A Y M Y A A Y D P L Q T F E L Y E F Q E Q Y
54294 ttgactccaggaactgaacaatgtgaagaatataacctggaaaaagtctcatgggttcttcataatattgagatgcctctaatt
253 L T P G T E Q C E E Y N L E K V S W V L H N I E M P L I
54378 aaagttctcttcgacatggaagtctacggtgtcgacttagaccaagataagctggcagaaattagagaacagtttactgccaat
281 K V L F D M E V Y G V D L D Q D K L A E I R E Q F T A N
54462 atgaacgaggctgagcaagagtttcaacagcttgtcagcgaatggcagcctgaaattgaagaacttcgacaaactaatttccag
309 M N E A E Q E F Q Q L V S E W Q P E I E E L R Q T N F Q
54546 agctatcaaaaactcgaaatggatgcaagaggtcgagtgacggtaagcatttccagtcctactcaattagcaattctgttttat
337 S Y Q K L E M D A R G R V T V S I S S P T Q L A I L F Y
54630 gatatcatgggattgaaaagtcctgaaagggataaacctagaggaacaggcgaaagtattgtcgagcattttgataacgatatc
365 D I M G L K S P E R D K P R G T G E S I V E H F D N D I
54714 tcaaaagcacttttgaaatatagaaaatatgcaaaattagtttcgacctatacaacacttgaccaacaccttgcaaagcctgac
393 S K A L L K Y R K Y A K L V S T Y T T L D Q H L A K P D
54798 aatcgaattcacactacattcaaacagtacggagctaagacagggcgtatgtcaagtgagaatcctaacttacagaatattcct
421 N R I H T T F K Q Y G A K T G R M S S E N P N L Q N I P
54882 tctcgcggtgagggtgcagtagttcgacaaatctttgcagccagtgaagggcattacattattggtagtgactactctcaacaa
449 S R G E G A V V R Q I F A A S E G H Y I I G S D Y S Q Q
54966 gaacctcgttcattggcggaattaagtggcgacgaaagtatgcgacatgcttacgaacaaaacctggacctatattcagttatc
477 E P R S L A E L S G D E S M R H A Y E Q N L D L Y S V I
55050 ggttcgaaactttatggtgttccctatgaagagtgtttagagttctatcccgacggaacgactaacaaggaaggaaaacttcga
505 G S K L Y G V P Y E E C _J E F Y P D G T T N K -E G_K -_, R~
55134 agaaattctgtcaagtccgttcttttaggtcttatgtacggccgcggggctaactcaatcgctgagcagatgaatgtatctgtc
533 R N S V K S V L L G L M Y G R G A N S I A E Q M N V S V
55218 aaagaagcgaataaggttattgaagatttcttcaccgagttccctaaagtggcagactatatcatattcgttcaacagcaggcg
561 K E A N K V I E D F F T E F P K V A D Y I I F V Q Q Q A
55302 caggacttgggatatgttcaaacagctaccggtcgaagaagaaggcttcctgatatgagtcttcctgaatacgagttcgagtat
589 Q D L G Y V Q T A T G R R R R L P D M S L P E Y E F E Y
55386 atcgacgctagcaagaacgaagatttcgacccctttaactttgacgcagaccaacagatggacgatactgttcctgaacatatt
617 I D A S K N E D F D P F N F D A D Q Q M D D T V P E H I
55470 atcgaaaaatattgggcccagctagatagagcctggggatttaagaagaagcaagaaattaaagaccaggcaaaagccgaagga
645 I E K Y W A Q L D R A W G F K K K Q E I K D Q A K A E G
55554 attcttattaaggataacggaggcaagatagctgatgctcagcgccaatgtttgaactcagttattcaaggaacggcagccgac
673 I L I K D N G G K I A D A Q R Q C L N S V I Q G T A A D
55638 atgactaagtacgcaatgattaaggtacacaatgacgctgaattgaaagaattaggattccatttaatgattccagttcacgat
701 M T Y A I K V H N D A E L K E L G F H L M I P V H D
55722 gagttactaggtgaggttcctatcaagaacgcaaaacggggagcagaaaggttgacagaagttatgattgaagcagccaaggac
729 E L L G E V P I K N A K R G A E R L T E V M I E A A K D
55806 attattagtcttccaatgaaatgtgaccccagtatagtagaaagatggtatggtgaagaaattgaaatctaa 55877
757 I I S L P M K C D P S I V E R W Y G E E I E I * dplORF004
40401 atgacaaaatttatcaactcatacggccctcttcacttgaacctttacgtcgaacaagttagtcaggacgtaacgaacaactcc
1 M T K F I N S Y G P L H L N L Y V E Q V S Q D V T N N S
40485 tcgcgagttagttggcgagctactgtcgaccgcgatggagcttatcgaacgtggacttatggaaatattagtaacctttccgta
29 S R V S W R A T V D R D G A Y R T W T Y G N I S N L S V
40569 tggttaaatggttcaagtgttcatagcagtcacccagactacgacacgtccggcgaagaggtaacgctcgcaagtggagaagtg
57 W L N G S S V H S S H P D Y D T S G E E V T L A S G E V
40653 actgttcctcacaatagtgacgggacaaagacaatgtccgtttgggcttcgtttgaccctaataacggcgttcacggaaatatc
85 T V P H N S D G T K T M S V W A S F D P N N G V H G N I
40737 actatctctactaattacactttagacagtattccaaggtctacacagatttctagttttgagggaaatcgaaatctaggatct
113 T I S T N Y T L D S I P R Ξ T Q I S S F E G N R N L G S
40821 ttacatacggttatctttaaccgaaaagtgaactcttttacgcatcaagtttggtaccgagttttcggtagcgactggatagat
141 L H T V I F N R K V N S F T H Q V W Y R V F G S D W I D
40905 ttaggtaagaaccatactactagcgtatcctttacgccgtcactggacttagcaaggtacttacctaaatcaagttccggaaca
169 L G K N H T T S V S F T P S L D L A R Y L P K S S S G T
40989 atggacatctgtattcgaacctataacggaactacgcaaattggtagtgacgtctattcaaacggatggaggttcaacatcccc
197 M D I C I R T Y N G T T Q I G S D V Y S N G R F N I P
41073 gattcagtacgtcctactttttcgggcatttctttagtagacacgacttcagcggttcgacagattttaacagggaacaacttc
225 D S V R P T F S G I S L V D T T S A V R Q I L T G N N F
41157 ctccaaatcatgtcgaacattcaagtcaacttcaacaatgcttccggcgcttacggatccactatccaagcatttcacgctgag
253 L Q I M S N I Q V N F N N A S G A Y G S T I Q A F H A E
41241 ctcgtaggtaaaaaccaagctatcaacgaaaacggcggcaaattgggtatgatgaactttaatggctccgctaccgtaagagca
281 L V G K N Q A I N E N G G K L G M M N F N G S A T V R A
41325 tgggttacagacacgcgaggaaaacaatcgaacgtccaagacgtatctatcaatgttatagaatactatggaccgtctatcaat
309 V T D T R G K Q S N V Q D V S I N V I E Y Y G P S I N
41409 ttctccgttcaacgtactcgtcaaaatcctgcaattatccaagctcttcgaaatgctaaggtcgcacctataacggtaggaggt
337 F S V Q R T R Q N P A I I Q A L R N A K V A P I T V G G
41493 caacagaaaaacatcatgcaaattaccttctccgtggcgccgttgaacactactaatttcacagaagatagaggttcggcgtca
365 Q Q K N I M Q I T F S V A P L N T T N F T E D R G S' A S
41577 gggacgttcactactatttccctaatgactaactcgtccgcgaacttagctggtaactacgggccggacaagtcttacatagtt
393 G T F T T I S L M T N S S A N -i A G N Y G P D K S Y I V
41661 aaggctaaaatccaagacaggttcacttcgactgaatttagtgctacggtagctaccgaatcagtagttcttaactatgacaag
421 K A K I Q D R F T S T E F S A T V A T E S V V L N Y D K
41745 gacggtcgacttggagttggtaaggttgtagaacaagggaaggcagggtcaattgatgcagcaggtgatatatatgctggaggt
449 D G R L G V G K V V E Q G K A G S I D A A G D I Y A G G
41829 cgacaagttcaacagtttcagctcactgataataatggagcattgaacaggggtcaatataacgatgtttggaataagcgtgaa
477 R Q V Q Q F Q L T D N N G A L N R G Q Y N D V W N K R E
41913 acagagtttacatggcgaagtaacaaatacgaggacaaccctacgggaactcgaggtgaatggggactatttcaaaatttctgg
505 T E F T W R S N K Y E D N P T G T R G E W G L F Q N F W
41997 ttagatagctggaaaatggttcaatccttcattacaatgtcaggaagaatgttcatcaggacagcgaacgatggaaacagctgg
533 L D S W K M V Q S F I T M S G R M F I R T A N D G N S W
42081 agacctaacaagtggaaagaggttctatttaagcaagacttcgaacagaataattggcagaaacttgttcttcaaagtgggtgg
561 R P N K W K E V L F K Q D F E Q N N W Q K L V L Q S G W
42165 aaccatcactcaacctatggcgacgcattctattcgaaaactcttgacggcatagtatatttgagaggaaatgtgcataaagga
589 N H H S T Y G D A F Y S K T L D G I V Y L R G N V H K G
42249 cttatcgacaaagaggctactattgcagtacttcctgaaggatttagaccgaaagtttcaatgtatcttcaggctctcaataac
617 Ii l D K E A T I A V L P E G F R P K V S M Y L Q A L N N
42333 tcatatggaaatgccattctatgtatatacactgacggaagacttgtggtgaaatcgaatgtagataattcttggttaaattta
645 S Y G N A I L C I Y T D G R L V V K S N V D N S W L N 1.
42417 gacaatgtctcatttcgtatttaa 42440
673 D N V S F R I * dplORF005
23674 atggctaaaaaatcaaaagctatctcacacacagacgaactgattagtcagtcgtttgacagccccttggcaaagaatcaaaag
1 M A K K S K A I S H T D E L I S Q S F D S P L A K N Q K
23758 ttcaagaaagagcttcaggaagttgaaaagtattatcaatacttcgacggatttgatgtcacggacttgaatactgactatggg
29 F K K E L Q E V E K Y Y Q Y F D G F D V T D N T D Y G
23842 caaacatggaagattgacgaagactcagtcgactataaacctactcgagaaattcgaaactatattcgacaacttatcaaaaag
57 Q T W K I D E D S V D Y K P T R E I R N Y I R-Q L_I -K "K"
23926 caatcacgctttatgatgggtaaagagccagagcttatctttagtccagttcaagacaatcaagatgaacaggctgagaacaag
85 Q S R F M M G K E P E L I F S P V Q D N Q D E Q A E N K
24010 cgtattctattcgactctattttaaggaattgtaaattctggagcaaaagtacaaatgcattagtcgacgccacagtaggtaag
113 R I L F D S I L R N C K F W S K S T N A L V D A T V G K
24094 cgggtattgatgacagtagtagcaaatgccgctcaacaaattgacgtccagttttattcaatgcctcagttcacctatacagtt
141 R V L M T V V A N A A Q Q I D V Q F Y S M P Q F T Y T V
24178 gaccctagaaacccttccagcttgctttctgttgacattgtttatcaggacgagcgtacaaaaggaatgagcactgaaaaacaa
169 D P R N P S S L L S V D I V Y Q D E R T K G M S T E K Q
24262 ctttggcatcattatagatatgaaatgaaagctggaacaagtcaatcaggaattgcaacagctttagaagacattgaagaacaa
197 L W H H Y R Y E M K A G T S Q S G I A T A L E D I E E Q
24346 tgttggctcacttatgccttaacggatggagagtcgaaccaaatctatatgacagaaagtggccaaactactatcaaggagaca
225 C W L T Y A L T D G E S N Q I Y M T E S G Q T T I K E T
24430 gaggctaaacttgtagaaattgaagacaacctaggaaacaagattgaagttcctttaaaagttcaagaatccgccccaaccggc
253 E A K L V E I E D N L G N K I E V P L K V Q E S A P T G
24514 ttgaagcaaattccttgtcgagttattcttaatgaaccattgactaatgacatatacgggacaagcgatgtcaaagaccttatc
281 L K Q I P C R V I L N E P L T N D I Y G T S D V K D L I
24598 acagtagcagataacttgaacaaaactattagtgacttacgagattcacttcgatttaaaatgttcgagcagcctgttatcatt
309 T V A D N L N K T I S D L R D S L R F K M F E Q P V I I
24682 gatggctcttctaagtcaattcaaggaatgaagattgcgccaaacgctttggtcgaccttaagagtgaccctacttcctcaatc
337 D G S S K S I Q G M K I A P N A L V D L K S D P T S S I
24766 ggcggtactggaggcaagcaagctcaagtcacttccatttcaggaaacttcaacttccttccagcggctgaatattatttagag
365 G G T G G K Q A Q V T S I S G N F N F L P A A E Y Y L E
24850 ggcgctaagaaagccatgtatgaactaatggaccagccaatgcctgaaaaggtacaggaggcgccatcaggaattgcaatgcag
393 G A K K A M Y E L M D Q P M P E K V Q E A P S G I A M Q
24934 ttcttattctacgacctaatttctcgatgtgacggaaaatggattgagtgggatgatgctattcaatggctcattcaaatgctg
421 F L F Y D L I S R C D G K W I E W D D A I Q W L I Q M L
25018 gaagaaattttagcaacagtgaatgttgacttgggaaatattcctcaagatattcaatcaagttatcaaacacttacgacaatg
449 E E I L A T V N V D L G N I P Q D I Q S S Y Q T L T T
25102 actatcgaacaccactatccaattcctagcgatgaactttctgctaagcaacttgcgctcactgaagttcaaactaatgtacgc
477 T I E H H Y P I P S D E L S A K Q L A L T E V Q T N V R
25186 agccaccaatcttacattgaagaattcagtaagaaggaaaaggcggacaaggaatgggaacgcattttggaagaacttgctcag
505 S H Q S Y I E E F S K K E K A D K E E R I L E E L A Q
25270 cttgacgaaatctcagctggagcattgcctgtattagcaaacgaattaaacgaacaagaggagcctcaagatgaaacgagtgaa
533 L D E I S A G A L P V L A N E L N E Q E E P Q D E T S E
25354 gaagacgaagttgatgacaaagaaaaagaacaaactgaacaaccaaccgaagaaggagtcgacccagacgttcaaggttaa 25434
561 E D E V D D K E K E Q T E Q P T E E G V D P D V Q G * dplORFOOβ
45296 atgattgaaatcgttatagcacgttcgaaagctaggcgaggtcgaaccctatttattgaaacatgggcaagcactgatgaagat
1 M I E I V I A R S K A R R G R T L F I E T W A S T D E D
45380 gcagttaaaatggcagaaaagatttccagcttgcccaatgtagtcgagacgtcttctaataacttcgaactaccttataagtat
29 A V K M A E K I S S L P N V V E T S S N N F E L P Y K Y
45464 ttcaataatgttatagacgctctagatgaatgggagcttcacatcttcggcgaacttgataaagatgttcaagactacattgac
57 F N N V I D A L D E W E L H I F G E L D K D V Q D Y I D
45548 tctcgaaaccgaatagcttcttcaagcaatgagcagttttcgttcaagactactccattcgcgcaccaggttgaatgtttcgaa
85 S R N R I A S S S N E Q F S F K T T P F A H Q V E C F E
45632 tacgcacaagagcatccatgtttccttttaggcgatgagcaaggtttagggaaaactaaacaggcaattgatattgcagttagc
113 Y A Q E H P C F L L G D E Q G L G K T K Q A I D I A V S
45716 aggaaggcaagtttcaaacattgtttaatcgtatgttgcatatcagggctcaaatggaattgggcaaaagaagtaggtattcat
141 R K A S F K H C L I V C C I S G L K W N W A K E V G I H
45800 tcaaatgagtcagctcatattttaggaagtcgagtcactaaagatgggaaattagtgattgacggagtttctaaacgggcagaa
169 S N E S A H I L G S R V T K D G K L V I D G V S K R A E
45884 gacttgcttggtggccacgacgaattcttccttatcactaacattgaaactcttcgcgatgctgtgttcattaaatacttaaat
197 D L L G G H D E F F L I T N I E T L R D A V F I K Y L N
45968 gaactgacaaaaagcggagaaattggaatggttattattgacgagattcacaagtgtaagaacccttcaagtaagcaaggggct
225 E L T K S G E I G M V I I D E I H K C K N P S S K Q G A
46052 tcaattcaaaagctccaaagttattacaagatgggacttacaggaactcctctaatgaataacccaatcgatgtattcaatgtt
253 S I Q K L Q S Y Y K M G L T G T P L M N N P I D V F N V
46136 atgaagtggctaggggcggaacatcatacactgactcagttcaaagagcgatactgtatcgtcgaccagttcaatcaaatcact
281 K L G A E H H T L T Q F K E R Y C I V D Q F N Q I T
46220 ggatatcgaaatctagctgaacttcgcgagcttgtcaacgactacatgcttagaagaacgaaggaagaagttttagacctgcct
309 G Y R N L A E L R E L V N D Y M L R R T K E E V L D L P
46304 gaaaagattcgagtcacagagtatgtcgacatgaactcgaaacagtcaaaaatctataaggaagttttgactaaacttgttcaa
337 E K I R V T E Y V D M N S K Q S K I Y K E V L T K L V Q
46388 gaaatagataaagtcaagctcatgcctaaccctctagccgaaacgattcgacttcgacaagcgactggaaatccttcgatttta
365 E I D K V K L P N P L A E T I R L R Q A T G N P Ξ I L
46472 actactcaagatgtcaagtcttgcaagttcgaaagatgtatcgaaattgtcgaggaatgtatccagcaaggaaagtcctgcgtg
393 T T Q D V K S C K F E R C I E I V E E C I Q Q G K S C V
46556 atatttagcaattgggaaaaggttattgaacctcttgctaagatactttcgaagacagtcaaatgcaacctggtaacaggagaa
421 I F S N W E K V I E P L A K I L S K T V K C N L V T G E
46640 accgcagataagttcaacgaaattgaagaatttatgaatcacagaaaggcttctgttattttaggaactataggtgcgctagga
449 T A D K F N E I E E F M N H R K A S V I L G T I G A L G
46724 acaggatttactttgacgaaagcggatacggttattttcttagatagtccgtggacacgcgcagaaaaggaccaagccgaagat
477 T G F T L T K A D T V I F L D S P W T R A E K D Q A E D
46808 aggtgtcatagaattggcgcaaaaagttctgtcactatctacacgcttgtcgccaaaggtactgttgacgaacgtatagaagac
505 R C H R I G A K S S V T I Y T L V A K G T V D E -J. I E D
46892 cttattgaacggaaaggagaattagcagattatatcgtagatggtaagcctatgaaatctaaaattggtaaccttttcgatatc
533 L I E R K G E L A D Y I V D G K P M K S K I G N L F D I
46976 ctgcttaaatag 46987
561 L K * dplORF007
22230 atgacaataagcctgagaaataaactacctaagttcaacttcgtcccttttagtaagaaacaactccagctcctaacatggtgg
1 M T I Ξ L R N K L P K F N F V P F S K K Q L Q L L T W W
22314 acaaagggctcaccttttcgaactttcgatatcgtcatagcagacggttccattcgttcaggaaaaacagtatcgatggctctt
29 T K G S P F R T F D I V I A D G S I R S G K T V S M A L
22398 tcattttccctttgggccatgacggaattcaacggacaaaactttgccatctgtggtaagacaattcactcagctcgacgaaat
57 S F S L W A M T E F N G Q N F A I C G K T I H S A R R N
22482 gttattcagcctctaaagcaaatgctcacaagtcgcgggtatgaaattcgagatgttcgaaatgaaaatctacttattattaga 85 V I Q P L K Q L T S R G Y E I R D V R N E N L L I I R 22566 cactttagaaatggcgaagaaattgtcaactacttctatatatttggaggaaaagatgagtcgagtcaagaccttatacagggg 113 H F R N G E E I V N Y F Y I F G G K D E S S Q D L I Q G 22650 gtaacattagcaggtatcttctgtgatgaggtggcactgatgcctgaatcgtttgtcaaccaagcgacagggcgctgttccgta 141 V T L A G I F C D E V A L M P E S F V N Q A T G R C S V 22734 acaggttcgaaaatgtggttctcttgtaacccggccaatcctaatcactacttcaagaagaactggattgacaaacaggtcgaa 169 T G S K M W F S C N P A N P N H Y F K K N W I D K Q V E 22818 aagcgtatcttatatcttcactttacaatggacgacaaccctagcttgacggatagcattaaaaggcgctatgagaaaatgtat 197 K R I L Y L H F T M D D N P S L T D S I K R R Y E K M Y 22902 gctggagtcttcaggaaaagatttattctcggcctttgggtaacagcagatggtctagtttattcaatgttcaatgaagagcag
225 A G V F R K R F I L G Li W V T A D G L V Y S M F N E E Q 22986 catgtcaaaaagctcaatatagaattcgaccgtttattcgtagcaggcgactttggtatctataatgcaacaaccttcggcctt 253 H V K K L N I E F D R L F V A G D F G I Y N A T T F G IJ 23070 tatggattctcgaaacgtcataagcgctaccatctaattgagtcatactaccactcagggcgcgaggcggaagagcaactaact 281 Y G F S K R H K R Y H L I E S Y Y H S G R E A E E Q L T 23154 gaggcggatgttaattcgaatattcaatttagttcagttctacaaaagactactaaagagtacgcaaatgatttagtcgatatg 309 E A D V N S N I Q F S S V L Q K T T K E Y A N D L V D M 23238 atacgaggaaagcaaatcgaatatataattctcgacccgtctgcttctgctatgattgttgaacttcaaaagcatccttatata 337 I R G K Q I E Y I I L D P S A S A M I V E L Q K H P Y I 23322 gctagaaagaatatccctatcattcctgctcgaaatgacgtgacgcttggcatttcatttcacgctgaactcttggctgagaat 365 A R K N I P I I P A R N D V T L G I S F H A E L L A E N 23406 agatttacactcgaccctagcaacacgcacgacattgatgaatactatgcttacagctgggacagtaaagcgagccaaacggga 393 R F T L D P S N T H D I D E Y Y A Y S W D S K A S Q T G 23490 gaagatagagtcattaaagagcatgaccactgcatggataggaacagatatgcctgtctcactgacgctctaatcaacgatgac 421 E D R V I K E H D H C M D R N R Y A C L T D A L I N D D 23574 ttcggtttcgaaatacaaatattatccggaaaaggcgctagaaactaa 23621
449 F G F E I Q I L S G K G A R N * dplORFOOδ
49624 gtgatacagcttcaagtcttaaataaagttctcgaagaaaagagcttatccattttagaaaataatggaattgaccaagaatac
1 V I Q L Q V L N K V L E E K S L S I L E N N G I D Q E Y
49708 ttcacggattatttagacgagtatcaatttattcaagaacacttttcgagatatggaagagttccggacgacgaaactattctc
29 F T D Y L D E Y Q F I Q E H F S R Y G R V P D D E T I L
49792 gaccattttcctggattcgaatttttcgaaattggcgaaactgatgaataccttatcgacaagctaaaagaggagcatctatat
57 D H F P G F E F F E I G E T D E Y L I D K L K E E H L Y
49876 aattcacttgttccaattttaacggaagcggctgaggacattcaagtagatagtaacattgcgattgcgaatataattccaaaa
85 N S L V P I L T E A A E D I Q V D S N I A I A N I I P K
49960 ctagaagaacttttcaatcgctctaaattcgtaggcggactagacattgctcgaaatgctaaacttcgactagactgggcgaat
113 L E E L F N R S K F V G G L D I A R N A K L R L D W A N
50044 actattagaaaccatgacggtgaaagacttggaatatcgacagggtttgaactattggacgacgtgcttggaggcttacttcct
141 T I R N H D G E R L G I S T G F E L L D D V L G G L L P
50128 ggtgaggatttgattgtcataatggctcgacctggacaaggtaagtcgtggactattgataaaatgcttgcaactgcttggaag
169 G E D L I V I M A R P G Q G K S W T I D K M L A T A W K
50212 aacgggcatgatgtccttctatatagcggggaaatgagtgaaatgcaagttggtgctcgtatagatactattctttcgaatgtt
197 N G H D V L L Y S G E M S E M Q V G A R I D T I L S N V
50296 agcatcaattcaattaccaaagggatttggaacgaccatcagttcgaaaaatatgaggaccatattcaagcaatgactgaggct
225 S I N S I T K G I W N D H Q F E K Y E D H I Q A M T E A
50380 gaaaattcccttgtggtagtcacgccctttatgattggaggaaagaaccttacccctgcaattttagatagcatgatatctaaa
253 E N S L V V V T P F M I G G K N L T P A I L D S M I S K
50464 tatagaccatctgtggtggggattgaccagctttcactcatgagcgagtcttatccaagcagggagcagaagcgaatccagtac
281 Y R P S V V G I D Q L S L M S E S Y P S R E Q K R I Q Y
50548 gccaacatcaccatggacctatataagatttctgctaaatatggaattcctattgtgcttaatgtccaagcagggcgttcggct
309 A N I T M D L Y K I S A K Y G I P I V L N V Q A G R S A
50632 aaaactgaaggcgctgaaagtatggaactagaacatatagcagaaagtgatggagtaggtcaaaatgctagcagagttatcgct
337 K T E G A E S M E L E H I A E S D G V G Q N A S R V I A
50716 atgaagcgtgacgaaaaatccggcatacttgaactatctgtcgttaaaaaccgatatggcgaagaccgaaaaatcatcgaatat
365 M K R D E K S G I L E L S V V K N R Y G E D R K I I E Y
50800 atgtgggacgttgaaactggaacctatactcttataggattcaaagaggaaggcgaagaaggaactgaaaaaggcgaaagctct
393 M W D V E T G T Y T L I G F K E E G E E G T E K G E S S
50884 ccattgaaagcaaaagcctctaggtcgactgctcgtcttcgaagtaaggttacaagggaaggagttgaagcattttga 50961
421 P L K A K A S R S T A R L R S K V T R E G V E A F * dplORF009
13160 atgacagactttaaaaaacgcttcaagaaagcagtaacagaaacaatcaatcgtgacggtatcgagaaccttatggattggctc
1 M T D F K K R F K K A V T E T I N R D G I E N-L M_D -W V
13244 gaaaatgataccaatttcttctcaagtccagcaagcactcgataccatggaagctatgaaggtggacttgtcgagcactcatta
29 E N D T N F F S S P A S T R Y H G S Y E G G L V E H S L
13328 aacgtgttcaatcaactacttttcgaaatggataccatggtaggcaaaggctgggaagacatttacccaatggaaacagttgca
57 N V F N Q L L F E M D T M V G K G E D I Y P M E T V A
13412 atcgtagcactatttcacgacctttgcaaagttggtcagtatcgtgaaactgaaaaatggcgcaagaacagcgacggtgaatgg
85 I V A L F H D L C K V G Q Y R E T E K W R K N S D G E W
13496 gaaagctatttagcatatgaatacgaccctgagcaacttacaatgggacatggtgcaaaatctaatttccttcttcaacgtttc
113 E S Y L A Y E Y D P E Q L T M G H G A K S N F L L Q R F
13580 attcaactcacgccagttgaagctcaagcaattttctggcatatgggagcctatgatattagtccttatgcaaatttgaatgga
141 I Q L T P V E A Q A I F W H M G A Y D I S P Y A N L N G
13664 tgtggagcagccttcgaaactaatccacttgcattcttaatccatcgcgcagatatggccgcaacttatgtagtcgaaaatgaa
169 C G A A F E T N P L A F L I H R A D M A A T Y V V E N E
13748 aacttcgaatactctcaaggtccagttgaacaagaggctgaggttgaagaagtagttgaagaaaaacctaagagttcaactcgt
197 N F E Y S Q G P V E Q E A E V E E V V E E K P K S S T R
13832 aagaaacctgcgcctaaggaagaaaaagttgaagaggctgaagaaaaaccaaaagctggaatcactcgacgtcgcaaacctgcg
225 K K P A P K E E K V E E A E E K P K A G I T R R R K P A
13916 ccaaaagaggaagaggtagaagagcctaaagaagagcctaagaaagcatcttctaaaattcgaatgcctaaaaagactgaaaag
253 P K E E E V E E P K E E P K K A S S K I R M P K K T E K
14000 gtcgaagaggtagaaagcgcagacgagccgaaagttgaagaagcagaggacgacaatgtggtggtacctgctggatatgttcga
281 V E E V E S A D E P K V E E A E D D N V V V P A G Y V R
14084 gatgtctactacttctacagtgaagtcgctgacgtttactacaagaaagatgtcgacgagcctgacgatgacagcgacattctt
309 D V Y Y F Y S E V A D V Y Y K K D V D E P D D D S D I L
14168 gtagacgaagaagagtacatggacgcaatgtgtcctgtattagaagaagacttcttctacgaacttgacggcaaggttcacaaa
337 V D E E E Y M D A M C P V L E E D F F Y E L D G K V H K
14252 ttagcaaaaggtgaacgcttgccggaagaatacgacgaagaaacttgggaacctatcactgaagcagaatacatcaagcgaaca
365 L A K G E R L P E E Y D E E T E P I T E A E Y I K R T
14336 gaaaaacctaaagcagttgcaaaacctactcgaaaaactccagcgccttctcgtcgccctcgcccttaa 14404
393 E K P K A V A K P T R K T P A P S R R P R P * dplORFOlO
8699 atgaaattggaacagttgatgaaggactggaataaggattcgaaagctcttgtagcagttcaaggacttgaacgtgaagcgctt
1 M K L E Q L M K D W N K D S K A L V A V Q G L E R E A L
8783 ccaagaatccctttttctgcgccttctatgaattatcaaacctacggcgggctccctcgaaaaagggtagttgaattcttcggt
29 P R I P F S A P S M N Y Q T Y G G L P R K R V V E F F G
8867 cctgagtcaagtgggaaaactacttcagctctcgacattgtcaagaatgcgcaaatggtatttgagcaggaatgggaacagaag
57 P E S S G K T T S A L D I V K N A Q M V F E Q E W E Q K
8951 actgaagaactcaaggaaaagctggaaaatgcgcgtgcatccaaagctagcaagactgctgtcaaggaacttgaaatgcaactc
85 T E E L K E K L E N A R A S K A S K T A V K E L E M Q L
9035 gatagtcttcaagagcctcttaagattgtatatcttgaccttgagaatacattagacactgagtgggctaaaaagattggagtc
113 D S L Q E P L K I V Y L D L E N T L D T E W A K K I G V
9119 gatgttgacaatatttggatagttcgccctgaaatgaacagcgctgaagaaatacttcaatatgttttagacattttcgaaaca
141 D V D N I I V R P E M N S A E E I L Q Y V L D I F E T
9203 ggtgaagttggcctagtagttctagattccttgccttacatggtcagtcaaaaccttattgatgaagagttgactaaaaaggcc
169 G E V G L V V L D S L P Y V S Q N L I D E E L T K K A
9287 tatgcaggaatctcagcgcctttgactgaatttagtcgaaaggttactcctcttcttactcgctacaatgcaatattcctaggc
197 Y A G I S A P L T E F S R K V T P L L T R Y N A I F L G
9371 atcaatcaaattcgagaagatatgaatagtcagtacaatgcctattcaactccaggcggaaagatgtggaagcatgcttgtgca
225 I N Q I R E D M N S Q Y N A Y S T P G G K M W K H A C A
9455 gttcgacttaaatttagaaaaggtgactaccttgacgaaaacggtgcatcattgacccgtactgctcgaaaccctgcagggaat
253 V R L K F R K G D Y L D E N G A S L T R T A R N P A G N
9539 gtagtagagtcattcgtcgagaagaccaaagcatttaagccggacagaaaattagtttcctatacgctttcctatcatgatgga
281 V V E S F V E K T K A F K P D R K L V S Y T L S Y H D G
9623 attcaaattgaaaatgaccttgtagatgtcgctgtcgaatttggagtcattcaaaaggcaggggcatggttcagtatcgtcgac
309 I Q I E N D L V D V A V E F G V I Q K A G A W F S I V D
9707 cttgaaactggagaaattatgacagatgaagacgaagaaccattgaagttccaaggcaaggcaaatctagttcgacgcttcaag
337 L E T G E I M T D E D E E P L K F Q G K A N L V R R F K
9791 gaggatgactacttattcgacatggtgatgactgcggttcacgaaattatcactcgagaagaaggctaa 9859
365 E D D Y L F D M V M T A V H E I I T R E E G * dplORFOll
28017 atgaatatttatgattatatcaacgcaggggagattgctagctacattcaagcacttccttcaaacgctcttcaataccttgga
1 M N I Y D Y I N A G E I A S Y I Q A L P S N A L Q Y L G
28101 ccaactcttttccctaatgctcaacaaacagggacagacatttcatggctcaagggtgcaaataatttgccagtaactatccag
29 P T L F P N A Q Q T G T D I S W L K G A N N L P V T I Q
28185 ccatctaactacgacgcgaaagcaagtcttcgtgaacgtgctggatttagcaaacaagctactgagatggcattcttccgtgag
57 P S N Y D A K A S L R E R A G F S K Q A T E M A F F R E
28269 tctatgcgacttggtgaaaaagaccgtcaaaacttgcaaatgctattgaaccaaagttcagctcttgcccaaccacttatcact
85 S M R L G E K D R Q N L Q M L L N Q S S A L A Q P L I T
28353 caactctataatgatactaagaaccttgtagacggtgttgaagcgcaagcagaatacatgcgtatgcaattgcttcaatacggt
113 Q L Y N D T K N L V D G V E A Q A E Y M R M Q L L Q Y G
28437 aaattcactgtcaaatcaactaacagcgaggctcaatacacttacgactacaacatggatgctaagcaacaatatgcagtcact
141 K F T V K S T N S E A Q Y T Y D Y N M D A K Q Q Y A V T
28521 aagaaatggactaacccagctgaaagtgaccctatcgctgacattttagcagcaatggatgacatcgaaaatcgtacaggtgtt
169 K K W T N P A E S D P I A D I L A A M D D I E N R T G V
28605 cgccctactcgaatggtcttgaaccgaaacacttataaccaaatgactaagagtgactctatcaagaaagctcttgcaattggt
197 R P T R M V L N R N T Y N Q M T K S D S I K K A L A I G
28689 gttcaaggttcttgggaaaacttcttgcttcttgcaagtgacgctgagaaattcatcgctgaaaaaacaggtcttcaa&tcgct
225 V Q G S W E N F L L L A S D A E K F I A E K T G Ji Q I A
28773 gtctactctaagaaaattgctcagttcgctgacgctgacaaacttcctgacgttggtaacattcgtcagttcaacttgattgac
253 V Y S K K I A Q F A D A D K L P D V G N I R Q F N L I D
28857 gacggtaaagtggtattgcttccacctgacgcagttggtcacacttggtacggtactactccagaagcattcgacttggcttca
281 D G K V V L L P P D A V G H T W Y G T T P E A F D L A S
28941 ggcggaacagacgctcaagttcaagttctttcaggcggacctaccgttacaacttatcttgaaaaacatcctgtcaacattgca
309 G G T D A Q V Q V L S G G P T V T T Y L E K H P V N I A
29025 acagttg atcagctgttatgattccatcattcgaaggaattgactatgtaggagttctcacaactaattag 29096
337 T V V S A V M I P S F E G I D Y V G V L T T N * dplORF012
5346 atgagtattaagttcaaaaccgaagaactttcaaaaattgtttctcagctcaataagttgaagcctagcaagttgctagaaatc
1 M S I K F K T E E L S K I V S Q L N K L K P S K L L E I
5430 acaaactattggcatatttttggtgacggcgaatgcgtcatgtttacagcgtatgatggctcaaacttccttcgatgcattatc
29 T N Y W H I F G D G E C V M F T A Y D G S N F L R C I I
5514 gacagcgatgttgaaattgacgtgattgtgaaagcagagcagtttggaaaacttgtagaaaagaccacggccgcaaccgtcaca
57 D S D V E I D V I V K A E Q F G K L V E K T T A A T V T
5598 ttagttcctgaagaatcttcgctaaaagttattgggaatggtgagtacaatattgatattgttacagaagatgaagagtaccct
85 L V P E E S S L K V I G N G E Y N I D I V T E D E E Y P
5682 acattcgaccacttgctcgaagacgtgagtgaagaaaatgctctcactttgaaaagctcgctgttctacggaatcgccaatatc
113 T F D H L L E D V S E E N A L T L K S S L F Y G I A N I
5766 aacgattctgcggtatctaaatcaggagcagatggaatttataccggcttcctgttaaaaggcggaaaagcaattactacagac
141 N D S A V S K S G A D G I Y T G F L L K G G K A I T T D
5850 atcattcgcgtatgtatcaaccctatcaaggaaaagggactagaaatgctcattccttacaacctaatgagtattttagcaagt
169 I I R V C I N P I K E K G L E M L I P Y N L M S I L A S
5934 attcctgatgagaagatgtacttctggcaaattgacgatactactgtctatatttcatcggcttcagtcgaaatttatggaaaa
197 I P D E K M Y F W Q I D D T T V Y I S S A S V E I Y G K
6018 ttgatggaaggtatggaagattatgaagacgtttcacagcttgactcaattgagtttgaagatgatgcggctatccctacagca
225 L M E G M E D Y E D V S Q L D S I E F E D D A A I P T A
6102 gaaatcctgagcgtattagaccgccttgtactattcacttcagcctttgacaaaggaaccgtcgaattcttattcttgaaagac
253 E I L S V L D R L V L F T S A F D K G T V E F L F L K D
6186 cgacttcgaattaaaacttctactagcagttatgaagacatcatgtacgcatctgctggcaagaaagtttcgaagaaagaattc
281 R L R I K T S T S S Y E D I M Y A S A G K K V S K K E F
6270 acttgccaccttaacagcttac cttgaaggaaattgtatcaaccgtcaccgaagaaaacttcactgtctcttatggaagcgaa
309 T C H L N S L L L K E I V S T V T E E N F T V S Y G S E
6354 accgcaattaagatttcatcgaatggtgtcgtttacttcctagcacttcaagagccggaagaataa 6419
337 T A I K I S S N G V V Y F L A L Q E P E E * dplORF013
10215 atgaatttagcttctaaataccgtcctcaaactttcgaggaagtggtagctcaagaatatgtcaaagaaattcttttgaatcaa
1 M N L A S K Y R P Q T F E E V V A Q E Y V K E I L L N Q
10299 ttacaaaatggcgctatcaaacacggctatctattctgtggtggcgctggaactggtaaaaccactactgctcgaattttcgcg
29 L Q N G A I K H G Y L F C G G A G T G K T T T A R I F A
10383 aaggatgtgaacaaaggacttggctctcctattgaaattgatgctgcttctaataatggggtagaaaatgttcgaaacattatt
57 K D V N K G L G S P I E I D A A S N N G V E N V R N I I
10467 gaagattctagatacaagtctatggacagcgagttcaaagtttacatcattgacgaggttcatatgctttcaaccggagcattt
85 E D S R Y K S M D S E F K V Y I I D E V H M L S T G A F
10551 aatgcgctgttgaaaacattagaagagccctcatcgggaaccgtgttcattctatgtactactgaccctcaaaagattcctgac
113 N A L L K T L E E P S S G T V F I L C T T D P Q K I P D
10635 actattctcagtcgagttcaacggtttgactttactcgaattgataatgacgacatcgttaatcaacttcaatttattatcgaa
141 T I L S R V Q R F D F T R I D N D D I V N Q L Q F I I E
10719 agtgaaaatgaagaaggagctggttatagttatgagcgtgacgccctttcgtttattgggaaacttgcaaatggaggaatgcgt
169 S E N E E G A G Y Ξ Y E R D A L S F I G K L A N G G M R
10803 gacagtatcacaaggctcgaaaaagtccttgattatagtcatcacgttgacatggaagccgtttctaatgcactaggagttccg
197 D S I T R L E K V L D Y S H H V D M E A V S N A L G V P
10887 gactacgaaacattcgcttcacttgttgaagctattgccaactatgacggctcaaagtgtttagaaattgtaaatgacttccac
225 D Y E T F A S L V E A I A N Y D G S K C L E I V N D F H
10971 tactcaggaaaagacttgaaattagtgactcgaaactttacagacttccttttagaggtttgtaagtattggctagttcgagat
253 Y S G K D L K L V T R N F T D F L L E V C K Y W L V R D
11055 atttcaatcactcaacttcctgctcattttgaaagtaagctagagcaattctgtgaggcttttcaatatcctactctattgtgg
281 I S I T Q L P A H F E S K L E Q F C E A F Q Y P T L L W
11139 atgctagaagaaatgaatgaacttgctggagttgttaaatgggagcctaatgctaaaccgataattgaaaccaaacttcttttg
309 M L E E M N E -J A G V V K W E P N A K P I I E T K L L L
11223 atgagcaaggaggagtga 11240
337 M S K E E * dplORF014
50961 atgaaagtaaatggtcttcaaattgaagcgactcctgaacaaataattgaaaaactttcgagacaacttgaagacgaaggaaca
1 M K V N G L Q I E A T P E Q I I E K L S R Q L E D E G T
51045 ttcatttttagacgaactaagtcgcttggaagcaactatcaattctcatgcccgtttcatgcaggagggactgaaaagcatccc
29 F I F R R T K S L G S N Y Q F S C P F H A G G T E K H P
51129 tcttgtggcatgagtagaaatccttcttattcaggaagtaaggtgacggaagctggaacggttcactgtttcacttgcggctac
57 S C G M S R N P Ξ Y S G S K V T E A G T V H C F T C G Y
51213 acttcaggactaactgaattcgtctcgaatgtattaggtcgaaacgatggagggttctatggaaaccagtggctgaaaaggaat
85 T S G L T E F V S N V L G R N D G G F Y G N Q W L K R N
51297 tttggaacatctagcgaagtagttaggcaaggcgtcagccctgaagcgtttcgaagaaatgggagaactgaaaaagtcgagcat
113 F G T S S E V V R Q G V S P E A F R R N G R T E K V E H
51381 aaaatcattcctgaagaggaacttgataaataccggtttattcatccttatatgtatgaacggaaattgacggacgagctcatc
141 K I I P E E E L D K Y R F I H P Y M Y E R K L~T D _E -i, I "
51465 gagatgtttgatgtaggttatgacaaactgcatgattgcatcacctttccagtacggaacctcaagggcgaaacagtattcttc
169 E M F D V G Y D K L H D C I T F P V R N L K G E T V F F
51549 aaccgtcgaagtgttcgttctaagtttcaccagtacggtgaagatgaccctaaaacggaatttctttatggccaatatgagctt
197 N R R S V R S K F H Q Y G E D D P K T E F L Y G Q Y E L
51633 gtagcatttcgagactattttgaaaaacctattagtcaagtattcgtgactgagtctgttatcaactgcttgactctttggtca
225 V A F R D Y F E K P I S Q V F V T E S V I N C L T L W S
51717 atgaagattccagcagtcgctcttatgggagtaggtggaggaaatcaaatcaatttactaaaacgacttccttatagaaatatt
253 M K I P A V A L M G V G G G N Q I N L L K R L P Y R N I
51801 gttctagcacttgaccctgataacgctgggcagacagcgcaggaaaaactctaccgacagttaaagcgaagcaaggtcgttaga
281 V L A L D P D N A G Q T A Q E K L Y R Q L K R S K V V R
51885 tttttgaactaccctaaagagttctatgataataagtgggatataaacgaccatccggaattattaaattttaatgatttagtc
309 F L N Y P K E F Y D N K W D I N D H P E L L N F N D L V
51969 ttgtag 51974
337 L * dplORF015
3793 atgggatttaatctatacttcgcaggaggtcacgctattagcactgacgattatttgaaggaaagaggagccaatcgcctattc
1 G F N L Y F A G G H A I S T D D Y L E R G A N R L F
3877 aatcaactgtacgaaagaaacgggattggcaaaaggtggattgagcataagaaaaccaatccaagcactacttcaaaactattc
29 N Q L Y E R N G I G K R I E H K K T N P S T T S K L F
3961 gtcgactctagtgcatattctgctcataccaaaggggctgaagttgacattgacgcctatatcgaatacgtgaatgataacgtg
57 V D S S A Y S A H T K G A E V D I D A Y I E Y V N D N V
4045 ggaatgtttgactgtatcgccgaactcgataaaattcctggtgtatttagacagcctaagacacgtgaacagcttttggaagca
85 G M F D C I A E L D K I P G V F R Q P K T R E Q L L E A
4129 ccacaaatttcttgggataattatctatacatgcgcgagcgaatggttgagaaagacaagctcttacctattttccatatggga
113 P Q I S W D N Y L Y M R E R M V E K D K L L P I F H M G
4213 gaagactttaaatggctcaacttgatgctcgaaactacattcgaaggcggaaagcatattccttacattggaatttcaccagcc
141 E D F K L N L M L E T T F E G G K H I P Y I G I S P A
4297 aatgactcgactacgaagcataaagacaagtggatggaaagagtattcgaagttattcgaaacagttctaatccagacgttaag
169 N D S T T K H K D K W M E R V F E V I R N S S N P D V K
4381 actcacgcatttgggatgacagttactagccaattagagcgtcacccattctatagcgccgactctacttctgtactgctcaca
197 T H A F G M T V T S Q L E R H P F Y S A D S T S V L L T
4465 ggagcgatgggaaacattatgacgtcaaaaggattagttgacttgtcacagaagaatggaggaattgatgctgtccgtaggctg
225 G A M G N I M T S K G L V D L S Q K N G G I D A V R R L
4549 ccaaaaccggttcaagttgaaattgaatccattatcgaagaaactggagcgcattttagcctagagcaattagttgaggactat
253 P K P V Q V E I E S I I E E T G A H F S L E Q L V E D Y
4633 aaacttcgagcattgttcaatgttcaatacatgctgaattgggcagagaactatgaattcaagggaattaaaaatcgtcaacgt
281 K L R A L F N V Q Y M L N A E N Y E F K G I K N R Q R
4717 cgactattttag 4728
309 R L F * dplORF016
43413 atgggagtcgatattgaaaaaggcgttgcgtggatgcaggcccgaaagggtcgagtatcttatagcatggactttcgagacggt
1 M G V D I E K G V A M Q A R K G R V S Y S M D F R D G
43497 cctgatagctatgactgctcaagttctatgtactatgctctccgctcagccggagcttcaagtgctggatgggcagtcaatact
29 P D S Y D C S S S M Y Y A L R S A G A S S A G W A V N T
43581 gagtacatgcacgcatggcttattgaaaacggttatgaactaattagtgaaaatgctccgtgggatgctaaacgaggcgacatc
57 E Y H A W L I E N G Y E L I S E N A P W D A K R G D I
43665 ttcatctggggacgcaaaggtgctagcgcaggcgctggaggtcatacagggatgttcattgacagtgataacatcattcactgc
85 F I W G R K G A S A G A G G H T G M F I D S D N I I H C
43749 aactacgcctacgacggaatttccgtcaacgaccacgatgagcgttggtactatgcaggtcaaccttactactacgtctatcgc
113 N Y A Y D G I S V N D H D E R W Y Y A G Q P Y Y Y V Y R 43833 ttgactaacgcaaatgctcaaccggctgagaagaaacttggctggcagaaagatgctactggtttctggtacgctcgagcaaac 141 L T N A N A Q P A E K K L G W Q K D A T G F W Y A R A N 43917 ggaacttatccaaaagatgagttcgagtatatcgaagaaaacaagtcttggttctactttgacgaccaaggctacatgctcgct 169 G T Y P K D E F E Y I E E N K S F Y F D D Q G Y M L A 44001 gagaaatggttgaaacatactgatggaaattggtattggttcgaccgtgacggatacatggctacgtcatggaaacggattggc 197 E K L K H T D G N Y F D R D G Y M A T S K R I G 44085 gagtcatggtactacttcaatcgcgatggttcaatggtaaccggttggattaagtattacgataattggtattattgtgatgct 225 E S Y Y F N R D G S M V T G I K Y Y D N W Y Y C D A 44169 accaacggcgacatgaaatcgaatgcgtttatccgttataacgacggctggtatctactattaccggacggacgtctggcagat 253 T N G D M K S N A F I R Y N D G Y L L L P D G R L A D 44253 aaacctcaattcaccgtagagccggacgggctcattactgctaaagtttaa 44303
281 K P Q F T V E P D G L I T A K V * dplORF017
11242 atgattggacagggacttgttaaatctaccatttcgaaatggaaacaacttccaaaatatataatcgtcgaaggtgaagtaggt
1 M I G Q G L V K S T I S K K Q L P K Y I I V E G E V G
11326 tcaggacggaagaccttaatccgttatattgcttcgaaatttgacgctgattctattgtagtaggaacgagtgtagatgacatt
29 S G R K T L I R Y I A S K F D A D S I V V G T S V D D I
11410 cgaaacatcattcaggatgcacagactattttcaaggcgagaatctacgtgatagacggaaatagcctgtcaatgtcagctctt
57 R N I I Q D A Q T I F K A R I Y V I D G N S L S M S A L
11494 aactcgcttttgaagatagcggaagagccacctttaaactgtcatatagccatgactgttgatagcatcaataatgctttacct 85 N S L L K I A E E P P L N C H I A M T V D S I N N A L P 11578 acgcttgcaagtagagcaaaagttctaaccatgctaccttatactaatgaagagaaaatgcagtttgtcaagtcctacaagaag 113 T L A S R A K V L T M L P Y T N E E K M Q F V K S Y K K 11662 gtagatacttcaggaattgacgaccgagcgattgtagactattgcaatcttgccagcaatcttcaaatgcttgaagacatatta 141 V D T S G I D D R A I V D Y C N L A S N L Q M -L E_D . L" 11746 gaatatggcgcagaagagctatttgaaaaggttacaacattttatgacttaatatgggaggcaagtgctagtjaattcgctaaag 169 E Y G A E E L F E K V T T F Y D L I W E A S A S N S L K 11830 gttactaattggctcaaatttaaggaaactgatgaaggaaaaattgagcctaaacttttcctcaactgtcttttaaattggtcg 197 V T N W L K F K E T D E G K I E P K L F L N C L L N W S 11914 acagttgtcatcaggaagcactatgtagaaatgtctttcgaagaacttgaggcccatgaccttttagtgagggaagcatctagg 225 T V V I R K H Y V E M S F E E L E A H D L L V R E A S R
11998 tgtttgcgaaaggtatctaaaaagggctcaaatgcgcgtgtctgcgtgaacgaatttatcaggagggtcaaacaagttgagtga 12081
253 C L R K V S K K G S N A R V C V N E F I R R V K Q V E * dplORF018
35847 atggctagcagacagacgctattggtcgacggaattgaccttgtcgacaaaggtgcaaccgtgctagaatatgtaggactcact
1 M A S R Q T L L V D G I D L V D K G A T V L E Y V G L T
35931 ttcgcaggatttaaggactcaggatttaaaaaccctgaaggcatagacggagtattagattctccgtctaatgctatgtccgct
29 F A G F K D S G F K N P E G I D G V L D S P S N A S A
36015 cttactggaagcgtgaccttaatgttccacggagaaaccgaaaagcaagttaatcaaaaatacaggcagttcaaacaatttatt
57 L T G S V T L M F H G E T E K Q V N Q K Y R Q F K Q F I
36099 cgctcgaagtcattttggagaatttcgacacttgaagaccctggatactatcgaacgggaaaatttttaggagaaaccgagcaa
85 R S K S F W R I S T L E D P G Y Y R T G K F L G E T E Q
36183 ggaaaacttgtagacgttcaagcctttaaagatacttcccttgtagttaaattagggattcagttcaaagatgcttacgagtac
113 G K L V D V Q A F K D T S L V V K L G I Q F K D A Y E Y
36267 agcgactcaactgttcgaaaggtttataagtttcaacccgctttgggaggcgatagcttacctaacccaggaagacctactcga
141 S D S T V R K V Y K F Q P A L G G D S L P N P G R P T R
36351 caatttagagtagaaataagaactacttctcaaatcaaaggatattttcgaattggcgaaaaaagttcaggacagtttgttgag
169 Q F R V E I R T T S Q I K G Y F R I G E K S S G Q F V E
36435 ttcggtactaattcagtattgatggaaagtggctcgattattattctaaatcttggaacttttgaacttattaaaattagcagt
197 F G T N S V L M E S G S I I I L N L G T F E L I K I S S
36519 gcaaatcaagcgactaacttatttagatacattaaacgaggcgcattcttcaagattcctaatggaaattcaacaattaccatt
225 A N Q A T N L F R Y I K R G A F F K I P N G N S T I T I
36603 gaataccgagccgatgacgcagcagcttggacctctactcttcccgctcaagttgaactgtttctaaatccgtcttactattag 36686
253 E Y R A D D A A A W T S T L P A Q V E L F L N P S Y Y * dplORF019
12161 atgaatgtttatctcaatcaaatgggaaatgtagttcgagaaacttcggtttcaacagtctggaaaaccctcactcaaaaaggg
1 M N V Y L N Q M G N V V R E T S V S T V W K T L T Q K G
12245 ctcgtttctaatcatcgaatattcgctgttcgagatgataaggagtttctgtctaatgagtcgaggtggaaaaggcttccggat
29 L V S N H R I F A V R D D K E F L S N E S R K R L P D
12329 gttagatatgggacacttgttttgatggttactaaaattgacaagcgaagcaagttgctaaaggcctttcctgataattgtgtt
57 V R Y G T L V L M V T K I D K R S K L L K A F P D N C V
12413 gagtttgagaaaatgactgacgcgcagttgaaaaggcattttgtgtctaaatactcgactattgatagcgacatgattgacatg
85 E F E K M T D A Q L K R H F V S K Y S T I D S D M I D
12497 gttatccagttctgtctaaacgattactctagaattgacaatgaattggacaagctgtcgcgattgaaaaaggttgacgcatca
113 V I Q F C L N D Y S R I D N E L D K L S R L K K V D A S
12581 gtagttgaatccattgtcaagcacaagaccgaaattgacattttcagcctagttgatgatgtattggaatataggccggagcag
141 V V E S I V K H K T E I D I F S L V D D V L E Y R P E Q
12665 gcaattatgaaagtgactgaacttttagccaaaggagaaagtcctattggattgcttaccttgctttatcaaaattttaataac
169 A I M K V T E L L A K G E S P I G L L T L L Y Q N F N N
12749 gcttgtcttgtgctaggagccgatgagcctaaagaagccaatctaggcattaagcagttcttaatcaataagattgtctataac
197 A C L V L G A D E P K E A N L G I K Q F L I N K I V Y N
12833 tttcaatacgagctggactcagcctttgaaggcatggctattttaggtcaagctatcgagggcataaagaatggtcgctataca
225 F Q Y E L D S A F E G M A I L G Q A I E G I K N G R Y T
12917 gaaagttcagtggtctatatttctttgtataaaattttttcacttacttaa 12967
253 E S S V V Y I S L Y K I F S L T * dplORF020
1864 atggttaatcaatacaatcagcctgaaagaggcaagattcgaatcaatgttcgcgaccctgagaaaatgcctatcatggaaatt
1 M V N Q Y N Q P E R G K I R I N V R D P E K M P I M E I
1948 ttcggtcctacaattcaaggtgaaggaatggttataggtcaaaagactattttcattcgaactggtggatgcgactatcattgc
29 F G P T I Q G E G M V I G Q K T I F I R T G G C D Y H C
2032 aactggtgtgactcagcctttacctggaacggtactactgagccggaatatatcacaggcaaagaagctgctagtcgaatcttg
57 N W C D S A F T W N G T T E P E Y I T G K E A A S R I L
2116 aaactagctttcaatgataaaggtgaacagatttgtaaccacgtgacattgactggaggaaatcctgccttaatcaacgagcct
85 K L A F N D K G E Q I C N H V T L T G G N P A L I N E P
2200 atggctaagatgatttcgattctaaaagaacatggattcaagtttggtctcgaaactcaaggaactcgattccaagaatggttc
113 M A K M I S I L K E H G F K F G L E T Q G T R F Q E W F
2284 aaagaagtaagcgatatcactattagtcctaaaccgccttcaagtggaatgagaactaatatgaaaattcttgaagctattgta
141 K E V S D I T I S P K P P S S G M R T N M K I L E A I V
2368 gatagaatgaatgatgaaaaccttgactggtcatttaaaatcgttatctttgacgaaaatgacctagcttatgcgcgtgatatg
169 D R M N D E N L D S F K I V I F D E N D L A Y A R D M
2452 tttaaaactttcgaaggcaagttacgtccagtgaactacctttcagttgggaatgcaaacgcatacgaagaaggaaaaatcagt
197 F K T F E G K L R P V N Y L S V G N A N A Y E E G K I S
2536 gataggcttcttgaaaagttgggatggctttgggataaagtgtatgaagacccagctttcaacaatgttcgacctttaccgcaa
225 D R L L E K L G W L W D K V Y E D P A F N N V R P L P Q
2620 cttcatacacttgtttatgataataaaagaggagtataa 2658
253 L H T L V Y D N K R G V * dplORF021
2504 atgcaaacgcatacgaagaaggaaaaatcagtgataggcttcttgaaaagttgggatggctttgggataaagtgtatgaagacc
1 M Q T H T K K E K S V I G F K S W D G F G I K _C -f~ K T
2588 cagctttcaacaatgttcgacctttaccgcaacttcatacacttgtttatgataataaaagaggagtataaaatgaaaattgag
29 Q L S T M F D L Y R N F I H L F M I I K E E Y K M K I E
2672 catctagataaaatcggtaacgtattagggagagagaacggatgggcttcccttaagccggatgaaattgtaaccttggacaat
57 H L D K I G N V L G R E N G A S L K P D E I V T L D N
2756 actgaggcagccgttcaaagactttttggtctattaggcgaggacgcagaacgtgacgggttgcaagatactccattccgtttt
85 T E A A V Q R L F G L L G E D A E R D G L Q D T P F R F
2840 gttaaagcactcgctgaacataccgtagggtatcgagaagaccctaaacttcatctcgaaaaaacattcgacgtcgaccatgaa
113 V K A L A E H T V G Y R E D P K L H L E K T F D V D H E
2924 gaccttgttcttgtgaaagacattccattcaattctttatgtgagcatcatttagctccgttcgtagggaaggtgcatattgca
141 D L V L V K D I P F N S L C E H H L A P F V G K V H I A
3008 tacattcctaaggataagattacaggtctttcaaaattcggtcgagtggttgaaggatacgctaaacgacttcaagtacaagag
169 Y I P K D K I T G L S K F G R V V E G Y A K R L Q V Q E
3092 cgcttgactcaacaaatcgctgacgctattcaggaagttctaaatcctcaagcagttgcggtcatcgtagaggctgagcatact
197 R L T Q Q I A D A I Q E V L N P Q A V A V I V E A E H T
3176 tgcatgagcggacgcggtattaagaagcacggggcaacgacagtgacttcaactatgcgaggtcttttccaagatgacgcatct
225 C M S G R G I K K H G A T T V T Ξ T M R G L F Q D D A S
3260 gctcgagcagaattgcttcagttgattaaaaagtag 3295
253 A R A E L L Q L I K K * dplORF022
30896 atgagtaaagacattctttacggaatcaagctcgtgcaaatcgaggagcttgacccattgactcagttgccaaaagtcggcgga
1 M S K D I L Y G I K L V Q I E E L D P L T Q L P K V G G
30980 gctaactttgtcgtagatacggcagaaacagcagaactcgaagccgtgacctcggagggaactgaagatgtgaaacgcaatgac
29 A N F V V D T A E T A E L E A V T S E G T E D V K R N D
31064 acgcgcattcttgctatcgtgcgtactccagaccttttatacggttatgacttaacattcaaggacaacacgtttgaccctgaa
57 T R I L A I V R T P D L L Y G Y D L T F K D N T F D P E
31148 atcatggccctaattgaaggtggtacagtacgtcaacaaggcggaactattgctggatacgacaccccaatgcttgcacaaggt
85 I M A L I E G G T V R Q Q G G T I A G Y D T P M L A Q G
31232 gcttctaatatgaaaccatttagaatgaacatctatgtgccaaactatgtaggtgactcaattgtcaactacgtgaaaatcact
113 A S N M K P F R N I Y V P N Y V G D S I V N Y V K I T
31316 ttgaataactgtaccggtaaagctccagggctttcaatcgggaaagagttctacgctcctgagttcaacatcaaggcacgtgaa
141 L N N C T G A P G L S I G K E F Y A P E F N I K A R E
31400 gcaaccaaagcaggtttgccagttaagtcaatggactatgtggcacaacttccagcggttcttcgtcgcgtgacattcgatttg
169 A T K A G L P V K S M D Y V A Q L P A V L R R V T F D L
31484 aacggtggaacaggaaccgccgacgcagttcgagttgaagcaggtaagaagatttctccaaaaccagttgaccctaccttaaca
197 N G G T G T A D A V R V E A G K K I S P K P V D P T L T
31568 ggtaaggctttcaaaggctggaaagttgaaggagaatcaactatttgggacttcgacaaccacatgatgcctgaccgagacgtc
225 G K A F K G W K V E G E S T I W D F D N H M M P D R D V
31652 aaactcgtagcacaatttgcatag 31675
253 K L V A Q F A * dplORF023
6419 atggccaagtccaatttaactagaattgcaaagatggttagagcaggaaacagtgaaggtcctgcttcatcttttgtcaattcg
1 M A K S N L T R I A K M V R A G N S E G P A S S F V N S
6503 ctgacccgggttattgaacgaactcagcctgaatataatccttcgacatattataagcccagcggggttggtggatgtattcga
29 L T R V I E R T Q P E Y N P S T Y Y K P S G V G G C I R
6587 aaaatgtatttcgaaagaatcggtgagtctattatagataacgcagattctaacctaattgcaatgggcgaagctggaacattt
57 K M Y F E R I G E S I I D N A D S N L I A M G E A G T F
6671 aggcacgaagttctccaagagtacatggttaaaatggctgaaatcgatgaggactttgaatggttgaatgtagcagagttcttg
85 R H E V L Q E Y M V K M A E I D E D F E L N V A E F L
6755 aaagaaaatccagttgaaggaactatcgtcgacgagcgtttcaagaaaaacgattatgaaacgaagtgtaagaacgaacttctt
113 K E N P V E G T I V D E R F K K N D Y E T K C K N E L L
6839 caactttcattcttgtgtgacggactagttcgatataaaggcaagctctacattttagagattaagactgaaaccatgttcaag
141 Q L S F L C D G L V R Y K G K L Y I L E I K T E T M F K
6923 ttcactaaacatactgagccctatgaagaacacaagatgcaagcaacttgctacggaatgtgtctaggagtcgatgatgtcatt
169 F T K H T E P Y E E H K M Q A T C Y G M C L G V D D V I
7007 ttcctttatgaaaatcgagataacttcgaaaagaaagcctacacgtttcacatcacagacgagatgaaaaatcaagtccttgga
197 F L Y E N R D N F E K K A Y T F H I T D E M K N Q V L G
7091 aaaattatgacctgcgaagagtatgtagagaaaggcgaaagtcctaaaatctattgctcttcagcctattgcccatattgtaga
225 K I M T C E E Y V E K G E S P K I Y C S S A Y C P Y C R
7175 aaggaaggtcgaaatctgtga 7195
253 K E G R N L * dplORF024
25992 atgaacgcagtagatggccaggtagttcatattctacaagtattagcagaagatggaaatgctacggctgaaaagttcgaaaag
1 M N A V D G Q V V H I L Q V L A E D G N A T A E K F E K
26076 gaagtcagggctgcatctttagtattttcacgaagagcagccgaggcagttgtcaaaggtgaaatctataaggacggcaaaaac
29 E V R A A S L V F S R R A A E A V V K G E I Y K D G K N
26160 ctctcgaaacgtgtttggtcttcagccgcacgcgcaggaaatgatgttcaacaaatagtcacacaaggcctagcaagtggaatg
57 L S K R V W S S A A R A G N D V Q Q I V T Q G L A S G M
26244 tctgctacagatatggctaaaatgctcgagaaatatatcgaccctaaggttcgaaaagattgggactttgataagatagctgag
85 S A T D M A K M L E K Y I D P K V R K D W D F D K I A E
26328 aagctagggaaacctgctgctcataaatatcaaaatctcgaatacaatgcccttcgacttgctcgaactaccattagccattcc
113 K L G K P A A H K Y Q N L E Y N A L R L A R T T I S H S
26412 gccacagctggagtgagacaatggggcaaggttaatccttatgctcgaaaagttcaatggcattctgttcacgctccaggtcga
141 A T A G V R Q W G K V N P Y A R K V Q H S V H A P G R
26496 acgtgtcaagcgtgtatcgatttagatggtgaagtatttcctatcgaagaatgtcctttcgaccatcctaatggaatgtgctac
169 T C Q A C I D L D G E V F P I E E C P F D H P N G M C Y -
26580 caaactgtatggtacgaaaactcactcgaagaaatcgctgatgagttgagaggctgggtagacggagaacctaaigatgtat a
197 Q T V Y E N S L E E I A D E L R G W V D G E P - D V L
26664 gacgaatggtacgacgatttaagttcaggaaaagttgagaaatacagcgacctcgactttgttaaaagttattag 26738
225 D E W Y D D L S S G K V E K Y S D L D F V K S Y * dplORF025
18778 atggcaaagaacaaaaagcgaaaaaaagtaaatgtcaaaaggaaaatgcttatccctacaaatctctcgaaaaaagtaaatgta
1 M A K N K K R K K V N V K R K M L I P T N L Ξ K V N V
18694 aaagcaatcgcttatagaaaagtcactgttaagtggctgcctaatacagatgaaattcaagtatatttcgacctttatataaat
29 K A I A Y R K V T V K W L P N T D E I Q V Y F D L Y I N
18610 aaaaacaggctgacaatgttaggcactattgacccggacaagagctattttgaaggaattaggattgtttgtaagaaacctcag
57 K N R L T M L G T I D P D K S Y F E G I R I V C K K P Q
18526 ccttggatgactgttaaggagctccaggttgcgcgtgcagacgccccaggtttttttgcagttcttaaagcctattgtcacacg
85 P W M T V K E J. Q V A R A D A P G F F A V L K A Y C H T
18442 gttggcgatgtactagatagcggagcagagcctactgaaattgttcaaggtattatgtataaagacggtgaactatttaaggac
113 V G D V L D Ξ G A E P T E I V Q G I M Y K D G E L F K D
18358 agtgaaattgtcagccttttcaaatacgatgtcaaagagccttatgagtttccaaaggaccttcctataaccttggacaacttt
141 S E I V S L F K Y D V K E P Y E F P K D L P I T L D N F
18274 ttagagttcattatgtctagccagcatactagagcacttgttttgcgttgtgctaatataggtgagttttccaagaattggcgg
169 L E F I M S S Q H T R A L V L R C A N I G E F S K N R
18190 aaatggcaaaaagctatccagctcctgctcgactatgccaaggcggatgactttaaagtagacgaaactgtttgggacttttca
197 K Q K A I Q L L L D Y A K A D D F K V D E T V D F S
18106 cccggctctaaagctggaaaggtagcacgtcgtaaaggctatgaggcaattcaacaagcccttgagcagataaataaataa 18026
225 P G S K A G K V A R R K G Y E A I Q Q A L E Q I N K * dplORF026
21512 atggcgaaagctactggaccaaaagttcgaagaggaaaaactcctccacggccaaaagacaaaaaaggaatcaaagcaaatgcg
1 M A K A T G P K V R R G K T P P R P K D K K G I K A N A
21596 cgtgtcaataaagaccagttcgtagagtatgactataaaggcatcaagatgacaattaaggaacgtgatgctagaatgaaattg
29 R V N K D Q F V E Y D Y K G I K M T I K E R D A R M K L
21680 gaatttattagaggcatgactattcaggaaattgcagcccgctatggattaaatgaaaagcgtgttggcgaaatacgggctcgc
57 E F I R G M T I Q E I A A R Y G L N E K R V G E I R A R
21764 gataaatgggtgaaggctaagaaagagttcgagaatgaaaaggctcttgttactaatgatacattgactcaaatgtatgcaggg
85 D K W V K A K K E F E N E K A L V T N D T L T Q M Y A G
21848 tttaaagtctcagtcaatattaaatatcacgccgcctgggagaaactaatgaacatcgtcgaaatgtgtttagataatcctgac
113 F K V S V N I K Y H A A W E K L M N I V E M C L D N P D
21932 agatatttatttactaaagaaggaaatattagatggggcgcattagatgtcctttcgaaccttatagatagagctcaaaaagga
141 R Y L F T K E G N I R G A L D V L S N L I D R A Q K G
22016 caagaaagagcgaatggaatgcttccggaagaggttcgatatagactacaaattgagcgcgagaaaattacattgctccgggcc
169 Q E R A N G M L P E E V R Y R L Q I E R E K I T L L R A
22100 aaaatgggcgaccaggaaattgaaggcgaggttaaagataacttcgtagaagcactagataaagcagctcaagccgtttggcaa
197 K G D Q E I E G E V K D N F V E A L D K A A Q A V W Q
22184 gaatttagtgacgcaacaggttcctacattaaaggagtgactgataatgacaataagcctgagaaataa 22252
225 E F S D A T G S Y I K G V T D N D N K P E K * dplORF027
52762 atgggaaaagtatcaattcaaaaatcaggaacatttagctcagggtctaataacgagtttttcacactcgctgaccacggtgac
1 M G K V S I Q K S G T F S S G S N N E F F T L A D H G D
52846 agcgcaattgtcactctattgtatgatgacccggaaggcgaagacatggattatttcgtagtccacgaagcagacgttgacggt
29 S A I V T L L Y D D P E G E D M D Y F V V H E A D V D G
52930 cgtcgacgctatatcaattgcaatgctattggcgaagacggggaaacagtccatcctgataattgtccattatgccaaaacgga
57 R R R Y I N C N A I G E D G E T V H P D N C P L C Q N G
53014 ttccctcgtattgaaaaactatttcttcaactttacaaccatgatacgggaaaagttgaaacatgggaccgaggccgttcttat
85 F P R I E K L F L Q L Y N H D T G K V E T W D R G R S Y
53098 gttcaaaagattgttacatttatcaataaatatggaagccttgtgactcagccttttgaaattattcgttcaggagctaaaggt
113 V Q K I V T F I N K Y G S L V T Q P F E I I R S G A K G
53182 gaccaacgaactacttatgaattccttccagagcgtccggaagacagtgctactcttgaagattttccagaaaagagcgaactt
141 D Q R T T Y E F L P E R P E D S A T L E D F P E K S E L
53266 cttggaactctaattttagacctcgacgaagaccaaatgtttgacgtggttgacggcaagttcactcttcaagaagagcgttct
169 L G T L I L D L D E D Q M F D V V D G K F T L Q E E R S
53350 tcaagtcgttcaaattcacgtagaggagcatctcctgcgcctagacgaggttccggtcgagaatcttcacaaggtcgaacagct
197 S S R S N S R R G A S P A P R R G S G R E S S Q G R T A
53434 gaaagaactccttcagttagtcgaagaactcctccaacacgaggtcgaggattctaa 53490
225 E R T P S V S R R T P P T R G R G F * dplORF028
44595 atgtcaaaaattaaattcgaaaaccttaaaaaaggcgatgttgtgctacgagctaaatctcaaacgaagtttaaaatcgtttca
1 S K I K F E N L K K G D V V L R A K S Q T K F K I V S
44679 attttagcagacgaaaagaaagcagaccttgaatcattagaagacggaggtgaacttcacctttcagcttcaactctcgaacgt
29 I L A D E K K A D L E S L E D G G E L H L S A S T L E R
44763 tggtacacaatggaagatgaaactgaacctaaaaaagaagaagctgctaaacctgctaaaaaggctgctcctgcagttgctcga
57 W Y T M E D E T E P K K E E A A K P A K K A A P A V A R
44847 cctgctcgaaaaggtagagtcgttcccaaacctaaaaaagaagtccttgaggaagaaattcctgaagttaaggaacagccggaa
85 P A R K G R V V P K P K K E V L E E E I P E V K E Q P E
44931 gaagttggttcagttagtgagaaatctactgttcgaaaacctgctcctaaaaaagaaagcgtgatggcgattactaaggctctt
113 E V G S V S E K S T V R K P A P K K E S V M A I T K A L
45015 gaaagtcgaattgttgaagcctttcctgcgtctactcgaatcgtcactcagtcttacatcgcctatcgctctaagaagaacttc
141 E S R I V E A F P A S T R I V T Q S Y I A Y R S K K N F
45099 gttactatcgaagaaactcgaaaaggtgtttctattggagttcgcgcaaaagggttgacagaagaccaaaagaaacfrtcttgca
169 V T I E E T R K G V S I G V R A K G L T E D Q K_-K L L A
45183 tctattgctcctgcatcttacgaatgggcgattgacggaatttttaaactcgtcaaggaagaagatattgacaccgcaatggaa
197 S I A P A S Y E A I D G I F K L V K E E D I D T A M E
45267 ttgattgaagcttctcacctttcttcgctatga 45299
225 L I E A S H L S S L * dplORF029
662 atgaaatcagtagttttattatccggcggagtcgactcagccacttgtttagcaattgaagttgacaagtggggttctaaaaat
1 K S V V L L S G G V D S A T C L A I E V D K W G S K N
746 gttcatgctatagcattcaattacggacaaaagcatgaagcagaacttgaaaatgctgctaatgttgcaatgttctacggagtc
29 V H A I A F N Y G Q K H E A E -J E N A A N V A M F Y G V
830 aagttcaccattcttgaaattgactcgaaaatctactcaagctctagctcttccttattacaaggaaaaggcgaaatttcacat
57 K F T I L E I D S K I Y S S Ξ S S S L L Q G K G E I S H
914 ggaaaatcttacgctgaaatcctagcagagaaggaagtagttgacacctatgttccatttagaaatggactaatgctttcacag
85 G K S Y A E I L A E K E V V D T Y V P F R N G L M L S Q
998 gctgcggcttatgcttattcggttggagcttcttacgtcgtatatggtgctcacgcagacgatgcggctggaggtgcttaccct
113 A A A Y A Y S V G A S Y V V Y G A H A D D A A G G A Y P
1082 gattgcactcctgagttctataattcaatgtcaaatgcaatggaatatggaactggaggcaaggtaacccttgtcgctcctcta
141 D C T P E F Y N S S N A M E Y G T G G K V T L V A P L
1166 cttactctaaccaaggcgcaagtcgttaaatggggaattgatttagatgttccttatttcttgactcgttcatgttatgaaagt
169 L T L T K A Q V V K W G I D L D V P Y F L T R S C Y E S
1250 gacgctgaaagttgtggaacttgcgcaacttgtatcgaccgcaaaaaggcattcgaagaaaatggaatgactgaccctattcat
197 D A E S C G T C A T C I D R K K A F E E N G M T D P I H
1334 tataaggagaattga 1348
225 Y K E N * dplORF030
20088 atgaataacgaaaaaattattgaaaaaattaaaaat ttattcaattagcaaatgacaacccgagtgacgaagaggggcaaact
1 M N N E K I I E K I K N L I Q L A N D N P Ξ D E E G Q T
20004 gcccttcttatggctcaaaagttgatgctaaagaataatatcgcacttgctcaagttgaacaatttgatgaacctaaacagttc
29 A L L M A Q K L M L K N N I A L A Q V E Q F D E P K Q F
19920 gagacttctcaagctgttgggaaagaagcaggtcgaatattttggtgggaacgtgaacttggtcatattctcgcgactaatttt
57 E T S Q A V G K E A G R I F W W E R E L G H I L A T N F
19836 aggtgcttttgtattaatcagcgtgatatgcgcttgaataaaagtcgaataattttcttcggcgaaaaacaagacgctgaatta
85 R C F C I N Q R D M R L N K S R I I F F G E K Q D A E L,
19752 gtgtctaaaatatatgaggctgctttgctttatcttcgttaccgtattgaccgacttcctactcgcgaaccttcctacaagaat
113 V S K I Y E A A L L Y L R Y R I D R L P T R E P S Y K N
19668 tcatacctcaaaggctttttgtcagccttagccattcgatttaaaaagcaggtggaagaatattcacttatggtcctacctagc
141 S Y L K G F L S A L A I R F K K Q V E E Y S L M V L P S
19584 gagcaaacaaaaaatgcgcttcaggacacatttcgaaatttaaagaaggaaggaattgacagacctcaacatgacttcaatctt
169 E Q T K N A L Q D T F R N L K K E G I D R P Q H D F N L
19500 gaagcgtatattgaagggcggtttcatggcgagaatgcaaagattatgcccgatgaaattttggaaggcggtaactaa 19423
197 E A Y I E G R F H G E N A K I M P D E I L E G G N *
dplORF031
26943 atggcttatcaattagaagacttgttaaaaggtctagatgaaccaactatcaaacaggtgaaggaaattatttcgaaaacttcg
1 M A Y Q Ii E D L L K G L D E P T I K Q V K E I I S K T S
27027 aaagaactcgatgctaaaattttcattgacggcgacggtcaacattttgtacctcacgcacgtttcgatgaagttgttcaacag
29 K E L D A K I F I D G D G Q H F V P H A R F D E V V Q Q
27111 cgcgatgcagctaacggctcaattaattcttataaagaacaagtcgcgacgctttctaaacaggtcaaagataacggtgatgcg
57 R D A A N G S I N S Y K E Q V A T L S K Q V K D N G D A
27195 cagaccactatccaaaaccttcaagagcaactcgacaagcagtctcaacttgcaaaaggcgctgtgattacttcagctcttcat
85 Q T T I Q N L Q E Q L D K Q S Q L A K G A V I T S A L H
27279 ccgttgattagtgactccattgctccagcagcagacattcttggatttatgaaccttgacaacattacggtcgaaagtgacggt
113 P L 1 S D S I A P A A D I L G F M N L D N I T V E S D G
27363 aaagttaaaggtcttgatgaagagttgaaagctgttcgtgagtctcgtaaatacttattcaaagaagtcgaagttcccgcagaa
141 K V K G L D E E L K A V R E S R K Y L F K E V E V P A E
27447 caagaggctcaagctaagtcgccagccgggactggaaatttaggaaatccaggtcgtgtcggtggtggtgttcccgaacctcgt
169 Q E A Q A K S P A G T G N Ii G N P G R V G G G V P E P R
27531 gaaatcggctcttttggtaagcaacttgctgctgctcaacaaacggcaggagcacaagaacaatcatcattctttaaataa 27611
197 E I G S F G K Q L A A A Q Q T A G A Q E Q S S F F K * dplORF032
52033 atgaaagaagcgaatagactagtttctagctatgtaggattcgaatgctggactgacgaagaatgtatcaggaactttgaacta
1 K E A N R L V S S Y V G F E C W T D E E C I R N F E L
52117 gaccctgatatgtcaattgcgtctgc tatcatcgttattttgggatgctttattcctatgcaaaaaggtttaaatgcttatct
29 D P D M S I A S A Y H R Y F G M L Y S Y A K R F K C L S
52201 cgacatgacattgaaagcattgcattcgagactatttcaaaatgtttggcaacgttcaaatcaaaccaaggggccaagttttca
57 R H D I E S I A F E T I S K C L A T F K S N Q G A K F S
52285 acttaccttacaagactcttcaagaatagaatagtcttagaatataggtacctaaatgcaccttccatgaatcgaaattggtat
85 T Y L T R L F K N R I V L E Y R Y Ii N A P S M N R N Y
52369 gtagaagtgacgttcgatagcgtttcgacaaatgaagaaggcgacgattttagtatcctatcgacagttggctattgtgaagac
113 V E V T F D S V S T N E E G D D F S I L S T V G Y C E D
52453 tacggaaaaattgaaattgaagcaagtcttgacttcatgacgctttctaatacagagtatgcttatatctcgtctgtcattcaa
141 Y G K I E I E A S L D F M T L S N T E Y A Y I S S V I Q
52537 aacggtccttcagtaagcgacgcagaaattgcgcgtgaaattggagtaagσaggtctgctattagtcagtctaagaagtcacta
169 N G P S V S D A E I A R E I G V S R S A I S Q S j_ R-- S* L
52621 aaaaataaattaaaagattttatataa 52647 _r
197 K N K L D F I * dplORF033
7670 atggcaagacctaagttacctcaaattgatattcgagaagaagaaatacgagatgctcaagacgtagcagactcgtatggtgcg
1 M A R P K L P Q I D I R E E E I R D A Q D V A D S Y G A
7754 attatcaataaagtagtcgacgaaattgttgaagcagcttgcggttcacttgaccaggcaatggaagaaattcaaatagttgta
29 I I N K V V D E I V E A A C G S L D Q A M E E I Q I V V
7838 agccaaaatcctgtcattatggaagaccttaactactacattggctatcttcccactcttctttatttcgccgcagatagggcg
57 S Q N P V I M E D L N Y Y I G Y L P T Ii li Y F A A D R A
7922 gaaatggtgggaatacaaatggattcaagttctgctatcaggaaagaaaaatacgataatctatacattttagccgccgggaaa
85 E M V G I Q M D S S S A I R K E K Y D N L Y I L A A G K
8006 actattcctgacaagcaagcagaaactcgaaaacttgtcatgaatgaagaagtcatcgaaaatgcttacaagcgagcctacaag
113 T I P D K Q A E T R K L V N E E V I E N A Y K R A Y K
8090 aaagttcaattaaagctagaacaggccgataaggtattagcatctttaaaacgaattcaaacctggcaactagcagagttagaa
141 K V Q L K L E Q A D K V L A S -i K R I Q T W Q Ii A E L E
8174 actcagtcaaataattcaaaaggagtattattaaatgcaaaaagacgtagacgtgaaaatgattga 8239
169 T Q S N N S K G V L L N A K R R R R E N D * dplORF034
131 atgagtcaaaacactacacgcactgacgctgaattgacaggcgttactcttttaggaaaccaagacaccaaatacgattatgac
1 M S Q N T T R T D A E L T G V T L L G N Q D T K Y D Y D 215 tataatccagacgtccttgaaactttccctaacaaacatcctgaaaataattacctagtaacatttgacggatatgaattcact 29 Y N P D V L E T F P N K H P E N N Y L V T F D G Y E F T
299 tccctttgccctaaaacaggacagcctgacttcgcgaatgttttcattagttacattccaaacgaaaagatggttgaatctaaa 57 S L C P K T G Q P D F A N V F I S Y I P N E K M V E S K 383 tcattgaaattgtacttattcagtttccgtaaccacggtgacttccacgaagattgcatgaacattattttgaatgacttgtat 85 S L K L Y L F S F R N H G D F H E D C M N I I L N D L Y 467 gaattgatggaacctaagtacattgaagtcatgggcctattcactcctcgtggtggaatttcaatttacccattcgtcaacaaa 113 E L M E P K Y I E V M G L F T P R G G I S I Y P F V N K 551 gtgaatcctcaatttgcaactcctgaacttgaacagcttcaacttcaacgcaaattgaacttccttggaaatgttcaaggtctt 141 V N P Q F A T P E L E Q L Q L Q R K L N F L G N V Q G L 635 ggacgagctattcgatag 652
169 G R A I R * dplORF035
17425 atgcacctaatgaaggattcgaagatgttgaggacatggaagtccttagcattcgagttcgaaacgaaggtgaggacgacgagt
1 M H L M K D S K M L R T W K S Ii A F E F E T K V R T T S
17341 gggttgaagttatcgcctgctatgaaaacgatgacgaggacgaagatttggaagggttataaaatgaaggtatttatcaacaat
29 G L K L S P A M K T M T R T K I W K G Y K M K V F I N N
17257 catactgaagctgatattgactacaaagatattctaaattttgtagcttatcgaaactctcctaaccctcaaattcaaatcact
57 H T E A D I D Y K D I Ii N F V A Y R N S P N P Q I Q I T
17173 agctggaacgctttgctttcctgctatacacggaatgagctttcttataaaggagtttcaataacggacttttttgaagccatt
85 S W N A L L S C Y T R N E Ii S Y K G V S I T D F F E A I
17089 caaactattgcaagttccttcactcacctagactcgaaaacaattgatacacaaaatgaaaagcgactcgaaaggattgaggaa
113 Q T I A S S F T H L D S K T I D T Q N E K R Ii E R I E E
17005 cttcagtcaagaataggtcattgtaactgtactatcgacgaacttaaaaaaggagtccacgaaatgccggatattgaatcagct
141 L Q S R I G H C N C T I D E L K K G V H E M P D I E S A
16921 atttcttaccagtacggacagattcttgcttatgaagatgaacttaattttctgctaaactaa 16859
169 I S Y Q Y G Q I Ii A Y E D E L N F L Ii N * dplORF036
48808 gtgttagtcgaacgaaaagccgacaaggaatgttgggaatggctagaagctgttcgagcaaatatagtcgaagaagttcgaaac
1 V L V E R K A D K E C W E W L E A V R A N I V E E V R N
48892 ggtcttagcattgttattgcttcgaatactgtcgggaatgggaaaactagctgggcggttcgacttttgcaacgctatttagca
29 G L S I V I A S N T V G N G K T Ξ W A V R L L Q R Y L A
48976 gaaactgcacttgacggaagaattgttgagaaaggaatgtttgtagtgtcagctcaactattgactgagttcggcgactataat
57 E T A Ii D G R I V E K G M F V V S A Q L L T E F G D Y N
49060 tattttcaaaccatgcaagaatttctcgaacgtttcgagcgccttaagacttgtgagctattagtcatagacgaaataggtgga
85 Y F Q T M Q E F L E R F E R L K T C E L L V I D E I G G
49144 ggttccttaaccaaggcctcttatccttatctgtatgacttggttaattatagggttgacaataacttgtcgactatttatacg
113 G S L T K A S Y P Y Ii Y D L V N Y R V D N N L S T I Y T
49228 actaattatactgacgatgaaattattgaccttttaggccaaaggctttatagtcgtatatatgatacttcagtggttctagat
141 T N Y T D D E I I D L L G Q R L Y S R I Y D T S V V L D
49312 tttcaggcaagcaatgtaagaggattggaggtaagcgaaattgaatcatag 49362
169 F Q A S N V R G L E V S E I E S * dplORF037
55855 atggtgaagaaattgaaatctaaaatctattcagttgcatatataattctagtagttattgcgaaccttgtgacaatttatttc
1 M V K K L K S K I Y S V A Y I I L V V I A N L V T I Y F
55939 gaacctttaaatgtgaaaggaattttaattcctccaagcagttggtttatgggattcactttcctgcttataaatctaataagc
29 E P L N V K G I L I P P S S W F M G F T F Li L I N L I S
56023 aagtacgagaagccaaaatttgcaggttctttgatatgggtagggttattccttacctcgttgatttgctttatgcaaaaccta
57 K Y E K P K F A G S L I V G L F L T S L I C F M Q N L
56107 ccacaatcgcttgtcgtggcttcaggagttgcattttggataagtcaaaaagcaagtgtctttatat cgacaagctctcgaat
85 P Q S L V V A S G V A F W I S Q K A S V F I F D K L S N
56191 aaattagactcgaagattgcaaatgctttgtctagcaacatcggttctattatagacgcaaccatatggatttcattaggactg
113 K L D S K I A N A L S S N I G S I I D A T I W I S L G L
56275 agtcctcttggaattggaacggttgcatatatagatattccgtcagccgtactaggccaagttctagttcagtttatcttgcag
141 S P L G I G T V A Y I D I P S A V L G Q V L V Q F I L^Q
56359 tcaattgcttcgagatatttgaaaaagtag 56388 - —
169 S I A S R Y L K K * _, dplORF038
1350 atgagagtttctaaaaccttaacattcgacgcagctcatcaactagttggacattttggaaaatgcgcaaatttgcacgggcat
1 M R V S K T L T F D A A H Q L V G H F G K C A N L H G H
1434 acttacaaagtcgaaatttcattagcaggcggaacttatgaccacggttcgagtcaagggatggttgttgacttttatcacgtc
29 T Y K V E I S L A G G T Y D H G S S Q G V V D F Y H V
1518 aagaaaatcgcaggtacattcattgacagacttgaccacgctgttcttcttcaagggaatgaaccaatcgctttagcaaatgca
57 K K I A G T F I D R L D H A V L Ii Q G N E P I A L A N A
1602 gttgacaccaagcgagttctatttggatttagaactacggctgagaatatgtcaagattccttacctggactctcacggagctt
85 V D T K R V L F G F R T T A E N S R F L T T L T E L
1686 atgtggaagcatgctcgtatcgactctatcaaactatgggaaactcctacaggttgcgcagaatgtacttactacgagattttc
113 M W K H A R I D S I K Ii W E T P T G C A E C T Y Y E I F
1770 acagaagacgagattgaaatgttcaagaacgtaacctttatcgacaaagacgaaaagattactgtccgcgaaattttagagcag
141 T E D E I E M F K N V T F I D K D E K I T V R E I L E Q
1854 gagcaggataatggttaa 1871
169 E Q D N G * dplORF039
3306 atgaataaaagtgcaaccttttggcttgttcgaacagctcttattgcggctctatatgtgacattgaccgttgcattttctgct
1 M N K S A T F W L V R T A Ii l A A L Y V T L T V A F S A
3390 attagttatggacctattcaatttagagtcagtgaagccttgattcttctacctttatggaaccatagatggactccggggatt
29 I S Y G P I Q F R V S E A L I L L P L W N H R W T P G I
3474 gtattaggaacaattattgcaaacttcttttcacctcttggactgattgacgttttattcggttcacttgctaccttccttgga
57 V L G T I I A N F F S P L G -i l D V Ii F G S -i A T F Ii G
3558 gtagtggcaatggtgaaagttgctaagatggcaagtcctctatattcacttatctgtccagttcttgctaatgcttaccttatt
85 V V A V K V A K M A S P L Y S Ii l C P V L A N A Y L I
3642 gcgctggaacttcgaatagtttactctttacctttttgggaatctgtcatctatgtaggaattagtgaagcgattatcgtttta
113 A L E L R I V Y Ξ L P F E S V I Y V G I S E A I I V L
3726 atttcatacttccttatttccacgctggcgaagaacaatcattttagaacactgataggagcgaaaaatgggatttaa 3803
141 I S Y F L I S T L A K N N H F R T L I G A K N G I * dplORF040
7192 gtgagctatactggaaaaatgttcgaggaagactttttcgaaggtgcaaaagactttgagaaagatgctttcacggtccgtcta
1 V S Y T G K M F E E D F F E G A K D F E K D A F T V R L
7276 tatgataccactaatggatttcgaggagttgcaaatccctgcgattatatagccgcaactaactttgggaccttgtttattgaa
29 Y D T T N G F R G V A N P C D Y I A A T N F G T L F I E
7360 ctgaaaactactaaagaagcttctttgagctttaataacatcactgataatcaatggttccagctatcacgcgcagatggatgc
57 L K T T K E A S L S F N N I T D N Q W F Q L S R A D G C
7444 aaatttattctcgccggaattttagtgtatttccaaaagcatgaaaagattatatggtatccaatttcaagccttgaaaaaatt
85 K F I L A G I L V Y F Q K H E K I I W Y P I S S L E K I
7528 aaacggtctggagttaaaagcgtcaacccaaacttcatcgatgcagggtatgaagtttcttacaagaagcgtcgaactagattg
113 K R S G V K S V N P N F I D A G Y E V S Y K K R R T R L
7612 accattcctttccaaaatgttctagatgcagttgagcttcattacaaggagaaaagcaatggcaagacctaa 7683
141 T I P F Q N V L D A V E Ii H Y K E K S N G K T * dplORF041
8208 atgcaaaaagacgtagacgtgaaaatgattgaccctaaacttgaccgattaaaatacacaggtgattgggttgatgtacgaatt
1 M Q K D V D V K M I D P K L D R L K Y T G D W V D V R I
8292 agttctatcactaaaattgacgccgacagcgccgatgtctcaagatgtcgaaaagtgcttcaaaaggctcaagtatattcagtg
29 S S I T K I D A D S A D V S R C R K V L Q K A Q V Y S V
8376 gcggcaggtgaatgcattaaaattgcacacggatttgctcttgaacttcctaagggatatgaagcaatcttgcatcctcgttcc
57 A A G E C I K I A H G F A L E Ii P K G Y E A I -i H P R S
8460 agtctttttaagaaaactggtctaatcttcgtttctagcggagtgattgacgaaggttacaaaggtgacactgatgaatggttc
85 S L F K K T G L I F V Ξ S G V I D E G Y K G D T D E F
8544 tcagtttggtatgctactcgtgacgcagatatcttctacgaccaaagaattgcccaatttagaattcaggaaaagcaacctgct
113 S V W Y A T R D A D I F Y D Q R I A Q F R I Q E K Q P A
8628 atcaagttcaatttcgtagaatctttaggaaatgcggctcgtggaggccatggaagtacaggtgatttctaa 8699
141 I K F N F V E Ξ L G N A A R G G H G S T G D F * dplORF042
48082 gtggcaaggcaaagaataggcaattcaggaaagcctaaaaatgaaattgaactaacattcaaagacaagcctaaaactcgttct
1 V A R Q R I G N S G K P K N E I E L T F K D K P K T R S
48166 accttattcaagaaggacgtggcaacaggtctttcaaaagtcgagcatgattattttcaaatagttgaagcacttaacggaaaa
29 T L F K K D V A T G Ii S K V E H D Y F Q I V E A L N G K
48250 caattcgaacctaatatgaagcaggtgtcatctttctttatagttcagtatgaatttattttcaatattaagtgcatcgattat
57 Q F E P N M K Q V S S F F I V Q Y E F I F N I K C I D Y
48334 aactggttcaacttttcgagcactatgaaaaatgttcgaacttatttaaacattgagtcgaacattgaactttgtcgattttta
85 N W F N F S S T M K N V R T Y L N I E S N I E L C R F L
48418 gctgaaagttttgttaaatatgaaaatgttcgaaaaagattgaacctaagcgaaaggttcataacggtctcgactttcaaaaga
113 A E S F V K Y E N V R K R L N L S E R F I T V S T F K R
48502 gcctggattttggacgaactcgaaggaaaaacgggttcaaaattcgaaggattttattag 48561
141 A I L D E L E G K T G S K F E G F Y * dplORF043
31699 atgactaatattatcacagctgagcagtttaagcaacttgcatttcaaatcatcgcacttccaggattttcaaaaggtagtgaa
1 M T N I I T A E Q F K Q L A F Q I I A L P G F S K G S E
31783 cctatccatgttaaaattcgagcagcaggtgtcatgaacctaatcgctaacgggaaaatccctaatacgcttttaggtaaagtg
29 P I H V K I R A A G V M N L I A N G K I P N T Ii L G K V
31867 acagaactgtttggagaaacttcgacagtcactaaagacaatgctagtctagcatcaattactgaccaacagaagaaagaagcg
57 T E L F G E T S T V T K D N A S L A S I T D Q- Q K K"— E A
31951 ctcgaccgattgaacaaaaccgataccggtattcaagacatggctgaacttcttcgagtattcgcagaagθttcaatggtagag
85 L D R L N K T D T G I Q D M A E L L R V F A E A S M V E
32035 cctacttacgctgaagtcggcgagtatatgacagatgagcaacttatgacaatcttcagtgcaatgtacggtgaagtgactcaa
113 P T Y A E V G E Y M T D E Q L M T I F S A M Y G E V T Q
32119 gctgaaacctttcgtacagacgaaggaaatgtctaa 32154
141 A E T F R T D E G N V *
dplORF044
25666 atggtaagtgttttgattagcagcagctcctttttgaagttcctgcttcattttagctcgacaagtatttctaaatcgaataag
1 M V S V L I S S S S F L K F L L H F S S T S I S K S N K
25582 gttttcaatttccttgtttcctacataagtggtgaaccgataatggcacttaggacattcgaagaatctccactctacgccctt
29 V F N F L V S Y I S G E P I M A L R T F E E S P L Y A L
25498 ttcgatatgtttcgaaataatctgtttagatgtaaggtcgaacttatgctcacaatggtcacaattaaccttgaacgtctgggt
57 F D M F R N N L F R C K V E L M -i T M V T I N Ii E R L G
25414 cgactccttcttcggttggttgttcagtttgttctttttctttgtcatcaacttcgtcttcttcactcgtttcatcttgaggct
85 R L Ii li R L V V Q F V L F L C H Q L R L Ii H S F H L E A
25330 cctcttgttcgtttaattcgtttgctaatacaggcaatgctccagctgagatttcgtcaagctgagcaagttcttccaaaatgc
113 P L V R L I R L L I Q A M L Q L R F R Q A E Q V L P K C
25246 gttcccattccttgtccgccttttccttcttactga 25211
141 V P I P C P P F P S Y * dplORF045
25340 atgaaacgagtgaagaagacgaagttgatgacaaagaaaaagaacaaactgaacaaccaaccgaagaaggagtcgacccagacg
1 M K R V K K T K Ii M T K K K N K Ii N N Q P K K E S T Q T
25424 ttcaaggttaattgtgaccattgtgagcataagttcgaccttacatctaaacagattatttcgaaacatatcgaaaagggcgta
29 F K V N C D H C E H K F D L T S K Q I I S K H I E K G V
25508 gagtggagattcttcgaatgtcctaagtgccattatcggttcaccacttatgtaggaaacaaggaaattgaaaaccttattcga
57 E W R F F E C P K C H Y R F T T Y V G N K E I E N L I R
25592 tttagaaatacttgtcgagctaaaatgaagcaggaacttcaaaaaggagctgctgctaatcaaaacacttaccattcatatcga
85 F R N T C R A K M K Q E L Q K G A A A N Q N T Y H S Y R
25676 attcaggatgagcaagctgggcataaaatctcagggcttatggcgaagctaaagaaggagataaacattgaaaaacgagaaaaa
113 I Q D E Q A G H K I S G L M A K L K K E I N I E K R E K
25760 gaatgggtatctatatag 25777
141 E V S I * dplORF046
42774 atgccaatgtggctaaacgacacagcagtcttgacgacgattattacagcgtgcagcggagtgcttactgtcctactaaataag
1 M P M W Ii N D T A V Ii T T I I T A C S G V Ii T V Ii L N K
42858 ttattcgaatggaaatcgaataaagccaagagcgttttagaggatatctctacaactcttagcactcttaaacagcaggtcgac
29 L F E K S N K A K S V L E D I S T T L S T L K Q Q V D
42942 gggattgaccaaacgacagtagcaatcaatcaccaaaatgacgtcattcaagacggaactagaaaaattcaacgttaccgtctt
57 G I D Q T T V A I N H Q N D V I Q D G T R K I Q R Y R L
43026 tatcacgacttaaaaagggaagtgataacaggctatacaactctcgaccattttagagagctctctattttattcgaaagttat
85 Y H D Ii K R E V I T G Y T T Ii D H F R E L S I L F E S Y
43110 aagaaccttggcggaaatggtgaagttgaagccttgtatgaaaaatacaagaaattaccaattagggaggaagatttagatgaa
113 K N L G G N G E V E A L Y E K Y K K Ii P I R E E D Ii D E
43194 actatctaa 43202
141 T I * dplORF047
47542 atgaaatttgaagatgaaaaacagttcatcgctgcaattgaagaagccggtgaattaaatgctaccaaaggcgacatggagaaa
1 M K F E D E K Q F I A A I E E A G E L N A T K G D M E K
47626 caagtcaaaagtcttcgtgatgctctaaaagagtacatgaaagaaaatgacattgaatctgctcaaggtaagcacttttctgct
29 Q V K S Ii R D A L K E Y M K E N D I E S A Q G K H F S A
47710 accttctacacgacagagcgctcaactatggacgaagaacgcttgaaagaaattatcgaaaaattagttgacgaagccgagacg
57 T F Y T T E R S T M D E E R L K E I I E K L V D E A E T
47794 gaagaaatgtgtgaaaaactttcagggcttatcgaatacaagcctgtcatcaatacgaaacttctcgaggatatgatttatcac
85 E E M C E K L S G L I E Y K P V I N T K L L E D M I Y H
47878 ggcgagattgaccaagaagcaattcttccagcagttgtcatttctgttacagaaggcattcgttttggaaaggctaaaatttag
47961
113 G E I D Q E A I L P A V V I S V T E G I R F G K A K I * dplORF048
16709 atggaaacaacactttatttcggttatcttacagcagattggaaagacggtcacaagaactacactttccactatgaaagcatt
1 M E T T L Y F G Y L T A D K D G H K N Y T F H Y E S I
16625 cctgtaaaagaaactgagaaacaatataaggtcactggaatcaatcctaacttgtacttagacctaggctcagttattagaaag
29 P V K E T E K Q Y K V T G I N P N Ii Y L D Ii G S V I R K
16541 agcgaacttgacattgcagtattcaaagcatgtcctgtcgctgaaactggagtcacacttactcgcgacatggaagttgatgct
57 S E Ii D I A V F K A C P V A E T G V T L T R D M E V D A
16457 agaattgaaatcatcaagaaattaactacaagaatcgaacgccttaacgaaagaattaaagcaagaaatgaacaaggtaaacaa
85 R I E I I K K L T T R I E R L N E R I K A R N E Q G K Q
16373 gaaagccgccacctagtatctgcgctagaagattgcgctcgtcaaattgctggaatttatcaataa 16308
113 E S R H L V S A L E D C A R Q I A G I Y Q * dplORF049
44018 atgtttcaaccatttctcagcgagcatgtagccttggtcgtcaaagtagaaccaagacttgttttcttcgatatactcgaactc
1 M F Q P F L S E H V A Ii V V K V E P R L V F F D I L E L
43934 atcttttggataagttccgtttgctcgagcgtaccagaaaccagtagcatctttctgccagccaagtttcttctcagccggttg
29 I F I S S V C S S V P E T S S I F L P A K F L L S R L
43850 agcatttgcgttagtcaagcgatagacgtagtagtaaggttgacctgcatagtaccaacgctcatcgtggtcgttgacggaaat
57 S I C V S Q A I D V V V R L T C I V P T I V V V D G _N
43766 tccgtcgtaggcgtagttgcagtgaatgatgttatcactgtcaatgaacatccctgtatgacctccagcgcctgcgcragcacc
85 S V V G V V A V N D V I T V N E H P C M T S S A^C A S T
43682 tttgcgtccccagatgaagatgtcgcctcgtttagcatcccacggagcattttcactaattag 43620
113 F A S P D E D V A S F S I P R S I F T N * dplORF050
15081 atgaacaatcagcgaaagcaaatgaacaaacgaatcgtcgaacttcgcgaagactatcaacgtgcaagaggtcgaataaacttc
1 M N N Q R K Q M N K R I V E L R E D Y Q R A R G R I N F
15165 cttcttgctgtaaaggaccacggcgaagaactcgaaaaccttgaagcctttgtgggatacattgacaatctagtcgaatgtttt
29 L L A V K D H G E E L E N L E A F V G Y I D N L V E C F
15249 cctgaaagccaacgaaatgtcttgaggctatgtgtattagatgaccttccagtcactaatgcggccgctgaaattggataccac
57 P E S Q R N V L R L C V L D D L P V T N A A A E I G Y H
15333 tatacatgggttcaccaacttcgagacaaagcagttgaaacacttgaagaaattttagatggggataacattattcgctctaaa
85 Y T W V H Q L R D K A V E T L E E I Ii D G D N I I R S K
15417 cacggaatcgaaattaaggagaaacttgatgaattatatggtaaaagtcattctagttag 15476
113 H G I E I K E K L D E L Y G K S H S S * dplORF051
29765 atgagttatgacgtgaattatgttaagaatcaagttcgtagagccattgaaaccgctcctactaaaatcaaggtacttcgaaac
1 M S Y D V N Y V K N Q V R R A I E T A P T K I K V L R N
29849 tcttgggtcagtgatggatatggaggaaagaaaaaggataaagcgaatgaagtcgtagcagacgaccttgtttgtttagttgat
29 S W V S D G Y G G K K K D K A N E V V A D D Ii V C L V D
29933 aattcaactgttcctgaccttttagccaattctactgacgcgggaaaaatttttgcccaaaatggagtgaaaattttcattcta
57 N S T V P D L L A N S T D A G K I F A Q N G V K I F I L
30017 tatgatgaaggcaaaatcattcaacgagccgatactatcgaaattaaaaactcaggaagacggtacagggtagtagaaacccac
85 Y D E G K I I Q R A D T I E I K N S G R R Y R V V E T H
30101 aatcttctcgagcaagacattttgatagaacttaaattggaggtgaacgactaa 30154
113 N L L E Q D I L I E L K Ii E V N D * dplORF052
30516 atgactaaacgaacgacaatgatggacagattgaaggaaattcttcctacatttcagctctcgcctgctcctatgcttccagga
1 M T K R T T M M D R L K E I Ii P T F Q L S P A P M Ii P G
30600 gttgaatttgacgagcaagatacagataggccggatgactacattgttcttcgatatagtcatagaatgcccagcgcaacaaat
29 V E F D E Q D T D R P D D Y I V L R Y S H R P S A T N
30684 agcctaggaagttttgcttattggaaagttcaaatctacgtccattcaaactcaattattggtatcgacgaatatagcagaaag
57 S L G S F A Y K V Q I Y V H S N S I I G I D E Y S R K
30768 gttcgaaacattatcaaggacatgggctacgaagtaacctatgcagaaactggtgactacttcgacacaatgctttctagatac
85 V R N I I K D M G Y E V T Y A E T G D Y F D T M L S R Y
30852 cgactagaaatcgaatatagaattccacaaggaggaaactaa 30893
113 R L E I E Y R I P Q G G N * dplORF053
50300 atgctaacattcgaaagaatagtatctatacgagcaccaacttgcatttcactcatttccccgctatatagaaggacatcatgc
1 M L T F E R I V S I R A P T C I S L I S P L Y R R T S C
50216 ccgttcttccaagcagttgcaagcattttatcaatagtccacgacttaccttgtccaggtcgagccattatgacaatcaaatcc
29 P F F Q A V A S I L S I V H D Ii P C P G R A I M T I K S
50132 tcaccaggaagtaagcctccaagcacgtcgtccaatagttcaaaccctgtcgatattccaagtctttcaccgtcatggtttcta
57 S P G S K P P S T S S N S S N P V D I P S L S P S W F L
50048 atagtattcgcccagtctagtcgaagtttagcatttcgagcaatgtctagtccgcctacgaatttagagcgattgaaaagttct S P P T N Ii E R L K S S a 49917
dplORF054
14423 atgtgtgaaaattgtcaaaacgaaacattcaatactagaattttcaatgaagatgaaagtggctatgtcgacgcctcattcact
1 M C E N C Q N E T F N T R I F N E D E S G Y V D A S F T
14507 tacaaggagattcgcgacaccgcagcagctattagcaatcgagcggtagaaaagaaagaccgtgacagccttttagtcgctaca
29 Y K E I R D T A A A I S N R A V E K K D R D S L L V A T
14591 gttatggctcttcccgtttctcacgcagaagatttaggcaagagactttgtattgcaaattctcgattggaagcatttcgtgaa
57 V M A L P V S H A E D L G K R L C I A N S R L E A F R E
14675 gctgttcaagaggctctcgagaatgaaaaggctgaagatttaaaggacgttatcttaggtcttatcgacgttgacaaaaaaatt
85 A V Q E A L E N E K A E D L K D V I L G L I D V D K K I
14759 ggcaaccttgcattgcaattagttgaatcaggagcattataa 14800
113 G N Ii A Ii Q Ii V E S G A Ii * dplORF055
27627 atgcctaatgtgcgagttaagaaaactgattttaatcaaaccactcgaagcattgtcgcaattcctgaccactacgttgctttg
1 M P N V R V K K T D F N Q T T R S I V A I P D H Y V A L
27711 gctgctcaaattccagctaccgcagcaactcaagtagggaacaagaaatacattcttgccggaacttgcgtgaaaaatgctact
29 A A Q I P A T A A T Q V G N K K Y I L A G T C V K N A T
27795 acatttgaaggacgcaaaactggactcgaagtagtatctaccggtgaacaattcgacggagttatcttcgctgaccaagaagtg
57 T F E G R K T G L E V V S T G E Q F D G V I F A D Q E V
27879 tttgaaggtgaagaaaaagtaaccgtgacagtattagttcacggattcgtcaaatatgcagcccttcgaaaagttggcgatgct
85 F E G E E K V T V T V L V H G F V K Y A A L R K V G D A
27963 gtgcctgaatctaaaaacgcaatgattcttgtcgttaaatag 28004
113 V P E S K N A M I L V V K * dplORF056
19151 atggaaaataaatggaaagttatccattttcaaaactcatgtattaaacaagtagacgatgaaaaaaggaggctcctgttcgaa
1 M E N K W K V I H F Q N S C I K Q V D D E K R R L Ii F E
19067 gttccaggaactccttatcgtctacaagtttgggtgaaaatgagcttagttaaaattgaaacacgcgcaggaaatggctattat
29 V P G T P Y R L Q V W V K S L V K I E T R A G N G Y Y
18983 aaaaggctagtatgccaagacgattttgtattttatggtaaggagtcaatagatggttacttaattgacgccaccafcaactggc
57 K R L V C Q D D F V F Y G K E S I D G Y L I D A^ f I T G
18899 aaatctttggcggaatattgtgagcctatgaacaggcatattctcgaaactattgcatcgcgagaagcagctgaactgaacaga
85 K S L A E Y C E P M N R H I L E T I A S R E A A E L N R
18815 gctaaaaagcaagaccaacagaaatggagatactag 18780
113 A K K Q D Q Q K W R Y * dplORF057
9859 atgcaaaaatctctatttggacctaagctagtgcctgctagttcaaggcgcaagaaaagaacggttccaaaacctaaacctaaa
1 M Q K S L F G P K L V P A S S R R K K R T V P K P K P K
9943 atcgatgagcaagtggttgagcttatgaaccgcagagagcgtcaagtgcttgttcatagttgcatctattattattttaatgac
29 I D E Q V V E L M N R R E R Q V L V H S C I Y Y Y F N D
10027 tcaattatagcagacgggcagtatgacaaatggagccacgaactatattctcttatagtttcgcaccctgatgagtttcgacag
57 S I I A D G Q Y D K W S H E L Y S Ii l V Ξ H P D E F R Q
10111 actgttctctataacgagtttaaacagtttgacggaaatactggaatgggtcttccatacgactgtcagtttgctgtaagggtc
85 T V L Y N E F Q F D G N T G M G L P Y D C Q F A V R V
10195 gcagaaaggcttttaagaaaatga 10218
113 A E R L L R K * dplORF058
15633 atgacatcacgcgcatacaaaccaattcccacgcgcagagctagtgctaaacaagagaaggcagttgctaagcagttgggagga
1 T S R A Y K P I P T R R A S A K Q E K A V A K Q L G G
15717 aaagtacagcctaattcaggagccactgactactacaaaggtgacgtcgtaacagactcaatgcttatagaatgcaagacagtt
29 K V Q P N S G A T D Y Y K G D V V T D S M L I E C K T V
15801 atgaagccacaaagttcagtcagcttgaaaaaggaatggttcctaaaaaatgaacaggaaaggttcgctcaaaaactcgactat
57 M K P Q S S V S L K K E W F L K N E Q E R F A Q K L D Y
15885 tctgctatcgctttcgactttggtgacggaggcgaacagtatatagcaatgtctataagtcagttcaagcgaatattagaggat
85 S A I A F D F G D G G E Q Y I A M S I S Q F K R I L E D
15969 agaaatgataaccttatttaa 15989
113 R N D N L I * dplORF059
30154 atgtctcagcctgaattagtatggaagcctgaagaatttgttagtaactgtgaacggtatcgaaacaagtttcaagtcgctgtc
1 S Q P E L V K P E E F V S N C E R Y R N K F Q V A V
30238 ataacagtctgcgaagtcgctgctactaagatggaagaatacgcaaagacgcatgctatttggacagaccgtacagggaatgct
29 I T V C E V A A T K M E E Y A K T H A I T D R T G N A
30322 cgacagaaactcaaaggagaagctgcttgggtaagcgcagaccaaatcatgatagctgtatcacatcacatggactacgggttt
57 R Q K L K G E A A W V S A D Q I M I A V S H H M D Y G F
30406 tggctagaactagctcatggtcgaaaatacaaaattctcgaacaggctgtagaagacaatgtcgaagaactttttagagcgttg
85 W -i E -i A H G R K Y K I L E Q A V E D N V E E L F R A L
30490 agaaggttattagactag 30507
113 R R L L D * dplORF060
38070 gtgatagctgtatctgctatccctactccgctctttccaggtacaccgtcgactccatcacgcccaggagctcccggtaaacct
1 V I A V S A I P T P L F P G T P S T P S R P G A P G K P
37986 gcgtcacctttaggaccttctagtcgaatccatgtaaagtcgtcaggaactaattcgctcggtttcttattagtattaaggaca
29 A S P L G P S S R I H V K S S G T N S L G F L L V Ii R T
37902 ccaatgtatttcccagattctgcattaaaattagtccctaaaatgtcatctgcgtatctaataacaacttgggactcatttaca
57 P M Y F P D S A L K L V P K M S S A Y Ii l T T W D S F T
37818 gtttcccctgaaaggactccttcgccgtcctcatttagcaagtccatcaagtcttttcgagggtcttggaaaatgatagtagag
85 V S P E R T P S P S S F S K S I K S F R G S W K M I V E
37734 tttgaaaggtcgtcgtag 37717
113 F E R S S * dplORFOδl
19475 atggcgagaatgcaaagattatgcccgatgaaattttggaaggcggtaactaaaatgaaattcgaagtttattctgcgcgacta
1 M A R M Q R L C P M K F K A V T K M K F E V Y S A R L
19391 tttgacgaagaggcgacatatgataggtatcgtgaagcactagagaaagttggaaatgtcgcttacttttgtgaaattgatact
29 F D E E A T Y D R Y R E A L E K V G N V A Y F C E I D T
19307 ggcaaccttgtaatcgaactcgagctagacagcctagatgacctaatcgcgctttcaaatgtagtgggaactggactaaaatta
57 G N L V I E Ii E L D S Ii D D L I A L S N V V G T G L K L
19223 tcacggccttatagagaagataagccttttcaattatggattgttgacgggtacatggaataa 19161
85 S R P Y R E D K P F Q L W I V D G Y M E * dplORF062
45284 gtgagaagcttcaatcaattccattgcggtgtcaatatcttcttccttgacgagtttaaaaattccgtcaatcgcccattcgta
1 V R S F N Q F H C G V N I F F L D E F K N S V N R P F V
45200 agatgcaggagcaatagatgcaagaagtttcttttggtcttctgtcaacccttttgcgcgaactccaatagaaacaccttttcg
29 R C R S N R C K K F L L V F C Q P F C A N S N R N T F S
45116 agtttcttcgatagtaacgaagttcttcttagagcgataggcgatgtaagactgagtgacgattcgagtagacgcaggaaaggc
57 S F F D S N E V L L R A I G D V R L S D D S S R R R K G
45032 ttcaacaattcgactttcaagagccttagtaatcgccatcacgctttcttttttaggagcaggttttcgaacagtagatttctc
85 F N N S T F K S L S N R H H A F F F R S R F S N S R F L
44948 actaactga 44940
113 T N * dplORF063
47200 atgaaattcactgaaggaaaaaattggtataaagttggagagatatgtcaaatgttgaaccgctctctatctacgattaatgtt
1 M K F T E G K N Y K V G E I C Q M L N R S L S T I N V
47284 tggtatgaagcaaaagacttcgctgaagaaaataacattcacttcccgtttgttcttcctgaacctagaacagaccttgaccat
29 Y E A K D F A E E N N I H F P F V L P E P R T D L D H
47368 cgtggttctcgattctgggatgacgaaggcgtgaacaaactcaaacgatttagggacaacctaatgcgcggtgactt^gcattc
57 R G S R F D D E G V N K L K R F R D N L M R G-,D L A F
47452 tacactcgaactcttgtagggaaaactgaaagggaagcaattcaagaagatgctaaagcatttaaacgtgaacatggattggag
85 Y T R T L V G K T E R E A I Q E D A K A F K R E H G L E
47536 aattaa 47541
113 N * dplORF064
29108 atggctacattgaaagctcttagcaccttaatcgtttccggagcagtagtgcattcagggtcggtattttcttgccctgaagcg
1 M A T L K A L S T L I V S G A V V H S G S V F S C P E A
29192 cttgcttcgtctttaattgaacgcaattttgcgttcgagattaaggcggctgaagatggagaaacggtagaaactgttcc caa
29 L A S S L I E R M F A F E I K A A E D G E T V E T V P Q
29276 acaattgaatcagttgaagaaattgacgaagttgaacaaatgcgcgaagagtatgcggctaaaaccgttcctgagctcgttgaa
57 T I E S V E E I D E V E Q M R E E Y A A K T V P E I. V E
29360 ttagcaagagctaatggaattgacatttcttcaatttctcgaaaaagcgaatatatcgacgctttaattaagtacgaactagga
85 li A R A N G I D I S S I S R K S E Y I D A Ii l K Y E L G
29444 gagtaa 29449
113 E * dplORF065
51497 atgcagtttgtcataacctacatcaaacatctcgatgagctcgtccgtcaatttccgttcatacatataaggatgaataaaccg
1 M Q F V I T Y I K H L D E L V R Q F P F I H I R M N K P
51413 gtatttatcaagttcctcttcaggaatgattttatgctcgactttttcagttctcccatttcttcgaaacgcttcagggctgac
29 V F I K F L F R N D F M L D F F S S P I S S K R F R A D
51329 gccttgcctaactacttcgctagatgttccaaaattccttttcagccactggtttccatagaaccctccatcgtttcgacctaa
51246
57 A L P N Y F A R C S K I P F Q P L V S I E P S I V S T * dplORF066
28898 gtgaccaactgcgtcaggtggaagcaataccactttaccgtcgtcaatcaagttgaactgacgaatgttaccaacgtcaggaag
1 V T N C V R W K Q Y H F T V V N Q V E L T N V T N V R K 28814 tttgtcagcgtcagcgaactgagcaattttcttagagtagacagcgatttgaagacctgttttttcagcgatgaatttctcagc 29 F V Ξ V S E L S N F L R V D S D Ii K T C F F S D E F Ii S
28730 gtcacttgcaagaagcaagaagttttcccaagaaccttgaacaccaattgcaagagctttcttgatagagtcactcttagtcat 57 V T C K K Q E V F P R T L N T N C K S F Ii D R V T L S H 28646 ttggttataagtgtttcggttcaagaccattcgagtagggcgaacacctgtacgattttcgatgtcatccattgctgctaa 28566
85 li V I S V Ξ V Q D H S S R A N T C T I F D V I H C C * dplORF067
45061 gtgacgattcgagtagacgcaggaaaggcttcaacaattcgactttcaagagccttagtaatcgccatcacgctttctttttta
1 V T I R V D A G K A S T I R L S R A Ii V I A I T li S F L
44977 ggagcaggttttcgaacagtagatttctcactaactgaaccaacttcttccggctgttccttaacttcaggaatttcttcctca
29 G A G F R T V D F S L T E P T S S G C S L T S G I S S S
44893 aggacttcttttttaggtttgggaacgactctaccttttcgagcaggtcgagcaactgcaggagcagcctttttagcaggttta
57 R T S F L G L G T T Ii P F R A G R A T A G A A F L A G Ii
44809 gcagcttcttcttttttaggttcagtttcatcttccattgtgtaccaacgttcgagagttgaagctgaaaggtga 44735
85 A A S S F L G S V S S S I V Y Q R S R V E A E R * dplORF068
29451 atggcagctcaaacggacattgaattagtcaaaatcaatatcgataacgataattctccgtcaccaatgactgaccaaagtatc
1 M A A Q T D I E L V K I N I D N D N S P S P M T D Q S I
29535 tcagctcttttagacaagcataaatctgtcgcctatgttagttatatgatttgcttaatgaagacccggaatgacgtggtaacc
29 S A Ii L D K H K S V A Y V S Y M I C Ii K T R N D V V T
29619 cttggacctatcagtctaaaaggtgacgcagactactggaaacaaatggcgcaattctattatgaccaatataagcaagaacag
57 L G P I S L K G D A D Y K Q M A Q F Y Y D Q Y K Q E Q
29703 cttgaaactgatgaaaagtcgaacgctggttcgacaatcttaatgaaaagggctgatgggacatga 29768
85 L E T D E K S N A G S T I L K R A D G T * dplORF069
20411 atgaaactttatcacgccactgattttgataatcttggtaaaattctagctgaaggattgaagccttcagctggagttatttac
1 M K L Y H A T D F D N L G K I Ii A E G Ii K P S A G V I Y
20327 ctagcagaaagttatgaaaaggctctagcctttttatcgcttcgaaatgttgatactattgtcgttctcgaacttgaagtagat
29 L A E S Y E K A L A F L S L R N V D T I V V L E L E V D
20243 attgaaaaatgtactgaaagtttcgaccataatgaaaagatgttttgtagcctatttcatttcgacacttgtcgcgcttggact
57 I E K C T E S F D H N E K M F C S L F H F D T C R A W T
20159 tatgacaagacaattgaagtagacgacattgacttttcgaaagctcgaaaatatgatagaaagtga 20094
85 Y D K T I E V D D I D F S K A R K Y D R K * dplORF070
15973 atgataaccttatttaaaataaacagtgaaggaacagttactccaattaaagggtcagccatgcaactgtacgcagaccttatt
1 M I T L F K I N S E G T V T P I K G S A M Q Ii Y A D L I
16057 cctatacaagaggacgatatacagttcgttgatataactggacttgaccctattgttcgagaaaacgtacttgagctcatttca
29 P I Q E D D I Q F V D I T G Ii D P I V R E N V Ii E L I S
16141 cggagccgtgtaggagtttcaaaatatggtacaaacctcgaccagaatgatgtcgacgatttcctacagcacgccaaagaagaa
57 R S R V G V S K Y G T N L D Q N D V D D F L Q H A K E E
16225 gcgctcgactttgctaactacctaaccaagctacaaagtcaacaaaagcaaaataaatag 16284
85 A L D F A N Y L T K L Q S Q Q K Q N K * dplORF071
38904 gtgaaacaggtcctagaggagttcaaggtcttcaaggtcctcaagggcttcaaggaattcctggacctgcaggagctgacggac
1 V K Q V L E E F K V F K V L K G F K E F L D L Q E L T D
38988 gttcgcaatatactcacctcgctttctctaatagtccaaacggtgagggatttagtcatactgacagcggacgagcatacgtcg
29 V R N I L T S L S L I V Q T V R D L V I L T A D _E_ H~ T~ S
39072 gtcagtatcaagatttcaatcccgtccattcaaaagaccctgcagcctatacatggacgaaatggaaggggaatgacggagctc 57 V S I K I S I P S I Q K T Ii Q P I H G R N G R G M T E L 39156 aagggatacccgggaagccaggcgcagacggtaagactaattatttccatatag 39209
85 K G Y P G S Q A Q T V R L I I S I * dplORF072
51045 atgttccttcgtcttcaagttgtctcgaaagtttttcaattatttgttcaggagtcgcttcaatttgaagaccatttactttca
1 M F L R -i Q V V S K V F Q Ii F V Q E S L Q F E D H Ii li S
50961 tcaaaatgcttcaactccttcccttgtaaccttacttcgaagacgagcagtcgacctagaggcttttgctttcaatggagagct
29 S K C F N S F P C N L T S K T S S R P R G F C F Q W R A
50877 ttcgcctttttcagttccttcttcgccttcctctttgaatcctataagagtataggttccagtttcaacgtcccacatatattc
57 F A F F Ξ S F F A F L F E S Y K S I G S S F N V P H I F
50793 gatgatttttcggtcttcgccatatcggtttttaacgacagatag 50749
85 D D F S V F A I S V F N D R * dplORF073
14262 gtgaacgcttgccggaagaatacgacgaagaaacttgggaacctatcactgaagcagaatacatcaagcgaacagaaaaaccta
1 V N A C R K N T T K K L G N L S L K Q N T S S E Q K N L
14346 aagcagttgcaaaacctactcgaaaaactccagcgccttctcgtcgccctcgcccttaaaagaaaggttgaaataaaatgtgtg
29 K Q Ii Q N L L E K Ii Q R -i L V A L A Ii K R K V E I K C V
14430 aaaattgtcaaaacgaaacattcaatactagaattttcaatgaagatgaaagtggctatgtcgacgcctcattcacttacaagg
57 K I V K T K H S I L E F S M K M K V A M S T P H S L T R
14514 agattcgcgacaccgcagcagctattagcaatcgagcggtag 14555
85 R F A T P Q Q L L A I E R * dplORF074
32298 gtgacgaaaagaaaaatccaggattgcaaatgcttatggagtgactattttcagtcgctcctctttttgtatatagaaaggaaa
1 V T K R K I Q D C K C L S D Y F Q S L L F L Y I E R K
32382 ttacatggattttgggtcaattgcagcaaaaatgactttggatatctcaaacttcacaagtcaattaaatcttgctcaaagtca
29 li H G F W V N C S K N D F G Y L K Ii H K S I K S C S K S
32466 agcgcaacggctcgcactagagtcttcgaagtcctttcaaattggttctgctttaacaggattagggaaaggacttacgactgc
57 S A T A R T R V F E V L S N F C F N R I R E R T Y D C
32550 ggttacccttcctcttatgggatttgcagccgcctctattaa 32591
85 G Y P S S Y G I C S R L Y * dplORF075
22447 atggcaaagttttgtccgttgaattccgtcatggcccaaagggaaaatgaaagagccatcgatactgtttttcctgaacgaatg
1 M A K F C P L N S V M A Q R E N E R A I D T V F P E R M
22363 gaaccgtctgctatgacgatatcgaaagttcgaaaaggtgagccctttgtccaccatgttaggagctggagttgtttcttacta
29 E P S A M T I S K V R K G E P F V H H V R S W S C F L Ii
22279 aaagggacgaagttgaacttaggtagtttatttctcaggcttattgtcattatcagtcactcctttaatgtaggaacctgttgc
57 K G T K L N L G S L F L R Ii l V I I S H S F N V G T C C
22195 gtcactaaattcttgccaaacggcttgagctgctttatctag 22154
85 V T K F L P N G L S C F I * dplORF076
5728 gtgagagcattttcttcactcacgtcttcgagcaagtggtcgaatgtagggtactcttcatcttctgtaacaatatcaatattg
1 V R A F S S L T S S S K S N V G Y S S S S V T I S I L
5644 tactcaccattcccaataacttttagcgaagattcttcaggaactaatgtgacggttgcggccgtggtcttttctacaagtttt
29 Y S P F P I T F S E D S S G T N V T V A A V V F S T S F
5560 ccaaactgctctgctttcacaatcacgtcaatttcaacatcgctgtcgataatgcatcgaaggaagtttgagccatcatacgct
57 P N C S A F T I T S I S T S L S I M H R R K F E P S Y A
5476 gtaaacatgacgcattcgccgtcaccaaaaatatgccaatag 5435
85 V N M T H S P S P K I C Q * dplORF077
14800 atggaacgaataaagacgctatttcacgtgatttatgctaacggcactcatttagaagtagcagctttgttcgataccgttgat
1 M E R I K T -i F H V I Y A N G T H Ii E V A A L F D T V D
14884 gattatgatgacgttatagaggacatccaggggtatattgatacccctgacctttataatcaaaggagcattagaatggcgcct
29 D Y D D V I E D I Q G Y I D T P D L Y N Q R S I R M A P
14968 tacaatcctgacatcaatggtgacgctattgctactgacattttactacgactagatgatattatctacgtcgacgcaacttgt
57 Y N P D I N G D A I A T D I L L R L D D I I Y V D A T C
15052 gaaactattaaatacgaggagcctattgcatga 15084
85 E T I K Y E E P I A * dplORF078
17507 atggcaacagtaaaggaaacagtaaaatttgacggacgtcttgtaactatcttcgactacgacgatttagagtgggaaggatat
1 M A T V K E T V K F D G R L V T I F D Y D D L E W E G Y
17423 gcacctaatgaaggattcgaagatgttgaggacatggaagtccttagcattcgagttcgaaacgaaggtgaggacgacgagtgg
29 A P N E G F E D V E D M E V L S I R V R N E G E D D E
17339 gttgaagttatcgcctgctatgaaaacgatgacgaggacgaagatttggaagggttataa 17280
57 V E V I A C Y E N D D E D E D L E G Ii * dplORF079
35288 atggaactgataccattgataaatcctcgaacaaggttgacccctgcgcttaccatttgtccagcgaatccagtaaccttagaa
1 M E L I P Ii l N P R T R Ii T P A L T I C P A N P V T L E
35204 acaattgaagttcccatgctgccaattttagagacagctgaaccaatcattgacccaataccactaatgaagtttcgaatcagg
29 T I E V P M Ii P I L E T A E P I I D P I P Ii M K F R I R
35120 ttcgcacctcctgaaaccatctgtcccacaaagctagcaatcttgctaactaatgatgaaagcatgtttccagctgtcgataaa
57 F A P P E T I C P T K Ii A I L Ii T N D E S M F P A V D K
35036 agtgagccgagaagtgaagcaataccttga 35007
85 S E P R S E A I P * dplORF080
42490 atgttgaaccttacaaaatcgcgccaaattgtggcagagttcactattggacaaggagctgaaaagaaacttgfccaaaacaacg
1 M L N L T K S R Q I V A E F T I G Q G A E K K V K T T
42574 attgtgaacattgatgcaaacgcagtatcaaccgtctctgaaactcttcatgacccagacttgtatgctgcgaaccgtcgagaa
29 I V N I D A N A V S T V S E T L H D P D L Y A A N R R E
42658 cttcgagctgacgagcaaaaacttcgcgaaactcgttacgcaatcgaagatgaaattctagctgaacagtcaaagactgaaaca
57 L R A D E Q K L R E T R Y A I E D E I L A E Q S K T E T
42742 gctctaacagctgaataa 42759
85 A L T A E * dplORFOβl
55466 atgttcaggaacagtatcgtccatctgttggtctgcgtcaaagttaaaggggtcgaaatcttcgttcttgctagcgtcgatata
1 M F R N S I V H Ii L V C V K V K G V E I F V L A S V D I
55382 ctcgaactcgtattcaggaagactcatatcaggaagccttcttcttcgaccggtagctgtttgaacatatcccaagtcctgcgc
29 L E L V F R K T H I R K P S S S T G S C L N I S Q V L R
55298 ctgctgttgaacgaatatgatatagtctgccactttagggaactcggtgaagaaatcttcaataaccttattcgcttctttgac
57 L li L N E Y D I V C H F R E Ii G E E I F N N L I R F F D
55214 agatacattcatctgctcagcgattga 55188
dplORF082
44728 gtgaacttcacctttcagcttcaactctcgaacgttggtacacaatggaagatgaaactgaacctaaaaaagaagaagctgcta
1 V N F T F Q L Q Ii S N V G T Q W K M K Ii N Li K K K K Ii L
44812 aacctgctaaaaaggctgctcctgcagttgctcgacctgctcgaaaaggtagagtcgttcccaaacctaaaaaagaagtccttg
29 N L L K R L L L Q L L D L L E K V E S F P N L K K K S L
44896 aggaagaaattcctgaagttaaggaacagccggaagaagttggttcagttagtgagaaatctactgttcgaaaacctgctccta
57 R K K F L K L R N S R K K L V Q L V R N L L F E N L L L
44980 aaaaagaaagcgtga 44994
85 K K K A * dplORF083
35974 atgccttcagggtttttaaatcctgagtccttaaatcctgcgaaagtgagtcctacatattctagcacggttgcacctttgtcg
1 P S G F Ii N P E S L N P A K V S P T Y S S T V A P L S
35890 acaaggtcaattccgtcgaccaatagcgtctgtctgctagccatctatttctcctttacggtgttacaatgttaccaaaccctg
29 T R S I P S T N S V C L L A I Y F S F T V L Q C Y Q T Ii
35806 atagagtttctttacttctattatacaatcctctcgacagtttgtcaacgtcgtcattgtttcgaactacgattgttccaatgt
57 I E F L Y F Y Y T I L S T V C Q R R H C F E L R L F Q C
35722 tga 35720
85 * dplORF084
15445 atgaattatatggtaaaagtcattctagttagtgtctttgtactgtcagccttttgcatgacttgctcaatggtttatttggtt
1 M N Y M V K V I L V S V F V Ii S A F C M T C S M V Y Ii V
15529 acaggtaagcaagaggaccaccgtagtaccgtcgcccttgtatttggcgctctcgtaagctctgcggcgttctattcgacactc
29 T G K Q E D H R S T V A L V F G A L V S S A A F Y S T L
15613 tttatcctcgcctatctgccatga 15636
dplORF085
10847 gtgatgactataatcaaggactttttcgagccttgtgatactgtcacgcattcctccatttgcaagtttcccaataaacgaaag
1 V M T I I K D F F E P C D T V T H S S I C K F P N K R K
10763 ggcgtcacgctcataactataaccagctccttcttcattttcactttcgataataaattgaagttgattaacgatgtcgtcatt
29 G V T Ii l T I T S S F F I F T F D N K Ii K Ii l N D V V I
10679 atcaattcgagtaaagtcaaaccgttgaactcgactgagaatagtgtcaggaatcttttgagggtcagtagtacatag 10602
57 I N S S K V K P L N S T E N S V R N L L R V S S T * dplORF086
52760 atatgggaaaagtatcaattcaaaaatcaggaacatttagctcagggtctaataacgagtttttcacactcgctgaccacggtg
1 I W E K Y Q F K N Q E H L A Q G L I T S F S H S L T T V
52844 acagcgcaattgtcactctattgtatgatgacccggaaggcgaagacatggattatttcgtag 52906
29 T A Q L S L Y C M M T R K A K T W I I S * dplORF087
30036 atgattttgccttcatcatatagaatgaaaattttcactccattttgggcaaaaatttttcccgcgtcagtagaattggctaaa
1 M I P S S Y R M K I F T P F W A K I F P A S V E Ii A K
29952 aggtcaggaacagttgaattatcaactaaacaaacaaggtcgtctgctacgacttcattcgctttatcctttttctttcctcca
29 R S G T V E L S T K Q T R S S A T T S F A L S F F F P P
29868 tatccatcactgacccaagagtttcgaagtaccttgattttagtaggagcggtttcaatggctctacgaacttga 29794
57 Y P S L T Q E F R S T L I L V G A V S M A Ii R T * dplORF088
5040 atgaaaaaagttcaaacttatcaagaatatctaaaactagttgagttcaaacgtcaactttctttaaatcttcgagaaggaaaa
1 M K K V Q T Y Q E Y L K L V E F K R Q L S Ii N Ii R E G K
5124 ataggagtcgatgaagcggttattcaattattcaccttctatagtttcaacaatatcgaggaacctcctttcattgtactcaaa
29 I G V D E A V I Q L F T F Y S F N N I E E P P F I V Ii K
5208 atgcaagaggctgccgtgaacgggacttatgaagcaaaactcaatatgcttaaaagatttaaaattatttag 5279
57 M Q E A A V N G T Y E A K L N M L K R F K I I * dplORF089
12495 atgtcaatcatgtcgctatcaatagtcgagtatttagacacaaaatgccttttcaactgcgcgtcagtcattttctcaaactca
1 M S I M S Ii S I V E Y Ii D T K C L F N C A S V I F S N S
12411 acacaattatcaggaaaggcctttagcaacttgcttcgcttgtcaattttagtaaccatcaaaacaagtgtcccat tctaaca
29 T Q L S G K A F S N L Ii R L S I L V T I K T S V P Y L T
12327 tccggaagccttttccacctcgactcattagacagaaactccttatcatctcgaacagcgaatattcgatga 12256
57 S G S L F H L D S Ii D R N S Ii S S R T A N I R * dplORF090 " "'
27037 atgctaaaattttcattgacggcgacggtcaacattttgtacctcacgcacgtttcgatgaagttgttcaacagcgcgatgcag
1 M L K F S L T A T V N I L Y L T H V S M K L F N S A M Q
27121 ctaacggctcaattaattcttataaagaacaagtcgcgacgctttctaaacaggtcaaagataacggtgatgcgcagaccacta
29 L T A Q Ii l li l K N K S R R F L N R S K I T V M R R P L
27205 tccaaaaccttcaagagcaactcgacaagcagtctcaacttgcaaaaggcgctgtga 27261
57 S K T F K S N S T S S L N L Q K A L *
dplORF091
43189 atgaaactatctaacgaacaatatgacgtagcaaagaacgtggtaaccgtagtcgttccagcagcgattgcactaattacaggt
1 M K Ii S N E Q Y D V A K N V V T V V V P A A I A L I T G
43273 cttggagcgttgtatcaatttgacactactgctatcacaggaaccattgcacttcttgcaacttttgcaggtactgttctagga
29 L G A L Y Q F D T T A I T G T I A Ii L A T F A G T V Ii G
43357 gtttctagccgaaactaccaaaaggaacaagaagctcaaaacaatgaggtggaataa 43413
57 V S S R N Y Q K E Q E A Q N N E V E * dplORF092
46989 atgaaaactatctccatattaaggaaagacactaaaaggaagccggacaggaacggaagaaaaactgcactcgaactagctcaa
1 M K T I S I Ii R K D T K R K P D R N G R K T A Ii E L A Q
47073 gagattgatatgtcacctagtgagttagcagagctccttcaaattcctgaaaggacggcaaccagaattttaaaactcgacaaa
29 E I D M S P S E Ii A E Ii li Q I P E R T A T R I Ii K L D K
47157 ctgctcaacaaagagcaatgctcaataatagaaaggtatataaatgaaattcactga 47213
57 L L N K E Q C S I I E R Y I N E I H * dplORF093
45756 atgcaacatacgattaaacaatgtttgaaacttgccttcctgctaactgcaatatcaattgcctgtttagttttccctaaacct
1 M Q H T I K Q C L K L A F L L T A I S I A C Ii V F P K P
45672 tgctcatcgcctaaaaggaaacatggatgctcttgtgcgtattcgaaacattcaacctggtgcgcgaatggagtagtcttgaac
29 C S S P K R K H G C S C A Y S K H S T W C A N G V V L N
45588 gaaaactgctcattgcttgaagaagctattcggtttcgagagtcaatgtag 45538
57 E N C S L L E E A I R F R E S * dplORF094
8281 atgtacgaattagttctatcactaaaattgacgccgacagcgccgatgtctcaagatgtcgaaaagtgcttcaaaaggctcaag
1 M Y E L V Ii S L K Li T P T A P M S Q D V E K C F K R Ii K
8365 tatattcagtggcggcaggtgaatgcattaaaattgcacacggatttgctcttgaacttcctaagggatatgaagcaatcttgc
29 Y I Q R Q V N A L K L H T D L L L N F L R D M K Q S C
8449 atcctcgttccagtctttttaagaaaactggtctaa 8484
dplORF095
8877 gtgggaaaactacttcagctctcgacattgtcaagaatgcgcaaatggtatttgagcaggaatgggaacagaagactgaagaac
1 V G K L L Q L S T L S R M R K W Y L S R N G N R R L K N
8961 tcaaggaaaagctggaaaatgcgcgtgcatccaaagctagcaagactgctgtcaaggaacttgaaatgcaactcgatagtcttc
29 S R K S W K M R V H P K Ii A R Ii li S R N L K C N S I V F
9045 aagagcctcttaagattgtatatcttgaccttgagaatacattag 9089
dplORF096
46681 gtgattcataaattcttcaatttcgttgaacttatctgcggtttctcctgttaccaggttgcatttgactgtcttcgaaagtat
1 V I H K F F N F V E L I C G F S C Y Q V A F D C Ii R K Y
46597 cttagcaagaggttcaataaσcttttcccaattgctaaatatcacgcaggactttccttgctggatacattcctcgacaatttc
29 li S K R F N N L F P I A K Y H A G Ii S L L D T F Ii D N F
46513 gatacatctttcgaacttgcaagacttgacatcttgagtagttaa 46469
57 D T S F E L A R L D I L S S * dplORF097
39100 atggacgggattgaaatcttgatactgaccgacgtatgctcgtccgctgtcagtatgactaaatccctcaccgtttggactatt
1 M D G I E I L I L T D V C S S A V S M T K S Ii T V W T I
39016 agagaaagcgaggtgagtatattgcgaacgtccgtcagctcctgcaggtccaggaattccttgaagcccttgaggaccttgaag
29 R E S E V S I L R T S V S S C R S R N S L K P Ii R T L K
38932 accttgaactcctctaggacctgtttcacctatcttggaaactga 38888
57 T L N S S R T C F T Y L G N * dplORF098
43627 gtgaaaatgctccgtgggatgctaaacgaggcgacatcttcatctggggacgcaaaggtgctagcgcaggcgctggaggtcata
1 V K M L R G M L N E A T S S S G D A K V Ii A Q A L E V I
43711 cagggatgttcattgacagtgataacatcattcactgcaactacgcctacgacggaatttccgtcaacgaccacgatgagcgtt
29 Q G C S L T V I T S F T A T T P T T E F P Ξ T T T M S V
43795 ggtactatgcaggtcaaccttactactacgtctatcgcttga 43836
57 G T M Q V N L T T T S I A * dplORF099
38298 atgcaagttcgccatctgctactgaagctccagctggtggatggtctacgcaagttcctaccgtcccaggtggtcagtatttat
1 M Q V R H Ii L L K L Q Ii V D G L R K F Ii P S Q V V S I Y
38382 ggactcgaacaagatggcgctacactgaccaaactgatgaaattggatattcagtttcaagaatgggcgagcagggtcctaaag
29 G li E Q D G A T L T K L M K Ii D I Q F Q E W A S R V L K
38466 gtgacgcaggtcgtgacggtattgcaggaaagaacggaatag 38507
57 V T Q V V T V L Q E R T E * dplORFlOO
1597 atgcagttgacaccaagcgagttctatttggatttagaactacggctgagaatatgtcaagattccttacctggactctcacgg
1 M Q L T P S E F Y L D L E L R L R I C Q D S L P G L S R
1681 agcttatgtggaagcatgctcgtatcgactctatcaaactatgggaaactcctacaggttgcgcagaatgtacttactacgaga
29 S li C G S M L V S T L S N Y G K Ii Li Q V A Q N V L T T .
1765 ttttcacagaagacgagattgaaatgttcaagaacgtaa 1803 - —
57 F S Q K T R L K C S R T * _ dplORFlOl
19220 gtgataattttagtccagttcccactacatttgaaagcgcgattaggtcatctaggctgtctagctcgagttcgattacaaggt
1 V I I Ii V Q F P L H L K A R Ii G H L G C L A R V R L Q G
19304 tgccagtatcaatttcacaaaagtaagcgacatttccaactttctctagtgcttcacgatacctatcatatgtcgcctcttcgt
29 C Q Y Q F H K S K R H F Q Ii S Ii V L H D T Y H M S P L R
19388 caaatagtcgcgcagaataaacttcgaatttcattttag 19426
57 Q I V A Q N K L R I S F * dplORF102
4034 atgataacgtgggaatgtttgactgtatcgccgaactcgataaaattcctggtgtatttagacagcctaagacacgtgaacagc
1 M I T W E C L T V S P N S I K F L V Y L D S L R H V N S
4118 ttttggaagcaccacaaatttcttgggataattatctatacatgcgcgagcgaatggttgagaaagacaagctcttacctattt
29 F W K H H K F Ii G I I I Y T C A S E W Ii R K T S S Y L F
4202 tccatatgggagaagactttaaatggctcaacttga 4237
57 S I W E K T L N G S T * dplORF103
49352 ttgaatcatagatatagtaacatcacaactatttttctttggcagattgtctttctttgtatttgctgcgcggtgtcctattgt
1 L N H R Y S N I T T I F L W Q I V F Ii C I C C A V S Y C
49436 gcaggagtgcataatgagcgagagtctcaagataaggtgattcaaagttataagcagaaagaaaagtcagccgtctacttgaca
29 A G V H N E R E S Q D K V I Q S Y K Q K E K S A V Y L T
49520 gtcgatagttcaggagcttggctaggaagtgctccgggagccaaggaaagtcctctctacaatgaaaagggacagcatgtagga
57 V D S S G A W Ii G S A P G A K E S P L Y N E K G Q H V G
49604 aaattgaaagaggtgggagagtga 49627
85 K L K E V G E * dplORF104
21427 atgagaaaaagagtgattttgaagctaaaaaggttgaactggtatgtccttaattcctactctcgaatggttgagtttttcgaa
1 R K R V I L K L K R L N Y V L N S Y S R M V E F F E
21343 cttttgaacttttcgaatggttcgacttttcgaaggattgaggttttcgaaccggttgagtttttcgagcattctcgacttttc
29 L L N F S N G S T F R R I E V F E P V E F F E H S R L F
21259 gacccctttctatgctcgacttttcgagtgttttga 21224
57 D P F L C S T F R V F * dplORF105
2028 atgatagtcgcatccaccagttcgaatgaaaatagtcttttgacctataaccattccttcaccttgaattgtaggaccgaaaat
1 M I V A S T S S N E N S L L T Y N H S F T L N C R T E N
1944 ttccatgataggcattttctcagggtcgcgaacattgattcgaatcttgcctctttcaggctgattgtattgattaaccattat
29 F H D R H F Ii R V A N I D S N L A S F R Ii l V L I N H Y
1860 cctgctcctgctctaaaatttcgcggacagtaa 1828
57 P A P A L K F R G Q * dplORF106
10529 atgaacctcgtcaatgatgtaaactttgaactcgctgtccatagacttgtatctagaatcttcaataatgtttcgaacattttc
1 M N Ii V N D V N F E Ii A V H R L V S R I F N N V S N I F
10445 taccccattattagaagcagcatcaatttcaataggagagccaagtcctttgttcacatccttcgcgaaaattcgagcagtagt
29 Y P I I R S S I N F N R R A K S F V H I L R E N S S S S
10361 ggttttaccagttccagcgccaccacagaatag 10329
57 G F T S S S A T T E * dplORF107
10750 atgagcgtgacgccctttcgtttattgggaaacttgcaaatggaggaatgcgtgacagtatcacaaggctcgaaaaagtccttg
1 S V T P F R Ii li G N L Q M E E C V T V S Q G S K K S L
10834 attatagtcatcacgttgacatggaagccgtttctaatgcactag 10878
29 I I V I T L T W K P F L M H * dplORF108
49447 atgcactcctgcacaataggacaccgcgcagcaaatacaaagaaagacaatctgccaaagaaaaatagttgtgatgttactata
1 M H S C T I G H R A A N T K K D N L P K K N S C D V T I
49363 tctatgattcaatttcgcttacctccaatcctcttacattgcttgcctgaaaatctagaaccactgaagtatcatatatacgac
29 S M I Q F R L P P I L L H C L P E N L E P L K Y H I Y D
49279 tataaagcctttggcctaaaaggtcaataa 49250
57 Y K A F G L K G Q * dplORF109
31632 atgtggttgtcgaagtcccaaatagttgattctccttcaactttccagcctttgaaagccttacctgttaaggtagggtcaact
1 M L S K S Q I V D S P S T F Q P L K A L P V K V G S T
31548 ggttttggagaaatcttcttacctgcttcaactcgaactgcgtcggcggttcctgttccaccgttcaaatcgaatgtcacgcga
29 G F G E I F L P A S T R T A S A V P V P P F K S N V T R
31464 cgaagaaccgctggaagttgtgccacatag 31435
57 R R T A G S C A T * dplORFHO
16444 atgatttcaattctagcatcaacttccatgtcgcgagtaagtgtgactccagtttcagcgacaggacatgctttgaatactgca
1 M I S I L A S T S M S R V S V T P V S A T G H A L N T A
16528 atgtcaagttcgctctttctaataactgagcctaggtctaagtacaagttaggattgattccagtgaccttatattgtttctca
29 M S S S L F L I T E P R S K Y K L G L I P V T L Y C F Ξ
16612 gtttcttttacaggaatgctttcatag 16638
57 V S F T G M L S * dplORFlll
28657 gtgactctatcaagaaagctcttgcaattggtgttcaaggttcttgggaaaacttcttgcttcttgcaagtgacgctgagaaat
1 V T L S R K L L Q L V F K V L G K T S C F L Q V T L R N
28741 tcatcgctgaaaaaacaggtcttcaaatcgctgtctactctaagaaaattgctcagttcgctgacgctgacaaacttcctgacg
29 S S L K K Q V F K S L S T L R K L L S S L T L T _N_ F~ if T
28825 ttggtaacattcgtcagttcaacttga 28851 _,
57 L V T F V S S T * dplORF112
32207 atgcaaactgatttaggcaaatactgcttcgacgcagcagccgttgcttatattagatatttgcaggaagacaagactcctagg
1 M Q T D L G K Y C F D A A A V A Y I R Y L Q E D K T P R
32291 tatcctggtgacgaaaagaaaaatccaggattgcaaatgcttatggagtga 32341
29 Y P G D E K K N P G L Q M L M E * dplORF113
17715 atgaaaacagttaaagaagcaatcaaacaattcggtgatgaatggtggtacgaaattatcaacgaaaacggccaaatgattcaa
1 M K T V K E A I K Q F G D E W W Y E I I N E N G Q M I Q
17631 gacggaagaatcgaagacatgggcgaatacatggaagaaacggtcgaccaagttaagttcatcaactatggtgacatcgaatct
29 D G R I E D M G E Y E E T V D Q V K F I N Y G D I E S
17547 caaattatcaaactatatatcgcataa 17521
57 Q I I K L Y I A * dplORF114
52952 atgctattggcgaagacggggaaacagtccatcctgataattgtccattatgccaaaacggattccctcgtattgaaaaactat
1 M L L A K T G K Q S I L I I V H Y A K T D S L V L K N Y
53036 ttcttcaactttacaaccatgatacgggaaaagttgaaacatgggaccgaggccgttcttatgttcaaaagattgttacattta
29 F F N F T T M I R E K Ii K H G T E A V L M F K R Ii li H Ii
53120 tcaataaatatggaagccttgtga 53143
57 S I N M E A L * dplORFllδ
5342 atgagcctcctttttttgatatatataatatacacgaatt tcgcgagtttgtaaagccgtttctaaataattttaaatctttt
1 M S L L F L I Y I I Y T N Y R E F V K P F L N N F K S F
5258 aagcatattgagttttgcttcataagtcccgttcacggcagcctcttgcattttgagtacaatgaaaggaggttcctcgatatt
29 K H I E F C F I S P V H G S L Ii H F E Y N E R R F Ii D I
5174 gttgaaactatagaaggtgaataa 5151
57 V E T I E G E * dplORFllδ
20662 atgaaattttcaaactttgctaaagcacttactaatgaatacctaatggtagtgaacaatgaccaagctgaagtcttaggcgca
1 M K F S N F A K A Ii T N E Y L M V V N N D Q A E V L G A
20578 ggaaatatcgaaaacattctcaacggttcgaactttgctaatgttgtagctgaagcgacagttttaaaactcgaaaaactcagc
29 G N I E N I li N G S N F A N V V A E A T V L K L E K L S
20494 gaagaggaagctattgagtag 20474
57 E E E A I E * dplORF117
24680 atgataacaggctgctcgaacattttaaatcgaagtgaatctcgtaagtcactaatagttttgttcaagttatctgctactgtg
1 M I T G C S N I L N R S E S R K S L I V Ii F K L S A T V
24596 ataaggtctttgacatcgcttgtcccgtatatgtcattagtcaatggttcattaagaataactcgacaaggaatttgcttcaag
29 I R S Ii T S L V P Y M S Ii V N G S Ii R I T R Q G I C F K
24512 ccggttggggcggattcttga 24492
57 P V G A D S * dplORFllβ
15023 atgatattatctacgtcgacgcaacttgtgaaactattaaatacgaggagcctattgcatgaacaatcagcgaaagcaaatgaa
1 M I Ii S T S T Q Ii V K Ii li N T R S Ii L H E Q S A K A N E
15107 caaacgaatcgtcgaacttcgcgaagactatcaacgtgcaagaggtcgaataaacttccttcttgctgtaaaggaccacggcga
29 Q T N R R T S R R L S T C K R S N K L P S C C K G P R R
15191 agaactcgaaaaccttga 15208
57 R T R K P * dplORF119
41054 atggaggttcaacatccccgattcagtacgtcctactttttcgggcatttctttagtagacacgacttcagcggttcgacagat
1 M E V Q H P R F S T S Y F F G H F F S R H D F S G S T D
41138 tttaacagggaacaacttcctccaaatcatgtcgaacattcaagtcaacttcaacaatgcttccggcgcttacggatccactat
29 F N R E Q L P P N H V E H S S Q L Q Q C F R R L R I H Y
41222 ccaagcatttcacgctga 41239
57 P S I S R * dplORF120
28387 gtgttgaagcgcaagcagaatacatgcgtatgcaattgcttcaatacggtaaattcactgtcaaatcaactaacagcgaggctc
1 V L K R K Q N T C V C N C F N T V N S L S N Q L T A R L
28471 aatacacttacgactacaacatggatgctaagcaacaatatgcagtcactaagaaatggactaacccagctgaaagtgacccta
29 N T L T T T T M L S N N M Q S L R N G L T Q Ii K V T L
28555 tcgctgacattttag 28569
57 S L T F * dplORF121
39222 gtgcagacggatcacgtgagttcagtttggaagataataatcaacaatatatgggttattactccgattatgagcaagcagata
1 V Q T D H V S S V W K I I I N N I W V I T P I M S K Q I
39306 gcagggatcgaactaagtatcgatggtttgaccgccttgccaatgttcaagtgggaggtcgaaacgagttccttaattctttat
29 A G I E L S I D G L T A L P M F K W E V E T S S L I L Y
39390 ttgaatttggtttaa 39404 dplORF122
40402 atgttattctccttatcctacataccgaatcacgttcatgtctggattaaacgagtattgttccgttctaaatcggccgacttg
1 M L F S L S Y I P N H V H V W I K R V L F R S K S A D L
40318 aatggattgggtaaagatcccgttatcgatgtgaatgaacccttgcgtaaggtacataacttcattccctgcggagaacataga
29 N G L G K D P V I D V N E P L R K V H N F I P- C G E— H ~R
40234 aattcggtcacttga 40220 →
57 N S V T * dplORF123
21327 atggttcgacttttcgaaggattgaggttttcgaaccggttgagtttttcgagcattctcgacttttcgacccctttctatgct
1 M V R L F E G L R F S N R L S F S S I Ii D F S T P F Y A
21243 cgacttttcgagtgttttgaggttttcgagcaggttcgacttttcgagaaattgagtttttcgacctctaaattaggctcgatt
29 R L F E C F E V F E Q V R Ii F E K Ii S F S T S K L G S I
21159 attcgaaaagtttag 21145
57 I R K V * dplORF124
17891 atggtaaaagttaaagatttgcaagtaggaatgaaagttgtaaatgcaaaaggtactgaatttaaagtaactgaccgtcaaggt
1 M V K V K D Ii Q V G M K V V N A K G T E F K V T D R Q G
17807 cgtaaatgggtaagcctagaacgtcttagtgatggacgtattcggttctatgataacgaatcactaatggacgaaaaagtggag
29 R K W V S Ii E R L S D G R I R F Y D N E S L M D E K V E
17723 gtagtaaaatga 17712
57 V V K * dplORFl25
49916 atgtcctcagccgcttccgttaaaattggaacaagtgaattatatagatgctcctcttttagcttgtcgataaggtattcatca
1 M S S A A S V K I G T S E L Y R C S S F S L S I R Y S S
49832 gtttcgccaatttcgaaaaattcgaatccaggaaaatggtcgagaatagtttcgtcgtccggaactcttccatatctcgaaaag
29 V S P I S K N S N P G K W S R I V S S S G T L P Y L E K
49748 tgttcttga 49740
57 C S * dplORF126
16136 atgagctcaagtacgttttctcgaacaatagggtcaagtccagttatatcaacgaactgtatatcgtcctcttgtataggaata
1 M S S S T F S R T I G S S P V I S T N C I S S S C I G I
16052 aggtctgcgtacagttgcatggctgaccctttaattggagtaactgttccttcactgtttattttaaataaggttatcatttct
29 R S A Y S C M A D P L I G V T V P S L F I L N K V I I S
15968 atcctctaa 15960
57 I L * dplORF127
13511 atgctaaatagctttcccattcaccgtcgctgttcttgcgccatttttcagtttcacgatactgaccaactttgcaaaggtcgt
1 M L N S F P I H R R C S C A I F Q F H D T D Q Ii C K G R
13427 gaaatagtgctacgattgcaactgtttccattgggt aaatgt cttcccagcctttgcctaccatggtatccatttcgaaaagta
29 E I V L R L Q Ii F P L G K C L P S Ii C L P Y P F R K V
13343 gttgattga 13335
57 V D * dplORF128
4852 atgacagcagttcaacaagttaagttctacttagaagaagccggcgctcactttctaaaagatgttgagtacagtgacaactta
1 M T A V Q Q V K F Y L E E A G A H F L K D V E Y S D N L
4936 gagcaagcaattatgaaagatattcttaaatggaatggcgctcatagagatgagcacgatatgaaaataacttcatacgaagta
29 E Q A I K D I L K W N G A H R D E H D M K I T S Y E V
5020 ttatag 5025
57 L * dplORF129
25133 atgaactttctgctaagcaacttgcgctcactgaagttcaaactaatgtacgcagccaccaatcttacattgaagaattcagta
1 M N F L L S N L R S L K F K L M Y A A T N L T L K N S V
25217 agaaggaaaaggcggacaaggaatgggaacgcattttggaagaacttgctcagcttgacgaaatctcagctggagcattgcctg
29 R R K R R T R N G N A F W K N L Ii S Iι T K S Q -i E H C -i
25301 tattag 25306
57 Y * dplORF130
16789 gtgcttgactttattcctttattatcgtataatcataatataaataaaacaagcgtcaaggacgcagaaagaggtcaattatgg
1 V L D F I P L L S Y N H N I N K T S V K D A E R G Q L W
16705 aaacaacactttatttcggttatcttacagcagattggaaagacggtcacaagaactacactttccactatgaaagcattcctg
29 K Q H F I S V I L Q Q I G K T V T R T T L S T M K A F L
16621 taa 16619
57 * dpl0RF131
43846 atgctcaaccggctgagaagaaacttggctggcagaaagatgctactggtttctggtacgctcgagcaaacggaacttatccaa
1 M L N R Ii R R N Ii A G R K M Ii li V S G T Ii E Q T E Ii l Q
43930 aagatgagttcgagtatatcgaagaaaacaagtcttggttctactttgacgaccaaggctacatgctcgctgagaaatggttga
44013
29 K M S S S I S K K T S L G S T Ii T T K A T C S Ii R N G * dplORF132
15304 gtgactggaaggtcatctaatacacatagcctcaagacatttcgttggctttcaggaaaacattcgactagattgtcaatgtat
1 V T G R S S N T H S L K T F R W Ii S G K H S T R L S M Y
15220 cccacaaaggcttcaaggttttcgagttcttcgccgtggtcctttacagcaagaaggaagtttattcgacctcttgcacgttga
15137
29 P T K A S R F S S S S P W S F T A R R K F I R P L A R * dplORF133
8061 atgacttcttcattcatgacaagttttcgagtttctgcttgcttgtcaggaatagttttcccggcggctaaaatgtatagatta
1 M T S S F M T S F R V S A C L S G I V F P A A~K M_Y T. L
7977 tcgtatttttctttcctgatagcagaacttgaatccatttgtattcccaccatttccgccctatctgcggcβfaaataa 7900
29 S Y F S F L I A E Ii E S I C I P T I S A L S A A K * dplORF134
498 atgacttcaatgtacttaggttccatcaattcatacaagtcattcaaaataatgttcatgcaatcttcgtggaagtcaccgtgg
1 M T S M Y L G S I N S Y K S F K I M F M Q S S W K S P W
414 ttacggaaactgaataagtacaatttcaatgatttagattcaaccatcttttcgtttggaatgtaa 349
29 L R K L N K Y N F N D L D S T I F S F G M * dplORF135
780 atgaagcagaacttgaaaatgctgctaatgttgcaatgttctacggagtcaagttcaccattcttgaaattgactcgaaaatct
1 M K Q N Ii K M L L M L Q C S T E S S S P F L K L T R K S
864 actcaagctctagctcttccttattacaaggaaaaggcgaaatttcacatggaaaatcttacgctgaaatcctag 938
29 T Q A L A L P Y Y K E K A K F H M E N Ii T L K S * dplORF136
55252 gtgaagaaatcttcaataaccttattcgcttctttgacagatacattcatctgctcagcgattgagttagccccgcggccgtac
1 V K K S S I T L F A S L T D T F I C S A I E L A P R P Y
55168 ataagacctaaaagaacggacttgacagaatttcttcgaagttttccttccttgttagtcgttccgtcgggatag 55094
29 I R P K R T D L T E F L R S F P S Ii li V V P S G * dplORF137
37146 atgcttcgaacttgtttgttagcaccgtcaggaggacaaactagtcgaacccattcacctgcgtctttgataatatctagcgcg
1 M li R T C L Ii A P S G G Q T S R T H S P A S Ii l l S S A
37062 acagcgcctacagaagaagcaacgtgtttcaacttcctaggcaagccttctgctagttcataccataatgcgtag 36988
29 T A P T E E A T C F N F L G K P S A S S Y H N A * dplORF138
30662 atgactatatcgaagaacaatgtagtcatccggcctatctgtatcttgctcgtcaaattcaactcctggaagcataggagcagg
1 T I S K N N V V I R P I C I L L V K F N S K H R S R
30578 cgagagctgaaatgtaggaagaatttccttcaatctgtccatcattgtcgttcgtttagtcatgttcactcctag 30504
29 R E L K C R K N F L Q S V H H C R S F S H V H S * dplORF139
12092 atgatactaaatcactcaacttgtttgaccctcctgataaattcgttcacgcagacacgcgcatttgagccctttttagatacc
1 M I L N H S T C L T L L I N S F T Q T R A F E P F L D T
12008 tttcgcaaacacctagatgcttccctcactaaaaggtcatgggcctcaagttcttcgaaagacatttctacatag 11934
29 F R K H L D A S L T K R S W A S S S S K D I S T * dplORF140
20562 atgttttcgatatttcctgcgcctaagacttcagcttggtcattgttcactaccattaggtattcattagtaagtgctttagca
1 M F S I F P A P K T S A W S F T T I R Y S Ii V S A Ii A
20646 aagtttgaaaatttcattttattttccctttatttgtttttctttatactattattatacaataatgattga 20717
29 K F E N F I L F S L Y Ii F F F I Ii -i li Y N N D * dplORF141
42922 gtgctaagagttgtagagatatcctctaaaacgctcttggctttattcgatttccattcgaataacttatttagtaggacagta
1 V Ii R V V E I S S K T L Ii A Ii F D F H S N N I- F S R T V
42838 agcactccgctgcacgctgtaataatcgtcgtcaagactgctgtgtcgtttagccacattggcatagattga 42767
29 S T P L H A V I I V V K T A V S F S H I G I D * dplORF142
31898 gtgactgtcgaagtttctccaaacagttctgtcactttacctaaaagcgtattagggattttcccgttagcgattaggttcatg
1 V T V E V S P N S S V T L P K S V Ii G I F P L A I R F M
31814 acacctgctgctcgaattttaacatggataggttcactaccttttgaaaatcctggaagtgcgatgatttga 31743
29 T P A A R I Ii T I G S L P F E N P G S A M I * dplORF143
7565 atgaagtttgggttgacgcttttaactccagaccgtttaattttttcaaggcttgaaattggataccatataatcttttcatgc
1 M K F G L T L L T P D R L I F S R L E I G Y H I I F S C
7481 ttttggaaatacactaaaattccggcgagaataaatttgcatccatctgcgcgtgatagctggaaccattga 7410
29 F W K Y T K I P A R I N L H P S A R D S W N H * dpl0RF144
36517 gtgcaaatcaagcgactaacttatttagatacattaaacgaggcgcattcttcaagattcctaatggaaattcaacaattacca
1 V Q I K R L T Y L D T L N E A H S S R F L M E I Q Q Ii P
36601 ttgaataccgagccgatgacgcagcagcttggacctctactcttcccgctcaagttgaactgtttctaa 36669
29 L N T E P M T Q Q Ii G P L L F P L K Ii N C F * dplORF145
42067 atggaaacagctggagacctaacaagtggaaagaggttctatttaagcaagacttcgaacagaataattggcagaaacttgttc
1 M E T A G D Ii T S G K R F Y L S K T S N R I I G R N Ii F
42151 ttcaaagtgggtggaaccatcactcaacctatggcgacgcattctattcgaaaactcttgacggcatag 42219
29 F K V G G T I T Q P M A T H S I R K L Ii T A * dplORF146
51484 atgacaaactgcatgattgcatcacctttccagtacggaacctcaagggcgaaacagtattcttcaaccgtcgaagtgttcgtt
1 M T N C M I A S P F Q Y G T S R A K Q Y S S T V E V F V
51568 ctaagtttcaccagtacggtgaagatgaccctaaaacggaatttctttatggccaatatgagcttgtag 51636
29 L S F T S T V K M T L K R N F F M A N M S Ii * dplORF147
55207 atgtatctgtcaaagaagcgaataaggttattgaagatttcttcaccgagttccctaaagtggcagactatatcatattcgttc
1 M Y L S K K R I R L Ii K I S S P S S Ii K W Q T I S Y S F
55291 aacagcaggcgcaggacttgggatatgttcaaacagctaccggtcgaagaagaaggcttcctgatatga 55359
29 N S R R R T W D M F K Q L P V E E E G F L I * dplORF148
28636 gtgtttcggttcaagaccattcgagtagggcgaacacctgtacgattttcgatgtcatccattgctgctaaaatgtcagcgata
1 V F R F K T I R V G R T P V R F S M S S I A A ~K M— S- "j. I
28552 gggtcactttcagctgggttagtccatttcttagtgactgcatattgttgcttagcatccatgttgtag 28-484
29 G S L S A G L V H F L V T A Y C C L A S M L * dplORF149
26474 atgccattgaacttttcgagcataaggattaaccttgccccattgtctcactccagctgtggcggaatggctaatggtagttcg
1 M P L N F S S I R I N L A P L S H S S C G G M A N G S S
26390 agcaagtcgaagggcattgtattcgagattttgatatttatgagcagcaggtttccctag 26331
29 S K S K G I V F E I L I F M S S R F P * dplORF150
15185 gtggtcctttacagcaagaaggaagtttattcgacctcttgcacgttgatagtcttcgcgaagttcgacgattcgtttgttcat
1 V V L Y S K K E V Y S T S C T Ii l V F A K F D D S F V H
15101 ttgctttcgctgattgttcatgcaataggctcctcgtatttaatagtttcacaagttgcgtcgacgtag 15033
29 li li S L I V H A I G S S Y L I V S Q V A Ξ T * dplORF151
28027 atgattatatcaacgcaggggagattgctagctacattcaagcacttccttcaaacgctcttcaataccttggaccaactcttt
1 M I I S T Q G R L Ii A T F K H F L Q T Ii F N T L D Q Ii F
28111 tccctaatgctcaacaaacagggacagacatttcatggctcaagggtgcaaataatttgccagtaa 28176
29 S li M Ii N K Q G Q T F H G S R V Q I I C Q * dplORF152
42235 atgtgcataaaggacttatcgacaaagaggctactattgcagtacttcctgaaggatttagaccgaaagtttcaatgtatcttc
1 M C I K D L S T K R L L L Q Y F L K D L D R K F Q C I F
42319 aggctctcaataactcatatggaaatgccattctatgtatatacactgacggaagacttgtggtga 42384
29 R li S I T H M E M P F Y V Y T L T E D L W * dplORF153
22307 atggtggacaaagggctcaccttttcgaactttcgatatcgtcatagcagacggttccattcgttcaggaaaaacagtatcgat
1 M V D K G L T F S N F R Y R H S R R F H S F R K N S I D
22391 ggctctttcattttccctttgggccatgacggaattcaacggacaaaactttgccatctgtggtaa 22456
29 G S F I F P L G H D G I Q R T K Ii C H Ii W * dplORF154
18446 gtgacaataggctttaagaactgcaaaaaaacctggggcgtctgcacgcgcaacctggagctccttaacagtcatccaaggctg
1 V T I G F K N C K K T W G V C T R N Ii E L Ii N S H P R L
18530 aggtttcttacaaacaatcctaattccttcaaaatagctcttgtccgggtcaatagtgcctaa 18592
29 R F L T N N P N S F K I A L V R V N S A * dplORF155
13512 atgaatacgaccctgagcaacttacaatgggacatggtgcaaaatctaatttccttcttcaacgtttcattcaactcacgccag
1 M N T T L, S N L Q W D t. V Q N Ii I S F F N V S F N S R Q
13596 ttgaagctcaagcaattttctggcatatgggagcctatgatattagtccttatgcaaatttga 13658
29 li K L K Q F S G I W E P M I Ii V Ii M Q I * dplORF156
18777 atgctagtatctccatttctgttggtcttgctttttagctctgttcagttcagctgcttctcgcgatgcaatagtttcgagaat
1 M li V S P F Ii li V Ii L F S S V Q F S C F S R C N S F E N
18861 atgcctgttcataggctcacaatattccgccaaagatttgccagttatggtggcgtcaattaa 18923
29 M P V H R Ii T I F R Q R F A S Y G G V N * dplORF157
13281 gtgcttgctggacttgagaagaaattggtatcattttcgagccaatccataaggttctcgataccgtcacgattgattgtttct
1 V -i A G Ii E K K L V S F S S Q S I R F S I P S R Ii l V S
13197 gttactgctttcttgaagcgttttttaaagtctgtcatattagacccctttcattttctataa 13135
29 V T A F L K R F L K S V I L D P F H F Ii * dplORF158
40727 gtgaacgccgttattagggtcaaacgaagcccaaacggacattgtctttgtcccgtcactattgtgaggaacagtcacttctcc
1 V N A V I R V K R S P N G H C L C P V T I V R N S H F S
40643 acttgcgagcgttacctcttcgccggacgtgtcgtagtctgggtgactgctatgaacacttga 40581
29 T C E R Y L F A G R V V V W V T A M N T * dplORF159
30371 atgatttggtctgcgcttacccaagcagcttctcctttgagtttctgtcgagcattccctgtacggtctgtccaaatagcatgc
1 M I W S A L T Q A A S P L S F C R A F P V R S V Q I A C
30287 gtctttgcgtattcttccatcttagtagcagcgacttcgcagactgttatgacagcgacttga 30225
29 V F A Y S S I L V A A T S Q T V M T A T * dplORF160
41324 atgggttacagacacgcgaggaaaacaatcgaacgtccaagacgtatctatcaatgttatagaatactatggaccgtctatcaa
1 G Y R H A R K T I E R P R R I Y Q C Y R I L T V Y Q
41408 tttctccgttcaacgtactcgtcaaaatcctgcaattatccaagctcttcgaaatgctaa 41467
29 F L R S T Y S S K S C N Y P S S S K C * dplORF161
52175 atgcaaaaaggtttaaatgcttatctcgacatgacattgaaagcattgcattcgagactatttcaaaatgtttggcaacgttca
1 Q K G Ii N A Y L D M T L K A L H S R L F Q N V W Q R S
52259 aatcaaaccaaggggccaagttttcaacttaccttacaagactcttcaagaatagaatag 52318
29 N Q T K G P S F Q L T L Q D S S R I E * dplORF162
13020 atgacagaagttgcggtaaatagcccgcaaaaggtgagagtagttatggtcgggaatattgaatttctcgaatatttaaaaagg
1 M T E V A V N S P Q K V R V V M V G N I E F Ii E Y L K R
13104 aagtacggaacagaaacttccatcagttatattatagaaaatgaaaggggtctaatatga 13163
29 K Y G T E T Ξ I S Y I I E N E R G L I * dplORF163
40224 gtgaccgaatttctatgttctccgcagggaatgaagttatgtaccttacgcaagggttcattcacatcgataacgggatcttta
1 V T E F Ii C Ξ P Q G M K Ii C T L R K G S F T S I T G S L
40308 cccaatccattcaagtcggccgatttagaacggaacaatactcgtttaatccagacatga 40367 - --
29 P N P F K S A D L E R N N T R L I Q T * _. dpl0RF164
6696 atgtactcttggagaacttcgtgcctaaatgttccagcttcgcccattgcaattaggttagaatctgcgttatctataatagac
1 M Y S R T S C L N V P A S P I A I R L E S A L S I I D
6612 tcaccgattctttcgaaatacatttttcgaatacatccaccaaccccgctgggcttataa 6553
29 S P I L S K Y I F R I H P P T P L G L *
dplORF165
50504 atgagtgaaagctggtcaatccccaccacagatggtctatatttagatatcatgctatctaaaattgcaggggtaaggttcttt
1 M S E S W S I P T T D G L Y Ii D I M Ii S K I A G V R F F
50420 cctccaatcataaagggcgtgactaccacaagggaattttcagcctcagtcattgcttga 50361
29 P P I I K G V T T T R E F S A S V I A * dplORF166
23519 gtggtcatgctctttaatgactctatcttctcccgtttggctcgctttactgtcccagctgtaagcatagtattcatcaatgtc
1 V V M L F N D S I F S R L A R F T V P A V S I V F I N V
23435 gtgcgtgttgctagggtcgagtgtaaatctattctcagccaagagttcagcgtgaaatga 23376
29 V R V A R V E C K S I L S Q E F S V K * dplORF167
1008 atgcttattcggttggagcttcttacgtcgtatatggtgctcacgcagacgatgcggctggaggtgcttaccctgattgcactc
1 M L I R Ii E Ii L T S Y V -i T Q T M R Ii E V L T Ii l A L
1092 ctgagttctataattcaatgtcaaatgcaatggaatatggaactggaggcaaggtaa 1148
29 L S S I I Q C Q M Q W N M E Ii E A R * dplORF168
54345 atgagactttttccaggttatattcttcacattgttcagttcctggagtcaagtattgttcttgaaattcatagagttcgaaag
1 M R Ii F P G Y I H I V Q F L E S S I V Ii E I H R V R K
54261 tttgcaaagggtcataggccgcatacatataggcaacatcaggaggaattaaactaa 54205
29 F A K G H R P H T Y R Q H Q E E L N * dplORF169
45954 atgaacacagcatcgcgaagagtttcaatgttagtgataaggaagaattcgtcgtggccaccaagcaagtcttctgcccgttta
1 M N T A S R R V S M L V I R K N S S W P P S K S S A R Ii
45870 gaaactccgtcaatcactaatttcccatctttagtgactcgacttcctaaaatatga 45814
29 E T P S I T N F P S L V T R Ii P K I * dplORF170
27600 atgatgattgttcttgtgctcctgccgtttgttgagcagcagcaagttgcttaccaaaagagccgatttcacgaggttcgggaa
1 M M I V Ii V Ii li P F V E Q Q Q V A Y Q K S R F H E V R E
27516 caccaccaccgacacgacctggatttcctaaatttccagtcccggctggcgacttag 27460
29 H H H R H D D F Ii N F Q S R Ii A T * dplORF171
47678 atgtcattttctttcatgtactcttttagagcatcacgaagacttttgacttgtttctccatgtcgcctttggtagcatttaat
1 M S F S F M Y S F R A S R R L Ii T C F S M S P L V A F N
47594 tcaccggcttcttcaattgcagcgatgaactgtttttcatcttcaaatttcatttaa 47538
29 S P A S S I A A M N C F S S S N F I * dplORF172
10462 atgtttcgaacattttctaccccattattagaagcagcatcaatttcaataggagagccaagtcctttgttcacatccttcgcg
1 M F R T F S T P Ii L E A A S I S I G E P S P L F T S F A
10378 aaaattcgagcagtagtggttttaccagttccagcgccaccacagaatagatag 10325
29 K I R A V V V L P V P A P P Q N R * dplORF173
32160 atgacattagacatttccttcgtctgtacgaaaggtttcagcttgagtcacttcaccgtacattgcactgaagattgtcataag
1 M T L D I S F V C T K G F S L S H F T V H C T E D C H K
32076 ttgctcatctgtcatatactcgccgacttcagcgtaagtaggctctaccattga 32023
29 L li l C H I L A D F S V S R Ii Y H * dplORF174
29766 atgtcccatcagcccttttcattaagattgtcgaaccagcgttcgacttttcatcagtttcaagctgttcttgcttatattggt
1 M S H Q P F S L R L S N Q R S T F H Q F Q A V L A Y I G
29682 cataatagaattgcgccatttgtttccagtagtctgcgtcaccttttagactga 29629
29 H N R I A P F V S S S L R H L L D * dplORF175
15648 atgcgcgtgatgtcatggcagataggcgaggataaagagtgtcgaatagaacgccgcagagcttacgagagcgccaaatacaag
1 M R V M S W Q I G E D K E C R I E R R R A Y E S A K Y K
15564 ggcgacggtactacggtggtcctcttgcttacctgtaaccaaataaaccattga 15511
29 G D G T T V V L L L T C N Q I N H * dplORF176
43031 gtgataaagacggtaacgttgaatttttctagttccgtcttgaatgacgtcattttggtgattgattgctactgtcgtttggtc
1 V I K T V T L N F S S S V Ii N D V I Ii V I D C Y C R L V
42947 aatcccgtcgacctgctgtttaagagtgctaagagttgtagagatatcctctaa 42894
29 N P V D L Ii F K S A K S C R D I L * dplORF177
19937 atgaacctaaacagttcgagacttctcaagctgttgggaaagaagcaggtcgaatattttggtgggaacgtgaacttggtcata
1 M N L N S S R Ii L K Ii li G K K Q V E Y F G G N V N Ii V I
19853 ttctcgcgactaattttaggtgcttttgtattaatcagcgtgatatgcgcttga 19800
29 F S R L I L G A F V L I S V I C A * dplORF178
11924 atgacaactgtcgaccaatttaaaagacagttgaggaaaagtttaggctcaatttttccttcatcagtttccttaaatttgagc
1 M T T V D Q F K R Q L R K S L G S I F P S S V- S L_N - S
11840 caattagtaacctttagcgaattgctagcacttgcctcccatattaagtcataa 11787 _,
29 Q li V T F S E L L A L A S H I K S * dplORF179
56058 atgggtagggttattccttacctcgttgatttgctttatgcaaaacctaccacaatcgcttgtcgtggcttcaggagttgcatt
1 M G R V I P Y L V D Ii L Y A K P T T I A C R G F R S C I
56142 ttggataagtcaaaaagcaagtgtctttatattcgacaagctctcgaataa 56192
29 L D K S K S K C Ii Y I R Q A Ii E * dplORF180
41176 atgttcgacatgatttggaggaagttgttccctgttaaaatctgtcgaaccgctgaagtcgtgtctactaaagaaatgcccgaa
1 M F D M I R K Ii F P V K I C R T A E V V S T K E M P E
41092 aaagtaggacgtactgaatcggggatgttgaacctccatccgtttgaatag 41042
29 K V G R T E S G M L N L H P F E * dplORFlβl
13126 atggaagtttctgttccgtacttcctttttaaatattcgagaaattcaatattcccgaccataactactctcaccttttgcggg
1 M E V S V P Y F L F K Y S R N S I F P T I T T L T F C G
13042 ctatttaccgcaacttctgtcataggctgtcctcctttgcttatactgtaa 12992
29 -. F T A T S V I G C P P L L I Ii * dplORF182
45369 gtgcttgcccatgtttcaataaatagggttcgacctcgcctagctttcgaacgtgctataacgatttcaatcatagcgaagaaa
1 V li A H V S I N R V R P R Ii A F E R A I T I S I I A K K
45285 ggtgagaagcttcaatcaattccattgcggtgtcaatatcttcttccttga 45235
dplORF183
13896 gtgattccagcttttggtttttcttcagcctcttcaactttttcttccttaggcgcaggtttcttacgagttgaactcttaggt
1 V I P A F G F S S A S S T F S S L G A G F Ii R V E L L G
13812 ttttcttcaactacttcttcaacctcagcctcttgttcaactggaccttga 13762
29 F S S T T S S T S A S C S T G P * dplORF184
53330 gtgaacttgccgtcaaccacgtcaaacatttggtcttcgtcgaggtctaaaattagagttccaagaagttcgctcttttctgga
1 V N L P S T T S N I S S S R S K I R V P R S S L F S G
53246 aaatcttcaagagtagcactgtcttccggacgctctggaaggaattcataa 53196
29 K S S R V A L S S G R S G R N S * dplORF185
22522 atgaaattcgagatgttcgaaatgaaaatctacttattattagacactttagaaatggcgaagaaattgtcaactacttctata
1 M K F E M F E K I Y Ii li li D T L E M A K K Ii S T T S I
22606 tatttggaggaaaagatgagtcgagtcaagaccttatacagggggtaa 22653
29 Y L E E K M S R V K T L Y R G * dplORF186
21272 atgctcgaaaaactcaaccggttcgaaaacctcaatccttcgaaaagtcgaaccattcgaaaagttcaaaagttcgaaaaactc
1 M L E K L N R F E N L N P S K S R T I R K V Q K F E K L
21356 aaccattcgagagtaggaattaaggacataccagttcaacctttttag 21403
29 N H S R V G I K D I P V Q P F * dplORF187
34415 atggtcttgttcaatctcttcctactatcattcaagcagctgttcaaattatcactgctttattcaatggtcttgttcaggcac
1 M V L F N L F L L S F K Q L F K L S L L Y S M V L F R H
34499 ttcctacgcttattcaagcaggtcttcaaattttgtcagctctcataa 34546
dplORF188
35609 atgttcgtaaagcagccggttcgcctcgagtggacttgttcaatacaggaagtgacaaccctaaccaacctcagtcacaatcta
1 M F V K Q P V R L E W T C S I Q E V T T L T N Ii S H N L
35693 aaaacaatcaaggcgagcaaaccgttgtcaacattggaacaatcgtag 35740
29 K T I K A S K P L S T L E Q S * dplORF189
42587 atgcaaacgcagtatcaaccgtctctgaaactcttcatgacccagacttgtatgctgcgaaccgtcgagaactt cgagctgacg
1 M Q T Q Y Q P S Ii K L F M T Q T C M Ii R T V E N F E L T
42671 agcaaaaacttcgcgaaactcgttacgcaatcgaagatgaaattctag 42718
dplORF190
39786 atgtattcactcaaagttgttcagtgtggctcaatcatattaaaatcgaacttggtaatatctctactccttttagtgaagcag
1 M Y S L K V V Q C G S I I L K S N L V I S Ii li L Ii V K Q
39870 aggaagaccttaaatatcgaattgactcaaaagccgatcaaaagctaa 39917
29 R K T L N I E L T Q K P I K S * dplORF191
40996 atgtccattgttccggaacttgatttaggtaagtaccttgctaagtccagtgacggcgtaaaggatacgctagtagtatggttc
1 M S I V P E L D L G K Y Ii A K S S D G V K D T L V V W F
40912 ttacctaaatctatccagtcgctaccgaaaactcggtaccaaacttga 40865
dplORF192
2920 atggtcgacgtcgaatgttttttcgagatgaagtttagggtcttctcgataccctacggtatgttcagcgagtgctttaacaaa
1 M V D V E C F F E M K F R V F S I P Y G M F S E C F N K
2836 acggaatggagtatcttgcaacccgtcacgttctgcgtcctcgcctaa 2789
29 T E W S I L Q P V T F C V L A * dplORF193
42456 atgatttcagctcaaattaaatacgaaatgagacattgtctaaatttaaccaagaattatctacattcgattt_caceacaagtc
1 M I S A Q I K Y E M R H C L N L T K N Y I, H S I, S P Q V
42372 ttccgtcagtgtatatacatagaatggcatttccatatgagttattga 42325
29 F R Q C I Y I E H F H M S Y * dplORF194
40284 atgaacccttgcgtaaggtacataacttcattccctgcggagaacatagaaattcggtcacttgataccttaatggtagagcta
1 M N P C V R Y I T S F P A E N I E I R S L D T Ii M V E L
40200 ccgtcgttcttaccgataattagaccttcattagaagagctcatgtaa 40153
29 P S F L P I I R P S L E E L M * dplORF195
42584 atgttcacaatcgttgttttgacaagtttcttttcagctccttgtccaatagtgaactctgccacaatttggcgcgattttgta
1 M F T I V V L T S F F S A P C P I V N S A T I W R D F V
42500 aggttcaacatagttctcacctcctttctaaaaaatattataacatga 42453
29 R F N I V L T S F L K N I I T * dplORF196
11273 atggtagatttaacaagtccctgtccaatcatgtcactcc ccttgctcatcaaaagaagtttggtttcaattatcggtttagc
1 M V D L T S P C P I M S L L L A H Q K K F G F N Y R F S
11189 attaggctcccatttaacaactccagcaagttcattcatttcttctag 11142
29 I R L P F N N S Ξ K F I H F F * dplORF197
7484 atgaaaagattatatggtatccaatttcaagccttgaaaaaattaaacggtctggagttaaaagcgtcaacccaaacttcatcg
1 M K R L Y G I Q F Q A L K K L N G L E L K A S T Q T S S
7568 atgcagggtatgaagtttcttacaagaagcgtcgaactagattga 7612
dplORF198
24119 atgccgctcaacaaattgacgtccagttttattcaatgcctcagttcacctatacagttgaccctagaaacccttccagcttgc
1 M P Ii N K Ii T S S F I Q C L S S P I Q L T Ii E T L P A C
24203 tttctgttgacattgtttatcaggacgagcgtacaaaaggaatga 24247
dplORF199
15742 gtggctcctgaattaggctgtacttttcctcccaactgcttagcaactgccttctcttgtttagcactagctctgcgcgtggga
1 V A P E L G C T F P P N C L A T A F S C L A L A L R V G
15658 attggtttgtatgcgcgtgatgtcatggcagataggcgaggataa 15614
29 I G L Y A R D V M A D R R G * dplORF200
47843 atgacaggcttgtattcgataagccctgaaagtttttcacacatttcttccgtctcggcttcgtcaactaatttttcgataatt
1 M T G L Y S I S P E S F S H I S S V S A S S T N F S I I
47759 tctttcaagcgttcttcgtccatagttgagcgctctgtcgtgtag 47715
29 S F K R S S S I V E R S V V * dplORF201
38569 atgggcttcacaagttccttctttaatcaaaggtcaatatctttggactcgaactatttggacctataccgattcaactaccga
1 M G F T S S F F N Q R S I S L D S N Y L D L Y R F N Y R
38653 aacgggctatcaaaaaacctacattccaaaagacgggaatga 38694
29 N G L S K N L H S K R R E * dplORF202
44483 gtggggcgtttattttttataaaaattttttacaaaatgcttgacaacattcactcattatcgtataatacaattataaaaata
1 V G R L F F I K I F Y K M Ii D N I H S L S Y N T I I K I
44567 aataaagccgaaaggcgaggaggacattatgtcaaaaattaa 44608
29 N A E R R G G H Y V K N * dplORF203
22781 gtgattaggattggccgggttacaagagaaccacattttcgaacctgttacggaacagcgccctgtcgcttggttgacaaacga
1 V I R I G R V T R E P H F R T C Y G T A P C R L V D K R
22697 ttcaggcatcagtgccacctcatcacagaagatacctgctaa 22656
29 F R H Q C H L I T E D T C * dplORF204
1471 atgaccacggttcgagtcaagggatggttgttgacttttatcacgtcaagaaaatcgcaggtacattcattgacagacttgacc
1 M T T V R V K G W Ii L T F I T S R K S Q V H S L T D L T
1555 acgctgttcttcttcaagggaatgaaccaatcgctttag 1593
29 T L F F F K G M N Q S L * dplORF205
8524 gtgacactgatgaatggttctcagtttggtatgctactcgtgacgcagatatcttctacgaccaaagaattgcccaatttagaa
1 V T L M N G S Q F G M L L V T Q I S S T T K E L P N L E
8608 ttcaggaaaagcaacctgctatcaagttcaatttcgtag 8646
29 F R K S N L L S S S I S * dplORF206
19855 atgaccaagttcacgttcccaccaaaatattcgacctgcttctttcccaacagcttgagaagtctcgaactgtttaggttcatc
1 T K F T F P P K Y S T C F F P N Ξ L R S L E I. F R F I
19939 aaattgttcaacttgagcaagtgcgatattattctttag 19977
29 K L F N L S K C D I I L * dplORF207
27502 gtgtcggtggtggtgttcccgaacctcgtgaaatcggctcttttggtaagcaacttgctgctgctcaacaaacggcaggagcac
1 V S V V V F P N L V K S A L L V S N L L L L N K R Q E H
27586 aagaacaatcatcattctttaaataataggaggaactaa 27624
29 K N N H H S L N N R R N * dplORF208 - —
47279 atgtttggtatgaagcaaaagacttcgctgaagaaaataacattcacttcccgtttgttcttcctgaacctagaacagaccttg
1 M F G M K Q K T S L K K I T F T S R L F F L N L E Q T L
47363 accatcgtggttctcgattctgggatgacgaaggcgtga 47401
29 T I V V L D S G T K A * dplORF209
29784 atgttaagaatcaagttcgtagagccattgaaaccgctcctactaaaatcaaggtacttcgaaactcttgggtcagtgatggat
1 M L R I K F V E P L K P L L L K S R Y F E T L G S V M D
29868 atggaggaaagaaaaaggataaagcgaatgaagtcgtag 29906
29 M E E R K R I K R M K S * dplORF210
53077 atgtttcaacttttcccgtatcatggttgtaaagttgaagaaatagtttttcaatacgagggaatccgttttggcataatggac
1 M F Q L F P Y H G C K V E E I V F Q Y E G I R F G I M D
52993 aattatcaggatggactgtttccccgtcttcgccaatag 52955
29 N Y Q D G L F P R L R Q * dplORF211
20959 gtgctcgacttttatgtcgcccctaatttttgtttttacttacggactatgggatttgtaggtattttcagggcgcttttttat
1 V L D F Y V A P N F C F Y L R T M G F V G I F R A L F Y
20875 ttacttattaagtccttttctatattagattgtttataa 20837
29 L L I K S F S I L D C L * dplORF212
52983 atggactgtttccccgtcttcgccaatagcattgcaattgatatagcgtcgacgaccgtcaacgtctgcttcgtggactacgaa
1 M D C F P V F A N S I A I D I A S T T V N V C F V D Y E
52899 ataatccatgtcttcgccttccgggtcatcatacaatag 52861
29 I I H V F A F R V I I Q * dplORF213
30291 atgcgtctttgcgtattcttccatcttagtagcagcgacttcgcagactgttatgacagcgacttgaaacttgtttcgataccg
1 M R L C V F F H L S S S D F A D C Y D S D L K L V S I P
30207 ttcacagttactaacaaattcttcaggcttccatactaa 30169
29 F T V T N K F F R L P Y * dplORF214
24273 atgatgccaaagttgtttttcagtgctcattccttttgtacgctcgtcctgataaacaatgtcaacagaaagcaagctggaagg
1 M M P K L F F S A H S F C T L V L I N N V N R K Q A G R
24189 gtttctagggtcaactgtataggtgaactgaggcattga 24151
29 V S R V N C I G E L R H * dplORF215
35822 atgttaccaaaccctgatagagtttctttacttctattatacaatcctctcgacagtttgtcaacgtcgtcattgtttcgaact
1 M L P N P D R V Ξ L L L L Y N P L D S L Ξ T S S L F R T
35738 acgattgttccaatgttgacaacggtttgctcgccttga 35700
29 T I V P L T T V C S P * dplORF216
32849 atggcctcggagctcgcggccacatctcctccagatacggcagccaggtcaagtacccctggcatagcgtccatgatttcattt
1 M A Ξ E L A A T S P P D T A A R S S T P G I A S M I S F
32765 acctggaaaccggctgaagctagattttccataccttga 32727
29 T W K P A E A R F S I P * dplORF217
23443 atgaatactatgcttacagctgggacagtaaagcgagccaaacgggagaagatagagtcattaaagagcatgaccactgcatgg
1 M N T M L T A G T V K R A K R E K I E S L K S M T T A
23527 ataggaacagatatgcctgtctcactgacgctctaa 23562
29 I G T D M P V S L T L * dplORF218
22029 atggaatgcttccggaagaggttcgatatagactacaaattgagcgcgagaaaattacattgctccgggccaaaatgggcgacc
1 M E C F R K R F D I D Y K L S A R K L H C S G P K W A T
22113 aggaaattgaaggcgaggttaaagataacttcgtag 22148
29 R K L K A R L K I T S * dplORF219
51388 atgattttatgctcgactttttcagttctcccatttcttcgaaacgcttcagggctgacgccttgcctaactacttcgctagat
1 M I L C S T F S V L P F L R N A S G L T P C L T T S L D
51304 gttccaaaattccttttcagccactggtttccatag 51269
29 V P K F L F S H W F P * dplORF220
6334 gtgaagttttcttcggtgacggttgatacaatttccttcaagagtaagctgttaaggtggcaagtgaattctttcttcgaaact
1 V K F S S V T V D T I Ξ F K S K L L R Q V N S F F E T
6250 ttcttgccagcagatgcgtacatgatgtcttcataa 6215
29 F L P A D A Y M M S S * dplORF221
43507 atgactgctcaagttctatgtactatgctctccgctcagccggagcttcaagtgctggatgggcagtcaatactgagtacatgc
1 M T A Q V L C T M L S A Q P E L Q V L D G Q S I L S T C
43591 acgcatggcttattgaaaacggttatgaactaa 43623
29 T H G L L K T V M N * dplORF 22
13212 gtgacggtatcgagaaccttatggattggctcgaaaatgataccaatttcttctcaagtccagcaagcactcgataccatggaa
1 V T V S R T L W I G S K M I P I S S Q V Q Q A L D T M E
13296 gctatgaaggtggacttgtcgagcactcattaa 13328
29 A M K V D L S S T H * - -- dplORF223 -.
14055 atgtggtggtacctgctggatatgttcgagatgtctactacttctacagtgaagtcgctgacgtttactacaagaaagatgtcg
1 M W W Y L L D M F E M S T T S T V K S L T F T T R K M S
14139 acgagcctgacgatgacagcgacattcttgtag 14171
29 T S L T M T A T F L *
dplORF224
13621 atgccagaaaattgcttgagcttcaactggcgtgagttgaatgaaacgttgaagaaggaaattagattttgcaccatgtcccat
1 M P E N C L S F N W R E L N E T L K K E I R F C T M S H
13537 tgtaagttgctcagggtcgtattcatatgctaa 13505
29 C K L L R V V F I C * dplORF225
32991 gtgagcaacgggtgcgacgtatttcatcgcctctgccatgtcgctagtttctgcgttcgtatcagctgctgctcgagcaaatac
1 V S N G C D V F H R L C H V A S F C V R I S C C S S K Y
32907 gtcagccacgtgacccgcctggtttgcctctaa 32875
dplORF226
25191 gtggctgcgtacattagtttgaacttcagtgagcgcaagttgcttagcagaaagttcatcgctaggaattggatagtggtgttc
1 V A A Y I S L N F S E R K L L S R K F I A R N W I V V F
25107 gatagtcattgtcgtaagtgtttgataacttga 25075
29 D S H C R K C L I T * dplORF227
23115 atgactcaattagatggtagcgcttatgacgtttcgagaatccataaaggccgaaggttgttgcattatagataccaaagtcgc
1 T Q L D G S A Y D V S R I H K G R R L L H Y R Y Q S R
23031 ctgctacgaataaacggtcgaattctatattga 22999
29 L L R I N G R I L Y * dplORF228
10450 atgttcgaaacattattgaagattctagatacaagtctatggacagcgagttcaaagtttacatcattgacgaggttcatatgc
1 M F E T L L K I L D T S L W T A S S K F T S L T R F I C
10534 tttcaaccggagcatttaatgcgctgttga 10563
29 F Q P E H L M R C * dplORF229
27634 atgtgcgagttaagaaaactgattttaatcaaaccactcgaagcattgtcgcaattcctgaccactacgttgctttggctgctc
1 M C E L R K L I L I K P L E A L S Q F L T T T L L W L L
27718 aaattccagctaccgcagcaactcaagtag 27747
29 K F Q L P Q Q L K * dplORF230
50723 gtgacgaaaaatccggcatacttgaactatctgtcgttaaaaaccgatatggcgaagaccgaaaaatcatcgaatatatgtggg
1 V T K N P A Y L N Y L S L K T D M A K T E K S S N I C G
50807 acgttgaaactggaacctatactcttatag 50836
29 T L K L E P I L L * dplORF231
31071 atgcgcgtgtcattgcgtttcacatcttcagttccctccgaggtcacggcttcgagttctgctgtttctgccgtatctacgaca
1 M R V S L R F T S S V P S E V T A S S S A V S A V S T T
30987 aagttagctccgccgacttttggcaactga 30958
29 K L A P P T F G N * dplORF232
29385 atgtcaattccattagctcttgctaattcaacgagctcaggaacggttttagccgcatactcttcgcgcatttgttcaacttcg
1 M S I P L A L A N S T S S G T V L A A Y S S R I C S T S
29301 tcaatttcttcaactgattcaattgtttga 29272
29 S I S S T D S I V * dplORF233
52892 atgtcttcgccttccgggtcatcatacaatagagtgacaattgcgctgtcaccgtggtcagcgagtgtgaaaaactcgttatta
1 S S P S G S S Y N R V T I A L S P W S A S V K N S L L
52808 gaccctgagctaaatgttcctgatttttga 52779
29 D P E L N V P D F * dplORF234
36253 atgcttacgagtacagcgactcaactgttcgaaaggtttataagtttcaacccgctttgggaggcgatagcttacctaacccag
1 M L T S T A T Q L F E R F I S F N P L E A I A Y L T Q
36337 gaagacctactcgacaatttagagtag 36363
29 E D L L D N L E * dplORF235
32768 atgaaatcatggacgctatgccaggggtacttgacctggctgccgtatctggaggagatgtggccgcgagctccgaggccatgg
1 M K S W T L C Q G Y L T W L P Y L E E M W P R A P R P W
32852 ctagttcacttcgagcctttggattag 32878
29 L V H F E P L D * dplORF236
37528 atgttcgtcgcttttagatttagcaatatatcgaggcttcatgtggcgtgtagtaaaccacgaaacatcaatgagatattcact
1 M F V A F R F S N I S R L H V A C S K P R N I N E I F T
37444 tccattgttgatagaagcaaacgttaa 37418
29 S I V D R S K R * dplORF237
1678 gtgagagtccaggtaaggaatcttgacatattctcagccgtagttctaaatccaaatagaactcgcttggtgtttaactgcattt
1 V R V Q V R N L D I F S A V V L N P N R T R L V^S T A F
1594 gctaaagcgattggttcattcccttga 1568
29 A K A I G S F P * dplORF238
1301 atgcctttttgcggtcgatacaagttgcgcaagttccacaactttcagcgtcactttcataacatgaacgagtcaagaaataag
1 M P F C G R Y K L R K F H N F Q R H F H N M N E S R N K
1217 gaacatctaaatcaattccccatttaa 1191
29 E H L N Q F P I * dplORF239
26521 atggtgaagtatttcctatcgaagaatgtcctttcgaccatcctaatggaatgtgctaccaaactgtatggtacgaaaactcac
1 M V K Y F L S K N V L S T I L M E C A T L Y G T K T H
26605 tcgaagaaatcgctgatgagttga 26628
29 S K K S L M S * dplORF240
41893 atgtttggaataagcgtgaaacagagtttacatggcgaagtaacaaatacgaggacaaccctacgggaactcgaggtgaatggg
1 M F G I S V K Q S L H G E V T N T R T T R E L E V N G
41977 gactatttcaaaatttctggttag 42000
29 D Y F K I S G * dplORF241
47020 gtgtctttccttaatatggagatagttttcattctatttaagcaggatatcgaaaaggttaccaattttagatttcataggctt
1 V S F L N M E I V F I L F K Q D I E K V T N F R F H R L
46936 accatctacgatataatctgctaa 46913
29 T I Y D I I C * dplORF242
41338 gtgtctgtaacccatgctcttacggtagcggagccattaaagttcatcatacccaatttgccgccgttttcgttgatagcttgg
1 V S V T H A -i T V A E P L K F I I P N L P P F S L I A W
41254 tttttacctacgagctcagcgtga 41231
29 F L P T S S A * dpl0RF243
51306 atgttccaaaattccttttcagccactggtttccatagaaccctccatcgtttcgacctaatacattcgagacgaattcagtta
1 M F Q N S F S A T G F H R T L H R F D L I H S R R I Q L
51222 gtcctgaagtgtagccgcaagtga 51199
29 V L K C S R K * dplORF244
27083 gtgaggtacaaaatgttgaccgtcgccgtcaatgaaaattttagcatcgagttctttcgaagttttcgaaataatttccttcac
1 V R Y K M L T V A V N E N F S I E F F R S F R N N F L H
26999 ctgtttgatagttggttcatctag 26976
29 L F D S W F I * dplORF245
6278 gtggcaagtgaattctttcttcgaaactttcttgccagcagatgcgtacatgatgtcttcataactgctagtagaagttttaat
1 V A S E F F L R N F L A S R C V H D V F I T A S R S F N
6194 tcgaagtcggtctttcaagaataa 6171
29 S K S V F Q E * dplORF246
2831 atggagtatcttgcaacccgtcacgttctgcgtcctcgcctaatagaccaaaaagtctttgaacggctgcctcagtattgtcca
1 M E Y L A T R H V L R P R L I D Q K V F E R L P Q Y C P
2747 aggttacaatttcatccggcttaa 2724
29 R L Q F H P A * dplORF247
29641 gtgacgcagactactggaaacaaatggcgcaattctattatgaccaatataagcaagaacagcttgaaactgatgaaaagtcga
1 V T Q T T G N K W R N S I M T N I S K N S L K L M K S R
29725 acgctggttcgacaatcttaa 29745
29 T L V R Q S * dplORF248
53560 gtgcaaagcctcgttctagcaagaagaacgatgctcagttacttgctcaacggaaaaacaggaagcctgcagttgaggttactt
1 V Q S L V L A R R T M L S Y L L N G K T G S L Q L R L L
53644 acatttcaggaaacgctctaa 53664
29 T F Q E T L * dplORF249
2012 gtggatgcgactatcattgcaactggtgtgactcagcctttacctggaacggtactactgagccggaatatatcacaggcaaag
1 V D A T I I A T G V T Q P L P G T V L L S R N I S Q A K
2096 aagctgctagtcgaatcttga 2116
29 K L V E S * dplORF250
23837 atgggcaaacatggaagattgacgaagactcagtcgactataaacctactcgagaaattcgaaactatattcgacaacttatca
1 M G K H G R T K T Q S T I N Ii E K F E T I F D N L S
23921 aaaagcaatcacgctttatga 23941
29 K S N H A L * dplORF251
39205 atggaaataattagtcttaccgtctgcgcctggcttcccgggtatcccttgagctccgtcattccccttccatttcgtccatgt
1 M E I I S L T V C A W L P G Y P L S S V I P L P F R P C
39121 ataggctgcagggtcttttga 39101
29 I G C R V F * dplORF252 " ~"
54771 gtgttgtataggtcgaaactaattttgcatattttctatatttcaaaagtgcttttgagatatcgttatβaaaatgctcgacaa
1 V L Y R S K L I L H I F Y I S K V L R Y R Y Q N A R Q
54687 tactttcgcctgttcctctag 54667
29 Y F R L F L * dpl0RF253
56255 atggttgcgtctataatagaaccgatgttgctagacaaagcatttgcaatcttcgagtctaatttattcgagagcttgtcgaat
1 M V A Ξ I I E P M L L D K A F A I F E S N L F E S L S N
56171 ataaagacacttgctttttga 56151
29 I K T L A F * dplORF254
48479 atgaacctttcgcttaggttcaatctttttcgaacattttcatatttaacaaaactttcagctaaaaatcgacaaagttcaatg
1 M N L S L R F N L F R T F S Y L T K L S A K N R Q S S M
48395 ttcgactcaatgtttaaataa 48375
29 F D S M F K * dplORF255
9572 atgctttggtcttctcgacgaatgactctactacattccctgcagggtttcgagcagtacgggtcaatgatgcaccgttttcgt
1 M L S S R R M T L L H S L Q G F E Q Y G S M M H R F R
9488 caaggtagtcaccttttctaa 9468
29 Q G S H L F * dplORF256
15289 atgaccttccagtcactaatgcggccgctgaaattggataccactatacatgggttcaccaacttcgagacaaagcagttgaaa
1 M T F Q S L M R P L K L D T T I H G F T N F E T K Q L K
15373 cacttgaagaaattttag 15390
29 H L K K F * dplORF257
28216 gtgaacgtgctggatttagcaaacaagctactgagatggcattcttccgtgagtctatgcgacttggtgaaaaagaccgtcaaa
1 V N V L D L A N K L Ii R W H S S V S L C D L V K K T V K
28300 acttgcaaatgctattga 28317
29 T C K C Y * dplORF258
44023 atggaaattggtattggttcgaccgtgacggatacatggctacgtcatggaaacggattggcgagtcatggtactacttcaatc
1 M E I G I G S T V T D T L R H G N G L A S H G T T S I
44107 gcgatggttcaatggtaa 44124
29 A M V Q * d lORF259
4298 atgactcgactacgaagcataaagacaagtggatggaaagagtattcgaagttattcgaaacagttctaatccagacgttaaga
1 M T R L R S I K T S G W K E Y S K L F E T V L I Q T L R
4382 ctcacgcatttgggatga 4399
29 L T H L G * dplORF260
24746 gtgaccctacttcctcaatcggcggtactggaggcaagcaagctcaagtcacttccatttcaggaaacttcaacttccttccag
1 V T L L P Q S A V L E A S K L K S L P F Q E T S T S F Q
24830 cggctgaatattatttag 24847
dplORF261
288 atgaattcacttccctttgccctaaaacaggacagcctgacttcgcgaatgttttcattagttacattccaaacgaaaagatgg
1 M N S L P F A L K Q D S L T S R M F S L V T F Q T K R
372 ttgaatctaaatcattga 389
dplORF262
9408 atgcctattcaactccaggcggaaagatgtggaagcatgcttgtgcagttcgacttaaatttagaaaaggtgactaccttgacg
1 M P I Q Ii Q A E R C G S M L V Q F D Ii N Ii E K V T T Ii T
9492 aaaacggtgcatcattga 9509
29 K T V H H *
dplORF263
27052 atgaaaattttagcatcgagttctttcgaagttttcgaaataatttccttcacctgtttgatagttggttcatctagacctttt
1 M K I L A S S S F E V F E I I S F T C Ii l V G S S R P F
26968 aacaagtcttctaattga 26951
29 N K S S N * d lORF264
6139 gtgaatagtacaaggcggtctaatacgctcaggatttctgctgtagggatagccgcatcatcttcaaactcaattgagtcaagc
1 V N S T R R S N T L R I S A V G I A A S S S N S I E S S
6055 tgtgaaacgtcttcataa 6038
29 C E T S S * dplORF265
4801 gtgaataaagtcaagcgtttttgtataaaaagttcatttttttttaaaaaaaataagagcgaaaagctcttatctaaaatagtc
1 V N K V K R F C I K S S F F F K K N K S E K Ii L S K I V
4717 gacgttgacgatttttaa 4700
29 D V D D F * dplORF266
50220 atgcccgttcttccaagcagttgcaagcattttatcaatagtccacgacttaccttgtccaggtcgagccattatgacaatcaa
1 M P .V P S S C K H F I N S P R L T L S R S S H Y D N Q
50136 atcctcaccaggaagtaa 50119
29 I L T R K * - -- dplORF267 -_
47367 atggtcaaggtctgttctaggttcaggaagaacaaacgggaagtgaatgttattttcttcagcgaagtcttttgcttcatacca
1 M V K V C S R F R K N K R E V N V I F F S E V F C F I P
47283 aacattaatcgtagatag 47266
29 N I N R R * dplORF268
12621 atgtcaatttcggtcttgtgcttgacaatggattcaactactgatgcgtcaacctttttcaatcgcgacagcttgtccaattca
1 M S I S V L C Ii T M D S T T D A S T F F N R D S L S N S
12537 ttgtcaattctagagtaa 12520
29 L S I L E * d lORF269
53834 gtgaatagtatcgagtccatcagtttctacgtcaatagaacctattccgtcttcaatcattttgtctacatactgctcgagttt
1 V N S I E S I S F Y V N R T Y S V F N H F V Y I L L E F
53750 tgcttcctcagtgattaa 53733
dplORF270
50792 atgatttttcggtcttcgccatatcggtttttaacgacagatagttcaagtatgccggatttttcgtcacgcttcatagcgata
1 M I F R S S P Y R F Ii T T D S S S M P D F S S R F I A I
50708 actctgctagcattttga 50691
dplORF271
19739 atgaggctgctttgctttatcttcgttaccgtattgaccgacttcctactcgcgaaccttcctacaagaattcatacctcaaag
1 M R L L C F I F V T V L T D F L L A N L P T R I H T S
19655 gctttttgtcagccttag 19638
29 A F C Q P * dplORF272
1556 gtggtcaagtctgtcaatgaatgtacctgcgattttcttgacgtgataaaagtcaacaaccatcccttgactcgaaccgtggtc
1 V V K S V N E C T C D F L D V I K V N N H P Ii T R T V V
1472 ataagttccgcctgctaa 1455
29 I S S A C * dplORF273
56256 atggatttcattaggactgagtcctcttggaattggaacggttgcatatatagatattccgtcagccgtactaggccaagttct
1 M D F I R T E S S W N W N G C I Y R Y S V S R T R P S S
56340 agttcagtttatcttgcagtcaattgcttcgagatatttgaaaaagtagtcaggaaaattcctgattatcttgcagtcaattgc
29 S S V Y Ii A V N C F E I F E K V V R K I P D Y L A V N C
56424 ttcgagatatttgaaaaagtagtcaggaaaattcctgattattttttttacaaaaacgcttga 56486
57 F E I F E K V V R K I P D Y F F Y K N A *
Table 31
Query= aid| 114822 | lan|dplORF001 Phage dpi ORF | 36698-40390 | 2 (1230 letters)
>gi|928828 (L44593) ORF1904; putative [Lactococcus lactis phage BK5-T] Length = 1904
Score = 427 bits (1086) , Expect = e-118
Identities = 226/475 (47%), Positives = 281/475 (58%), Gaps = 45/475 (9%)
Query: 395 AESGKYIGVLNTNKKPSELVPDDFTWIRLEGPKGDAGLPGAPGRDGVOGVPGKSGVGIAD 454
A+ YIG + P D+T + +G+ G GA G+DGV GK GVGI Sbjct: 820 ADYPSYIGQYTDFIQYDSAKPSDYT SLI---RGNDGKDGATGKDGV AGKDGVGIKT 873
Query: 455 TAITYAVSVSGTQEPENG SEQVPELIKGRFLWT TF RYTDGSHETGYSVAYIGQDGNS 514
T ITYA+S SGT +P GW+ QVP L+KG++LWTKT W YTD S ETGYSV YI +DGN+ Sbjct: 874 TVITYALSSSGTDKPNTGWTSQVPTLVKGQYL TKTVWTYTDSSSETGYSVTYIAKDG N 933
Query: 515 GKDGIAGKDGVGIAATEVMYASSPSATEAPAGGWSTQVPTVPGGQYLWTRTRWRYTDQTD 574
G DGIAGKDGVGI T + YA S T APA GW++QVP VP GQ+LWT+T YTD T Sbjct: 934 GOTGIAGKDGVGIKKTTITYAVGTSGTTAPASGVroSQVPWPAGQFLWTKTVWTYTDNTS 993
Query: 575 EIGYSVSRMGEQGPKGDAGR---DGIAGKNGIGLKSTSVSYGISPTDSAIP-GVWASQVP 630
E GYSV+ MG +G KGD G +GIAGK+G G+K+T+++Y SP + P G W++ VP Sbjct: 994 ETGYSVAMMGVKGDKGDPGNNGTNGIAGKDGKGIKATAITYQASPNGTTAPTGTWSASVP 1053
Query: 631 SLIKGQYLWTRTIWTYTDSTTETGYQ TYIPKDGNDGKNGIAGKDGVGIKSTTITYAGST 690
+ KG +LWTRTIWTYTD+TTETGY Y+ +GN+G +G GKDG GIK+TTITYAGST Sbjct: 1054 PVAKGSFLWTRTIWTYTDNTTETGYAVAYMGTNGNNGHDGFPGKDGTGIKTTTITYAGST 1113
Query: 691 SGTVAPTSNWTSAIPWQPGFFLWTKTVTOr_TDDTSETGYSVSKIGETXXX_-X__XXXXXX 750
SGT P + TS +P V G +LWTKTVW YTD+TSETGYSV+ +G Sbjct: 1114 SGTTPP.MG TSTVPTVAEGNYL TKTV TYTDNTSETGYSVAMMG VKGDKGDP 1167
Query: 751 XXXXXXXXXXADGRS-QYTHIiAFSNSPNGEGFSHTDSGRAYVGQYQDFNPVHSKDPAAYT 809
DG+ + T + + SPNG A G + P +K +T
Sbjct: 1168 GN GTNGIAGKDGKGIKATAITYQASPNGT TAPTGT SASVPPVAKGSFLWT 1219
Query: 810 TKW--- KGNDGAQGIPGKPGADGKTNYFHIAYASSADGS 846
T GN+G G PGK G KT I YA S G+
Sbjct: 1220 RTI TYTD TTETGYAVAYMGTNG NGHDGFPGKDGTGIKTT--TITYAGSTSGT 1272
Score = 396 bits (1007) , Expect = e-109
Identities = 208/449 (46%), Positives = 260/449 (57%), Gaps = 42/449 (9%)
Query: 421 IRLEGPKGDAGLPGAPGRDGVDGVPGKSGVGIADTAITYAVSVSGTQEPENGWSEQVPEL 480
+ + G KGD G PG +G +G+ GK G GI TAITY S +GT P WS VP + Sbjct: 1155 VAMMGVKGDKG---DPGNNGTNGIAGKDGKGIKATAITYQASPNGTTAPTGTWSASVPPV 1211
Query: 481 IKGRFLWTKTFWRYTDGSHETGYSVAYIGQDGNSGKDGIAGKDGVGIAATEVMYASSPSA 540
KG FL T+T YTD + ETGY+VAY+G +GN+G DG GKDG GI T + YA S S Sbjct: 1212 AKGSFLWTRTIWTYTDNTTETGYAVAYMGTNGNNGHDGFPGKDGTGIKTTTITYAGSTSG 1271
Query: 541 TEAPAGGWSTQVPTVPGGQYLWTRTRWRYTDQTDEIGYSVSRMGEQGPKGDAGR DGI 597
T P GW++ VPTV G YL T+T YTD T E GYSV+ MG +G KGD G +GI Sbjct: 1272 TTPP NGπ,STVPTVAEG^r TKTV^^_TDNTSETGYS AM GVKGDKGD G NGTNGI 1331
Query: 598 AGKNGIGLKSTSVSYGISPTDSAIP-GVWASQVPSLIKGQYLWTRTIWTYTDSTTETGYQ 656
AGK+G G+K+T+++Y SP + P G W++ VP + KG +LWTRTI TYTD+TTETGY Sbjct: 1332 AGKDGKGIKATAITYQASPNGTTAPTGT SASVPPVAKGSFL TRTI TYTDNTTETGYA 1391
Query: 657 KTYIPKDG DGKNGIAGKDGVGIKSTTITYAGSTSGTVAPTSN TSAIPNVQPGFFL TK 716
Y+ +GN+G +G GKDG GIK+TTITYAGSTSGT P + WTS +P V G +LWTK Sbjct: 1392 VAYMGTNG NGHDGFPGKDGTGIKTTTITYAGSTSGTTPP NGWTSTVPTVAEGNYLWTK 1451 _
Query: 717 TVWNYTDDTSETGYSVSKIGETXXXXXXXXXXXXXXXXXXXXXXADGRS-QYTHLAFSNS 775
TVW YTD+TSETGYSV+ +G DG+ + T + + S
Sbjct: 1452 TVWTΎTDNTSETGYSVAMMG VKGDKGDPGNNGTNGIAGKDGKGIKATAITYQAS 1505
Query: 776 PNGEGFSHTDSGRAYVGQYQDF PVHSKDPAAYTWTKW KGND 817
PNG A G + P +K +T T W GN+
Sbjct: 1506 PNGT TAPTGTWSASVPPVAKGSFLWTRTIWTYTDNTTETGYAVAYMGTNGNN 1557
Query: 818 GAQGIPGKPGADGKTNYFHIAYASSADGS 846
G G PGK G KT I YA S G+ Sbjct: 1558 GHDGFPGKDGTGIKTT--TITYAGSTSGT 1584
Score = 384 bits (977) , Expect = e-105
Identities = 179/322 (55%), Positives = 222/322 (68%), Gaps = 7/322 (2%)
Query: 421 IRLEGPKGDAGLPGAPGRDGVDGVPGKSGVGIADTAITYAVSVSGTQEPENGWSEQVPEL 480
+ + G KGD G PG +G +G+ GK G GI TAITY S +GT P WS VP + Sbjct: 1311 VAMMGVKGDKG---DPGNNGTNGIAGKDGKGIKATAITYQASPNGTTAPTGTWSASVPPV 1367
Query: 481 IKGRFLWTKTFWRYTDGSHETGYSVAYIGQDGNSGKDGIAGKDGVGIAATEVMYASSPSA 540
KG FLWT+T W YTD + ETGY+VAY+G +GN+G DG GKDG GI T + YA S S Sbjct: 1368 AKGSFLWTRTIWTYTDNTTETGYAVAYMGTNGNNGHDGFPGKDGTGIKTTTITYAGSTSG 1427
Query: 541 TEAPAGGWSTQVPTVPGGQYLWTRTRWRYTDQTDEIGYSVSRMGEQGPKGDAGR DGI 597
T P GW++ VPTV G YLWT+T W YTD T E GYSV+ MG +G KGD G +GI Sbjct: 1428 TTPP1INGWTSTVPTVAEGNYLWTKTVWTYTDNTSETGYSVAMMGVKGDKGDPGNNGTNGI 1487
Query: 598 AGKNGIGLKSTSVSYGISPTDSAIP-GVWASQVPSLIKGQYLWTRTIWTYTDSTTETGYQ 656
AGK+G G+K+T+++Y SP + P G W++ VP + KG +LWTRTIWTYTD+TTETGY Sbjct: 1488 AGKDGKGIKATAITYQASPNGTTAPTGTWSASVPPVAKGSFLWTRTIWTYTDNTTETGYA 1547
Query: 657 KTYIPKDGNDGKNGIAGKDGVGIKSTTITYAGSTSGTVAPTSNWTSAIPNVQPGFFLWTK 716
Y+ +GN+G +G GKDG GIK+TTITYAGSTSGT P + WTS +P V G +LWTK Sbjct: 1548 VAYMGTNGNNGHDGFPGKDGTGIKTTTITYAGSTSGTTPPNNGWTSTVPTVAEGNYLWTK 1607
Query: 717 TVWNYTDDTSETGYSVSKIGET 738
TVW YTD++ ETGYSV K+G T Sbjct: 1608 TVWAYTDNSFETGYSVGKMGNT 1629
Score = 201 bits (507) , Expect = 2e-50
Identities = 121/297 (40%), Positives = 156/297 (51%), Gaps = 19/297 (6%)
Query: 421 IRLEGPKGDAGLPGAPGRDGVDGVPGKSGVGIADTAITYAVSVSGTQEPENGWSEQVPEL 480
+ + G KGD G PG +G +G+ GK G GI TAITY S +GT P WS VP + Sbjct: 1467 VAMMGVKGDKG DPGNNGTNGIAGKDGKGIKATAITYQASPNGTTAPTGTWSASVPPV 1523
Query: 481 IKGRFLWTKTFWRYTDGSHETGYSVAYIGQDGNSGKDGIAGKDGVGIAATEVMYASSPSA 540
KG FLWT+T W YTD + ETGY+VAY+G +GN+G DG GKDG GI T + YA S S Sbjct: 1524 AKGSFLWTRTIWTYTDNTTETGYAVAYMGTNGNNGHDGFPGKDGTGIKTTTITYAGSTSG 1583
Query: 541 TEAPAGGWSTQVPTVPGGQYLWTRTRWRYTDQTDEIGYSVSRMGEQGPKGDAGRDGIAGK 600
T P GW++ VPTV G YLWT+T W YTD + E GYSV +MG GP AG +G GK Sbjct: 1584 TTPPNNGWTSTVPTVAEGNYLWTKTVWAYTDNSFETGYSVGKMGNTGP AGSNGNPGK 1640
Query: 601 NGIGLKSTSVSYGISPTDSAIPGVWASQVPSLIKG-QYLWTRTIWTYTDSTTE--TGYQK 657
+ T+ G++ S + + ++ G +Y W W + G Sbjct: 1641 WSDTEPTTKFKGLTWKYSGWDMPLGNGTKILAGTEYYWNGNNWALYEINAH INGDNL 1700
Query: 658 TYIPKDGNDGK-NGIAGKDGVGIKSTTITYAGS TSGTVAPTSNWTSAIPNVQ 708
+ DGK I G +GV + T T GS +S + T N T AI Q Sbjct: 1701 SVTNGTFKDGKIESIWGSNGV---NGTTTIEGSHLQIHSSDSTTNTEN-TLAIDNRQ 1753
Query= sid | 114823 | Ian |dplORF002 Phage dpi ORF| 32386-35835 | 1 (1149 letters)
>dbj |BAA31888| (AB009866) orf 15 [bacteriophage phi PVL] Length = 694
Score = 280 bits (709), Expect = 3e-74
Identities = 157/465 (33%), Positives = 257/465 (54%), Gaps = 28/465 (6%)
Query: 40 QIGSALTGLGKGLTTAVTLPLMGFAAASIKVGNEFQAQMSRVQAIAGATAEELGRMKTQA 99
+IG+++ +G+ +T VT P++ A + K G EF M +V+A +GAT EE +K +A Sbjct: 151 EIGNSMKNVGRNMTMYVTAPWAGFAVAAKKGIEFDDSMRKVKATSGATGEEFEALKKKA 210
Query: 100 IDLGAKTAFSAKEAAQGMENI_.SAGFQVNEI^1DAMPGVLDIi -_X X__X_aαXXX ASSL 159
++GA T FSA ++A+ + +A AG+ ++M+ + GV+DL + L
Sbjct: 211 REMGATTKFSASDSAEALNYMALAGWDSKQMMEGLSGVMDLAAASGEELGAVSDIVTDGL 270
Query: 160 RAFGLEANQAGHVADVFARAAADTNAETSDMAEAMKYVAPVAHSMGLSLEETAASIGIMA 219
AFGL+A +GH+ADV A+ ++ N + + EA KYVAPVA ++G ++E+T+ +IG+M+ Sbjct: 271 TAFGLKAKDSGHLADVLAQTSSKANTDVRGLGEAFKYVAPVAGALGYTIEDTSIAIGLMS 330
Query: 220 DAGIKGSQAGTTLRGALSRIAKPTKAMVKSMQELGVSFYDANGNMIPLREQIAQLKTATA 279
+AGIKG +AGT LR + ++ PT+AM M+ LG+S D+NG MIP+R+ + QL+ Sbjct: 331 NAGIKGEKAGTALRTMFTNLSSPTRAMGNEMERLGISITDSNGKMIPMRKLLDQLREKFK 390
Query: 280 GLTQEER-IRHLVTLYGQNSLSGMIiALIiDAGPEKLDKMTNALVNSDGAAKEMAETMQDNIA 339
L+++++ T++G+ ++SG LA+++A E K+T ++ +S GA+K MA+TM+ L Sbjct: 391 HLSKDQQASSAATIFGKEAMSGALAIINASDEDYQKLTKSIDSSTGASKRMADTMESGLG 450
Query: 340 SKIEQMGGAFESVAIIVQQILEPALAKIVGAITKVLEAFVNMSPIGQKMWIFAGMVAAL 399
K+ + E +A+ + +EPAL IV A +KV+ + Q W F VA L Sbjct: 451 GKLRTLRSQLEEI_ iTIYDRIEPALKIIVSAFSKVVTWVTKLPTSIQIιAVVGFGLFVAVL 510
Query: 400 GPLLLIAGM VMTTIVKLRIAIQFLGPAFMGTMGTIAGVIAIF 441
GPL+ + G+ MT + L I + F IA ++ +F Sbjct: 511 GPLVFMFGLFISVMGNAMTVLGPLLINVNKASGLFAFLRTKIASLVKLFPILGVSISSLT 570
Query: 442 --YALVAV FMIAYTKSERFRNFINSLAPAIKAGFGGA 476
ALV + F AY +SE FRN +N + F A Sbjct: 571 LPITLIVGALVGIGIAFYQAYKRSETFRNIVNQAISGVANAFKAA 615
Query= sid| 114824 | lan|dplORF003 Phage dpi ORF| 53538-55877 | 3 (779 letters)
>sp|P4374l|DP01_HAEIN DNA POLYMERASE I (POL I) >gi | 1074025 |pir| | E64098 DNA polymerase I (polA) homolog - Hae ophilus influenzae (strain Rd KW20) >gi 11573871 (U32767) DNA polymerase I (polA) [Haemophilus influenzae Rd] Length = 930
Score = 191 bits (481) , Expect = le-47
Identities = 148/553 (26%) , Positives = 262/553 (46%) , Gaps = 60/553 (10%)
Query: 63 RLELITEEAKLEQYVDKMIEDGIGSIDVETDGLDTIHDELAGVCLYSPSQKGIYAPVNHV 122
+ E + +A L ++++K+ + ++D ETD LD + L G+ + + Y P+ Sbjct: 333 KYETLLTQADLTRWIEKLNAAKLIAVDTETDSLDYMSANLVGISFALENGEAAYLPLQLD 392
Query: 123 SNMTKMRIKNQISPEFMKKMLQRIVDSGIPVIYHNSKFDMKSIYWRLGVKMNEPAWDTYL 182
++ + +K +L+ + I I N KFD +SI+ R G+++ +DT L Sbjct: 393 YLDAPKTLEKSTALAAIKPILE NPNIHKIGQNIKFD-ESIFARHGIELQGVEFDTML 448
Query: 183 AAMLLNENESHSLKSLHSKYVRNEENAEVAKFNDLFKGIPFSLIPPDVAYMYAAYDPLQT 242
+ LN H++ L +Y+ +E A + + F+ IP + A YAA D T Sbjct: 449 LSYTLNSTGRHNMDDLAKRYLGHETIAFESLAGKGKSQLTFNQIPLEQATEYAAEDADVT 508
Query: 243 FELYEFQEQYLTPGTEQCEEYNLEKVSWVLHNIEMPLIKVLFDMEVYGVDLDQDKLAEIR 302
+L + L E Y +E+PL+ VL ME GV +D D L
Sbjct: 509 MKLQQALWLKLQEEPTLVELYK TMELPLLHVLSRMERTGVLIDSDALFMQS 559
Query: 303 EQFTANMNEAEQEFQQLVSEWQPEIEELRQTNFQSYQKLEMDARGRVTVSISSPTQLAIL 362
+ + + E++ L + +++S QL +
Sbjct: 560 NEIASRLTALEKQAYALAGQ - - -PFNLASTKQLQEI 592
Query: 363 FYDIMGLKSPERDKPRG---TGESIVEH--FDNDISXXXXXXXXXXXXVSTYTT-LDQHL 416
+D + L ++ P+G T E ++E + +++ STYT L Q +
Sbjct: 593 LFDKLELPVLQKT-PKGAPSTNEEVLEELSYSHELPKILVKHRGLSKLKSTYTDKLPQMV 651
Query: 417 AKPDNRIHTTFKQYGAKTGRMSSENPNLQNIPSRGE-GAWRQIFAASEGHYIIGSDYSQ 475
R+HT++ Q TGR+SS +PNLQNIP R E G +RQ F A EG+ 1+ +DYSQ Sbjct: 652 NSQTGRVHTSYHQAVTATGRLSSSDPNLQNIPIRNEEGRHIRQAFIAREGYSIVAADYSQ 711
Query: 476 QEPRSLAELSGDESMRHAYEQNLDLYSVIGSKLYGVPYEECLEFYPDGTTNKEGKLRRNS 535 --
E R +A LSGD+ + +A+ Q D++ ++++GV +E T+++ R + Sbjct: 712 IELRIMAHLSGDQGLINAFSQGKDIHRSTAAEIFGVSLDE VTSEQ RRN 759
Query: 536 VKSVLLGLMYGRGANSIAEQMNVSVKEANKVIEDFFTEFPKVADYIIFVQQQAQDLGYVQ 595 K++ GL+YG A ++ Q+ +S +A K ++ +F +P V ++ ++++A+ GYV+
Sbjct: 760 AKAINFGLIYGMSAFGLSRQLGISRADAQKYMDLYFQRYPSVQQFMTDIREKAKAQGYVE 819
Query: 596 TATGRRRRLPDMS 608
T GRR LPD++ Sbjct: 820 TLFGRRLYLPDIN 832
Score = 46.9 bits (109), Expect = Se-04
Identities = 34/123 (27%) , Positives = 66/123 (53%) , Gaps = 16/123 (13%)
Query: 663 EIKDQAKAEGI --LIKDNGGKIADAQRQCLNSVIQGTAADMTKYAMIKV 709
+I+++AKA+G + N + A+R +N+ +QGTAAD+ K AMIK+
Sbjct: 807 DIREKAKAQGYVETLFGRRLYLPDINSSNAMRRKGAERVAINAPMQGTAADIIKRAMIKL 866
Query: 710 HNDAELKELGFHLMIPVHDELLGEVPIKNAKRGAERLTEVMIEAAKDIISLPMKCDPSIV 769
++ + +++ VHDEL+ EV + E++ + M EAA +++ +P+ + + Sbjct: 867 -DEVIRHDPDIEMIMQVHDELVFEVRSEKVAFFREQIKQHM-EAAAELV-VPLIVEVGVG 923
Query: 770 ERW 772
+ W Sbjct: 924 QNW 926
Query= sid| 114825 | lan|dplORF004 Phage dpi ORF| 40401-42440 | 3 (679 letters)
>emb|CAB0798l| (Z93946) hypothetical protein [bacteriophage Dp-1] Length = 532
Score = 1011 bits (2585), Expect = 0.0
Identities = 497/499 (99%) , Positives = 498/499 (99%)
Query: 1 MTKFINSYGPLHLNLYVEQVSQDVTNNSSRVSWRATVDRDGAYRTWTYGNISNLSVWLNG 60
MTKFINSYGPLHLNLYVEQVSQDVTNNSSRVSWRATVDRDGAYRTWTYGNISNLSVWLNG Sbjct: 1 MTKFINSYGPLHIiNLYVEQVSQDVTNNSSRVSWRATVDRDGAYRTWTYGNISNLSVWLNG 60
Query: 61 SSVHSSHPDYDTSGEEVTLASGEVTVPHNSDGTKTMSVWASFDPNNGVHGNITISTNYTL 120
SSVHSSHPDYDTSGEEVTLASGEVTVPHNSDGTKTMSVWASFDPNNGVHGNITISTNYTL Sbjct: 61 SSVHSSHPDYDTSGEEVTLASGEVTVPHNSDGTKTMSVWASFDPNNGVHGNITISTNYTL 120
Query: 121 DSIPRSTQISSFEGNRNLGSLHTVIFNRKVNSFTHQVWYRVFGSDWIDLGKNHTTSVSFT 180
DSIPRSTQISSFEGNRNLGSLHTVIFNRKVNSFTHQVWYRVFGSDWIDLGKNHTTSVSFT Sbjct: 121 DSIPRSTQISSFEGNRNLGSLHTVIFNRKVNSFTHQVWYRVFGSDWIDLGKNHTTSVSFT 180
Query: 181 PSLDLARYLPKSSSGTMDICIRTYNGTTQIGSDVYSNGWRFNIPDSVRPTFSGISLVDTT 240
PSLDLARYLPKSSSGTMDICIRTYNGTTQIGSDVYSNGWRFNIPDSVRPTFSGISLVDTT Sbjct: 181 PSIiDLARYLPKSSSGTMDICIRTYNGTTQIGSDVYSNGWRFNIPDSVRPTFSGISLVDTT 240
Query: 241 SAVRQILTGNNFLQIMSNIQVNFNNASGAYGSTIQAFHAELVGKNQAINENGGKLGMMNF 300
SAVRQILTGNNFLQIMSNIQVNFNNASGAYGSTIQAFHAELVGKNQAINENGGKLGMMNF Sbjct: 241 SAVRQILTGNNFLQIMSNIQVNFNNASGAYGSTIQAFHAELVGKNQAINENGGKLGMMNF 300
Query: 301 NGSATVRAWVTDTRGKQSNVQDVSINVIEYYGPSINFSVQRTRQNPAIIQALRNAKVAPI 360
NGSATVRAWVTDTRGKQSNVQDVSINVIEYYGPSINFSVQRTRQNPAIIQALRNAKVAPI Sbjct: 301 NGSATVRAWVTDTRGKQSNVQDVSINVIEYYGPSINFSVQRTRQNPAIIQALRNAKVAPI 360
Query: 361 TVGGQQKNIMQITFSVAPLNTTNFTEDRGSASGTFTTISLMTNSSANLAGNYGPDKSYIV 420
TVGGQQKNIMQITFSVAPLNTTNFTEDRGSASGTFTTISL+TNSSANIiAGNYGPDKSYIV Sbjct: 361 TVGGQQKNIMQITFSVAPLNTTNFTEDRGSASGTFTTISLLTNSSANLAGNYGPDKSYIV 420
Query: 421 KAKIQDRFTSTEFSATVATESVVLNYDKDGRLGVGKVVEQGKAGSIDAAGDIYAGGRQVQ 480
KAKIQDRFTSTEFSATV TESWLNYDKDGRLGVGKWEQGKAGSIDAAGDIYAGGRQVQ Sbjct: 421 KAKIQDRFTSTEFSATVPTESWLNYDKDGRLGVGKWEQGKAGSIDAAGDIYAGGRQVQ 480
Query: 481 QFQLTDNNGALNRGQYNDV 499
QFQLTDNNGALNRGQYNDV Sbjct: 481 QFQLTDNNGALNRGQYNDV 499
Query= sid | 114827 | Ian |dplORF006 Phage dpi ORF| 45296-46987 | 2 (563 letters)
>gb|AAD18987| (AE001666) SWI/SNF family helicase_2 [Chlamydia pneumoniae] Length = 1166
Score = 171 bits (429) , Expect = le-41
Identities = 150/522 (28%), Positives = 254/522 (47%), Gaps = 55/522 (10%)
Query: 46 SSNNFE-LPYKYFNNVIDALDEWELHIFGELDIDVQDYIDSRNRIASSSNEQFSFKTTPF 104
S + FE LP + ++ + L E + I GE++ D QD + T
Sbjct: 659 SLDQFEALPVNF--SMSERLIEIQKQIRGEIEFDFQD VPQQIQATLRSYQTEG 709
Query: 105 AHQVECFEYAQEHPCFLLGDEQGLGKTKQAIDIAVSRKASFKH--CLIVCCISGLKWNWA 162
H +E + H +L D+ GLGKT QAI IAV++ K C ++ C + L +NW Sbjct: 710 VHWLE--RLRKMHLNGILADDMGLGKTLQAI-IAVTQSKLEKGSGCSLIVCPTSLVYNWK 766
Query: 163 KEVGIHSNESAHILGSRVTKDGKLVIDGV-SKRAEDLLGGHDEFFLITNIETLRDAVFIK 221
+E + E LVIDGV S+R + L D IT+ L+ V Sbjct: 767 EEFRKFNPEFR TLVIDGVPSQRRKQLTALADRDVAITSYNLLQKDV 812
Query: 222 YLNELTKSGEIGMVIIDEIHKCKNPSSKQGASIQKLQSYYKMGLTGTPLMNNPIDVFNVM 281
EL KS V++DE H KN +++ S++ +QS +++ LTGTP+ N+ +++++ Sbjct: 813 ELYKSFRFDYWLDEAHHIKNRTTRNAKSVKMIQSDHRLILTGTPIENSLEELWSLF 869
Query: 282 KWLGAEHHTLTQFKERYCIVDQFNQITGYR NLAELRELVNDYMLRRTKEEVL-DL 335
+L L +R+ V ++ + Y N+ L++ V+ ++LRR KE+VL DL Sbjct: 870 DFLMPG LLSSYDRF--VGKYIRTGNYMGNKADNMVALKKKVSPFILRRMKEDVLKDL 924
Query: 336 PEKIRVTEYVDMNSKQSKIY KEVLTKLVQEIDKVKLMPNPLAETIRLRQATGN 388
P + + + Q ++Y K+ L++LV++ ++ + LA RL+Q + Sbjct: 925 PPVSEILYHCHLTESQKELYQSYAASAKQELSRLVKQEGFERIHIHVLATLTRLKQICCH 984
Query: 389 PSILTTQDVK---SCKFERCIEIVEECIQQGKSCVIFSNWEKVIEPLAKIL-SKTVKCNL 444
P+I + S K++ ++++ + G V+FS + K++ + K L S+ + Sbjct: 985 PAIFAKDAPEPGDSAKYDMLMDLLSSLVDSGHKTWFSQYTKMLGIIKKDLESRGIPFVY 1044
Query: 445 VTGETADKFNEIEEFMNHRKASVILGTIGALGTGFTLTKADTVIFLDSPWTRAEKDQAED 504
+ G T ++ + + +F V L ++ A GTG L ADTVI D W A ++QA D Sbjct: 1045 LDGSTKNRLDLVNQFNEDPSLLVFLISLKAGGTGLNLVGADTVIHYDMWWNPAVENQATD 1104
Query: 505 RCHRIGAKSSVTIYTLVAKGTVDERIEDLIERKGELADYIVD 546
R HRIG SV+ Y LV T++E+I L RK L +++ Sbjct: 1105 RVHRIGQSRSVSSYKLVTLNTIEEKILTLQNRKKSLVKKVIN 1146
Query= sid | 114828 | lan|dplORF007 Phage dpi ORF| 22230-2362l| 3 (463 letters)
>gi 12444105 (U88974) ORF26 [Streptococcus thermophilus temperate bacteriophage O1205] Length = 411
Score = 88.9 bits (217), Expect = 7e-17
Identities = 80/315 (25%) , Positives = 133/315 (41%) , Gaps = 48/315 (15%)
Query: 139 QGVTLAGIFCDEVALMPESFVNQATGRCSVTGSKMWFSCNPANPNHYFKKNWIDKQVEKR 198
+G T G + +E +L E + RCS G+++ + NP NPNH+ +++I K + + Sbjct: 121 RGFTAFGAYVNEASLANELVFKEIISRCSGDGARWWDSNPDNPNHWLNRDYIGKN-DGK 179
Query: 199 ILYLHFTMDDNPSLT DSIKRRYEKMYAGVFRKRFILGLWVTADGLVYSMFNEEQHV 254
1+ F +DDN L+ DSIK K G F R ILGLW A+G +Y+ ++ + HV Sbjct: 180 IIDFSFKLDDNTFLSKRYIDSIKAATPK---GKFYDRDILGLWTVAEGAIYADYDSKIHV 236
Query: 255 KKLNIEFDRLFVAGDFGIYNATTFGLYGFSKRHKRYHLIESYYHSGREAEEQLTEADVNS 314
E R F D+G + + + G ++L++ +E + + +A Sbjct: 237 VDELPEMKRYFGGIDWGYTHYGSIVIVG-EGVDNNFYLVDGVAAQFKEIDWWVEQA 291
Query: 315 NIQFSSVLQKTTKEYANDLVDMIRGKQIEYIILDPSASAMIVELQKHPYIAR KNIPI 371
+K T Y N + + ++AR + I
Sbjct: 292 RKLTGIYGN IPFYADSARPEHVARFENEGFDI 323
Query: 372 IPARNDVTLGISFHAELLAENRFTLDPSNT-HDIDEYYAYSWDSKASQTGEDRVIKEHDH 430
+ A V GI A+L E + + DE Y Y W ++ +D +KE D
Sbjct: 324 MNANKSVIAGIELIAKLFKEKKLYVKRGFVPRFFDEIYQYRWKENST---KDEPLKEFDD 380
Query: 431 CMDRNRYACLTDALI 445
+D RYA +D +1 Sbjct: 381 VLDSVRYAIYSDYVI 395
Query= sid| 114829 | Ian|dplORF008 Phage dpi ORF| 49624-50961 | 1 (445 letters)
>gb|AAD1990l| (AF100420) DnaB replication fork helicase [Thermus aquaticus]
Length = 444
Score = 67.5 bits (162) , Expect = 2e-10
Identities = 69/248 (27%) , Positives = 111/248 (43%) , Gaps = 14/248 (5%)
Query: 147 GERLGISTGFEXXXXXXXXXXXXXXXIVIMARPGQGKS-WTIDKMLATAWKNGHDVLLYS 205
GE G+ TGF+ I I ARP GK+ + + A K G V +YS
Sbjct: 178 GEVAGVRTGFKELDQLIGTLGPGSLNI-IAARPAMGKTAFALTIAQNAALKEGVGVGIYS 236
Query: 206 GEMSEMQVGARIDTILSNVSINSITKGIWNDHQFEKYEDHIQAMTEAENSLVWTPFMIG 265
EM Q+ R+ + + +N + G D F + D ++EA + TP + Sbjct: 237 LEMPAAQLTLRMMCSEARIDMNRVRLGQLTDRDFSRLVDVASRLSEAP-IYIDDTPDLTL 295
Query: 266 GKNLTPAILDSMISKYRPSWGIDQLSLMS--ESYPSREQKRIQYANITMDLYKISAKYG 323
+ A ++S+ + ++ ID L LMS S S E ++ + A 1+ L ++ + G Sbjct: 296 ME--VRARARRLVSQNQVGLIIIDYLQLMSGPGSGKSGENRQQEIAAISRGLKAIiARELG 353
Query: 324 IPIVLNVQAGRSAKTEGAESMELEHIAESDGVGQNASRVIAMKRD EKSGILEL 376
IPI+ Q R+ + + L + ES + Q+A V+ + RD EK+GI E+ Sbjct: 354 IPIIALSQLΞRAVEARPNKRPMLSDLRESGSIEQDADLVMFIYRDEYYNPHSEKAGIAEI 413
Query: 377 SWKNRYG 384
V K R G Sbjct: 414 IVGKQRNG 421
Query= sid | 114831 | Ian |dplORF010 Phage dpi ORF| 8699-9859 | 2 (386 letters)
>gi 12760912 (AF037258) RecA protein [Chlorobium tepidum] Length = 346
Score = 133 bits (331) , Expect = 2e-30
Identities = 99/340 (29%) , Positives = 164/340 (48%) , Gaps = 66/340 (19%)
Query: 44 GGLPRKRWEFFGPESSGKTTSALDIVKNAQMVFXXXXXXXXXXXXXXXXNARASKASKT 103
GGLPR RV E +GPESSGKTT AL + AQ Sbjct: 67 GGLPRGRVTEIYGPESSGKTTIiALHAIAEAQ KNG 100
Query: 104 AVKELEMQLDSLQEPLKIVYLDLENTLDTEWAKKIGVDVDNIWIVRPEMNSAEEILQYVL 163
+ L +D E+ D +A+K+GVD++ + + +PE S E+ L V
Sbjct: 101 GIAAL VDAEHAFDPTYARKLGVDINALLVSQPE--SGEQALSIVE 143
Query: 164 DIFETGEVGLWLDSLPYMVSQNLIDEELTKKAYAGISAPLTEFSRKVTPLLTRYNAIFL 223
+ +G V ++V+DS+ +V Q ++ E+ + +++ RK+T +++ +++ L Sbjct: 144 TLVRSGAVDIIVIDSVAALVPQAELEGEMGDSWGLQARLMSQALRKLTGAISKSSSVCL 203
Query: 224 GINQIREDMNSQYNA-YSTPGGKMWKHACAVRLKFRKGDYLDENGASLTRTARNPAGNW 282
INQ+R+ + Y + +T GGK K +VRL RK + ++G L GN Sbjct: 204 FINQLRDKIGVMYGSPETTTGGKALKFYSSVRLDIRKIAQI-KDGEELV GNRT 255
Query: 283 ESFVEKTKAFKPDRKLVSYTLSYHDGIQIENDLVDVAVEFGVIQKAGAWFSIVDLETGEI 342
+ V K K P K + + Y +GI + +L+D+AVEFG+I+K+GAWFS + G Sbjct: 256 KVKWKNKV-APPFKTAEFDILYGEGISVLGELIDLAVEFGIIKKSGAWFSYGTEKLG-- 312
Query: 343 MTDEDEEPLKFQGKANLVRRFKEDDYLFDMVMTAVHEIIT 382
QG+ N+ + KED+ L + + V +++T Sbjct: 313 QGRENVKKLLKEDETLRNTIRQQVRDMLT 341
Query= sid | 114832 | Ian |dplORF011 Phage dpi ORF| 28017-29096 | 3 (359 letters)
>gi 12444110 (U88974) ORF31 [Streptococcus thermophilus temperate bacteriophage O1205] Length = 348
Score = 187 bits (469) , Expect = le-46
Identities = 118/358 (32%) , Positives = 187/358 (51%) , Gaps = 21/358 (5%)
Query: 3 IYDYINAGEIASYIQALPSNALQYLGPTLFPNAQQTGTDISWLKGANNLPVTIQPSNYDA 62
IYD + A IA Y AL N LG ++FP +Q GT +S++KGA+ V ++ + +D Sbjct: 4 IYDKVTASNIAGYFNALQENVSSTLGESIFPARKQLGTKLSYIKGASGQSVALKAAAFDT 63
Query: 63 KASLRERAGFSKQATEMAFFRESMRLGEKDRQNLQMLLNQSSA-LAQPLITQLYNDTKNL 121
++R+R +M FF+E+M + E DRQ L ++ + +A L ++ ++ND L Sbjct: 64 NVTIRDRVSAEMHDEQMPFFKEAMLVKENDRQQLNLVKDSGNAVLVNTIVAGIFNDNLTL 123
Query: 122 VDGVEAQAEYMRMQLLQYGKFTVKSTNSEAQYTYDYNMDAKQQYAVTKKWTNPAESDPIA 181
V+G A+ E MRMQ+L GK S Y D K+Q V+K W P + P+A Sbjct: 124 VNGARARLEAMRMQVLATGKIAFTSDGVNKDIDYGVKPDHKKQ--VSKSWAEPG-ATPLA 180
Query: 182 DILAAMDDIENRTGVRPTRMVLNRNTYNQMTKΞDSIKKAL-AIGVQGSWENFLLLASDAE 240
D+ A+ + G+ P R V+N T+ + K+ S K + + GS + ++ E Sbjct: 181 DLEDAI-ETARELGLNPERAVMNAKTFGLIRKAASTVKVIKPLAGDGS AVTKAELE 235
Query: 241 KFIAEKTGLQIAVYSKKIAQFADADKLPDVGNIRQFNLIDDGKWLLPPDAVGHTWYGTT 300
+IA+ G+ I + + D G + +F DG + L+P +G+T +GTT
Sbjct: 236 NYIADNFGVSIVLENGTYRN- - DKGEVSKF--YPDGHLTLIPNGPLGNTVFGTT 285
Query: 301 PEAFDLASGGT-DAQVQVLSGGPTVTTYLEKHPVNIATWSAVMIPSFEGIDYVGVLT 357
PE DL + T +A+V+++ G VTT PVN+ T VS V +PSFE +D V +LT Sbjct: 286 PEESDLFADNTVNAEVEIVDNGIAVTTTKTTDPVNVQTKVSMVALPSFERLDDVYMLT 343
Query= sid| 114834 | lan|dplORF013 Phage dpi ORF| 10215-11240 | 3 (341 letters)
>sp|P09122|DP3X_BACSU DNA POLYMERASE III SUBUNITS GAMMA AND TAU Length = 563
Score = 182 bits (458) , Expect = 2e-45
Identities = 118/353 (33%), Positives = 176/353 (49%), Gaps = 31/353 (8%)
Query: 7 YRPQTFEEVVAQEYVKEILIiNQLQNGAIKHGYLFCXXXXXXXXXXXRIFAKDVN- 60
+RPQ FE+W QE++ + L N L H YLF +IFAK VN
Sbjct: 10 FRPQRFEDWGQEHITKTLQNALLQKKFSHAYLFSGPRGTGKTSAAKIFAKAVNCEHAPV 69
Query: 61 KGL GSPIEIDAASNNGVENVRNIIEDSRYKSMDSEFKVYIIDEVH 105
KG+ IEIDAASNNGV+ +R+I + ++ +KVYIIDEVH Sbjct: 70 DEPCNECAACKGITNGSISDVIEIDAASNNGVDEIRDIRDKVKFAPSAVTYKVYIIDEVH 129
Query: 106 MLSTGAFNALLKTLEEPSSGTVFILCTTDPQKIPDTILSRVQRFDFTRIDNDDIVNQLQF 165
MLS GAFNALLKTLEEP +FIL TT+P KIP TI+SR QRFDF Rl + IV ++ Sbjct: 130 MLSIGAFNALLKTLEEPPEHCIFILATTEPHKIPLTIISRCQRFDFKRITSQAIVGRMNK 189
Query: 166 IIESENEEGAGYSYERDALSFIGKLANGGMRDSITRLEKVLDYSHHVDMEAVSNAL G 222
I+++E E +L I A+GGMRD+++ L++ + +S D+ V +AL G
Sbjct: 190 IVDAEQ LQVEEGSLEIIASAAHGGMRDALSLLDQAISFSG--DILKVEDALLITG 242
Query: 223 VPDYETFASLVEAIANYDGSKCLEIVNDFHYSGKDLKLVTRNFTDFLLEVCKYWLVRDIS 282
L +++ + + S LE +N+ GKD + + + ++ Y + Sb]Ct: 243 AVSQLYIGKLAKSLHDKNVSDALETLNELLQQGKDPAKLIEDMIFYFRDMLLYKTAPGLE 302
Query: 283 ITQLPAHFESKLEQFCEAFQYPTLLWMLEEMNELAGWKWEPNAKPIIETKLL 335
+ + E L M++ +N+ +KW + + E ++ Sbjct: 303 GVLEKVKVDETFRELSEQIPAQALYEMIDILNKSHQEMKWTNHPRIFFEVAW 355
Query= si | 114835 | Ian |dplORF014 Phage dpi ORF | 50961-51974 | 3 (337 letters)
>sp|P47492|PRIM_MYCGE DNA PRIMASE >gi | 1361496 | pir| | F64227 DNA primase (dnaE) homolog MG250 - Mycoplasma genitalium (SGC3) >gi|3844848 (U39704) DNA primase (dnaE) [Mycoplasma genitalium] Length = 607
Score = 57.0 bits (135), Expect = 2e-07
Identities = 53/190 (27%), Positives = 89/190 (45%), Gaps = 17/190 (8%)
Query: 146 EELDKYRFIHP YMYERKLTDELIEMFDVGYDK--LHDCITFPVRNLKGETVFF 196
E +++Y FI+P Y++ K + + FD K + I P+ + G V F Sbjct: 170 ESMERYPFINPKIKPSELYLFS-KTNQQGLGFFDFNTKKATFQNQIMIPIHDFNGNPVGF 228
Query: 197 NRRSVRSKFHQYGEDDPKTEFLYGQYELVAFRDYFEKPISQVFVTESVINCLTLWSMKIP 256 - _
+ RSV + ++ EF + + EL+ K ++Q+F+ E + TL + K _
Sbjct: 229 SARSVDNINKLKYKNSADHEF-FKKGELLFNFHRLNKNLNQLFIVEGYFDVFTLTNSKFE 287
Query: 257 AVALMGVGGGN-QINLLKR--LPYRNIVLALDPDNAGQTAQEKLYRQLKRSK-VVRFIiNY 312
AVALMG+ + QI +K + +VLALD D +GQ A L +L + +V + + Sbjct: 288 AVALMGLALNDVQIKAIKAHFKELQTLVLALDNDASGQNAVFSLIEKLNNNNFIVEIVQW 347
Query: 313 PKEFYDNKWD 322
+ D WD Sbjct: 348 EHNYKD--WD 355
Query= sid| 114837 | lan|dplORF016 Phage dpi ORF| 43413-44303 | 3 (296 letters)
>emb|CAB07986 I (Z93946) N-acetylmuramoyl-L-alanine amidase [bacteriophage Dp-1] Length = 296
Score = 661 bits (1686), Expect = 0.0
Identities = 296/296 (100%) , Positives = 296/296 (100%)
Query: 1 MGVDIEKGVAWMQARKGRVSYSMDFRDGPDSYDCSSSMYYALRSAGASSAGWAVNTEYMH 60
MGVDIEKGVAWMQARKGRVSYSMDFRDGPDSYDCSSSMYYALRSAGASSAGWAVNTEYMH Sbjct: 1 MGVDIEKGVAWMQARKGRVSYSMDFRDGPDSYDCSSSMYYALRSAGASSAGWAVNTEYMH 60
Query: 61 AWLIENGYELISENAPWDAKRGDIFIWGRKGASAGAGGHTGMFIDSDNIIHCNYAYDGIS 120
AWLIENGYELISENAPWDAKRGDIFIWGRKGASAGAGGHTGMFIDSDNIIHCNYAYDGIS Sbjct: 61 AWLIENGYELISENAPWDAKRGDIFIWGRKGASAGAGGHTGMFIDSDNIIHCNYAYDGIS 120
Query: 121 VNDHDERWYYAGQPYYYVYRLTNANAQPAEKKLGWQKDATGFWYARANGTYPKDEFEYIE 180
VNDHDERWYYAGQPYYYVYRLTNANAQPAEKKLGWQKDATGFWYARANGTYPKDEFEYIE Sbjct: 121 VNDHDERWYYAGQPYYYVYRLTNANAQPAEKKLGWQKDATGFWYARANGTYPKDEFEYIE 180
Query: 181 ENKSWFYFDDQGYMLAEKWLKHTDGNWYWFDRDGYMATSWKRIGESWYYFNRDGSMVTGW 240
ENKSWFYFDDQGYMLAEKWLKHTDGNWYWFDRDGYMATSWKRIGESWYYFNRDGSMVTGW Sbjct: 181 ENKSWFYFDDQGYMLAEKWLKHTDGNWYWFDRDGYMATSWKRIGESWYYFNRDGΞMVTGW 240
Query: 241 IKYYDNWYYCDATNGDMKSNAFIRYNDGWYLLLPDGRLADKPQFTVEPDGLITAKV 296
IKYYDNWYYCDATNGDMKSNAFIRYNDGWYLLLPDGRLADKPQFTVEPDGLITAKV Sbjct: 241 IKYYDNWYYCDATNGDMKSNAFIRYNDGWYLLLPDGRLADKPQFTVEPDGLITAKV 296
Query= sid| 114841 | lan|dplORF020 Phage dpi ORF| 1864-2658 | 1 (264 letters)
>emb|CAB13247 I (Z99111) similar to coenzyme PQQ synthesis [Bacillus subtilis] Length = 243
Score = 217 bits (548), Expect = 5e-56
Identities = 117/248 (47%) , Positives = 163/248 (65%) , Gaps = 15/248 (6%)
Query: 23 MPIMEIFGPTIQGEGMVIGQKTIFIRTGGCDYHCNWCDSAFTWNGTTEPE--YITGKEAA 80
+P++EIFGPTIQGEGMVIGQKT+F+RT GCDY C+WCDSAFTW+G+ + + ++T +E Sbjct: 5 IPVLEIFGPTIQGEGMVIGQKTMFVRTAGCDYSCSWCDSAFTWDGSAKKDIRWMTAEEIF 64
Query: 81 SRILKLAFNDKGEQICNHVTLTGGNPALINEPMAKMISILKEHGFKFGLETQGTRFQEWF 140
+ + D G +HVT++GGNPAL+ + + I +LKE+ + LETQGT +Q+WF Sbjct: 65 AEL KDIGGDAFSHVTISGGNPALLKQ-LDAFIELLKENNIRAALETQGTVYQDWF 118
Query: 141 KEVSDITISPKPPSSGMRTNMKILEAIVDRM--NDENLDWSFKIVIFDENDIiAYARDMFK 198
+ D+TISPKPPSS M TN + L+ 1+ + ND S K+VIF++ DL +A+ + K Sbjct: 119 TLIDDLTISPKPPSSKMVTNFQKLDHILTSLQENDRQHAVSLKWIFNDEDLEFAKTVHK 178
Query: 199 TFEGKLRPVNYLSVGNANAY--EEGKIΞDRLLEKLGWLWDKVYEDPAFNNVRPLPQLHTL 256
+ G YL VGN + + ++ + LL K L DKV D N VR LPQLHTL Sbjct: 179 RYPG---IPFYLQVGNDDVHTTDDQSLIAHLLGKYEALVDKVAVDAELNLVRVLPQLHTL 235
Query: 257 VYDNKRGV 264
++ NKRGV Sbjct: 236 LWGNKRGV 243
Query= sid | 114842 | Ian|dplORF021 Phage dpi ORF| 2504-3295 | 2 (263 letters)
>sp|P19465|GCHl_BACSU GTP CYCLOHYDROLASE I (GTP-CH-I) >gi | 98411 |pir | |A38256 GTP cyclohydrolase I (EC 3.5.4.16) - Bacillus subtilis >gi|l43231 (M37320) regulatory protein [Bacillus subtilis] >gi 1143799 (M80245) MtrA [Bacillus subtilis] >gi|2634696|emb|CAB14194| (Z99115) GTP cyclohydrolase I [Bacillus subtilis] Length = 190
Score = 208 bits (523), Expect = 4e-53
Identities = 103/185 (55%) , Positives = 133/185 (71%) , Gaps = 1/185 (0%)
Query: 80 VTLDNTEaAVQRLFGLLGEDAERDGLQDTPFRFVKALAEHTVGYREDPKLHLEKTFDVDH 139
V + E AV+++ +GED R+GL DTP R K AE G EDPK H + F +H Sbjct: 4 VNKEQIEQAVRQILEAIGEDPNREGLLDTPKRVAKMYAEVFSGLNEDPKEHFQTIFGENH 63
Query: 140 EDLVLVKDIPFNSLCEHHLAPFVGKVHIAYIPKD-KITGLSKFGRWEGYAKRLQVQERL 198
E+LVLVKDI F+S+CEHHL PF GK H+AYIP+ K+TGLSK R VE AKR Q+QER+ Sbjct: 64 EELVLVKDIAFHSMCEHHLVPFYGKAHVAYIPRGGKVTGLΞKLARAVEAVAKRPQLQERI 123
Query: 199 TQQIADAIQEVLNPQAVAVIVEAEHTCMSGRGIKKHGATTVTSTMRGLFQDDASARAELL 258
T IA++I E L+P V V+VEAEH CM+ RG++K GA TVTS +RG+F+DDA+ARAE+L Sbjct: 124 TSTIAESIVETIiDPHGVMVVVEAEHMCMTMRGVRKPGAKTVTSAVRGVFKDDAAARAEVL 183
Query: 259 QLIKK 263
+ IK+ Sbjct: 184 EHIKR 188
Query= sid| 114843 | lan|dplORF022 Phage dpi ORF| 30896-31675 | 2 (259 letters)
>gi 12347102 (U77367) internalin [Listeria monocytogenes] Length = 821
Score = 55.0 bits (130), Expect = 5e-07
Identities = 44/149 (29%) , Positives = 63/149 (41%) , Gaps = 13/149 (8%)
Query: 119 FRMNIYVPNYVG--DSIVNYVKITLNNCTGKAPGLSIGKEFYAPEFNIKAREATKAGLPV 176
F + VPN + D + + NN T AP L Y PE +K + K + Sbjct: 383 FSKTLSVPNNITSIDGTLIAPETISNNGTYDAPNLKWSLPNYLPE--VKYTFSQKIPIGT 440
Query: 177 KSMDYVAQLPAVLR RVTFDLNGGTGTADAVRVEAGKKISPKPVDPTLTGKAFKGW 231
+ +Y + L+ +VTF++ G T + V E + P+P PT G F GW Sbjct: 441 GTSNYSGFITQPLKELLDYKVTFNVEGNTSEVETVTEE---NLIPEPTSPTKQGYTFDGW 497
Query: 232 -KVEGESTIWDFDNHMMPDRDVKLVAQFA 259
E T WDF MP D+ L A F+ Sbjct: 498 YDAETGGTKWDFTTGQMPANDLTLYAHFS 526
Score = 43.4 bits (100), Expect = 0.002
Identities = 47/195 (24%), Positives = 73/195 (37%), Gaps = 12/195 (6%)
Query: 72 YDLTFKDNTFDPEIMALIEGGTVRQQGGTIAGYDT-PMLAQGASNMKPFRMNIYVPNY-- 128
YD + T + +G + GG + T M A + F +N Y N+ Sbjct: 547 YDALLNEPTTPTKQGYTFDGWYDAETGGNKWDFKTMKMPANDVAFYAHFTINNYQANFDI 606
Query: 129 ---VGDSIVNYVKITLNNCTGKAPGLSIGKEFYAPEFNIKAREATKAGLPVKSMDYVAQL 185
V + + Y + T G + + A K TK +P + A Sbjct: 607 DGEVKNETIAYDTLLNEPTTPTKQGYTFDGWYDAETGGTKWDFKTKE-MPANDVTLYAHF 665
Query: 186 PAVLRRVTFDLNGGTGTADAVRVEAGKKISPKPVDPTLTGKAFKGW-KVEGESTIWDFDN 244
+ FD++G T + V +A + P+P P+ TG +GW E T WDF Sbjct: 666 TINNYQANFDIDGAV-TEEWNYDA---LIPEPTSPSKTGFTLEGWYDAEVGGTKWDFKT 721
Query: 245 HMMPDRDVKLVAQFA 259
MP D+ L A F+ Sbjct: 722 MKMPANDITLYAHFS 736
Score = 38.3 bits (87), Expect = 0.057
Identities = 42/169 (24%) , Positives = 59/169 (34%) , Gaps = 10/169 (5%)
Query: 96 QQGGTIAGYDT-PMLAQGASNMKPFRMNIYVPNYVGDSIVNYVKIT LNNCTGKAPG 150
+ GGT + T MA + F +N Y + D +V + LN T Sbjct: 501 ETGGTKWDFTTGQMPANDLTLYAHFSVNSYQANFDIDGWTNEAWYDALLNEPTTPTKQ 560
Query: 151 LSIGKEFYAPEFNIKAREATKAGLPVKSMDYVAQLPAVLRRVTFDLNGGTGTADAVRVEA 210
+Y E + +P + + A + FD++G A
Sbjct: 561 GYTFDGWYDAETGGNKWDFKTMKMPANDVAFYAHFTINNYQANFDIDGEVKNETI A 616
Query: 211 GKKISPKPVDPTLTGKAFKGW-KVEGESTIWDFDNHMMPDRDVKLVAQF 258
+ +P PT G F GW E T WDF MP DV L A F Sbjct: 617 YDTLLNEPTTPTKQGYTFDGWYDAETGGTKWDFKTKEMPANDVTLYAHF 665
Query= sid| 114850 | lan|dplORF029 Phage dpi ORF| 662-1348 | 2 (228 letters)
>gi 12650185 (AE001074) succinoglycan biosynthesis regulator (exsB) [Archaeoglobus fulgidus] Length = 239
Score = 119 bits (295) , Expect = 2e-26
Identities = 79/224 (35%) , Positives = 113/224 (50%) , Gaps = 11/224 (4%)
Query: 1 MKSWLLSGGVDSATCLAIEVDKWGSKNVHAIAFNYGQKHEAELENAANVAMFYGVKFTI 60
MK+V+LLSGG+DS+T L +D G VHA+ F YGQKH E+E+A VA V+ Sbjct: 1 MKAVMLLSGGIDSSTLLYYLLD--GGYEVHALTFFYGQKHSKEIESAEKVAKAAKVRHLK 58
Query: 61 LEIDSKIYXXXXXXLLQGKGEISHGKSYAEILAEKEWDTYVPFRNGLMLSQXXXXXXXX 120
++I S 1+ L G+ E+ Y+E + + T VP RN ++LS Sbjct: 59 VDI-STIHDLISYGALTGEEEVPKA-FYSEEVQRR TIVPNRNMILLS--IAAGYAV 110
Query: 121 XXXXXXXXJCXXXXXXXXXXPDCTPEFYNSMSNAMEYGT-GGKVTLVAPLLTLTKAQVVKW 179
PDC EF ++ A+ V + AP + +TKA +V+ Sbjct: 111 KIGAKEVHYAAHLSDYSIYPDCRIEFVKALDTAVYLANIWTPVEVRAPFVDMTKADIVRL 170
Query: 180 GIDLDVPYFLTRSCYESDAESCGTCATCIDRKKAFEENGMTDPI 223
G+ L VPY LT SCYE C +C TC++R +AF NG+ DP+ Sbjct: 171 GLKLGVPYELTWSCYEGGDRPCLSCGTCLERTEAFLANGVKDPL 214
Query= sid| 114855 | lan|dplORF034 Phage dpi ORF| 131-652 | 2 (173 letters)
>emb|CAB13248 I (Z99111) similar to hypothetical proteins [Bacillus subtilis] Length = 165
Score = 220 bits (556) , Expect = 4e-57
Identities = 103/139 (74%) , Positives = 117/139 (84%)
Query: 5 TTRTDAELTGVTLLGNQDTKYDYDYNPDVLETFPNKHPENNYLVTFDGYEFTSLCPKTGQ 64
TTR ++EL GVTLLGNQ T Y ++Y PDVLE+FPNKH +Y V F+ EFTSLCPKTGQ Sbjct: 2 TTRKESELEGVTLLGNQGTNYLFEYAPDVLESFPNKHVNRDYFVKFNCPEFTSLCPKTGQ 61
Query: 65 PDFANVFISYIPNEKMVESKSLKLYLFSFRNHGDFHEDCMNIILNDLYELMEPKYIEVMG 124
PDFA ++ISYIP+EKMVESKSLKLYLFSFRNHGDFHEDCMNII+NDL ELM+P+YIEV G Sbjct: 62 PDFATIYISYIPDEKMVESKSLKLYLFSFRNHGDFHEDCMNIIMNDLIELMDPRYIEVWG 121
Query: 125 LFTPRGGISIYPFVNKVNP 143
FTPRGGISI P+ N P Sbjct: 122 KFTPRGGISIDPYTNYGKP 140
Query= sid| 114857 | lan|dplORF036 Phage dpi ORF| 48808-4936211 (184 letters)
>gi 11353529 (U38906) ORF12 [Bacteriophage rlt] Length = 296
Score = 53.5 bits (126), Expect = le-06
Identities = 42/149 (28%), Positives = 70/149 (46%), Gaps = 9/149 (6%)
Query: 34 IASNTVGNGKTSWAVRLLQRYLAETALDGRIVEKGMFWSAQLLTEFGDYNYFQTMQEFL 93
+ S G GK+ A+ +L+ L T L ++ V + F + + F + + F+ Sbjct: 155 WSGPAGTGKSHLAMSILKDCLQHTDLT--VIFASWSEVLHLIKDSFDNKDSFYSTEYFM 212
Query: 94 ERFERLKTCELLVIDEIGGGSLTKASYPYLYDLVNYRVDNNLSTIYTTNYTDDEIIDLLG 153
E F + +LLVID+IG +T+ S L ++++ R TI TTN DEI Sbjct: 213 EVF RNTDLLVIDDIGSEKITEWSMSLLTEVLDART KTIITTNLKSDEIRKKYH 265
Query: 154 QRLYSRIYDTSWLDFQASNVRGLEVSEI 182
R YSR++ F N++ VS++ Sbjct: 266 NRTYSRLFRGIGKKAFNFENIKDKRVSQL 294
Query= sid | 114859 | Ian |dplORF038 Phage dpi ORF| 1350-1871 |3 (173 letters)
>sp|P44123|YB90_HAEIN HYPOTHETICAL PROTEIN HI1190 >gi | 1074675 | ir | | F64021 hypothetical protein HI1190 - Haemophilus influenzae (strain Rd KW20) >gi 11574117 (U32798) 6-pyruvoyl tetrahydrobiopterin synthase, putative [Haemophilus influenzae Rd] Length = 141
Score = 100 bits (247) , Expect = 6e-21
Identities = 59/143 (41%) , Positives = 83/143 (57%) , Gaps = 10/143 (6%)
Query: 2 RVSKTLTFDAAHQLVGHFGKCANLHGHTYKVEISLAGGTYDHGSSQGMWDFYHVKKIA- 60
++SK +FD AH L GH GKC NLHGHTYK+++ ++G Y G+ + MV+DF +K I Sbjct: 3 KISKEFSFDMAHLLDGHDGKCQNLHGHTYKLQVEISGDLYKSGAKKAMVIDFSDLKSIVK 62
Query: 61 GTFIDRLDHAVLL-QGNEP IALANAVDTKRVLFGFRTTAENMSRFLTWTLTELMWK 115
+D +DHA + Q NE L +++K FRTTAE ++RF+ L + Sbjct: 63 KVILDPMDHAFIYDQTNERESQIATLLQKLNSKTFGVPFRTTAEEIARFIFNRLKH--DE 120
Query: 116 HARIDSIKLWETPTGCAECTYYE 138
I SI+LWETPT + C Y E Sbjct: 121 QLSISSIRLWETPT--SFCEYQE 141
Query= sid| 114860 | lan|dplORF039 Phage dpi ORF| 3306-3803 | 3 (165 letters)
>emb| CAA68244 I (X99978) ORF7; hydophobic protein [Lactobacillus plantarum] Length = 168
Score = 64.4 bits (154), Expect = 5e-10
Identities = 49/156 (31%) , Positives = 84/156 (53%) , Gaps = 9/156 (5%)
Query: 8 WLVRTALIAALYVTLTVAFSAISY--GPIQFRVSEALILLPLWNHRWTPGIVLGTIIANF 65
W++ AL+AA+YV L + +A S G IQFRVSE L L ++N ++ GIV G 1+ + Sbjct: 9 WIIN-ALVAAMYWLCLGPAAFSLASGAIQFRVSEGIiNHLAVFNRKYIWσiVAGVILFDA 67
Query: 66 FSP-LGLIDVLFGSLATFLGXXXXXXXXXXXSPLYSLICPVLA NAYLIALELRIVY 120
F P L++VLFG + L ++ + +A + ++IAL + ++
Sbjct: 68 FGPGASLLNVLFGGGQSLLALLVLTWLAPKLKTVWQRMLLNIALFTVSMFMIALMITMMS 127
Query: 121 S-LPFWESVIYVGISEAIIVLIΞYFLISTLAKNNHF 155
S + FW + + +SE 11+ 1+ ++ +L + HF Sbjct: 128 SGVAFWPTYLTTALSELIIMSITAPIMYSLDRVLHF 163
Query= sid| 114862 | lan|dplORF041 Phage dpi ORF| 8208-8699| 3 (163 letters)
>gi 12522313 (AF012906) dUTPase homolog [Bacillus subtilis] >gi|2634394|emb|CAB13893| (Z99114) similar to deoxyuridine 5 ' -triphosphate nucleotidohydrolase [Bacillus subtilis] >gi| 3025643 (AF020713) putative dUTPase [Bacteriophage SPBc2] Length = 142
Score = 108 bits (267) , Expect = 2e-23
Identities = 65/160 (40%) , Positives = 83/160 (51%) , Gaps = 25/160 (15%)
Query: 5 VDVKMIDPKLDRLKYT--GDWVDVRISSITKIDADSADVSRCRKVLQKAQVYSVAAGECI 62
+ +K +D R+ GDW+D+R + I D + - -
Sbjct: 3 IKIKYLDETQTRINKMEQGDWIDLRAAEDVAIKKDEFKL 41 --
Query: 63 KIAHGFALELPKGYEAILHPRSSLFKKTGLIFVSS-GVIDEGYKGDTDEWFSVWYATRDA 121
+ G A+ELP+GYEA + PRSS +K G+I +S GVIDE YKGD D WF YA RD Sbjct: 42 -VPLGVAMELPEGYEAHWPRSSTYKNFGVIQTNSMGVIDESYKGDNDFWFFPAYALRDT 100
Query: 122 DIFYDQRIAQFRIQEKQPAIKFNFVESLGNAARGGHGSTG 161
I Rl QFRI +K PA+ V+ LGN RGGHGSTG Sbjct: 101 KIKKGDRICQFRIMKKMPAVDLIEVDRLGNGDRGGHGSTG 140
Query= si | 114867 | Ian |dplORF046 Phage dpi ORF | 42774-43202 | 3 (142 letters)
>emb|CAB07984 I (Z93946) hypothetical protein [bacteriophage Dp-1] Length = 142
Score = 287 bits (728) , Expect = 2e-77
Identities = 142/142 (100%) , Positives = 142/142 (100%)
Query: 1 MPMWLNDTAVLTTIITACSGVLTVLLNKLFEWKSNKAKSVLEDISTTLSTLKQQVDGIDQ 60
MPMWLNDTAVLTTIITACSGVLTVLLNKLFEWKSNKAKSVLEDISTTLSTLKQQVDGIDQ Sbjct: 1 MPMWIiNDTAVLTTIITACSGVLTVLLNKLFEWKSNKAKSVLEDISTTLSTLKQQVDGIDQ 60
Query: 61 TTVAINHQNDVIQDGTRKIQRYRLYHDLKREVITGYTTLDHFRELSILFESYKNLGGNGE 120
TTVAINHQNDVIQDGTRKIQRYRLYHDLKREVITGYTTLDHFRELSILFESYKNLGGNGE Sbjct: 61 TTVAINHQNDVIQDGTRKIQRYRLYHDLKREVITGYTTLDHFRELSILFESYKNLGGNGE 120
Query: 121 VEALYEKYKKLPIREEDLDETI 142
VEALYEKYKKLPIREEDLDETI Sbjct: 121 VEALYEKYKKLPIREEDLDETI 142
Query= sid| 11490l| Ian| dplORFO80 Phage dpi ORF| 42490-42759 | 1 (89 letters)
>emb|CAB07983 I (Z93946) hypothetical protein [bacteriophage Dp-1] Length = 124
Score = 147 bits (367) , Expect = le-35
Identities = 75/75 (100%) , Positives = 75/75 (100%)
Query: 1 MLNLTKSRQIVAEFTIGQGAEKKLVKTTIVNIDANAVSTVSETLHDPDLYAANRRELRAD 60
MI-^LTKSRQIVAEFTIGQGAEKKLVKTTIVNIDANAVSTVSETLHDPDLYAANRRELRAD Sbjct: 1 MLNLTKSRQIVAEFTIGQGAEKKLVKTTIVNIDANAVSTVSETLHDPDLYAANRRELRAD 60
Query: 61 EQKLRETRYAIEDEI 75
EQKLRETRYAIEDEI Sbjct: 61 EQKLRETRYAIEDEI 75
Query= si | 114912 | Ian|dplORF091 Phage dpi ORF| 43189-43413 | 1 (74 letters)
>emb|CAB07985| (Z93946) holin [bacteriophage Dp-1] Length = 74
Score = 63.2 bits (151), Expect = 2e-10 Identities > 34/74 (45%) , Positives = 34/74 (45%)
Query: 1 MKLSNEQYDXXXX_-XX-_X_-_XX>_XXXXXXXXXYQFDX_-XXX^ 60
MKLSNEQYD YQFD VLGVSSR
Sbjct: 1 MKLSNEQYDVAKNVVTVWPAAIALITGLGALYQFDTTAITGTIALLATFAGTVLGVSSR 60
Query: 61 NYQKEQEAQNNEVE 74
NYQKEQEAQNNEVE Sbjct: 61 NYQKEQEAQNNEVE 74
Condensed listing of homology information from above
Phage: dpi
Database: nr
Program- Blastp
Query= sιd| 114822 | lan|dplORF001 Phage dpi ORF| 36698-40390 | 2 (1230 letters) i 2444124 (U88974) ORF45 [Streptococcus thermophilus temperate 467 e-130 gi 928828 (L44593) ORF1904; putative [Lactococcus lactis phage B. 427 e-118
91 2935676 (AF032121) unknown [Streptococcus thermophilus bacter 309 le-82 91 2935691 (AF032122) unknown [Streptococcus thermophilus bacter. 306 7e-82 9i 3540289 (AF057033) putative anti-receptor [Streptococcus ther 279 6e-74
9 4530154 |gb|AAD21894.11 (AF085222) putative tail-host specific. 220 3e-56 g 930045 |emb|CAA33387| (X15332) alpha-1 (III) collagen [Homo sa. 58 4e-07 gi 1070603 I ir | | CGHU7L collagen alpha l(III) chain precursor - h. 58 4e-07 gi 4502951 jref |NP_000081.1|PCOL3A1| collagen, type III, alpha 1 58 4e-07
91 115290|spjP04258|CA13_BOVIN COLLAGEN ALPHA l(III) CHAIN >gι | 7. 58 4e-07 gi 575322 |emb|CAA36279| (X52046) type III collagen [Mus musculus] 57 8e-07 g 2119163 |pιr I |S59856 collagen alpha l(III) chain precursor - m. 57 8e-07 gi 543912|sp|P1394l|CA13_RAT COLLAGEN ALPHA 1 (III) CHAIN >gι|543. 57 ie-06 91 3171998|emb|CAA06510| (A 005395) collagen alpha 1 (III) [Ratt. 57 le-06 gi 3947565 |emb|CAA90250| (Z49967) similar to collagen; cDNA EST . 54 7e-06 91 423403 |pιr I |A46053 bullous pemphigoid antigen, BPAG2, type XV. 53 9e-06
91 115410|sp|P12114|CCSl_CAEEL CUTICLE COLLAGEN SQT-1 >gι| 84437 | 53 9e-06 9 387380l|emb|CAA90084 I (Z49907) cuticle collagen SQT-1; cDNA E. 53 9e-06
Query= sιd| 114823 I lan|dplORF002 Phage dpi ORF| 32386-3583511 (1149 letters)
91 33419221 dbj |BAA31888| (AB009866) orf 15 [bacteriophage phi PVL] ] 280 3e-74 9i 4126622 j db;) |BAA36642.l| (AB016282) ORF36 [bacteriophage phi-105 5]] 232 le-59 91 1369948 |emb|CAA59194 I (X84706) host interacting protein [Bact. 201 3e-50 9 3139112 (AF063097) gpT [Bacteriophage P2J 188 2e-46 9 3337272 (U32222) G protein (Bacteriophage 186] 161 3e-38 9 4063799|db_ |BAA36253 | (AB008550) orf25; similar to T gene of . 159 8e-38 91 3172274 (AF022214) minor tail subunit; putative tape-measure . 123 6e-27 9 465127|sp|Q05233|VG26_BPML5 MINOR TAIL PROTEIN GP26 >g | 41904. 108 2e-22 9i 3540284 (AF057033) putative minor tail protein [Streptococcus. 99 2e-19 9 2444119 (U88974) ORF40 [Streptococcus thermophilus temperate 90 6e-17 9 2634555|emb|CAB14053| (Z99115) yo l [Bacillus subtilis] >gι | 3. 66 le-09 9 2392838 (AF011378) unknown [Bacteriophage ski] 64 5e-09 9i 2764873 | emb | CAA66557 | (X97918) gene 18.1 [Bacteriophage SPP1] 62 3e-08 9i 1353559 (U38906) ORF42 [Bacteriophage rlt] 61 6e-08 91 630841 |pιr I |S39079 puff C-8 protein - fungus gnat (Rhynchosci. 55 2e-06 91 1730865 |sp|P51731|Y027_BPHPl HYPOTHETICAL 72.8 KD PROTEIN IN . 53 8e-06 9i 224288 I prf j 11101273J ORF 7 [Bacteriophage HP1] S3 le-05
Query= sid | 114824 | lan|dplORF003 Phage dpi ORF| 53538-55877 | 3 (779 letters)
9 118825|sp|P00582|DPOl_ECOLI DNA POLYMERASE I (POL I) >gi|6705.. 193 3e-48 91 2982102 |pdb|lKFS| A Cham A, All-Oxygen Dna Complexed To The 3 . 193 3e-48 91 229889|pdb| 1DPI I DNA Polymerase I (Klenow Fragment) (E.C.2... 193 3e-48 gi 1169402 |sp|P43741|DP01_HAEIN DNA POLYMERASE I (POL I) >gi|l07 191 le-47
91 2688462 (AE001156) DNA polymerase I (polA) [Borrelia burgdorf .. 190 3e-47 g 809180 |pdb|lKLN|A Escherichia coli 190 3e-47 gi 1913934|emb|CAA72997| (Y12328) DNA-directed DNA polymerase I 189 8e-47 g 4090935 (AF028719) DNA polymerase type I [Rhodothermus sp. 'I. 175 le-42 gi 473157l|gb|AAD28505.1|AF121780_l (AF121780) DNA polymerase I . 174 2e-42 gi 1633576 (U57757) similar to proofreading 3' -5' exonuclease an. 173 4e-42 3322368 (AE001195) DNA polymerase I (polA) [Treponema pallidum] 172 9e-42 gi 1006595 | db_ |BAA10748| (D64005) DNA polymerase I [Synechocysti.. 171 2e-41 gi 585062|sp|Q07700|DPOl_MYCTU DNA POLYMERASE I (POL I) >gι|4161.. 163 5e-39 gi 4376908|gb|AAD1875l| (AE001645) DNA Polymerase I [Chlamydia p . 157 2e-37 1169403 |sp|P46835|DP01_MYCLE DNA POLYMERASE I (POL I) >gι|l07 152 7e-36 gi 2145839 |pιr I I S72949 DNA polymerase I - Mycobacterium leprae >.. 152 7e-36 gi 1405438 |emb|CAA67184| (X98575) DNA-dependent DNA polymerase [ . 152 9e-36 gi 2506365 I sp I P80194 I DP01_THECA DNA POLYMERASE I, THERMOSTABLE (.. 147 2e-34 gi 3328929 (AE001322) DNA Polymerase I [Chlamydia trachomatis] 147 3e-34
91 3913510 I sp I 052225 I DP01_THEFI DNA POLYMERASE I THERMOSTABLE ( 146 7e--34 91 1205984 (U33536) DNA polymerase I [Bacillus stearothermophilus] 146 7e- -34 gi 118827|sp|P13252|DP01_STRPN DNA POLYMERASE I (POL I) >gι|9802 145 9e- -34 gi 1942202|pdb|lJXE| Ξtoffel Fragment Of Taq Dna Polymerase I 145 le- -33 91 1943520 |pdb|lKTQ| Dna Polymerase 145 le- -33 9 1084022|pιrj |JX0359 DNA-directed DNA polymerase (EC 2 7 7 7) 145 le- -33 91 5078911 db] |BAA06775| (D32013) DNA Polymerase [Thermus aquaticus] 145 le- -33 gi 118828|sp|P19821|DP01_THEAQ DNA POLYMERASE I THERMOSTABLE (T 145 le- -33 91 1706502 I sp|P52028|DPOl_THETH DNA POLYMERASE I, THERMOSTABLE ( 144 2e- -33 91 10972111 prf | | 2113329A DNA polymerase [Thermus aquaticus therm 144 2e- -33 g 2098289|pdbj 1TAU|A Cham A, Structure Of Dna Polymerase 143 3e- -33
Query= sιd| 114825 | lan| dplORF004 Phage dpi ORF| 40401-42440 | 3 (679 letters) gι|l93476l|emb|CAB07981| (Z93946) hypothetical prote [bacterio 1011 0 0 gij 3540290 (AF057033) putative minor structural protein [Strepto 346 2e- -94 gi 12444125 (U88974) ORF46 [Streptococcus thermophilus temperate 339 3e- -92 gι|l934762|emb|CAB07982| (Z93946) hypothetical protem [bacterio 300 2e- -80 gι|4530155|gb|AAD21895 l| (AF085222) unknown [Streptococcus ther 276 4e- -73 gi 12935677 (AF032121) unknown [Streptococcus thermophilus bacter 250 3e- -65 gi 12935692 (AF032122) unknown [Streptococcus thermophilus bacter 250 3e- •65 gi 11136289 (U42597) histidine kinase A [Dictyostelium discoideum] 50 7e-05
Query= sιd| 114827 | lan| dplORF006 Phage dpi ORF| 45296-46987 | 2 (563 letters)
9i 4377165 gb|AAD18987| (AE001666) SWI/SNF family helιcase_2 [Ch 171 le- -41 gi 1769947 emb|CAA67095| (X98455) SNF [Bacillus cereus] 160 3e- -38 g 3329163 (AE001341) SWF/SNF family helicase [Chlamydia trachom 159 6e- -38 gi 4377149 gb|AAD18973| (AE001664) SWI/SNF family helιcase_l [Ch 157 2e- -37 91 3328995 (AE001326) SWI/SNF family helicase [Chlamydia trachom 153 2e- -36 gi 2493354 sp|P75093|Y018_MYCPN HYPOTHETICAL HELICASE MG018/MG01 146 4e- -34 gi 1653748 db] |BAA18659| (D90916) helicase of the snf2/rad54 fam 143 3e- -33 gi 1763712 emb|CAB05939| (Z83337) member of the SNF2 helicase fa 143 4e- -33 gi 2636153 emb|CAB15645 l| (Z99122) similar to SNF2 helicase [Ba 143 4e- -33 gi 2909552 emb|CAA17284| (AL021924) helZ [Mycobacterium tubercul 140 2e- -32 gi 3844627 (U39681) ATP-dependent RNA helicase, putative [Mycopl 136 3e- -31 g 1351463 sp|P47264|Y018_MYCGE HYPOTHETICAL HELICASE MG018 136 4e- -31 91 2660669 (AC002342) human Mι-2 autoantigen-like prote [Arabi 131 2e- -29 gi 1361537 pir I 1164201 helicase (motl) homolog - Mycoplasma gem 129 4e- -29 91 3482977 embjCAA20533 l| (AL031369) putative protein [Arabidop 128 9e- -29 gi 3298562 (U91543) zinc-finger helicase [Homo sapiens] 120 2e- -26 gi 3875971 emb|CAB0249l| (Z80344) similar to helicase, cDNA EST 120 2e- -26 gi 4557451 ref |NP_001263 11 PCHD3 | chromodomam helicase DNA bind 120 2e- -26 gi 2645435 (AF007780) CHD3 [Drosophila melanogaster] 118 le- -25 91 3875165 I emb| CAA91798 I (Z67881) Similarity to Mouse Chromodoma 118 le- ■25
Query= sιd| 114828 | lan|dplORF007 Phage dpi ORF| 22230-23621 | 3 (463 letters) gι| 2444105 (U88974) ORF26 [Streptococcus thermophilus temperate 89 7e-17 gι|3318666 (U19754) BBA31 homolog [Borrelia burgdorferi] 59 7e-08 gij 2690260 (AE000790) conserved hypothetical protein [Borrelia b 56 5e-07
Query= sιd| 114829 | lan|dplORF008 Phage dpi ORF] 49624-50961 | 1 (445 letters) gι|4406210|gb|AAD19901| (AF100420) DnaB replication fork helicas 68 2e- -10 gi j 3121983 |spj 025916 |DNAB_HELPY REPLICATIVE DNA HELICASE >gι|231 67 2e- -10 gι|4416322JgbJAAD20314| (AF106032) replicative helicase, DnaB [B 65 9e- -10 gij 4155895 (AE001551) REPLICATIVE DNA HELICASE [Helicobacter pyl 60 4e- -08 gij 3322317 (AE001191) replicative DNA helicase (dnaB) [Treponema 58 le- -07 gιjl3803l|sp|P04530|VG41_BPT4 PRIMASE-HELICASE (PROTEIN GP41) >g 53 3e- -06 gι|2983861 (AE000742) replicative DNA helicase [Aquifex aeolicus] 51 le- •05
Query= sιd| 114831 | lan|dplORF010 Phage dpi ORF| 8699-9859 | 2 (386 letters) gi 12760912 (AF037258) RecA protem [Chlorobium tepidum] 133 2e- -30 gιJ3219851|sp|P94666|RECA_CLOPE RECA PROTEIN >gι| 1698591 (U61497 129 3e- -29 gιjl350566|spjP48295|RECA_STRVL RECA PROTEIN >gι|508860 (U04837) 128 7e- •29 gι|744163 |prf j |2014250A recA-like protein [Streptomyces violaceus] 126 3e- -28 gι|730487Jsp|P41054|RECA_STRAM RECA PROTEIN >gι | 511133 | emb | CAA82 125 4e- •28 gιJ2687334|emb|CAA15875| (AL020958) RecA protein [Streptomyces c 125 6e- -28 g jl350565|sp|P48294|RECA_STRLI RECA PROTEIN >gι | 481482 |pιr| | S38 125 6e- •28
gi 464599| sp|P33542|RECA_AQUPY RECA PROTEIN >gι 11086167 | pir \ |A55.. 123 2e-27
91 417636J sp|P32725|RECA_RHOSH RECA PROTEIN >gι j 541307 | pir | j S415.. 123 2e-27
91 2984348 (AE000775) recombination protein RecA [Aquifex aeolicus ] 123 2e-27 gi 3219854 |sp|P95846|RECA_STRRM RECA PROTEIN >gι 11729800 | emb \ CAA.. 122 4e-27 gi 2500086 jsp|Q5956θJRECA_MYCSM RECA PROTEIN >gι | 1430892 j emb j CAA.. 122 4e-27 gi 1350567 jspjp48296JRECA_THEAQ RECA PROTEIN >gι j 1072963 jpir j | A5... 122 6e-27 gi 625663| pirj I X0292 recA protein - Thermus aquaticus (strain HB8) ) 121 le-26 gi 1172880 |spjP42440|RECA_CAMJE RECA PROTEIN _gι | 2119991 |pιr | | 14. 120 2e-26 gi 4154654 (AE001453) RECA PROTEIN. [Helicobacter pylori J99] 120 2e-26
91 1072968 |pιr||C55020 recA protein - Thermus sp >gι | 458472 |db_ | . 120 2e-26 gi 3219852 |sp|P95469|RECA_PARDE RECA PROTEIN >gι| 1825468 (U59631. 119 3e-26 gi 2507284 jsp|P42445|RECA_HELPY RECA PROTEIN >gι | 2313235 | gb |AAD0. 119 4e-26
91 1172890 jspJQ02350|RECA_STAAU RECA PROTEIN >gι|463285 (L25893). 118 5e-26
91 4416209 jgb|AAD2026l| (AF094756) RecA prote [Bifidobacterium . 118 5e-26 gi 2500084 jsp|Q59180|RECA_BORBU RECA PROTEIN _gι| 1276443 (U23457 118 5e-26
Query= sιd| 114832 | lan|dplORF011 Phage dpi ORF| 28017-29096 | 3 (359 letters) gi 12444110 (U88974) ORF31 [Streptococcus thermophilus temperate . 187 le-46 gι| 3320438 (AF057033) gp348 [Streptococcus thermophilus bacterio. 179 2e-44 gij 479514 |pir I I S34244 hypothetical prote p38 - act ophage VWB. 62 8e-09
Query= sid | 114834 ] Ian | dplORF013 Phage dpi ORF| 10215-11240 | 3 (341 letters) gι|580855|emb|CAA29958| (X06803) dnaZX-like ORF put. DNA polymer. 182 2e-45 gι|118807|sp|P09122|DP3X_BACSU DNA POLYMERASE III SUBUNITS GAMMA. 182 2e-45 gιJ98292|pιrj |S13786 DNA-directed DNA polymerase (EC 2.7.7.7) II. 182 2e-45 gιjl527142 (U66040) DNA polymerase III gamma subunit [Salmonella. 172 4e-42 gιJ2494197|sp|P74876|DP3X_SALTY DNA POLYMERASE III SUBUNITS GAMM. 172 4e-42 gιjll8808|sp|P06710|DP3X_ECOLI DNA POLYMERASE III SUBUNITS GAMMA, 170 le-41 gij 4155207 (AE001497) DNA POLYMERASE III SUBUNITS GAMMA AND TAU . 169 2e-41 gιJ231384l|gb|AAD07767.l| (AE000584) DNA polymerase III gamma an. 168 4e-41 gij 2583049 (AF025391) DNA polymerase III holoenzyme tau subunit . 166 3e-40 gij 2984127 (AE000759) DNA polymerase III gamma subunit [Aquifex . 166 3e-40 giJ3861390|emb|CAA15289| (AJ235273) DNA POLYMERASE III SUBUNITS . 165 5e-40 gιjll69397|sp|P43746|DP3X_HAEIN DNA POLYMERASE III SUBUNITS GAMM. 156 2e-37 gij 1293572 (U49738) DNA polymerase III tau homolog DnaX [Cauloba. 151 8e-36 gij 3328753 (AE001306) DNA Pol III Gamma and Tau [Chlamydia trach. 148 4e-35 gιJ4376294|gb|AAD18193| (AE001589) DNA Polymerase III Gamma and . 148 5e-35 gij 581255 I emb JCAA28175J (X04487) alternate dnaZX protein (AA 1-6. 146 3e-34 gιJ2688379 (AE001151) DNA polymerase III, subunits gamma and tau. 140 2e-32 gij 3323329 (AE001268) DNA polymerase III, subunits gamma and tau. 137 le-31
Query= sid| 114835 | lan|dplORF014 Phage dpi ORF| 50961-51974 | 3 (337 letters) gι|1346796|sp|P47492|PRIM_MYCGE DNA PRIMASE >gι | 1361496 | pir | | F64.. 57 2e-07 gij 740008 I rf j I 2004290A primase [Haemophilus influenzae] 51 le-05 gi j 1172619 I spJQ08346 I PRIM_HAEIN DNA PRIMASE >gι 11074033 | pir | |A64.. 51 le-05 gi ] 1709769 j sp j Q04505 j PRIM_LACLA DNA PRIMASE >gι j 1075726 | pir | j JC2... 51 le-05 gi j 639846 | b3 JBAA03516| (D14690) DNA primase [Lactococcus lactis] 51 le-05 Query= si | 114837 | Ian|dplORF016 Phage dpi ORF| 43413-44303 | 3 (296 letters)
L| 1934766 | emb | CAB07986 | (Z93946) N-acetylmuramoyl-L-alanine ami. 661 0.0 L| 113676 |sp|P06653|ALYS_STRPN AUTOLYSIN (N-ACETYLMURAMOYL-L-ALA. 221 4e-57 LJ282326 jpir I |A42935 N-acetylmuramoyl-L-alanme amidase (EC 3.5. 219 3e-56 .| 416618 jsp|P32762|ALYS_BPHB3 LYTIC AMIDASE (N-ACETYLMURAMOYL-L. 212 2e-54 .J285273 jpir I |A42936 N-acetylmuramoyl-L-alanine amidase (EC 3.5. 212 2e-54 . j 127787 jsp I P15057 I LYCA_BPCP1 LYSOZYME (ENDOLYSIN) (MURAMIDASE). 162 4e-39 . j 67761 |pιr I IMUBPCP N-acetylmuramoyl-L-alanine amidase (EC 3.5.. 162 4e-39 .|127789|sp|P19386|LYCA_BPCP9 LYSOZYME (ENDOLYSIN) (MURAMIDASE) 160 le-38 .| 928832 (L44593) ORF259; putative [Lactococcus lactis phage BK. 119 2e-26 J 2511705 | emb |CAA717831 (Y10818) sigA binding prote [Streptoc. 111 9e-24 LJ 4097980 (U72655) surface protein C [Streptococcus pneumoniae] 107 le-22 L 12351768 (U89711) PspA [Streptococcus pneumoniae] 105 4e-22 LJ 2425109 (AF019904) choline binding protein A [Streptococcus p. 104 6e-22 LJ282335 |pιr| |A41971 surface protein pspA precursor - Streptoco. 104 le-21 LJ 2576331 | emb ]CAA05158| (AJ002054) SpsA protein [Streptococcus . 103 2e-21 LJ 2127295 jpirj I S57962 cspC protein - Clostridium acetobutylicum. 85 6e-16 .j 2576333 j emb JCAA05159 I (AJ002055) SpsA protein [Streptococcus . 84 le-15 LJ4106522Jgb|AAD02874.l| (AF097909) excreted protein FibB [Pept. 83 3e-15 J1361406 jpir I |SS7714 cspB protein - Clostridium acetobutylicum. 82 4e-15 tj 1914872 j emb JCAB04758 I (Z82001) PCPA [Streptococcus pneumoniae] 81 9e-15
gi | 3168594 | db;) |BAA28613| (AB012763) SpaA [Erysipelothπx rhusiop 81 le -14 gij 2292750 j emb JCAA64942 j (X95646) homology to orf259 of lactococ 80 3e 14 gij 2935696 (AF032122) putative lysm [Streptococcus thermophilus 80 3e- -14 gij 4586910 I db;) |BAA76540 1| (AB017447) protective antigen SpaA 1 80 3e 14 gij 3540294 (AF057033) lysin [Streptococcus thermophilus bacterio 79 5e- 14
Query= sιd| 114841 | lan|dplORF020 Phage dpi 0RF| 1864-2658 | 1 (264 letters) gi 2633745|emb|CAB13247| (Z99111) similar to coenzyme PQQ synthe 217 5e -56 gi 2808502 j emb JCAA12532J (A 225561) ExsD protein [Sinorhizobium 163 le -39 gi 3861151 j emb I CAA1505lj (A 235272) unknown [Rickettsia prowazekii] 82 6e -15 gi 1652793 j db_ JBAA17712J (D90908) hypothetical protein [Synechoc 76 3e -13 gi 1723815 |sp|P55139|YGCF_ECOLI HYPOTHETICAL 25 0 KD PROTEIN IN 70 2e -11 gi 2984272 (AE000769) hypothetical protein [Aquifex aeolicus] 66 4e 10 91 4155435 (AE001516) putative [Helicobacter pylori J99] 57 le- 07 gi 2127833 |pιr| |C64505 coenzyme PQQ synthesis protein III homolo 55 5e- 07 gi 2622338 (AE000890) coenzyme PQQ synthesis protein III [Methan 54 9e- 07 91 3257042 | db_ |BAA29725| (AP000003) 2S4aa long hypothetical prot 53 2e- 06 gi 2314068 jgb|AAD07976 l| (AE000602) conserved hypothetical prot 52 6e- 06 91 1723816Jsp|P45097|YGCF_HAEIN HYPOTHETICAL PROTEIN HI1189 >gι | 50 2e- 05
Query= sιd| 114842 | lan| dplORF021 Phage dpi ORF| 2504-3295 | 2 (263 letters) gi 127481|sp|P19465|GCHl_BACSU GTP CYCLOHYDROLASE I (GTP-CH-I) > 208 4e -53 gi 3242315|emb|CAA04237| (AJ000685) GTP cyclohydrolase [Streptoc 191 4e -48 2494695Jsp|Q54769|GCHl_SYNP7 GTP CYCLOHYDROLASE I (GTP-CH-I) 189 2e -47 g 255061|bbsjll2832 (S44049) GTP cyclohydrolase I {clone hGCH-1 187 7e -47 gi 4503949 I ref |NP_000152 11 PGCH11 GTP cyclohydrolase 1 (dopa-res 187 7e -47 gi 2113967 j emb JCAB08935 I (Z95557) folE [Mycobacterium tuberculosis] 187 7e -47 g 173024θjsp|P50141|GCHl_CHICK GTP CYCLOHYDROLASE I (GTP-CH-I) 185 3e -46 91 2494696 j sp j QS5759 JGCH1_SYNY3 GTP CYCLOHYDROLASE I (GTP-CH-I) 184 5e -46 gi 121061|sp|P22288|GCHl_RAT GTP CYCLOHYDROLASE I PRECURSOR (GTP 184 6e -46 gi 3183014 I sp I 013774 I GCH1_SCHP0 GTP CYCLOHYDROLASE I (GTP-CH-I) 184 6e -46 g 3097224Jemb|CAA18795| (AL023093) GTP cyclohydrolase I [Mycoba 182 2e •45 91 2494697Jsp|Q19980|GCHl_CAEEL PROBABLE GTP CYCLOHYDROLASE I (G 182 2e -45 gi 462167|sp|Q05915|GCHl_MOUSE GTP CYCLOHYDROLASE I PRECURSOR (G 180 7e -45 gi 1669664|emb|CAA89808| (Z49706) GTP cyclohydrolase I [Dictyost 180 le- -44 gi 2981082 (AF052048) GTP-cyclohydrolase [Ostertagia ostertagi] 178 3e 44 gi 31954|emb|CAA78908| (Z16418) GTP cyclohydrolase I [Homo sapi 177 8e- 44 gi 551344 |bbs 1150280 (S71373) GTP cyclohydrolase I [mice, Peptid 174 5e- ■43 gi 1730247|spjp51601|GCHl_YEAST GTP CYCLOHYDROLASE I (GTP-CH-I) 174 7e- 43 gi 1246912Jemb|CAA87397| (Z47201) GTP cyclohydrolase 1 [Saccharo 172 2e- 42 gi 1730246Jsp|P51595|GCHl_STRPN GTP CYCLOHYDROLASE I (GTP-CH-I) 168 3e- 41 g 2982951 (AE000680) GTP cyclohydrolase I [Aquifex aeolicus] 164 6e- 40
Query= sid | 114843 | Ian |dplORF022 Phage dpi 0RF| 30896-31675 | 2 (259 letters) gi 12347102 (U77367) internalin [Listeria monocytogenes] 55 5e-07 gιJ3123226|sp|P25146|lNLA_LISMO INTERNALIN A PRECURSOR >gι| 48705 52 4e-06 gi 1149674 (M67471) internalin [Listeria monocytogenes] 52 4e-06
Query= sιd| 114850 | lan|dplORF029 Phage dpi ORF| 662-1348 | 2 (228 letters) gi 12650185 (AE001074) succ oglycan biosynthesis regulator (exsB 119 2e -26 gi J386123l|emb|CAA15131 I (AJ235272) unknown [Rickettsia prowazekii] 117 8e -26 gij 2622210 (AE000881) conserved protem [Methanobacterium thermo 108 4e -23 gij 2983380 (AE000709) trans-regulatory protein ExsB [Aquifex aeo 88 6e -17 gij 1001327 I db_ |BAA10814| (D64006) ExsB [Synechocystis sp ] 88 6e -17 gi J2128055 jpir j |B64468 hypothetical protem homolog MJ1347 - Met 83 le •15 gij 4155143 (AE001491) putative [Helicobacter pylori J99] 82 4e -15 gιJ2313760|gb|AAD07701 l| (AE000578) conserved hypothetical prot 80 2e 14 gιJ2120814 |pιr I |S60183 protein ExsB - Rhizobium mel loti >gι|ll4 76 3e- 13 gi j 2633743 j emb JCAB13245 I (Z99111) similar to hypothetical protei 75 5e- 13 gιjll75543Jsp|P44124|YBAX_HAEIN HYPOTHETICAL PROTEIN HI1191 >gι | 74 le 12 gij 2495537 jsp|P77756 I YBAX_ECOLI HYPOTHETICAL 25 5 KD PROTEIN IN 71 5e- 12 gi j 3256471 jdb3 I BAA29154 l| (AP000001) 269aa long hypothetical pr 67 le- 10 gι| 2921156 (AF022216) aluminum resistance prote [Arthrobacter 54 le- 06
Query= sid | 114855 | Ian | dplORF034 Phage dpi ORF| 131-652 | 2 (173 letters) gι|2633746|emb|CAB13248| (Z99111) similar to hypothetical protei 220 4e-57
L 14155926 (AE001554) putative [Helicobacter pylori J99] 162 le -39
.j 2314588 |gb|AAD08456.1| (AE000642) conserved hypothetical prot ... 161 3e -39
.j 2983458 (AE000714) hypothetical protein [Aquifex aeolicus] 103 9e -22
. j 1006604 I dbj |BAA10757| (D64005) hypothetical protein [Synechoc... 87 6e -17
.j 2967529 (U11045) unknown [Buchnera aphidicola] 79 2e -14
. J2495654|sp|Q46920|YQCD_ECOLI HYPOTHETICAL 32.6 KD PROTEIN IN ... 69 2e -11
.j 1175604 j spj P44153 JYQCD_HAEIN HYPOTHETICAL PROTEIN HI1291 >gi | ... 63 le -09
. J3860642 jemb|CAA14543| (A 235270) unknown [Rickettsia prowazekii] 56 le- -07
Query= sid| 114857 | lan|dplORF036 Phage dpi ORF| 48808-49362 | 1 (184 letters) gi 11353529 (U38906) ORF12 [Bacteriophage rlt] 53 le-06
Query= sid| 114859 | lan| dplORF038 Phage dpi ORF| 1350-187113 (173 letters) gi|1175542|sp|P44123|YB90_HAEIN HYPOTHETICAL PROTEIN HI1190 >gi | ... 100 6e-21 gij 2982977 (AE000681) hypothetical protein [Aquifex aeolicus] 67 7e-ll gi J3860744 | emb | CAA14645 | (AJ235270) unknown [Rickettsia prowazekii] 65 3e-10 gi]2650193 (AE001074) conserved hypothetical protein [Archaeoglo... 58 4e-08 gi I 3258383 | dbj |BAA31066.l| (AP000007) 157aa long hypothetical pr... 55 2e-07 gij 1001713 I dbj JBAA10550| (D64004) hypothetical protein [Synechoc... 50 8e-06 gij 4155434 (AE001516) putative [Helicobacter pylori J99] 50 le-05
Query= sid| 114860 | lan| plORF039 Phage dpi ORF| 3306-3803 | 3 (165 letters) gi|l922884|emb|CAA68244 I (X99978) ORF7 ; hydophobic protein [Lact... 64 5e-10
Query= sid| 114862 | lan|dplORF041 Phage dpi ORF| 8208-8699 | 3 (163 letters) gi 12522313 (AF012906) dUTPase homolog [Bacillus subtilis] >gi|26. 108 2e-23 giJ2634150|emb|CAB13650| (Z99113) similar to deoxyuridine 5'-tri. 108 3e-23 gij 3913546 jsp I 054134 I DUT_STRCO DEOXYURIDINE 5 ' -TRIPHOSPHATE NUCL. 56 2e-07 gij 3913542 j spj 048500 I DUT_BPT5 DEOXYURIDINE 5 ' -TRIPHOSPHATE NUCLE . 52 3e-06 gij 3913548 j spj 068992 I DUT_CHLTE DEOXYURIDINE 5 ' -TRIPHOSPHATE NUCL. 50 le-05
Query= sid| 114867 | lan|dplORF046 Phage dpi ORF| 42774-43202 | 3 (142 letters) gi|1934764 | emb | CAB07984 | (Z93946) hypothetical protein [bacterio... 287 2e-77
Query= sid| 114901 | lan|dplORF080 Phage dpi ORF| 42490-42759 | 1 (89 letters) gi|1934763|emb|CAB07983] (Z93946) hypothetical protein [bacterio.., 147 le-35
Query= sid 1114912 | Ian | plORF091 Phage dpi 0RF| 43189-4341311 (74 letters) gi|1934765|emb|CAB07985| (Z93946) holin [bacteriophage Dp-1] 63 2e-10
Table 32
Sequence of Dpi published by Sheehan and al.. 4731 nucleotides.
1 tttaaatttt ttgacaaagt taattcaaat tgtaccgctg aagcaatttt ccatgtattc actcaaagtt
71 gttcagtgtg gctcaatcat attaaaatcg aacttggtaa tatctctact ccttttagtg aagcagagga
141 agaccttaaa tatcgaattg actcaaaagc cgatcaaaag ctaactaacc aacagttgac ggcactcacg
211 gaaaaggctc aactacatga cgcagaactg aaagctaagg ctacaatgga gcagttaagt aacttagaaa
281 aggcttatga aggtagaatg aaagctaatg aagaagctat caacaaatcg gaacccgacc taatcttagc
351 ggcaagtcga attgaagcta ctatccaaga acttggcggg ctacgggaac tgaagaagtt cgtcgacagt 421 tgcatgagct cttctaatca aggtctaatt atcggtaaga acgacggtag ctctaccatt aaggtatcaa
491 gtgaccgaat ttctatgttc tccgcaggga atgaagttat gtaccttacg caagggttca ttcacatcga 561 taacgggatc tttacccaat ccattcaagt cggccgattt agaacggaac aatactcgtt taatccagac 631 atgaacgtga ttcggtatgt aggataagga gaataacatg acaaaattta tcaactcata cggccctctt 701 cacttgaacc tttacgtcga acaagttagt caggacgtaa cgaacaactc ctcgcgagtt agttggcgag 771 ctactgtcga ccgcgatgga gcttatcgaa cgtggactta tggaaatatt agtaaccttt ccgtatggtt 841 aaatggttca agtgttcata gcagtcaccc agactacgac acgtccggcg aagaggtaac gctcgcaagt 911 ggagaagtga ctgttcctca caatagtgac gggacaaaga caatgtccgt ttgggcttcg tttgacccta 981 ataacggcgt tcacggaaat atcactatct ctactaatta cactttagac agtattccaa ggtctacaca 1051 gatttctagt tttgagggaa atcgaaatct aggatcttta catacggtta tctttaaccg aaaagtgaac 1121 tcttttacgc atcaagtttg gtaccgagtt ttcggtagcg actggataga tttaggtaag aaccatacta 1191 ctagcgtatc ctttacgccg tcactggact tagcaaggta cttacctaaa tcaagttccg gaacaatgga 1261 catctgtatt cgaacctata acggaactac gcaaattggt agtgacgtct attcaaacgg atggaggttc 1331 aacatccccg attcagtacg tcctactttt tcgggcattt ctttagtaga cacgacttca gcggttcgac 1401 agattttaac agggaacaac ttcctccaaa tcatgtcgaa cattcaagtc aacttcaaca atgcttccgg 1471 cgcttacgga tccactatcc aagcatttca cgctgagctc gtaggtaaaa accaagctat caacgaaaac 1541 ggcggcaaat tgggtatgat gaactttaat ggctccgcta ccgtaagagc atgggttaca gacacgcgag 1611 gaaaacaatc gaacgtccaa gacgtatcta tcaatgttat agaatactat ggaccgtcta tcaatttctc 1681 cgttcaacgt actcgtcaaa atcctgcaat tatccaagct cttcgaaatg ctaaggtcgc acctataacg 1751 gtaggaggtc aacagaaaaa catcatgcaa attaccttct ccgtggcgcc gttgaacact actaatttca 1821 cagaagatag aggttcggcg tcagggacgt tcactactat ttccctactg actaactcgt ccgcgaactt 1891 agctggtaac tacgggccgg acaagtctta catagttaag gctaaaatcc aagacaggtt cacttcgact 1961 gaatttagtg ctacggtacc taccgaatca gtagttctta actatgacaa ggacggtcga cttggagttg 2031 gtaaggttgt agaacaaggg aaggcagggt caattgatgc agcaggtgat atatatgctg gaggtcgaca 2101 agttcaacag tttcagctca ctgataataa tggagcattg aacaggggtc aatataacga tgttggaata 2171 agcgtgaaac agagtttaca tggcgaagta acaaatacga ggacaaccct acgggaactc gaggtgaatg 2241 gggactattt caaaatttct ggttagatag ctggaaaatg gttcaatcct tcattacaat gtcaggaaga 2311 atgttcatca ggacagcgaa cgatggaaac agctggagac ctaacaagtg gaaagaggtt ctatttaagc 2381 aagacttcga acagaataat tggcagaaac ttgttcttca aagtgggtgg aaccatcact caacctatgg 2451 cgacgcattc tattcgaaaa ctcttgacgg catagtatat ttgagaggaa atgtgcataa aggacttatc 2521 gacaaagagg ctactattgc agtacttcct gaaggattta gaccgaaagt ttcaatgtat cttcaggctc 2591 tcaataactc atatggaaat gccattctat gtatatacac tgacggaaga cttgtggtga aatcgaatgt 2661 agataattct tggttaaatt tagacaatgt ctcatttcgt atttaatttg agctgaaatc atgttataat 2731 attttttaga aaggaggtga gaactatgtt gaaccttaca aaatcgcgcc aaattgtggc agagttcact 2801 attggacaag gagctgaaaa gaaacttgtc aaaacaacga ttgtgaacat tgatgcaaac gcagtatcaa 2871 ccgtctctga aactcttcat gacccagact tgtatgctgc gaaccgtcga gaacttcgag ctgacgagca 2941 aaaacttcgc gaaactcgtt acgcaatcga agatgaaatt aatagctgga gcgggggaaa aaagggggag 3011 cccggctcta acaggctgaa taaggaggcg tcaatctatg ccaatgtggc taaacgacac cgcagtcttg 3081 acgacgatta ttacagcgtg cagcggagtg cttactgtcc tactaaataa gttattcgaa tggaaatcga 3151 ataaagccaa gagcgtttta gaggatatct ctacaactct tagcactctt aaacagcagg tcgacgggat 3221 tgaccaaacg acagtagcaa tcaatcacca aaatgacgtc attcaagacg gaactagaaa aattcaacgt 3291 taccgtcttt atcacgactt aaaaagggaa gtgataacag gctatacaac tctcgaccat tttagagagc 3361 tctctatttt attcgaaagt tataagaacc ttggcggaaa tggtgaagtt gaagccttgt atgaaaaata 3431 caagaaatta ccaattaggg aggaagattt agatgaaact atctaacgaa caatatgacg tagcaaagaa 3501 cgtggtaacc gtagtcgttc cagcagcgat tgcactaatt acaggtcttg gagcgttgta tcaatttgac 3571 actactgcta tcacaggaac cattgcactt cttgcaactt ttgcaggtac tgttctagga gtttctagcc 3641 gaaactacca aaaggaacaa gaagctcaaa acaatgaggt ggaataatgg gagtcgatat tgaaaaaggc 3711 gttgcgtgga tgcaggcccg aaagggtcga gtatcttata gcatggactt tcgagacggt cctgatagct 3781 atgactgctc aagttctatg tactatgctc tccgctcagc cggagcttca agtgctggat gggcagtcaa 3851 tactgagtac atgcacgcat ggcttattga aaacggttat gaactaatta gtgaaaatgc tccgtgggat 3921 gctaaacgag gcgacatctt catctgggga cgcaaaggtg ctagcgcagg cgctggaggt catacaggga 3991 tgttcattga cagtgataac atcattcact gcaactacgc ctacgacgga atttccgtca acgaccacga 4061 tgagcgttgg tactatgcag gtcaacctta ctactacgtc tatcgcttga ctaacgcaaa tgctcaaccg 4131 gctgagaaga aacttggctg gcagaaagat gctactggtt tctggtacgc tcgagcaaac ggaacttatc 4201 caaaagatga gttcgagtat atcgaagaaa acaagtcttg gttctacttt gacgaccaag gctacatgct
4271 cgctgagaaa tggttgaaac atactgatgg aaattggtat tggttcgacc gtgacggata catggctacg ... - -
4341 tcatggaaac ggattggcga gtcatggtac tacttcaatc gcgatggttc aatggtaacc ggttggattϊ~
4411 agtattacga taattggtat tattgtgatg ctaccaacgg cgacatgaaa tcgaatgcgt ttatccgtta
4481 taacgacggc tggtatctac tattaccgga cggacgtctg gcagataaac ctcaattcac cgtagagccg
4551 gacgggctca ttactgctaa agtttaaaat atagagagga ggaagctctt ttcttaatat tgtttctctt
4621 aatcccgcaa ggtttcgacc ctgcggggtt tatgtgtcgt gaattactct atttacttat tcgaagattt
4691 caattataat taaataatca acgagattca taattggagg aatg
Table 33
Streptococcus accession numbers gi|5776553|gb|AF026471.2|AF026471 [5776553] gi|5231200|gb|AF157824.1|AF157824 [5231200] gi|5410470|gb|AF139890.1|AF139890 [5410470] gi|5231197|gb|AF157823.1|AF157823 [5231197] gi|5410468|gb|AF139889.1|AF139889 [5410468] gi|5231194|gb|AFl 57822.1|AF 157822 [5231194] gi|5410466|gb|AF139888.1|AF139888 [5410466] gi|5231191 |gb|AF 157821.11 AF 157821 [5231191 ] gi|5410464|gb|AF139887.1|AF139887 [5410464] gi|5231188|gb|AFl 57820.11 AF 157820 [5231188] gi|5410462|gb|AF139886.1|AF139886 [5410462] gi|5231185|gb|AF157819.1|AF157819 [5231185] gi|5410460|gb|AF139885.1|AF139885 [5410460] gi|5231182|gb|AF157818.1|AF157818 [5231182] gi|5410458|gb|AF139884.1|AF139884 [5410458] gi|5231179|gb|AF157817.1|AF157817 [5231179] gi|5410456|gb|AF139883.1|AF139883 [5410456] gi|4336851|gb|AF106138.1|AF106138 [4336851] gi|3093394|emb|AJ005697.1|SPN5697 [3093394] gi|4336848|gb|AF106137.1|AF106137 [4336848] gi|5759208|gb|AF171873.1|AF171873 [5759208] gi|4336845|gb|AF106136.1|AF106136 [4336845] gi|5758311|gb|AF162664.1|AF162664 [5758311] gi|4336842|gb|AF106135.1|AF106135 [4336842] gij5739313|gb|AF161701.1|AF161701 [5739313] gi|4336839|gb|AF106134.1|AF106134 [4336839] gi|5739310|gb|AF161700.1|AF161700 [5739310] gi|4336836jgb|AF106133.1|AF106133 [4336836] gi|5726354|gb|AF159448.1|AF159448 [5726354] gi|4336833|gb|AF106132.1|AF106132 [4336833] gi|5726290|gb|AF127143.1|AF127143 [5726290] gi|3907597|gb|AF094575.1|AF094575 [3907597] gi|5712666|gb|AF140784.1|AF140784 [5712666] gi|5030425|gb|AF061748.2|AF061748 [5030425] gi|4218525|emb|AJ009639.1|SPAJ9639 [4218525] gi|4902881|emb|AJ239004.1|SPN239004 [4902881] gi|5616524|gb|AF169483.1|AF169483 [5616524] gi|5579395|gb| AF 162656.1 |AF 162656 [5579395] gi|5001710|gb|AF112358.1|AFl 12358 [5001710] gi|5579393|gb|AF162655.1|AF162655 [5579393] gi|5001690|gb|AF106539.1|AF106539 [5001690] gi|4973271|gb|AF144420.1|AF144420 [4973271] gi|5578890|emb|AJ131985.1|SPN131985 [5578890] gi|4973269|gb|AF144419.1|AF144419 [4973269] gi|5566442|gb|AF167442.1|AF167442 [5566442] gi|4973267|gb|AF144418.1|AF144418 [4973267] gi|5459332|emb|AJ243540.1|EVE243540 gi|4928190|gb|AF129757.1|AF129757 [4928190] [5459332] gi|4927743 |gb| AF 126061.11 AF 126061 [4927743] gi|5305398|gb|AF072811.1 |AF072811 [5305398] gi|4927742|gb| AF 126060.11 AF 126060 [4927742] gi|5295921|emb|AJ242698.1|SPN242698 [5295921] gi|4927741 |gb| AF 126059.11 AF 126059 [4927741 ] gi|5295920|emb|AJ242697.1|SPN242697 gi|4495247|emb|AJ240675.1|SPN240675 [4495247] [5295920] gi|5295919|emb|AJ242696.1 |SPN242696 gi|4495245|emb|AJ240670.1|SPN240670 [5295919] [4495245] gi|5295918|emb| AJ242695.1 |SPN242695 gi|4495243|emb|AJ240669.1|SPN240669 [5295918] [4495243] gi|4583522|gb|AF140356.1 |AF 140356 [4583522] gi|4495241|emb|AJ240668.1|SPN240668 [4495241] gi|5231206|gb| AF 157826.11 AF 157826 [5231206] gi|4495239|emb|AJ240667.1|SPN240667 gi|5231203|gb|AF157825.1|AF157825 [5231203] [4495239]
gi|4495237|emb|AJ240666.1|SPN240666 gi|4495189|emb|AJ240640.1 |SPN240640 [4495237] [4495189] gi|4495235|emb|AJ240665.1|SPN240665 gi|4495187|emb|AJ240639.1 |SPN240639 [4495235] [4495187] gi|4495233|emb|AJ240664.1|SPN240664 gij4495185|emb|AJ240638.1|SPN240638 [4495233] [4495185] gi|4495231 |emb| AJ240663.1 |SPN240663 gi|4495183|emb|AJ240637.1 |SPN240637 [4495231] [4495183] gi|4495229|emb| AJ240662.1 |SPN240662 gi|4495181|emb|AJ240636.1|SPN240636 [4495229] [4495181] gi|4495227|emb|AJ240661.1 |SPN240661 gi|4495179|emb|AJ240635.1|SPN240635 [4495227] [4495179] gi|4495225|emb|AJ240660.1 |SPN240660 gi|4495177|emb|AJ240634.1 |SPN240634 [4495225] [4495177] gi|4495223|emb|AJ240659.1|SPN240659 gi|4495175|emb|AJ240633.1|SPN240633 [4495223] [4495175] gi|4495221 |emb|AJ240658.1 |SPN240658 gi|4495173 |emb|AJ240630.1 |SPN240630 [4495221] [4495173] gi|4495219|emb|AJ240657.1|SPN240657 gi|4495171|emb|AJ240629.1|SPN240629 [4495219] [4495171] gi|4495217|emb|AJ240656.1|SPN240656 gi|4495169|emb|AJ240628.1|SPN240628 [4495217] [4495169] gi|4495215|emb|AJ240655.1|SPN240655 gi|4495167|emb|AJ240627.1 |SPN240627 [4495215] [4495167] gi|4495213|emb|AJ240654.1 |SPN240654 gi|4495165 |emb| AJ240626.1 |SPN240626 [4495213] [4495165] gi|4495211 |emb| AJ240653.1 |SPN240653 gi|4495163|emb|AJ240625.1 |SPN240625 [4495211] [4495163] gi|4495209|emb|AJ240652.1|SPN240652 gi|4495161 |emb|AJ240624.1 |SPN240624 [4495209] [4495161] gi|4495207|emb|AJ240651.1 |SPN240651 gi|4495159|emb|AJ240623.1 |SPN240623 [4495207] [4495159] gi|4495205|emb|AJ240650.1|SPN240650 gi|4495157|emb|AJ240622.1|SPN240622 [4495205] [4495157] gi|4495203 |emb| AJ240649.1 |SPN240649 gi|4495155|emb|AJ240621.1|SPN240621 [4495203] [4495155] gi|4495201 |emb|AJ240648.1 |SPN240648 gi|4495153 |emb| AJ240620.1 |SPN240620 [4495201] [4495153] gi|4495199|emb|AJ240647.1|SPN240647 gi|4495151|emb|AJ240619.1|SPN240619 [4495199] [4495151] gi|4495197|emb| AJ240644.1 |SPN240644 gi|4495149|emb|AJ240616.1|SPN240616 [4495197] [4495149] gi|4495195|emb|AJ240643.1|SPN240643 gi|4495147|emb|AJ240615.1|SPN240615 [4495195] [4495147] gi|4495193 |emb|AJ240642.1 |SPN240642 gi|4495145|emb|AJ240614.1|SPN240614 [4495193] [4495145] gi|4495191 |emb|AJ240641.1 |SPN240641 gi|4495143|emb|AJ240613.1|SPN240613 [4495191] [4495143]
gi|4495141jemb|AJ240612.1|SPN240612 gi|4538797|emb|AJ240781.1|SPN240781 [4495141] [4538797] gi|4495139|emb| A J240611.1 |SPN240611 gi|4538794|emb|AJ240780.1|SPN240780 [4495139] [4538794] gi|4495137|emb| A J240610.1 |SPN240610 gi|4538791|emb|AJ240779.1|SPN240779 [4495137] [4538791] gi|4495135|emb|AJ240609.1 JSPN240609 gi|4538788|emb|AJ240778.1|SPN240778 [4495135] [4538788] gi|4495133 |embj A J240608.1 |SPN240608 gi|4538785|emb|AJ240777.1|SPN240777 [4495133] [4538785] gi|4495131 |emb| AJ240607.1 [SPN240607 gi|4538782|emb|AJ240776.1|SPN240776 [4495131] [4538782] gi|4495129|emb|AJ240606.1|SPN240606 gi|4538779|emb|AJ240775.1|SPN240775 [4495129] [4538779] gi|4883698|gb|AF079807.1|AF079807 [4883698] gi|4538776|emb|AJ240774.1|SPN240774 [4538776] gi|4838562|gb|AF145055.1|AF145055 [4838562] gi|4063727|gb|L29324.1|STRINTE [4063727] gi|4538773|emb|AJ240773.1|SPN240773 [4538773] gi|3093401|emb|AJ005619.1|SPAJ5619 [3093401] gi|4538770|emb|AJ240772.1|SPN240772 gij4103889|gb|AF029368.1|AF029368 [4103889] [4538770] gi|2897689|dbj|D63805.1 |D63805 [2897689] gi|4538767|emb|AJ240771.1 |SPN240771 [4538767] gi|4566771|gb|AFl 17741.1|AF117741 [4566771] gi|4538764|emb|AJ240770.1|SPN240770 gi|4566768|gb|AF 117740.11 AF 117740 [4566768] [4538764] gi|4538836|emb|AJ240793.1|SPN240793 gi|4538761|emb|AJ240769.1|SPN240769 [4538836] [4538761] gi|4538832|emb|AJ240792.1|SPN240792 gi|4538758|emb|AJ240768.1|SPN240768 [4538832] [4538758] gi|4538828|emb|AJ240791.1|SPN240791 gi|4538755|emb|AJ240767.1|SPN240767 [4538828] [4538755] gi|4538824|emb|AJ240790.1|SPN240790 gi|4538752|emb|AJ240766.1 JSPN240766 [4538824] [4538752] gi|4538821|emb|AJ240789.1|SPN240789 gi|4538749|emb|AJ240765.1|SPN240765 [4538821] [4538749] gi|4538818|emb|AJ240788.1|SPN240788 gi|4538746|emb|AJ240761.1 |SPN240761 [4538818] [4538746] gi|4538815|emb|AJ240787.1|SPN240787 gi|4538743|emb|AJ240760.1|SPN240760 [4538815] [4538743] gi|4538812|emb|AJ240786.1|SPN240786 gi|4538740|emb|AJ240759.1|SPN240759 [4538812] [4538740] gi|4538809|emb|AJ240785.1|SPN240785 gi|4538737|emb|AJ240758.1|SPN240758 [4538809] [4538737] gi|4538806|emb|AJ240784.1 |SPN240784 gi|4538734|emb|AJ240757.1|SPN240757 [4538806] [4538734] gi|4538803|emb|AJ240783.1[SPN240783 gi|4538731|emb|AJ240756.1|SPN240756 [4538803] [4538731] gi|4538800|emb|AJ240782.1|SPN240782 gi|4538728|emb|AJ240755.1|SPN240755 [4538800] [4538728]
gi|4538725|emb|AJ240754.1|SPN240754 gi|4519233|dbj|AB011207.1|AB011207 [4519233] [4538725] gi|4519231|dbj|AB011206.1|AB011206 [4519231] gi|4538722|emb|AJ240753.1|SPN240753 gi|4519229|dbj|AB011205.1|AB011205 [4519229] [4538722] gi|4538719|emb|AJ240752.1|SPN240752 gi|4519227|dbj|AB011204.1 |AB011204 [4519227] [4538719] gi|4519225|dbjjAB011203.1 |AB011203 [4519225] gi|4538716|emb| AJ240751.1 |SPN240751 gi|4519223 jdbj | ABO 11202.11 ABO 11202 [4519223] [4538716] gi|4519221|dbj|AB011201.1|AB011201 [4519221] gi|4538713|emb|AJ240750.1 |SPN240750 [4538713] gi|4519219|dbj|AB011200.1|AB011200 [4519219] gi|4538710|emb| AJ240749.1 |SPN240749 gi|4519217|dbj|AB011199.1|AB011199 [4519217] [4538710] gi|4519215|dbj|AB011198.1|AB011198 [4519215] gi|4538707|emb|AJ240748.1|SPN240748 gi|4495127|emb|AJ240605.1 |SPN240605 [4538707] [4495127] gi|4538704|emb|AJ240747.1|SPN240747 gi|4468031|emb|AJ132957.1|SPN132957 [4538704] [4468031] gi|4538701 |emb|AJ240746.1 |SPN240746 gi|4468029|emb|AJ132956.1|SPN132956 [4538701] [4468029] gi|4538698|emb|AJ240745.1|SPN240745 gi|4218532|emb|AJ010312.1|SPN010312 [4538698] [4218532] gi|4538695|emb|AJ240744.1|SPN240744 gi|4456852|emb|AJ236792.1|SPN236792 [4538695] [4456852] gi|4538692|emb|AJ240743.1|SPN240743 gi|4456850|emb|AJ236791.1|SPN236791 [4538692] [4456850] gi|4538689|emb|AJ240742.1|SPN240742 g_|4456848|emb|AJ236790.1|SPN236790 [4538689] [4456848] gi|4538686|emb| AJ240741.1 |SPN240741 gi|4456846|emb|AJ236789.1|SPN236789 [4538686] [4456846] gi|4538683[emb|AJ240740.1|SPN240740 gi|3550644|emb|AJ006987.1|SPAJ6987 [3550644] [4538683] gi|3550625|emb|AJ006986.1 |SPAJ6986 [3550625] gi|4538680|emb|AJ240739.1|SPN240739 [4538680] gi|4416518|gb|AF014458.2|AF014458 [4416518] gi|4538677|emb|AJ240738.1|SPN240738 gi|4406260|gb|AF 105116.1 |AF 105116 [4406260] [4538677] gi|4406257|gbj AF 105115.11 AF 105115 [4406257] gi|4530444 |gb| AF 118229.11 AF 118229 [4530444] gi|4406254|gb|AF105114.1|AF105114 [4406254] gi|4519253|dbj|AB015852.1|AB015852 [4519253] gi|4406246|gb|AF 105113.11 AF 105113 [4406246] gi|4519251|dbj|AB015851.1|AB015851 [4519251] gi|4406243|gb|AF105112.1|AF105112 [4406243] gi|4519249|dbj|AB015850.1|AB015850 [4519249] gi|4138533|emb|AJ005815.1|SPN5815 [4138533] gi|4519247|dbj|AB015849.1|AB015849 [4519247] gi|3821726|emb|AJ232433.1|SPN232433 gi|4519245|dbj|AB015848.1|AB015848 [4519245] [3821726] gi|4519243|dbj|AB015847.1|AB015847 [4519243] gi|3821724|emb|AJ232432.1|SPN232432 [3821724] gi|4519241|dbj|AB015846.1|AB015846 [4519241] gi|3821722|emb|AJ23243U|SPN232434— " gi|4519239|dbj|AB011210.1JAB011210 [4519239] [3821722] gi|4519237|dbj|AB011209.1|AB011209 [4519237] gi|3821720|emb|AJ232430.1 |SPN232430 gi|4519235|dbj|AB011208.1|AB011208 [4519235] [3821720]
gij3821718|emb|AJ232429.1|SPN232429 gi|3821670|emb|AJ232405.1|SPN232405 [3821718] [3821670] gi|3821716|emb|AJ232428.1|SPN232428 gi|3821668|emb|AJ232404.1 |SPN232404 [3821716] [3821668] gi|3821714|emb|AJ232427.1|SPN232427 gi|3821666|emb|AJ232403.1 ISPN232403 [3821714] [3821666] gi|3821712|emb|AJ232426.1|SPN232426 gi|3821664|emb|AJ232402.1|SPN232402 [3821712] [3821664] gi|3821710|emb|AJ232425.1|SPN232425 gi|3821662|emb|AJ232401.1 |SPN232401 [3821710] [3821662] gi|3821708|emb|AJ232424.1|SPN232424 gi|3821660|emb|AJ232399.1|SPN232399 [3821708] [3821660] gi|3821706|emb|AJ232423.1|SPN232423 gi|3821658|emb|AJ232398.1 |SPN232398 [3821706] [3821658] gi|3821704|emb|AJ232422.1|SPN232422 gi|3821656|emb|AJ232397.1 |SPN232397 [3821704] [3821656] gi|3821702|emb|AJ232421.1 |SPN232421 gi|3821654|embjAJ232396.1|SPN232396 [3821702] [3821654] gi|3821700|emb|AJ232420.1|SPN232420 gi|3821652|emb|AJ232395.1|SPN232395 [3821700] [3821652] gi|3821698|emb|AJ232419.1|SPN232419 gi|3821650|emb|AJ232394.1 |SPN232394 [3821698] [3821650] gi|3821696|emb|AJ232418.1|SPN232418 gi|3821648|emb|AJ232393.1 |SPN232393 [3821696] [3821648] gi|3821694|emb|AJ232417.1|SPN232417 gi|3821646|emb|AJ232392.1|SPN232392 [3821694] [3821646] gi|3821692|emb|AJ232416.1|SPN232416 gi|3821644|emb|AJ232391.1|SPN232391 [3821692] [3821644] gi|3821690|emb|AJ232415.1|SPN232415 gi|3821642|emb|AJ232390.1|SPN232390 [3821690] [3821642] gi|3821688|emb|AJ232414.1|SPN232414 gi|3821640|emb|AJ232389.1|SPN232389 [3821688] [3821640] gi|3821686|emb|AJ232413.1|SPN232413 gi|3821638|emb|AJ232388.1|SPN232388 [3821686] [3821638] gi|3821684|emb|AJ232412.1|SPN232412 gi|3821636|emb|AJ232387.1 |SPN232387 [3821684] [3821636] gi|3821682|emb|AJ232411.1|SPN232411 gi|3821634!emb|AJ232386.1|SPN232386 [3821682] [3821634] gi|3821680|emb|AJ232410.1|SPN232410 gi|3821632|emb|AJ232385.1|SPN232385 [3821680] [3821632] gi|3821678|emb|AJ232409.1|SPN232409 gi|3821630|emb|AJ232384.1|SPN232384 [3821678] [3821630] gij3821676|emb|AJ232408.1|SPN232408 gij3821628|emb|AJ232383.1|SPN232383 [3821676] [3821628] gi|3821674|emb| AJ232407.1 |SPN232407 gi|3821626|emb|AJ232382.1|SPN232382 [3821674] [3821626] gi|3821672|emb|AJ232406.1|SPN232406 gi|3821624|emb|AJ232381.1|SPN232381 [3821672] [3821624]
gi|3821622|emb|AJ232380.1 SPN232380 gi|3821576|emb|AJ232356.1|SPN232356 [3821622] [3821576] gi|3821620|emb|AJ232379.1 SPN232379 gi|3821574|emb|AJ232355.1|SPN232355 [3821620] [3821574] gi|3821618|emb|AJ232378.1 SPN232378 gi|3821572|emb|AJ232353.1|SPN232353 [3821618] [3821572] gi|3821616|emb|AJ232377.1 SPN232377 gij3821570|emb|AJ232352.1ISPN232352 [3821616] [3821570] gi|3821614|emb|AJ232376.1 SPN232376 gi|3821568|emb|AJ232351.1|SPN232351 [3821614] [3821568] gi|3821612|emb|AJ232375.1 SPN232375 gi|3821566|emb|AJ232350.1|SPN232350 [3821612] [3821566] gi|3821610|emb|AJ232373.1 SPN232373 gi|3821564|emb|AJ232349.1|SPN232349 [3821610] [3821564] gi|3821608|emb|AJ232372.1 SPN232372 gi|3821562|emb|AJ232348.1|SPN232348 [3821608] [3821562] gi|3821606|emb|AJ232371.1 SPN232371 gi|3821560|emb|AJ232347.1|SPN232347 [3821606] [3821560] gi|3821604|emb|AJ232370.1 SPN232370 gi|3821558|emb|AJ232346.1|SPN232346 [3821604] [3821558] gi|3821602|emb|AJ232369.1 SPN232369 gi|3821556|emb|AJ232345.1|SPN232345 [3821602] [3821556] gi|3821600|emb|AJ232368.1 SPN232368 gi|3821554|emb|AJ232344.1|SPN232344 [3821600] [3821554] gi|3821598|emb|AJ232367.1| SPN232367 gi|3821552|emb|AJ232343.1|SPN232343 [3821598] [3821552] gi|3821596|emb|AJ232366.1 SPN232366 gi|3821550|emb|AJ232342.1|SPN232342 [3821596] [3821550] gi|3821594|emb|AJ232365.1 SPN232365 gi|3821548|emb|AJ232341.1|SPN232341 [3821594] [3821548] gi|3820454|emb|AJ007367.1 SPN7367 [3820454] gi|3821546|emb| A J232340.1 |SPN232340 [3821546] gi|3821592|emb|AJ232364.1 SPN232364 [3821592] gi|3821544|emb|AJ232339.1|SPN232339 [3821544] gi|3821590|emb|AJ232363.1 SPN232363 [3821590] gi|3821542|emb|AJ232338.1|SPN232338 [3821542] gi|3821588|emb|AJ232362.1 SPN232362 [3821588] gi|3821540|emb|AJ232337.1|SPN232337 [3821540] gi|3821586|emb|AJ232361.1 SPN232361 [3821586] gi|3821538|emb|AJ232336.1|SPN232336 [3821538] gi|3821584|emb|AJ232360.1l SPN232360 [3821584] gi|3821536|emb|AJ232335.1|SPN232335 [3821536] gi|3821582|emb|AJ232359.1 SPN232359 [3821582] gi|3821534|emb|AJ232334.1|SPN232334 [3821534] gi|3821580|emb|AJ232358.1 SPN232358 [3821580] gi|3821532|emb|AJ232333.1|SPN2 2333 [3821532] gi|3821578|emb|AJ232357.1 SPN232357 [3821578] gi|3821530|emb|AJ232332.1|SPN232332 [3821530]
gi|3821528|emblAJ232331.1 JSPN232331 gi|3821480|emb|AJ232306.1ISPN232306 [3821528] [3821480] gi|3821526|emblAJ232330.1jSPN232330 gi|3821478|emb|AJ232305.1 |SPN232305 [3821526] [3821478] gi|3821524|emb|AJ232329.1|SPN232329 gi|3821476|emb|AJ232304.1|SPN232304 [3821524] [3821476] gij3821522|emblAJ232328.1|SPN232328 gi|3821474|emb|AJ232303.1|SPN232303 [3821522] [3821474] gi|3821520|emb!AJ232327.1|SPN232327 gi|3821472|emb|AJ232302.1|SPN232302 [3821520] [3821472] gi|3821518|emb|AJ232326.1|SPN232326 gi|3821470|emb|AJ232301.1|SPN232301 [3821518] [3821470] gi|3821516|emb|AJ232325.1 |SPN232325 gi|3821468|emb|AJ232300.1|SPN232300 [3821516] [3821468] gi|3821514|emb|AJ232324.1|SPN232324 gi|3821466|emb|AJ232299.1|SPN232299 [3821514] [3821466] gi|3821512|emb|AJ232322.1|SPN232322 gij3821464|emb|AJ232298.1|SPN232298 [3821512] [3821464] gi|3821510|emb|AJ232321.1|SPN232321 gi|3821462|emb|AJ232297.1|SPN232297 [3821510] [3821462] gi|3821508|emb|AJ232320.1 [SPN232320 gi|3821460|emb|AJ232295.1|SPN232295 [3821508] [3821460] gi|3821506|emb|AJ232319.1|SPN232319 gi|3821458|emb|AJ232294.1|SPN232294 [3821506] [3821458] gi|3821504|emb|AJ232318.1|SPN232318 gi|3821456|emb|AJ232293.1|SPN232293 [3821504] [3821456] gi|3821502|emb|AJ232317.1|SPN232317 gi|3821454|emb|AJ232292.1|SPN232292 [3821502] [3821454] gi|3821500|emb| AJ232316.1 [SPN232316 gi|3821452|emb|AJ232291.1 |SPN232291 [3821500] [3821452] gi|3821498|emb|AJ232315.1|SPN232315 gi|3821450|emb|AJ232290.1|SPN232290 [3821498] [3821450] gi|3821496]emb|AJ232314.1|SPN232314 gi|3821448|emb|AJ232289.1|SPN232289 [3821496] [3821448] gi|3821494|emb|AJ232313.1|SPN232313 gi|3821446|emb|AJ232288.1|SPN232288 [3821494] [3821446] gi|3821492|emb|AJ232312.1|SPN232312 gi|3821444|emb|AJ232287.1|SPN232287 [3821492] [3821444] gi|3821490|emb|AJ232311.1|SPN232311 gi|3821442|emb|AJ232286.1jSPN232286 [3821490] [3821442] gi|3821488|emb|AJ232310.1|SPN232310 gi|3821440|emb|AJ232285.1|SPN232285 [3821488] [3821440] gi|3821486|emb|AJ232309.1|SPN232309 gi|3821438|emb|AJ232284.1|SPN232284 [3821486] [3821438] gi|3821484|emb|AJ232308.1|SPN232308 gi|3821436|emb|AJ232283.1|SPN232283 [3821484] [3821436] gi|3821482|emb|AJ232307.1|SPN232307 gi|3821434jemb|AJ232282.1|SPN232282 [3821482] [3821434]
gi|3821432|emb|AJ232281.1|SPN232281 gi|3821384|emb|AJ232256.1 SPN232256 [3821432] [3821384] gi|3821430|emb|AJ232280.1|SPN232280 gi|3821382|emb|AJ232255.1 SPN232255 [3821430] [3821382] gi|3821428|emb|AJ232279.1|SPN232279 gi|3821380|emb|AJ232254.1 SPN232254 [3821428] [3821380] gi|3821426|emb|AJ232278.1|SPN232278 gij3821378|emb|AJ232253.1 SPN232253 [3821426] [3821378] gi|3821424|emb|AJ232276.1|SPN232276 gi|3821376|emb|AJ232252.1 SPN232252 [3821424] [3821376] gi|3821422|emb|AJ232275.1|SPN232275 g_|3821374|emb|AJ232251.1 SPN232251 [3821422] [3821374] gi|3821420|emb|AJ232274.1|SPN232274 gi|3821372|emb|AJ232250.1 SPN232250 [3821420] [3821372] gi|3821418|emb|AJ232273.1|SPN232273 gi|3821370|emb|AJ232249.1 SPN232249 [3821418] [3821370] gij3821416|emb|AJ232272.1|SPN232272 gi|3821367|emb|AJ232248.1 SPN232248 [3821416] [3821367] gi|3821414|emb|AJ232271.1|SPN232271 gi|3821365|emb|AJ232247.1 SPN232247 [3821414] [3821365] gi|3821412|emb|AJ232270.1|SPN232270 gi|3821363|emb|AJ232246.1! SPN232246 [3821412] [3821363] gi|3821410|emb|AJ232269.1|SPN232269 gi|3821361|emb|AJ232245.1 SPN232245 [3821410] [3821361] gi|3821408|emb|AJ232268.1|SPN232268 gi|3821359|emb|AJ232244.1 SPN232244 [3821408] [3821359] gi|3821406|emb|AJ232267.1|SPN232267 gi|3821357|emb|AJ232243.1 SPN232243 [3821406] [3821357] gi|3821404|emb|AJ232266.1|SPN232266 gi|3821355|emb|AJ232241.1 SPN232241 [3821404] [3821355] gij3821402|emb|AJ232265.1|SPN232265 gi|2921842|gb|AF047385.1 |AF047385 [2921842] [3821402] gi|2909863|gb|AF047696.1|AF047696 [2909863] gi|3821400|emb|AJ232264.1|SPN232264 gij4193353|gb|AF055088.1|AF055088 [4193353] [3821400] gi|3821398|emb|AJ232263.1|SPN232263 gi|4185242|gb| AH007276.1 |SEG_SPTNJUNC [4185242] [3821398] gi|4185241 |gb| AF066797.1 |SPTNJUNC2 gi|3821396|emb|AJ232262.1|SPN232262 [4185241] [3821396] gi|4185240|gb|AF066796.1 |SPTNJUNC1 gi|3821394|emb|AJ232261.1 |SPN232261 [4185240] [3821394] gi|3821392|emb|AJ232260.1 |SPN232260 gi|4097979|gb|U72655.1 |SPU72655 [4097979] [3821392] gi|4063720|gb|L29323.1 |ST MTR [4063720] gi|3821390|emb|AJ232259.1|SPN232259 gi|1657605|gb|U66846.1|SPU66846 [1657605] _ _ [3821390] gi|1657602|gb|U66845.1|SPU668454l557602] gi|3821388|emb|AJ232258.1|SPN232258 [3821388] gi|4009485|gb|AF068903.1|AF068903 [4009485] gij3821386|emb|AJ232257.1|SPN232257 gi|4009477|gb| AF068902.11 AF068902 [4009477] [3821386] gi|4009462|gb| AF068901.11 AF068901 [4009462]
gi|3947767|emb|AJ233896.1|SPN233896 gij 1498294jgb|U41735.1 |SPU41735 [1498294] [3947767] gi|1213493|gb|U47687.1|SPU47687 [1213493] gi|3947765|emb|AJ233895.1|SPN233895 gi|1163109|gb|U43526.1|SPU43526 [1163109] [3947765] gi|3947763|emb|AJ233894.1|SPN233894 gi|556001|gb|U15171.1|SPU15171 [556001] [3947763] gi|455063|gb|U02920.1 |SPU02920 [455063] gi|3947761 |emb|AJ233893.1 |SPN233893 gi|784896|gb|L36923.1|STRSTRH [784896] [3947761] gi|3320386|gb|AF030373.1 |AF030373 [3320386] gi|3947759|emb|AJ233892.1|SPN233892 gi|2804772|gb|AF030374.1|AF030374 [2804772] [3947759] gi|3947757|emb|AJ233891.1|SPN233891 gi|2804762|gb|AF030372.1|AF030372 [2804762] [3947757] gi|2804756|gb|AF030371.1|AF030371 [2804756] gi|3947755|emb|AJ233890.1|SPN233890 gi|2804750|gb| AF030370.1 |AF030370 [2804750] [3947755] gi|2804745|gb|AF030369.1|AF030369 [2804745] gi|3947753|emb|AJ233889.1|SPN233889 [3947753] gi|2804739|gb|AF030368.1 |AF030368 [2804739] gi|3947751|emb|AJ233888.1|SPN233888 gi|2804732|gb|AF030367.1 |AF030367 [2804732] [3947751] gi|2804726|gb|AF030366.1 |AF030366 [2804726] gi|3947749|emb|AJ233887.1|SPN233887 gi|2804720|gb|AF030365.1|AF030365 [2804720] [3947749] gi|2804713|gb|AF030364.1|AF030364 [2804713] gi|3947730|emb|AJ233886.1|SPN233886 [3947730] gi|2804707|gb|AF030363.1|AF030363 [2804707] gi|3758891|emb|Z71552.1|SPADCA [3758891] gi|2804701|gb|AF030362.1|AF030362 [2804701] gi|3818479|gb| AF057294.1 |AF057294 [3818479] gi|2804694|gb|AF030361.1|AF030361 [2804694] gi|2351 67|gb|U89711.1 |SPU89711 [2351767] gi|2804688|gb|AF030360.1|AF030360 [2804688] gi|3395661|dbj|AB006879.1|AB006879 [3395661] gi|2804682|gb|AF030359.1 |AF030359 [2804682] gi|3395659|dbj|AB006878.1|AB006878 [3395659] gi|3550979|dbj|AB010387.1|AB010387 [3550979] gi|3395657|dbj|AB006877.1|AB006877 [3395657] gi|2275100|emb|AJ000336.1|SPR6LDH [2275100] gi|3395655|dbj] AB006876.1 |AB006876 [3395655] gi|3551853|gb|AF076029.1|AF076029 [3551853] gi|3395653|dbj|AB006875.1|AB006875 [3395653] gi|3551773|gb|U94770.1 |SPU94770 [3551773] gi|3395651 |dbj| AB006874.11 AB006874 [3395651 ] gi|3550617|emb|AJ004869.1|SPAJ4869 [3550617] gi|3395649|dbj|AB006873.1|AB006873 [3395649] gi|3513563|gb|AF055727.1|AF055727 [3513563] gi|3395647|dbj|AB006872.1|AB006872 [3395647] gi|3513561 |gb| AF055726.11 AF055726 [3513561] gi|3395645|dbj|AB006871.1 |AB006871 [3395645] gi|3513559|gb|AF055725.1|AF055725 [3513559] gi|3395643|dbj|AB006870.1|AB006870 [3395643] gi|3513557|gb|AF055724.1|AF055724 [3513557] gi|3395641|dbj|AB006869.1|AB006869 [3395641] gij3513555|gb|AF055723.1|AF055723 [3513555] gi|3395639|dbj|AB006868.1|AB006868 [3395639] gi|3513553|gb|AF055722.1|AF055722 [3513553] gi|2315992|gb|U87092.1 |SPU87092 [2315992] gi|3513549|gb|AF055721.1 |AF055721 [3513549] gi|2209338|gb|U93576.1|SPU93576 [2209338] gi|3513545 |gb| AF055720.11 AF055720 [3513545] gi|2109442|gb|AF000658.1|SPDNAARG gi|1914869|emb|Z82001.1|SPZ82001 [1914869] - [2109442] gi|2911421|gb|AF046238.1|AF046238" 2911421] gi|1881538|gb|U09239.1|SPU09239 [1881538] gi|2911419|gb|AF046237.1!AF046237 [2911419] gi| 1666904|gb|U76218.1 |SPU76218 [ 1666904] gi|2911417|gb|AF046236.1 |AF046236 [2911417] gi|1613766|gb|U33315.1|SPU33315 [1613766] gi|2911415|gb|AF046235.1|AF046235 [2911415]
gi|2911413|gbjAF046234.1|AF046234 [2911413] gi|2765992|embjZ99825.1 ISPZ99825 [2765992] gi|2911411 |gb| AF046233.1 j AF046233 [2911411] gi|2765990|emb|Z99824.1 ISPZ99824 [2765990] gi|2911409|gb|AF046232.1|AF046232 [2911409] gi|2765988|emb|Z99823.1 ISPZ99823 [2765988] gi|2911407|gb|AF046231.1 |AF046231 [2911407] gi|2765986|emb|Z99822.1 ISPZ99822 [2765986] gi|2911405 |gb| AF046230.11 AF046230 [2911405] gi|2765984|emb|Z99821.1 ISPZ99821 [2765984] gi|3258601|gb|U40786.1|SPU40786 [3258601] gij2765982|emb|Z99820.1 ISPZ99820 [2765982] gi|3211756|gb| AF052209.1 |AF052209 [3211756] gi|2765980|embjZ99819.1 ISPZ99819 [2765980] gi|3211752|gb|AF052208.1|AF052208 [3211752] gi|2765978|emb|Z99818.1 ISPZ99818 [2765978] gi|3211747|gb|AF052207.11 AF052207 [3211747] gi|2765976|emb|Z99817.1 ISPZ99817 [2765976] gi|3220194|gb|AF053121.1 JAF053121 [3220194] gi|2765974|emb|Z99816.1 ISPZ99816 [2765974] gi|2766052|emb|Z99863.1 ISPZ99863 [2766052] gi|2765972|emb|Z99815.1 ISPZ99815 [2765972] gi|2766050|emb|Z99862.1 |SPZ99862 [2766050] gi|2765970|emb|Z99814.1 ISPZ99814 [2765970] gi|2766048|emb|Z99861.1 ISPZ99861 [2766048] gi|2765968|emb|Z99813.1 ISPZ99813 [2765968] gi|2766046|emb|Z99860.1 ISPZ99860 [2766046] gi|2765966|emb|Z99812.1 ISPZ99812 [2765966] gi|2766044|emb|Z99859.1 (SPZ99859 [2766044] gi|2765964|emb|Z99811.1 ISPZ99811 [2765964] gi|2766042|embjZ99858.1 ISPZ99858 [2766042] gi|2765962|emb|Z99810.1 ISPZ99810 [2765962] gi|2766040|emb|Z99857.1 |SPZ99857 [2766040] gi|2765960|emb|Z99809.1 ISPZ99809 [2765960] gi|2766038|emb|Z99856.1 ISPZ99856 [2766038] gi|2765958|emb|Z99808.1 ISPZ99808 [2765958] gi|2766036|emb|Z99855.1 ISPZ99855 [2766036] gi|2765956|emb|Z99807.1 ISPZ99807 [2765956] gi|2766034|emb|Z99854.1 |SPZ99854 [2766034] gij2765954|emb|Z99806.1 ISPZ99806 [2765954] gi|2766032|emb|Z99853.1 |SPZ99853 [2766032] gi|2765952|emb|Z99805.1 ISPZ99805 [2765952] gi|2766030|emb|Z99852.1 |SPZ99852 [2766030] gi|2765950|emb|Z99804.1 |SPZ99804 [2765950] gi|2766028|emb|Z99851.1 |SPZ99851 [2766028] gi|2765948|embjZ99803.1 JSPZ99803 [2765948] gi|2766026|emb|Z99850.1 ISPZ99850 [2766026] gi|2894104|emb|X77249.1|SPR6CIARH [2894104] gi|2766024|emb|Z99849.1 ISPZ99849 [2766024] gi|3153897|gb[ AF067128.11 AF067128 [3153897] gij2766022|embjZ99848.1 |SPZ99848 [2766022] gi|3152712|gb|AF065153.1|AF065153 [3152712] gi|2766020|emb|Z99847.1 ISPZ99847 [2766020] gi|3152710|gb| AF065152.11 AF065152 [3152710] gi|2766018|emb|Z99846.1 |SPZ99846 [2766018] gi|3152708|gb| AF065151.11 AF065151 [3152708] gi|2766016|emb|Z99845.1 ISPZ99845 [2766016] gi|3116426|gb|U84387.1 |SPU84387 [3116426] gij2766014|emb|Z99844.1 ISPZ99844 [2766014] gi|2385403|emb|AJ001247.1|SP7465RR3 [2385403] gi|2766012|emb|Z99843.1 ISPZ99843 [2766012] gi|2342540|emb|AJ001250.1|SP7978RR5 gi|2766010|emb|Z99842.1 ISPZ99842 [2766010] [2342540] gi|2766008|emb|Z99841.1 |SPZ99841 [2766008] gi|2342539|emb|AJ001251.1 |SP7978RR3 gi|2766006|emb|Z99840.1 |SPZ99840 [2766006] [2342539] gi|2766004|emb|Z99839.1 |SPZ99839 [2766004] gi|2342538|emb|AJ001248.1|SP7466RR5 [2342538] gi|2766002|emb|Z99838.1 ISPZ99838 [2766002] gi|2342537|emb|AJ001249.1|SP7466RKr "" gi|2766000|emb|Z99837.1 |SPZ99837 [2766000] [2342537] gi|2765998|embjZ99828.1 |SPZ99828 [2765998] gi|3065896|gb|AF058920.1 |AF058920 [3065896] gij2765996|emb|Z99827.1 |SPZ99827 [2765996] gi|2982647|emb|AJ002294.1 JSPAJ2294 [2982647] gij2765994|emb|Z99826.1 JSPZ99826 [2765994]
gi|2982645|emb|AJ002293.1|SPAJ2293 [2982645] gi|2766116|embjZ99895.1 SPZ99895 [2766116] gij2982643|emb|AJ002292.1|SPAJ2292 [2982643] gi|2766114|emb|Z99894.1 ISPZ99894 [2766114] gi|2982641 |emb| AJ002291.1 |SPAJ2291 [2982641 ] gi|2766112|emb|Z99893.1 SPZ99893 [2766112] gi| 1620466jemb|X99400.1 |SPDACAO [ 1620466] gi|2766110|emb|Z99892.1 SPZ99892 [2766110] gi|2196665 |emb|Z84381.1 |HSZ84381 [2196665] gi|2766108|emb|Z99891.1 SPZ99891 [2766108] gi|2196663 |emb|Z84380.1 |HSZ84380 [2196663] gi|2766106|emb|Z99890.1 [SPZ99890 [2766106] gi|2196661 |emb|Z84379.1 |HSZ84379 [2196661 ] gi|2766104|emb|Z99889.1 ISPZ99889 [2766104] gi|2196659|emb|Z84378.1|HSZ84378 [2196659] gi|2766102[emb|Z99888.1 SPZ99888 [2766102] gi|625175|gb|L36131.1|STREXP10A [625175] gi|2766100|emb|Z99887.1 |SPZ99887 [2766100] gi|3004945|gb|AF036624.1 |AF036624 [3004945] gij2766098jemb|Z99886.1l ISPZ99886 [2766098] gi|3004943|gb|AF036623.1|AF036623 [3004943] gi|2766096|emb|Z99885.1! SPZ99885 [2766096] gi|3004941|gb|AF036622.1|AF036622 [3004941] gi|2766094|emb|Z99884.1! SPZ99884 [2766094] gij3004939|gb| AF036621.11 AF036621 [3004939] gi|2766092|emb|Z99883.1 SPZ99883 [2766092] gi|3004937|gb|AF036620.1|AF036620 [3004937] gi|2766090|emb|Z99882.1 ISPZ99882 [2766090] gi|3004935|gb|AF036619.1|AF036619 [3004935] gi|2766088|emb|Z99881.1 SPZ99881 [2766088] gi|2370572|emb|Z86112.1|SPZ86112 [2370572] gi|2766086|emb|Z99880.1 ISPZ99880 [2766086] gi|2765946|emb|Z99802.1 |SPZ99802 [2765946] gi|2766084|emb|Z99879.1 ;SPZ99879 [2766084] gi|2398824|emb|Z34303.1|SPCINREC [2398824] gi|2766082|emb|Z99878.1 SPZ99878 [2766082] gi|2894512|emb|AJ223491.1 |SPPPR3 [2894512] gi|2766080|emb|Z99877.1 SPZ99877 [2766080] gi|2198539|emb|X85787.1 ISPCPS 14E [2198539] gi|2766078|emb|Z99876.1 |SPZ99876 [2766078] gi|2766156|emb|Z99915.1 |SPZ99915 [2766156] gi|2766076|emb|Z99875.1 |SPZ99875 [2766076] gi|2766154|emb|Z99914.1|SPZ99914 [2766154] gi|2766074|emb|Z99874.1 ISPZ99874 [2766074] gi|2766152|emb|Z99913.1|SPZ99913 [2766152] gi|2766072|emb|Z99873.1 SPZ99873 [2766072] gi|2766150|emb|Z99912.1|SPZ99912 [2766150] gi|2766070|emb|Z99872.1 SPZ99872 [2766070] gi|2766148|emb|Z99911.1|SPZ99911 [2766148] gi|2766068|emb|Z99871.1| ISPZ99871 [2766068] gi|2766146|emb|Z99910.1|SPZ99910 [2766146] gi|2766066|emb|Z99870.1| ISPZ99870 [2766066] gi|2766144|emb|Z99909.1|SPZ99909 [2766144] gi|2766064|emb|Z99869.1 ISPZ99869 [2766064] gij2766142|emb|Z99908.1|SPZ99908 [2766142] gi|2766062|emb|Z99868.1 iSPZ99868 [2766062] gi|2766140|emb|Z99907.1|SPZ99907 [2766140] gi|2766060|emb|Z99867.1 SPZ99867 [2766060] gi|2766138|emb|Z99906.1|SPZ99906 [2766138] gi|2766058|emb|Z99866.1 SPZ99866 [2766058] gi|2766136|emb|Z99905.1|SPZ99905 [2766136] gi|2766056|emb|Z99865.1 SPZ99865 [2766056] gi|2766134|emb|Z99904.1 |SPZ99904 [2766134] gi|2766054|emb|Z99864.1 ISPZ99864 [2766054] gi|2766132|emb|Z99903.1|SPZ99903 [2766132] gi|2765906|emb|Z99206.1 ISPZ99206 [2765906] gi|2766130|emb|Z99902.1 |SPZ99902 [2766130] gij2765904|emb|Z99205.1 ISPZ99205 [2765904] gi|2766128|eπ_b|Z99901.1|SPZ99901 [2766128] gi|2765902|emb|Z99204.1 SPZ99204 [2765902] gi|2766126|emb|Z99900.1 JSPZ99900 [2766126] gij2765900|embjZ99203.1 SPZ99203 [27659.00] - gi|2766124|emb|Z99899.1 |SPZ99899 [2766124] gi|2765898|emb|Z99202.1! SPZ99202 [2765898] gi|2766122|emb|Z99898.1 [SPZ99898 [2766122] gi|2765896jemb|Z99201.1| |SPZ99201 [2765896] gi|2766120|emb|Z99897.1|SPZ99897 [2766120] gi|2765894|emb|Z99200.1 ISPZ99200 [2765894] gi|2766118jemb|Z99896.1|SPZ99896 [2766118] gi|2708631 |gb| AF036951. 1|AF036951 [2708631]
gi|886956|emb|Z49097.1|SPCS1112X [886956] gi|1161269|gb|L39074.1|STRSPXB [1161269] gi|2656093|gb|L21856.1|STRMALR [2656093] gi|1460093|emb|X94909.1|SPIGAlPRT [1460093] gi|2576332|emb|AJ002055.1|SPSPSA47 [2576332] gijl750263|gb|U72720.1|SPU72720 [1750263] gi|2576330|emb|AJ002054.1|SPSPSA2 [2576330] gi|298649|gb|S56948.1 |S56948 [298649] gi|2511704|emb|Y10818.1|SPY10818 [2511704] gi|254537|gb|S43511.1 |S43511 [254537] gi|1944619|emb|Z83335.1|SPZ83335 [1944619] gi|245227|gb|S81051.1 |S81051 [245227] gi|2425108|gb|AF019904.11 AFO 19904 [2425108] gi|245226|gb|S81045.1 |S81045 [245226] gi|2385404]emb| AJ001246.1 |SP7465RR5 gi|245225|gb|S81043.1|S81043 [245225] [2385404] gi| 1150618|emb|Z49988.1 ISPMMSAGEN gi|438213|emb|Z16082.1|PNALIB [438213] [1150618] gi|2149613|gb|U90721.1|SPU90721 [2149613] gi|47456|emb|X01138.1|SPTN917A [47456] gi|49391 |emb|Z21841.1 |SPPBP2BB [49391 ] gi|1658316|emb|Z47210.1|SPDEXCAP [1658316] gi|2209207|gb|AF004325.1|AF004325 [2209207] gijl550802|emb|X95385.1|SPCOMCGEN [1550802] gi|2293061|emb|Z95914.1|SPZ95914 [2293061] gi|47457|emb|X01137.1|SPTN917B [47457] gi|2276393|gb|U16156.1|SPU16156 [2276393] gi|975714|emb|X90941.1 |SPTRJ5251 [975714] gi|2183314|gb|AF003930.1 |AF003930 [2183314] gi|2182093|emb|X95717.1|SPPARECGN gi|975713|emb|X90940.1|SPTLJ5251 [975713] [2182093] gi|975709|emb|X90939.1|SPDNATETM [975709] gi|984230jemb|Z49095.1|SPCSl 111A [984230] gi| 1524346|emb|Z79691.1 |SOORFS [ 1524346] gi|886954|emb|Z49096.1|SPCS1092X [886954] gij 1553054|emb|X98364.1 |SPPBPHU9 [ 1553054] gi|1181613|dbj|D82873.1|STRPBP2BE [1181613] gi|1553052|emb|X98367.1|SPPBPHU13 [1553052] gi|1181612|dbj|D82871.1|STRPBP2BCZ gi|1553050|embjX98366.1|SPPBPHU12 [1553050] [1181612] gi|1553048|emb|X98365.1|SPPBPHUl l [1553048] gi|1181611|dbj|D82870.1|STRPBP2BB2 [1181611] gi| 1575029|gb|U53509.1 |SPU53509 [1575029] gi|1181579|dbj|D82869.1|STRPBP2BAl [1181579] gij 1542968|gb|U49088.1|SPU49088 [1542968] gi|1181192|dbj|D82872.1|STRPBP2BD [1181192] gij 1542966|gb|U49087.1 |SPU49087 [ 1542966] gi|575595|dbj|D42075.1|STRPBP2B2 [575595] gi|1536961|emb|Y07845.1|SPGYRA [1536961] gi|1339971|dbj|D42074.1|STRPBP2Bl [1339971] gi|47391|emb|X16367.1|SPPBPX [47391] gi|2108329|embjY11463.1|SPDNAGCPO gi|1490398|emb|Z67739.1|SPPARCETP [1490398] [2108329] gi| 1490395|emb|Z67740.1 |SPGYRBORF gi| 1944115|dbj|AB002522.1 |AB002522 [ 1944115] [1490395] gi|1666669|emb|Z77727.1|SPIS1381C [1666669] gij 1431589|emb|Z74777.1 |SPTMRDHFR gi| 1666668|emb|Z77726.1 |SPIS 1381 B [ 1666668] [1431589] gi|1666667|emb|Z77725.1|SPIS1381A [1666667] gi|408145|emb|Z21702.1|SPUNGMUTX [408145] gi|1914873|emb|Z82002.1|SPZ82002 [1914873] gi|47461 |emb|X61025.1 |SPXISINT [47461 ] gi|1431584|emb|Z74778.1|SPDHFR [1431584] gi|47459|emb|X55651.1 |SPUNGG [47459] gi|47452|emb|Z15120.1|SPSTRG [47452] gi|47454|emb|X52632.1|SPT1545E [47454] _ . gi|581717|emb|Z12159.1|SPCP131G [581717] gi|47421 |emb|Zl 7307.1 |SPRECA [424211 gi|47342|emb|X17337.1|SPAMILOC [47342] gi|47419|emb|X67873.1|SPPONA8 [47419] gi| 1800300|gb|U83667.1 |SPU83667 [1800300] gi|47417|emb|X67872.1|SPPONA7 [47417] gi|1532066|emb|Y07780.1|SPTET0GEN [1532066] gi|47415|emb|X67871.1|SPPONA6 [47415]
gi|47413|emb|X67870.1|SPPONA5 [47413] gi|47331 |emb|X65133.1 |SP577PBPX [47331] gi|47411 |emb|X67869.1 |SPPONA4 [47411 ] gi|559527|emb|X65136.1 |SP110PBPX [559527] gij47409|emb|X67867.1jSPPONA2 [47409] gpl 1415|emb|Z22807.1|SP16SRNAA [311415] gi|47407|emb|X67866.1|SPPONAl [47407] gi|47329|emb|X65135.1 |SP531PBPX [47329] gi|47405|emb|X67868.1|SPPNA3 [47405] gi|47307|emb|X65131.1 |SP290PBPX [47307] gi|47403|emb|X52474.1|SPPLY [47403] gi|47295|emb|X58312.1|SP16SRNA [47295] gi|984232|emb|X16022.1|SPPENA [984232] gi|854614|emb|Z49109.1|SPGADAGN [854614] gi|517190|emb|X78215.1|SPPBPXG [517190] gi|556428|gb|L36660.1|STRORFl [556428] gi|295840|emb|Z22230.1 |SPPBP2BBA [295840] gi|511062|embjZ35135.1|SPALIAG [511062] gi|288981|emb|Z22185.1|SPPBP2BAC [288981] gi|1208737|gb|U47625.1|SPU47625 [1208737] gi|288979|emb|Z22184.1 |SPPBP2BAB [288979] gi|530062|gb|U12567.1|SPU12567 [530062] gi|288466|emb|Z21981.1 |SPPBP2BAA [288466] gi| 153656|gb|M29686.1 |STRHEXB [ 153656] gi|49390|emb|Z21813.1 |SPPBP2XD [49390] gi| 153654|gb|Ml 8729.1 |STRHEXA [153654] gi|49389|emb|Z21812.1 |SPPBP2XC [49389] gi|153608|gb|M14339.1|STRDPN2A [153608] gi|49387|emb|Z21811.1|SPPBP2BJ [49387] gi|153605|gb|M14340.1|STRDPNlA [153605] gi|49385|emb|Z21810.1|SPPBP2BI [49385] gi|643543|gb|U20084.1|SPU20084 [643543] gi|49382|emb|Z21808.1 |SPPBP2BH [49382] gi|643541 |gb|U20083.1 |SPU20083 [643541 ] gi|49380|emb|Z21807.1 |SPPBP2BG [49380] gi|643539|gb|U20082.1 |SPU20082 [643539] gi|49379|emb|Z21806.1 |SPPBP2BF [49379] gi|643537|gb|U20081.1|SPU20081 [643537] gi|49377|emb|Z21805.1 |SPPBP2BE [49377] gi|643535jgb|U20080.1 |SPU20080 [643535] gi|49376|emb|Z21804.1 |SPPBP2XB [49376] gi|643533|gbjU20079.1|SPU20079 [643533] gi|49375|emb|Z21803.1 |SPPBP2XA [49375] gi|643531 |gb|U20078.1 |SPU20078 [643531 ] gij49374|emb|Z21802.1 |SPPBP2BD [49374] gi|643529|gb|U20077.1 |SPU20077 [643529] gij49372|emb|Z21801.1|SPPBP2BC [49372] gi|643527|gb|U20076.1|SPU20076 [643527] gi|49369|emb|Z21799.1 |SPPBP2B A [49369] gi|643525|gb|U20075.1|SPU20075 [643525] gi|47399|emb|X13137.1|SPPENASE [47399] gi|643523|gb|U20074.1|SPU20074 [643523] gi|47397|emb|X13136.1|SPPENARE [47397] gi|643521 |gb|U20073.1 |SPU20073 [643521 ] gi| 1052802|emb|X83917.1 |SPGYRBG [ 1052802] gi|643519|gb|U20072.1|SPU20072 [643519] gi|587550|emb|X72967.1|SPNANA [587550] gi|643517|gb|U20071.1 |SPU20071 [643517] gi|49384|emb|Z21809.1|SPPBPlAB [49384] gi|643515|gbjU20070.1 |SPU20070 [643515] gi[49371 |emb|Z21800.1 |SPPBP 1 AA [49371 ] gi|643513|gb|U20069.1|SPU20069 [6435131 gi|984228|emb|Z49094.1|SPCS1091A [984228] gi|643511 jgb|U20068.1 |SPU20068 [643511 ] gi|47372|emb|X54225.1|SPENDA [47372] gi|643509|gb|U20067.1 |SPU20067 [643509] gij806590|emb|Z49246.1 |SP667SOD [806590] gi|1017802|gb|U37560.1|SPU37560 [1017802] gij407172|emb|Z26851.1 |SPATPAS2 [407172] gi|663277|gb|M36180.1 |STRCOMAA [663277] gi|407166|emb|Z26850.1 |SPATPAS 1 [407166] gi|437704|gb|L20670.1 |STRHYALURO [437704] gi|47353|emb|X63602.1|SPBOX [47353] gi| 153849|gb|L07751.1 |TRNTN5252RTF53849] gi|47348|emb|X05577.1 |SPAPHA3 [47348] gi|153855|gb|M25519.1|STRVAl [153855] gi|47337|emb|X65132.1|SP824PBPX [47337] gi|153853|gb|M80215.1|STRUVS402A [153853] gi|47335|emb|X65134.1 JSP669PBPX [47335] gij 153848|gb|L07750.1 |STRTN5252L [153848]
gij 153840|gb|M74122.1 |STRSURPROA [ 153840] gi|153796|gb|M60763.1|STRRRNAA [153796] gi|l 5379 l|gb|M31296.1ISTRRECP [153791] gi|516639|gb|L20556.1 |STRPLPA [516639] gi|153783|gb|M28679.1|STRPROMB [153783] gi|153782|gb|M28678.1|STRPROMA [153782] gi| 153766|gb|M90527.1 |STRPONA [ 153766] gi| 153764|gb| J04479.1 |STRPOLA [ 153764] gi|153752|gbjM25515.1|STRNG4369 [153752] gi| 153722|gb|L08611.1 |STRMLTODX [ 153722] gi|153702|gb|J01796.1|STRMALMXP [153702] gi|153701|gb|J01795.1|STRMALMX [153701] gi|153693|gb|M13812.1|STRLYTPN [153693] gi| 153691 |gbjM 17717.1 |STRLYS [ 153691 ] gi|153667|gb|M25525.1|STRKAG73 [153667] gi|398102jgb|L20564.1|STREXP9B [398102] gi|398100|gb|L20563.1 |STREXP9A [398100] gi|398098|gb|L20562.1|STREXP8A [398098] gi|398096|gb|L20561.1 |STREXP7A [398096] gi|398094|gb|L20560.1 |STREXP6A [398094] gij398092|gb|L20559.1 |STREXP5A [398092] gi|398090|gb|L20558.1|STREXP4A [398090] gi| 153626|gb| J04234.1 |STREXOA [ 153626] gi|153612|gb|M11226.1|STRDPNM [153612] gi| 153603|gb|M25521.1 |STRDN87669 [153603] gi|153601|gb|M25526.1|STRDN87577 [153601] gi|153599|gb|M25522.1|STRDN179 [153599] gi| 153594|gb|M37688.1 ISTRDACA [ 153594] gi|153582|gb|L07752.1|STRATTB [153582] gi|466514|gb|L31413.1|STRlRRA [466514] gi|153551|gb|M25520.1|STR8249 [153551] gi|153549|gb|M25524.1|STR5313972 [153549] gi| 153547|gb|M25517.1 |STR29044 [ 153547] gi|153545|gb|M25523.1|STR181071 [153545] gi|153541|gb|M25518.1|STR121 [153541] gi| 153539|gb|M25516.1 [STR110K70 [153539] - - gi|506632|gb|U04047.1|SPU04047 [506632] gi|393267jgb|L19055.1|STRPAPA [393267] gi|442066|gbjS62272.1 |S62272 [442066] gi|295191 jgbjL 15190.1 jSTRPURIS YN [295191 ]