NUCLEIC ACID BASED MODULATORS OF GENE EXPRESSION Background ofthe Invention
This invention relates to reagents useful as inhibitors of gene expression relating to diseases such as cancers, diabetes, obesity, Alzheimer's disease, cardiac diseases, age- related diseases, and/or hepatitis B infections and related conditions.
Summary of the Invention
The invention features novel nucleic acid-based techniques [e.g., enzymatic nucleic acid molecules (ribozymes), antisense nucleic acids, 2-5A antisense chimeras, triplex DNA, antisense nucleic acids containing RNA cleaving chemical groups (for example, Cook et al., U.S. Patent 5,359,051)] and methods for their use to modulate the expression of molecular targets impacting the development and progression of cancers, diabetes, obesity, Alzheimer's disease, cardiac diseases, age-related diseases, and or hepatitis B infections and related conditions In a preferred embodiment, the invention features novel nucleic acid-based techniques [e.g., enzymatic nucleic acid molecules (ribozymes), antisense nucleic acids, 2- 5A antisense chimeras, triplex DNA, antisense nucleic acids containing RNA cleaving chemical groups (for exaple, Cook et al., U.S. Patent 5,359,051)] and methods for their use for inhibiting the expression of disease related genes, e.g., Protein-Tyrosine-Phosphatase- lb (PTP-IB, Genbank accession No. NM_002827), Methionine Aminopeptidase (MetAP- 2, Genbank accession No. U29607), beta-Secretase (BACE, Genbank accession No. AF190725), Presenilin-1 (ps-1, Genbank accession No. L76517), Presenilin-2 (ps-2, Genbank accession No. L43964), Human Epidermal Growth Factor Receptor-2 (HER2/c- erb2/neu, Genbank accession No. X03363), Phosphola ban (PLN, Genbank accession No. NM_002667), Telomerase (TERT, Genbank accession No. NM_003219) and Hepatitis B virus genes (HBV, Genbank accession No. AF 100308.1). Such ribozymes can be used in a method for treatment of diseases caused by the expression of these genes in man and other animals, including other primates.
Thus, in an additional preferred embodiment, the invention features novel nucleic acid-based techniques such as enzymatic nucleic acid molecules and antisense molecules and methods for their use to down regulate or inhibit the expression of genes encoding Protein-Tyrosine-Phosphatase-lb (PTP-IB), Methionine Aminopeptidase (MetAP -2),
beta-Secretase (BACE), Presenilin-1 (ps-1), Presenilin-2 (ps-2), Human Epidermal Growth Factor Receptor-2 (HER2/c-erb2/neu), Phospholamban (PLN), Telomerase (hTERT) PKC alpha, and Hepatitis B (HBV) proteins. In particular, applicant describes the selection and function of nucleic acid molecules capable of cleaving RNAs encoded by these genes and their use to reduce levels of PTP-IB, MetAP-2, BACE, ps-1 , ps-2, HER2, PLN, TERT, and/or HBV proteins in various tissues to treat the diseases discussed herein. Such nucleic acid molecules are also useful for diagnostic uses.
In a preferred embodiment, the invention features the use of one or more ofthe nucleic acid-based techniques independently or in combination to inhibit the expression of the genes encoding PTP-IB, MetAP-2, BACE, ps-1 , ps-2, HER2, PLN, TERT, and/or HBV. Specifically, the invention features the use of nucleic acid-based techniques to - specifically inhibit the expression of PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, PKC alpha, and/or HBV genes.
In yet another preferred embodiment, the invention features the use of an enzymatic nucleic acid molecule, preferably in the hammerhead, NCH (Inozyme), G-cleaver, amberzyme, zinzyme, and/or DNAzyme motif, to inhibit the expression of PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, PKC alpha and/or HBV RNA.
Applicant indicates that these nucleic acid molecules are able to inhibit expression of PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, PKC alpha, and/or HBV genes. Those of ordinary skill in the art, will find that it is clear from the examples described that other nucleic acid molecules that inhibit target PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV encoding mRNAs may be readily designed and are within the scope ofthe invention.
By "inhibit" it is meant that the activity of target genes or level of mRNAs or equivalent RNAs encoding target genes is reduced below that observed in the absence of the nucleic acid molecules of the instant invention (e.g., enzymatic nucleic acid molecules), antisense nucleic acids, 2-5A antisense chimeras, triplex DNA, antisense nucleic acids containing RNA cleaving chemical groups). In one embodiment, inhibition with an enzymatic nucleic acid molecule preferably is below that level observed in the presence of an enzymatically attenuated nucleic acid molecule that is able to bind to the same site on the mRNA, but is unable to cleave that RNA. In another embodiment, inhibition with nucleic acid molecules, including enzymatic nucleic acid and antisense
molecules, is preferably greater than that observed in the presence of, for example, an oligonucleotide with scrambled sequence or with mismatches. In another embodiment, inhibition of target genes with the nucleic acid molecule ofthe instant invention is greater than in the presence ofthe nucleic acid molecule than in its absence. According to the invention, the activity of telomerase enzyme or the level of RNA encoding one or more portein subunits ofthe telomerase enzyme is inhibited if it is at least 10% less, 20% less, 50% less, 75% less or even not active or present at all, in the presence of a nucleic acid of the invention relative to the level in the absence of such a nucleic acid.
By "enzymatic nucleic acid molecule" it is meant a nucleic acid molecule which has complementarity in a substrate binding region to a specified gene target, and also has an enzymatic activity which is active to specifically cleave target RNA. That is, the enzymatic nucleic acid molecule is able to intermolecularly cleave RNA and thereby inactivate a target RNA molecule. These complementary regions allow sufficient hybridization ofthe enzymatic nucleic acid molecule to the target RNA and thus permit cleavage. One hundred percent complementarity is preferred, but complementarity as low as 50-75% may also be useful in this invention. The nucleic acids may be modified at the base, sugar, and/or phosphate groups. The term enzymatic nucleic acid is used interchangeably with phrases such as ribozymes, catalytic RNA, enzymatic RNA, catalytic DNA, aptazyme or aptamer-binding ribozyme, regulatable ribozyme, catalytic oligonucleotides, nucleozyme, DNAzyme, RNA enzyme, endoribonuclease, endonuclease, minizyme, leadzyme, oligozyme or DNA enzyme. All of these terminologies describe nucleic acid molecules with enzymatic activity. The specific enzymatic nucleic acid molecules described in the instant application are not meant to be limiting and those skilled in the art will recognize that all that is important in an enzymatic nucleic acid molecule of this invention is that it have a specific substrate binding site which is complementary to one or more ofthe target nucleic acid regions, and that it have nucleotide sequences within or surrounding that substrate binding site which impart a nucleic acid cleaving activity to the molecule (Cech et al., U.S. Patent No. 4,987,071 ; Cech et al., 1988, JAMA 260:20 3030-4). By "nucleic acid molecule" as used herein is meant a molecule having nucleotides.
The nucleic acid can be single, double, or multiple stranded and may comprise modified or unmodified nucleotides or non-nucleotides or various mixtures and combinations thereof.
An example of a nucleic acid molecule according to the invention is a gene which encodes for a macromolecule such as a protein.
By "enzymatic portion" or "catalytic domain" is meant that portion/region ofthe enzymatic nucleic acid molecule essential for cleavage of a nucleic acid substrate (for example see Figures 1-5).
By "substrate binding arm" or "substrate binding domain" is meant that portion/region of a ribozyme which is complementary to (i.e., able to base-pair with) a portion of its substrate. Generally, such complementarity is 100%, but can be less if desired. For example, as few as 10 bases out of 14 may be base-paired. Such arms are shown generally in Figures 1-5. That is, these arms contain sequences within a ribozyme which are intended to bring ribozyme and target RNA together through complementary base-pairing interactions. The ribozyme ofthe invention may have binding arms that are contiguous or non-contiguous and may be of varying lengths. The length ofthe binding arm(s) are preferably greater than or equal to four nucleotides and of sufficient length to stably interact with the target RNA; specifically 12-100 nucleotides; more specifically 14- 24 nucleotides long. If two binding arms are chosen, the design is such that the length of the binding arms are symmetrical (i.e., each ofthe binding arms is ofthe same length; e.g., five and five nucleotides, six and six nucleotides or seven and seven nucleotides long) or asymmetrical (i.e., the binding arms are of different length; e.g., six and three nucleotides; three and six nucleotides long; four and five nucleotides long; four and six nucleotides long; four and seven nucleotides long; and the like). Binding arms can be complementary to the specified substrate, to a portion ofthe indicated substrate, to the indicated substrate sequence and additional adjacent sequence, or a portion ofthe indicated sequence and additional adjacent sequence. By "NCH" or "Inozyme" motif is meant, an enzymatic nucleic acid molecule comprising a motif as described in Ludwig et al, USSN No. 09/406,643, filed September 27, 1999, entitled "COMPOSITIONS HAVING RNA CLEAVING ACTIVITY", and International PCT publication Nos. WO 98/58058 and WO 98/58057, all incorporated by reference herein in their entirety, including the drawings. By "G-cleaver" motif is meant, an enzymatic nucleic acid molecule comprising a motif as described in Eckstein et al, International PCT publication No. WO 99/16871, incoφorated by reference herein in its entirety, including the drawings.
By "zinzyme" motif is meant, a class II enzymatic nucleic acid molecule comprising a motif as described herein and in Beigelman et al, International PCT publication No. WO 99/55857, incoφorated by reference herein in its entirety, including the drawings. By "amberzyme" motif is meant, a class I enzymatic nucleic acid molecule comprising a motif as described herein and in Beigelman et al, International PCT publication No. WO 99/55857, incoφorated by reference herein in its entirety, including the drawings.
By 'DNAzyme' is meant, an enzymatic nucleic acid molecule lacking a ribonucleotide (2' -OH) group. In particular embodiments, the enzymatic nucleic acid molecule may have an attached linker(s) or other attached or associated groups, moieties, or chains containing one or more nucleotides with 2'-OH groups. A DNAzyme can be synthesized chemically or can be expressed by means of a single stranded DNA vector or equivalent thereof.
By "sufficient length" is meant an oligonucleotide of greater than or equal to 3 nucleotides that is of a length great enough to provide the intended function under the expected condition. For example, for binding arms of enzymatic nucleic acid "sufficient length" means that the binding arm sequence is long enough to provide stable binding to a target site under the expected binding conditions. Preferably, the binding arms are not so long as to prevent useful turnover. By "stably interact" is meant, interaction ofthe oligonucleotides with target nucleic acid (e.g., by forming hydrogen bonds with complementary nucleotides in the target under physiological conditions).
By "equivalent" RNA to PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV is meant to include those naturally occurring RNA molecules having homology (partial or complete) to PTP-IB, MetAP-2, BACE, ps-1 , ps-2, HER2, PLN, TERT, and/or HBV proteins or encoding for proteins with similar function as PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV in various organisms, including human, rodent, primate, rabbit, pig, protozoans, fungi, plants, and other microorganisms and parasites. The equivalent RNA sequence also includes in addition to the coding region, regions such as 5 '-untranslated region, 3 '-untranslated region, introns, intron-exon junction and the like in HBV.
By "homology" is meant the nucleotide sequence of two or more nucleic acid molecules is partially or completely identical.
By "antisense nucleic acid", it is meant a non-enzymatic nucleic acid molecule that binds to target RNA by means of RNA-RNA or RNA-DNA or RNA-PNA (protein nucleic acid; Egholm et al, 1993 Nature 365, 566) interactions and alters the activity ofthe target RNA (for a review, see Stein and Cheng, 1993 Science 261, 1004 and Woolf et al, US patent No. 5,849,902). Typically, antisense molecules will be complementary to a target sequence along a single contiguous sequence ofthe antisense molecule. However, in certain embodiments, an antisense molecule may bind to substrate such that the substrate molecule forms a loop, and/or an antisense molecule may bind such that the antisense molecule forms a loop. Thus, the antisense molecule may be complementary to two (or even more) non-contiguous substrate sequences or two (or even more) non-contiguous sequence portions of an antisense molecule may be complementary to a target sequence or both. For a review of current antisense strategies, see Schmajuk et al, 1999, J. Biol. Chem., 27 A, 21783-21789, Delihas et al, 1997, Nature, 15, 751-753, Stein et al, 1997, Antisense N. A. Drug Dev., 7, 151, Crooke, 1998, Biotech. Genet. Eng. Rev., 15, 121-157, Crooke, 1997, Ad. Pharmacol, 40, 1-49. In addition, antisense DNA can be used to target RNA by means of DNA-RNA interactions, thereby activating RNase H, which digests the target RNA in the duplex. Antisense DNA can be synthesized chemically or can be expressed via the use of a single stranded DNA expression vector or the equivalent thereof.
By "2-5A antisense chimera" it is meant, an antisense oligonucleotide containing a 5'-phosphorylated 2'-5'-linked adenylate residue. These chimeras bind to target RNA in a sequence-specific manner and activate a cellular 2-5 A-dependent ribonuclease which, in turn, cleaves the target RNA (Torrence et al, 1993 Proc. Natl. Acad. Sci. USA 90, 1300). By "triplex DNA" it is meant an oligonucleotide that can bind to a double-stranded DNA in a sequence-specific manner to form a triple-strand helix. Formation of such triple helix structure has been shown to inhibit transcription ofthe targeted gene (Duval- Valentin et al, 1992, Proc. Natl. Acad. Sci.USA, 89, 504). By "gene" it is meant a nucleic acid that encodes a RNA.
By "complementarity" is meant that a nucleic acid can form hydrogen bond(s) with another RNA sequence by either traditional Watson-Crick or other non-traditional types. In reference to the nucleic molecules ofthe present invention, the binding free energy for a nucleic acid molecule with its target or complementary sequence is sufficient to allow the relevant function ofthe nucleic acid to proceed, e.g., ribozyme cleavage, antisense or triple helix inhibition. Determination of binding free energies for nucleic acid molecules is well known in the art (see, e.g., Turner et al., 1987, CSH Symp. Quant. Biol. LJJ pp.123-133; Frier et al., 1986, Proc. Nat. Acad. Sci. USA 83:9373-9377; Turner et al., 1987, J. Am. Chem. Soc. 109:3783-3785). A percent complementarity indicates the percentage of contiguous residues in a nucleic acid molecule which can form hydrogen bonds (e.g.,
Watson-Crick base pairing) with a second nucleic acid sequence (e.g., 5, 6, 7, 8, 9, 10 out of 10 being 50%, 60%, 70%, 80%, 90%, and 100% complementary). "Perfectly complementary" means that all the contiguous residues of a nucleic acid sequence will hydrogen bond with the same number of contiguous residues in a second nucleic acid sequence.
At least seven basic varieties of naturally-occurring enzymatic RNAs are known presently. Each can catalyze the hydrolysis of RNA phosphodiester bonds in trans (and thus can cleave other RNA molecules) under physiological conditions. Table I summarizes some ofthe characteristics of these ribozymes. In general, enzymatic nucleic acids act by first binding to a target RNA. Such binding occurs through the target binding portion of a enzymatic nucleic acid which is held in close proximity to an enzymatic portion ofthe molecule that acts to cleave the target RNA. Thus, the enzymatic nucleic acid first recognizes and then binds a target RNA through complementary base-pairing, and once bound to the correct site, acts enzymatically to cut the target RNA. Strategic cleavage of such a target RNA will destroy its ability to direct synthesis of an encoded protein. After an enzymatic nucleic acid has bound and cleaved its RNA target, it is released from that RNA to search for another target and can repeatedly bind and cleave new targets. Thus, a single ribozyme molecule is able to cleave many molecules of target RNA. In addition, the ribozyme is a highly specific inhibitor of gene expression, with the specificity of inhibition depending not only on the base-pairing mechanism of binding to the target RNA, but also on the mechanism of target RNA cleavage. Single mismatches,
or base-substitutions, near the site of cleavage can completely eliminate catalytic activity of a ribozyme.
The enzymatic nucleic acid molecule that cleave the specified sites in PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV-specific RNAs represent a novel therapeutic approach to treat a variety of pathologic indications, including, HBV infection, hepatitis, hepatocellular carcinoma, tumorigenesis, cirrhosis, liver failure, cancers including breast, ovarian, prostate, and esophogeal cancer, tumorigenesis, retinopathy, arthritis, psoriasis, female reproduction, restinosis, certain infectious diseases, transplant rejection and autoimmune disease such as multiple sclerosis, lupus, and AIDS, age related diseases such as macular degeneration and skin ulceration, Alzheimer's disease, dementia, diabetes, obesity and any other condition related to the level of PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and or HBV in a cell or tissue. In one ofthe preferred embodiments ofthe inventions described herein, the enzymatic nucleic acid molecule is formed in a hammerhead or haiφin motif, but may also be formed in the motif of a hepatitis delta virus, group I intron, group U intron or RNase P RNA (in association with an RNA guide sequence), Neurospora VS RNA, DNAzymes, NCH cleaving motifs, or G-cleavers. Examples of such hammerhead motifs are described by Dreyfus, supra, Rossi et al, 1992, AIDS Research and Human Retroviruses 8, 183. Examples of haiφin motifs are described by Hampel et al, EP0360257, Hampel and Tritz, 1989 Biochemistry 28, 4929, Feldstein et al, 1989, Gene 82, 53, Haseloff and Geriach, 1989, Gene, 82, 43, Hampel et al, 1990 Nucleic Acids Res. 18, 299; and Chowrira & McSwiggen, US. Patent No. 5,631,359. The hepatitis delta virus motif is described by Perrotta and Been, 1992 Biochemistry 31, 16. The RNase P motif is described by Guerrier-Takada et al, 1983 Cell 35, 849; Forster and Airman, 1990, Science 249, 783; and Li and Airman, 1996, Nucleic Acids Res. 24, 835. The Neurospora VS RNA ribozyme motif is described by Collins (Saville and Collins, 1990 Cell 61, 685-696; Saville and Collins, 1991 Proc. Natl. Acad. Sci. USA 88, 8826-8830; Collins and Olive, 1993 Biochemistry 32, 2795-2799; and Guo and Collins, 1995, EMBO. J. 14, 363). Group II introns are described by Griffin et al, 1995, Chem. Biol. 2, 761; Michels and Pyle, 1995, Biochemistry 34, 2965; and Pyle et al, International PCT Publication No. WO 96/22689. The Group I intron is described by Cech et al, U.S. Patent 4,987,071. DNAzymes are described by Usman et al, International PCT Publication No. WO 95/11304; Chartrand et
al., 1995, NAR 23, 4092; Breaker et al, 1995, Chem. Bio. 2, 655; and Santoro et al, 1997, PNAS 94, 4262. NCH cleaving motifs are described in Ludwig & Sproat, International PCT Publication No. WO 98/58058; and G-cleavers are described in Kore et al, 1998, Nucleic Acids Research 26, 4116-4120 and Eckstein et al, International PCT Publication No. WO 99/16871. Additional motifs include the Aptazyme (Breaker et al, WO 98/43993), Amberzyme (Class I motif; Figure 3; Beigelman et al, International PCT publication No. WO 99/55857) and Zinzyme (Beigelman et al, International PCT publication No. WO 99/55857), all these references are incoφorated by reference herein in their totalities, including drawings and can also be used in the present invention. These specific motifs are not limiting in the invention and those skilled in the art will recognize that all that is important in an enzymatic nucleic acid molecule of this invention is that it has a specific substrate binding site which is complementary to one or more ofthe target gene RNA regions, and that it have nucleotide sequences within or surrounding that substrate binding site which impart an RNA cleaving activity to the molecule (Cech et al, U.S. Patent No. 4,987,071).
In preferred embodiments ofthe present invention, a nucleic acid molecule, e.g., an antisense molecule, a triplex DNA, or a ribozyme, is 13 to 100 nucleotides in length, e.g., in specific embodiments 35, 36, 37, or 38 nucleotides in length (e.g., for particular ribozymes or antisense). In particular embodiments, the nucleic acid molecule is 15-100, 17-100, 20-100, 21-100, 23-100, 25-100, 27-100, 30-100, 32-100, 35-100, 40-100, 50-100, 60-100, 70-100, or 80-100 nucleotides in length. Instead of 100 nucleotides being the upper limit on the length ranges specified above, the upper limit ofthe length range can be, for example, 30, 40, 50, 60, 70, or 80 nucleotides. Thus, for any ofthe length ranges, the length range for particular embodiments has lower limit as specified, with an upper limit as specified which is greater than the lower limit. For example, in a particular embodiment, the length range can be 35-50 nucleotides in length. All such ranges are expressly included. Also in particular embodiments, a nucleic acid molecule can have a length which is any ofthe lengths specified above, for example, 21 nucleotides in length. In a preferred embodiment, the invention provides a method for producing a class of nucleic acid-based gene inhibiting agents which exhibit a high degree of specificity for the RNA of a desired target. For example, the enzymatic nucleic acid molecule is preferably targeted to a highly conserved sequence region of target RNAs encoding PTP-IB, MetAP-
2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV proteins (specifically PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV RNA) such that specific treatment of a disease or condition can be provided with either one or several nucleic acid molecules ofthe invention. Such nucleic acid molecules can be delivered exogenously to specific tissue or cellular targets as required. Alternatively, the nucleic acid molecules (e.g., ribozymes and antisense) can be expressed from DNA and/or RNA vectors that are delivered to specific cells.
As used in herein "cell" is used in its usual biological sense, and does not refer to an entire multicellular organism, e.g., specifically does not refer to a human. The cell may be present in an organism which may be a human but is preferably a non-human multicellular organism, e.g., birds, plants and mammals such as cows, sheep, apes, monkeys, swine, dogs, and cats. The cell may be prokaryotic (e.g., bacterial cell) or eukaryotic (e.g., mammalian or plant cell).
By "PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV proteins" is meant, a protein or a mutant protein derivative thereof, comprising sequence expressed and/or encoded by PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, genes and/or the HBV genome respectively.
By "highly conserved sequence region" is meant a nucleotide sequence of one or more regions in a target gene does not vary significantly from one generation to the other or from one biological system to the other.
The enzymatic nucleic acid-based inhibitors of PTP-IB, MetAP-2, BACE, ps-1, ps- 2, HER2, PLN, TERT, and/or HBV expression are useful for the prevention ofthe diseases and conditions including HBV infection, hepatitis, hepatocellular carcinoma, tumorigenesis, cirrhosis, liver failure, cancers including breast, ovarian, prostate, and esophogeal cancer, tumorigenesis, retinopathy, arthritis, psoriasis, female reproduction, restinosis, certain infectious diseases, transplant rejection and autoimmune disease such as multiple sclerosis, lupus, and AIDS, age related diseases such as macular degeneration and skin ulceration, Alzheimer's disease, dementia, diabetes, obesity and any other condition related to the level of PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV in a cell or tissue, and any other diseases or conditions that are related to the levels of PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV in a cell or tissue.
By "related" is meant that the reduction of PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV expression (specifically PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV genes) RNA levels and thus reduction in the level ofthe respective protein will relieve, to some extent, the symptoms ofthe disease or condition.
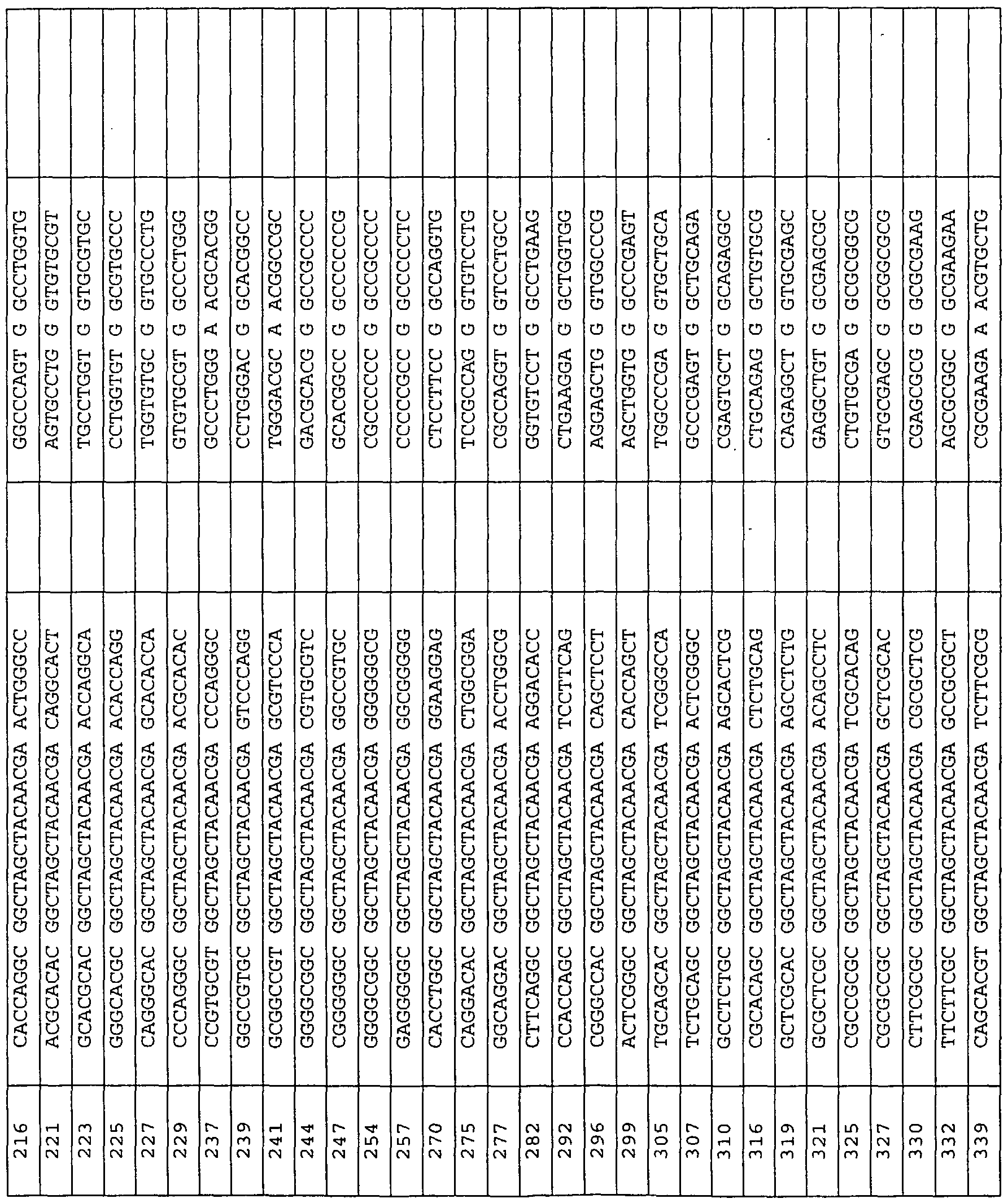
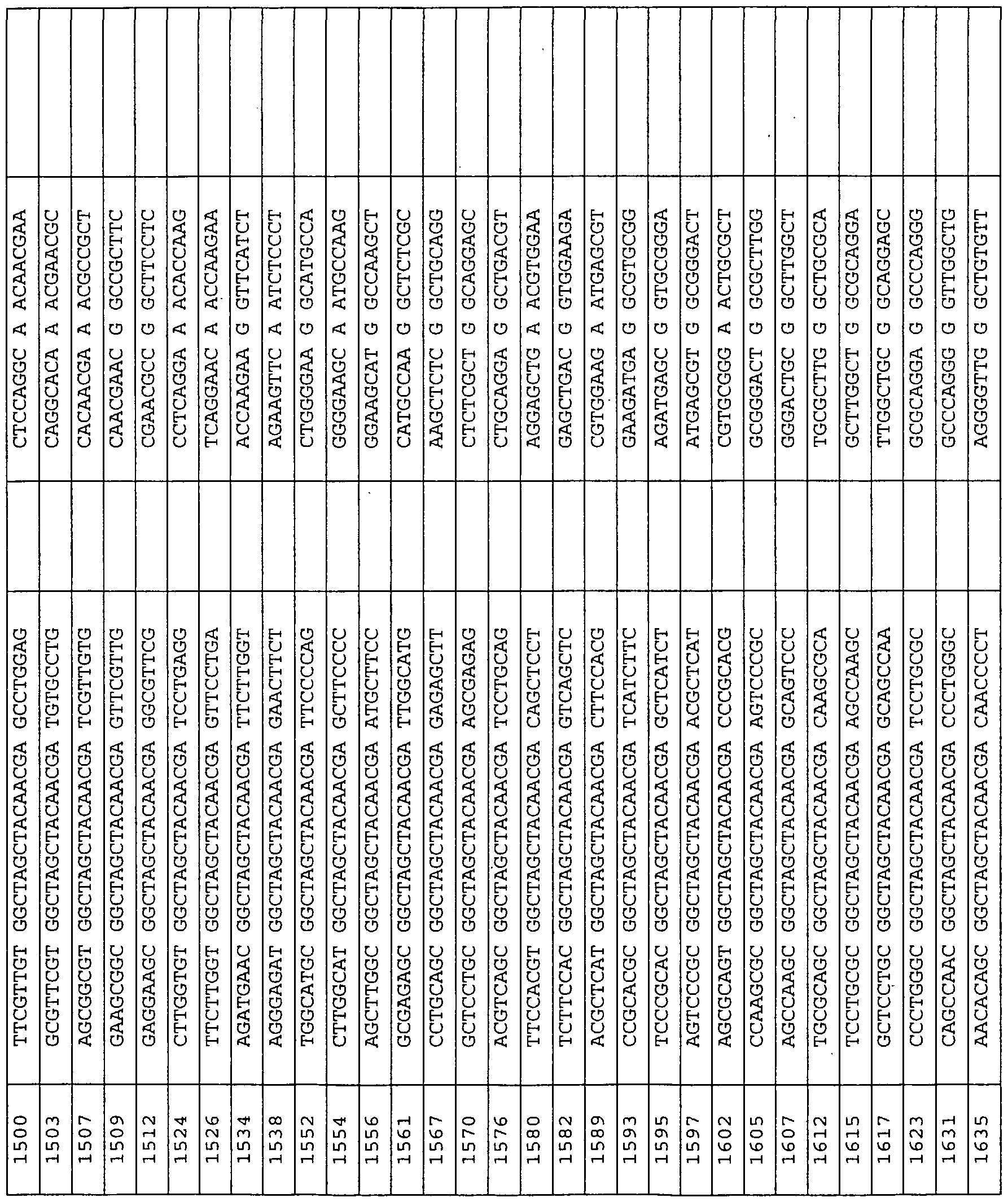
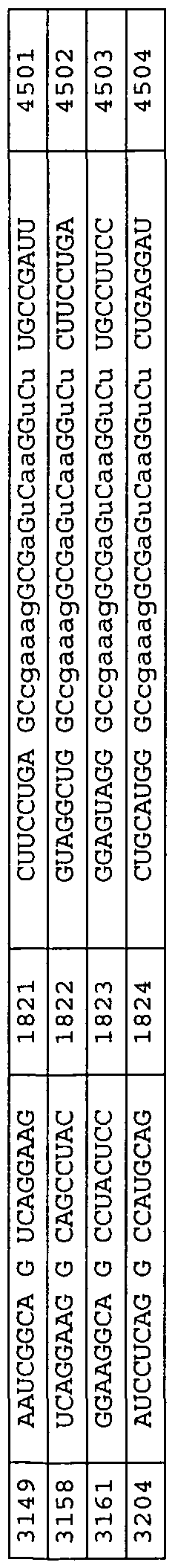
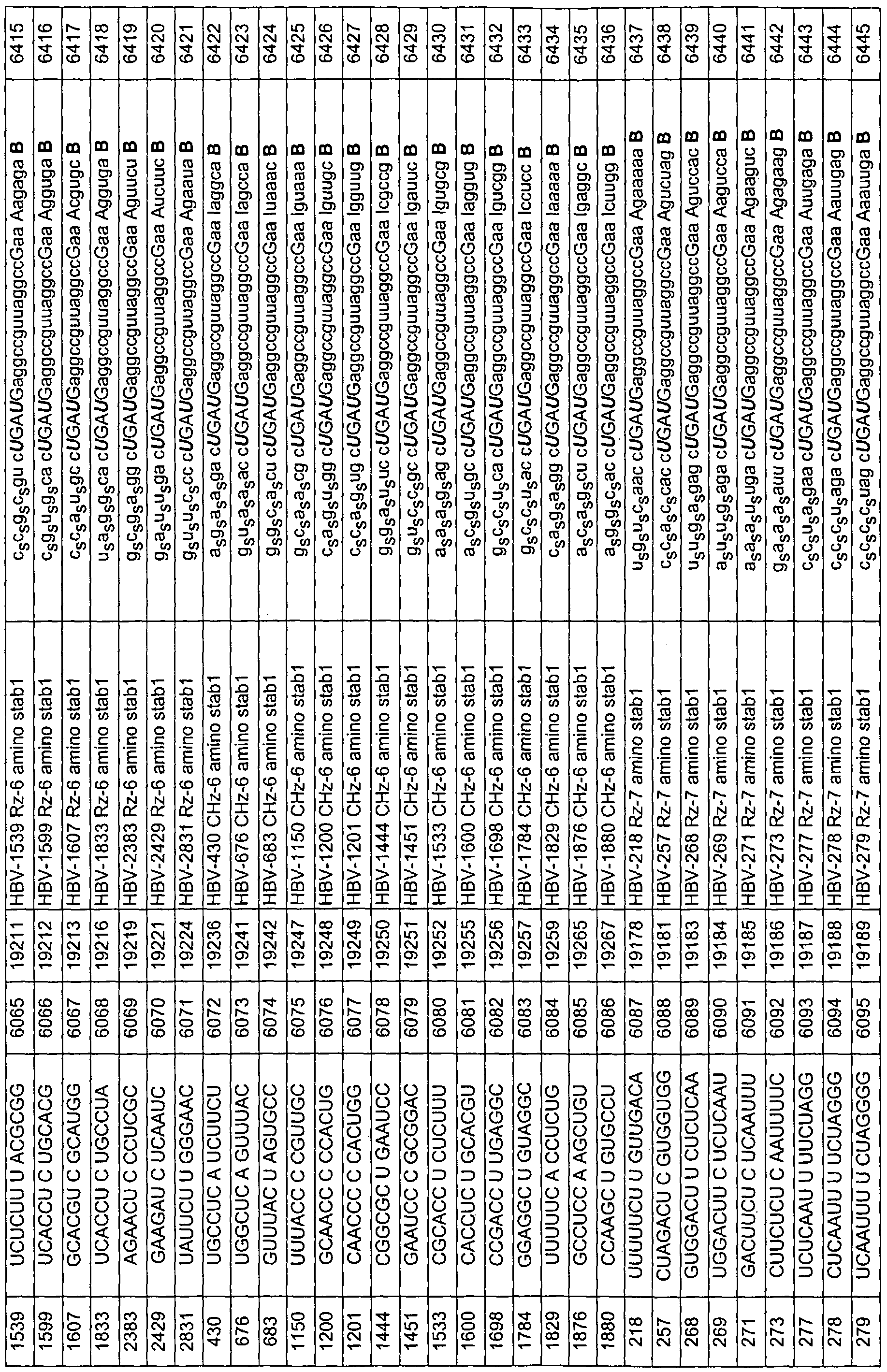
The nucleic acid-based inhibitors ofthe invention are added directly, or can be complexed with cationic lipids, packaged within liposomes, or otherwise delivered to target cells or tissues. The nucleic acid or nucleic acid complexes can be locally administered to relevant tissues ex vivo, or in vivo through injection, infusion pump or stent, with or without their incoφoration in biopolymers. In preferred embodiments, the enzymatic nucleic acid inhibitors comprise sequences, which are complementary to the subsfrate sequences in Tables 3-31, 33, 34, 36-43, 56, 58, 59, 62, 63. Examples of such enzymatic nucleic acid molecules also are shown in Tables 3-29, 31, 33, 34, 37-43, 56, 58, 59, 62, 63. Examples of such enzymatic nucleic acid molecules consist essentially of sequences defined in these tables. hi yet another embodiment, the invention features antisense nucleic acid molecules including sequences complementary to the substrate sequences shown in Tables 3-31, 33, 34, 36, 37-43, 56, 58, 59, 62, 63. Such nucleic acid molecules can include sequences as shown for the binding arms ofthe enzymatic nucleic acid molecules in Tables 3-29, 31, 33, 34, 37-43, 56, 58, 59, 62, 63. Similarly, triplex molecules can be provided targeted to the corresponding DNA target regions, and containing the DNA equivalent of a target sequence or a sequence complementary to the specified target (subsfrate) sequence. Typically, antisense molecules will be complementary to a target sequence along a single contiguous sequence ofthe antisense molecule. However, in certain embodiments, an antisense molecule may bind to subsfrate such that the subsfrate molecule forms a loop, and/or an antisense molecule may bind such that the antisense molecule forms a loop. Thus, the antisense molecule may be complementary to two (or even more) noncontiguous substrate sequences or two (or even more) non-contiguous sequence portions of an antisense molecule may be complementary to a target sequence or both. In another aspect, the invention provides mammalian cells containing one or more nucleic acid molecules and/or expression vectors of this invention. The one or more nucleic acid molecules may independently be targeted to the same or different sites.
By "consists essentially of is meant that the active nucleic acid molecule ofthe invention, for example, an enzymatic nucleic acid molecule, contains an enzymatic center or core equivalent to those in the examples, and binding arms able to bind mRNA such that cleavage at the target site occurs. Other sequences may be present which do not interfere with such cleavage. Thus, a core region may, for example, include one or more loop or stem-loop structures, which do not prevent enzymatic activity. "X" in the sequences in Tables 3, 4, 9, 10, 13, 14, 18, 19, 24, 25, 33, 34, 37, 38, 63 can be such a loop. A core sequence for a hammerhead ribozyme can be CUGAUGAG X CGAA where X=GCCGUUAGGC or other stem II region as specifically or generally known in the art. In another aspect ofthe invention, ribozymes or antisense molecules that interact with target RNA molecules and inhibit PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV (specifically PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV RNA) activity are expressed from transcription units inserted into DNA or RNA vectors. The recombinant vectors are preferably DNA plasmids or viral vectors. Ribozyme or antisense expressing viral vectors could be constructed based on, but not limited to, adeno-associated virus, retrovirus, adenovirus, or alphavirus. Preferably, the recombinant vectors capable of expressing the ribozymes or antisense are delivered as described above, and persist in target cells. Alternatively, viral vectors may be used that provide for transient expression of ribozymes or antisense. Such vectors might be repeatedly administered as necessary. Once expressed, the ribozymes or antisense bind to the target RNA and inhibit its function or expression. Delivery of ribozyme or antisense expressing vectors could be systemic, such as by intravenous or intramuscular administration, by administration to target cells ex -planted from the patient followed by reinfroduction into the patient, or by any other means that would allow for introduction into the desired target cell. Antisense DNA can be expressed via the use of a single stranded DNA intracellular expression vector.
By RNA is meant a molecule comprising at least one ribonucleotide residue. By "ribonucleotide" is meant a nucleotide with a hydroxyl group at the 2' position of a β-D- ribo-furanose moiety. By "vectors" is meant any nucleic acid- and/or viral-based technique used to deliver a desired nucleic acid.
By "patient" is meant an organism, which is a donor or recipient of explanted cells or the cells themselves. "Patient" also refers to an organism to which the nucleic acid molecules ofthe invention can be administered. Preferably, a patient is a mammal or mammalian cells. More preferably, a patient is a human or human cells. The nucleic acid molecules ofthe instant invention, individually, or in combination or in conjunction with other drugs, can be used to treat diseases or conditions discussed above. For example, to treat a disease or condition associated with PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV, the patient may be treated, or other appropriate cells may be treated, as is evident to those skilled in the art, individually or in combination with one or more drugs under conditions suitable for the treatment.
In a further embodiment, the described molecules, such as antisense or ribozymes, can be used in combination with other known treatments to treat conditions or diseases discussed above. For example, the described molecules could be used in combination with one or more known therapeutic agents to treat HBV infection, hepatitis, hepatocellular carcinoma, tumorigenesis, cirrhosis, liver failure, cancers including breast, ovarian, prostate, and esophogeal cancer, tumorigenesis, retinopathy, arthritis, psoriasis, female reproduction, restinosis, certain infectious diseases, transplant rejection and autoimmune disease such as multiple sclerosis, lupus, and AIDS, age related diseases such as macular degeneration and skin ulceration, Alzheimer's disease, dementia, diabetes, and/or obesity. In another preferred embodiment, the invention features nucleic acid-based inhibitors (e.g., enzymatic nucleic acid molecules (ribozymes), antisense nucleic acids, triplex DNA, antisense nucleic acids containing RNA cleaving chemical groups) and methods for their use to down regulate or inhibit the expression of RNA (e.g., PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and or HBV) capable of progression and/or maintenance of HBV infection, hepatitis, hepatocellular carcinoma, tumorigenesis, cirrhosis, liver failure, cancers including breast, ovarian, prostate, and esophogeal cancer, tumorigenesis, retinopathy, arthritis, psoriasis, female reproduction, restinosis, certain infectious diseases, fransplant rejection and autoimmune disease such as multiple sclerosis, lupus, and ADDS, age related diseases such as macular degeneration and skin ulceration, Alzheimer's disease, dementia, diabetes, and/or obesity.
In another preferred embodiment, the invention features nucleic acid-based techniques (e.g., enzymatic nucleic acid molecules (ribozymes), antisense nucleic acids, triplex DNA, antisense nucleic acids containing RNA cleaving chemical groups) and methods for their use to down regulate or inhibit the expression of PTP-IB, MetAP-2, BACE, ps- 1 , ps-2, HER2, PLN, TERT, and or HBV RNA expression.
By "comprising" is meant including, but not limited to, whatever follows the word "comprising". Thus, use ofthe term "comprising" indicates that the listed elements are required or mandatory, but that other elements are optional and may or may not be present. By "consisting of is meant including, and limited to, whatever follows the phrase "consisting of. Thus, the phrase "consisting of indicates that the listed elements are required or mandatory, and that no other elements may be present. By "consisting essentially of is meant including any elements listed after the phrase, and limited to other elements that do not interfere with or contribute to the activity or action specified in the disclosure for the listed elements. Thus, the phrase "consisting essentially of indicates that the listed elements are required or mandatory, but that other elements are optional and may or may not be present depending upon whether or not they affect the activity or action ofthe listed elements.
Other features and advantages ofthe invention will be apparent from the following description ofthe preferred embodiments thereof, and from the claims.
Description Of The Preferred Embodiments
The drawings will first briefly be described.
Drawings: Figure 1 shows the secondary structure model for seven different classes of enzymatic nucleic acid molecules. Arrow indicates the site of cleavage. indicate the target sequence. Lines interspersed with dots are meant to indicate tertiary interactions. - is meant to indicate base-paired interaction. Group I Intron: P1-P9.0 represent various stem-loop structures (Cech et al, 1994, Nature Struc. Bio., 1, 273). RNase P (M1RNA): EGS represents external guide sequence (Forster et al, 1990, Science, 249, 783; Pace et al, 1990, J. Biol. Chem., 265, 3587). Group II Intron: 5'SS means 5' splice site; 3'SS means 3 '-splice site; IBS means intron binding site; EBS means exon binding site (Pyle et
al, 1994, Biochemistry, 33, 2716). VS RNA: I- VI are meant to indicate six stem-loop structures; shaded regions are meant to indicate tertiary interaction (Collins, International PCT Publication No. WO 96/19577). HDV Ribozyme: I-IV are meant to indicate four stem-loop structures (Been et al, US Patent No. 5,625,047). Hammerhead Ribozyme: I- HI are meant to indicate three stem- loop structures; stems I-HI can be of any length and may be symmetrical or asymmetrical (Usman et al, 1996, Curr. Op. Struct. Bio., 1, 527). Hairpin Ribozyme: Helix 1, 4 and 5 can be of any length; Helix 2 is between 3 and 8 base-pairs long; Y is a pyrimidine; Helix 2 (H2) is provided with a least 4 base pairs (i.e., n is 1, 2, 3 or 4) and helix 5 can be optionally provided of length 2 or more bases (preferably 3 - 20 bases, i.e., m is from 1 - 20 or more). Helix 2 and helix 5 may be covalently linked by one or more bases (i.e., r is > 1 base). Helix 1, 4 or 5 may also be extended by 2 or more base pairs (e.g., - 20 base pairs) to stabilize the ribozyme structure, and preferably is a protein binding site. In each instance, each N and N' independently is any normal or modified base and each dash represents a potential base- pairing interaction. These nucleotides may be modified at the sugar, base or phosphate.
Complete base-pairing is not required in the helices, but is preferred. Helix 1 and 4 can be of any size (i.e., o and p is each independently from 0 to any number, e.g., 20) as long as some base-pairing is maintained. Essential bases are shown as specific bases in the structure, but those in the art will recognize that one or more may be modified chemically (abasic, base, sugar and/or phosphate modifications) or replaced with another base without significant effect. Helix 4 can be formed from two separate molecules, i.e., without a connecting loop. The connecting loop when present may be a ribonucleotide with or without modifications to its base, sugar or phosphate, "q" > is 2 bases. The connecting loop can also be replaced with a non-nucleotide linker molecule. H refers to bases A, U, or C. Y refers to pyrimidine bases. " " refers to a covalent bond. (Burke et al, 1996,
Nucleic Acids & Mol Biol, 10, 129; Chowrira et al, US Patent No. 5,631,359).
Figure 2 shows examples of chemically stabilized ribozyme motifs. HH Rz, represents hammerhead ribozyme motif (Usman et al, 1996, Curr. Op. Struct. Bio., 1, 527); NCH Rz represents the NCH ribozyme motif (described herein and in Ludwig & Sproat, International PCT Publication No. WO 98/58058); G-Cleaver, represents G- cleaver ribozyme motif (Kore et α/., 1998, Nucleic Acids Research, 26, 4116-4120). N or
n, represent independently a nucleotide which may be same or different and have complementarity to each other; rl, represents ribo-Inosine nucleotide; arrow indicates the site of cleavage within the target. Position 4 ofthe HH Rz and the NCH Rz is shown as having 2'-C-allyl modification, but those skilled in the art will recognize that this position can be modified with other modifications well known in the art, so long as such modifications do not significantly inhibit the activity ofthe ribozyme.
Figure 3 shows an example ofthe Amberzyme ribozyme motif that is chemically stabilized (see, for example, Beigelman et al, International PCT publication No. WO 99/55857; also referred to as Class I Motif). The Amberzyme motif is a class of enzymatic nucleic acid molecules that do not require the presence of a ribonucleotide (2'-OH) group for activity.
Figure 4 shows an example ofthe Zinzyme A ribozyme motif that is chemically stabilized (see, for example, International PCT publication No. WO 99/55857; also referred to as Class A Motif). The Zinzyme motif is a class of enzymatic nucleic acid molecules that do not require the presence of a ribonucleotide (2' -OH) group for activity. Figure 5 shows an example of a DNAzyme motif described by Santoro et al., 1997, PNAS, 94, 4262.
Figure 6 is a diagrammatic representation ofthe hammerhead ribozyme motif known in the art and the NCH motif. Stem JJ can be 2 base-pair long, preferably, 2, 3, 4, 5, 6, 7, 8, and 10 base-pairs long. Each N and N' is independently any base or non- nucleotide as used herein; X is adenosine, cytidine or uridine; Stem I-HI are meant to indicate three stem-loop structures; stems I-HI can be of any length and may be symmetrical or asymmetrical (Usman et al, 1996, Curr. Op. Struct. Bio., 1, 527); arrow indicates the site of cleavage in the target RNA; Rz refers to ribozyme; Loop II may be present or absent. If Loop II is present it is greater than or equal to three nucleotides, preferably four nucleotides. The Loop II sequence is preferably 5'-GAAA-3' or 5'- GUUA-3'.
Figure 7 shows examples of chemically stabilized ribozyme motifs. HH Rz, represents hammerhead ribozyme motif (Usman et al, 1996, Curr. Op. Struct. Bio., 1, 527); NCH-Inosine Rz represents the NCH ribozyme motif with riboinosine at 15.1 position; NCH-Xylo Rz represents the NCH ribozyme with xylo inosine at 15.1 position. N or n, represent independently a nucleotide which may be same or different and may have
complementarity to each other; ri, represents ribo-Inosine nucleotide; xl represent xylo- inosine; arrow indicates the site of cleavage within the target. Position 4 ofthe HH Rz and the NCH Rzs is shown as having 2'-C-allyl modification, but those skilled in the art will recognize that this position can be modified with other modifications well known in the art, so long as such modifications do not significantly inhibit the activity ofthe ribozyme. Figure 8 is a graphical representation of data showing inhibition of cell proliferation mediated by NCH and HH ribozymes targeted against HER2/new/ErbB2 gene. Untreated, refers to cells not treated with ribozymes; HH RZ refers to hammerhead ribozyme; NCX RZ refers to the NCH ribozymes ofthe invention; LA refers to catalytically inactive or attenuated ribozyme used as a confrol.
Figure 9 is a schematic diagram ofthe process for the synthesis of beta-D- xylofuranosyl hypoxantine 3 '-phosphoramidite.
Figure 10 displays a schematic representation of NTP synthesis using nucleoside substrates. Figure 11 shows a scheme for an in vitro selection method. A pool of nucleic acid molecules is generated with a random core region and one or more region(s) with a defined sequence. These nucleic acid molecules are bound to a column containing immobilized oligonucleotide with a defined sequence, where the defined sequence is complementary to region(s) of defined sequence of nucleic acid molecules in the pool. Those nucleic acid molecules capable of cleaving the immobilized oligonucleotide (target) in the column are isolated and converted to complementary DNA (cDNA), followed by transcription using NTPs to form a new nucleic acid pool.
Figure 12 shows a scheme for a two column in vitro selection method. A pool of nucleic acid molecules is generated with a random core and two flanking regions (region A and region B) with defined sequences. The pool is passed through a column which has immobilized oligonucleotides with regions A' and B' that are complementary to regions A and B ofthe nucleic acid molecules in the pool, respectively. The column is subjected to conditions sufficient to facilitate cleavage ofthe immobilized oligonucleotide target. The molecules in the pool that cleave the target (active molecules) have A' region ofthe target bound to their A region, whereas the B region is free. The column is washed to isolate the active molecules with the bound A' region ofthe target. This pool of active molecules may also contain some molecules that are not active to cleave the target (inactive
molecules) but have dissociated from the column. To separate the contaminating inactive molecules from the active molecules, the pool is passed through a second column (column 2) which contains immobilized oligonucleotides with the A' sequence but not the B' sequence. The inactive molecules will bind to column 2 but the active molecules will not bind to column 2 because their A region is occupied by the A' region ofthe target oligonucleotide from column 1. Column 2 is washed to isolate the active molecules for further processing as described in the scheme shown in Figure 11.
Figure 13 is a diagram of a novel 48 nucleotide enzymatic nucleic acid motif which was identified using in vitro methods described in the instant invention. The molecule shown is only exemplary. The 5' and 3' terminal nucleotides (referring to the nucleotides ofthe subsfrate binding arms rather than merely the single terminal nucleotide on the 5' and 3' ends) can be varied so long as those portions can base-pair with target substrate sequence. In addition, the guanosine (G) shown at the cleavage site ofthe subsfrate can be changed to other nucleotides so long as the change does not eliminate the ability of enzymatic nucleic acid molecules to cleave the target sequence. Substitutions in the nucleic acid molecule and/or in the subsfrate sequence can be readily tested, for example, as described herein.
Figure 14 is a schematic diagram of HCV luciferase assay used to demonstrate efficacy of class I enzymatic nucleic acid molecule motif. Figure 15 is a graph indicating the dose curve of an enzymatic nucleic acid molecule targeting site 146 on HCV RNA.
Figure 16 is a bar graph showing enzymatic nucleic acid molecules targeting 4 sites within the HCV RNA are able to reduce RNA levels in cells.
Figure 17 shows secondary structures and cleavage rates for characterized Class π enzymatic nucleic acid motifs.
Figure 18 is a diagram of a novel 35 nucleotide enzymatic nucleic acid motif which was identified using in vitro methods described in the instant invention. The molecule shown is only exemplary. The 5' and 3' terminal nucleotides (referring to the nucleotides ofthe subsfrate binding arms rather than merely the single terminal nucleotide on the 5' and 3' ends) can be varied so long as those portions can base-pair with target substrate sequence. In addition, the guanosine (G) shown at the cleavage site ofthe subsfrate can be changed to other nucleotides so long as the change does not eliminate the ability of
enzymatic nucleic acid molecules to cleave the target sequence. Substitutions in the nucleic acid molecule and/or in the substrate sequence can be readily tested, for example, as described herein.
Figure 19 is a bar graph showing subsfrate specificities for Class π (zinzyme) ribozymes.
Figure 20 is a bar graph showing Class II enzymatic nucleic acid molecules targeting 10 representative sites within the HER2 RNA in a cellular proliferation screen.
Figure 21 is a synthetic scheme outlining the synthesis of 5-[3- aminopropynyl(propyl)]uridine 5'-triphosphates and 4-imidazoleaceticacid conjugates. Figure 22 is a synthetic scheme outlining the synthesis of 5-[3-(N-4- imidazoleacetyl)aminopropynyl(propyl)]uridine 5 '-triphosphates.
Figure 23 is a synthetic scheme outlining the synthesis of carboxylate tethered uridine 5'-triphosphoates.
Figure 24 is a synthetic scheme outlining the synthesis of 5-(3-aminoalkyl) and 5- [3(N-succinyl)aminopropyl] functionalized cytidines.
Figure 25 is a diagram of a class I ribozyme stem truncation and loop replacement analysis.
Figure 26 is a diagram of class I ribozymes with truncated stem(s) and/or non- nucleotide linkers used in loop structures. Figure 27 is a diagram of "no-ribo" class LI ribozymes.
Figure 28 is a graph showing cleavage reactions with class II ribozymes under differing divalent metal concentrations.
Figure 29 is a diagram of differing class π ribozymes with varying ribo content and their relative rates of catalysis. Figure 30 is a graph showing class II ribozyme (zinzyme) mediated reduction of
HER2 RNA in SKBR3 breast carcinoma cells. Cells were treated with 100 nm, and 200 nm of zinzyme (RPI 18656) targeting site 972 of HER2 RNA and a corresponding scrambled attenuated confrol complexed with 2.5 μg/ml of lipid. Active zinzymes and scrambled attenuated controls were compared to untreated cells after 24 hours post treatment.
Figure 31 is a graph showing class II ribozyme (zinzyme) mediated dose response anti-prolferation assay in SKBR3 breast carcinoma cells. Cells were freated with 100 nm, and 200 nm of zinzyme (RPI 18656) targeting site 972 of HER2 RNA and a corresponding scrambled attenuated control complexed with 2.0 μg/ml of lipid. Active zinzymes and scrambled attenuated controls were compared to untreated cells after 24 hours post treatment.
Figure 32 is a graph which shows the dose dependent reduction of HER2 RNA in SKOV-3 cells treated with RPI 19293 from 0 to 100 nM with 5.0 μg/ml of cationic lipid.
Figure 33 is a graph which shows the dose dependent reduction of HER2 RNA and inhibition of cellular proliferation in SKBR-3 cells freated with RPI 19293 from 0 to 400 nM with 5.0 μg/ml of cationic lipid.
Figure 34 shows a non-limiting example ofthe replacement of a 2'-O-methyl 5'- CA-3' with a ribo G in the class LI (zinzyme) motif. The representative motif shown for the puφose ofthe figure is a "seven-ribo" zinzyme motif, however, the interchangeability of a G and a CA in the position shown in Figure 25 of the class II (zinzyme) motif extends to any combination of 2-O-methyl and ribo residues. For instance, a 2'-O-methyl G can replace the 2'-O-methyl 5'-CA-3' and vise versa.
Figure 35 is a graph which shows a screen of class II ribozymes (zinzymes) targeting site 972 of HER2 RNA which contain ribo-G reductions (RPI 19727 = no ribo, RPI 19728 = one ribo, RPI 19293 = two ribo, RPI 19729 = three ribo, RPI 19730 = four ribo, 19731 = five ribo, and RPI 19292 = seven ribo) for anti-proliferative activity in SKBR3 cells.
Figure 36 summarizes the results of functional group modification studies in which various nucleoside analogs were tested for activity in the NCH ribozyme motif. Kreι values describe the cleavage values of a given substituent at position 15.1 relative the Inosine at position 15.1 (1-15.1).
Figure 37 summarizes reported functional group modification studies performed at the A 15.1 residue in the A-15.1 »U-16.1 context of NUH cleaving ribozymes. Kreι values describe the cleavage values of a given substituent at position 15.1 relative the adenosine at position 15.1 (A-15.1).
Mechanism of action of Nucleic Acid Molecules ofthe Invention
Antisense: Antisense molecules may be modified or unmodified RNA, DNA, or mixed polymer oligonucleotides and primarily function by specifically binding to matching sequences resulting in inhibition of peptide synthesis (Wu-Pong, Nov 1994, BioPharm, 20-33). The antisense oligonucleotide binds to target RNA by Watson Crick base-pairing and blocks gene expression by preventing ribosomal translation ofthe bound sequences either by steric blocking or by activating RNase H enzyme. Antisense molecules may also alter protein synthesis by interfering with RNA processing or transport from the nucleus into the cytoplasm (Mukhopadhyay & Roth, 1996, Crit. Rev. in Oncogenesis 7, 151 - 190).
In addition, binding of single stranded DNA to RNA may result in nuclease degradation ofthe heteroduplex (Wu-Pong, supra; Crooke, supra). To date, the only backbone modified DNA chemistry which will act as substrates for RNase H are phosphorothioates, phosphorodithioates, and borontrifluoridates. Recently, it has been reported that 2'-arabino and 2'-fluoro arabino- containing oligos can also activate RNase H activity.
A number of antisense molecules have been described that utilize novel configurations of chemically modified nucleotides, secondary structure, and/or RNase H substrate domains (Woolf et al., International PCT Publication No. WO 98/13526; Thompson et al, International PCT Publication No. WO 99/54459 ; Hartmann et al, International PCT Publication No. WO 00/17346) all of these are incoφorated by reference herein in their entirety.
Antisense DNA can be used to target RNA by means of DNA-RNA interactions, thereby activating RNase H, which digests the target RNA in the duplex. Antisense DNA can be chemically synthesized or can be expressed via the use of a single stranded DNA intracellular expression vector or the equivalent thereof.
Triplex Forming Oligonucleotides (TFO): Single stranded DNA may be designed to bind to genomic DNA in a sequence specific manner. TFOs are comprised of pyrimidine- rich oligonucleotides which bind DNA helices through Hoogsteen Base-pairing (Wu-Pong, supra). The resulting triple helix composed ofthe DNA sense, DNA antisense, and TFO disrupts RNA synthesis by RNA polymerase. The TFO mechanism may result in gene expression or cell death since binding may be irreversible (Mukhopadhyay & Roth, supra)
2'-5' Oligoadenylates: The 2-5 A system is an interferon-mediated mechanism for RNA degradation found in higher vertebrates (Mifra et al, 1996, Proc Nat Acad Sci USA 93, 6780-6785). Two types of enzymes, 2-5 A synthetase and RNase L, are required for RNA cleavage. The 2-5 A synthetases require double stranded RNA to form 2'-5' oligoadenylates (2-5 A). 2-5A then acts as an allosteric effector for utilizing RNase L which has the ability to cleave single stranded RNA. The ability to form 2-5 A structures with double stranded RNA makes this system particularly useful for inhibition of viral replication.
(2'-5') oligoadenylate structures may be covalently linked to antisense molecules to form chimeric oligonucleotides capable of RNA cleavage (Torrence, supra). These molecules putatively bind and activate a 2-5 A dependent RNase, the oligonucleotide/enzyme complex then binds to a target RNA molecule which can then be cleaved by the RNase enzyme. The covalent attachment of 2'-5' oligoadenylate structures is not limited to antisense applications, and can be further elaborated to include attachment to nucleic acid molecules ofthe instant invention.
Enzymatic Nucleic Acid: Seven basic varieties of naturally-occurring enzymatic RNAs are presently known. In addition, several in vitro selection (evolution) strategies (Orgel, 1979, Proc. R. Soc. London, B 205, 435) have been used to evolve new nucleic acid catalysts capable of catalyzing cleavage and ligation of phosphodi ester linkages (Joyce, 1989, Gene, 82, 83-87; Beaudry et al, 1992, Science 257, 635-641 ; Joyce, 1992, Scientific American 267, 90-97; Breaker et al, 1994, TIBTECH 12, 268; Bartel et al, 1993, Science 261:1411-1418; Szostak, 1993, TIBS 17, 89-93; Kumar et al, 1995, FASEB J., 9, 1183; Breaker, 1996, Curr. Op. Biotech., 7, 442; Santoro et al, 1997, Proc. Natl. Acad. Sci., 94, 4262; Tang et al, 1997, RNA 3, 914; Nakamaye & Eckstein, 1994, supra; Long & Uhlenbeck, 1994, supra; Ishizaka et al., 1995, supra; Vaish et al, 1997, Biochemistry 36, 6495; all of these are incoφorated by reference herein). Each can catalyze a series of reactions including the hydrolysis of phosphodiester bonds in trans (and thus can cleave other RNA molecules) under physiological conditions.
In general, enzymatic nucleic acids act by first binding to a target RNA. Such binding occurs through the target binding portion of an enzymatic nucleic acid which is held in close proximity to an enzymatic portion ofthe molecule that acts to cleave the target RNA. Thus, the enzymatic nucleic acid first recognizes and then binds a target
RNA through complementary base-pairing, and once bound to the correct site, acts enzymatically to cut the target RNA. Strategic cleavage of such a target RNA will destroy its ability to direct synthesis of an encoded protein. After an enzymatic nucleic acid has bound and cleaved its RNA target, it is released from that RNA to search for another target and can repeatedly bind and cleave new targets.
Nucleic acid molecules of this invention will block to some extent PTP-IB, MetAP- 2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV protein expression and can be used to treat disease or diagnose disease associated with the levels of PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV. The enzymatic nature of a ribozyme has significant advantages, such as the concentration of ribozyme necessary to affect a therapeutic treatment is low. This advantage reflects the ability ofthe ribozyme to act enzymatically. Thus, a single ribozyme molecule is able to cleave many molecules of target RNA. In addition, the ribozyme is a highly specific inhibitor, with the specificity of inhibition depending not only on the base-pairing mechanism of binding to the target RNA, but also on the mechanism of target RNA cleavage. Single mismatches, or base-substitutions, near the site of cleavage can be chosen to completely eliminate catalytic activity of a ribozyme.
Nucleic acid molecules having an endonuclease enzymatic activity are able to repeatedly cleave other separate RNA molecules in a nucleotide base sequence-specific manner. Such enzymatic nucleic acid molecules can be targeted to virtually any RNA transcript, and achieve efficient cleavage in vitro (Zaug et al, 324, Nature, 429 1986 ; Uhlenbeck, 1987 Nature, 328, 596; Kim et al., 84 Proc. Natl. Acad. Sci. USA, 8788, 1987; Dreyfus, 1988, Einstein Quart. J. Bio. Med., 6, 92; Haseloff and Geriach, 334 Nature, 585, 1988; Cech, 260 JAMA, 3030, 1988; Jefferies et al., 17 Nucleic Acids Research, 1371, 1989; and Santoro et al, 1997 supra).
Because of their sequence specificity, trans-cleaving ribozymes show promise as therapeutic agents for human disease (Usman & McSwiggen, 1995 Ann. Rep. Med. Chem. 30, 285-294; Christoffersen and Marr, 1995 J. Med. Chem. 38, 2023-2037). Ribozymes can be designed to cleave specific RNA targets within the background of cellular RNA. Such a cleavage event renders the RNA non-functional and abrogates protein expression from that RNA. In this manner, synthesis of a protein associated with a disease state can be selectively inhibited (Warashina et al, 1999, Chemistry and Biology, 6, 237-250.
The nucleic acid molecules ofthe instant invention are also referred to as GeneBloc™ reagents, which are essentially nucleic acid molecules (e.g.; ribozymes, antisense) capable of down-regulating gene expression.
Target sites
Targets for useful ribozymes and antisense nucleic acids can be determined as disclosed in Draper et al, WO 93/23569; Sullivan et al, WO 93/23057; Thompson et al, WO 94/02595; Draper et al, WO 95/04818; McSwiggen et al, US Patent No. 5,525,468, and all hereby incoφorated in their entireties by reference herein. Other examples include the following PCT applications, which concern inactivation of expression of disease- related genes: WO 95/23225, WO 95/13380, WO 94/02595, all incoφorated by reference herein. Rather than repeat the guidance provided in those documents here, below are provided specific examples of such methods, not limiting to those in the art. Ribozymes and antisense to such targets are designed as described in those applications and synthesized to be tested in vitro and in vivo, as also described. The sequence of human PTP-IB, MetAP-2, BACE, ps-1, ps-2, HER2, PLN, TERT, and/or HBV RNAs (for example, GenBank accession Nos. (PTP-IB,. NM_002827), (MetAP-2, U29607), (BACE, AF190725), (ps-1, L76517), (ps-2, L43964), (HER2/c-erb2/neu, X03363), (PLN, NM_002667), (TERT, NM_003219) and (HBV, AF100308.1, HBV strain 2-18; additionally, other HBV strains can be screened by one skilled in the art, see Table 35 for other possible strains) were screened for optimal enzymatic nucleic acid and antisense target sites using a computer-folding algorithm. Antisense, hammerhead, DNAzyme, NCH (Inozyme), amberzyme, zinzyme or G-Cleaver ribozyme binding/cleavage sites were identified. These sites are shown in Tables 3-29, 31, 33, 34, 37-43, 56, 58, 59, 62, 63 (all sequences are 5' to 3' in the tables; X can be any base-paired sequence, the actual sequence is not relevant here). The nucleotide base position is noted in the Tables as that site to be cleaved by the designated type of enzymatic nucleic acid molecule. Table 36 shows substrate positions selected from Renbo et al, 1987, Sci. Sin., 30, 507, used in Draper, US patent No. 6,017,756 entitled "METHOD AND REAGENT FOR INHIBITING HEPATITIS B VIRUS REPLICATION" and Draper et al., International PCT publication No. WO 93/23569, filed April 29, 1993, entitled "METHOD AND REAGENT FOR INHIBITING VIRAL REPLICATION". While human sequences can be
screened and enzymatic nucleic acid molecule and/or antisense thereafter designed, as discussed in Stinchcomb et al, WO 95/23225, mouse targeted ribozymes may be useful to test efficacy of action ofthe enzymatic nucleic acid molecule and/or antisense prior to testing in humans. Antisense, hammerhead, DNAzyme, NCH (Inozyme), amberzyme, zinzyme or G-
Cleaver ribozyme binding/cleavage sites were identified, as discussed above. The nucleic acid molecules were individually analyzed by computer folding (Jaeger et al, 1989 Proc. Natl. Acad. Sci. USA, 86, 7706) to assess whether the sequences fold into the appropriate secondary structure. Those nucleic acid molecules with unfavorable intramolecular interactions such as between the binding arms and the catalytic core were eliminated from consideration. Varying binding arm lengths can be chosen to optimize activity.
Antisense, hammerhead, DNAzyme, NCH, amberzyme, zinzyme or G-Cleaver ribozyme binding/cleavage sites were identified and were designed to anneal to various sites in the RNA target. The binding arms are complementary to the target site sequences described above. The nucleic acid molecules were chemically synthesized. The method of synthesis used follows the procedure for normal DNA/RNA synthesis as described below and in Usman et al, 1987 J. Am. Chem. Soc, 109, 7845; Scaringe et al, 1990 Nucleic Acids Res., 18, 5433; Wincott et al, 1995 Nucleic Acids Res. 23, 2677-2684; and Caruthers et al, 1992, Methods in Enzymology 211,3-19.
Synthesis of Nucleic acid Molecules
Synthesis of nucleic acids greater than 100 nucleotides in length is difficult using automated methods, and the therapeutic cost of such molecules is prohibitive. In this invention, small nucleic acid motifs ("small refers to nucleic acid motifs no more than 100 nucleotides in length, preferably no more than 80 nucleotides in length, and most preferably no more than 50 nucleotides in length; e.g., antisense oligonucleotides, hammerhead or the NCH ribozymes) are preferably used for exogenous delivery. The simple structure of these molecules increases the ability ofthe nucleic acid to invade targeted regions of RNA structure. Exemplary molecules ofthe instant invention are chemically synthesized, and others can similarly be synthesized.
Oligonucleotides (e.g.; antisense GeneBlocs) are synthesized using protocols known in the art as described in Caruthers et al, 1992, Methods in Enzymology 211, 3-19, Thompson et al, Intemational PCT Publication No. WO 99/54459, Wincott et al, 1995, Nucleic Acids Res. 23, 2677-2684, Wincott et al, 1997, Methods Mol. Bio., 74, 59, Brennan et al, 1998, Biotechnol Bioeng., 61, 33-45, and Brennan, US patent No.
6,001 ,311. All of these references are incoφorated herein by reference. The synthesis of oligonucleotides makes use of common nucleic acid protecting and coupling groups, such as dimethoxytrityl at the 5'-end, and phosphoramidites at the 3'-end. In a non-limiting example, small scale syntheses are conducted on a 394 Applied Biosystems, Inc. synthesizer using a 0.2 μmol scale protocol with a 2.5 min coupling step for 2'-O- methylated nucleotides and a 45 sec coupling step for 2'-deoxy nucleotides. Table II outlines the amounts and the contact times ofthe reagents used in the synthesis cycle. Alternatively, syntheses at the 0.2 μmol scale can be performed on a 96-well plate synthesizer, such as the instrument produced by Protogene (Palo Alto, CA) with minimal modification to the cycle. A 33-fold excess (60 μL of 0.11 M = 6.6 μmol) of 2'-O-methyl phosphoramidite and a 105-fold excess of S-ethyl tefrazole (60 μL of 0.25 M = 15 μmol) can be used in each coupling cycle of 2'-O-methyl residues relative to polymer-bound 5'- hydroxyl. A 22-fold excess (40 μL of 0.11 M = 4.4 μmol) of deoxy phosphoramidite and a 70-fold excess of S-ethyl tefrazole (40 μL of 0.25 M = 10 μmol) can be used in each coupling cycle of deoxy residues relative to polymer-bound 5'-hydroxyl. Average coupling yields on the 394 Applied Biosystems, Inc. synthesizer, determined by colorimetric quantitation ofthe trityl fractions, are typically 97.5-99%. Other oligonucleotide synthesis reagents for the 394 Applied Biosystems, Inc. synthesizer include the following: detritylation solution is 3% TCA in methylene chloride (ABI); capping is performed with 16% N-methyl imidazole in THF (ABI) and 10% acetic anhydride/10% 2,6-lutidine in THF (ABI); and oxidation solution is 16.9 mM I2, 49 mM pyridine, 9% water in THF (PERSEPTIVE™). Burdick & Jackson Synthesis Grade acetonitrile is used directly from the reagent bottle. S-Ethyltefrazole solution (0.25 M in acetonitrile) is made up from the solid obtained from American International Chemical, Inc. Alternately, for the introduction of phosphorothioate linkages, Beaucage reagent (3H- l,2-Benzodithiol-3-one 1,1 -dioxide, 0.05 M in acetonitrile) is used.
Deprotection ofthe antisense oligonucleotides is performed as follows: the polymer- bound trityl-on oligoribonucleotide is transferred to a 4 mL glass screw top vial and suspended in a solution of 40% aq. methylamine (1 mL) at 65 °C for 10 min. After cooling to -20 °C, the supematant is removed from the polymer support. The support is washed three times with 1.0 mL of EtOH:MeCN:H2O/3 : 1 : 1 , vortexed and the supernatant is then added to the first supernatant. The combined supernatants, containing the oligoribonucleotide, are dried to a white powder.
The method of synthesis used for normal RNA including certain enzymatic nucleic acid molecules follows the procedure as described in Usman et al, 1987, J. Am. Chem. Soc, 109, 7845; Scaringe et al, 1990, Nucleic Acids Res., 18, 5433; and Wincott et al, 1995, Nucleic Acids Res. 23, 2677-2684 Wincott et al, 1997, Methods Mol. Bio., 74, 59, and makes use of common nucleic acid protecting and coupling groups, such as dimethoxytrityl at the 5'-end, and phosphoramidites at the 3'-end. In a non-limiting example, small scale syntheses are conducted on a 394 Applied Biosystems, Inc. synthesizer using a 0.2 μmol scale protocol with a 7.5 min coupling step for alkylsilyl protected nucleotides and a 2.5 min coupling step for 2'-O-methylated nucleotides. Table II outlines the amounts and the contact times ofthe reagents used in the synthesis cycle. Alternatively, syntheses at the 0.2 μmol scale can be done on a 96-well plate synthesizer, such as the instrument produced by Protogene (Palo Alto, CA) with minimal modification to the cycle. A 33-fold excess (60 μL of 0.11 M = 6.6 μmol) of 2'-O-methyl phosphoramidite and a 75-fold excess of S-ethyl tefrazole (60 μL of 0.25 M = 15 μmol) can be used in each coupling cycle of 2'-O-methyl residues relative to polymer-bound 5'- hydroxyl. A 66-fold excess (120 μL of 0.11 M = 13.2 μmol) of alkylsilyl (ribo) protected phosphoramidite and a 150-fold excess of S-ethyl tefrazole (120 μL of 0.25 M = 30 μmol) can be used in each coupling cycle of ribo residues relative to polymer-bound 5'-hydroxyl. Average coupling yields on the 394 Applied Biosystems, Inc. synthesizer, determined by colorimetric quantitation ofthe trityl fractions, are typically 97.5-99%. Other oligonucleotide synthesis reagents for the 394 Applied Biosystems, Inc. synthesizer include the following: detritylation solution is 3% TCA in methylene chloride (ABI); capping is performed with 16% N-methyl imidazole in THF (ABI) and 10% acetic anhydride/10%. 2,6-lutidine in THF (ABI); oxidation solution is 16.9 mM I2, 49 mM pyridine, 9% water in THF (PERSEPTIVE™). Burdick & Jackson Synthesis Grade
acetonitrile is used directly from the reagent bottle. S-Ethyltetrazole solution (0.25 M in acetonitrile) is made up from the solid obtained from American International Chemical, Inc. Alternately, for the introduction of phosphorothioate linkages, Beaucage reagent (3H- l,2-Benzodithiol-3-one l,l-dioxide0.05 M in acetonitrile) is used. Deprotection ofthe RNA is performed using either a two-pot or one-pot protocol.
For the two-pot protocol, the polymer-bound trityl-on oligoribonucleotide is transferred to a 4 mL glass screw top vial and suspended in a solution of 40% aq. methylamine (1 mL) at 65 °C for 10 min. After cooling to -20 °C, the supernatant is removed from the polymer support. The support is washed three times with 1.0 mL of EtOH:MeCN:H2O/3 : 1 : 1 , vortexed and the supernatant is then added to the first supematant. The combined supernatants, containing the oligoribonucleotide, are dried to a white powder. The base deprotected oligoribonucleotide is resuspended in anhydrous TEA/HF/NMP solution (300 μL of a solution of 1.5 mL N-methylpyrrolidinone, 750 μL TEA and 1 mL TEA»3HF to provide a 1.4 M HF concentration) and heated to 65 °C. After 1.5 h, the oligomer is quenched with 1.5 M NH4HCO3.
Alternatively, for the one-pot protocol, the polymer-bound trityl-on oligoribonucleotide is transferred to a 4 mL glass screw top vial and suspended in a solution of 33% ethanolic methylamine/DMSO: 1/1 (0.8 mL) at 65 °C for 15 min. The vial is brought to r.t. TEA*3HF (0.1 mL) is added and the vial is heated at 65 °C for 15 min. The sample is cooled at -20 °C and then quenched with 1.5 M NH4HCO3.
For purification ofthe trityl-on oligomers, the quenched NH4HCO3 solution is loaded onto a C-18 containing cartridge that had been prewashed with acetonitrile followed by 50 mM TEAA. After washing the loaded cartridge with water, the RNA is detritylated with 0.5% TFA for 13 min. The cartridge is then washed again with water, salt exchanged with 1 M NaCl and washed with water again. The oligonucleotide is then eluted with 30% acetonitrile.
Inactive hammerhead ribozymes or binding attenuated confrol (B AC) oligonucleotides) are synthesized by substituting a U for G5 and a U for A14 (numbering from Hertel, K. J., et al, 1992, Nucleic Acids Res.. 20, 3252). Similarly, one or more nucleotide substitutions can be introduced in other enzymatic nucleic acid molecules to inactivate the molecule and such molecules can serve as a negative confrol.
The average stepwise coupling yields are typically >98% (Wincott et al, 1995 Nucleic Acids Res. 23, 2677-2684). Those of ordinary skill in the art will recognize that the scale of synthesis can be adapted to be larger or smaller than the example described above including but not limited to 96-well format, all that is important is the ratio of chemicals used in the reaction.
Alternatively, the nucleic acid molecules ofthe present invention can be synthesized separately and joined together post-synthetically, for example, by ligation (Moore et al, 1992, Science 256, 9923; Draper et al, International PCT publication No. WO 93/23569; Shabarova et al, 1991, Nucleic Acids Research 19, 4247; Bellon et al, 1997, Nucleosides & Nucleotides, 16, 951; Bellon et al, 1997, Bioconjugate Chem. 8, 204).
The nucleic acid molecules ofthe present invention are modified extensively to enhance stability by modification with nuclease resistant groups, for example, 2'-amino, 2'- C-allyl, 2'-flouro, 2'-O-methyl, 2'-H (for a review see Usman and Cedergren, 1992, TIBS 17, 34; Usman et al, 1994, Nucleic Acids Symp. Ser. 31, 163). Ribozymes are purified by gel elecfrophoresis using general methods or are purified by high pressure liquid chromatography (HPLC; see Wincott et al, supra, the totality of which is hereby incoφorated herein by reference) and are re-suspended in water.
The sequences ofthe ribozymes and antisense constructs that are chemically synthesized, useful in this study, are shown in Tables 3-31, 33, 34, 37-43, 56, 58, 59, 62, 63. Those in the art will recognize that these sequences are representative only of many more such sequences where the enzymatic portion ofthe ribozyme (all but the binding arms) is altered to affect activity. The ribozyme and antisense construct sequences listed in Tables 3-31, 33, 34, 37-43, 56, 58, 59, 62, 63 maybe formed of ribonucleotides or other nucleotides or non-nucleotides. Such ribozymes with enzymatic activity are equivalent to the ribozymes described specifically in the Tables.
Optimizing Activity ofthe nucleic acid molecule ofthe invention.
Chemically synthesizing nucleic acid molecules with modifications (base, sugar and/or phosphate) that prevent their degradation by serum ribonucleases may increase their potency (see e.g., Eckstein et al, International Publication No. WO 92/07065; Perrault et al, 1990 Nature 344, 565; Pieken et al., 1991, Science 253, 314; Usman and Cedergren, 1992, Trends in Biochem. Sci. 17, 334; Usman et al, International Publication No. WO 93/15187; Rossi et al, International Publication No. WO 91/03162; Sproat, US Patent No.
5,334,711 ; and Burgin et al, supra; all of these describe various chemical modifications that can be made to the base, phosphate and/or sugar moieties ofthe nucleic acid molecules herein and are all hereby incoφorated by reference herein). Modifications which enhance their efficacy in cells, and removal of bases from nucleic acid molecules to shorten oligonucleotide synthesis times and reduce chemical requirements are desired. There are several examples in the art describing sugar, base and phosphate modifications that can be introduced into nucleic acid molecules (e.g., enzymatic nucleic acid molecules) without significantly effecting catalysis and with significant enhancement in their nuclease stability and efficacy. Enzymatic nucleic acid molecules are modified to enhance stability and/or enhance catalytic activity by modification with nuclease resistant groups, for example, 2'-amino, 2"-C-allyl, 2'-fluoro, 2'-O-methyl, 2'-O-allyl, 2'-H, nucleotide base modifications (for a review see Usman and Cedergren, 1992 TIBS 17, 34; Usman et al, 1994 Nucleic Acids Symp. Ser. 31, 163; Burgin et al, 1996 Biochemistry 35, 14090). Sugar modification of enzymatic nucleic acid molecules have been extensively described in the art (see Eckstein et al, International Publication PCT No. WΟ 92/07065; Perrault et al. Nature 1990, 344, 565-568; Pieken et al. Science 1991, 253, 314-317; Usman and Cedergren, Trends in Biochem. Sci. 1992, 17, 334-339; Usman et al. International Publication PCT No. WO 93/15187; Sproat, US Patent No. 5,334,711 and Beigelman et al, 1995 J. Biol. Chem. 270, 25702; all ofthe references are hereby incoφorated in their totality by reference herein). Such publications describe general methods and strategies to determine the location of incoφoration of sugar, base and/or phosphate modifications and the like into enzymatic nucleic acid molecules without inhibiting catalysis, and are incoφorated by reference herein. The 2'-position ofthe sugar in a nucleotide present in the nucleic acid molecules ofthe instant invention which tolerates substitution is selected from the group comprising -H, -OH, -COOH, -CONH , - CONHR1, -CONR'R2, -NΉ^ -NHR'. -NR'R2, -NHCOR1, -SH, SR1, -F, -ONH2, - ONHR1, -ONR'R2, -NHOH, -NHOR1, -NR2OH, -NR2OR1, substituted or unsubstituted CI-CΪO straight chain or branched alkyl, substituted or unsubstituted C2-Cιo straight chain or branched alkenyl, substituted or unsubstituted C2-C10 straight chain or branched alkynyl, substituted or unsubstituted Cr o straight chain or branched alkoxy, substituted or unsubstituted C2-C10 straight chain or branched alkenyloxy, and substituted or unsubstituted C2-C10 straight chain or branched alkynyloxy. The substituents for sugar 2'
position preferably are independently halogen, cyano, amino, carboxy, ester, ether, carboxamide, hydroxy, or mercapto. R1 and R2 can be substituted or unsubstituted alkyl, alkenyl, or alkynyl groups, where the substituents are independently halogen, cyano, amino, carboxy, ester, ether, carboxamide, hydroxy, or mercapto. In view of such teachings, similar modifications can be used as described herein to modify the nucleic acid molecules ofthe instant invention. Such publications describe general methods and strategies to determine the location of incoφoration of sugar, base and/or phosphate modifications and the like into ribozymes without inhibiting catalysis, and are incoφorated by reference herein. In view of such teachings, similar modifications can be used as described herein to modify the nucleic acid molecules ofthe instant invention.
Some ofthe non-limiting examples of base modifications that can be introduced into enzymatic nucleic acids without significantly effecting their catalytic activity include, inosine, purine, pyridin-4-one, pyridin-2-one, phenyl, pseudouracil, 2, 4, 6-trimethoxy benzene, 3-methyluracil, dihydrouridine, naphthyl, aminophenyl, 5-alkylcytidines (e.g., 5-methylcytidine), 5-alkyluridines (e.g., ribothymidine), 5-halouridine (e.g., 5-bromouridine) or 6-azapyrimidines or 6-alkylpyrimidines (e.g. 6-methyluridine) and others (Burgin et al, 1996, Biochemistry, 35, 14090). By "modified bases" in this aspect is meant nucleotide bases other than adenine, guanine, cytosine and uracil at 1' position or their equivalents; such bases may be used within the catalytic core ofthe enzyme and/or in the substrate-binding regions.
The nucleic acid bases can be hypoxanthin-9-yl, or a functional equivalent thereof, in position15 'of the ribozyme; the base at other positions may be guanin-9-yl, hypoxanthin-9-yl or 7-deazaguanin-9-yl in positions 5, 8 and 12 in the ribozyme; adenin-9- yl, 2,6-diaminopurin-9-yl, purin-9-yl or 7-deaza adenin-9-yl in positions 6, 9, 13 and 14; uracil-1-yl, uracil-5-yl, thymin-1-yl or 5-propynyluracil-l-yl in position 4; cytosin-1-yl, 5- methylcytosin-1-yl or 5-propynylcytosin-l-yl in position 3; and adenin-9-yl, cytosin-1-yl, guanin-9-yl, uracil-1-yl, uracil-5-yl, hypoxanthin-9-yl, thymin-1-yl, 5-methylcytosin-l-yl, 2,6-diaminopurin-9-yl, purin-9-yl, 7-deaza adenin-9-yl, 7-deazaguanin-9-yl, 5- propynylcytosin-1-yl, 5-propynyluracil-l-yl, isoguanin-9-yl, 2-aminopurin-9-yl, 6- methyluracil-1-yl, 4-thiouracil-l-yl, 2-pyrimidone-l-yl, quinazoline-2,4-dione-l-yl, xanthin-9-yl, N2-dimethylguanin-9-yl, or a functional equivalent thereof in position 7. The
base at position 15.1 is preferably hypoxanthin-9-yl or an analog where no hydrogen bond can form between any group at the 2 position ofthe base and the 2-oxo group of C16'1. Preferably, B is not guanin-9-yl in position 15.1.
In particular, the invention features modified ribozymes having a base substitution selected from pyridin-4-one, pyridin-2-one, phenyl, pseudouracil, 2, 4, 6-trimethoxy benzene, 3-methyluracil, dihydrouracil, naphthyl, 6-methyl-uracil and aminophenyl. While chemical modification of oligonucleotide intemucleotide linkages with phosphorothioate, phosphorothioate, and/or 5'-methylphosphonate linkages improves stability, too many of these modifications may cause some toxicity. Therefore, when designing nucleic acid molecules, the amount of these intemucleotide linkages should be minimized. The reduction in the concentration of these linkages should lower toxicity resulting in increased efficacy and higher specificity of these molecules.
Nucleic acid molecules having chemical modifications which maintain or enhance activity are provided. Such nucleic acid molecules are also generally more resistant to nucleases than unmodified nucleic acid. Thus, in a cell and/or in vivo the activity may not be significantly lowered. Therapeutic nucleic acid molecules delivered exogenously must optimally be stable within cells until translation ofthe target RNA has been inhibited long enough to reduce the levels ofthe undesirable protein. This period of time varies between hours to days depending upon the disease state. Clearly, nucleic acid molecules must be resistant to nucleases in order to function as effective infracellular therapeutic agents.
Improvements in the chemical synthesis of RNA and DNA (Wincott et al, 1995 Nucleic Acids Res. 23, 2677; Caruthers et al, 1992, Methods in Enzymology 211,3-19 (all are incoφorated by reference herein) have expanded the ability to modify nucleic acid molecules by introducing nucleotide modifications to enhance their nuclease stability as described above.
Use of these the nucleic acid-based molecules ofthe invention will lead to better treatment ofthe disease progression by affording the possibility of combination therapies (e.g., multiple antisense or enzymatic nucleic acid molecules targeted to different genes, nucleic acid molecules coupled with known small molecule inhibitors, or intermittent treatment with combinations of molecules (including different motifs) and/or other chemical or biological molecules). The treatment of patients with nucleic acid molecules may also include combinations of different types of nucleic acid molecules.
Therapeutic nucleic acid molecules (e.g., enzymatic nucleic acid molecules and antisense nucleic acid molecules) delivered exogenously must optimally be stable within cells until translation ofthe target RNA has been inhibited long enough to reduce the levels ofthe undesirable protein. This period of time varies between hours to days depending upon the disease state. Clearly, these nucleic acid molecules must be resistant to nucleases in order to function as effective intracellular therapeutic agents. Improvements in the chemical synthesis of nucleic acid molecules described in the instant invention and in the art have expanded the ability to modify nucleic acid molecules by introducing nucleotide modifications to enhance their nuclease stability as described above.
By "enhanced enzymatic activity" is meant to include activity measured in cells and/or in vivo where the activity is a reflection of both catalytic activity and ribozyme stability. In this invention, the product of these properties is increased or not significantly (less than 10- fold) decreased in vivo compared to an all RNA ribozyme or all DNA enzyme.
In yet another preferred embodiment, nucleic acid catalysts having chemical modifications which maintain or enhance enzymatic activity are provided. Such nucleic acid catalysts are also generally more resistant to nucleases than unmodified nucleic acid. Thus, in a cell and/or in vivo the activity may not be significantly lowered. As exemplified herein such ribozymes are useful in a cell and/or in vivo even if activity over all is reduced 10 fold (Burgin et al, 1996, Biochemistry, 35, 14090). Such ribozymes herein are said to "maintain" the enzymatic activity of an all RNA ribozyme.
In another aspect the nucleic acid molecules comprise a 5' and/or a 3'- cap structure. By "cap structure" is meant chemical modifications, which have been incoφorated at either terminus ofthe oligonucleotide (see, for example, Wincott et al, WO 97/26270, incoφorated by reference herein). These terminal modifications protect the nucleic acid molecule from exonuclease degradation, and may help in delivery and/or localization within a cell. The cap may be present at the 5'-terminus (5'-cap) or at the 3'-terminal (3'- cap) or may be present on both termini. In non-limiting examples: the 5 '-cap is selected from the group comprising inverted abasic residue (moiety); 4',5'-methylene nucleotide; 1- (beta-D-erythrofuranosyl) nucleotide, 4'-thio nucleotide; carbocyclic nucleotide; 1,5-
anhydrohexitol nucleotide; L-nucleotides; alpha-nucleotides; modified base nucleotide; phosphorodithioate linkage; tλreo-pentofuranosyl nucleotide; acyclic 3',4'-seco nucleotide; acyclic 3,4-dihydroxybutyl nucleotide; acyclic 3,5-dihydroxypentyl nucleotide, 3'-3'- inverted nucleotide moiety; 3'-3'-inverted abasic moiety; 3'-2'-inverted nucleotide moiety; 3'-2'-inverted abasic moiety; 1,4-butanediol phosphate; 3'-phosphoramidate; hexylphosphate; aminohexyl phosphate; 3'-phosphate; 3'-phosphorothioate; phosphorodithioate; or bridging or non-bridging methylphosphonate moiety (for more details, see Wincott et al, International PCT publication No. WO 97/26270, incoφorated by reference herein). Ln yet another preferred embodiment, the 3 '-cap is selected from a group comprising, 4',5'-methylene nucleotide; l-(beta-D-erythrofuranosyl) nucleotide; 4'-thio nucleotide, carbocyclic nucleotide; 5'-amino-alkyl phosphate; l,3-diamino-2 -propyl phosphate; 3-aminopropyl phosphate; 6-aminohexyl phosphate; 1,2-aminododecyl phosphate; hydroxypropyl phosphate; 1,5-anhydrohexitol nucleotide; L-nucleotide; alpha- nucleotide; modified base nucleotide; phosphorodithioate; t/zreo-pentofuranosyl nucleotide; acyclic 3',4'-seco nucleotide; 3,4-dihydroxybutyl nucleotide; 3,5- dihydroxypentyl nucleotide, 5'-5 '-inverted nucleotide moiety; 5'-5'-inverted abasic moiety; 5'-phosphoramidate; 5'-phosphorothioate; 1,4-butanediol phosphate; 5'-amino; bridging and/or non-bridging 5'-phosphoramidate, phosphorothioate and/or phosphorodithioate, bridging or non bridging methylphosphonate and 5'-mercapto moieties (for more details see Beaucage and Iyer, 1993, Tetrahedron 49, 1925; incoφorated by reference herein).
An "alkyl" group refers to a saturated aliphatic hydrocarbon, including straight- chain, branched-chain, and cyclic alkyl groups. Preferably, the alkyl group has 1 to 12 carbons. More preferably it is a lower alkyl of from 1 to 7 carbons, more preferably 1 to 4 carbons. The alkyl group may be substituted or unsubstituted. When substituted the substituted group(s) is preferably, hydroxyl, cyano, alkoxy, =O, =S, NO2 or N(CH3)2, amino, or SH. The term also includes alkenyl groups which are unsaturated hydrocarbon groups containing at least one carbon-carbon double bond, including straight-chain, branched-chain, and cyclic groups. Preferably, the alkenyl group has 1 to 12 carbons. More preferably it is a lower alkenyl of from 1 to 7 carbons, more preferably 1 to 4 carbons. The alkenyl group may be substituted or unsubstituted. When substituted the
substituted group(s) is preferably, hydroxyl, cyano, alkoxy, =O, =S, NO2, halogen, N(CH3)2, amino, or SH. The term "alkyl" also includes alkynyl groups which have an unsaturated hydrocarbon group containing at least one carbon-carbon triple bond, including straight-chain, branched-chain, and cyclic groups. Preferably, the alkynyl group has 1 to 12 carbons. More preferably it is a lower alkynyl of from 1 to 7 carbons, more preferably 1 to 4 carbons. The alkynyl group may be substituted or unsubstituted. When substituted the substituted group(s) is preferably, hydroxyl, cyano, alkoxy, =O, =S, NO2 or N(CH3)2, amino or SH.
Such alkyl groups may also include aryl, alkylaryl, carbocyclic aryl, heterocyclic aryl, amide and ester groups. An "aryl" group refers to an aromatic group which has at least one ring having a conjugated pi electron system and includes carbocyclic aryl, heterocyclic aryl and biaryl groups, all of which may be optionally substituted. The preferred substituent(s) of aryl groups are halogen, trihalomethyl, hydroxyl, SH, OH, cyano, alkoxy, alkyl, alkenyl, alkynyl, and amino groups. An "alkylaryl" group refers to an alkyl group (as described above) covalently joined to an aryl group (as described above). Carbocyclic aryl groups are groups wherein the ring atoms on the aromatic ring are all carbon atoms. The carbon atoms are optionally substituted. Heterocyclic aryl groups are groups having from 1 to 3 heteroatoms as ring atoms in the aromatic ring and the remainder ofthe ring atoms are carbon atoms. Suitable heteroatoms include oxygen, sulfur, and nifrogen, and include furanyl, thienyl, pyridyl, pyrrolyl, N-lower alkyl pyrrolo, pyrimidyl, pyrazinyl, imidazolyl and the like, all optionally substituted. An "amide" refers to an -C(O)-NH-R, where R is either alkyl, aryl, alkylaryl or hydrogen. An "ester" refers to an -C(O)-OR', where R is either alkyl, aryl, alkylaryl or hydrogen.
By "nucleotide" as used herein is as recognized in the art to include natural bases (standard), and modified bases well known in the art. Such bases are generally located at the 1' position of a nucleotide sugar moiety. Nucleotides generally comprise a base, sugar and a phosphate group. The nucleotides can be unmodified or modified at the sugar, phosphate and/or base moiety, (also referred to interchangeably as nucleotide analogs, modified nucleotides, non-natural nucleotides, non-standard nucleotides and other; see, for example, Usman and McSwiggen, supra; Eckstein et al, International PCT Publication No. WO 92/07065; Usman et al, International PCT Publication No. WO 93/15187;
Uhlman & Peyman, supra, all are hereby incoφorated by reference herein). There are several examples of modified nucleic acid bases known in the art as summarized by Limbach et al, 1994, Nucleic Acids Res. 22, 2183. Some ofthe non-limiting examples of base modifications that can be introduced into nucleic acid molecules include, inosine, purine, pyridin-4-one, pyridin-2-one, phenyl, pseudouracil, 2, 4, 6-trimethoxy benzene, 3- methyl uracil, dihydrouridine, naphthyl, aminophenyl, 5-alkylcytidines (e.g., 5-methylcytidine), 5-alkyluridines (e.g., ribothymidine), 5-halouridine (e.g., 5-bromouridine) or 6-azapyrimidines or 6-alkylpyrimidines (e.g. 6-methyluridine), propyne, and others (Burgin et al, 1996, Biochemistry, 35, 14090; Uhlman & Peyman, supra).
By "modified bases" in this aspect is meant nucleotide bases other than adenine, guanine, cytosine and uracil at 1' position or their equivalents; such bases may be used at any position, for example, within the catalytic core of an enzymatic nucleic acid molecule and/or in the substrate-binding regions ofthe nucleic acid molecule. Such modified nucleotides include dideoxynucleotides which have pharmaceutical utility well known in the art, as well as utility in basic molecular biology methods such as sequencing. In a preferred embodiment, the invention features modified ribozymes with phosphate backbone modifications comprising one or more phosphorothioate, phosphorodithioate, methylphosphonate, moφholino, amidate carbamate, carboxymethyl, acetamidate, polyamide, sulfonate, sulfonamide, sulfamate, formacetal, thioformacetal, and/or alkylsilyl, substitutions. For a review of oligonucleotide backbone modifications, see Hunziker and Leumann, 1995, Nucleic Acid Analogues: Synthesis and Properties, in Modern Synthetic Methods, VCH, 331-417, and Mesmaeker et al, 1994, Novel Backbone Replacements for Oligonucleotides, in Carbohydrate Modifications in Antisense Research, ACS, 24-39. These references are hereby incoφorated by reference herein.
By "abasic" is meant sugar moieties lacking a base or having other chemical groups in place of a base at the 1' position, (for more details, see Wincott et al, International PCT publication No. WO 97/26270).
By "unmodified nucleoside" or "unmodified nucleotide" is meant one ofthe bases adenine, cytosine, guanine, thymine, uracil joined to the 1' carbon of β-D-ribo-furanose.
By "modified nucleoside" or "modified nucleotide" is meant any nucleotide base which contains a modification in the chemical structure of an unmodified nucleotide base, sugar and/or phosphate.
In connection with 2'-modified nucleotides as described for the present invention, by "amino" is meant 2'-NH2 or 2'-O- NH2, which may be modified or unmodified. Such modified groups are described, for example, in Eckstein et al., U.S. Patent 5,672,695 and Matulic-Adamic et al., WΟ 98/28317, which are both incoφorated by reference in their entireties.
Various modifications to nucleic acid (e.g., antisense and ribozyme) structure can be made to enhance the utility of these molecules. Such modifications will enhance shelf-life, half-life in vitro, stability, and ease of introduction of such oligonucleotides to the target site, e.g., to enhance penetration of cellular membranes, and confer the ability to recognize and bind to targeted cells.
Use of these molecules will lead to better treatment ofthe disease progression by affording the possibility of combination therapies (e.g., multiple ribozymes targeted to different genes, ribozymes coupled with known small molecule inhibitors, or intermittent treatment with combinations of ribozymes (including different ribozyme motifs) and/or other chemical or biological molecules). The treatment of patients with nucleic acid molecules may also include combinations of different types of nucleic acid molecules. Therapies may be devised which include a mixture of ribozymes (including different ribozyme motifs), antisense and/or 2-5 A chimera molecules to one or more targets to alleviate symptoms of a disease.
Administration of Nucleic Acid Molecules Methods for the delivery of nucleic acid molecules are described in Akhtar et al,
1992, Trends Cell Bio., 2, 139; and Delivery Strategies for Antisense Oligonucleotide Therapeutics, ed. Akhtar, 1995 which are both incoφorated herein by reference. Sullivan et al, PCT WO 94/02595, further describes the general methods for delivery of enzymatic RNA molecules. These protocols may be utilized for the delivery of virtually any nucleic acid molecule. Nucleic acid molecules may be administered to cells by a variety of methods known to those familiar to the art, including, but not restricted to, encapsulation in liposomes, by iontophoresis, or by incoφoration into other vehicles, such as hydrogels,
cyclodextrins, biodegradable nanocapsules, and bioadhesive microspheres. For some indications, nucleic acid molecules may be directly delivered ex vivo to cells or tissues with or without the aforementioned vehicles. Alternatively, the nucleic acid/vehicle combination is locally delivered by direct injection or by use of a catheter, infusion pump or stent. Many examples in the art describe CNS delivery methods of oligonucleotides by osmotic pump, (see Chun et al, 1998, Neuroscience Letters, 257, 135-138, D'Aldin et al, 1998, Mol. Brain Research, 55, 151-164, Dryden et α/., 1998, J. Endocrinol, 157, 169- 175, Ghirnikar et al, 1998, Neuroscience Letters, 247, 21-24) or direct infusion (Broaddus et al, 1997, Neurosurg. Focus, 3, article 4). Other routes of delivery include, but are not limited to oral (tablet or pill form) and/or intrathecal delivery (Gold, 1997, Neuroscience, 76, 1153-1158). For a comprehensive review on drug delivery strategies including broad coverage of CNS delivery, see Jain, Drug Delivery Systems: Technologies and Commercial Opportunities, Decision Resources, 1998. Other routes of delivery include, but are not limited to, infravascular, intramuscular, subcutaneous or joint injection, aerosol inhalation, oral (tablet or pill form), topical, systemic, ocular, intraperitoneal and/or intrathecal delivery. More detailed descriptions of nucleic acid delivery and administration are provided in Sullivan et al, supra, Draper et al, PCT WO93/23569; Beigelman et al, PCT WO99/05094, and Klimuk et al, PCT WO99/04819 all of which are incoφorated by reference herein. The molecules ofthe instant invention can be used as pharmaceutical agents.
Pharmaceutical agents prevent, inhibit the occurrence, or treat (alleviate a symptom to some extent, preferably all ofthe symptoms) of a disease state in a patient.
The negatively charged polynucleotides ofthe invention can be administered (e.g., RNA, DNA or protein) and introduced into a patient by any standard means, with or without stabilizers, buffers, and the like, to form a pharmaceutical composition. When it is desired to use a liposome delivery mechanism, standard protocols for formation of liposomes can be followed. The compositions ofthe present invention may also be formulated and used as tablets, capsules or elixirs for oral administration; suppositories for rectal administration; sterile solutions; suspensions for injectable administration; and the other compositions known in the art.
The present invention also includes pharmaceutically acceptable formulations ofthe compounds described. These formulations include salts ofthe above compounds, e.g., acid addition salts, for example, salts of hydrochloric, hydrobromic, acetic acid, and benzene sulfonic acid. A pharmacological composition or formulation refers to a composition or formulation in a form suitable for administration, e.g., systemic administration, into a cell or patient, preferably a human. Suitable forms, in part, depend upon the use or the route of entry, for example, oral, transdermal, or by injection. Such forms should not prevent the composition or formulation from reaching a target cell (i.e., a cell to which the negatively charged polymer is desired to be delivered to). For example, pharmacological compositions injected into the blood stream should be soluble. Other factors are known in the art, and include considerations such as toxicity and forms which prevent the composition or formulation from exerting its effect.
By "systemic administration" is meant in vivo systemic absoφtion or accumulation of drugs in the blood stream followed by distribution throughout the entire body.
Administration routes which lead to systemic absoφtion include, without limitations: intravenous, subcutaneous, intraperitoneal, inhalation, oral, infrapulmonary and intramuscular. Each of these administration routes expose the desired negatively charged polymers, e.g., nucleic acids, to an accessible diseased tissue. The rate of entry of a drug into the circulation has been shown to be a function of molecular weight or size. The use of a liposome or other drug carrier comprising the compounds ofthe instant invention can potentially localize the drug, for example, in certain tissue types, such as the tissues ofthe reticular endothelial system (RES). A liposome formulation which can facilitate the association of drug with the surface of cells, such as, lymphocytes and macrophages is also useful. This approach may provide enhanced delivery ofthe drug to target cells by taking advantage ofthe specificity of macrophage and lymphocyte immune recognition of abnormal cells, such as cancer cells.
By pharmaceutically acceptable formulation is meant, a composition or formulation that allows for the effective distribution ofthe nucleic acid molecules ofthe instant invention in the physical location most suitable for their desired activity. Nonlimiting examples of agents suitable for formulation with the nucleic acid molecules ofthe instant invention include: P-glycoprotein inhibitors (such as Pluronic P85) which can enhance
entry of drugs into the CNS (Jolliet-Riant and Tillement, 1999, Fundam. Clin. Pharmacol, 13, 16-26); biodegradable polymers, such as poly (DL-lactide-coglycolide) microspheres for sustained release delivery after intracerebral implantation (Emerich, DF et al, 1999, Cell Transplant, 8, 47-58) Alkermes, Inc. Cambridge, MA; and loaded nanoparticles, such as those made of polybutylcyanoacrylate, which can deliver drugs across the blood brain barrier and can alter neuronal uptake mechanisms (Prog Neuropsychopharmacol Biol Psychiatry, 23, 941-949, 1999). Other non-limiting examples of delivery strategies for the nucleic acid molecules ofthe instant invention include material described in Boado et al, 1998, J. Pharm. Sci., 87, 1308-1315; Tyler et al, 1999, FEBS Lett., 421, 280-284; Pardridge et al, 1995, PNAS USA., 92, 5592-5596; Boado, 1995, Adv. Drug Delivery Rev., 15, 73-107; Aldrian-Herrada et al, 1998, Nucleic Acids Res., 26, 4910-4916; and Tyler et al, 1999, PNAS USA., 96, 7053-7058.
The invention also features the use ofthe composition comprising surface-modified liposomes containing poly (ethylene glycol) lipids (PEG-modified, or long-circulating liposomes or stealth liposomes). These formulations offer a method for increasing the accumulation of drugs in target tissues. This class of drug carriers resists opsonization and elimination by the mononuclear phagocytic system (MPS or RES), thereby enabling longer blood circulation times and enhanced tissue exposure for the encapsulated drug (Lasic et al. Chem. Rev. 1995, 95, 2601-2627; Ishiwata et al, Chem. Pharm. Bull. 1995, 43, 1005- 1011). Such liposomes have been shown to accumulate selectively in tumors, presumably by extravasation and capture in the neovascularized target tissues (Lasic et al, Science 1995, 267, 1275-1276; Oku et al., 995, Biochim. Biophys. Acta, 1238, 86-90). The long- circulating liposomes enhance the pharmacokinetics and pharmacodynamics of DNA and RNA, particularly compared to conventional cationic liposomes which are known to accumulate in tissues ofthe MPS (Liu et al, J. Biol. Chem. 1995, 42, 24864-24870; Choi et al, International PCT Publication No. WO 96/10391; Ansell et al, Intemational PCT Publication No. WO 96/10390; Holland et al, International PCT Publication No. WO 96/10392; all of which are incoφorated herein by reference). Long-circulating liposomes are also likely to protect drugs from nuclease degradation to a greater extent compared to cationic liposomes, based on their ability to avoid accumulation in metabolically aggressive MPS tissues such as the liver and spleen.
The present invention also includes compositions prepared for storage or administration which include a pharmaceutically effective amount ofthe desired compounds in a pharmaceutically acceptable carrier or diluent. Acceptable carriers or diluents for therapeutic use are well known in the pharmaceutical art, and are described, for example, in Remington's Pharmaceutical Sciences, Mack Publishing Co. (A.R.
Gennaro edit. 1985) hereby incoφorated by reference herein. For example, preservatives, stabilizers, dyes and flavoring agents may be provided. These include sodium benzoate, sorbic acid and esters ofp-hydroxybenzoic acid. In addition, antioxidants and suspending agents may be used. A pharmaceutically effective dose is that dose required to prevent, inhibit the occurrence, or treat (alleviate a symptom to some extent, preferably all ofthe symptoms) of a disease state. The pharmaceutically effective dose depends on the type of disease, the composition used, the route of administration, the type of mammal being freated, the physical characteristics ofthe specific mammal under consideration, concurrent medication, and other factors which those skilled in the medical arts will recognize. Generally, an amount between 0.1 mg/kg and 100 mg/kg body weight/day of active ingredients is administered dependent upon potency ofthe negatively charged polymer. The nucleic acid molecules ofthe present invention may also be administered to a patient in combination with other therapeutic compounds to increase the overall therapeutic effect. The use of multiple compounds to freat an indication may increase the beneficial effects while reducing the presence of side effects.
Alternatively, certain ofthe nucleic acid molecules ofthe instant invention can be expressed within cells from eukaryotic promoters (e.g., Izant and Weintraub, 1985, Science, 229, 345; McGarry and Lindquist, 1986, Proc. Natl. Acad. Sci., USA 83, 399; Scanlon et al, 1991, Proc. Natl. Acad. Sci. USA, 88, 10591-5; Kashani-Sabet et al, 1992, Antisense Res. Dev., 2, 3-15; Dropulic et al, 1992, J. Virol, 66, 1432-41; Weerasinghe et al, 1991, J. Virol, 65, 5531-4; Ojwang et α/., 1992, Proc. Natl. Acad. Sci. USA, 89, 10802-6; Chen et al, 1992, Nucleic Acids Res., 20, 4581-9; Sarver et α/., 1990 Science, 247, 1222-1225; Thompson et al, 1995, Nucleic Acids Res., 23, 2259; Good et al, 1997, Gene Therapy, 4, 45; all of these references are hereby incoφorated herein, in their totalities, by reference). Those skilled in the art realize that any nucleic acid can be expressed in eukaryotic cells from the appropriate DNA/RNA vector. The activity of such
nucleic acids can be augmented by their release from the primary transcript by a ribozyme (Draper et al, PCT WO 93/23569, and Sullivan et al, PCT WO 94/02595; Ohkawa et al, 1992, Nucleic Acids Symp. Ser., 27, 15-6; Taira et α/., 1991, Nucleic Acids Res., 19, 5125- 30; Ventura et al, 1993, Nucleic Acids Res., 21, 3249-55; Chowrira et al, 1994, J. Biol. Chem., 269, 25856; all of these references are hereby incoφorated in their totality by reference herein).
In another aspect ofthe invention, RNA molecules ofthe present invention are preferably expressed from transcription units (see, for example, Couture et al, 1996, TIG., 12, 510) inserted into DNA or RNA vectors. The recombinant vectors are preferably DNA plasmids or viral vectors. Ribozyme expressing viral vectors could be constructed based on, but not limited to, adeno-associated virus, retrovirus, adenovirus, or alphavirus. Preferably, the recombinant vectors capable of expressing the nucleic acid molecules are delivered as described above, and persist in target cells. Alternatively, viral vectors may be used that provide for transient expression of nucleic acid molecules. Such vectors might be repeatedly administered as necessary. Once expressed, the nucleic acid molecule binds to the target mRNA. Delivery of nucleic acid molecule expressing vectors could be systemic, such as by intravenous or intra-muscular administration, by administration to target cells ex-planted from the patient followed by reinfroduction into the patient, or by any other means that would allow for introduction into the desired target cell (for a review see Couture et al, 1996, TIG, 12, 510).
In one aspect, the invention features an expression vector comprising a nucleic acid sequence encoding at least one ofthe nucleic acid molecules ofthe instant invention is disclosed. The nucleic acid sequence encoding the nucleic acid molecule ofthe instant invention is operably linked in a manner which allows expression of that nucleic acid molecule.
In another aspect the invention features an expression vector comprising: a) a transcription initiation region (e.g., eukaryotic pol I, II or HI initiation region); b) a transcription termination region (e.g., eukaryotic pol I, LI or LU termination region); c) a nucleic acid sequence encoding at least one ofthe nucleic acid catalyst ofthe instant invention; and wherein said sequence is operably linked to said initiation region and said termination region, in a manner which allows expression and/or delivery of said nucleic acid molecule. The vector may optionally include an open reading frame (ORF) for a
protein operably linked on the 5' side or the 3'-side ofthe sequence encoding the nucleic acid catalyst ofthe invention; and/or an intron (intervening sequences).
Transcription ofthe nucleic acid molecule sequences are driven from a promoter for eukaryotic RNA polymerase I (pol I), RNA polymerase II (pol H), or RNA polymerase HI (pol HI). Transcripts from pol π or pol LU promoters will be expressed at high levels in all cells; the levels of a given pol LI promoter in a given cell type will depend on the nature of the gene regulatory sequences (enhancers, silencers, etc.) present nearby. Prokaryotic RNA polymerase promoters are also used, providing that the prokaryotic RNA polymerase enzyme is expressed in the appropriate cells (Elroy-Stein and Moss, 1990, Proc. Natl. Acad. Sci. USA, 87, 6743-7; Gao and Huang 1993, Nucleic Acids Res., 21, 2867-72;
Lieber et al., 1993, Methods Enzymol, 217, 47-66; Zhou et al., 1990, Mol. Cell. Biol, 10, 4529-37). All of these references are incoφorated by reference herein. Several investigators have demonstrated that nucleic acid molecules, such as ribozymes expressed from such promoters can function in mammalian cells (e.g. Kashani-Sabet et al., 1992, Antisense Res. Dev., 2, 3-15; Ojwang et al., 1992, Proc. Natl. Acad. Sci. USA, 89,
10802-6; Chen et al, 1992, Nucleic Acids Res., 20, 4581-9; Yu et al., 1993, Proc. Natl Acad. Sci. USA, 90, 6340-4; L'Huillier et α/., 992, EMBOJ., 11, 4411-8; Lisziewicz et al, 1993, Proc. Natl. Acad. Sci. U. S. A, 90, 8000-4; Thompson et al, 1995, Nucleic Acids Res., 23, 2259; Sullenger & Cech, 1993, Science, 262, 1566). More specifically, transcription units such as the ones derived from genes encoding U6 small nuclear
(snRNA), transfer RNA (tRNA) and adenovirus VA RNA are useful in generating high concentrations of desired RNA molecules such as ribozymes in cells (Thompson et al, supra; Couture and Stinchcomb, 1996, supra; Noonberg et al, 1994, Nucleic Acid Res., 22, 2830; Noonberg et al, US Patent No. 5,624,803; Good et al, 1997, Gene Ther., A, 45; Beigelman et al, International PCT Publication No. WO 96/18736; all of these publications are incoφorated by reference herein. The above ribozyme transcription units can be incoφorated into a variety of vectors for introduction into mammalian cells, including but not restricted to, plasmid DNA vectors, viral DNA vectors (such as adenovirus or adeno-associated virus vectors), or viral RNA vectors (such as retroviral or alphavirus vectors) (for a review see Couture and Stinchcomb, 1996, supra).
In yet another aspect, the invention features an expression vector comprising nucleic acid sequence encoding at least one ofthe nucleic acid molecules ofthe invention, in a manner which allows expression of that nucleic acid molecule. The expression vector comprises in one embodiment; a) a transcription initiation region; b) a transcription termination region; c) a nucleic acid sequence encoding at least one said nucleic acid molecule; and wherein said sequence is operably linked to said initiation region and said termination region, in a manner which allows expression and/or delivery of said nucleic acid molecule. In another preferred embodiment the expression vector comprises: a) a transcription initiation region; b) a transcription termination region; c) an open reading frame; d) a nucleic acid sequence encoding at least one said nucleic acid molecule, wherein said sequence is operably linked to the 3 '-end of said open reading frame; and wherein said sequence is operably linked to said initiation region, said open reading frame and said termination region, in a manner which allows expression and/or delivery of said nucleic acid molecule. In yet another embodiment, the expression vector comprises: a) a transcription initiation region; b) a transcription termination region; c) an infron; d) a nucleic acid sequence encoding at least one said nucleic acid molecule; and wherein said sequence is operably linked to said initiation region, said intron and said termination region, in a manner which allows expression and/or delivery of said nucleic acid molecule. In another embodiment, the expression vector comprises: a) a transcription initiation region; b) a transcription termination region; c) an infron; d) an open reading frame; e) a nucleic acid sequence encoding at least one said nucleic acid molecule, wherein said sequence is operably linked to the 3'-end of said open reading frame; and wherein said sequence is operably linked to said initiation region, said infron, said open reading frame and said termination region, in a manner which allows expression and/or delivery of said nucleic acid molecule.
Examples:
The following are non-limiting examples showing the selection, isolation, synthesis and activity of nucleic acids ofthe instant invention.
Example 1 : Telomerase
The ribonucleoprotein enzyme telomerase consists of an RNA template subunit and one or more protein subunits including telomerase reverse transcriptase (TERT), which function together to direct the synthesis of telomeres. Telomeres exist as non-nucleosome DNA/protein complexes at the physical ends of eukaryotic chromosomes. These capping structures maintain chromosome stability and replicative potential (Zakian, V. A., 1995, Science, 270, 1601-1607). Telomere structure is characterized by tandem repeats of conserved DNA sequences rich in G-C base pairs. Additional conserved telomere elements include a terminal 3 '-overhang in the G-rich strand and non-histone structural proteins that are complexed with telomeric DNA in the nucleus. (Blackburn, "E., 1990, JBC, 265, 5919-5921.). Observed shortening of telomeres coincides with the onset of cellular senescence in most somatic cell lines lacking significant levels of telomerase. This finding has had a profound impact on our views concerning the mechanisms of aging, age related disease, and cancer. Conventional DNA polymerases are unable to fully replicate the ends of linear chromosomes (Watson, J. D., 1972, Nature, 239, 197-201). This inability stems from the 3' G-rich overhang that is a product of ribonuclease cleavage ofthe RNA primer used in DNA replication. The overhang prevents DNA polymerase replication since the recessed C-rich parent strand cannot be used as a template. Telomerase overcomes this limitation by extending the 3' end ofthe chromosome using deoxyribonucleotides as substrates and a sequence within the telomerase RNA subunit as a template. (Lingner, J., 1995, Science, 269, 1533-1534). As such, telomerase is considered a reverse transcriptase that is responsible for telomere maintenance.
Telomerase was first discovered by in Tetrahymena thermophila in 1985 (Greider, C. W., 1995, Cell, 43, 405-413). The RNA subunits and their respective genes were later discovered and characterized in protozoa, budding yeast, and mammals. Genetic studies of these genes confirmed the role of telomerase RNA (TR) in determining telomere sequence by mutating genes which encode the telomeric RNA (Yu, G. L., 1990, Nature, 344, 126-132), (Singer, M. S., 1994, Science, 266, 404-409), (Blasco, M. A., 1995, Science, 269, 1267-1270). These studies showed that telomerase activity parallels TR expression in protozoa, yeast and mice. However, the expression of human telomerase RNA (hTR) does not correlate well with telomerase activity in mammalian cells. Many
human tissues express hTR but are devoid of telomerase activity (Feng, J., 1995, Science, 269, 1236-1241). Knockout mice, in which the mTR gene has been deleted from germline cells, have been shown to be viable for at least six generations. Cells from later generations of these mice showed chromosomal abnormalities consistent with telomere degradation, indicating that mTR is necessary for telomere length maintenance, but is not required for embryonic development, oncogenic transformation, or tumor formation in mice (Blasco, M. A., 1997, Cell, 91, 25-34).
The first catalytically active subunit of telomerase (pi 23) was isolated from Euplotes aediculatus along with another subunit (p43) and a 66-kD RNA subunit (Linger, J., 1996, Proc. Natl. Acad. Sci., 93, 10712-10717). Subsequent studies revealed telomerase catalytic subunit homologs from fission yeast (Est2p) and human genes (TRT1). The human homolog, TRT1 encoding hTERT, expressed mRNA with a strong correlation to telomerase activity in human cells (Nakamura, T. M., 1997, Science, 277, 955-959). Reconstitution of telomerase activity with in vitro transcribed and translated hTERT and hTR, either co-synthesized or simply mixed, demonstrated that hTERT and hTR represent the minimal components of telomerase. Furthermore, transient expression of hTERT in normal diploid human cells restored telomerase activity, demonstrating that hTERT is the only component necessary to restore telomerase activity in normal human cells (Weinrich, S. L., 1997, Nature Genetics, 17, 498-502). The introduction of telomerase into normal human cells using hTERT expression via fransfection has resulted in the extension of life span in these cells. Such findings indicate that telomere loss in the absence of telomerase is the "mitotic clock" that controls the replicative potential of a cell prior to senescence (Bodnar, A. G., 1998, Science, 279, 349-352).
Expression of telomerase is observed in germ cell and most cancer cell lines. These "immortal" cell lines continue to divide without shortening of their telomeres (Kim, N. W., 1994, Science, 266, 2011-2015). A model of tumor progression has evolved from these findings, suggesting a role for telomerase expression in malignant transformation. Successful malignant transformation in human cells was accomplished for the first time by ectopic expression of hTERT in combination with two oncogenes, SV40 large-T and H- ras. Injection of nude mice with cells expressing these oncogenes and hTERT resulted in rapid growth of tumors. These observations indicate that hTERT mediated telomere
maintenance is essential for the formation of human tumor cells (Hahn, W. C, 1999, Nature, 400, 464-468).
Various methods have been developed to assay telomerase activity in vitro. The most widely used method to characterize telomerase activity is the telomeric repeat amplification protocol (TRAP). TRAP utilizes RT-PCR of cellular exfracts to measure telomerase activity by making the amount of PCR target dependant upon the biochemical activity ofthe enzyme (Kim, N. W., 1997, Nucleic Acids Research, 25, 2595-2597, which is incoφorated by reference herein).
A method based on Kim is as follows. Briefly, for the telomerase assay, 2μg of protein exfract is used. The exfract is assayed in 50μl of reaction mixture containing 0.1 μg TS substrate primer (5'-AATCCGTCGAGCAGAGTT-3', end-labeled using alpha-32P- ATP and T4 polynucleotide kinase), 0.1 μg ACX return primer(5'-GCGCGG[CTTACC]3 CTAACC-3'), 0.1 μg NT internal control primer (5*-ATCGCTTCTCGGCCTTTT-3'), 0.01 micromol TSNT internal confrol template (5'- AATCCGTCGAGCAGAGTTAAAAGGCCGAGAACGAT-3), 50 μM each deoxynucleoside triphosphate, 2 U of Taq DNA polymerase, and 2 μl CHAPS protein exfract, all in IX TRAP buffer (20 mM Tris (pH 8.3), 68 mM KC1, 1.5 mM MgCl2, 1 mM EGTA, 0.05% Tween 20). Each reaction is placed in a thermocycler block preheated to 30 C and incubated at 30 C for 10 minutes, then cycled for 27 cycles of 94 degrees C for 30 seconds, 60 degrees C for 30 seconds. Reaction products are separated on a denaturing 8% polyacrylamide gel, followed by drying ofthe gel and autoradiography. The internal control (to control for possible Taq polymerase inhibition) generates a band of 36 nt. Comparison of radioactive signal integrated (e.g., by phoφhorimager analysis) for telomerase-extended bands with the radioactive signal from a reaction performed with a known amount of quantification standard template (termed R8; 5'-
AATCCGTCGAGCAGAGTTAG [GGTTAG]7-3) allows expression of telomerase activity as an absolute value. The absolute value = TPG (total product generated) =[(TP- TPi)/TI]/[(R8-B)/RI)] x 100, where TP = telomerase products from test exfract, TPi = telomerase products from a heat-inactivated (75 C, 10 minutes) extract reaction, Tl = the signal from the internal control, R8 = the signal from the R8 qualification standard template reaction, B = signal from a lysis buffer-only blank reaction, and RI = the internal confrol value for the reaction containing R8 template and NT and TSNT confrol primers.
TPG values of 0-10,000 are possible, with the linear range being from approximately 1 to 1000 TPG. The range of 1 to 1000 TPG encompasses the minimum and maximum levels of telomerase activity in most tumor samples tested, while non-tumor cells most often have no telomerase activity (TPG approximately zero). Telomerase activity may also be assayed as follows. Samples to be assayed for telomerase activity are prepared by extraction into CHAPS lysis buffer (lOmM Tris pH 7.5, ImM MgCl2, ImM EGTA, 0.1 mM PMSF, 5mM -mercaptoethanol, ImM DTT, 0.5% 3-[(3-cholamidopropyl)-dimethyl-amino]-l- propanesulfonate (CHAPS), 10% glycerol and 40 U/ml RNAse inhibitor (Promega, Madison, WI, U.S.A.). Cells are suspended in CHAPS lysis buffer and incubated on ice for 30 minutes, which allows lysis of 90-100%) of cells. Lysate is then transferred to polyallomer centrifuge tubes and spun at 100,000 x g for 1 hour at 4 degrees C. The supernatant is the protein exfract, and concentration ranges of 4-10 μg/μl are suitable for telomerase assay. Extracts may be concentrated if necessary using a Microcon Microfilter 30 (Amicron, Beverly, MA U.S.A.) according to the manufactureris instructions. Exfracts may be stored frozen at -80 degrees C until assayed.
A variety of animal models have been designed to assay telomerase activity in vivo. Inhibition of telomerase activity has been analyzed in rats via cell proliferation studies with MNU (N-methyl-N-nitosurea) induced mammary carcinomas in response to treatment with 4-(hydroxyphenyl)retinamide (4-HPR), a known inhibitor of mammary carcinogenesis in animal models and premenopausal women (Bednarek, A., 1999, Carcinogenesis, 20, 879-883). Additional studies have focused on the up-regulation of telomerase in transformed cell lines from animal and human model systems (Zhang, P. B., 1998, Leuk. Res., 22, 509-516), (Chadeneau, C, 1995, Oncogene, 11, 893-898), (Greenberg, R., 1999, Oncogene, 18, 1219-1226).
Human cell culture studies have been established to assay inhibition of telomerase activity in human carcinomas responding to various therapeutics. A human breast cancer model for studying telomerase inhibitors is described (Raymond, E., 1999, Br. J. Cancer, 80, 1332-1341). Human studies of telomerase expression as related to various other cancers are described including cervical cancer (Nakano, K., 1998, Am. J. Pathol, 153, 857-864), endometrial cancer (Kyo, S., 1999, Int. J. Cancer, 80, 60-63), meningeal carcinoma (Kleinschmidt-DeMasters, B. K., 1998, J. Neurol. Sci., 161, 124-134), lung
carcinoma (Yashima, K., 1997, Cancer Reseach, 57, 2372-2377), testicular cancer in response to cisplatin (Burger, A. M., 1997, Eur. J. Cancer, 33, 638-644), and ovarian carcinoma (Counter, C. M., 1994, Proc. Natl. Acad. Sci., 91, 2900-2904).
Particular degenerative and disease states that can be associated with telomerase expression modulation include but are not limited to:
Cancer: Almost all human tumors have detectable telomerase activity (Shay, J. W., 1997, Eur. J. Cancer, 33, 787-791). Treatment with telomerase inhibitors may provide effective cancer therapy with minimal side effects in normal somatic cells that lack telomerase activity. The therapeutic potential exists for the treatment of a wide variety of cancer types.
Restinosis: Telomerase inhibition in vascular smooth muscle cells may inhibit restinosis by limiting proliferation of these cells.
Infectious disease: Telomerase inhibition in infectious cell types that express telomerase activity may provide selective anti-infectious agent activity. Such treatment may prove especially effective in protozoan-based infection such as Giardia and Lesh Meniesis.
Transplant rejection: Telomerase inhibition in endothelial cell types may demonstrate selective immunnosuppressant activity. Activation of telomerase in transplant cells could benefit grafting success through increased proliferative potential. Autoimmune disease: Telomerase modulation in various immune cells may prove beneficial in treating diseases such as multiple sclerosis, lupus, and AIDS.
Age related disease: Activation of telomerase expression in cells at or nearing senescence as a result of advanced age or premature aging could benefit conditions such as macular degeneration, skin ulceration, and rheumatoid arthritis. The present body of knowledge in telomerase research indicates the need for methods to assay telomerase activity and for compounds that can regulate telomerase expression for research, diagnostic, trait alteration, animal health and therapeutic use.
Gemcytabine and cyclophosphamide are non-limiting examples of chemotherapeutic agents that can be combined with or used in conjunction with the nucleic acid molecules (e.g. ribozymes and antisense molecules) ofthe instant invention. Those skilled in the art will recognize that other drugs such as anti-cancer compounds and therapies can be similarly be readily combined with the nucleic acid molecules ofthe instant invention
(e.g. ribozymes and antisense molecules) and are hence within the scope ofthe instant invention. Such compounds and therapies are well known in the art (see for example Cancer: Principles and Pranctice of Oncology, Volumes 1 and 2, eds Devita, V.T., Hellman, S., and Rosenberg, S.A., J.B. Lippincott Company, Philadelphia, USA; incoφorated herein by reference) and include, without limitations, antifolates; fluoropyrimidines; cytarabine; purine analogs; adenosine analogs; amsacrine; topoisomerase I inhibitors; anthrapyrazoles; retinoids; antibiotics such as bleomycin, anthacyclins, mitomycin C, dactinomycin, and mithramycin; hexamethylmelamine; dacarbazine; 1-asperginase; platinum analogs; alkylating agents such as nitrogen mustard, melphalan, chlorambucil, busulfan, ifosfamide, 4-hydroperoxycyclophosphamide, nifrosoureas, thiotepa; plant derived compounds such as vinca alkaloids, epipodophyllotoxins, taxol; Tomaxifen; radiation therapy; surgery; nutritional supplements; gene therapy; radiotherapy such as 3D-CRT; immunotoxin therapy such as ricin, monoclonal antibodies herceptin; and the like. For combination therapy, the nucleic acids ofthe invention are prepared in one of two ways. First, the agents are physically combined in a preparation of nucleic acid and chemotherapeutic agent, such as a mixture of a nucleic acid ofthe invention encapsulated in liposomes and ifosfamide in a solution for intravenous administration, wherein both agents are present in a therapeutically effective concentration (e.g., ifosfamide in solution to deliver 1000-1250 mg/m2/day and liposome-associated nucleic acid ofthe invention in the same solution to deliver 0.1-100 mg/kg/day). Alternatively, the agents are administered separately but simultaneously in their respective effective doses (e.g., 1000-1250 mg/m2/d ifosfamide and 0.1 to 100 mg/kg/day nucleic acid ofthe invention).
Gaeta et al, US patents No. 5,760,062; 5,767,278; 5,770,613 have described small molecule inhibitors of human telomerase RNA (hTR) subunit.
Blasco et al, 1995, Science, 269, 1267-1270 describe the synthesis and testing of antisense oligonucleotides targeted against a specific region ofthe mouse telomerase RNA (mTR) subunit and reported reduction in telomerase activity in mice.
Bisoffi et al, 1998, Eur. J. Cancer, 34, 1242-1249 have studied the down regulation of human telomerase activity by a refro virus vector expressing antisense RNA targeted against the hTR RNA.
Norton et al, 1996, Nature Biotechnology, 14, 615-619 have reported the use of a peptide nucleic acid (PNA) molecule targeting hTR RNA to down regulate telomerase activity in human immortal breast epithelial cells.
Yokoyama et al, 1998, Cancer Research, 58, 5406-5410 have reported the synthesis and testing of hammerhead ribozyme constructs targeting hTR RNA resulting in a decrease in the telomerase activity in Ishikawa cells.
Henderson, European Patent Application No. 666,313-A2 describes methods of identifying and cloning hTR gene for use in gene therapy approaches for creating aberrant telomeric sequences in transfected human tumor cells. A ribozyme based gene therapy approach to inhibit the expression of hTR gene is described as well. The intended result of such therapies involves incurred genetic instability based on non-native telomeric sequences resulting in rapid cell death ofthe freated cells.
West et al, US patent No. 5,489,508 describe methods for determining telomere length and telomerase activity in cells. Inhibitors of hTR RNA, including oligonucleotides and/or small molecules are described.
These foregoing approaches of targeting the telomerase RNA subunit (TR) may not be very beneficial, because as demonstrated by Feng et al, (Feng, J., 1995, Science, 269, 1236-1241), telomerase activity in humans does not correlate well to hTR concentration.
Collins et al, International PCT publication No. WO 98/01542 describes assays for the detection of telomerase activity. Four human telomerase subunit proteins are described called pi 40, pi 05, p48 and p43. In addition, hybridization probes and primers are described as inhibitors of telomerase gene function. Antibody based inhibitors of telomerase protein subunits are described.
A more attractive approach to telomerase regulation would involve the regulation of human telomerase by modulating the expression ofthe protein subunits ofthe enzyme, preferably the reverse transcriptase (hTERT) subunit. Based of reconstitution experiments, hTERT and hTR represent the minimal components of telomerase. Since hTR expression does not correlate well with telomerase activity in human cells and since many human cells express hTR without telomerase activity, targeting hTERT may prove more beneficial than targeting hTR. hTERT is the only component necessary to restore telomerase activity in normal human cells. A study in which the three major subunits of telomerase (hTR, TPI, and hTERT were assayed in normal and malignant endometrial
tissues determined that hTERT is a rate limiting determinant of enzymatic activity of human telomerase (Kyo, S., 1999, Int. J. Cancer, 80, 60-63). Additional protein subunits that have been isolated most likely serve only a structural role in telomerase activity, but may be important in enhancing the activity ofthe telomerase enzyme. As such, hTERT is one ofthe better targets for the ectopic regulation of telomerase activity.
Cech et al, International PCT publication No. WO 98/14593 describe compositions and methods related to hTERT for diagnosis, prognosis and treatment of human diseases, for altering proliferative capacity in cells and organisms, and for screening compounds and treatments with potential use as human therapeutics. Cech et al, International PCT publication No. WO 98/14592 describe nucleic acid and amino acid sequences encoding various telomerase protein subunits and motifs of Euplotes aediculatus, and related sequences from Schizosaccharomyces, Saccharomyces sequences, and human telomerase. The polypeptides comprising telomeric subunits and functional polypeptides and ribonucleoproteins that contain these subunits are described as well. Cech et al, International PCT Publication No. WO 98/14592, mentions in general terms the the possibility of using antisense and ribozymes to down regulate the expression of human telomerase reverse transcriptase enzyme.
Identification of Potential Target Sites in Human TERT RNA The sequence of human TERT was screened for accessible sites using a computer folding algorithm. Regions ofthe RNA that did not form secondary folding structures and contained potential ribozyme and/or antisense binding/cleavage sites were identified. The sequences of these cleavage sites are shown in Tables 13-17.
Selection of Enzymatic Nucleic Acid Cleavage Sites in Human TERT RNA
To test whether the sites predicted by the computer-based RNA folding algorithm corresponded to accessible sites in TERT RNA, 10 hammerhead ribozyme and three G- Cleaver ribozyme sites were selected for further analysis (Table 17). Ribozyme target sites were chosen by analyzing sequences of Human TERT (Nakamura et al, 1997 Science 277, 955-959; Genbank sequence accession number: NM 003219) and prioritizing the sites on the basis of folding. Ribozymes were designed that could bind each target and were individually analyzed by computer folding (Christoffersen et al,
1994 J. Mol. Struc. Theochem, 311, 273; Jaeger et al, 1989, Proc. Natl. Acad. Sci. USA, 86, 7706) to assess whether the ribozyme sequences fold into the appropriate secondary structure. Those ribozymes with unfavorable intramolecular interactions between the binding arms and the catalytic core were eliminated from consideration. As noted below, varying binding arm lengths can be chosen to optimize activity. Generally, at least 5 bases on each arm are able to bind to, or otherwise interact with, the target RNA.
Chemical Synthesis and Purification of Ribozymes for Efficient Cleavage of TERT RNA Ribozymes were designed to anneal to various sites in the RNA message. The binding arms are complementary to the target site sequences described above. The ribozymes were chemically synthesized. The method of synthesis used followed the procedure for normal RNA synthesis as described above and in Usman et al., (1987 J. Am. Chem. Soc, 109, 7845), Scaringe et al., (1990 Nucleic Acids Res., 18, 5433) and Wincott et al., supra, and made use of common nucleic acid protecting and coupling groups, such as dimethoxytrityl at the 5'-end, and phosphoramidites at the 3'-end. The average stepwise coupling yields were >98%.
Ribozymes were also synthesized from DNA templates using bacteriophage T7 RNA polymerase (Milligan and Uhlenbeck, 1989, Methods Enzymol. 180, 51). Ribozymes were purified by gel electrophoresis using general methods or were purified by high pressure liquid chromatography (HPLC; See Wincott et al., supra; the totality of which is hereby incoφorated herein by reference) and were resuspended in water. The sequences of the chemically synthesized ribozymes used in this study are shown below in Table 13-17.
Ribozyme Cleavage of TERT RNA Target in vitro Ribozymes targeted to the human TERT RNA are designed and synthesized as described above. These ribozymes can be tested for cleavage activity in vitro, for example using the following procedure. The target sequences and the nucleotide location within the
TERT RNA are given in Tables 13-17.
Cleavage Reactions: Full-length or partially full-length, internally-labeled target RNA for ribozyme cleavage assay is prepared by in vitro transcription in the presence of
[a.32p] CTP, passed over a G 50 Sephadex column by spin chromatography and used as
substrate RNA without further purification. Alternately, substrates are 5'-32p-en(i labeled using T4 polynucleotide kinase enzyme. Assays are performed by pre- warming 15 μl of a 2X concentration of purified ribozyme in ribozyme cleavage buffer (50 mM Tris-HCI, pH 7.5 at 37°C, 10 mM MgC ) and the cleavage reaction was initiated by adding the 2X ribozyme mix to an equal volume (15 μl) of subsfrate RNA (maximum of 1-5 nM; 5 x 105 to 1 x 107 cpm) that was also pre- warmed in cleavage buffer. As an initial screen, assays o are carried out for 1 hour at 37 C using a final concentration of either 40 nM or 1 mM ribozyme, i.e., ribozyme excess. The reaction is quenched by the addition of an equal volume (30 μl) of 95% formamide, 20 mM EDTA, 0.05% bromophenol blue and 0.05% o xylene cyanol after which the sample is heated to 95 C for 2 minutes, quick chilled and loaded onto a denaturing polyacrylamide gel. Subsfrate RNA and the specific RNA cleavage products generated by ribozyme cleavage are visualized on an autoradiograph of the gel. The percentage of cleavage is determined by Phosphor Imager® quantitation of bands representing the intact subsfrate and the cleavage products.
Example 2: PTP-IB
Protein tyrosine phosphorylation and dephosphorylation are important mechanisms in the regulation of signal transduction pathways that control the processes of cell growth, proliferation, and differentiation (Fantl, W. J., 1993, Annu. Rev. Biochem., 62, 453-481). Cooperative enzyme classes regulate protein tyrosine phosphorylation and dephosphorylation events. These broad classes of enzymes consist ofthe protein tyrosine kinases (PTKs) and protein tyrosine phosphatases (PTPs). PTKs and PTPs can exist as both receptor-type transmembrane proteins and as cytoplasmic protein enzymes. Receptor tyrosine kinases propagate signal transduction events via extracellular receptor-ligand interactions that result in the activation ofthe tyrosine kinase portion ofthe PTK in the cytoplasmic domain. Receptor-like transmembrane PTPs function through extracellular ligand binding that modulates dephosphorylation of infracellular phosphotyrosine proteins via cytoplasmic phosphatase domains. Cytoplasmic PTKs and PTPs exert enzymatic activity without receptor-mediated ligand interactions, however, phosphorylation can regulate the activity of these enzymes.
Protein tyrosine phosphatase IB, a cytoplasmic PTP, was the first PTP to be isolated in homogeneous form (Tonks, N. K., 1988, J. Biol. Chem., 263, 6722-6730), characterized (Tonks, N. K., 1988, J. Biol. Chem., 263, 6731-6737), and sequenced (Charbonneau, H., 1989, Biochemistry, 86, 5252-5256). Cytoplasmic and receptor-like PTPs both share a catalytic domain characterized by eleven conserved amino acids containing cysteine and arginine residues that are critical for phosphatase activity (Streuli, M., 1990, EMBO, 9, 2399-2407). A cysteine residue at position 215 is responsible for the covalent attachment of phosphate to the enzyme (Guan, K, 1991, J. Biol. Chem., 266, 17026-17030). The crystal structure of human PTP IB defined the phosphate binding site ofthe enzyme as a glycine rich cleft at the surface of the molecule with cysteine 215 positioned at the base of this cleft. The location of cysteine 215 and the shape ofthe cleft provide specificity of PTPase activity for tyrosine residues but not for serine or threonine residues (Barford, D., 1994, Science, 263, 1397-1404).
Receptor tyrosine kinase and protein tyrosine phosphatase localization plays a key role in the regulation of phosphotyrosine mediated signal fransduction. PTP-IB activity and specificity against a panel of receptor tyrosine kinases demonstrated clear differences between substrates, suggesting that cellular comparrmentalization is a determinant in defining the activity and function ofthe enzyme (Lammers, R.,1993, J. Biol. Chem., 268, 22456-22462). Experiments have indicated that PTP-IB is localized predominantly in the endoplasmic reticulum via its 35 amino acid carboxyterminal sequence. PTP-IB is also tightly associated with microsomal membranes with its catalytic phosphatase domain oriented towards the cytoplasm (Frangioni, J. V., 1992, Cell, 68, 545-560).
PTP-IB has been identified as a negative regulator ofthe insulin response. PTP-IB is widely expressed in insulin sensitive tissues (Goldstein, B. J., 1993, Receptor, 3, 1-15). Isolated PTP-IB dephosphorylates the insulin receptor in vitro (Tonks, N. K., 1988, J. Biol. Chem., 263, 6731-6737). PTP-IB dephosphorylation of multiple phosphotyrosine residues ofthe insulin receptor proceeds sequentially and with specificity for the three tyrosine residues that are critical for receptor autoactivation (Ramachandran, C, 1992, Biochemistry, 31, 4232-4238). In addition to insulin receptor dephosphorylation, PTP-IB also dephosphorylates the insulin related subfrate 1 (IRS-1), a principal substrate ofthe insulin receptor (Lammers, R., 1993, J. Biol. Chem., 268, 22456-22462).
Microinjection of PTP IB into Xenopus oocytes results in the inhibition of insulin stimulated tyrosine phosphorylation of endogenous proteins, including the β-subunit ofthe insulin and insulin-like growth factor receptor proteins. The resulting 3 to 5 fold increase over endogenous PTPase activity also blocks the activation of an S6 peptide kinase (Cicirelli, M. F., 1990, Proc, Natl. Acad. Sci., 87, 5514-5518). Inactivation of recombinant rat PTP-IB with antibody immunoprecipitation results in the dramatic increase in insulin stimulated DNA synthesis and phosphatidylinositol 3 '-kinase activity. Insulin stimulated receptor autophosphorylation and insulin receptor substrate 1 tyrosine phosphorylation are increased dramatically as well through PTP-IB inhibition (Ahmad, F., 1995, J. Biol. Chem., 270, 20503-20508).
Increased PTP-IB expression correlates with insulin resistance in hyperglycemic cultured fibroblasts. In this study, desensitized insulin receptor function was observed via impaired insulin-induced autophosphorylation ofthe receptor. Treatment with insulin sensitivity normalizing thiazolidine derivatives resulted in the amelioration ofthe hyperglycemic insulin resistance via a normalization in PTP-IB expression (Maegawa, H., 1995, J. Biol. Chem., 270, 7724-7730). A murine model of insulin resistance with a knockout of the hefrerotrimeric GTP-binding protein subunit Giα2 provides a type 2 diabetis phenotype that correlates with the increased expression of PTP-IB (Moxam, C. M., 1996, Nature, 379, 840-844). PTP-IB interacts directly with the activated insulin receptor β-subunit. An inactive homolog of PTP-IB was used to precipitate the activated insulin receptor in both purified receptor preparations and whole-cell lysates. Phosphorylation ofthe insulin receptor's triple tyrosine residues in the kinase domain is necessary for PTP-IB interaction. Furthermore, insulin stimulates tyrosine phosphorylation of PTP-IB (Seely, B. L., 1996, Diabetes, 45, 1379-1385). A similar study confirmed the direct interaction of PTP-IB with the insulin receptor β-subunit as well as the required multiple phosphorylation sites within the receptor and PTP-IB (Bandyopadhyay, D., J. Biol. Chem., 272, 1639-1645).
Knockout mice lacking the PTP-IB gene (both homozygous PTP-IB"''" and heterozygous PTP-1B+/") have been used to study the specific role of PTP-IB relating to insulin action in vivo. The resulting PTP-IB deficient mice were healthy and, in the fed state, had lower blood glucose and circulating insulin levels that were half that of their
PTP-1B+/+ expressing littermates. These PTP-IB deficient mice demonstrated enhanced insulin sensitivity in glucose and insulin tolerance tests. At the physiological level, the PTP-IB deficient mice showed increased phosphorylation ofthe insulin receptor after insulin administration. When fed a high fat diet, the PTP-IB deficient mice were resistant to weight gain and remained insulin sensitive as opposed to normal PTP-IB expressing mice, who rapidly gained weight and become insulin resistant (Elchebly, M., 1999, Science, 283, 1544-1548). As such, modulation of PTP-IB expression could be used to regulate autophosphorylation ofthe insulin receptor and increase insulin sensitivity in vivo. This modulation could prove beneficial in the treatment of insulin related disease states. In light ofthe above findings, particular disease states that involve PTP-IB expression include but are not limited to:
Diabetes: Both type 1 and type 2 diabetes may be treated by modulation of PTP-IB expression. Type 2 diabetes correlates to desensitized insulin receptor function (White et al, 1994). Disruption ofthe PTP-IB dephosphorylation ofthe insulin receptor in vivo manifests in insulin sensitivity and increased insulin receptor autophosphorylation (Elchebly et al, 1999). Insulin dependant diabetes, type 1, may respond to PTP-IB modulation through increased insulin sensitivity.
Obesity: Elchebly et al, 1999, demonstrated that PTP-IB deficient mice were resistant to weight gain when fed a high fat diet compared to normal PTP-IB expressing mice. This finding suggests that PTP-IB modulation may be beneficial in the treatment of obesity. Ahmad et al, 1997, Metab. Clin. Exp., 46, 1140-1145, describe reduced PTPs in adipose tissue and improved insulin sensitivity in obese subjects following weight loss.
Troglitazone is a non-limiting example of a pharmaceutical agent that can be combined with or used in conjunction with the nucleic acid molecules (e.g. ribozymes and antisense molecules) ofthe instant invention. Those skilled in the art will recognize that other drugs such as anti-diabetes and anti-obesity compounds and therapies can be similarly be readily combined with the nucleic acid molecules ofthe instant invention (e.g. ribozymes and antisense molecules) are hence within the scope ofthe instant invention. Methods have been developed to assay PTP-IB activity.
Maegawa et al, 1995, J. Biol. Chem., 270, 7724-7730, describe a tissue culture model in which Rat 1 fibroblasts expressing human insulin receptors can be used to model
hyperglycemia induced insulin resistance. Maegawa et al. also describe assays to measure PTPase activity using labeled phosphorylated insulin receptors and by immunoenzymatic techniques.
Moxham et al, 1996, Nature, 379, 840-844, describe a murine animal and tissue culture model employing Giα2 deficiency to study hyperinsulinaemia, impaired glucose tolerance and resistance to insulin in vivo. Assays for PTPase activity and tyrosine phosphorylation of insulin-receptor subsfrate 1 are described.
Khandelwal et al, 1995, Molecular and Cellular Biochemistry, 153, 87-94, describe four different animal models for studying insulin dependent and insulin resistant diabetes mellitus. These models were used to study the effect of vanadate, an insulin mimetic and PTPase inhibitor, on the insulin-stimulated phosphorylation ofthe insulin receptor and its tyrosine kinase acitivity.
Wang et al, 1999, Biochim. Biophys. Acta, 1431, 14-23, describe fluorescein monophosphates as fluorogenic substrates for PTPs. Various methods and compounds have been developed to inhibit protein tyrosine phosphatase activity.
Wrobel et al, 1999, J. Med. Chem., 42, 3199-3202, describe PTP-IB inhibition and antihyperglycemic activity in the ob/ob mouse model by 1 l-arylbenzo[b]naphtho[2,3- d]furans and arylbenzo[b]naphtho[2,3-d]thiophenes. Andersen et al, International PCT publication No. WO 98/DK407 describe the preparation of thienopyridzinones and thienochromenones as modulators of PTPases.
Taing et al, 1999, Biochemistry, 38, 3793-3803, describe potent and highly selective inhibitors of PTP-IB comprising an array of bis(aryldifluorophosphonates).
Ham et al, 1999, Bioorg. Med. Chem. Lett., 9, 185-186, describe selective inactivation of PTP- 1 B by a sulfone analog of naphthoquinone.
Desmarais et al, 1999, Biochem, J., 337, 219-223, describe [Difluro(phosphono)methyl]phenylalanine-containing peptide inhibitors of PTPs.
Taylor et al, 1998, Bioorg. Med. Chem., 6, 2235, describe potent non-peptidyl inhibitors of PTP-IB. Kotoris et al, 1998, Bioorg. Med. Chem. Lett., 8, 3275-3280, describe novel phosphate mimetics for the design of non-peptidyl inhibitors of PTPs.
Groves et al, 1998, Biochemistry, 37, 17773-17783, describe the structural basis for PTP-IB inhibition by the phosphotyrosine peptide mimetics
(difluoronaphthylmethyl)phosphonic acid and the fluoromalonyl tyrosines with complexed crystal structures. Yao et al, 1998, Bioorgl Med. Chem., 6, 1799-1810, describe the structure-based design and synthesis of small molecule PTP-IB inhibitors comprising novel naphthyldifluoromethyl phosphonic acids 1 and 2.
Taylor et al, 1998, Bioorg. Med. Chem., 6, 1457-1468, describe potent non-peptidyl inhibitors of PTP-IB. Desmarais et al, 1998, Arch. Biochem. Biophys., 354, 225-231, describe inhibition of PTP-IB and CD45 by sulfotyrosyl peptides.
Mjalli et al, application US 96-766114, cont. in part of US patent No. 543,630, describe the preparation of heterocyclic compounds as modulators of proteins with phosphotyrosine recognition units. Wang et al, 1998, Bioorg. Med. Chem. Lett., 8, 345-350, describe naphthalenebis[α,α-difluoromethylenephosphonates] as potent inhibitors of PTPs.
Rice et al, 1997, Biochemistry, 36, 15965-15974, describe a targeted library of small molecule tyrosine and dual-specificity phosphatase inhibitors with random side chain variation from a rational core design. Olefsky, International PCT publication No. WO 97/US2752 describes a method and phosphopeptides used for the treatment of insulin resistance based on the association of PTP-IB with the activated insulin receptor. Also included is a method for determining whether a compound inhibits PTP-IB binding to the insulin receptor.
Huyer et al, 1997, J. Biol. Chem., 272, 843-851, describe the mechanism of inhibition of PTPases by vanadate and pervanadate.
Burke et al, 1996, Biochemistry, 35, 15989-15996, describe the structure-based design of PTP-IB inhibitors.
Tonks et al, International PCT publication No. WO 97/US13016, describe subsfrate-frapping protein PTPase mutants for identification of tyrosine-phosphorylated protein substrates and their clinical uses.
The human genome is thought to contain up to 100 PTPases, each varying slightly in chemistry but vastly in function. Compounds designed to inhibit PTP-IB activity specifically by covalent binding to or modification of PTP-IB have the potential for multiple side effects. Conventional drug substances that will potently suppress PTP-IB activity with few or no side effects from interaction with other PTPs are difficult to envision. A more attractive approach to PTP-IB modulation would involve the specific regulation of PTP-IB expression with oligonucleotides.
Identification of Potential Target Sites in Human PTP-IB RNA The sequence of human PTP-IB was screened for accessible sites using a computer folding algorithm. Regions ofthe RNA that did not form secondary folding structures and contained potential ribozyme and/or antisense binding/cleavage sites were identified. The sequences of these cleavage sites are shown in Tables 3-8.
Selection of Enzymatic Nucleic Acid Cleavage Sites in Human PTP-IB RNA
To test whether the sites predicted by the computer-based RNA folding algorithm corresponded to accessible sites in PTP-IB RNA, 10 hammerhead ribozyme, five NCH and three G-Cleaver ribozyme sites were selected for further analysis (Table 8). Ribozyme target sites were chosen by analyzing sequences of Human PTP-IB (Genbank accession number M33689) and prioritizing the sites on the basis of folding. Ribozymes were designed that could bind each target and were individually analyzed by computer folding (Christoffersen et al, 1994 J. Mol. Struc. Theochem, 311, 273; Jaeger et al, 1989, Proc. Natl. Acad. Sci. USA, 86, 7706) to assess whether the ribozyme sequences fold into the appropriate secondary structure. Those ribozymes with unfavorable intramolecular interactions between the binding arms and the catalytic core were eliminated from consideration. As noted below, varying binding arm lengths can be chosen to optimize activity. Generally, at least 5 bases on each arm are able to bind to, or otherwise interact with, the target RNA.
Chemical Synthesis and Purification of Ribozymes for Efficient Cleavage of PTP-IB RNA
Ribozymes were designed to anneal to various sites in the RNA message. The binding arms are complementary to the target site sequences described above. The ribozymes were chemically synthesized. The method of synthesis used followed the procedure for normal RNA synthesis as described above and in Usman et al., (1987 J. Am. Chem. Soc, 109, 7845), Scaringe et al., (1990 Nucleic Acids Res., 18, 5433) and Wincott et al., supra, and made use of common nucleic acid protecting and coupling groups, such as dimethoxytrityl at the 5'-end, and phosphoramidites at the 3'-end. The average stepwise coupling yields were >98%. Ribozymes were also synthesized from DNA templates using bacteriophage T7
RNA polymerase (Milligan and Uhlenbeck, 1989, Methods Enzymol. 180, 51). Ribozymes were purified by gel electrophoresis using general methods or were purified by high pressure liquid chromatography (HPLC; see Wincott et al., supra; the totality of which is hereby incoφorated herein by reference) and were resuspended in water. The sequences of the chemically synthesized ribozymes used in this study are shown below in Tables 3-8.
Ribozyme Cleavage of PTP-IB RNA Target in vitro
Ribozymes targeted to the human PTP-IB RNA are designed and synthesized as described above. These ribozymes can be tested for cleavage activity in vitro, for example, using the following procedure. The target sequences and the nucleotide location within the PTP-IB RNA are given in Tables 3-8.
Cleavage Reactions: Full-length or partially full-length, internally-labeled target RNA for ribozyme cleavage assay is prepared by in vitro transcription in the presence of [α-32p] CTP, passed over a G 50 Sephadex column by spin chromatography and used as subsfrate RNA without further purification. Alternately, substrates are 5'-32p-end labeled using T4 polynucleotide kinase enzyme. Assays are performed by pre-warming a 2X concentration of purified ribozyme in ribozyme cleavage buffer (50 mM Tris-HCI, pH 7.5 at 37°C, 10 mM MgCl2) and the cleavage reaction was initiated by adding the 2X ribozyme mix to an equal volume of substrate RNA (maximum of 1-5 nM) that was also pre-warmed in cleavage buffer. As an initial screen, assays are carried out for 1 hour at o
37 C using a final concentration of either 40 nM or 1 mM πbozyme, i.e., nbozyme excess.
The reaction is quenched by the addition of an equal volume of 95% formamide, 20 mM EDTA, 0.05%) bromophenol blue and 0.05% xylene cyanol after which the sample is o heated to 95 C for 2 minutes, quick chilled and loaded onto a denatuπng polyacrylamide gel. Substrate RNA and the specific RNA cleavage products generated by ribozyme cleavage are visualized on an autoradiograph ofthe gel. The percentage of cleavage is determined by Phosphor Imager® quantitation of bands representing the intact subsfrate and the cleavage products.
Example 3: MetAP-2 Methionyl aminopeptidases are metalloproteases that are known to possess post- franslational enzymatic activity by hydrolytically cleaving amino-terminal methionine residues from nascent peptide substrates in a non-processive manner (Kendall, R. L., 1992, J. Biol. Chem., 267, 20667-20673). This family of enzymes is divided into two classes (type 1 and type 2) based on differences in sequence, although the overall structure ofthe two classes are similar (Liu, S., 1998, Science, 282, 1324-1327). Methionine aminopeptidase expression appears to be involved in the confrol of cellular proliferation. Deletion ofthe MetAP gene from E. Coli is lethal (Chang, S. Y., 1989, J. Bacteriol., 171, 4071-4072). In Saccharomyces cerevisiae, deletion ofthe gene that codes for either MetAP- 1 or 2 results in a slow growth phenotype while deletion of both genes is lethal (Li, X., 1995, Proc. Natl. Acad. Sci., 92, 12357-12361). (Human methionine aminopeptidase- 1, MetAP-1, accession No. P53582).
The aminopeptidase function of this class of enzymes may serve a regulatory role in activating signal peptides in conjunction with N-myristoyl transferase (NMT) activity. NMT is expressed from a lethal gene in yeast (Duronio, R. J., 1989, Science, 243, 796- 800). NMT is responsible for amino-terminal ligation of myristic acid onto nascent peptides and cannot act on peptides with an amino-terminal methionine residue (Resh, M. D., 1996, Cell. Signal., 8, 403-412). Myristoylation of proteins correlates to intracellular localization events that may determine why certain signaling proteins are dependent on NMT for activity (Taunton, J., 1997, Chemistry & Biology, 4, 493-496). Protein tyrosine kinase Src is dependant on myristoylation for activity and has been identified as an upstream regulator of human vascular endothelial growth factor (VEGF) expression
through hypoxic induction in solid tumors (Mukhopadhyay, D., 1995, Nature, 375, 577- 581). MetAPs may therefore regulate the activation of signal peptides (such as VEGF) through cofranslational modification of nascent peptides with NMT. Disruption of protein myristoylation by MetAP inhibition could result in the improper localization of signaling proteins resulting in inhibition of cell growth. (Human N-myristoyltransferase, hNMT, accession No. AF043324.)
Fumagillin, a sesquiteφene diepoxide metabolite ofthe fungus Aspergillus fumigatus, and a related compound TNP-470, are strong inhibitors of growth in cultured endothelial cells. The antiproliferative and angiostatic activity of fumagillin was originally discovered by the serendipitous contamination of Aspergillus fumigatus in an endothelial cell culture dish in which cells closest to the fungal colony displayed growth inhibition. Synthetic analogs of fumagillin were later synthesized resulting in the discovery of TNP- 470, which is 50 times more potent of an inhibitor than fumagillin and is less toxic in mice (Ingber, D., 1990, Nature, 348, 555-557). Treatment of endothelial cells with these compounds results in late Gl phase arrest. TNP-470 inhibits the signaling pathway of retinoblastoma gene product phosphorylation, cyclin dependent kinases cdk2 and cdk4 activation, and cyclins E and A expression (Abe, J., 1994, Cancer Res., 54, 3407-3412). TNP-470 has also been shown to potently inhibit endothelial cell proliferation induced by the growth factors VEGF and bFGF (Toi, M., 1994, Oncology Reports, 1, 423-426). The bifunctional protein MetAP-2 has been identified as the molecular target for fumagillin and related compounds that demonstrate antiproliferative activity in endothelial cells. The use of affinity chromatography with a fumagillin-biotin conjugate resulted in the isolation of a 67-kDa mammalian protein through covalent interaction with the bound subsfrate. Analysis of digested peptide fragments from the isolated protein revealed MetAP-2 as the covalently bound substrate. Subsequent growth inhibition studies in yeast utilizing MetAP- 1 and MetAP-2 deletion strains determined that MetAP-2 is selectively inhibited by fumagillin in vivo (Sin, N., 1997, Proc. Natl. Acad. Sci., 94, 6099-6103). A similar study with TNP-470 and ovalicin, another potent inhibitor of neovascularization, determined that MetAP-2 is the molecular target for these fumagillin-related compounds (Griffith, E. C, 1997, Chemistry & Biology, 4, 461-471).
MetAP-2 expression correlates with cellular growth. Non-dividing cells in culture have no detectable levels ofthe 67-kDa MetAP-2 protein by immunoassay. MetAP-2 has been shown to affect franslational initiation by association with eukaryotic initiation factor 2α (eIF-2 ) (Ray, M. K., 1992, Proc. Natl. Acad. Sci., 89, 539-543). The binding of MetAP-2 with eLF-2α inhibits the heme-regulated inhibitor kinase (HRI) phosphorylation of eLF-2α in vitro in reticulocyte lysates (Datta, B., 1988, Proc. Natl. Acad. Sci., 85, 3324- 3328). MetAP-2/eIF-2α binding results in the partial reversal of protein synthesis inhibition by double stranded RNA dependent kinase mediated phosphorylation in vivo (Wu, S., 1996, Biochemistry, 35, 8275-8280). Griffith et al. also determined that covalent binding of TNP-470 and ovalicin, while potently inhibiting methionine aminopeptidase type 2 activity specifically, did not affect the regulatory activity of MetAP-2 on eIF-2α. This finding by Griffith et al. rules out the possibility that confrol of eLF-2α phosphorylation by MetAP-2 is responsible for the inhibition of endothelial cell proliferation by fumagillin related compounds. Particular angiogenesis related degenerative and disease states that can be associated with MetAP expression modulation include but are not limited to: Cancer: Solid tumors are unable to grow or metastasize without the formation of new blood vessels (Hanahan, D., 1996, Cell, 86, 353-364). Inhibition of angiogenesis via MetAP modulation can potentially be used to freat a wide variety of cancers.
Diabetic retinopathy and age related macular degeneration: Ocular neovascularization is observed in diabetic retinopathy, which is mediated by up-regulation of VEGF (Adamis, A. P., 1994, Amer. J. Ophthal, 118, 445-450). The requirement of protein kinase Src in hypoxia induced VEGF expression (Mukhopadhyay, D., 1995, Nature, 375, 577-581) indicates that MetAP modulation of aminopeptidase activity can potentially be used to freat conditions involving ocular neovascularization.
Arthritis: The ingrowth of a vascular pannus in arthritis may be mediated by the overexpression of angiogenic factors from infiltrating inflammatory cells, macrophages, and immune cells (Peacock, D. J., 1992, J. exp. Med., 175, 1135-1138). Angiogenesis inhibition through MetAP modulation can potentially be used to freat arthritis.
Psoriasis: Angiogenesis has been implicated in psoriasis due to overexpression of the angiogenic polypeptide interleukin-8 and decreased expression ofthe angiogenesis inhibitor thrombospondin (Nickoloff, B. J., 1994, Amer. J. Pathol. 44, 820-828). Angiogenesis inhibition through MetAP modulation can potentially be used to treat psoriasis.
Female reproduction: Angiogenesis in the female reproductive system has been implicated in several disorders ofthe reproductive tract (Reynolds, L. P., 1992, FASEB, 6, 886-892). Modulation of angiogenesis through confrol of MetAP may have various applications in the area of female reproduction and fertility. Various methods have been developed to assay MetAP activity.
Griffith et al, 1998, Proc. Natl. Acad. Sci., 95, 15183-15188, describe an enzymatic assay for MetAP-2 activity in vitro and an endothelial cell culture proliferation assay for MetAP-2 activity in vivo.
Weber et al, 1999, International PCT publication No. WO 98/US-21231 describe novel fluorescent reporter molecules and an enzymatic assay that can be used for determining the activity of MetAP-2 for drug screening and determining the chemosensitivity of human cancer cells to treatment with chemotherapeutic drugs.
Larrabee, J. A. et al, 1999, Anal. Biochem, 269, 194-198, describe the use of a high- pressure liquid chromatographic (HPLC) method for assaying MetAP-2 activity with application to the study of enzymic inactivation.
Quantitative methods have been developed to assay the efficacy of antiangiogenic therapies.
Wantanabe et al, 1992, Molec Biol. Cell, 3, 324a, describe the quantitation of angiogenic peptides (bFGF) in human serum as a prognostic test for breast cancer. Nguyen et al, 1994, J. Narn. Cancer Inst., 86, 356-361, describe the quantitation of angiogenic peptides (bFGF) in the urine of patients with a wide spectrum of cancers. Li et al, 1994, The Lancet, 344, 82-86, describe the quantitation of angiogenic peptides (bFGF) in the cerebrospinal fluid of children with brain tumors. This work also describes determining the extent of neovascularization in histological sections by utilizing microvessel count.
The present body of knowledge in angiogenesis research indicates the need for compounds that can modulate MetAP activity for research, diagnostic, trait alteration, animal health and therapeutic use.
Griffith et al, International PCT publication No. WO 9856372 describe small molecule inhibitors of MetAP2 and uses thereof.
D'Amato et al, International PCT publication No. WO 9805293 describe the use of AGM-1470 (TNP-470) as an angiogenesis inhibitor for use in regulating the female reproductive system and for treating diseases ofthe reproductive tissue.
Davidson et al, US patent No. 5,801,146 describe a compound and method for inhibiting angiogenesis using mammalian kringle 5 protein.
Cao et al, US patent No. 5,854,221 describe a protein-based endothelial cell proliferation inhibitor and its method of use.
Chang et al, US patent No. 5,888,796 describe a clone of a nucleotide sequence encoding a protein having two functions comprising methionine aminopeptidase activity and anti eIF-2 phosphorylation activity.
Wang et al, 1998, Proc. Am. Assoc Cancer Res., 39, 98 (absfr.) describe blocked proliferation of human endothelial cells by human MetAP-2 antisense oligonucleotides.
A rat corneal model has been developed to study ribozyme inhibition of VEGF receptor-mediated angiogenesis (Pavco, P. A., 1999, Nucleic Acids Research, 27, 2569- 2577). A similar study employing MetAP-2 inhibition could be used to study ribozyme based inhibition of MetAP-2 induced angiogenesis in vivo.
Identification of Potential Target Sites in Human MetAP-2 RNA
The sequence of human MetAP-2 was screened for accessible sites using a computer- folding algorithm. Regions ofthe RNA that did not form secondary folding structures and contained potential ribozyme and/or antisense binding/cleavage sites were identified. The sequences of these cleavage sites are shown in Tables 9-12.
Selection of Enzymatic Nucleic Acid Cleavage Sites in Human MetAP-2 RNA To test whether the sites predicted by the computer-based RNA folding algorithm corresponded to accessible sites in MetAP-2 RNA, 11 hammerhead ribozyme, 4 NCH and three G-Cleaver ribozyme sites were selected for further analysis (Table 12). Ribozyme
target sites were chosen by analyzing sequences of Human MetAP-2 (Genbank accession number HSU29607) and prioritizing the sites on the basis of folding. Ribozymes were designed that could bind each target and were individually analyzed by computer folding (Christoffersen et al, 1994 J. Mol. Struc. Theochem, 311, 273; Jaeger et al, 1989, Proc. Natl. Acad. Sci. USA, 86, 7706) to assess whether the ribozyme sequences fold into the appropriate secondary structure. Those ribozymes with unfavorable intramolecular interactions between the binding arms and the catalytic core were eliminated from consideration. As noted below, varying binding arm lengths can be chosen to optimize activity. Generally, at least 5 bases on each arm are able to bind to, or otherwise interact with, the target RNA.
Chemical Synthesis and Purification of Ribozymes for Efficient Cleavage of MetAP-2 RNA
Ribozymes were designed to anneal to various sites in the RNA message. The binding arms are complementary to the target site sequences described above. The ribozymes were chemically synthesized. The method of synthesis used followed the procedure for normal RNA synthesis as described above and in Usman et al., (1987 J. Am. Chem. Soc, 109, 7845), Scaringe et al., (1990 Nucleic Acids Res., 18, 5433) and Wincott et al., supra, and made use of common nucleic acid protecting and coupling groups, such as dimethoxytrityl at the 5'-end, and phosphoramidites at the 3'-end. The average stepwise coupling yields were >98%.
Ribozymes were also synthesized from DNA templates using bacteriophage T7 RNA polymerase (Milligan and Uhlenbeck, 1989, Methods Enzymol. 180, 51). Ribozymes were purified by gel electrophoresis using general methods or were purified by high pressure liquid chromatography (HPLC; see Wincott et al, supra; the totality of which is hereby incoφorated herein by reference) and were resuspended in water. The sequences of the chemically synthesized ribozymes used in this study are shown below in Table 9-12.
Ribozyme Cleavage of MetAP-2 RNA Target in vitro Ribozymes targeted to the human MetAP-2 RNA are designed and synthesized as described above. These ribozymes can be tested for cleavage activity in vitro, for
example, using the following procedure. The target sequences and the nucleotide location within the MetAP-2 RNA are given in Tables 9-12.
Cleavage Reactions: Full-length or partially full-length, internally-labeled target RNA for ribozyme cleavage assay is prepared by in vitro transcription in the presence of [a-32p] CTP, passed over a G 50 Sephadex column by spin chromatography and used as subsfrate RNA without further purification. Alternately, substrates are 5'-32p-end labeled using T4 polynucleotide kinase enzyme. Assays are performed by pre-warming a 2X concentration of purified ribozyme in ribozyme cleavage buffer (50 mM Tris-HCI, pH 7.5 at 37°C, 10 mM MgC ) and the cleavage reaction was initiated by adding the 2X ribozyme mix to an equal volume of substrate RNA (maximum of 1 -5 nM) that was also pre-warmed in cleavage buffer. As an initial screen, assays are carried out for 1 hour at o
37 C using a final concentration of either 40 nM or 1 mM ribozyme, i.e., ribozyme excess. The reaction is quenched by the addition of an equal volume of 95% formamide, 20 mM EDTA, 0.05%) bromophenol blue and 0.05% xylene cyanol after which the sample is o heated to 95 C for 2 minutes, quick chilled and loaded onto a denaturing polyacrylamide gel. Subsfrate RNA and the specific RNA cleavage products generated by ribozyme cleavage are visualized on an autoradiograph ofthe gel. The percentage of cleavage is determined by Phosphor Imager® quantitation of bands representing the intact substrate and the cleavage products.
Example 4: BACE, ps-1, ps-2
Alzheimer's disease (AD) is a progressive, degenerative disease ofthe brain which affects approximately 4 million people in the United States alone. An estimated 14 million Americans will have Alzheimer's disease by the middle ofthe next century if no cure or definitive prevention ofthe disease is found. Nearly one out often people over age 65 and nearly half of those over 85 have Alzheimer's disease. Alzheimer's disease is not confined to the elderly, a small percentage of people in their 30' s and 40 's are afflicted with early onset AD. Alzheimer's disease is the most common form of dementia, and amounts to the third most expensive disease in the US following heart disease and cancer. An estimated 100 billion dollars are spent annually on Alzheimer's disease (National Alzheimer's Association, 1999).
Alzheimer's disease is characterized by the progressive formation of insoluble plaques and vascular deposits in the brain consisting ofthe 4 kD amyloid β peptide (Aβ). These plaques are characterized by dysfrophic neurites that show profound synaptic loss, neurofibrillary tangle formation, and gliosis. Aβ arises from the proteolytic cleavage of the large type I transmembrane protein, β-amyloid precursor protein (APP) (Kang et al, 1987, Nature, 325, 733). Processing of APP to generate Aβ requires two sites of cleavage by a β-secretase and a γ-secretase. β-secretase cleavage of APP results in the cytoplasmic release of a 100 kD soluble amino-terminal fragment, APPsβ, leaving behind a 12 kD transmembrane carboxy-terminal fragment, C99. Alternately, APP can be cleaved by a α- secretase to generate cytoplasmic APPsα and transmembrane C83 fragments. Both remaining transmembrane fragments, C99 and C83, can be further cleaved by a γ- secretase, leading to the release and secretion of Alzheimer's related Aβ and a non- pathogenic peptide, p3, respectively (Vassar et al, 1999, Science, 286, 735-741). Early onset familial Alzheimer's disease is characterized by mutant APP protein with a Met to Leu substitution at position PI, characterized as the "Swedish" familial mutation (Mullan et al, 1992, Nature Genet., 1, 345). This APP mutation is characterized by a dramatic enhancement in β-secretase cleavage (Citron et al, 1992, Nature, 360, 672).
The identification of β-secretase, and γ-secretase constituents involved in the release of β-amyloid protein is of primary importance in the development of treatment strategies for Alzheimer's disease. Characterization of α-secretase is also important in this regard since α-secretase cleavage may compete with β-secretase cleavage resulting in non- pathogenic vs. pathogenic protein production. Involvement ofthe two metalloproteases, ADAM 10, and TACE has been demonstrated in α-cleavage of AAP (Buxbaum et al, 1999, J. Biol. Chem., 273, 27765, and Lammich et al, 1999, Proc. Natl. Acad. Sci. U.S.A., 96, 3922). Studies of γ-secretase activity have demonsfrated presenilin dependence (De Stooper et al, 1998, Nature, 391, 387, and De Stooper et al, 1999, Nature, 398, 518), and as such, presenilins have been proposed as γ-secretase even though presenilin does not present proteolytic activity (Wolfe et al, 1999, Nature, 398, 513).
Recently, Vassar et al, 1999, supra reported β-secretase cleavage of AAP by the transmembrane aspartic protease beta site APP cleaving enzyme, BACE. While other potential candidates for β-secretase have been proposed (for review see Evin et al, 1999, Proc. Natl. Acad. Sci. U.S.A., 96, 3922), none have demonstrated the full range of characteristics expected from this enzyme. Vassar et al, supra, demonstrate that BACE expression and localization are as expected for β-secretase, that BACE overexpression in cells results in increased β-secretase cleavage of APP and Swedish APP, that isolated BACE demonstrates site specific proteolytic activity on APP derived peptide substrates, and that antisense mediated endogenous BACE inhibition results in dramatically reduced β-secretase activity.
Current treatment strategies for Alzheimer's disease rely on either the prevention or the alleviation of symptoms and/or the slowing down of disease progression. Two drugs approved in the treatment of Alzheimer's, donepezil (Aricept®) and tacrine (Cognex®), both cholinomimetics, attempt to slow the loss of cognitive ability by increasing the amount of acetylcholine available to the brain. Antioxidant therapy through the use of antioxidant compounds such as alpha-tocopherol (vitamin E), melatonin, and selegeline (Eldepryl®) attempt to slow disease progression by minimizing free radical damage. Estrogen replacement therapy is thought to incur a possible preventative benefit in the development of Alzheimer's disease based on limited data. The use of anti-inflammatory drugs may be associated with a reduced risk of Alzheimer's as well. Calcium channel blockers such as Nimodipine® are considered to have a potential benefit in treating Alzheimer's disease due to protection of nerve cells from calcium overload, thereby prolonging nerve cell survival. Nootropic compounds, such as acetyl-L-carnitine (Alcar®) and insulin, have been proposed to have some benefit in treating Alzheimer's due to enhancement of cognitive and memory function based on cellular metabolism.
Whereby the above treatment strategies may all improve quality of life in
Alzheimer's patients, there exists an unmet need in the comprehensive treatment and prevention of this disease. As such, there exists the need for therapeutics effective in reversing the physiological changes associated with Alzheimer's disease, specifically, therapeutics that can eliminate and/or reverse the deposition of amyloid β peptide. The use of compounds to modulate the expression of proteases that are instrumental in the
release of amyloid β peptide, namely β-secretase (BACE), and γ-secretase (presenilin), is of therapeutic significance.
Tsai et al, 1999, Book of Absfrasts, 218th ACS National Meeting, New Orleans, Aug 22-26, describe substrate-based alpha-aminoisobutyric acid derivatives of difluoro ketone peptidomimetic inhibitors of amyloid β peptide through γ-secretase inhibition.
Czech et al, International PCT publication No. WO/9921886, describe peptides capable of inhibiting the interaction between presenilins and the β-amyloid peptide or its precursor for therapeutic use.
Fournier et al, International PCT publication No. WO/9916874, describe human brain proteins capable of interacting with presenilins and cDNAs encoding them toward therapeutic use.
St. George-Hyslop et al, International PCT publication No. WO/9727296, describe genes for proteins that interact with presenilins and their role in Alzheimer's disease toward therapeutic use. Vassar et al, 1999, Science, 286, 735-741, describe specific antisense oligonucleotides targeting BACE, used for inhibition studies of endogenous BACE expression in 101 cells and APPsw cells via lipid mediated fransfection.
Vassar et al, 1999, Science, 286, 735-741, describe a cell culture model for studying BACE inhibition. Specific antisense nucleic acid molecules targeting BACE mRNA were used for inhibition studies of endogenous BACE expression in 101 cells and APPsw
(Swedish type amyloid precursor protein expressing) cells via lipid mediated transfection. Antisense treatment resulted in dramatic reduction of both BACE mRNA by Northern blot analysis, and APPsβsw ("Swedish" type β-secretase cleavage product) by ELISA, with maximum inhibition of both parameters at 75-80%. This model was also used to study the effect of BACE inhibition on amyloid β-peptide production in APPsw cells.
Games et al, 1995, Nature, 373, 523-527, describe a transgenic mouse model in which mutant human familial type APP (Phe 717 instead of Val) is overexpressed. This model results in mice that progressively develop many ofthe pathological hallmarks of Alzheimer's disease, and as such, provides a model for testing therapeutic drugs. Particular degenerative and disease states that can be associated with BACE expression modulation include but are not limited to Alzheimer's disease and dementia.
Donepezil, tacrine, selegeline, and acetyl-L-carnitine are non-limiting examples of pharmaceutical agents that can be combined with or used in conjunction with the nucleic acid molecules (e.g. ribozymes and antisense molecules) ofthe instant invention. Those skilled in the art will recognize that other drugs such as diuretic and antihypertensive compounds and therapies can be similarly be readily combined with the nucleic acid molecules ofthe instant invention (e.g. ribozymes and antisense molecules) are hence within the scope ofthe instant invention.
Identification of Potential Target Sites in Human BACE RNA The sequence of human BACE was screened for accessible sites using a computer- folding algorithm. Regions ofthe RNA that did not form secondary folding structures and contained potential ribozyme and/or antisense binding/cleavage sites were identified. The sequences of these cleavage sites are shown in Tables 18-23.
Selection of Enzymatic Nucleic Acid Cleavage Sites in Human BACE RNA
Ribozyme target sites were chosen by analyzing sequences of Human BACE (Genbank sequence accession number: AF 190725) and prioritizing the sites on the basis of folding. Ribozymes were designed that could bind each target and were individually analyzed by computer folding (Christoffersen et al, 1994 J. Mol. Struc. Theochem, 311, 273; Jaeger et al, 1989, Proc. Natl. Acad. Sci. USA, 86, 7706) to assess whether the ribozyme sequences fold into the appropriate secondary structure. Those ribozymes with unfavorable intramolecular interactions between the binding arms and the catalytic core were eliminated from consideration. As noted below, varying binding arm lengths can be chosen to optimize activity. Generally, at least 5 bases on each arm are able to bind to, or otherwise interact with, the target RNA.
Chemical Synthesis and Purification of Ribozymes and Antisense for Efficient Cleavage and/or blocking of BACE RNA
Ribozymes and antisense constructs were designed to anneal to various sites in the RNA message. The binding arms ofthe ribozymes are complementary to the target site sequences described above, while the antisense constructs are fully complimentary to the target site sequences described above. The ribozymes and antisense constructs were
chemically synthesized. The method of synthesis used followed the procedure for normal RNA synthesis as described above and in Usman et al., (1987 J. Am. Chem. Soc, 109, 7845), Scaringe et al., (1990 Nucleic Acids Res., 18, 5433) and Wincott et al., supra, and made use of common nucleic acid protecting and coupling groups, such as dimethoxytrityl at the 5'-end, and phosphoramidites at the 3'-end. The average stepwise coupling yields were >98%.
Ribozymes and antisense constructs were also synthesized from DNA templates using bacteriophage T7 RNA polymerase (Milligan and Uhlenbeck, 1989, Methods Enzymo 180, 51). Ribozymes and antisense constructs were purified by gel elecfrophoresis using general methods or were purified by high pressure liquid chromatography (HPLC; See Wincott et al., supra; the totality of which is hereby incoφorated herein by reference) and were resuspended in water. The sequences ofthe chemically synthesized ribozymes and antisense constructs used in this study are shown below in Table 18-23.
Ribozyme Cleavage of BACE RNA Target in vitro
Ribozymes targeted to the human BACE RNA are designed and synthesized as described above. These ribozymes can be tested for cleavage activity in vitro, for example, using the following procedure. The target sequences and the nucleotide location within the BACE RNA are given in Tables 18-23.
Cleavage Reactions: Full-length or partially full-length, internally-labeled target RNA for ribozyme cleavage assay is prepared by in vitro transcription in the presence of [a-32p] CTP, passed over a G 50 Sephadex column by spin chromatography and used as substrate RNA without further purification. Alternately, substrates are 5'-32p-end labeled using T4 polynucleotide kinase enzyme. Assays are performed by pre-warming a 2X concentration of purified ribozyme in ribozyme cleavage buffer (50 mM Tris-HCI, pH 7.5 at 37°C, 10 mM MgCl2) and the cleavage reaction was initiated by adding the 2X ribozyme mix to an equal volume of substrate RNA (maximum of 1-5 nM) that was also pre-warmed in cleavage buffer. As an initial screen, assays are carried out for 1 hour at o 37 C using a final concentration of either 40 nM or 1 mM ribozyme, i.e., ribozyme excess. The reaction is quenched by the addition of an equal volume of 95% formamide, 20 mM
EDTA, 0.05%) bromophenol blue and 0.05% xylene cyanol after which the sample is heated to 95 C for 2 minutes, quick chilled and loaded onto a denaturing polyacrylamide gel. Substrate RNA and the specific RNA cleavage products generated by ribozyme cleavage are visualized on an autoradiograph ofthe gel. The percentage of cleavage is determined by Phosphor Imager® quantitation of bands representing the intact subsfrate and the cleavage products.
Example 5: Phospholamban
Cardiac disease leading to heart failure is the leading cause of combined morbidity and mortality in the developed world. Nearly twenty million people worldwide suffer from heart failure related disease. An estimated five million Americans are afflicted with congestive heart failure (CHF), with 400,000 new cases diagnosed each year. In the US, cardiac disease associated failure results in approximately 40,000 deaths per year, and is associated with an additional 250,000 deaths (Harnish, 1999, Drug & Market Development, 10, 114-119). Heart failure related disease represents a major public health issue due to an overall increase in prevalence and incidence in aging populations with a greater proportion of survivors of acute myocardial infarction (AMI) (Kannel et al, 1994, Br. Heart. J., 72 (suppl), 3). Heart failure related disease represents the most common reason for hospitalization of elderly patients in the US. The resulting life expectancy of these patients is less than that of many common cancers, with five year survival rates for men and women at only 25% and 38% respectively, and with one year mortality rates for severe heart failure at 50% (Ho et al, 1993, Circulation, 88, 107).
Heart disease is characterized by a progressive decrease in cardiac output resulting from insufficient pumping activity ofthe diseased heart. The resulting venous back- pressure results in peripheral and pulmonary dysfunctional congestion. The heart responds to a variety of mechanical, hemodynamic, hormonal, and pathological stimuli by increasing muscle mass in response to an increased demand for cardiac output. The resulting transformation of heart tissue (myocardial hypertrophy) can arise as a result of genetic, physiologic, and environmental factors, and represents an early indication of clinical heart disease and an important risk factor for subsequent heart failure (Hunter and Chien, 1999, New England J. of Medicine, 99, 313-322).
Coronary heart disease is a predominant factor in the development ofthe cardiac disease state, along with prior AMI, hypertension, diabetes mellitus, and valvular heart disease. Diagnosis of cardiac disease includes determination of coronary heart disease associated left ventricular systolic dysfunction (LVSD) and/or left ventricular diastolic dysfunction (LVDD) by echocaardiographic imaging (Cleland, 1997, Dis Management Health Outcomes, 1, 169). Promising diagnosis may also rely on assaying atrial natriuretic peptide (ANP) and brain natriuretic peptide (BNP) concentrations. ANP and BNP levels are indicative ofthe level of ventricular dysfunction (Davidson et al, 1996, Am. J. Cardiol., 77, 828). Current treatment strategies for cardiac disease associated failure are varied.
Diuretics are often used to reduce pulmonary edema and dyspnea in patients with fluid overload, and are usually used in conjunction with angiotensin converting enzyme (ACE) inhibitors for vasodilation. Digoxin is another popular choice for treating cardiac disease as an ionofropic agent, however, doubts remain concerning the long-term efficacy and safety of Digoxin (Harnish, 1999, Drug & Market Development, 10, 114- 119). Carvedilol, a beta-blocker, has been introduced to complement the above treatments in order to slow down the progression of cardiac disease. Antiarrhythmic agents can be used in order to reduce the risk of sudden death in patients suffering from cardiac disease. Lastly, heart transplants have been effective in the treatment of patients with advanced stages of cardiac disease, however, the limited supply of donor hearts greatly limits the scope of this treatment to the broad population (Hamish, 1999, Drug & Market Development, 10, 114- 119).
Whereby the above treatment strategies can all improve morbidity and mortality associated with cardiac disease, the only existing definitive approach to curing the diseased heart is replacement by transplant. Even a healthy, fransplanted heart can become diseased in response to the various stresses of mechanical, hemodynamic, hormonal, and pathological stimuli associated with extrinsic risk factors. As such there exists the need for therapeutics effective in reversing the physiological changes associated with cardiac disease. Myocardial hypertrophy and apoptosis are the underlying degenerative process associated with cardiac hypertrophy and failure. A variety of signaling pathways are involved in the progression of myocardial hypertrophy and myocardial apoptosis. Genetic
studies have been instrumental in elucidating these pathways and their involvement in cardiac disease through in vitro assays of cardiac muscle cells and in vivo studies of genetically engineered animals.
Studies in which the expression of specific genes have been altered in cardiac myocytes have shown that specific peptide hormones, growth factors, and cytokines can activate various features ofthe hyperfrophic response (Hunter and Chien, 1999, New England J of Medicine, 99, 313-322). Particular substances that have been characterized from these studies include potential therapeutic and molecular targets involved in heart failure. Hunter et al, in Chien, KR, ed. Molecular basis of heart disease: a companion to Braunwald 's Heart Disease, Philadelphia: W.B. Saunders, 1999:211 -250, describe classes of therapeutic and molecular targets involved in heart failure including:
1. Endothelin 1 and angiotensin π receptor antagonists, and antagonists of ras, p38, and c-jun Ν-terminal kinase (JΝK) for inhibition of pathologic hypertrophy.
2. Insulin like growth factor I and growth hormone receptor stimulation for promotion of physiologic hypertrophy.
3. beta- 1 -adrenergic receptor blockers for inhibition of neurohumoral over stimulation.
4. Phospholamban and Sarcolipin small molecule inhibitors for relief of sarcoplasmic reticulum calcium ATPase inhibition to provide enhancement of myocardial contractile and relaxation responses. 5. Small molecule inhibitors of β-adrenergic receptor kinase to counteract the desensitization of G protein coupled receptor kinases in order to provide enhancement of myocardial contractile and relaxation responses. 6. Enhancement of angiogenic growth factors (VEGF, FGF-5) for relief of energy deprivation in cardiac tissues. 7. Promoters of myocyte survival including gp 130 ligands (cardiofrophin 1), and
Νeuregulin for the inhibition of apoptosis of myocytes.
8. Inhibitors of apoptosis such as Caspase inhibitors for the inhibition of apoptosis of myocytes.
9. Inhibitors of cytokines such as TΝF-alpha for the inhibition of apoptosis of myocytes. Congestive heart failure, heart failure, dilated cardiomyopathy and pressure overload hypertrophy are nonlimiting examples of disorders and disease states that can be associated with the above classes of molecular targets.
The failure of cardiac contractile performance leading to cardiac disorders and disease, governed by impairment of cardiac excitation/contraction coupling, points to the importance ofthe signaling pathways involved in this process. The release and uptake of cytosolic Ca2+ by the sarcoplasmic reticulum plays an integral role in each cycle of cardiac confraction and excitation (Minamisawa et al, 1999, Cell, 99, 313-322). The process of Ca2+ reuptake is mediated by the cardiac sarcoplasmic reticulum Ca2+ ATPase (SERCA2a). SERCA2a activity is regulated by phospholamban, a p52 muscle specific sarcoplasmic reticulum phosphoprotein (Koss et al, 1996, Circ. Res., 79, 1059-1063, and Simmerman et al, 1998, Physiol. Rev., 78, 921-947). In its active, unphosphorylated state, phospholamban is a potent inhibitor of SERCA2a activity. Phosphorylation of phospholamban at serine 16 by cyclic AMP-dependent protein kinase (PKA) or calmodulin kinase, results in the inhibition of phospholamban interaction with SERCA2a. This phosphorylation event is predominantly responsible for the proportional increase in the rate of Ca2+ uptake into the sarcoplasmic reticulum and resultant ventricular relaxation (Tada et al, 1982, Mol. Cell Biochem., 46, 73-95, and Luo et al, 1998, J. Biol. Chem., 273, 4734-4739).
Since a proportional decrease in Ca2+ uptake is a hallmark feature of heart failure (Sordahl et al, 1973, Am. J. Physiol, 224, 497-502) and since an increase in the relative ratio of phospholamban to SERCA2a is an important determinant of sarcoplasmic reticulum dysfunction in heart failure (Hasenfuss, 1998, Cardiovasc. Res., 37, 279-289), the targeting of phospholamban and related regulatory factors as therapeutic targets for heart disorders should prove valuable for cardiac indications.
Pystynen et al, International PCT publication No. WO 99/00132, describe bisethers of 1-oxa, aza and thianaphthalen-2-ones as small molecule inhibitors of phospholamban for increasing coronary flow via direct dilation ofthe coronary arteries.
Pystynen et al, International PCT publication No. WO 99/15523, describe bisethers of 1-oxa, aza and thianaphthalen-2-ones as small molecule inhibitors of phospholamban that are useful for treating heart failure.
The efficacy ofthe above mentioned treatment strategies is limited. Small molecule inhibition of a molecular target is often limited by toxicity, which can restrict dosing and overall efficacy.
He et al, 1999, Circulation, 100, 974-980, describe endogenous expression of mutant phospholamban and phospholamban antisense RNA to investigate the corresponding effect on SERC A2a activity and cardiac myocyte contractility.
A more attractive approach to the treatment of heart disease would involve the use of ribozymes and/or antisense constructs to modulate the expression of target molecules involved in heart failure. The use of nucleic acid molecules ofthe instant invention permits highly specific regulation ofthe molecular targets of interest, including phospholamban (PLN) (GenBank accession No. NM_002667), sarcolipin (SLN) (GenBank accession No. NM_003063), angiotensin II receptor (GenBank accession No. U20860), endothelin 1 receptor (GenBank accession No. NM 001957), K-ras (GenBank accession No. NM 004985), p38 (GenBank accession No. AF092535), c-jun N-terminal kinase (GenBank accession No. NM_002750, L31951, NM_002753), growth hormone receptor (GenBank accession No. NM_000163), insulin- like growth factor I receptor (GenBank accession No. NM_000875), beta- 1 -adrenergic receptor (GenBank accession No. NM_000024), βl-adrenergic receptor kinase (GenBank accession No. NM_001619, NM_005160), VEGF receptor (GenBank accession No. U43368, M27281 X15997), fibroblast growth factor 5 (GenBank accession No. NM_004464), cardiofrophin I (GenBank accession No. NM_001330), neuregulin (GenBank accession No. AF009227), TNF-alpha (GenBank accession No. X02910 X02159), PI3 kinase (GenBank accession No. NM_006218, NM_006219, U86453, NM_002649, M61906), and AKT kinase (GenBank accession No. NM_005163, M77198).
Various methods have been developed to assay phospholamban activity in vitro and in vivo. Holt et al, 1999, J. Mol. Cell. Cardiol, 31, 645-656, describe a cell culture model in which thyroid hormone control of confraction and the Ca2+-ATPase/phospholamban complex is studied in adult rat ventricular myocytes. Slack et al. 1997, J. Biol. Chem., 272, 18862-18868, describe studies in which the ectopic expression of phospholamban in mouse fast-twitch skeletal muscle cells alters sarcoplasmic reticulum Ca2+ transport and muscle relaxation. MacLennan et al, 1996, Soc. Gen. Physiol. Ser., 51, 89-103, in a review of regulatory interactions between calcium ATPases and phospholamban describe phospholamban/ Ca2+- ATPase interactions in protein expressed in heterologous cell culture experiments. Comwell et al, 1991, Mol. Pharmacol, 40,923-931, describe the
regulation of sarcoplasmic reticulum protein phosphorylation by localized cyclic GMP- dependent protein kinase in vascular smooth muscle cells.
Minamisawa et al, 1999, Cell, 99, 313-322, describe a phospholamban knockout mouse model which affords protection from induced dilated cardiomyopathy. Dillmann et al, 1999, Am. J. Cardiol, 83, 89H-91H, describe a fransgenic rat model for the study of altered expression of calcium regulatory proteins, including phospholamban, and their effect on myocyte contractile response. LekanneDeprez et al, 1998, J. Mol. Cell. Cardiol, 30, 1877-1888, describe a rat pressure-overload model to investigate alterations in gene expression of phospholamban, atrial natriuretic peptide (ANP), sarcoplasmic endoplasmic reticular calcium ATPase 2 (SERCA2), collagen LLIαl, and calsequestrin (CSQ). Jones et al, 1998, J. Clin. Invest., 101, 1385-1393, describe a mouse model for investigating the regulation of calcium signaling in transgenic mouse cardiac myocytes overexpressing calsequestrin. In this study, the upregulation and downregulation of calcium uptake and release proteins were determined, including phospholamban. Lorenz et al, 1997, Am J. Physiol, 273, 6, describe a mouse model for the study of regulatory effects of phospholamban on cardiac function in intact mice. This study makes use of animal models with altered levels of phospholamban to permit in vivo evaluation ofthe physiological role of phospholamban. Arai et al, 1996, Saishin Igaku, 51, 1095-1104, presents a review article of gene targeted animal models expressing cardiovascular abnormalities. The study of phospholamban and other protein expression modification effects in mice is presented. Wankerl et al, 1995, J. Mol. Med., 73, 487-496, presents a review article describing the study of calcium transport proteins in the nonfailing and failing heart. Animal models investigating the major calcium handling myocardial proteins, including phospholamban, are described. These models, as well as others, may be used to evaluate the effect of treatment with nucleic acid molecules ofthe instant invention on cardiac function. Endpoints may be, but are not limited to, left ventricular pressure, left ventricular pressure as a function of time (LVdP/dt), and mean arterial blood pressure. Endpoints will be evaluated under basal and stimulated (cardiac load) conditions. Particular degenerative and disease states that can be associated with phospholamban expression modulation include but are not limited to congestive heart failure, heart failure, dilated cardiomyopathy and pressure overload hypertrophy:
Digoxin, Bendrofluazide, Dofetilide, and Carvedilol are non-limiting examples of pharmaceutical agents that can be combined with or used in conjunction with the nucleic acid molecules (e.g. ribozymes and antisense molecules) ofthe instant invention. Those skilled in the art will recognize that other drugs such as diuretic and antihypertensive compounds and therapies can be similarly be readily combined with the nucleic acid molecules ofthe instant invention (e.g. ribozymes and antisense molecules) are hence within the scope ofthe instant invention.
Identification of Potential Target Sites in Human phospholamban RNA The sequence of human phospholamban was screened for accessible sites using a computer folding algorithm. Regions ofthe RNA that did not form secondary folding structures and contained potential ribozyme and/or antisense binding/cleavage sites were identified. The sequences of these cleavage sites are shown in Tables 24-30.
Selection of Enzymatic Nucleic Acid Cleavage Sites in Human phospholamban RNA Ribozyme target sites were chosen by analyzing sequences of Human phospholamban (Genbank sequence accession number: NM 002667) and prioritizing the sites on the basis of folding. Ribozymes were designed that could bind each target and were individually analyzed by computer folding (Christoffersen et al, 1994 J. Mol. Struc. Theochem, 311, 273; Jaeger et al, 1989, Proc. Natl. Acad. Sci. USA, 86, 7706) to assess whether the ribozyme sequences fold into the appropriate secondary structure. Those ribozymes with unfavorable intramolecular interactions between the binding arms and the catalytic core were eliminated from consideration. As noted below, varying binding arm lengths can be chosen to optimize activity. Generally, at least 5 bases on each arm are able to bind to, or otherwise interact with, the target RNA.
Chemical Synthesis and Purification of Ribozymes and Antisense for Efficient Cleavage and/or blocking of phospholamban RNA
Ribozymes and antisense constructs were designed to anneal to various sites in the RNA message. The binding arms ofthe ribozymes are complementary to the target site sequences described above, while the antisense constructs are fully complimentary to the target site sequences described above. The ribozymes and antisense constructs were
chemically synthesized. The method of synthesis used followed the procedure for normal RNA synthesis as described above and in Usman et al., (1987 J. Am. Chem. Soc, 109, 7845), Scaringe et al., (1990 Nucleic Acids Res., 18, 5433) and Wincott et al., supra, and made use of common nucleic acid protecting and coupling groups, such as dimethoxytrityl at the 5 '-end, and phosphoramidites at the 3 '-end. The average stepwise coupling yields were >98%.
Ribozymes and antisense constructs were also synthesized from DNA templates using bacteriophage T7 RNA polymerase (Milligan and Uhlenbeck, 1989, Methods Enzymol. 180, 51). Ribozymes and antisense constructs were purified by gel elecfrophoresis using general methods or were purified by high pressure liquid chromatography (HPLC; see Wincott et al., supra; the totality of which is hereby incoφorated herein by reference) and were resuspended in water. The sequences ofthe chemically synthesized ribozymes and antisense constructs used in this study are shown below in Table 24-30.
Ribozyme Cleavage of phospholamban RNA Target in vitro
Ribozymes targeted to the human phospholamban RNA are designed and synthesized as described above. These ribozymes can be tested for cleavage activity in vitro, for example using the following procedure. The target sequences and the nucleotide location within the phospholamban RNA are given in Tables 24-30.
Cleavage Reactions: Full-length or partially full-length, internally-labeled target RNA for ribozyme cleavage assay is prepared by in vitro transcription in the presence of [a-32p] CTP, passed over a G 50 Sephadex column by spin chromatography and used as substrate RNA without further purification. Alternately, substrates are 5'-32p_end labeled using T4 polynucleotide kinase enzyme. Assays are performed by pre-warming a 2X concenfration of purified ribozyme in ribozyme cleavage buffer (50 mM Tris-HCI, pH 7.5 at 37°C, 10 mM MgC ) and the cleavage reaction was initiated by adding the 2X ribozyme mix to an equal volume of substrate RNA (maximum of 1-5 nM) that was also pre-warmed in cleavage buffer. As an initial screen, assays are carried out for 1 hour at
O 37 C using a final concenfration of either 40 nM or 1 mM ribozyme, i.e., ribozyme excess. The reaction is quenched by the addition of an equal volume of 95% formamide, 20 mM
EDTA, 0.05%o bromophenol blue and 0.05% xylene cyanol after which the sample is o heated to 95 C for 2 minutes, quick chilled and loaded onto a denaturing polyacrylamide gel. Subsfrate RNA and the specific RNA cleavage products generated by ribozyme cleavage are visualized on an autoradiograph ofthe gel. The percentage of cleavage is determined by Phosphor Imager® quantitation of bands representing the intact substrate and the cleavage products.
Tissue distribution of BrdU-labeled antisense in mice
CD1 mice were injected with a single bolus (30 mg/kg) of a BrdU-labeled antisense oligonucleotide or a similar molar amount of BrdU (as a control). At various time points (30 min, 2h and 6 h), mice were sacrificed and major tissues isolated and fixed. Distribution of antisense oligonucleotides was determined by probing with an anti-BrdU antibody and immunohistochemical staining. Tissue slices were probed with an anti-BrdU antibody followed by a reporter enzyme-conjugated second antibody and finally an enzyme subsfrate. Visualization ofthe colored product by microscopy indicated nuclear staining, demonstrating effective distribution of antisense oligonucleotide in cardiac tissue.
Tissue distribution of BrdU-labeled ribozymes in monkey
Rhesus monkeys were dosed with BrdU-labeled ribozyme by intravenous bolus injection at 0.1 , 1.0, and 10 mg/kg once daily over five days. Saline injection was used in control animals. Animals were sacrificed and major tissues isolated and fixed. Tissue samples were probed with an anti-BrdU antibody followed by a reporter enzyme- conjugated second antibody and finally an enzyme subsfrate. Significant quantities of chemically modified ribozyme are detected in cardiac tissue following this dosing regimen.
Example 6: HBV
Chronic hepatitis B is caused by an enveloped virus, commonly known as the hepatitis B virus or HBV. HBV is transmitted via infected blood or other body fluids, especially saliva and semen, during delivery, sexual activity, or sharing of needles contaminated by infected blood. Individuals may be "carriers" and transmit the infection to others without ever having experienced symptoms ofthe disease. Persons at highest
risk are those with multiple sex partners, those with a history of sexually transmitted diseases, parenteral drug users, infants bom to infected mothers, "close" contacts or sexual partners of infected persons, and healthcare personnel or other service employees who have contact with blood. Transmission is also possible via tattooing, ear or body piercing,
5 and acupuncture; the virus is also stable on razors, toothbrushes, baby bottles, eating utensils, and some hospital equipment such as respirators, scopes and instruments. There is no evidence that HBsAg positive food handlers pose a health risk in an occupational setting, nor should they be excluded from work. Hepatitis B has never been documented as being a food-borne disease. The average incubation period is 60 to 90 days, with a
! 0 range of 45 to 180; the number of days appears to be related to the amount of virus to which the person was exposed. However, determining the length of incubation is difficult, since onset of symptoms is insidious. Approximately 50% of patients develop symptoms of acute hepatitis that last from 1 to 4 weeks. Two percent or less of these individuals develop fulminant hepatitis resulting in liver failure and death.
5 The determinants of severity include: (1) The size ofthe dose to which the person was exposed; (2) the person's age with younger patients experiencing a milder form ofthe disease; (3) the status ofthe immune system with those who are immunosuppressed experiencing milder cases; and (4) the presence or absence of co-infection with the Delta virus (hepatitis D), with more severe cases resulting from co-infection. In symptomatic
!0 cases, clinical signs include loss of appetite, nausea, vomiting, abdominal pain in the right upper quadrant, arthralgia, and tiredness/loss of energy. Jaundice is not experienced in all cases, however, jaundice is more likely to occur if the infection is due to transfusion or percutaneous serum transfer, and it is accompanied by mild pruritus in some patients. Bilirubin elevations are demonsfrated in dark urine and clay-colored stools, and liver
15 enlargement may occur accompanied by right upper-quadrant pain. The acute phase ofthe disease may be accompanied by severe depression, meningitis, Guillain-Barre syndrome, myelitis, encephalitis, agranulocytosis, and/or thrombocytopenia.
Hepatitis B is generally self-limiting and will resolve in approximately 6 months. Asymptomatic cases can be detected by serologic testing, since the presence ofthe virus
>0 leads to production of large amounts of HBsAg in the blood. This antigen is the first and most useful diagnostic marker for active infections. However, if HBsAg remains positive for 20 weeks or longer, the person is likely to remain positive indefinitely and is now a
carrier. While only 10% of persons over age 6 who contract HBV become carriers, 90% of infants infected during the first year of life do so.
Hepatitis B virus (HBV) infects over 300 million people worldwide (Imperial, 1999, Gastroenterol. Hepatol, 14 (suppl), Sl-5). In the United States approximately 1.25 million individuals are chronic carriers of HBV as evidenced by the fact that they have measurable hepatitis B virus surface antigen HBsAg in their blood. The risk of becoming a chronic HBsAg carrier is dependent upon the mode of acquisition of infection as well as the age ofthe individual at the time of infection. For those individuals with high levels of viral replication, chronic active hepatitis with progression to cirrhosis, liver failure and hepatocellular carcinoma (HCC) is common, and liver transplantation is the only treatment option for patients with end-stage liver disease from HBV.
The natural progression of chronic HBV infection over a 10 to 20 year period leads to cirrhosis in 20-to-50% of patients and progression of HBV infection to hepatocellular carcinoma has been well documented. There have been no studies that have determined sub-populations that are most likely to progress to cirrhosis and/or hepatocellular carcinoma, thus all patients have equal risk of progression.
It is important to note that the survival for patients diagnosed with hepatocellular carcinoma is only 0.9 to 12.8 months from initial diagnosis (Takahashi et al, 1993, American Journal of Gastroenterology, 88, 240-243). Treatment of hepatocellular carcinoma with chemotherapeutic agents has not proven effective and only 10% of patients will benefit from surgery due to extensive tumor invasion ofthe liver (Trinchet et al, 1994, Presse Medicine, 23, 831-833). Given the aggressive nature of primary hepatocellular carcinoma, the only viable treatment alternative to surgery is liver transplantation (Pichlmayr et al, 1994, Hepatology., 20, 33S-40S). Upon progression to cirrhosis, patients with chronic HCV infection present with clinical features, which are common to clinical cirrhosis regardless ofthe initial cause (D'Amico et al, 1986, Digestive Diseases and Sciences, 31, 468-475). These clinical features may include: bleeding esophageal varices, ascites, jaundice, and encephalopathy (Zakim D, Boyer TD. Hepatology a textbook of liver disease, Second Edition Volume 1. 1990 W.B. Saunders Company. Philadelphia). In the early stages of cirrhosis, patients are classified as compensated, meaning that although liver tissue damage has occurred, the patient's liver is still able to detoxify metabolites in the blood-stream. In addition, most
patients with compensated liver disease are asymptomatic and the minority with symptoms report only minor symptoms such as dyspepsia and weakness. Ln the later stages of cirrhosis, patients are classified as decompensated meaning that their ability to detoxify metabolites in the bloodstream is diminished and it is at this stage that the clinical features
5 described above will present.
In 1986, D'Amico et al. described the clinical manifestations and survival rates in 1155 patients with both alcoholic and viral associated cirrhosis (D'Amico supra). Ofthe 1155 patients, 435 (37%) had compensated disease although 70% were asymptomatic at the beginning ofthe study. The remaining 720 patients (63%) had decompensated liver
10 disease with 78% presenting with a history of ascites, 31 % with jaundice, 17% had bleeding and 16% had encephalopathy. Hepatocellular carcinoma was observed in six (0.5%)) patients with compensated disease and in 30 (2.6%) patients with decompensated disease.
Over the course of six years, the patients with compensated cirrhosis developed
5 clinical features of decompensated disease at a rate of 10% per year. In most cases, ascites was the first presentation of decompensation. In addition, hepatocellular carcinoma developed in 59 patients who initially presented with compensated disease by the end of the six-year study.
With respect to survival, the D'Amico study indicated that the five-year survival rate
!0 for all patients on the study was only 40%. The six-year survival rate for the patients who initially had compensated cirrhosis was 54% while the six-year survival rate for patients who initially presented with decompensated disease was only 21%. There were no significant differences in the survival rates between the patients who had alcoholic cirrhosis and the patients with viral related cirrhosis. The major causes of death for the
!5 patients in the D'Amico study were liver failure in 49%; hepatocellular carcinoma in 22%; and, bleeding in 13% (D'Amico supra).
Hepatitis B virus is a double-stranded circular DNA virus. It is a member ofthe Hepadnaviridae family. The virus consists of a central core that contains a core antigen (HBcAg) surrounded by an envelope containing a surface protein surface antigen (HBsAg)
»0 and is 42 nm in diameter. It also contains an e antigen (HBeAg) which, along with HBcAg and HBsAg, is helpful in identifying this disease
In HBV virions, the genome is found in an incomplete double-stranded form. HBV uses a reverse transcriptase to transcribe a positive-sense full length RNA version of its genome back into DNA. This reverse transcriptase also contains DNA polymerase activity and thus begins replicating the newly synthesized minus-sense DNA strand. However, it appears that the core protein encapsidates the reverse-transcriptase/polymerase before it completes replication.
From the free-floating form, the virus must first attach itself specifically to a host cell membrane. Viral attachment is one ofthe crucial steps which determines host and tissue specificity. However, currently there are no in vitro cell-lines that can be infected by HBV. There are some cells lines, such as HepG2, which can support viral replication only upon transient or stable transfection using HBV DNA.
After attachment, fusion ofthe viral envelope and host membrane must occur to allow the viral core proteins containing the genome and polymerase to enter the cell. Once inside, the genome is translocated to the nucleus where it is repaired and cyclized. The complete closed circular DNA genome of HBV remains in the nucleus and gives rise to four transcripts. These transcripts initiate at unique sites but share the same 3'-ends. The 3.5-kb pregenomic RNA serves as a template for reverse transcription and also encodes the nucleocapsid protein and polymerase. A subclass of this transcript with a 5 '-end extension codes for the precore protein that, after processing, is secreted as HBV e antigen. The 2.4-kb RNA encompasses the pre-Sl open reading frame (ORF) that encodes the large surface protein. The 2.1-kb RNA encompasses the pre-S2 and S ORFs that encode the middle and small surface proteins, respectively. The smallest transcript (~0.8- kb) codes for the X protein, a transcriptional activator.
Multiplication ofthe HBV genome begins within the nucleus of an infected cell. RNA polymerase LI transcribes the circular HBV DNA into greater-than-full length mRNA. Since the mRNA is longer than the actual complete circular DNA, redundant ends are formed. Once produced, the pregenomic RNA exits the nucleus and enters the cytoplasm.
The packaging of pregenomic RNA into core particles is triggered by the binding of the HBV polymerase to the 5' epsilon stem-loop. RNA encapsidation is believed to occur as soon as binding occurs. The HBV polymerase also appears to require associated core protein in order to function. The HBV polymerase initiates reverse transcription from the
5' epsilon stem-loop three to four base pairs at which point the polymerase and attached nascent DNA are transferred to the 3' copy ofthe DR1 region. Once there, the (-)DNA is extended by the HBV polymerase while the RNA template is degraded by the HBV polymerase RNAse H activity. When the HBV polymerase reaches the 5' end, a small stretch of RNA is left undigested by the RNAse H activity. This segment of RNA is comprised of a small sequence just upstream and including the DR1 region. The RNA oligomer is then translocated and annealed to the DR2 region at the 5' end ofthe (-)DNA. It is used as a primer for the (+)DNA synthesis which is also generated by the HBV polymerase. It appears that the reverse transcription as well as plus strand synthesis may occur in the completed core particle.
Since the pregenomic RNA is required as a template for DNA synthesis, this RNA is an excellent target for ribozyme cleavage. Nucleoside analogues that have been documented to inhibit HBV replication target the reverse transcriptase activity needed to convert the pregenomic RNA into DNA. Ribozyme cleavage ofthe pregenomic RNA template would be expected to result in a similar inhibition of HBV replication. Further, targeting the 3 '-end ofthe pregenomic RNA that is common to all HBV transcripts could result in reduction of all HBV gene products and an additional level of inhibition of HBV replication.
As previously mentioned, HBV does not infect cells in culture. However, fransfection of HBV DNA (either as a head-to-tail dimer or as an "overlength" genome of >100%) into HuH7 or Hep G2 hepatocytes results in viral gene expression and production of HBV virions released into the media. Thus, HBV replication competent DNA would be co-fransfected with ribozymes in cell culture. Such an approach has been used to report infracellular ribozyme activity against HBV (zu Putlitz, et al, 1999, J. Virol, 73, 5381- 5387, and Kim et al, 1999, Biochem. Biophys. Res. Commun., 257, 759-765). In addition, stable hepatocyte cell lines have been generated that express HBV. In these cells only ribozyme would need to be delivered; however, a delivery screen would need to be performed. In addition, stable hepatocyte cell lines have been generated that express HBV. Infracellular HBV gene expression can be assayed by a Taqman® assay for HBV RNA or by ELISA for HBV protein. Exfracellular virus can be assayed by PCR for DNA or ELISA for protein. Antibodies are commercially available for HBV surface antigen and
core protein. A secreted alkaline phosphatase expression plasmid can be used to normalize for differences in transfection efficiency and sample recovery.
There are several small animal models to study HBV replication. One is the fransplantation of HBV-infected liver tissue into irradiated mice. Viremia (as evidenced
5 by measuring HBV DNA by PCR) is first detected 8 days after fransplantation and peaks between 18 - 25 days (Ilan et al, 1999, Hepatology, 29, 553-562).
Transgenic mice that express HBV have also been used as a model to evaluate potential anti-virals. HBV DNA is detectable in both liver and serum (Morrey et al, 1999, Antiviral Res., 42, 97-108).
10 An additional model is to establish subcutaneous tumors in nude mice with Hep G2 cells transfected with HBV. Tumors develop in about 2 weeks after inoculation and express HBV surface and core antigens. HBV DNA and surface antigen is also detected in the circulation of tumor-bearing mice (Yao et al, 1996, J. Viral Hepat., 3, 19-22).
Woodchuck hepatitis virus (WHV) is closely related to HBV in its virus structure,
15 genetic organization, and mechanism of replication. As with HBV in humans, persistent WHV infection is common in natural woodchuck populations and is associated with chronic hepatitis and hepatocellular carcinoma (HCC). Experimental studies have established that WHV causes HCC in woodchucks and woodchucks chronically infected with WHV have been used as a model to test a number of anti-viral agents. For example,
!0 the nucleoside analogue 3T3 was observed to cause dose dependent reduction in virus (50% reduction after two daily treatments at the highest dose) (Hurwitz et al, 1998. Antimicrob. Agents Chemother., 42, 2804-2809).
Current therapeutic goals of treatment are three-fold: to eliminate infectivity and transmission of HBV to others, to arrest the progression of liver disease and improve the
!5 clinical prognosis, and to prevent the development of hepatocellular carcinoma (HCC). Interferon alpha use is the most common therapy for HBV; however, recently Lamivudine (3TC) has been approved by the FDA. Interferon alpha (IFN-alpha) is one treatment for chronic hepatitis B. The standard duration of IFN-alpha therapy is 16 weeks, however, the optimal treatment length is still poorly defined. A complete response (HBV
>0 DNA negative HBeAg negative) occurs in approximately 25% of patients. Several factors have been identified that predict a favorable response to therapy including: High ALT , low HBV DNA , being female, and heterosexual orientation.
There is also a risk of reactivation ofthe hepatitis B virus even after a successful response, this occurs in around 5% of responders and normally occurs within 1 year.
Side effects resulting from treatment with type 1 interferons can be divided into four general categories including: Influenza-like symptoms, neuropsychiatric, laboratory abnormalities, and other miscellaneous side effects. Examples of influenza-like symptoms include, fatigue, fever; myalgia, malaise, appetite loss, tachycardia, rigors, headache and arthralgias. The influenza-like symptoms are usually short-lived and tend to abate after the first four weeks of dosing (Dusheiko et al, 1994, Journal of Viral Hepatitis, 1, 3-5). Neuropsychiatric side effects include irritability, apathy, mood changes, insomnia, cognitive changes, and depression. Laboratory abnormalities include the reduction of myeloid cells, including granulocytes, platelets and to a lesser extent, red blood cells. These changes in blood cell counts rarely lead to any significant clinical sequellae. In addition, increases in triglyceride concentrations and elevations in serum alaine and aspartate aminotransferase concentration have been observed. Finally, thyroid abnormalities have been reported. These thyroid abnormalities are usually reversible after cessation of interferon therapy and can be controlled with appropriate medication while on therapy. Miscellaneous side effects include nausea, diarrhea, abdominal and back pain, pruritus, alopecia, and rhinorrhea. In general, most side effects will abate after 4 to 8 weeks of therapy (Dushieko et al, supra ). Lamivudine (3TC) is a nucleoside analogue, which is a very potent and specific inhibitor of HBV DNA synthesis. Lamivudine has recently been approved for the treatment of chronic Hepatitis B. Unlike treatment with interferon, treatment with 3TC does not eliminate the HBV from the patient. Rather, viral replication is controlled and chronic administration results in improvements in liver histology in over 50%> of patients. Phase m studies with 3TC, showed that treatment for one year was associated with reduced liver inflammation and a delay in scarring ofthe liver. In addition, patients treated with Lamivudine (lOOmg per day) had a 98 percent reduction in hepatitis B DNA and a significantly higher rate of seroconversion, suggesting disease improvements after completion of therapy. However, stopping of therapy resulted in a reactivation of HBV replication in most patients. In addition recent reports have documented 3TC resistance in approximately 30% of patients.
Particular degenerative and disease states that can be associated with HBV expression modulation include but are not limited to, HBV infection, hepatitis, cancer, tumorigenesis, cirrhosis, liver failure and others.
Lamivudine (3TC), L-FMAU, adefovir dipivoxil, type 1 Interferon, therapeutic 5 vaccines, steriods, and 2 '-5' Oligoadenylates are non- limiting examples of pharmaceutical agents that can be combined with or used in conjunction with the nucleic acid molecules (e.g. ribozymes and antisense molecules) ofthe instant invention. Those skilled in the art will recognize that other drugs such as diuretic and antihypertensive compounds or other therapies can similarly and readily be combined with the nucleic acid molecules ofthe
10 instant invention (e.g. ribozymes and antisense molecules) and are, therefore, within the scope ofthe instant invention.
Current therapies for treating HBV infection, including interferon and nucleoside analogues, are only partially effective. In addition, drug resistance to nucleoside analogues is now emerging, making freatment of chronic Hepatitis B more difficult. Thus, a need
15 exists for effective freatment of this disease which utilizes antiviral inhibitors which work by mechanisms other than those currently utilized in the freatment of both acute and chronic hepatitis B infections.
Draper, US patent No. 6,017,756, describes the use of ribozymes for the inhibition of Hepatitis B Virus.
10 Passman et al, 2000, Biochem. Biophys. Res. Commun., 268(3), 728-733.; Gan et al, 1998, J. Med. Coll. PLA, 13(3), 157-159.; Li et al, 1999, Jiefangjun Yixue Zazhi, 24(2), 99-101.; Puftitz et α/., 1999, J. Virol, 73(7), 5381-5387.; Kim et al, 1999, Biochem. Biophys. Res. Commun., 257(3), 759-765.; Xu et al, 1998, Bingdu Xuebao, 14(4), 365-369.; Welch et al, 1997, Gene Ther., 4(7), 736-743.; Goldenberg et al, 1997,
>5 International PCT publication No. WO 97/08309, Wands et al, 1997, J. of Gastroenterology and Hepatology, 12(suppl.), S354-S369.; Ruiz et al, 1997, BioTechniques, 22(2), 338-345.; Gan et al, 1996, J. Med. Coll. PLA, 11(3), 171-175.; Beck and Nassal, 1995, Nucleic Acids Res., 23(24), 4954-62.; Goldenberg, 1995, International PCT publication No. WO 95/22600.; Xu et al, 1993, Bingdu Xuebao, 9(4),
$0 331-6.; Wang et al, 1993, Bingdu Xuebao, 9(3), 278-80, all describe ribozymes that are targeted to cleave a specific HBV target site.
The enzymatic nucleic acid molecules ofthe instant invention exhibit a high degree of specificity for only the viral mRNA in infected cells. Nucleic acid molecules ofthe instant invention targeted to highly conserved sequence regions allow the freatment of many strains of human HBV with a single compound. No freatment presently exists which specifically attacks expression ofthe viral gene(s) that are responsible for transformation of hepatocytes by HBV.
The methods of this invention can be used to treat human hepatitis B virus infections, which include productive virus infection, latent or persistent virus infection, and HBV-induced hepatocyte transformation. The utility can be extended to other species of HBV which infect non-human animals where such infections are of veterinary importance.
Preferred target sites are genes required for viral replication, a non-limiting example includes genes for protein synthesis, such as the 5' most 1500 nucleotides ofthe HBV pregenomic mRNAs. For sequence references, see Renbao et al, 1987, Sci. Sin., 30, 507. This region confrols the franslational expression ofthe core protein (C), X protein (X) and DNA polymerase (P) genes and plays a role in the replication ofthe viral DNA by serving as a template for reverse transcriptase. Disruption of this region in the RNA results in deficient protein synthesis as well as incomplete DNA synthesis (and inhibition of transcription from the defective genomes). Target sequences 5' ofthe encapsidation site can result in the inclusion ofthe disrupted 3' RNA within the core virion structure and targeting sequences 3' ofthe encapsidation site can result in the reduction in protein expression from both the 3' and 5' fragments.
Alternative regions outside ofthe 5' most 1500 nucleotides ofthe pregenomic mRNA also make suitable targets of enzymatic nucleic acid mediated inhibition of HBV replication. Such targets include the mRNA regions that encode the viral S gene.
Selection of particular target regions will depend upon the secondary structure ofthe pregenomic mRNA. Targets in the minor mRNAs can also be used, especially when folding or accessibility assays in these other RNAs reveal additional target sequences that are unavailable in the pregenomic mRNA species. A desirable target in the pregenomic RNA is a proposed bipartite stem-loop structure in the 3 '-end ofthe pregenomic RNA which is believed to be critical for viral replication (Kidd and Kidd-Ljunggren, 1996. Nuc. Acid Res. 24:3295-3302). The 5'end ofthe HBV
pregenomic RNA carries a cw-acting encapsidation signal, which has inverted repeat sequences that are thought to form a bipartite stem-loop structure. Due to a terminal redundancy in the pregenomic RNA, the putative stem-loop also occurs at the 3'-end. While it is the 5' copy which functions in polymerase binding and encapsidation, reverse franscription actually begins from the 3' stem-loop. To start reverse transcription, a 4 nt primer which is covalently attached to the polymerase is made, using a bulge in the 5' encapsidation signal as template. This primer is then shifted, by an unknown mechanism, to the DR1 primer binding site in the 3' stem-loop structure, and reverse franscription proceeds from that point. The 3' stem-loop, and especially the DR1 primer binding site, appear to be highly effective targets for ribozyme intervention.
Sequences ofthe pregenomic RNA are shared by the mRNAs for surface, core, polymerase, and X proteins. Due to the overlapping nature ofthe HBV transcripts, all share a common 3'-end. Ribozyme targeting this common 3'-end will thus cleave the pregenomic RNA as well as all ofthe mRNAs for surface, core, polymerase and X proteins.
In preferred embodiments, the invention features a method for the analysis of HBV proteins. This method is useful in determining the efficacy of HBV inhibitors. Specifically, the instant invention features an assay for the analysis of HBsAg proteins and secreted alkaline phosphatase (SEAP) confrol proteins to determine the efficacy of agents used to modulate HBV expression.
The method consists of coating a micro-titer plate with an antibody such as anti- HBsAg Mab (for example, Biostride B88-95-31ad,ay) at 0.1 to 10 μg/ml in a buffer (for example, carbonate buffer, such as Na2CO3 15 mM, NaHCO3 35 mM, pH 9.5) at 4°C overnight. The microtiter wells are then washed with PBST or the equivalent thereof, (for example, PBS, 0.05% Tween 20) and blocked for 0.1-24 hr at 37° C with PBST, 1% BSA or the equivalent thereof. Following washing as above, the wells are dried (for example, at 37° C for 30 min). Biotinylated goat anti-HBsAg or an equivalent antibody (for example, Accurate YVS1807) is diluted (for example at 1:1000) in PBST and incubated in the wells (for example, 1 hr. at 37° C). The wells are washed with PBST (for example, 4x). A conjugate, (for example, Streptavidin/ Alkaline Phosphatase Conjugate, Pierce 21324) is diluted to 10-10,000 ng/ml in PBST, and incubated in the wells (for example, 1 hr. at 37° C). After washing as above, a substrate (for example, p-nitrophenyl phosphate substrate,
Pierce 37620) is added to the wells, which are then incubated (for example, 1 hr. at 37° C). The optical density is then determined (for example, at 405 nm). SEAP levels are then assayed, for example, using the Great EscAPe® Detection Kit (Clontech K2041-1), as per the manufacturers instructions. In the above example, incubation times and reagent
5 concentrations may be varied to achieve optimum results, a non-limiting example is described in Example 6.
Comparison of this HBsAg ELISA method to a commercially available assay from World Diagnostics, Inc. 15271 NW 60th Ave, #201, Miami Lakes, FL 33014 (305) 827- 3304 (Cat. No. EL10018) demonstrates an increase in sensitivity (signal :noise) of 3-20
10 fold.
Identification of Potential Target Sites in Human HBV RNA
The sequence of human HBV was screened for accessible sites using a computer- folding algorithm. Regions ofthe RNA that did not form secondary folding structures and ! 5 contained potential ribozyme and/or antisense binding/cleavage sites were identified. The sequences of these cleavage sites are shown in Tables 36-43.
Selection of Enzymatic Nucleic Acid Cleavage Sites in Human HBV RNA
Ribozyme target sites were chosen by analyzing sequences of Human HBV
!0 (accession number: AF 100308.1) and prioritizing the sites on the basis of folding.
Ribozymes were designed that could bind each target and were individually analyzed by computer folding (Christoffersen et al, 1994 J. Mol. Struc. Theochem, 311, 273; Jaeger et al, 1989, Proc. Natl. Acad. Sci. USA, 86, 7706) to assess whether the ribozyme sequences fold into the appropriate secondary structure. Those ribozymes with unfavorable
!5 intramolecular interactions between the binding arms and the catalytic core were eliminated from consideration. As noted herein, varying binding arm lengths can be chosen to optimize activity. Generally, at least 5 bases on each arm are able to bind to, or otherwise interact with, the target RNA.
Chemical Synthesis and Purification of Ribozymes and Antisense for Efficient Cleavage and/or blocking of HBV RNA
Ribozymes and antisense constructs were designed to anneal to various sites in the RNA message. The binding arms ofthe ribozymes are complementary to the target site sequences described above, while the antisense constructs are fully complementary to the target site sequences described above. The ribozymes and antisense constructs were chemically synthesized. The method of synthesis used followed the procedure for normal RNA synthesis as described above and in Usman et al., (1987 J. Am. Chem. Soc, 109, 7845), Scaringe et al., (1990 Nucleic Acids Res., 18, 5433) and Wincott et al., supra, and made use of common nucleic acid protecting and coupling groups, such as dimethoxytrityl at the 5'-end, and phosphoramidites at the 3'-end. The average stepwise coupling yields were typically >98%.
Ribozymes and antisense constructs were also synthesized from DNA templates using bacteriophage T7 RNA polymerase (Milligan and Uhlenbeck, 1989, Methods Enzymol. 180, 51). Ribozymes and antisense constructs were purified by gel electrophoresis using general methods or were purified by high pressure liquid chromatography (HPLC; see Wincott et al., supra; the totality of which is hereby incoφorated herein by reference) and were resuspended in water. The sequences ofthe chemically synthesized ribozymes used in this study are shown below in Table 43.
!0
Ribozyme Cleavage of HBV RNA Target in vitro
Ribozymes targeted to the human HBV RNA are designed and synthesized as described above. These ribozymes can be tested for cleavage activity in vitro, for example using the following procedure. The target sequences and the nucleotide location within the !5 HBV RNA are given in Tables 36-43.
Cleavage Reactions: Full-length or partially full-length, internally-labeled target RNA for ribozyme cleavage assay is prepared by in vitro franscription in the presence of [α-32p] CTP, passed over a G 50 Sephadex® column by spin chromatography and used as substrate RNA without further purification. Alternately, substrates are 5'-32p-end labeled 0 using T4 polynucleotide kinase enzyme. Assays are performed by pre-warming a 2X concentration of purified ribozyme in ribozyme cleavage buffer (50 mM Tris-HCI, pH 7.5
at 37°C, 10 mM MgCl2) and the cleavage reaction was initiated by adding the 2X ribozyme mix to an equal volume of substrate RNA (maximum of 1-5 nM) that was also pre-warmed in cleavage buffer. As an initial screen, assays are carried out for 1 hour at o
37 C using a final concentration of either 40 nM or 1 mM ribozyme, i.e., ribozyme excess. The reaction is quenched by the addition of an equal volume of 95% formamide, 20 mM EDTA, 0.05% bromophenol blue and 0.05% xylene cyanol after which the sample is o heated to 95 C for 2 minutes, quick chilled and loaded onto a denaturing polyacrylamide gel. Substrate RNA and the specific RNA cleavage products generated by ribozyme cleavage are visualized on an autoradiograph ofthe gel. The percentage of cleavage is determined by Phosphor Imager® quantitation of bands representing the intact substrate and the cleavage products.
Transfection of HepG2 Cells with psHBV-1 and Ribozymes
The human hepatocellular carcinoma cell line Hep G2 was grown in Dulbecco's modified Eagle media supplemented with 10% fetal calf serum, 2 mM glutamine, 0.1 mM nonessential amino acids, 1 mM sodium pyruvate, 25 mM Hepes, 100 units penicillin, and 100 μg/ml streptomycin. To generate a replication competent cDNA, prior to fransfection the HBV genomic sequences are excised from the bacterial plasmid sequence contained in the psHBV-1 vector (Those skilled in the art understand that other methods may be used to generate a replication competent cDNA). This was done with an EcoRI and Hind IU restriction digest. Following completion ofthe digest, a ligation was performed under dilute conditions (20 μg/ml) to favor intermolecular ligation. The total ligation mixture was then concentrated using Qiagen spin columns.
Secreted alkaline phosphatase (SEAP) was used to normalize the HBsAg levels to confrol for transfection variability. The pSEAP2-TK control vector was constructed by ligating a Bgl II-Hind HI fragment ofthe pRL-TK vector (Promega), containing the heφes simplex virus thymidine kinase promoter region, into Bgl HIHind HI digested pSEAP2- Basic (Clontech). Hep G2 cells were plated (3 x IO4 cells/well) in 96-well microtiter plates and incubated overnight. A lipid/DNA/ribozyme complex was formed containing (at final concentrations) cationic lipid (15 μg/ml), prepared psHBV-1 (4.5 μg/ml), pSEAP2-TK (0.5 μg/ml), and ribozyme (100 μM). Following a 15 min. incubation at 37° C, the
complexes were added to the plated Hep G2 cells. Media was removed from the cells 96 hr. post-fransfection for HBsAg and SEAP analysis.
Transfection ofthe human hepatocellular carcinoma cell line, Hep G2, with replication competent HBV DNA results in the expression of HBV proteins and the production of virions. To investigate the potential use of ribozymes for the freatment of chronic HBV infection, a series of ribozymes that target the 3' terminus ofthe HBV genome have been synthesized. Ribozymes targeting this region have the potential to cleave all four major HBV RNA transcripts as well as the potential to block the production of HBV DNA by cleavage ofthe pregenomic RNA. To test the efficacy of these HBV ribozymes, they were co-transfected with HBV genomic DNA into Hep G2 cells, and the subsequent levels of secreted HBV surface antigen (HBsAg) were analyzed by ELISA. To confrol for variability in fransfection efficiency, a confrol vector which expresses secreted alkaline phosphatase (SEAP), was also co-transfected. The efficacy ofthe HBV ribozymes was determined by comparing the ratio of HBsAg: SEAP and/or HBeAg: SEAP to that of a scrambled attenuated confrol (SAC) ribozyme. Twenty-five ribozymes
(RPI18341, RPI18356, RPI18363, RPI18364, RPI18365, RPI18366, RPI18367, RPI18368, RPI18369, RPI18370, RPI18371, RPI18372, RPI18373, RPI18374, RPI18303, RPI18405, RPI18406, RPI18407, RPI18408, RPI18409, RPI18410, RPI18411, RPI18418, RPI18419, and RPI18422) have been identified which cause a reduction in the levels of HBsAg and/or HBeAg as compared to the corresponding SAC ribozyme.
Example 6: Analysis of HBsAg and SEAP Levels Following Ribozyme Treatment
Immulon 4 (Dynax) microtiter wells were coated overnight at 4° C with anti-HBsAg Mab (Biostride B88-95-31ad,ay) at 1 μg/ml in Carbonate Buffer (Na2CO3 15 mM, NaHCO3 35 mM, pH 9.5). The wells were then washed 4x with PBST (PBS, 0.05%
Tween® 20) and blocked for 1 hr at 37° C with PBST, 1% BSA. Following washing as above, the wells were dried at 37° C for 30 min. Biotinylated goat ant-HBsAg (Accurate YVS1807) was diluted 1:1000 in PBST and incubated in the wells for 1 hr. at 37° C. The wells were washed 4x with PBST. Streptavidin Alkaline Phosphatase Conjugate (Pierce 21324) was diluted to 250 ng/ml in PBST, and incubated in the wells for 1 hr. at 37° C. After washing as above, p-nitrophenyl phosphate substrate (Pierce 37620) was added to the wells, which were then incubated for 1 hr. at 37° C. The optical density at 405 nm was
then determined. SEAP levels were assayed using the Great EscAPe® Detection Kit (Clontech K2041-1), as per the manufacturers instructions.
Example 7: X-gene Reporter Assay The effect of ribozyme treatment on the level of fransactivation of a SV40 promoter driven firefly luciferase gene by the HBV X-protein was analyzed in transfected Hep G2 cells. As a confrol for variability in transfection efficiency, a Renilla luciferase reporter driven by the TK promoter, which is not transactivated by the X protein, was used. Hep G2 cells were plated (3 x IO4 cells/well) in 96-well microtiter plates and incubated overnight. A lipid/DNA ribozyme complex was formed containing (at final concentrations) cationic lipid (2.4 μg/ml), the X-gene vector pSBDR(2.5 μg/ml), the firefly reporter pSV40HCVluc (0.5 μg/ml), the Renilla luciferase control vector pRL-TK (0.5 μg/ml), and ribozyme (100 μM). Following a 15 min. incubation at 37° C, the complexes were added to the plated Hep G2 cells. Levels of firefly and Renilla luciferase were analyzed 48 hr. post transfection, using Promega' s Dual-Luciferase Assay System. The HBV X protein is a transactivator of a number of viral and cellular genes. Ribozymes which target the X region were tested for their ability to cause a reduction in X protein fransactivation of a firefly luciferase gene driven by the S V40 promoter in transfected Hep G2 cells. As a control for fransfection variability, a vector containing the Renilla luciferase gene driven by the TK promotor, which is not activated by the X protein, was included in the co-transfections. The efficacy ofthe HBV ribozymes was determined by comparing the ratio of firefly luciferase: Renilla luciferase to that of a scrambled attenuated confrol (SAC) ribozyme. Eleven ribozymes (RPI18365, RPI18367, RPI18368, RPI18371, RPI18372, RPI18373, RPI18405, RPI18406, RPI18411, RPI18418, RPI18423) were identified which cause a reduction in the level of fransactivation of a reporter gene by the X protein, as compared to the corresponding SAC ribozyme.
Example 8: HBV transgenic mouse study
A fransgenic mouse strain (founder strain 1.3.32 with a C57B1/6 background) that expresses HBV RNA and forms HBV viremia (Morrey et al, 1999, Antiviral Res., 42, 97- 108; Guidotti et al, 1995, J. Virology, 69, 10, 6158-6169) was utilized to study the in vivo
activity of ribozymes ofthe instant invention. This model is predictive in screening for anti-HBV agents. Ribozyme or the equivalent volume of saline was administered via a continuous s.c infusion using Alzet® mini-osmotic pumps for 14 days. Alzet® pumps were filled with test material(s) in a sterile fashion according to the manufacturer's instructions. Prior to in vivo implantation, pumps were incubated at 37°C overnight (> 18 hours) to prime the flow modulators. On the day of surgery, animals were lightly anesthetized with a ketamine/xylazine cocktail (94 mg/kg and 6 mg/kg, respectively; 0.3 ml, IP). Baseline blood samples (200 μl) were obtained from each animal via a retro- orbital bleed. A 2 cm area near the base ofthe tail was shaved and cleansed with betadine surgical scrub and sequentially with 70%> alcohol. A 1 cm incision in the skin was made with a #15 scalpel blade or a blunt pair of scissors near the base ofthe tail. Forceps were used to open a pocket rosfrally (i.e., towards the head) by spreading apart the subcutaneous connective tissue. The pump was inserted with the delivery portal pointing away from the incision. Wounds were closed with sterile 9-mm stainless steel clips or with sterile 4-0 suture. Animals were then allowed to recover from anesthesia on a warm heating pad before being returned to their cage. Wounds were checked daily. Clips or sutures were replaced as needed. Incisions typically healed completely within 7 days post-op. Animals were then deeply anesthetized with the ketamine/xylazine cocktail (150 mg/kg and 10 mg/kg, respectively; 0.5 ml, IP) on day 14 post pump implantation. A midline thoracotomy/ laparatomy was performed to expose the abdominal cavity and the thoracic cavity. The left ventricle was cannulated at the base and animals exsanguinated using a 23G needle and 1 ml syringe. Serum was separated, frozen and analyzed for HBV DNA and antigen levels. Experimental groups were compared to the saline confrol group in respect to percent change from day 0 to day 14. HBV DNA was assayed by quantitative PCR
Results
Table 44 is a summary ofthe group designation and dosage levels used in the HBV fransgenic mouse study. Baseline blood samples were obtained via a refroorbital bleed and animals (N=10/group) received anti-HBV ribozymes (100 mg/kg/day) as a continuous SC infusion. After 14 days, animals treated with a ribozyme targeting site 273 (RPI.18341) of
the HBV RNA showed a significant reduction in serum HBV DNA concenfration, compared to the saline treated animals as measured by a quantitative PCR assay. More specifically, the saline treated animals had a 69% increase in serum HBV DNA concentrations over this 2-week period while treatment with the 273 ribozyme (RPI.18341) resulted in a 60% decrease in serum HBV DNA concentrations. Ribozymes directed against sites 1833 (RPI.18371), 1873 (RPI.18418), and 1874 (RPI.18372) decreased serum HBV DNA concentrations by 49%, 15% and 16%, respectively.
Example 7: Activity of NCH Ribozyme to inhibit HER2 gene expression HER2 (also known as neu, erbB2 and c-erbB2) is an oncogene that encodes a 185- kDa transmembrane tyrosine kinase receptor. HER2 is a member ofthe epidermal growth factor receptor (EGFR) family and shares partial homology with other family members. In normal adult tissues HER2 expression is low. However, HER2 is overexpressed in at least 25-30% of breast (McGuire & Greene, 1989) and ovarian cancers (Berchuck, et al, 1990). Furthermore, overexpression of HER2 in malignant breast tumors has been correlated with increased metastasis, chemoresistance and poor survival rates (Slamon et al, 1987 Science 235: 177-182). Because HER2 expression is high in aggressive human breast and ovarian cancers, but low in normal adult tissues, it is an attractive target for ribozyme-mediated therapy (Thompson et al, supra). The greatest HER2 specific effects have been observed in cancer cell lines that express high levels of HER2 protein (as measured by ELISA). Specifically, in one study that freated five human breast cancer cell lines with the HER2 antibody (anti-erbB2-sFv), the greatest inhibition of cell growth was seen in three cell lines (MDA-MB-361, SKBR-3 and BT-474) that express high levels of HER2 protein. No inhibition of cell growth was observed in two cell lines (MDA-MB-231 and MCF-7) that express low levels of HER2 protein (Wright et al, 1997). Another group successfully used SKBR-3 cells to show HER2 antisense oligonucleotide-mediated inhibition of HER2 protein expression and HER2 RNA knockdown (Vaughn et al, 1995). Other groups have also demonstrated a decrease in the levels of HER2 protein, HER2 mRNA and/or cell proliferation in cultured cells using anti-HER2 ribozymes or antisense molecules (Suzuki, T. et al, 1997; Weichen, et al, 1997; Czubayko, F. et al, 1997; Colomer, et al, 1994; Befram et al, 1994). Because cell lines that express higher levels of HER2 have been more sensitive to anti-
HER2 agents, we are pursuing several medium to high expressing cell lines, including SKBR-3 and T47D, for ribozyme screens in cell culture.
A variety of endpoints have been used in cell culture models to look at HER2- mediated effects after freatment with anti-HER2 agents. Phenotypic endpoints include
5 inhibition of cell proliferation, apoptosis assays and reduction of HER2 protein expression. Because overexpression of HER2 is directly associated with increased proliferation of breast and ovarian tumor cells, a proliferation endpoint for cell culture assays will be our primary screen. There are several methods by which this endpoint can be measured. Following treatment of cells with ribozymes, cells are allowed to grow (typically 5 days)
0 after which either the cell viability, the incoφoration of [3H] thymidine into cellular DNA and/or the cell density can be measured. The assay of cell density is very straightforward and can be done in a 96-well format using commercially available fluorescent nucleic acid stains (such as Syto 13 or CyQuant). The assay using CyQuant is in place at RPI and is currently being employed to screen -100 ribozymes targeting HER2 (details below).
5 As a secondary, confirmatory endpoint a ribozyme-mediated decrease in the level of
HER2 protein expression can be evaluated using a HER2-specific ELISA.
Validation of Cell Lines and Ribozyme Treatment Conditions
Two human breast cancer cell lines (T47D and SKBR-3) that are known to express
!0 medium to high levels of HER2 protein, respectively, were considered for ribozyme screening. In order to validate these cell lines for HER2-mediated sensitivity, both cell lines were freated with the HER2 specific antibody, Herceptin® (Genentech) and its effect on cell proliferation was determined. Herceptin was added to cells at concentrations ranging from 0-8 μM in medium containing either no serum (OptiMem), 0.1% or 0.5%>
!5 FBS and efficacy was determined via cell proliferation. Maximal inhibition of proliferation (-50%) in both cell lines was observed after addition of Herceptin at 0.5 nM in medium containing 0.1 %> or no FBS. The fact that both cell lines are sensitive to an anti-HER2 agent (Herceptin) supports their use in experiments testing anti-HER2 ribozymes.
0 Prior to ribozyme screening, the choice ofthe optimal lipid(s) and conditions for ribozyme delivery was determined empirically for each cell line. Applicant has established a panel of proprietary lipids that can be used to deliver ribozymes to cultured cells and are
very useful for cell proliferation assays that are typically 3-5 days in length. Initially, this panel of proprietary lipid delivery vehicles was screened in SKBR-3 and T47D cells using previously established confrol oligonucleotides. Specific lipids and conditions for optimal delivery were selected for each cell line based on these screens. These conditions were used to deliver HER2 specific ribozymes to cells for primary (inhibition of cell proliferation) and secondary (decrease in HER2 protein) efficacy endpoints.
Primary Screen: Inhibition of Cell Proliferation
Although optimal ribozyme delivery conditions were determined for two cell lines, the SKBR-3 cell line were be used for the initial screen because it has the higher level of HER2 protein, and thus should be most susceptible to a HER2-specific ribozyme. Follow- up studies can be carried out in T47D cells to confirm leads as necessary.
Ribozyme screens were be performed using an automated, high throughput 96-well cell proliferation assay. Cell proliferation were measured over a 5-day freatment period using the nucleic acid stain CyQuant for determining cell density. The growth of cells treated with ribozyme/lipid complexes were compared to both untreated cells and to cells treated with Scrambled-arm attenuated core Confrols (SAC; or A; Figure 8). SACs can no longer bind to the target site due to the scrambled arm sequence and have nucleotide changes in the core that greatly diminish ribozyme cleavage. These SACs are used to determine non-specific inhibition of cell growth caused by ribozyme chemistry (i. e. multiple 2' O-Me modified nucleotides, a single 2'C-allyl uridine, 4 phosphorothioates and a 3' inverted abasic). Lead ribozymes are chosen from the primary screen based on their ability to inhibit cell proliferation in a specific manner. Dose response assays are carried out on these leads and a subset was advanced into a secondary screen using the level of HER2 protein as an endpoint.
Secondary Screen: Decrease in HER2 Protein
A secondary screen that measures the effect of anti-HER2 ribozymes on HER2 protein levels is used to support preliminary findings. A robust HER2 ELISA for both T47D and SKBR-3 cells has been established and is available for use as an additional endpoint.
Ribozyme Mechanism Assays
A Taqman assay for measuring the ribozyme-mediated decrease in HER2 RNA has also been established. This assay is based on PCR technology and can measure in real time the production of HER2 mRNA relative to a standard cellular mRNA such as GAPDH. This RNA assay is used to establish proof that lead ribozymes are working through an RNA cleavage mechanism and result in a decrease in the level of HER2 mRNA, thus leading to a decrease in cell surface HER2 protein receptors and a subsequent decrease in tumor cell proliferation.
Animal Models
Evaluating the efficacy of anti-HER2 agents in animal models is an important prerequisite to human clinical trials. As in cell culture models, the most HER2 sensitive mouse tumor xenografts are those derived from human breast carcinoma cells that express high levels of HER2 protein. In a recent study, nude mice bearing BT-474 xenografts were sensitive to the anti-HER2 humanized monoclonal antibody Herceptin, resulting in an 80%) inhibition of tumor growth at a 1 mg kg dose (ip, 2 X week for 4-5 weeks). Tumor eradication was observed in 3 of 8 mice freated in this manner (Baselga et al, 1998). This same study compared the efficacy of Herceptin alone or in combination with the commonly used chemotherapeutics, paclitaxel or doxorubicin. Although, all three anti- HER2 agents caused modest inhibition of tumor growth, the greatest antitumor activity was produced by the combination of Herceptin and paclitaxel (93 % inhibition of tumor growth vs 35% with paclitaxel alone). The above studies provide proof that inhibition of HER2 expression by anti-HER2 agents causes inhibition of tumor growth in animals. Lead anti-HER2 ribozymes chosen from in vitro assays are further tested in mouse xenograft models. Ribozymes are first tested alone and then in combination with standard chemotherapies.
Animal Model Development
Three human breast tumor cell lines (T47D, SKBR-3 and BT-474) were characterized to establish their growth curves in mice. These three cell lines have been implanted into the mammary papillae of both nude and SCLD mice and primary tumor volumes are measured 3 times per week. Growth characteristics of these tumor lines using
a Matrigel implantation format will also be established. In addition, the use of two other breast cell lines that have been engineered to express high levels of HER2 are also being used. The tumor cell line(s) and implantation method that supports the most consistent and reliable tumor growth is used in animal studies testing the lead HER2 ribozyme(s). Ribozyme are administered by daily subcutaneous injection or by continuous subcutaneous infusion from Alzet mini osmotic pumps beginning 3 days after tumor implantation and continuing for the duration ofthe study. Group sizes of at least 10 animals are employed. Efficacy is determined by statistical comparison of tumor volume of ribozyme-treated animals to a control group of animals freated with saline alone. Because the growth of these tumors is generally slow (45-60 days), an initial endpoint will be the time in days it takes to establish an easily measurable primary tumor (i.e. 50-100 mm3) in the presence or absence of ribozyme treatment.
Clinical Summary Breast cancer is a common cancer in women and also occurs in men to a lesser degree. The incidence of breast cancer in the United States is -180,000 cases per year and -46,000 die each year ofthe disease. In addition, 21,000 new cases of ovarian cancer per year lead to -13,000 deaths (data from Hung et al, 1995 and the Surveillance, Epidemiology and End Results Program, NCI). Ovarian cancer is a potential secondary indication for anti-HER2 ribozyme therapy.
A full review of breast cancer is given in the NCI PDQ for Breast Cancer. A brief overview is given here. Breast cancer is evaluated or "staged" on the basis of tumor size, and whether it has spread to lymph nodes and/or other parts ofthe body. In Stage I breast cancer, the cancer is no larger than 2 centimeters and has not spread outside ofthe breast. In Stage LI, the patient's tumor is 2-5 centimeters but cancer may have spread to the axillary lymph nodes. By Stage LU, metastasis to the lymph nodes is typical, and tumors are 5 centimeters. Additional tissue involvement (skin, chest wall, ribs, muscles etc.) may also be noted. Once cancer has spread to additional organs ofthe body, it is classed as Stage LV. Almost all breast cancers (>90%) are detected at Stage I or LI, but 31% of these are already lymph node positive. The 5-year survival rate for node negative patients (with standard surgery/radiation/chemotherapy /hormone regimens) is 97%; however,
involvement ofthe lymph nodes reduces the 5-year survival to only 77%. Involvement of other organs ( Stage HI) drastically reduces the overall survival, to 22% at 5 years. Thus, chance of recovery from breast cancer is highly dependent on early detection. Because up to 10% of breast cancers are hereditary, those with a family history are considered to be at high risk for breast cancer and should be monitored very closely.
Breast cancer is highly treatable and often curable when detected in the early stages. (For a complete review of breast cancer treatments, see the NCI PDQ for Breast Cancer.) Common therapies include surgery, radiation therapy, chemotherapy and hormonal therapy. Depending upon many factors, including the tumor size, lymph node involvement and location ofthe lesion, surgical removal varies from lumpectomy (removal ofthe tumor and some surrounding tissue) to mastectomy (removal ofthe breast, lymph nodes and some or all ofthe underlying chest muscle). Even with successful surgical resection, as many as 21% ofthe patients may ultimately relapse (10-20 years). Thus, once local disease is controlled by surgery, adjuvant radiation treatments, chemotherapies and/or hormonal therapies are typically used to reduce the rate of recurrence and improve survival. The therapy regimen employed depends not only on the stage ofthe cancer at its time of removal, but other variables such the type of cancer (ductal or lobular), whether lymph nodes were involved and removed, age and general health ofthe patient and if other organs are involved. Common chemotherapies include various combinations cytotoxic drugs to kill the cancer cells. These drugs include paclitaxel (Taxol), docetaxel, cisplatin, methotrexate, cyclophosphamide, doxorubin, fluorouracil etc. Significant toxicities are associated with these cytotoxic therapies. Well-characterized toxicities include nausea and vomiting, myelosuppression, alopecia and mucosity. Serious cardiac problems are also associated with certain ofthe combinations, e.g. doxorubin and paclitaxel, but are less common.
Testing for esfrogen and progesterone receptors helps to determine whether certain anti-hormone therapies might be helpful in inhibiting tumor growth. If either or both receptors are present, therapies to interfere with the action ofthe hormone ligands, can be given in combination with chemotherapy and are generally continued for several years. These adjuvant therapies are called SERMs, selective esfrogen receptor modulators, and they can give beneficial estrogen-like effects on bone and lipid metabolism while antagonizing estrogen in reproductive tissues. Tamoxifen is one such compound. The
primary toxic effect associated with the use of tamoxifen is a 2 to 7-fold increase in the rate of endometrial cancer. Blood clots in the legs and lung and the possibility of stroke are additional side effects. However, tamoxifen has been determined to reduce breast cancer incidence by 49% in high-risk patients and an extensive, somewhat controversial, clinical study is underway to expand the prophylactic use of tamoxifen. Another SERM, raloxifene, was also shown to reduce the incidence of breast cancer in a large clinical trial where it was being used to freat osteoporosis. In additional studies, removal ofthe ovaries and/or drugs to keep the ovaries from working are being tested.
Bone marrow fransplantation is being studied in clinical trials for breast cancers that have become resistant to traditional chemotherapies or where >3 lymph nodes are involved. Marrow is removed from the patient prior to high-dose chemotherapy to protect it from being destroyed, and then replaced after the chemotherapy. Another type of "fransplant" involves the exogenous freatment of peripheral blood stem cells with drugs to kill cancer cells prior to replacing the freated cells in the bloodstream. One biological freatment, a humanized monoclonal anti-HER2 antibody, Herceptin
(Genentech) has been approved by the FDA as an additional treatment for HER2 positive tumors. Herceptin binds with high affinity to the exfracellular domain of HER2 and thus blocks its signaling action. Herceptin can be used alone or in combination with chemotherapeutics (i.e. paclitaxel, docetaxel, cisplatin, etc.) (Pegram, et al, 1998). In Phase LTJ studies, Herceptin significantly improved the response rate to chemotherapy as well as improving the time to progression (Ross & Fletcher, 1998). The most common side effects attributed to Herceptin are fever and chills, pain, asthenia, nausea, vomiting, increased cough, diarrhea, headache, dyspnea, infection, rhinitis, and insomnia. Herceptin in combination with chemotherapy (paclitaxel) can lead to cardiotoxicity (Sparano, 1999), leukopenia, anemia, diarrhea, abdominal pain and infection.
HER2 Protein Levels for Patient Screening and as a Potential Endpoint
Because elevated HER2 levels can be detected in at least 30% of breast cancers, breast cancer patients can be pre-screened for elevated HER2 prior to admission to initial clinical trials testing an anti-HER2 ribozyme. Initial HER2 levels can be determined (by ELISA) from tumor biopsies or resected tumor samples.
During clinical trials, it may be possible to monitor circulating HER2 protein by ELISA (Ross and Fletcher, 1998). Evaluation of serial blood/serum samples over the course ofthe anti-HER2 ribozyme freatment period could be useful in determining early indications of efficacy. In fact, the clinical course of Stage TV breast cancer was correlated
5 with shed HER2 protein fragment following a dose-intensified paclitaxel monotherapy. In all responders, the HER2 serum level decreased below the detection limit (Luftner et al). Two cancer-associated antigens, CA27.29 and CA15.3, can also be measured in the serum. Both of these glycoproteins have been used as diagnostic markers for breast cancer. CA27.29 levels are higher than CA15.3 in breast cancer patients; the reverse is
10 true in healthy individuals. Of these two markers, CA27.29 was found to better discriminate primary cancer from healthy subjects. In addition, a statistically significant and direct relationship was shown between CA27.29 and large vs small tumors and node postive vs node negative disease (Gion, et al, 1999). Moreover, both cancer antigens were found to be suitable for the detection of possible metastases during follow-up (Rodriguez
5 de Patema et al, 1999). Thus, blocking breast tumor growth may be reflected in lower CA27.29 and/or CA15.3 levels compared to a control group. FDA submissions for the use of CA27.29 and CA15.3 for monitoring metastatic breast cancer patients have been filed (reviewed in Beveridge, 1999). Fully automated methods for measurement of either of these markers are commercially available.
!0
References
Baselga, J., Norton, L. Albanell, J., Kim, Y.M. and Mendelsohn, J. (1998) Recombinant humanized anti-HER2 antibody (Herceptin) enhances the antitumor activity of paclitaxel and doxorubicin against HER2/neu overexpressing human breast cancer 15 xenografts. Cancer Res. 15 : 2825-2831.
Berchuck, A. Kamel, A., Whitaker, R. et al. (1990) Overexpression of her-2/neu is associated with poor survival in advanced epithelial ovarian cancer. Cancer Research 50: 4087-4091.
Bertram, J. Killian, M., Brysch, W., Schlingensiepen, K.-H., and Kneba, M. (1994) »0 Reduction of erbB2 gene product in mamma carcinoma cell lines by erbB2 mRNA- specific and tyrosine kinase consensus phosphorothioate antisense oligonucleotides. Biochem. BioPhys. Res. Comm. 200: 661-667.
Beveridge, R.A. (1999) Review of clinical studies of CA27.29 in breast cancer management. Int. J. Biol. Markers 14: 36-39.
Colomer, R., Lupu, R., Bacus, S.S. and Gelmann, E.P. (1994) erbB-2 antisense ohgonucloetides inhibit the proliferation of breast carcinoma cells with erbB-2 oncogene amplification. British J. Cancer 70 : 819-825.
Czubayko, F., Downing, S.G., Hsieh, S.S., Goldstein, D.J., Lu P.Y., Trapnell, B.C. and Wellstein, A. (1997) Adenovirus-mediated fransduction of ribozymes abrogates HER- 2/neu and pleiofrophin expression and inhibits tumor cell proliferation. Gene Ther. A: 943-949. Gion, M., Mione, R., Leon, A.E. and Dittadi, R. (1999) Comparison ofthe diagnostic accuracy of CA27.29 and CA15.3 in primary breast cancer. Clin. Chem. 45: 630-637.
Hung, M.-C, Matin, A., Zhang, Y., Xing, X., Sorgi, F., Huang, L. and Yu, D. (1995) HER-2/neu-targeting gene therapy - a review. Gene 159: 65-71. Luftner, D., Schnabel. S. and Possinger, K. (1999) c-erbB-2 in serum of patients receiving fractionated paclitaxel chemotherapy. Int. J. Biol. Markers 14: 55-59.
McGuire, H.C. and Greene, M.I. (1989) The neu (c-erbB-2) oncogene. Semin. Oncol. 16: 148-155.
NCI PDQ/Treatment/Health Professionals/Breast Cancer: http://cancemet.nci.nih.gov/clinpdq/soa/Breast_cancer_Physician.hfrnl
NCI PDQ/Treatment/Patients/Breast Cancer: http ://cancernet.nci.nih. gov/clinpdq/pif/Breast cancer Patient.html
Pegram, M.D., Lipton, A., Hayes, D.F., Weber, B.L., Baselga, J.M., Tripathy, D., Baly, D., Baughman, S.A., Twaddell, T., Glaspy, J.A. and Slamon, D.J. (1998) Phase JJ study of receptor-enhanced chemosensitivity using recombinant humanized anti- pi 85HER2/neu monoclonal antibody plus cisplatin in patients with HER2/neu- overexpressing metastatic breast cancer refractory to chemotherapy treatment. J. Clin. Oncol. 16: 2659-2671.
Rodriguez de Paterna, L., Arnaiz, F., Estenoz, J. Ortuno, B. and Lanzos E. (1999) Study of serum tumor markers CEA, CA15.3, CA27.29 as diagnostic parameters in patients with breast carcinoma. Int. J. Biol. Markers 10: 24-29.
Ross, J.S. and Fletcher, J.A. (1998) The HER-2/neu oncogene in breast cancer: Prognostic factor, predictive factor and target for therapy. Oncologist 3: 1998.
Slamon, D.J., Clark, G.M., Wong, S.G., Levin, W.J., Ullrich, A. and McGuire, W.L. (1987) Human breast cancer: correlation of relapse and survival with amplification ofthe HER-2/neu oncogene. Science 235: 177-182.
Sparano, J.A. (1999) Doxorubicin/taxane combinations: Cardiac toxicity and pharmacokinetics. Semin. Oncol. 26: 14-19.
Surveillance, Epidemiology and End Results Program (SEER) Cancer Statistics Review: http://www.seer.ims.nci.nih.gov/Publications/CSRl 973_1996/ Suzuki T., Curcio, L.D., Tsai, J. and Kashani-Sabet M. (1997) Anti-c-erb-B-2
Ribozyme for Breast Cancer. In Methods in Molecular Medicine, Vol. 11, Therapeutic Applications of Ribozmes, Human Press, Inc., Totowa, NJ. aughn, J.P., Iglehart, J.D., Demirdji, S., Davis, P., Babiss, L.E., Caruthers, M.H., Marks, J.R. (1995) Antisense DNA downregulation ofthe ERBB2 oncogene measured by a flow cytometric assay. Proc Natl Acad Sci USA 92: 8338-8342.
Weichen, K., Zimmer, C. and Dietel, M. (1997) Selection of a high activity c-erbB-2 ribozyme using a fusion gene of c-erbB-2 and the enhanced green fluorescent protein. Cancer Gene Therapy 5: 45-51.
Wright, M., Grim, J., Deshane, J., Kim, M., Strong, T.V., Siegel, G.P., Curiel, D.T. (1997) An intracellular anti-erbB-2 single-chain antibody is specifically cytotoxic to human breast carcinoma cells overexpressing erbB-2. Gene Therapy 4: 317-322.
Applicant has designed, synthesized and tested several NCH ribozymes and HH ribozymes targeted against HER2 RNA (see for example Tables 31 and 34) in cell proliferation assays.
Proliferation assay:
The model proliferation assay used in the study can require a cell plating density of 2000 cells/well in 96-well plates and at least 2 cell doublings over a 5-day freatment period. To calculate cell density for proliferation assays, the FFPS (fluoro-imaging processing system) method well in the art was used. This method allows for cell density measurements after nucleic acids are stained with CyQuant dye, and has the advantage of
accurately measuring cell densities over a very wide range 1,000-100,000 cells/well in 96- well format.
Ribozymes (50-200 nM) were delivered in the presence of cationic lipid at 2.0 μg/mL and inhibition of proliferation was determined on day 5 post-treatment. Two full ribozyme screens were completed and 4 lead HH and 11 lead NCH ribozymes were chosen for further testing. Ofthe 15 lead Rzs chosen from primary screens, 4 NCH and 1 HH Rzs continued to inhibit cell proliferation in subsequent experiments. NCH Rzs against sites, 2001 (RPI No. 17236), 2783 (RPI No. 17249), 2939 (RPI No. 17251) or 3998 (RPI No. 17262) caused inhibition of proliferation ranging from 25-60% as compared to a scrambled confrol Rz (LA; RPI No. 17263). Ofthe five lead Rzs, the most efficacious is the NCH Rz (RPI No. 17251) against site 2939 of HER2 RNA. An example of results from cell culture assay is shown in Figure 8. Referring to Figure 8, NCH ribozymes and a HH ribozyme targeted against HER2 RNA, are shown to cause significant inhibition of proliferation of cells. This shows that ribozymes, for instance the NCH ribozymes are capable of inhibiting HER2 gene expression in mammalian cells.
Example 8: Activity of Class LI (Zinzyme) nucleic acid catalysts to inhibit HER2 gene expression
Applicant has designed, synthesized and tested several class π (zinzyme) ribozymes targeted against HER2 RNA (see, for example, Tables 58, 59, and 62) in cell proliferation RNA reduction assays.
Proliferation assay:
The model proliferation assay used in the study requires a cell-plating density of 2000-10000 cells/well in 96-well plates and at least 2 cell doublings over a 5-day treatment period. Cells used in proliferation studies were either human breast or ovarian cancer cells (SKBR-3 and SKOV-3 cells respectively). To calculate cell density for proliferation assays, the FLPS (fluoro-imaging processing system) method well known in the art was used. This method allows for cell density measurements after nucleic acids are stained with CyQuant® dye, and has the advantage of accurately measuring cell densities over a very wide range 1,000-100,000 cells/well in 96-well format.
Ribozymes (50-200 nM) were delivered in the presence of cationic lipid at 2.0-5.0 μg/mL and inhibition of proliferation was determined on day 5 post-treatment. Two full ribozyme screens were completed resulting in the selection of 14 ribozymes. Class IL (zinzyme) ribozymes against sites, 314 (RPI No. 18653), 443 (RPI No. 18680), 597 (RPI No. 18697), 659 (RPI No. 18682), 878 (RPI Nos. 18683 and 18654), 881 (RPI Nos. 18684 and 18685) 934 (RPI No. 18651), 972 (RPI No. 18656, 19292, 19727, 19728, and 19293), 1292 (RPI No. 18726), 1541 (RPI No. 18687), 2116 (RPI No. 18729), 2932 (RPI No. 18678), 2540 (RPI No. 18715), and 3504 (RPI No. 18710) caused inhibition of proliferation ranging from 25-80% as compared to a scrambled control ribozyme. An example of results from a cell culture assay is shown in Figure 20. Referring to Figure 20, Class II ribozymes targeted against HER2 RNA are shown to cause significant inhibition of proliferation of cells. This shows that ribozymes, for instance the Class II (zinzyme) ribozymes are capable of inhibiting HER2 gene expression in mammalian cells.
RNA assay:
RNA was harvested 24 hours post-treatment using the Qiagen RNeasy® 96 procedure. Real time RT-PCR (TaqMan® assay) was performed on purified RNA samples using separate primer/probe sets specific for either target HER2 RNA or control actin RNA (to normalize for differences due to cell plating or sample recovery). Results are shown as the average of triplicate determinations of HER2 to actin RNA levels post- treatment. Figure 30 shows class LI ribozyme (zinzyme) mediated reduction in HER2 RNA targeting site 972 vs a scrambled attenuated confrol.
Dose response assays: Active ribozyme was mixed with binding arm-attenuated confrol (BAC) ribozyme to a final oligonucleotide concenfration of either 100, 200 or 400 nM and delivered to cells in the presence of cationic lipid at 5.0 μg/mL. Mixing active and BAC in this manner maintains the lipid to ribozyme charge ratio throughout the dose response curve. HER2 RNA reduction was measured 24 hours post-treatment and inhibition of proliferation was determined on day 5 post-treatment. The dose response antiproliferation results are summarized in Figure 31 and the dose-dependent reduction of HER2 RNA results are
summarized in Figure 32. Figure 33 shows a combined dose response plot of both anti- proliferation and RNA reduction data for a class LI ribozyme targeting site 972 of HER2 RNA (RPI 19293).
Example 9: Compositions having RNA cleaving activity
Hammerhead ribozymes are an example of catalytic RNA molecules which are able to recognize and cleave a given specific RNA subsfrate (Hutchins et α/.,1986, Nucleic Acids Res. 14:3627; Keese and Symons, in Viroids and viroid-like pathogens (J.J. Semanchik, publ., CRC-Press, Boca Raton, Florida, 1987, pages 1-47). The catalytic center of hammerhead ribozymes is flanked by three stems and can be formed by adjacent sequence regions ofthe RNA or also by regions, which are separated from one another by many nucleotides. Figure 6 shows a diagram of such a catalytically active hammerhead structure. The stems have been denoted I, IL and ILI. The nucleotides are numbered according to the standard nomenclature for hammerhead ribozymes (Hertel et al, 1992, Nucleic Acids Res. 20:3252). In this nomenclature, bases are denoted by a number, which relates their position relative to the 5' side ofthe cleavage site. Furthermore, each base that is involved in a stem or loop region has an additional designation (which is denoted by a decimal point and then another number) that defines the position of that base within the stem or loop. A designation of A15 Λ would indicate that this base is involved in a paired region and that it is the first nucleotide in that stem going away from the core region. This accepted convention for describing hammerhead-derived ribozymes allows for the nucleotides involved in the core ofthe enzyme to always have the same number relative to all ofthe other nucleotides. The size ofthe stems involved in substrate binding or core formation can be any size and of any sequence, and the position of A9, for example, will remain the same relative to all ofthe other core nucleotides. Nucleotides designated, for example, NΛ12 or N9Λ represent an inserted nucleotide where the position ofthe caret (Λ) relative to the number denotes whether the insertion is before or after the indicated nucleotide. Thus, NΛ represents a nucleotide inserted before nucleotide position 12, and N9Λ represents a nucleotide inserted after nucleotide position 9. The consensus sequence ofthe catalytic core structure is described by Ruffher and
Uhlenbeck, 1990, Nucleic Acids Res. 18:6025-6029. Perriman et al, 1992, Gene 113:157- 163, have meanwhile shown that this structure can also contain variations, for example,
naturally occurring nucleotide insertions such as N9Λ and NΛ12. Thus, the positive strand ofthe satellite RNA ofthe tobacco ring-spot virus does not contain any ofthe two nucleotide insertions while the +RNA strand ofthe virusoid ofthe lucerne transient streak virus (vLTSV) contains a N9Λ = U insertion which can be mutated to C or G without loss ofactivity (Sheldon and Symons, 1989, Nucleic Acids Res. 17:5679-5685). Furthermore, in this special case, N7 = A and R15 1 = A. On the other hand, the minus strand ofthe carnation stunt associated viroid (-CarSV) is quite unusual since it contains both nucleotide insertions, that is NΛ12 = A and N9Λ = C (Hernandez et α/.,1992, Nucleic Acids Res. 20:6323-6329). In this viroid N7 = A and R15 1 = A. In addition, this special hammerhead structure exhibits a very effective self-catalytic cleavage despite the more open central stem.
Possible uses of hammerhead ribozymes include, for example, generation of RNA restriction enzymes and the specific inactivation ofthe expression of genes in, for example, animal, human or plant cells and prokaryotes, yeasts and plasmodia. A particular biomedical interest is based on the fact that many diseases, including many forms of tumors, are related to the overexpression of specific genes. Inactivating such genes by cleaving the associated mRNA represents a possible way to confrol and eventually treat such diseases. Moreover there is a great need to develop antiviral, antibacterial, and antifungal pharmaceutical agents. Ribozymes have potential as such anti-infective agents since RNA molecules vital to the survival ofthe organism can be selectively destroyed. In addition to needing the correct hybridizing sequences for subsfrate binding, substrates for hammerhead ribozymes have been shown to strongly prefer the triplet N i6.2 Tji6.i H i7 (NUH) where N can e my nucieotide, U is uridine, and H is either adenosine, cytidine, or uridine (Koizumi et al, 1988, FEBS Lett. 228, 228-230; Ruffner et al, 1990, Biochemistry 29, 10695-10702 ; Perriman et al, 1992, Gene 113, 157-163). NUH is sometimes designated as NUX. The fact that changes to this general rule for substrate specificity result in non- functional substrates implies that there are "non core compatible" structures which are formed when substrates are provided which deviate from the stated requirements. Evidence along these lines was recently reported by Uhlenbeck and co-workers (Uhlenbeck et al, 1997, Biochemistry 36: 1108-1114) when they demonsfrated that the substitution of a G at position 17 caused a functionally catastrophic base pair between G17 and C3 to form, both preventing the correct orientation ofthe
scissile bond for cleavage and the needed tertiary interactions of C3 (Murray et al, 1995, Biochem. J. 311 :487-494). The strong preference for a U at position 16.1 may exist for similar reasons. Many experiments have been done in an attempt to isolate ribozymes which are able to efficiently relieve the requirement of a U at position 16.1, however, attempts to find hammerhead type ribozymes which can cleave substrates having a base other than a U at position 16.1 have proven impossible (Perriman et al, 1992, Gene 113, 157-163).
Efficient catalytic molecules with reduced or altered requirements in the cleavage region are highly desirable because their isolation would greatly increase the number of available target sequences that molecules of this type could cleave. For example, it would be desirable to have a ribozyme variant that could efficiently cleave substrates containing triplets other than N U H since this would increase the number of potential target cleavage sites.
Chemically modified oligonucleotides which contain a block of deoxyribonucleotides in the middle region ofthe molecule have potential as pharmaceutical agents for the specific inactivation ofthe expression of genes (Giles et al, 1992, Nucleic Acids Res. 20:763-770). These oligonucleotides can form a hybrid DNA- RNA duplex in which the DNA bound RNA strand is degraded by RNase H. Such oligonucleotides are considered to promote cleavage ofthe RNA and so cannot be characterized as having an RNA-cleaving activity nor as cleaving an RNA molecule (the RNase H is cleaving). A significant disadvantage of these oligonucleotides for in vivo applications is their low specificity, since hybrid formation, and thus cleavage, can also take place at undesired positions on the RNA molecules.
Since, unmodified ribozymes are sensitive to degradation by RNases, chemically modified active substances have to be used in order to administer hammerhead ribozymes exogenously (discussed, for example, by Heidenreich et al, 1994, J. Biol. Chem. 269:2131-2138; Kiehntopf et al, \99A, EMBOJ. 13:4645-4652; Paolella et al, 1992, EMBOJ. 11:1913-1919; and Usman et al, \99 A, Nucleic Acids Symp. Ser. 31 :163-164). Sproat et al, U.S. Pat. No. 5,334,711, describe such chemically modified active substances based on synthetic catalytic oligonucleotide structures with a length of 35 to 40 nucleotides which are suitable for cleaving a nucleic acid target sequence and contain modified nucleotides that contain an optionally substituted alkyl, alkenyl or alkynyl group
with 1 - 10 carbon atoms at the 2'-O atom ofthe ribose. These oligonucleotides contain modified nucleotide building blocks and form a structure resembling a hammerhead structure. These oligonucleotides are able to cleave specific RNA subsfrates.
Usman et al, U.S. Patent No. 5,891,684, describe enzymatic nucleic acid molecules with one or more nucleotide base modification(s) in a subsfrate binding arm.
Thompson et al, US Patent No. 5,599,704 describe enzymatic RNA molecules targeted against ErbB2/«ew/Her2 RNA.
Sullivan et al, US Patent No. 5,616,490 describe enzymatic RNA molecules targeted against protein kinase C (PKC) RNA. Sioud, International PCT publication No. WO 99/63066 describe hammerhead ribozymes targeted against specific sites within protein kinase C alpha (PKC alpha), VEGF, and TNF alpha RNA.
Jarvis et al, International PCT publication No. WO 98/505030, describe the synthesis of xylo-ribonucleosides and oligonucleotides comprising xylo modifications. This invention relates to novel enzymatic nucleic acid molecules having an RNA- cleavage activity, as well as their use for cleaving RNA substrates in vitro and in vivo. The compositions contain an active center, the subunits of which are selected from nucleotides and/or nucleotide analogues, as well as flanking regions contributing to the formation of a specific hybridization with an RNA subsfrate. Preferred compositions form, in combination with an RNA substrate, a structure resembling a hammerhead structure. The active center ofthe disclosed compositions is characterized by the presence of I15 ' which allows cleavage of RNA substrates having C16 -1. It is therefore an object of the present invention to provide compositions that cleave RNA, and in particular to provide RNA- cleaving oligomers which at the same time have a high stability, activity, and specificity. This invention relates to novel nucleic acid molecules with catalytic activity, which are particularly useful for cleavage of RNA or DNA or combination thereof. The nucleic acid catalysts ofthe instant invention are distinct from other nucleic acid catalysts known in the art. Specifically, nucleic acid catalysts ofthe instant invention are capable of catalyzing an intermolecular or intramolecular endonuclease reaction. It is another object ofthe present invention to provide compositions that cleave RNA subsfrates having a cleavage site triplet other than N16,2U16 1H17 (NUH; Figure 6), where N is a nucleotide, U is uridine and H is adenosine, uridine or cytidine. H is used
interchangably with X. Specifically, the enzymatic nucleic acid molecule ofthe instant invention has an endonuclease activity to cleave RNA substrates having a cleavage triplet N i6.2 .i6.ι jji7 (Ncjj. FigUre 6), where N is a nucleotide, C is cytidine and H is adenosine, uridine or cytidine. H is used interchangeably with X. In another aspect the invention features an enzymatic nucleic acid molecule ofthe instant invention has an endonuclease activity to cleave RNA substrates having a cleavage triplet N162C16 1N17 (NCN; Figure 6), where N is a nucleotide, C is cytidine.
In a preferred embodiment, the invention features an enzymatic nucleic acid molecule having formula 1 :
^ — G— A — A — I D — 3.
L
^ (N) n~ (N)p— A — G— N — A — G — TJ — C ~ E~ 5' where N represents independently a nucleotide or a non-nucleotide linker, which may be same or different; D and E are independently oligonucleotides of length sufficient to stably interact (e.g., by forming hydrogen bonds with complementary nucleotides in the target) with a target nucleic acid molecule (the target can be an RNA, DNA or mixed polymers), preferably, the length of D and E are independently between 3-20 nucleotides long, specifically, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 17, and 20; o and n are integers independently greater than or equal to 1 and preferably less than about 100, specifically 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 50, wherein if (N)0 and (N)n are nucleotides, (N)o and (N)n are optionally able to interact by hydrogen bond interaction, in particular if n =1 and o=l then (N)n is preferably a purine (e.g., G, and A) and (N)o is preferably a pyrimidine (e.g., C and U) and (N)n preferably forms; • indicates base-paired interaction; L is a linker which may be present or absent (i.e., the molecule may be assembled from two separate oligonucleotides), but when present, is a nucleotide and/or a non-nucleotide linker, which may be a single-stranded and/or double-stranded region; p is an integer 0 or 1, when p=l, (N)p is preferably A or U; and represents a chemical linkage (e.g. a phosphate ester linkage, amide linkage, phosphorothioate linkage or others known in the art). A, U, I, C and G represent adenosine, uridine, inosine, cytidine and guanosine nucleotides, respectively. The N in 5'-CUGANGA-3' region of formula 1 is preferably U.
The nucleotides in the formula 1 are unmodified or modified at the sugar, base, and/or phosphate as known in the art.
In a preferred embodiment, the invention features an enzymatic nucleic acid molecule having formula 2:
G-A- A- I D- 3'
L
(N) n G- (N)P-A- G- N- A- G~ U- C~ E" 5'
where N represents independently a nucleotide or a non-nucleotide linker, which may be same or different; D and E are independently oligonucleotides of length sufficient to stably interact (e.g., by forming hydrogen bonds with complementary nucleotides in the target) with a target nucleic acid molecule (the target can be an RNA, DNA or mixed polymers), preferably, the length of D and E are independently between 3-20 nucleotides long, specifically, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 17, and 20; o and n are integers independently greater than or equal to 0 and preferably less than about 100, specifically 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 50, wherein if (N)0 and (N)n are nucleotides, (N)o and (N)n are optionally able to interact by hydrogen bond interaction; • indicates base- paired interaction; L is a linker which may be present or absent (i.e., the molecule may be assembled from two separate oligonucleotides), but when present, is a nucleotide and/or a non-nucleotide linker, which may be a single-stranded and/or double-stranded region; p is an integer 0 or 1, when p=l, (N)p is preferably A, C or U; and represents a chemical linkage (e.g. a phosphate ester linkage, amide linkage, phosphorothioate linkage or others known in the art). A, U, I, C and G represent adenosine, uridine, inosine, cytidine and guanosine nucleotides, respectively. The N in 5'-CUGANGA-3' region of formula 2 is preferably U. The nucleotides in the formula 2 are unmodified or modified at the sugar, base, and/or phosphate as known in the art. In a preferred embodiment, the I (inosine) in formula 1 and 2 is preferably a riboinosine or a xylo-inosine.
In yet another embodiment, the nucleotide linker (L) is a nucleic acid aptamer, such as an ATP aptamer, HIV Rev aptamer (RRE), HIV Tat aptamer (TAR) and others (for a review see Gold et al, 1995, Annu. Rev. Biochem., 64, 763; and Szostak & Ellington, 1993, in The RNA World, ed. Gesteland and Atkins, pp 511, CSH Laboratory Press). A "nucleic acid aptamer" as used herein is meant to indicate nucleic acid sequence capable of interacting with a ligand. The ligand can be any natural or a synthetic molecule, including but not limited to a resin, metabolites, nucleosides, nucleotides, drugs, toxins, transition state analogs, peptides, lipids, proteins, amino acids, nucleic acid molecules, hormones, carbohydrates, receptors, cells, viruses, bacteria and others. In a preferred embodiment L has the sequence 5 '-GAAA-3 ' or 5 '-GUUA-3 ' .
In yet another embodiment, the non-nucleotide linker (L) is as defined herein. The term "non-nucleotide", as used herein, includes either abasic nucleotide, polyether, polyamine, polyamide, peptide, carbohydrate, lipid, or polyhydrocarbon compounds. Specific examples include those described by Seela and Kaiser, Nucleic Acids Res. 1990, 75:6353 and Nucleic Acids Res. 1987, 75:3113; Cload and Schepartz, J. Am. Chem. Soc. 1991, 773:6324; Richardson and Schepartz, J. Am. Chem. Soc. 1991, 775:5109; Ma et al., Nucleic Acids Res. 1993, 27:2585 and Biochemistry 1993, 52:1751; Durand et al., Nucleic Acids Res. 1990, 75:6353; McCurdy et al., Nucleosides & Nucleotides 1991, 70:287; Jschke et al., Tetrahedron Lett. 1993, 34:301; Ono et al., Biochemistry 1991, 50:9914; Arnold et al, International Publication No. WO 89/02439; Usman et al, International Publication No. WO 95/06731 ; Dudycz et al, International Publication No. WO 95/11910 and Ferentz and Verdine, J. Am. Chem. Soc. 1991, 775:4000, all hereby incoφorated by reference herein. Non-nucleotide linkers can be any molecule, which is not an oligomeric sequence, that can be covalently coupled to an oligomeric sequence. Preferred non-nucleotide linkers are oligomeric molecules formed of non-nucleotide subunits. Examples of such non-nucleotide linkers are described by Letsinger and Wu, (J. Am. Chem. Soc. 117:7323-7328 (1995)), Benseler et al, (J. Am. Chem. Soc. 115:8483-8484 (1993)) and Fu et al. , (J. Am. Chem. Soc. 116:4591-4598 (1994)). Prefeπed non-nucleotide linkers, or subunits for non-nucleotide linkers, include substituted or unsubstituted Ci-Cio straight chain or branched alkyl, substituted or unsubstituted C2-C10 straight chain or branched alkenyl, substituted or unsubstituted C2- C10 straight chain or branched alkynyl, substituted or unsubstituted C do straight chain or
branched alkoxy, substituted or unsubstituted C2-C10 straight chain or branched alkenyloxy, and substituted or unsubstituted C2-Cι0 straight chain or branched alkynyloxy. The substituents for these prefeπed non-nucleotide linkers (or subunits) can be halogen, cyano, amino, carboxy, ester, ether, carboxamide, hydroxy, or mercapto. Thus, in a prefeπed embodiment, the invention features an enzymatic nucleic acid molecule having one or more non-nucleotide moieties, and having enzymatic activity to cleave an RNA or DNA molecule. By the term "non-nucleotide" is meant any group or compound which can be incoφorated into a nucleic acid chain in the place of one or more nucleotide units, including either sugar and/or phosphate substitutions, and allows the remaining bases to exhibit their enzymatic activity. The group or compound is abasic in that it does not contain a commonly recognized nucleotide base, such as adenosine, guanine, cytosine, uracil or thymine. The terms "abasic" or "abasic nucleotide" as used herein encompass sugar moieties lacking a base or having other chemical groups in place of nucleotide base at the 1' position. In a prefeπed embodiment, the invention features modified ribozymes with phosphate backbone modifications comprising one or more phosphorothioate, phosphorodithioate, methylphosphonate, moφholino, amidate carbamate, carboxymethyl, acetamidate, polyamide, sulfonate, sulfonamide, sulfamate, formacetal, thioformacetal, and/or alkylsilyl, substitutions. For a review of oligonucleotide backbone modifications see Hunziker and Leumann, 1995, Nucleic Acid Analogues: Synthesis and Properties, in Modern Synthetic Methods, VCH, 331-417, and Mesmaeker et al, 1994, Novel Backbone Replacements for Oligonucleotides, in Carbohydrate Modifications in Antisense Research, ACS, 24-39.
In a further prefeπed embodiment ofthe instant invention, an inverted deoxy abasic moiety is utilized at the 3' end ofthe enzymatic nucleic acid molecule.
By "pyrimidines" is meant nucleotides comprising modified or unmodified derivatives of a six membered pyrimidine ring. An example of a pyrimidine is modified or unmodified uridine.
In a prefeπed embodiment, the nucleosides ofthe instant invention include, 2'-O- methyl-2,6-diaminopurine riboside; 2'-deoxy-2'amino-2,6-diaminopurine riboside; 2'-(N- alanyl) amino-2'-deoxy-uridine; 2'-(N-phenylalanyl)amino-2'-deoxy-uridine; 2'-deoxy -2'- (N-beta-alanyl) amino ; 2'-deoxy-2'-(lysiyl) amino uridine; 2'-C-allyl uridine; 2'-O-amino-
uridine; 2'-O-methylthiomethyl adenosine; 2'-O-methylthiomethyl cytidine ; 2'-O- methylthiomethyl guanosine; 2'-O-methylthiomethyl-uridine; 2'-Deoxy-2'-(N-histidyl) amino uridine; 2'-deoxy-2'-amino-5-methyl cytidine; 2'-(N-β-carboxamidine-beta- alanyl)amino-2 '-deoxy-uridine; 2 ' -deoxy-2 ' -(Ν-beta-alanyl)-guanosine; 2 '-O-amino- adenosine; 2'-(N-lysyl)amino -2'-deoxy-cytidine; 2'-Deoxy -2'-(L-histidine) amino Cytidine; and 5-Imidazoleacetic acid 2'-deoxy-5'-triphosphate uridine.
By "oligonucleotide" as used herein is meant a molecule having two or more nucleotides. The polynucleotide can be single, double or multiple stranded and may have modified or unmodified nucleotides or non-nucleotides or various mixtures and combinations thereof.
In a prefeπed embodiment, the enzymatic nucleic acid molecule of formula 1 or 2 include at least three ribonucleotide residues, preferably 4, 5, 6, 7, 8, 9, and 10 ribonucleotide residues.
In prefeπed embodiments, the enzymatic nucleic acid ofthe instant invention includes one or more stretches of RΝA, which provide the enzymatic activity ofthe molecule, linked to the non-nucleotide moiety. The necessary RΝA components are known in the art (see for e.g., Usman et al, supra).
Thus, in one prefeπed embodiment, the invention features enzymatic nucleic acid molecules that inhibit gene expression and/or cell proliferation in vitro or in vivo (e.g. in patients). These chemically or enzymatically synthesized nucleic acid molecules contain substrate binding domains that bind to accessible regions of specific target nucleic acid molecules. The nucleic acid molecules also contain domains that catalyze the cleavage of target. Upon binding, the enzymatic nucleic acid molecules cleave the target molecules, preventing for example, translation and protein accumulation. In the absence ofthe expression ofthe target gene, cell proliferation, for example, is inhibited.
In another prefeπed embodiment, catalytic activity ofthe molecules described in the instant invention can be optimized as described by Draper et al., supra. The details will not be repeated here, but include altering the length ofthe ribozyme binding arms, or chemically synthesizing ribozymes with modifications (base, sugar and/or phosphate) that prevent their degradation by serum ribonucleases and/or enhance their enzymatic activity (see e.g., Eckstein et al, International Publication No. WO 92/07065; Peπault et al, 1990 Nature 344, 565; Pieken et al., 1991 Science 253, 314; Usman and Cedergren, 1992
Trends in Biochem. Sci. 17, 334; Usman et al, International Publication No. WO 93/15187; and Rossi et al, International Publication No. WO 91/03162; Sproat, US Patent No. 5,334,711; and Burgin et al, supra; all of these describe various chemical modifications that can be made to the base, phosphate and/or sugar moieties of enzymatic RNA molecules). Modifications which enhance their efficacy in cells, and removal of bases from stem loop structures to shorten RNA synthesis times and reduce chemical requirements are desired. (All these publications are hereby incoφorated by reference herein.).
By "nucleic acid catalyst" as used herein is meant a nucleic acid molecule (e.g., the molecule of formulae 1 and 2) capable of catalyzing (altering the velocity and/or rate of) a variety of reactions including the ability to repeatedly cleave other separate nucleic acid molecules (endonuclease activity) in a nucleotide base sequence-specific manner. Such a molecule with endonuclease activity may have complementarity in a subsfrate binding region to a specified gene target, and also has an enzymatic activity that specifically cleaves RNA or DNA in that target. That is, the nucleic acid molecule with endonuclease activity is able to inframolecularly or intermolecularly cleave RNA or DNA and thereby inactivate a target RNA or DNA molecule. This complementarity functions to allow sufficient hybridization ofthe enzymatic RNA molecule to the target RNA or DNA to allow the cleavage to occur. 100%> complementarity is prefeπed, but complementarity as low as 50-75%) may also be useful in this invention. The nucleic acids may be modified at the base, sugar, and/or phosphate groups. The term enzymatic nucleic acid as used herein is used interchangeably with phrases such as ribozymes, catalytic RNA, enzymatic RNA, catalytic oligonucleotides, nucleozyme, RNA enzyme, endoribonuclease, endonuclease, minizyme, oligozyme, finderon or nucleic acid catalyst. All of these terminologies describe nucleic acid molecules ofthe instant invention with enzymatic activity. The specific examples of enzymatic nucleic acid molecules described in the instant application are not limiting in the invention and those skilled in the art will recognize that all that is important in an enzymatic nucleic acid molecule of this invention is that it has a specific substrate binding site which is complementary to one or more ofthe target nucleic acid regions, and that it have nucleotide sequences within or suπounding that substrate binding site which impart a nucleic acid cleaving activity to the molecule (Cech et al., U.S. Patent No. 4,987,071; Cech et al, 1988, 260 JAMA 3030).
The enzymatic nucleic acid molecule of Formula 1 or 2 may independently comprise a cap structure which may independently be present or absent.
By "chimeric nucleic acid molecule" or "mixed polymer" is meant that, the molecule maybe comprised of both modified or unmodified nucleotides. In yet another prefeπed embodiment, the 3 '-cap is selected from a group comprising,
4', 5 '-methylene nucleotide; l-(beta-D-erythrofuranosyl) nucleotide; 4'-thio nucleotide, carbocyclic nucleotide; 5'-amino-alkyl phosphate; l,3-diamino-2-propyl phosphate, 3- aminopropyl phosphate; 6-aminohexyl phosphate; 1,2-aminododecyl phosphate; hydroxypropyl phosphate; 1,5-anhydrohexitol nucleotide; L-nucleotide; alpha-nucleotide; modified base nucleotide; phosphorodithioate; tλreσ-pentofuranosyl nucleotide; acyclic
3',4'-seco nucleotide; 3,4-dihydroxybutyl nucleotide; 3,5-dihydroxypentyl nucleotide, 5'-5'- inverted nucleotide moiety; 5'-5'-inverted abasic moiety; 5'-phosphoramidate; 5'- phosphorothioate; 1,4-butanediol phosphate; 5'-amino; bridging and/or non-bridging 5'- phosphoramidate, phosphorothioate and/or phosphorodithioate, bridging or non bridging methylphosphonate and 5'-mercapto moieties (for more details, see Beaucage and Iyer, 1993, Tetrahedron 49, 1925; incoφorated by reference herein). By the term "non- nucleotide" is meant any group or compound which can be incoφorated into a nucleic acid chain in the place of one or more nucleotide units, including either sugar and/or phosphate substitutions, and allows the remaining bases to exhibit their enzymatic activity. The group or compound is abasic in that it does not contain a commonly recognized nucleotide base, such as adenosine, guanine, cytosine, uracil or thymine. The terms "abasic" or "abasic nucleotide" as used herein encompass sugar moieties lacking a base or having other chemical groups in place of a base at the 1' position.
In a prefeπed embodiment, the invention features l-(beta-D-xylofuranosyl)- xypoxanthine phosphoramidite and a process for the synthesis thereof and incoφoration into oligonucleotides, such as enzymatic nucleic acid molecule.
In yet another prefeπed embodiment, the invention features enzymatic nucleic acid molecules targeted against HER2 RNA, specifically, ribozymes in the hammerhead and NCH motifs. In a prefeπed embodiment, the invention features enzymatic nucleic acid molecules targeted against PKC alpha RNA, specifically, ribozymes in the hammerhead and NCH motifs.
Targets, for example PKC alpha RNA, for useful ribozymes and antisense nucleic acids can be determined, for example, as described in Draper et al, WO 95/04818; McSwiggen et al, U.S. Patent Nos. 5,525,468 and 5,646,042, all are hereby incoφorated by reference herein in their totality. Other examples include the following PCT applications, which concern inactivation of expression of disease-related genes: WO 95/23225, WO 95/13380, WO 94/02595, all incoφorated by reference herein.
The specific enzymatic nucleic acid molecules described in the instant application are not limiting in the invention and those skilled in the art will recognize that all that is important in an enzymatic nucleic acid molecule of this invention is that it has a specific subsfrate binding site (e.g., D and E of Formula 1 above) which is complementary to one or more ofthe target nucleic acid regions, and that it have nucleotide sequences within or suπounding that substrate binding site which impart a nucleic acid cleaving activity to the molecule.
All naturally occurring hammerhead ribozymes have an A15 '-U16 Λ base pair. In addition, it is known that subsfrates for ribozymes based on the consensus hammerhead sequence strongly prefer a substrate that contains an N16'2U16 1H17 triplet in which H17 is not a guanosine (Koizumi et al, FEBS Lett. 228, 228-230 (1988); Ruffner et al, Biochemistry 29, 10695-10702 (1990); Perriman et al, Gene 113, 157-163 (1992)). Many experiments have been done in an attempt to isolate ribozymes which are able to efficiently relieve the requirement of a U at position 16.1, however, attempts to find ribozymes which can cleave subsfrates having a base other than a U at position 16.1 have proven largely unsuccessful (Perriman et al, Gene 113, 157-163 1992, Singh et al, Antisense and Nucleic Acid Drug Development 6:165-168 (1996)).
However, examination ofthe recently published X-ray crystal structures (Pley et al, Nαtwre 372:68-74 (1994), Scott et al, Cell 81 :991-1002 (1995), and Scott et al, Science 274:2065-2069 (1996)) led to the realization that the A15 '-U16 Λ interaction is a non- standard base pair with a single hydrogen bond between the exocyclic amine (Ν6) ofthe adenosine and the 4-oxo group ofthe uridine. Modeling studies (based on the crystal structure) then led to the discovery that the interaction ofthe wild-type A15 1-U16 1 base pair can be spatially mimicked by replacement with an ι15 1-C16,1 base pair that adopts an isostructural orientation and which preserves the required contact ofthe 2-keto group of C16 -1 with A6 ofthe uridine turn. In the model, the polarity ofthe stabilizing hydrogen
bond between positions 15.1 and 16.1 is reversed in the ι15 1_c16 1 interaction, but the coπect orientation ofthe bases around this bond is maintained.
It has been discovered that hammerhead ribozyme analogues containing an inosine at position 15.1 readily cleave RNA substrates containing an
triplet. Based on this, disclosed are compositions, preferably synthetic oligomers, which cleave a nucleic acid target sequence containing the triplet N
16'
2C
16 1H
17. It is prefeπed that H
17 is not guanosine, however, under certain circumstances, NCG triplet containing RNA can be cleaved by the ribozymes ofthe instant invention. The ability to cleave subsfrates having
1 Λ 7 1 Λ 1 17
N C X triplets effectively doubles the number of targets available for cleavage by compositions ofthe type disclosed.
Example 10: Synthesis of l-(beta-D-xylofuranosyl)-xypoxanthine phosphoramidite Referring to Figure 9, Inosine (1) was 5'-O-monomethoxyfritylated and 2'-O- silylated under standard conditions to afford 2 (Charubala, R; Pfleiderer, W. Heterocycles 1990, 30, 1141). Oxidation/reduction procedure afforded 3 in moderate yield (Matulic- Adamic, J.; Daniher, A.T.; Gonzalez, C; Beigelman, L. Bioorg. Med. Chem. Lett.. 1999, 9, 157): !H NMR (CDC13) δ 12.80 (br s, IH, NH), 8.11 (s, IH, H-8), 8.08 (s, IH, H-2), 7.45-6.80 (m, 14H, trityl), 5.85 (d, Jr 2>= 1.6, IH, H-l'), 4.83 (d, J2',3'=7.2, IH, H-2'), 4.46 (br s, IH, 3'-OH), 4.34 (m, IH, H-4'), 4.06 (m, IH, H-3'), 3.77 (s, 6H, 2 x OMe), 3.60 (app d, 2H, H-5', H-5"), 0.89 (s, 9H, t-Bu), 0.07 (s, 3H, Me), 0.06 (s, 3H, Me). Standard phosphitylation of 3 afforded the desired phosphoramidite 4. More acid stable 5'-O-MMT group is used in this particular case because applicant found that 5'-O-DMT protection is more labile in xylo nucleoside series than in ribo nucleoside series. The xylo-inosine was incoφorated into oligonucleotides using the standard procedures known in the art and as described herein.
Example 11 : Activity ofthe xylo-Inosine-modified NCH Ribozyme
Several NCH ribozymes with xylo-inosine at position 15.1 were designed (Figure 7) to cleave RNA containing GCA, ACA, UCA or the CCA triplet. These ribozymes were
synthesized and purified as described herein and tested using standard RNA cleavage reaction conditions (see Table 31, for example, and see below).
The ribozymes were chemically synthesized. The method of synthesis used followed the procedure for normal RNA synthesis as described above and in Usman et al., (1987 J. Am. Chem. Soc, 109, 7845), Scaringe et al., (1990 Nucleic Acids Res., 18, 5433) and
Wincott et al., supra, and made use of common nucleic acid protecting and coupling groups, such as dimethoxytrityl at the 5'-end, and phosphoramidites at the 3'-end. The average stepwise coupling yields were >98%>.
Ribozymes were purified by gel elecfrophoresis using general methods or were purified by high pressure liquid chromatography (HPLC; See Wincott et al., supra; the totality of which is hereby incoφorated herein by reference) and were resuspended in water. The sequences ofthe chemically synthesized ribozymes used in this study are shown below in Table 33.
Cleavage Reactions: Full-length or partially full-length, internally-labeled target RNA for ribozyme cleavage assay is prepared by in vitro transcription in the presence of
[alpha-32p] CTP, passed over a G 50 Sephadex column by spin chromatography and used as substrate RNA without further purification. Alternately, substrates were 5'-32p-end labeled using T4 polynucleotide kinase enzyme. Assays are performed by pre-warming a 2X concentration of purified ribozyme in ribozyme cleavage buffer (50 mM Tris-HCI, pH 7.5 at 37°C, 10 mM MgCtø) and the cleavage reaction was initiated by adding the 2X ribozyme mix to an equal volume of substrate RNA (maximum of 1-5 nM) that was also pre-warmed in cleavage buffer. As an initial screen, assays are carried out for 1 hour at o
37 C using a final concentration of 40 nM or 1 mM ribozyme, i.e., ribozyme excess. The reaction is quenched by the addition of an equal volume of 95% formamide, 20 mM EDTA, 0.05% bromophenol blue and 0.05% xylene cyanol after which the sample is heated to 95 C for 2 minutes, quick chilled and loaded onto a denaturing polyacrylamide gel. Substrate RNA and the specific RNA cleavage products generated by ribozyme cleavage are visualized on an autoradiograph ofthe gel. The percentage of cleavage is determined by Phosphor hnager® quantitation of bands representing the intact subsfrate and the cleavage products.
The results ofthe experiments are summarized in Table 32, which shows that NCH- xylo ribozymes are catalytically active to cleave target RNA.
Example 12: Activity of NCH Ribozyme variants The nucleic acid molecules ofthe instant invention allow for the ability to cleave a new set of 12 NCH triplets. Determination of single turnover rate constants at pH 6 of these ribozymes in the all ribo form show that with NCA type triplets, the cleavage rate is higher than at NUA sites. NCC and NUC site rates are similar, and NCU sites are slightly lower than NUU sites. Additional measurements of multiple turnover parameters ofthe all ribo ribozymes performed under non-saturating conditions using 5nM ribozyme and changing the substrate concentration from 50 to 500 nM at pH 7.4 with 10 mM Mg ++ at 37 °C gave Km= 100 nM and kcat=6.5 min -1 for GCA vs Km -30 nM and kcat =2.0 min _1 for GUA cleaving all ribo ribozymes. These data verify that the ribozymes with an I»C base pair are efficient catalysts in multiple turnover reactions and the relative order of activity between NCH and NUH cleavers established at pH 6 (Ludwig et al, 1998, Nucleic Acids Res., 26, 2279-2285) remains unchanged.
To gain more insight into the structural requirements ofthe 15.1- 16.1 base pair of the ribozymes ofthe instant invention, applicant synthesized several variants ofthe active 1-15.1 »C-16.1 structure and tested these ribozyme analogues with their coπesponding subsfrates. The influence of several core stabilization strategies on the activity ofthe NCH cleaving ribozymes was also investigated.
Various nucleoside analogs were incoφorated at position 15.1 ofthe ribozyme. Cleavage activity was tested with the complementary Fl* labeled substrates at pH 7.4 in the presence of 10 mM Mg ++ under conditions of ribozyme excess (i.e. single turnover conditions). The modified oligonucleotides were synthesized by standard oligonucleotide synthesis procedures. Xanthosine was protected using O-2 ,0-4pivaloyloxymethyl groups; N,N-dimethylguanosine with 6-O-( 2-nitrophenyl-)ethyl and 6-thio-inosine with S- cyanoethyl protecting groups. The cleavage activity ofthe ribozymes containing the 15.1 analogs is summarized in Figure 36. For comparison Figure 37 summarizes reported functional group modification studies performed at the A 15.1 residue in the A-15.1 »U- 16.1 context of NUH cleaving ribozymes.
Modifications at the purine 15.1 Nl and/or C6 positions (Figure 36 A, B, C)
In the 6-thio-inosine (A) (si) 15.1 substituted ribozyme, the original (1-15.1) position 6 O»H-N (C-16.1) bonds are replaced by weaker (sl-15.1) position 6 SΗ-N (C-16.1) hydrogen bonds while all other functional groups remain unchanged. Ribozymes with an adenosine (B) at position 15.1 (A- 15.1 ) are inactive with C-16.1 substrates since the ribozyme geometry requires the [A-15.1] position 6 amino group and the [C-16.1] position 4 amino group hydrogen-bond donor functional groups to be in close proximity. Similarly, low activity is observed with 1-15.1 ribozymes and U-16.1 subsfrates, where the [1-15.1] position 6 keto and [C-16.1] position 4 keto hydrogen-bond acceptor groups are opposed (Figure 37, B). Although inosine can form stable mismatch pairs with uridine in RNA duplexes or in tRNA anticodon-mRNA interactions, these results suggest that the geometry in the MJ mismatches differ from that ofthe A»U (or I*C) base pair in the active NUH ribozyme. Substitution of Nl-Methyl-inosine (C) in place of inosine at position 15.1 leads to complete loss of cleavage activity.
Modifications at the purine 15.1 C2 and/or N3 position (Figure 36 D, E, F)
The extremely low activity observed with the G-15.1 (D) substituted analog may be explained by the formation of a G-C Watson-Crick base pair. The replacement ofthe I«C pair with a G»C pair can significantly distort the geometry at the 15.1-16.1 position. G- 15.1 N2-alkylation (E) gives only minimal recovery of catalytic activity compared to G- 15.1, suggesting that the steric problems introduced by the bulky N-methyl groups may interfere with stacking interactions. The activity of this construct is significantly less than that of iso-G-15.1 (Figure 37, E) containing ribozymes in the standard A-U context. Xanthosine 15.1 (F) contains the same functional groups as inosine at the Nl and C6 sites but contains an additional hydrogen-bond donor site at position N3 along with a C2 carbonyl group. The complete lack ofactivity seen with this construct reinforces the importance ofthe purine N3 acceptor functionality in transition state formation. Similarly, 3-deaza-adenosine (Figure 37, F) containing ribozymes were also inactive. The C2 carbonyl ofthe 15.1 purine shows no significant negative interference in iso-guanosine containing 15.1 ribozymes.
Activity of modified core variants
To complete the characterization ofthe I*C pair containing ribozymes, the acceptance of various core substitution patterns was tested. Short subsfrates containing GCH and GUH (H= non G) triplets were compared using 3 different modified ribozymes. The acceptance of the U-42 '-O-alkyl substituent is the greatest with GCA triplets while U- 4 = 2'-deoxy-2'-amino uridine and U-4 = ribo uridine substituted ribozymes show a similar level of activity with NCH and NUH triplets. The results of this comparison are summarized in Table 64. In addition, a ribozyme construct in which ribo inosine replaces adenosine at positions 14 and 15.1 was tested which demonsfrated cleavage activity. Apart from the A-15.1 *U-16.1 to 1-15 .1 »C-16.1 change that reverses the polarity of an important H-bond in the ribozyme structure, no other functional group changes at the 15.1 purine residue seem to be compatible with the requirements of efficient catalysis. The 1-15.1 and A-15.1 ribozymes are equally suitable for practical applications because there are only minor differences in the acceptance of stabilizing residues.
Example 13: Activity of NCH Ribozyme to inhibit HER2 gene expression
Applicant has designed, synthesized and tested several NCH ribozymes and HH ribozymes targeted against HER2 RNA (see, for example, Tables 31 and 34) in cell proliferation assays.
Proliferation assay: The model proliferation assay used in the study can require a cell plating density of 2000 cells/well in 96-well plates and at least 2 cell doublings over a
5-day treatment period. To calculate cell density for proliferation assays, the FIPS (fluoro- imaging processing system) method well in the art was used. This method allows for cell density measurements after nucleic acids are stained with CyQuant® dye, and has the advantage of accurately measuring cell densities over a very wide range 1,000-100,000 cells/well in 96-well format.
Ribozymes (50-200 nM) were delivered in the presence of cationic lipid at 2.0 μg/mL and inhibition of proliferation was determined on day 5 post- treatment. Two full ribozyme screens were completed and 4 lead HH and 11 lead NCH ribozymes were chosen for further testing. Ofthe 15 lead Rzs chosen from primary screens, 4 NCH and 1
HHRzs continued to inhibit cell proliferation in subsequent experiments. NCH Rzs against sites, 2001 (RPI No. 17236), 2783 (RPI No. 17249), 2939 (RPI No. 17251) or 3998 (RPI No. 17262) caused inhibition of proliferation ranging from 25-60%) as compared to a scrambled confrol Rz (LA; RPI No. 17263). Ofthe five lead Rzs, the most efficacious is the NCH Rz (RPI No. 17251) against site 2939 of ΗER2 RNA. An example of results from cell culture assay is shown in Figure 3. Referring to Figure 3, NCH ribozymes and a HH ribozyme targeted against HER2 RNA are shown to cause significant inhibition of proliferation of cells. This shows that ribozymes, for instance, the NCH ribozymes are capable of inhibiting HER2 gene expression in mammalian cells.
Example 14: Activity of NCH Ribozyme to inhibit PKC alpha gene expression
The Protein Kinase C family contains twelve cuπently known isozymes divided into three classes: the classic, Ca^ dependent (PKCα, βl, βH, γ), the novel, non-Ca** dependent (PKCδ, ε, μ, η, θ) and the atypical (PKC ξ, i/λ); all of which are serine/threonine kinases. These isozymes show distinct and overlapping tissue, cellular, and subcellular distribution. They aid in the regulation of cell growth and differentiation through their response to second messenger products of lipid metabolism (Blobe, et al, 1996, Cancer Surveys, 27, 213-248). These second messengers include diacylglyceral (DAG), inositol-friphosphate (LP3), lysophospholipids, free fatty acids, and phosphatidate which act directly or in addition to changes in the Ca** concenfration. A simple model for PKCα activation follows a two step mechanism. First, membrane association of PKCα is through Ca*4" and phospholipid interactions and second, the kinase is activated by interaction with DAG. An example of a signal cascade subsequent to PKC activation is PKC's phosphorylation of c-Raf, which phosphorylates MEK, which phosphorylates MAP, which phosphorylates transcription factors such as Jun and thereby activates a mitogenic program in the nucleus. There are numerous substrates for the various PKC's, one which for PKCα ultimately stimulates franscription factors that activate P- glycoprotein (P-gp) causing the multi-drug resistant phenotype (MDR) (Blobe, et al, 1994, Cancer and Metastasis Reviews, 13, 411-431).
Cell Culture Review
PKC's have been implicated in tumor promotion since the discovery that these molecules can serve as receptors for tumor-promoting phorbol esters. An increase in PKC overexpression in numerous tumor cell lines and tumor tissues has also been demonsfrated. PKC overexpression has been shown to be associated with increased invasion and metastasis in mouse Lewis lung carcinoma, mouse B16 melanoma (Lee et al, 1997, Molecular Carcinogenesis, 18, 44-53), mouse mammary adenocarcinoma, mouse fibrosarcoma, human lung carcinoma (Wang and Liu, 1998, Acta Pharmacologica Sinica, 19, 265-268), human bladder carcinoma, human pancreatic cancer (Denham et al., 1998, Surgery, 124, 218-223), and human gastric cancer (Dean et al, 1996, Cancer Research, 56, 3499-3507). Mounting evidence suggests PKCα can stimulate adhesion molecule expression and can directly act on these membrane bound species as subsfrates, thereby modulating cellular adhesion to the extracellular matrix and increasing metastic potential. Furthermore, human surgical specimens have demonsfrated elevated PKC in breast tumors, thyroid carcinomas and melanomas (Becker et al, 1990, Oncogene, 5, 1133- 1139).
Utz et al, 1994, Int. J. Cancer, 57, 104-110, describe a cell proliferation assay in which small molecule inhibitors of PKC demonstrate anti-pro liferative activity in CCRF- VCR 1000 and KB-8511 cells with the multidrug resistant (MDR) phenotype. PKCα is overexpressed in tumor tissues that express the MDR phenotype. This phenotype is associated with the expression of a 170 kDa broad specificity drug efflux pump, P-gp. PKCα phosphorylation of P-gp has been shown in vitro. In addition, PKC expression coπelates with resistance to doxorubicin and high P-gp levels in human renal carcinoma and non-small cell lung carcinoma, inhibitors of PKC partially reverse the MDR phenotype and decrease phosphorylation of P-gp (Caponigro et al, 1997, Anti-Cancer Drugs, 8, 26-33).
Dean et al, 1994, Journal of Biological Chemistry, 269, 16416-24, describe cell culture studies in which antisense targeting of PKC α resulted in the potent inhibition of mRNA and protein expression in human lung carcinoma (A549) cells. In this study, PKC α inhibition resulted in the reduced induction of intercellular adhesion molecule 1 (ICAM- 1) mRNA by phorbol esters.
Yano et al, 1999, Endocrinology, 140, 4622-4632, describe a cell proliferation study in which down regulation of different PKC isoforms, including PKCα, results in the inhibition of insulin like growth factor I induced vascular smooth muscle cell proliferation, migration, and gene expression. Wang et al, 1999, Experimental Cell Research, 250, 253-263, describe cell culture studies in which antisense inhibition of PKCα results in the reversal ofthe transformed phenotype in human lung carcinoma (LTEPa-2) cells. In this study, the amounts of PKCα protein and total PKC activity were decreased when compared to control cells. Sioud and Sorensen, 1998, Nature Biotechnology, 16, 556-561, describe hammerhead ribozyme inhibition of PKCα in rat glioma cell lines (BT4C and BT4Cn). This study demonsfrated inhibition of malignant glioma cell proliferation along with the inhibition of regulatory Bcl-xL protein expression. BC1-XL is overexpressed in glioma cells and is an apoptosis inhibitor. The ribozyme mediated inhibition of cell proliferation presumably results from apoptosis induction of transformed glioma cells through suppression of PKCα and Bcl-xL (Leirdal and Sioud, 1999, British J. of Cancer, 80, 1558- 1564). Animal Models
Evaluating the efficacy of anti-PKCα agents in animal models is an important prerequisite to human clinical trials. A variety of mouse xenograft models using human tumor cell lines have been developed using cell lines which express high levels of PKCα protein. McGraw et α/, 1997 ', Anti-Cancer Drug Design, 12, 315-326, describe mouse xenograft models using human breast (MDA MB-321), prostate (Du-145), colon (Colo 205, WiDr), lung (NCI H69, H209, J460, H520, A549), bladder (T-24), and melanoma (SK-mel 1) carcinoma cells. Antisense oligonucleotides targeting PKCα administered infravenously following s.c. transplanted tumor cells resulted in dose dependant decreases in tumor size when compared to confrols in most cases. Similar studies using T-24 bladder carcinoma, non-small cell lung carcinoma (A549), and Colo 205 colon carcinoma mouse xenografts are described in Dean et al, 1996, Biochemical Society Transactions, 24, 623. Sioud and Sorensen, 1998, Nature Biotechnology, 16, 556-561, describe a rat model in which inbred syngeneic BDLX rats were inoculated subcutaneously with BT4Cn glioma cells. After approximately three weeks, rats were freated with a single injection of
ribozyme targeting PKCα resulting in inhibition of tumor growth as determined by tumor size and/or weight when compared to confrols. The above studies provide proof that inhibition of PKCα expression by anti-PKCα agents causes inhibition of tumor growth in animals. Lead anti-PKCα ribozymes chosen from in vitro assays can be further tested in mouse xenograft models. Ribozymes can be first tested alone and then in combination with standard chemotherapies. Animal Model Development
Human lung (A549, NCI H520) tumor and breast (MDA-MB 231) cell lines can be characterized to establish their growth curves in mice. These cell lines are been implanted into both nude and SCLD mice and primary tumor volumes are measured 3 times per week. Growth characteristics of these tumor lines using a Matrigel implantation format can also be established. In addition, the use of other cell lines that have been engineered to express high levels of PKCα can also be used. The tumor cell line(s) and implantation method that supports the most consistent and reliable tumor growth can be used in animal studies to test promising PKCα ribozyme(s). Ribozymes can be administered by daily subcutaneous injection or by continuous subcutaneous infusion from Alzet mini osmotic pumps beginning 3 days after tumor implantation and continuing for the duration ofthe study. Group sizes of at least 10 animals are employed. Efficacy is determined by statistical comparison of tumor volume of ribozyme-freated animals to a control group of animals freated with saline alone. Because the growth of these tumors is generally slow (45-60 days), an initial endpoint will be the time in days it takes to establish an easily measurable primary tumor (i.e. 50-100 mm3) in the presence or absence of ribozyme treatment. Clinical Summary Overview
Ribozymes targeting PKCα have strong potential to develop into useful therapeutics directed towards numerous cancer types. Lung cancer is the leading cause of cancer deaths for both men and women in the USA. The incidence of lung cancer in the United States is -172,000 cases per year, accounting for 14% of cancer diagnoses. Approximately 158,000 die each year of lung cancer, accounting for 28% of all cancer deaths. Numerous other
indications exist including cancers ofthe bladder, colon, breast, prostate, and ovary in addition to melanoma and glioblastoma.
McGraw et α/., 1997 , Anti-Cancer Drug Design, 12, 315-326, describe a Phase I trial for ISIS 3521/CGP 64128A, a PKC alpha antisense construct. In this trial, ISIS 3521/CGP 64128A was administered as either a two-hour i.v. infusion three times per week for three consecutive weeks, or as a continuous i.v. infusion for twenty-one consecutive days. The authors report that patients demonstrated excellent tolerance to the antisense compound when administered at doses of up to 2.5 mg/kg by the two-hour i.v. infusion and at 1.5 mg/kg/day by continuous i.v. infusion. In patients receiving the two-hour i.v. infusion schedule, the post-infusion plasma concenfration ofthe compound increased proportional to the dose, and metabolites were determined to have been cleared rapidly from plasma with a half-life of thirty to forty- five minutes. These metabolites were composed of chain- shortened oligonucleotides, consistent with exonuclease-mediated degradation. No evidence of accumulation, induction, or inhibition of metabolism was found after the administration of repetitive doses. Therapy
Treatment options for lung cancer are determined by the type and stage ofthe cancer and include surgery, radiation therapy, and chemotherapy. For many localized cancers, surgery is usually the treatment of choice. Because the disease has usually spread by the time it is discovered, radiation therapy and chemotherapy are often needed in combination with surgery. Chemotherapy alone or combined with radiation has replaced surgery as the treatment of choice for small cell lung cancer; on this regimen, a large percentage of patients experience remission, which in some cases is long-lasting. The 1-year relative survival rates for lung cancer have increased from 32% in 1973 to 41% in 1994, largely due to improvements in surgical techniques. The 5-year relative survival rate for all stages combined is only 14%. The survival rate is 50% for cases detected when the disease is still localized, but only 15% of lung cancers are discovered that early.
Common chemotherapies include various combinations of cytotoxic drugs to kill the cancer cells. These drugs include paclitaxel (Taxol), docetaxel, cisplatin, methofrexate, cyclophosphamide, doxorubin, fluorouracil etc. Significant toxicities are associated with these cytotoxic therapies. Well-characterized toxicities include nausea and vomiting,
myelosuppression, alopecia and mucosity. Serious cardiac problems are also associated with certain ofthe combinations, e.g. doxorubin and paclitaxel, but are less common.
Applicant has designed several NCH ribozymes targeted against PKCα RNA (Genebank accession No NM_002737) (see, for example, Table 63). These ribozymes are used first in a proliferation assay that is used to select ribozyme leads.
Proliferation assay: The model proliferation assay useful in the study can require a cell plating density of 2000 cells/well in 96-well plates and at least 2 cell doublings over a 5-day freatment period. To calculate cell density for proliferation assays, the FLPS (fluoro- imaging processing system) method well known in the art can be used. This method allows for cell density measurements after nucleic acids are stained with CyQuant® dye, and has the advantage of accurately measuring cell densities over a very wide range 1,000-100,000 cells/well in 96-well format.
Ribozymes (50-200 nM) are delivered in the presence of cationic lipid at 2.0 μg/mL and inhibition of proliferation is determined on day 5 post-treatment. Two full ribozyme screens are usually completed and lead ribozymes are chosen for further testing. Ofthe lead ribozymes chosen from primary screens, ribozymes which continue to inhibit cell proliferation in subsequent experiments are selected for PKCα RNA and protein inhibition studies.
Example 15: Nucleoside Triphosphates and their incoφoration into oligonucleotides
The synthesis of nucleotide triphosphates and their incoφoration into nucleic acids using polymerase enzymes has greatly assisted in the advancement of nucleic acid research. The polymerase enzyme utilizes nucleotide triphosphates as precursor molecules to assemble oligonucleotides. Each nucleotide is attached by a phosphodiester bond formed through nucleophilic attack by the 3' hydroxyl group ofthe oligonucleotide 's last nucleotide onto the 5' triphosphate ofthe next nucleotide. Nucleotides are incoφorated one at a time into the oligonucleotide in a 5' to 3' direction. This process allows RNA to be produced and amplified from virtually any DNA or RNA templates.
Most natural polymerase enzymes incoφorate standard nucleotide triphosphates into nucleic acid. For example, a DNA polymerase incoφorates dATP, dTTP, dCTP, and dGTP into DNA and an RNA polymerase generally incoφorates ATP, CTP, UTP, and
GTP into RNA. There are however, certain polymerases that are capable of incoφorating non-standard nucleotide triphosphates into nucleic acids (Joyce, 1997, PNAS 94, 1619- 1622, Huang et al., Biochemistry 36, 8231-8242).
Before nucleosides can be incoφorated into RNA transcripts using polymerase enzymes they must first be converted into nucleotide triphosphates which can be recognized by these enzymes. Phosphorylation of unblocked nucleosides by treatment with POCl and trialkyl phosphates was shown to yield nucleoside 5'- phosphorodichloridates (Yoshikawa et al, 1969, Bull. Chem. Soc. (Japan) 42, 3505). Adenosine or 2'-deoxyadenosine 5 '-triphosphate was synthesized by adding an additional step consisting of freatment with excess tri-n-butylammonium pyrophosphate in DMF followed by hydrolysis (Ludwig, 1981, Acta Biochim. et Biophys. Acad. Sci. Hung. 16, 131-133).
Non-standard nucleotide triphosphates are not readily incoφorated into RNA transcripts by traditional RNA polymerases. Mutations have been introduced into RNA polymerase to facilitate incoφoration of deoxyribonucleotides into RNA (Sousa & Padilla, 1995, EMBO J. 14,4609-4621, Bonner et al, 1992, EMBO J. 11, 3767-3775, Bonner et al., 1994, J. Biol. Chem. 42, 25120-25128, Aurup et al, 1992, Biochemistry 31, 9636-9641).
McGee et al, International PCT Publication No. WO 95/35102, describes the incoφoration of 2'-NH2-NTP's, 2'-F-NTP's, and 2'-deoxy-2'-benzyloxyamino UTP into RNA using bacteriophage T7 polymerase.
Wieczorek et al, 1994, Bioorganic & Medicinal Chemistry Letters A, 987-994, describes the incoφoration of 7-deaza-adenosine triphosphate into an RNA transcript using bacteriophage T7 RNA polymerase.
Lin et al, 1994, Nucleic Acids Research 22, 5229-5234, reports the incoφoration of 2'-NH2-CTP and 2'-NH2-UTP into RNA using bacteriophage T7 RNA polymerase and polyethylene glycol containing buffer. The article describes the use ofthe polymerase synthesized RNA for in vitro selection of aptamers to human neutrophil elastase (HNE).
This invention relates to novel nucleotide triphosphate (NTP) molecules, and their incoφoration into nucleic acid molecules, including nucleic acid catalysts. The NTPs of the instant invention are distinct from other NTPs known in the art. The invention further relates to incoφoration of these nucleotide triphosphates into oligonucleotides using an RNA polymerase; the invention further relates to novel transcription conditions for the
incoφoration of modified (non-standard) and unmodified NTP's, into nucleic acid molecules. Further, the invention relates to methods for synthesis of novel NTP's
In a first aspect, the invention features NTP's having the formula triphosphate-OR, for example the following formula 3:

where R is any nucleoside; specifically the nucleosides 2'-O-methyl-2,6- diaminopurine riboside; 2'-deoxy-2'amino-2,6-diaminopurine riboside; 2'-(N-alanyl) amino-2'-deoxy-uridine; 2'-(N-phenylalanyl)amino-2'-deoxy-uridine; 2'-deoxy -2'-(N-β- alanyl) amino ; 2'-deoxy-2'-(lysiyl) amino uridine; 2'-C-allyl uridine; 2'-O-amino-uridine; 2'-O-methylthiomethyl adenosine; 2'-O-methylthiomethyl cytidine ; 2'-O- methylthiomethyl guanosine; 2'-O-methylthiomethyl-uridine; 2'-deoxy-2'-(N-histidyl) amino uridine; 2'-deoxy-2'-amino-5-methyl cytidine; 2'-(N-β-carboxamidine-β- alanyl)amino-2 ' -deoxy-uridine; 2 ' -deoxy-2 ' -(Ν- β -alanyl)-guanosine; 2 ' -O-amino- adenosine; 2'-(N-lysyl)amino-2'-deoxy-cytidine; 2'-Deoxy -2'-(L-histidine) amino Cytidine; 5-Imidazoleacetic acid 2'-deoxy uridine, 5-[3-(Ν-4- imidazoleacetyl)aminopropynyl] -2 '-O-methyl uridine, 5-(3-aminopropynyl)-2'-O-methyl uridine, 5-(3-aminopropyl)-2'-O-methyl uridine, 5-[3-(N-4-imidazoleacetyl)aminopropyl]- 2'-O-methyl uridine, 5-(3-aminopropyl)-2'-deoxy-2-fluoro uridine, 2'-Deoxy-2'-(β-alanyl- L-histidyl)amino uridine, 2'-deoxy-2'-β-alaninamido-uridine, 3 -(2 '-deoxy-2' -fluoro- β-D- ribofuranosyl)piperazino[2,3-D]pyrimidine-2-one, 5-[3-(N-4- imidazoleacetyl)aminopropyl]-2'-deoxy-2'-fluoro uridine, 5-[3-(N-4- imidazoleacetyl)aminopropynyl] -2 ' -deoxy-2 ' -fluoro uridine, 5 -E-(2-carbox yvinyl-2 ' - deoxy-2 '-fluoro uridine, 5-[3-(N-4-aspartyl)aminopropynyl-2'-fluoro uridine, 5-(3- aminopropyl)-2'-deoxy-2-fluoro cytidine, and 5-[3-(N-4-succynyl)aminopropyl-2'-deoxy- 2-fluoro cytidine.
In a second aspect, the invention features inorganic and organic salts ofthe nucleoside triphosphates ofthe instant invention.
In a third aspect, the invention features a process for the synthesis of pyrimidine nucleotide triphosphate (such as UTP, 2'-O-MTM-UTP, dUTP and the like) including the steps of monophosphorylation where the pyrimidine nucleoside is contacted with a mixture having a phosphorylating agent (such as phosphorus oxychloride, phospho-tris-
5 triazolides, phospho-tris-triimidazolides and the like), trialkyl phosphate (such as triethylphosphate or trimethylphosphate or the like) and a hindered base (such as dimethylaminopyridine, DMAP and the like) under conditions suitable for the formation of pyrimidine monophosphate; and pyrophosphorylation where the pyrimidine monophosphate is contacted with a pyrophosphorylating reagent (such as
10 tributylammonium pyrophosphate) under conditions suitable for the formation of pyrimidine triphosphates.
By "nucleotide triphosphate" or "NTP" is meant a nucleoside bound to three inorganic phosphate groups at the 5' hydroxyl group ofthe modified or unmodified ribose or deoxyribose sugar where the 1 ' position ofthe sugar may comprise a nucleic acid base
15 or hydrogen. The triphosphate portion may be modified to include chemical moieties which do not destroy the functionality ofthe group (i.e., allow incoφoration into an RNA molecule).
In another prefeπed embodiment, nucleotide triphosphates (NTPs) ofthe instant invention are incoφorated into an oligonucleotide using an RNA polymerase enzyme.
!0 RNA polymerases include but are not limited to mutated and wild type versions of bacteriophage T7, SP6, or T3 RNA polymerases. Applicant has also found that the NTPs ofthe present invention can be incoφorated into oligonucleotides using certain DNA polymerases, such as Taq polymerase.
In yet another prefeπed embodiment, the invention features a process for
!5 incoφorating modified NTP's into an oligonucleotide including the step of incubating a mixture having a DNA template, RNA polymerase, NTP, and an enhancer of modified NTP incoφoration under conditions suitable for the incoφoration ofthe modified NTP into the oligonucleotide.
By "enhancer of modified NTP incoφoration" is meant a reagent which facilitates
SO the incoφoration of modified nucleotides into a nucleic acid transcript by an RNA polymerase. Such reagents include, but are not limited to, methanol, LiCl, polyethylene glycol (PEG), diethyl ether, propanol, methyl amine, ethanol, and the like.
In another prefeπed embodiment, the modified nucleotide triphosphates can be incoφorated by franscription into a nucleic acid molecules including enzymatic nucleic acid, antisense, 2-5A antisense chimera, oligonucleotides, triplex forming oligonucleotide (TFO), aptamers and the like (Stull et al, 1995 Pharmaceutical Res. 12, 465). By "triplex forming oligonucleotides (TFO)" it is meant an oligonucleotide that can bind to a double-stranded DNA in a sequence-specific manner to form a triple-strand helix. Formation of such triple helix structure has been shown to inhibit transcription of the targeted gene (Duval- Valentin et al, 1992 Proc. Natl. Acad. Sci. USA 89, 504).
In yet another prefeπed embodiment, the modified nucleotide triphosphates ofthe instant invention can be used for combinatorial chemistry or in vitro selection of nucleic acid molecules with novel function. Modified oligonucleotides can be enzymatically synthesized to generate libraries for screening.
In another prefeπed embodiment, the invention features nucleic acid based techniques (e.g., enzymatic nucleic acid molecules), antisense nucleic acids, 2-5 A antisense chimeras, triplex DNA, antisense nucleic acids containing RNA cleaving chemical groups) isolated using the methods described in this invention and methods for their use to diagnose, down regulate or inhibit gene expression.
In yet another prefeπed embodiment, the invention features enzymatic nucleic acid molecules targeted against HER2 RNA, specifically including ribozymes in the class II (zinzyme) motif.
Targets, for example HER2 RNA, for useful ribozymes and antisense nucleic acids can be determined, for example, as described in Draper et al, WO 93/23569; Sullivan et al, WO 93/23057; Thompson et al, WO 94/02595; Draper et al, WO 95/04818; McSwiggen et al, US Patent Nos. 5,525,468 and 5,646,042, all are hereby incoφorated by reference herein in their totalities. Other examples include the following PCT applications, which concern inactivation of expression of disease-related genes: WO 95/23225, and WO 95/13380; all of which are incoφorated by reference herein.
In yet another prefeπed embodiment, the invention features a process for incoφorating a plurality of compounds of formula 3. In yet another embodiment, the invention features a nucleic acid molecule with catalytic activity having formula 4:
In the formula shown above X, Y, and Z represent independently a nucleotide or a non-nucleotide linker, which may be same or different; • indicates hydrogen bond formation between two adjacent nucleotides which may or may not be present; Y' is a nucleotide complementary to Y; Z' is a nucleotide complementary to Z; 1 is an integer greater than or equal to 3 and preferably less than 20, more specifically 4, 5, 6, 7, 8, 9, 10, 11, 12, or 15; m is an integer greater than 1 and preferably less than 10, more specifically 2, 3, 4, 5, 6, or 7; n is an integer greater than 1 and preferably less than 10, more specifically 3, 4, 5, 6, or 7; o is an integer greater than or equal to 3 and preferably less than 20, more specifically 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, or 15; 1 and o may be the same length (1 = o) or different lengths (1 ≠ o); each X(l) and X(o) are oligonucleotides which are of sufficient length to stably interact independently with a target nucleic acid sequence (the target can be an RNA, DNA or RNA/DNA mixed polymers); W is a linker of > 2 nucleotides in length or may be a non-nucleotide linker; A, U, C, and G represent the nucleotides; G is a nucleotide, preferably 2'-O-methyl or ribo; A is a nucleotide, preferably 2'-O-methyl or ribo; U is a nucleotide, preferably 2'-amino (e.g., 2'-NH2 or 2'-O- NH2), 2'-O-methyl or ribo; C represents a nucleotide, preferably 2'-amino (e.g., 2'-NH2 or 2'-O-
NH2), and represents a chemical linkage (e.g. a phosphate ester linkage, amide linkage, phosphorothioate, phosphorodithioate or others known in the art).
In yet another embodiment, the invention features a nucleic acid molecule with catalytic activity having formula 5:
In the formula shown above X, Y, and Z represent independently a nucleotide or a non-nucleotide linker, which may be same or different; • indicates hydrogen bond formation between two adjacent nucleotides which may or may not be present; Z' is a nucleotide complementary to Z; 1 is an integer greater than or equal to 3 and preferably less than 20, more specifically 4, 5, 6, 7, 8, 9, 10, 11, 12, or 15; n is an integer greater than 1 and preferably less than 10, more specifically 3, 4, 5, 6, or 7; o is an integer greater than or equal to 3 and preferably less than 20, more specifically 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, or
15; 1 and o may be the same length (1 = o) or different lengths (1 ≠ o); each Xm and X(o) are oligonucleotides which are of sufficient length to stably interact independently with a target nucleic acid sequence (the target can be an RNA, DNA or RNA/DNA mixed polymers); preferably has a G at the 3 '-end, XQ) preferably has a G at the 5 '-end; W is a linker of ≥ 2 nucleotides in length or may be a non-nucleotide linker; Y is a linker of > 1 nucleotides in length, preferably G, 5' -CA-3', or 5' -CAA-3', or may be a non-nucleotide linker; A, U, C, and G represent nucleotides; G is a nucleotide, preferably 2'-O-methyl, 2'-deozy-2'-fluoro, or 2'-OH; A is a nucleotide, preferably 2'-O-methyl, 2'-deozy-2'-
fluoro, or 2'-OH; U is a nucleotide, preferably 2'-O-methyl, 2 '-deozy-2' -fluoro, or 2'-OH;
C represents a nucleotide, preferably 2'-amino (e.g., 2'-NH2 or 2'-O- NH2, and represents a chemical linkage (e.g. a phosphate ester linkage, amide linkage, phosphorothioate, phosphorodithioate or others known in the art). The enzymatic nucleic acid molecules of Formula 4 and Formula 5 may independently comprise a cap structure which may independently be present or absent.
In yet another prefeπed embodiment, the 3 '-cap is selected from a group comprising,
4',5'-methylene nucleotide; l-(beta-D-erythrofuranosyl) nucleotide; 4'-thio nucleotide; carbocyclic nucleotide; 5'-amino-alkyl phosphate; l,3-diamino-2-propyl phosphate; 3- aminopropyl phosphate; 6-aminohexyl phosphate; 1,2-aminododecyl phosphate; hydroxypropyl phosphate; 1,5-anhydrohexitol nucleotide; L-nucleotide; alpha-nucleotide; modified base nucleotide; phosphorodithioate; tΛreo-pentofuranosyl nucleotide; acyclic
3',4'-seco nucleotide; 3,4-dihydroxybutyl nucleotide; 3,5-dihydroxypentyl nucleotide; 5'-5'- inverted nucleotide moiety; 5'-5'-inverted abasic moiety; 5'-phosphoramidate; 5'- phosphorothioate; 1,4-butanediol phosphate 5'-amino; bridging and/or non-bridging 5'- phosphoramidate, phosphorothioate and/or hosphorodithioate; bridging or non bridging methylphosphonate and 5'-mercapto moieties (for more details, see Beaucage and Iyer,
1993, Tetrahedron 49, 1925; incoφorated by reference herein).
In another aspect, the invention provides mammalian cells containing one or more nucleic acid molecules and/or expression vectors of this invention. The one or more nucleic acid molecules may independently be targeted to the same or different sites.
Nucleotide Synthesis
Addition of dimethylaminopyridine (DMAP) to the phosphorylation protocols known in the art can greatly increase the yield of nucleotide monophosphates while decreasing the reaction time. Synthesis ofthe nucleosides ofthe invention have been described in several publications and Applicants previous applications (Beigelman et al, Intemational PCT publication No. WO 96/18736; Dudzcy et al, Int. PCT Pub. No. WO 95/11910; Usman et al, Int. PCT Pub. No. WO 95/13378; Matulic-Adamic et al, 1997, Tetrahedron Lett. 38, 203; Matulic-Adamic et al, 1997, Tetrahedron Lett. 38, 1669; all of which are incoφorated herein by reference). These nucleosides are dissolved in triethyl phosphate and chilled in an ice bath. Phosphorus oxychloride (POCl3) is then added
followed by the introduction of DMAP. The reaction is then warmed to room temperature and allowed to proceed for 5 hours. This reaction allows the formation of nucleotide monophosphates which can then be used in the formation of nucleotide triphosphates. Tributylamine is added followed by the addition of anhydrous acetonitrile and tributylammonium pyrophosphate. The reaction is then quenched with TEAB and stiπed overnight at room temperature (about 20°C). The triphosphate is purified using Sephadex® column purification or equivalent and/or HPLC and the chemical structure is confirmed using NMR analysis. Those skilled in the art will recognize that the reagents, temperatures ofthe reaction, and purification methods can easily be alternated with substitutes and equivalents and still obtain the desired product.
Nucleotide Triphosphates
The invention provides nucleotide triphosphates which can be used for a number of different functions. The nucleotide triphosphates formed from nucleosides found in Table 45 are unique and distinct from other nucleotide triphosphates known in the art.
Incoφoration of modified nucleotides into DNA or RNA oligonucleotides can alter the properties ofthe molecule. For example, modified nucleotides can hinder binding of nucleases, thus increasing the chemical half-life ofthe molecule. This is especially important if the molecule is to be used for cell culture or in vivo. It is known in the art that the introduction of modified nucleotides into these molecules can greatly increase the stability and thereby the effectiveness ofthe molecules (Burgin et al, 1996, Biochemistry 35, 14090-14097; Usman et al, 1996, Curr. Opin. Struct. Biol. 6, 527-533).
Modified nucleotides are incoφorated using either wild type or mutant polymerases. For example, mutant T7 polymerase is used in the presence of modified nucleotide triphosphate(s), DNA template and suitable buffers. Those skilled in the art will recognize that other polymerases and their respective mutant versions can also be utilized for the incoφoration of NTP's ofthe invention. Nucleic acid transcripts were detected by incoφorating radiolabelled nucleotides (α-32P NTP). The radiolabeled NTP contained the same base as the modified triphosphate being tested. The effects of methanol, PEG and LiCl were tested by adding these compounds independently or in combination. Detection and quantitation ofthe nucleic acid transcripts was performed using a Molecular Dynamics
Phosphorlmager. Efficiency of franscription was assessed by comparing modified nucleotide triphosphate incoφoration with all-ribonucleotide incoφoration control. Wild- type polymerase was used to incoφorate NTP's using the manufacturer's buffers and instructions (Boehringer Mannheim).
Transcription Conditions
Incoφoration rates of modified nucleotide triphosphates into oligonucleotides can be increased by adding to traditional buffer conditions, several different enhancers of modified NTP incoφoration. Applicant has utilized methanol and LiCl in an attempt to increase incoφoration rates of dNTP using RNA polymerase. These enhancers of modified NTP incoφoration can be used in different combinations and ratios to optimize transcription. Optimal reaction conditions differ between nucleotide triphosphates and can readily be determined by standard experimentation. Overall, however, Applicant has found that inclusion of enhancers of modified NTP incoφoration such as methanol or inorganic compound such as lithium chloride increase the mean transcription rates. Applicant synthesized pyrimidine nucleotide triphosphates using DMAP in the reaction. For purines, applicant utilized standard protocols previously described in the art (Yoshikawa et al supra;. Ludwig, supra). Described below is one example of a pyrimdine nucleotide triphosphate and one purine nucleotide triphosphate synthesis.
Synthesis of purine nucleotide triphosphates: 2'-O-methyl-guanosine-5 '-triphosphate
2'-O-methyl guanosine nucleoside (0.25 grams, 0.84 mmol) was dissolved in triethyl phosphate (5.0) ml by heating to 100°C for 5 minutes. The resulting clear, colorless solution was cooled to 0°C using an ice bath under an argon atmosphere. Phosphorous oxychloride (1.8 eq., 0.141 ml) was then added to the reaction mixture with vigorous stirring. The reaction was monitored by HPLC, using a sodium perchlorate gradient. After 5 hours at 0°C, tributylamine (0.65 ml) was added followed by the addition of anhydrous acetonitrile (10.0 ml), and after 5 minutes (reequilibration to 0°C) tributylammonium pyrophosphate (4.0 eq., 1.53 g) was added. The reaction mixture was quenched with 20 ml of 2M TEAB after 15 minutes at 0°C (HPLC analysis with above conditions showed consumption of monophosphate at 10 minutes) then stiπed overnight at room temperature, the mixture was evaporated in vacuo with methanol co-evaporation
(4x) then diluted in 50 ml 0.05M TEAB. DEAE sephadex purification was used with a gradient of 0.05 to 0.6 M TEAB to obtain pure triphosphate (0.52 g, 66.0% yield) (elutes around 0.3M TEAB); the purity was confirmed by HPLC and NMR analysis.
Synthesis of Pyrimidine nucleotide triphosphates: 2'-O-methylthiomethyl-uridine-5'- triphosphate
2'-O-methylthiomethyl uridine nucleoside (0.27 grams, 1.0 mmol) was dissolved in triethyl phosphate (5.0 ml). The resulting clear, colorless solution was cooled to 0°C with an ice bath under an argon atmosphere. Phosphorus oxychloride (2.0 eq., 0.190 ml) was then added to the reaction mixture with vigorous stirring. Dimethylaminopyridine
(DMAP, 0.2eq., 25 mg) was added, the solution warmed to room temperature and the reaction was monitored by HPLC, using a sodium perchlorate gradient. After 5 hours at 20°C, tributylamine (1.0 ml) was added followed by anhydrous acetonitrile (10.0 ml), and after 5 minutes tributylammonium pyrophosphate (4.0 eq., 1.8 g) was added. The reaction mixture was quenched with 20 ml of 2M TEAB after 15 minutes at 20°C (HPLC analysis with above conditions showed consumption of monophosphate at 10 minutes) then stiπed overnight at room temperature. The mixture was evaporated in vacuo with methanol co- evaporation (4x) then diluted in 50 ml 0.05M TEAB. DEAE fast flow Sepharose purification with a gradient of 0.05 to 1.0 M TEAB was used to obtain pure triphosphate (0.40 g, 44%) yield) (elutes around 0.3M TEAB) as determined by HPLC and NMR analysis.
Utilization of DMAP in Uridine 5 '-Triphosphate Synthesis
The reactions were performed on 20 mg aliquots of nucleoside dissolved in 1 ml of triethyl phosphate and 19 ul of phosphorus oxychloride. The reactions were monitored at 40 minute intervals automatically by HPLC to generate yield-of-product curves at times up to 18 hours. A reverse phase column and ammonium acetate/ sodium acetate buffer system (50mM & lOOmM respectively at pH 4.2) was used to separate the 5', 3', 2' monophosphates (the monophosphates elute in that order) from the 5 '-triphosphate and the starting nucleoside. The data is shown in Table 46. These conditions doubled the product yield and resulted in a 10-fold improvement in the reaction time to maximum yield (1200 minutes down to 120 minutes for a 90%> yield). Selectivity for 5'-monophosphorylation
was observed for all reactions. Subsequent triphosphorylation occuπed in nearly quantitative yield.
Materials Used in Bacteriophage T7 RNA Polymerase Reactions Buffer 1: Reagents are mixed together to form a 10X stock solution of buffer 1
(400 mM Tris-Cl [pH 8.1], 200 mM MgCl2, 100 mM DTT, 50 mM spermidine, and 0.1% triton® X-100). Prior to initiation ofthe polymerase reaction methanol, LiCl is added and the buffer is diluted such that the final reaction conditions for condition 1 consisted of : 40mM tris (pH 8.1), 20mM MgCl2, 10 mM DTT, 5 mM spermidine, 0.01% triton® X- 100, 10% methanol, and 1 mM LiCl.
BUFFER 2: Reagents are mixed together to form a 10X stock solution of buffer 2 (400 mM Tris-Cl [pH 8.1], 200 mM MgCl2, 100 mM DTT, 50 mM spermidine, and 0.1% triton® X-100). Prior to initiation ofthe polymerase reaction PEG, LiCl is added and the buffer is diluted such that the final reaction conditions for buffer 2 consisted of : 40mM tris (pH 8.1), 20mM MgCl2, 10 mM DTT, 5 mM spermidine, 0.01% triton® X-100, 4% PEG, and 1 mM LiCl.
BUFFER 3: Reagents are mixed together to form a 10X stock solution of buffer 3 (400 mM Tris-Cl [pH 8.0], 120 mM MgCl2, 50 mM DTT, 10 mM spermidine and 0.02% triton® X-100). Prior to initiation ofthe polymerase reaction PEG is added and the buffer is diluted such that the final reaction conditions for buffer 3 consisted of : 40mM tris (pH 8.0), 12 mM MgCl2, 5 mM DTT, 1 mM spermidine, 0.002% triton® X-100, and 4% PEG.
BUFFER 4: Reagents are mixed together to form a 10X stock solution of buffer 4 (400 mM Tris-Cl [pH 8.0], 120 mM MgCl2, 50 mM DTT, 10 mM spermidine and 0.02% triton® X-100). Prior to initiation ofthe polymerase reaction PEG, methanol is added and the buffer is diluted such that the final reaction conditions for buffer 4 consisted of :
40mM tris (pH 8.0), 12 mM MgCl2, 5 mM DTT, 1 mM spermidine, 0.002% triton® X- 100, 10% methanol, and 4%> PEG.
BUFFER 5: Reagents are mixed together to form a 10X stock solution of buffer 5 (400 mM Tris-Cl [pH 8.0], 120 mM MgCl2, 50 mM DTT, 10 mM spermidine and 0.02% triton® X-100). Prior to initiation ofthe polymerase reaction PEG, LiCl is added and the buffer is diluted such that the final reaction conditions for buffer 5 consisted of : 40mM
tris (pH 8.0), 12 mM MgCl2, 5 mM DTT, 1 mM spermidine, 0.002% triton® X-100, 1 mM LiCl and 4% PEG.
BUFFER 6: Reagents are mixed together to form a 10X stock solution of buffer 6 (400 mM Tris-Cl [pH 8.0], 120 mM MgCl2, 50 mM DTT, 10 mM spermidine and 0.02% triton® X-100). Prior to initiation ofthe polymerase reaction PEG, methanol is added and the buffer is diluted such that the final reaction conditions for buffer 6 consisted of : 40mM tris (pH 8.0), 12 mM MgCl2, 5 mM DTT, 1 mM spermidine, 0.002% triton® X- 100, 10% methanol, and 4% PEG.
BUFFER 7: Reagents are mixed together to form a 10X stock solution of buffer 6 (400 mM Tris-Cl [pH 8.0], 120 mM MgCl2, 50 mM DTT, 10 mM spermidine and 0.02% triton® X-100). Prior to initiation ofthe polymerase reaction PEG, methanol and LiCl is added and the buffer is diluted such that the final reaction conditions for buffer 6 consisted of : 40mM tris (pH 8.0), 12 mM MgCl2, 5 mM DTT, 1 mM spermidine, 0.002% triton® X-100, 10% methanol, 4% PEG, and 1 mM LiCl.
Screening of Modified nucleotide triphosphates with Mutant T7 RNA Polymerase
Modified nucleotide triphosphates were tested in buffers 1 through 6 at two different temperatures (25 and 37°C). Buffers 1-6 tested at 25°C were designated conditions 1-6 and buffers 1-6 tested at 37°C were designated conditions 7-12 (Table 47). In each condition, Y639F mutant T7 polymerase (Sousa and Padilla, supra) (0.3-2 mg/20 ml reaction), NTP's (2 mM each), DNA template (10 pmol), inorganic pyrophosphatase (5U/ml) and α-32P NTP (0.8 mCi/pmol template) were combined and heated at the designated temperatures for 1-2 hours. The radiolabeled NTP used was different from the modified triphosphate being testing. The samples were resolved by polyacrylamide gel elecfrophoresis. Using a Phosphorhnager (Molecular Dynamics, Sunnyvale, CA), the amount of full-length transcript was quantified and compared with an all-RNA confrol reaction. The data is presented in Table 48; results in each reaction are expressed as a percent compared to the all-ribonucleotide triphosphate (rNTP) control. The confrol was run with the mutant T7 polymerase using commercially available polymerase buffer (Boehringer Mannheim, Indianapolis, LN).
Incoφoration of Modified NTP's using Wild-type T7 RNA polymerase
Bacteriophage T7 RNA polymerase was purchased from Boehringer Mannheim at 0.4 U/μL concentration. Applicant used the commercial buffer supplied with the enzyme and 0.2 μCi alpha-32P NTP in a 50 μL reaction with nucleotides triphosphates at 2 mM each. The template was a double-stranded PCR fragment, which was used in previous screens. Reactions were carried out at 37°C for 1 hour. Ten μL ofthe sample was run on a 7.5%o analytical PAGE and bands were quantitated using a Phosphorhnager. Results are calculated as a comparison to an "all ribo" control (non-modified nucleotide triphosphates) and the results are in Table 49. Incoφoration of Multiple Modified nucleotide triphosphates Into Oligonucleotides Combinations of modified nucleotide triphosphates were tested with the transcription protocol described above, to determine the rates of incoφoration of two or more of these triphosphates. Incoφoration of 2 '-Deoxy-2 '-(L-histidine) amino uridine (2'- his-NH2-UTP) was tested with unmodified cytidine nucleotide triphosphates, rATP and rGTP in reaction condition number 9. The data is presented as a percentage of incoφoration of modified NTP's compared to the all rNTP control and is shown in Table 50a.
Two modified cytidines (2'-NH2-CTP or 2'dCTP) were incoφorated along with 2'- his-NH2-UTP with identical efficiencies. 2'-his-NH2-UTP and 2'-NH2-CTP were then tested with various unmodified and modified adenosine triphosphates in the same buffer (Table 50b). The best modified adenosine triphosphate for incoφoration with both 2'-his- NH2-UTP and 2'-NH2-CTP was 2'-NH2-DAPTP.
Optimization of Reaction conditions for Incoφoration of Modified Nucleotide Triphosphate
The combination of 2'-his-NH2-UTP, 2'-NH2-CTP, 2'-NH2-DAP, and rGTP was tested in several reaction conditions (Table 51) using the incoφoration protocol described above. The results demonstrate that ofthe buffer conditions tested, incoφoration of these modified nucleotide triphosphates occur in the presence of both methanol and LiCl.
Selection of Novel Enzymatic nucleic acid molecule Motifs using 2'-deoxy-2'amino Modified GTP and CTP
For selection of new enzymatic nucleic acid molecule motifs, pools of enzymatic nucleic acid molecules were designed to have two substrate binding arms (5 and 16 nucleotides long) and a random region in the middle. The subsfrate has a biotin on the 5' end, 5 nucleotides complementary to the short binding arm ofthe pool, an unpaired G (the desired cleavage site), and 16 nucleotides complementary to the long binding arm ofthe pool. The subsfrate was bound to column resin through an avidin-biotin complex. The general process for selection is shown in Figure 11. The protocols described below represent one possible method that may be utilized for selection of enzymatic nucleic acid molecules and are given as a non-limiting example of enzymatic nucleic acid molecule selection with combinatorial libraries.
Construction of Libraries: The oligonucleotides listed below were synthesized by Operon Technologies
(Alameda, CA). Templates were gel purified and then run through a Sep-Pak™ cartridge
(Waters, Millford, MA) using the manufacturers protocol. Primers (MST3, MST7c,
MST3del) were used without purification.
Primers: MST3 (30 mer): 5'- CAC TTA GCA TTA ACC CTC ACT AAA GGC CGT-3'
MST7c (33 mer): 5'-TAA TAC GAC TCA CTA TAG GAA AGG TGT GCA ACC-3'
MST3del (18 mer): 5'-ACC CTC ACT AAA GGC CGT-3'
Templates:
MSN60c (93 mer): 5'-ACC CTC ACT AAA GGC CGT (N)60 GGT TGC ACA CCT TTG-3'
MSN40c (73 mer): 5'-ACC CTC ACT AAA GGC CGT (N)40 GGT TGC ACA CCT
TTG-3'
MSN20c (53 mer): 5'-ACC CTC ACT AAA GGC CGT (N)20 GGT TGC ACA CCT
TTG-3' N60 library was constructed using MSN60c as a template and MST3/MST7c as primers. N40 and N20 libraries were constructed using MSN40c (or MSN20c) as template and MST3del/MST7c as primers.
Single-stranded templates were converted into double-sfranded DNA by the following protocol: 5 nmol template, 10 nmol each primer, in 10 ml reaction volume using standard PCR buffer, dNTP's, and taq DNA polymerase (all reagents from Boerhinger Mannheim). Synthesis cycle conditions were 94°C, 4 minutes; (94°C, 1 minute; 42°C, 1 minute; 72°C, 2 minutes) x 4; 72°C, 10 minutes. Products were checked on agarose gel to confirm the length of each fragment (N60=123 bp, N40=91 bp, N20=71 bp) and then were phenol/chloroform extracted and ethanol precipitated. The concenfration ofthe double-sfranded product was 25 μM.
Transcription ofthe initial pools was performed in a 1 ml volume comprising: 500 pmol double-sfranded template (3 x IO14 molecules), 40 mM tris-HCl (pH 8.0), 12 mM MgCl2, 1 mM spermidine, 5 mM DTT, 0.002% triton X-100, 1 mM LiCl, 4% PEG 8000, 10% methanol, 2 mM ATP (Pharmacia), 2 mM GTP (Pharmacia), 2 mM 2'-deoxy-2'- amino-CTP (USB), 2 mM 2' -deoxy-2 '-amino-UTP (USB), 5 U/ml inorganic pyrophosphatase (Sigma), 5 U/μl T7 RNA polymerase (USB; Y639F mutant was used in some cases at 0.1 mg/ml (Sousa and Padilla, supra)), 37°C, 2 hours. Transcribed libraries were purified by denaturing PAGE (N60=106 ntds, N40=74, N20=54) and the resulting product was desalted using Sep-Pak™ columns and then ethanol precipitated.
Initial column-Selection: The following biotinylated substrate was synthesized using standard protocols
(Usman et al, 1987 J. Am. Chem. Soc, 109, 7845; Scaringe et al, 1990 Nucleic Acids Res., 18, 5433; and Wincott et al, 1995 Nucleic Acids Res. 23, 2677-2684):
5'-biotin-C18 spacer-GCC GUG GGU UGC ACA CCU UUC C-C18 spacer-thiol- modifier C6 S-S-inverted abasic-3' Subsfrate was purified by denaturing PAGE and ethanol precipitated. 10 nmol of subsfrate was linked to a NeutrAvidin™ column using the following protocol: 400 μl UlfraLink Immobilized NeutrAvidin™ slurry (200 μl beads, Pierce, Rockford, LL) were loaded into a polystyrene column (Pierce). The column was washed twice with 1 ml of binding buffer (20 mM NaPO4 (pH 7.5), 150 mM NaCl) and then capped off (i.e., a cap was put on the bottom ofthe column to stop the flow). 200 μl ofthe subsfrate suspended in binding buffer was applied and allowed to incubate at room temperature for 30 minutes
with occasional vortexing to ensure even linking and distribution ofthe solution to the resin. After the incubation, the cap was removed and the column was washed with 1 ml binding buffer followed by 1 ml column buffer (50 mM tris-HCL (pH 8.5), 100 mM NaCl, 50 mM KC1). The column was then ready for use and capped off. 1 nmol ofthe initial pool RNA was loaded on the column in a volume of 200 μl column buffer. It was allowed to bind the substrate by incubating for 30 minutes at room temperature with occasional vortexing. After the incubation, the cap was removed and the column was washed twice with 1 ml column buffer and capped off. 200 μl of elution buffer (50 mM tris-HCl (pH 8.5), 100 mM NaCl, 50 mM KC1, 25 mM MgCl2) was applied to the column followed by 30 minute incubation at room temperature with occasional vortexing. The cap was removed and four 200 μl fractions were collected using elution buffer.
Second column (counter selection):
A diagram for events in the second column is generally shown in Figure 12 and subsfrate oligonucleotide used is shown below:
5'-GGU UGC ACA CCU UUC C-C18 spacer-biotin-inverted abasic-3' This column subsfrate was linked to UltraLink NeutrAvidin™ resin as previously described (40 pmol) which was washed twice with elution buffer. The eluent from the first column purification was then run on the second column. The use of this column allowed for binding of RNA that non-specifically diluted from the first column, while RNA that performed a catalytic event and had product bound to it, flowed through the second column. The fractions were ethanol precipitated using glycogen as carrier and rehydrated in sterile water for amplification.
Amplification:
RNA and primer MST3 (10-100 pmol) were denatured at 90°C for 3 minutes in water and then snap-cooled on ice for one minute. The following reagents were added to the tube (final concentrations given): IX PCR buffer (Boerhinger Mannheim), 1 mM dNTP's (for PCR, Boerhinger Mannheim), 2 U/μl RNase-Inhibitor (Boerhinger Mannheim), 10 U/μl Superscript™ LI Reverse Transcriptase (BRL). The reaction was incubated for 1 hour at 42°C, then at 95 °C for 5 minutes in order to destroy the
Superscript™. The following reagents were then added to the tube to increase the volume five-fold for the PCR step (final concentrations/amounts given): MST7c primer (10-100 pmol, same amount as in RT step), IX PCR buffer, taq DNA polymerase (0.025-0.05 U/μl, Boerhinger Mannheim). The reaction was cycled as follows: 94°C, 4minutes; (94°C, 30s; 42-54°C, 30s; 72°C, lminute) x 4-30 cycles; 72°C, 5minutes; 30°C, 30 minutes. Cycle number and annealing temperature were decided on a round by round basis. In cases where heteroduplex was observed, the reaction was diluted five-fold with fresh reagents and allowed to progress through 2 more amplification cycles. Resulting products were analyzed for size on an agarose gel (N60=123 bp, N40=103 bp, N20=83 bp) and then ethanol precipitated.
Transcriptions:
Transcription of amplified products was done using the conditions described above with the following modifications: 10-20% ofthe amplification reaction was used as template, reaction volume was 100-500 μl, and the products sizes varied slightly (N60=106 ntds, N40=86, N20=66). A small amount of 32P-GTP was added to the reactions for quantitation puφoses.
Subsequent rounds: Subsequent rounds of selection used 20 pmols of input RNA and 40 pmol ofthe 22 nucleotide substrate on the column.
Activity of pools:
Pools were assayed for activity under single turnover conditions every three to four rounds. Activity assay conditions were as follows: 50 mM tris-HCl (pH 8.5), 25 mM MgCl2, 100 mM NaCl, 50 mM KC1, trace 32P-labeled substrate, 10 nM RNA pool. 2X pool in buffer and, separately, 2X substrate in buffer were incubated at 90°C for 3 minutes, then at 37°C for 3 minutes. Equal volume 2X subsfrate was then added the 2X pool tube (t=0). Initial assay time points were taken at 4 and 24 hours: 5 μl was removed and quenched in 8 μl cold Stop buffer (96% formamide, 20 mM EDTA, 0.05% bromphenyl blue/xylene cyanol). Samples were heated 90°C, 3 minutes, and loaded on a 20%
sequencing gel. Quantitation was performed using a Molecular Dynamics Phosphorimager and ImageQuaNT™ software. The data is shown in Table 52.
Samples from the pools of oligonucleotide were cloned into vectors and sequenced using standard protocols (Sambrook et al, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press). The enzymatic nucleic acid molecules were transcribed from a representative number of these clones using methods described in this application. Individuals from each pool were tested for RNA cleavage from N60 and N40 by incubating the enzymatic nucleic acid molecules from the clones with 5/16 substrate in 2mM MgC12, pH 7.5, lOmM KCl at 37°C. The data in Table 54 shows that the enzymatic nucleic acid molecules isolated from the pool are individually active.
Kinetic Activity:
Kinetic activity ofthe enzymatic nucleic acid molecule shown in Table 54, was determined by incubating enzymatic nucleic acid molecule (10 nM) with substrate in a cleavage buffer (pH 8.5, 25 mM MgCl2, 100 mM NaCl, 50 mM KCl) at 37°C.
Magnesium Dependence:
Magnesium dependence of round 15 of N20 was tested by varying MgCl2 while other conditions were held constant (50 mM tris [pH 8.0], 100 mM NaCl, 50 mM KCl, single turnover, 10 nM pool). The data is shown in Table 55, which demonstrates increased activity with increased magnesium concentrations.
Selection of Novel Enzymatic nucleic acid molecule Motifs using 2 '-Deoxy-2 '-(N- histidyl) amino UTP. 2 '-Fluoro- ATP, and 2'-deoxy-2'-amino CTP and GTP The method used for selection of novel enzymatic nucleic acid molecule motifs using 2 '-deoxy-2 'amino modified GTP and CTP was repeated using 2'-Deoxy-2'-(N- histidyl) amino UTP, 2'-Fluoro-ATP, and 2 '-deoxy-2 '-amino CTP and GTP. However, rather than causing cleavage on the initial column with MgCl2, the initial random modified-RΝA pool was loaded onto substrate-resin in the following buffer; 5 mM ΝaOAc pH 5.2, 1 M ΝaCl at 4° C. After ample washing, the resin was moved to 22 ° C and the buffer switch 20 mM HEPES pH 7.4, 140 mM KCl, 10 mM ΝaCl, 1 mM CaCl2, 1 mM MgCl2. In one selection of Ν60 oligonucleotides, no divalent cations (MgCl2,
CaCl2) was used. The resin was incubated for 10 minutes to allow reaction and the eluant collected.
The enzymatic nucleic acid molecule pools were capable of cleaving l-3%> ofthe present subsfrate even in the absence of divalent cations, the background (in the absence of modified pools) was 0.2 - 0.4 %.
Synthesis of 5-substituted 2,-modified nucleosides
When designing monomeric nucleoside triphosphates for selection of therapeutic catalytic RNAs, one has to take into account nuclease stability of such molecules in biological sera. A common approach to increase RNA stability is to replace the sugar 2'- OH group with other groups like 2'-fluoro, 2'-O-methyl or 2'-amino. Fortunately such 2'- modified pyrimidine 5 'triphosphates are shown to be subsfrates for RNA polymerases. (Aurup, H.; Williams, D.M.; Eckstein, F. Biochemistry 1992, 57, 9637; and Padilla, R.; Sousa, R. Nucleic Acids Res. 1999, 27, 1561.) On the other hand it was shown that variety of substituents at pyrimidine 5-position is well tolerated by T7 RNA polymerase (Tarasow, T.M.; Eaton, B.E. Biopolymers 1998, 48, 29), most likely because the natural hydrogen-bonding pattern of these nucleotides is preserved. We have chosen 2'-fluoro and 2'-O-methyl pyrimidine nucleosides as starting materials for attachment of different functionalities to the 5-position ofthe base. Both rigid (alkynyl) and flexible (alkyl) spacers are used. The choice of imidazole, amino and carboxylate pendant groups is based on their ability to act as general acids, general bases, nucleophiles and metal ligands, all of which can improve the catalytic effectiveness of selected nucleic acids. Figures 21 - 24 relate to the synthesis of these compounds.
2'-O-methyluridine was 3',5'-bis-acetylated using acetic anhydride in pyridine and then converted to its 5-iodo derivative la using ceric ammonium nitrate reagent
(Asakura, J.; Robins, M.J. J. Org. Chem. 1990, 55, 4928) (Scheme 1). Both reactions proceeded in a quantitative yield and no chromatographic purifications were needed. Coupling between 1 and N-trifluoroacetyl propargylamine using copper(I) iodide and tefrakis(triphenylphosphine)palladium(0) catalyst as described by Hobbs (Hobbs, F.W., Jr. J. Org. Chem. 1989, 54, 3420) yielded 2a in 89% yield. Selective O-deacylation with aqueous ΝaOH afforded 3a which was phosphorylated with POClβ/triethylphosphate
(TEP) in the presence of l,8-bis(dimethylamino)naphthalene (Proton-Sponge) (Method A) (Kovacz, T; Otvδs, L. Tetrahedron Lett. 1988, 29, 4525). The intermediate nucleoside phosphorodichloridate was condensed in situ with tri-n-butylammonium pyrophosphate. At the end, the N-TFA group was removed with concentrated ammonia. 5'- Triphosphate was purified on Sephadex® DEAE A-25 ion exchange column using a linear gradient of 0.1-0.8M triethylammonium bicarbonate (TEAB) for elution. Traces of contaminating inorganic pyrophosphate are removed using C-18 RP HPLC to afford analytically pure material. Conversion into Νa-salt was achieved by passing the aqueous solution of triphosphate through Dowex 50WX8 ion exchange resin in Νa+ form to afford 4a in 45% yield. When Proton-Sponge was omitted in the first phosphorylation step, yields were reduced to 10-20%. Catalytic hydrogenation of 3a yielded 5-aminopropyl derivative 5a which was phosphorylated under conditions identical to those described for propynyl derivative 3a to afford triphosphate 6a in 50% yield.
For the preparation of imidazole derivatized triphosphates 9a and Ila, we developed an efficient synthesis of N-diphenylcarbamoyl 4-imidazoleacetic acid (ImAADPC): Transient protection of carboxyl group as TMS-ester using TMS- Cl/pyridine followed by DPC-C1 allowed for a clean and quantitative conversion of 4- imidazoleacetic acid (LmAA) to its N-DPC protected derivative.
Complete deacylation of 2a afforded 5-(3-aminopropynyl) derivative 8a which was condensed with 4-imidazoleacetic acid in the presence of 1 -(3- dimethylaminopropyl)-3-ethylcarbodiimide (EDC) to afford 9a in 68% yield. Catalytic hydrogenation of 8a yielded 5-(3-aminopropyl) derivative 10a which was condensed with ImAADPC to yield conjugate Ila in 32% yield. Yields in these couplings were greatly improved when 5' -OH was protected with DMT group (not shown) thus efficientlypreventing undesired 5 '-O-esterification. Both 9a and Ila failed to yield triphosphate products in reaction with POClβ/TEP/Proton-Sponge.
On the contrary, phosphorylation of 3'-O-acetylated derivatives 12a and 13a using 2-chloro-4H-l,3,2-benzodioxaphosphorin-4-one followed by pyrophosphate addition and oxidation (Method B, Scheme 2; Ludwig, J., Eckstein, F., J. Org. Chem. 1989, 54, 631) afforded the desired triphosphates 14a and 15a in 57% yield, respectively.
2 '-Deoxy-2 '-fluoro nucleoside 5 '-triphosphates containing amino- (4b, 6b) and imidazole- (14b, 15b) linked groups were synthesized in a manner analogous to that described for the preparation of 2'-O-methyl nucleoside 5 '-triphosphates (Schemes 1 and 2). Again, only Ludwig-Eckstein's phosphorylation worked for the preparation of 4- imidazoleacetyl derivatized triphosphates.
It is worth noting that when "one-pot-two-steps" phosphorylation reaction (Kovacz, T; Otvδs, L. Tetrahedron Lett. 1988, 29, 4525) of 5b was quenched with 40% aqueous methylamine instead of TEAB or H2O, the γ-amidate 7b was generated as the only detectable product. Similar reaction was reported recently for the preparation ofthe γ-amidate of pppA2'p5'A2'p5'A.12
Carboxylate group was introduced into 5-position of uridine both on the nucleoside level and post-synthetically (Method C) (Scheme 3). 5-Iodo-2'-deoxy-2'-fluorouridine (16) was coupled with methyl acrylate using modified Heck reaction13 to yield 17 in 85% yield. 5'-O-Dimethoxytritylation, followed by in situ 3'-O-acetylation and subsequent detritylation afforded 3 '-protected derivative 18. Phosphorylation using 2-chloro-4H- l,3,2-benzodioxa-phosphorin-4-one followed by pyrophosphate addition and oxidation (Ludwig, J.; Eckstein, F. J. Org. Chem. 1989, 54, 631) afforded the desired triphosphate in 54% yield. On the other hand, 5-(3-aminopropyl)uridine 5 '-triphosphate 6b was coupled with N-hydroxysuccinimide ester of Fmoc-Asp-OFm to afford, after removal of Fmoc and Fm groups with diethylamine, the desired aminoacyl conjugate 20 in 50% yield.
Cytidine derivatives comprising 3-aminopropyl and 3(N-succinyl)aminopropyl groups were synthesized according to Scheme 4. Peracylated 5-(3-aminopropynyl)uracil derivative 2b is reduced using catalytic hydrogenation and then converted in seven steps and 5%ι overall yield into 3'-acetylated cytidine derivative 25. This synthesis was plagued by poor solubility of intermediates and formation ofthe Ν4-cyclized byproduct during ammonia freatment ofthe 4-triazolyl intermediate. Phosphorylation of 25 as described in reference 11 yielded triphosphate 26 and N -cyclized product 27 in 1 : 1 ratio. They were easily separated on Sephadex DEAE A-25 ion exchange column using 0.1-0.8M TEAB gradient. It appears that under basic conditions the free primary amine can displace any remaining intact 4-NΗBz group leading to the cyclized product. This is similar to displacement of 4-triazolyl group by primary amine as mentioned above.
We reasoned that utilization of N4-unprotected cytidine will solve this problem. This lead to an improved synthesis of 26: Iodination of 2'-deoxy-2'-fluorocytidine (28) provided the 5-iodo derivative 29 in 58%> yield. This compound was then smoothly converted into 5-(3-aminopropynyl) derivative 30. Hydrogenation afforded 5-(3- aminopropyl) derivative 31 which was phosphorylated directly with POCI3/ PPi to afford
26 in 37%) yield. Coupling ofthe 5 '-triphosphate 26 with succinic anhydride yielded succinylated derivative 32 in 36% yield.
Synthesis of 5-Imidazoleacetic acid 2'-deoxy-5'-triphosphate uridine
5-dintrophenylimidazoleacetic acid 2'-deoxy uridine nucleoside (80 mg) was dissolved in 5 ml of triethylphosphate while stirring under argon, and the reaction mixture was cooled to 0°C. Phosphorous oxychloride (1.8 eq, 22 ml) was added to the reaction mixture at 0°C, three more aliquots were added over the course of 48 hours at room temperature. The reaction mixture was then diluted with anhydrous MeCΝ (5 ml) and cooled to 0°C, followed by the addition of tributylamine (0.65 ml) and tributylammonium pyrophosphate (4.0 eq, 0.24 g). After 45 minutes, the reaction was quenched with 10 ml aq. methyl amine for four hours. After co-evaporation with MeOH (3x), purified material on DEAE Sephadex followed by RP chromatography to afford 15 mg of triphosphate. Synthesis of 2'-(N-lvsyl)-amino-2'-deoxy-cytidine Triphosphate
2'-(Ν-lysyl)-amino-2'-deoxy cytidine (0.180 g, 0.22 mmol) was dissolved in triethyl phosphate (2.00 ml) under Ar. The solution was cooled to 0 °C in an ice bath.
Phosphorus oxychloride (99.999%, 3 eq., 0.0672 mL) was added to the solution and the reaction was stiπed for two hours at 0 °C. Tributylammonium pyrophosphate (4 eq., 0.400 g) was dissolved in 3.42 mL of acetonitrile and tribuytylamine (0.165 mL). Acetonitrile (1 mL) was added to the monophosphate solution followed by the pyrophosphate solution which was added dropwise. The resulting solution was clear. The reaction was allowed to warm up to room temperature. After stirring for 45 minutes, methylamine (5 mL) was added and the reaction and stiπed at room temperature for 2 hours. A biphasic mixture appeared (little beads at the bottom ofthe flask). TLC (7:1:2 iPrOH:NH4OH:H2O) showed the appearance of triphosphate material. The solution was concentrated, dissolved in water and loaded on a newly prepared DEAE Sephadex A-25 column. The column was washed with a gradient up to 0.6 M TEAB buffer and the product eluted off in fractions
90-95. The fractions were analyzed by ion exchange HPLC. Each fraction showed one triphosphate peak that eluted at -4.000 minutes. The fractions were combined and pumped down from methanol to remove buffer salt to yield 15.7 mg of product. Synthesis of 2'-deoxy-2'-(L-histidine)amino Cytidine Triphosphate 2'-[N-Fmoc, Nimid -dinifrophenyl-histidyl]amino-2' -cytidine (0.310 g, 4.04 mmol) was dissolved in triethyl phosphate (3 ml) under Ar. The solution was cooled to 0 °C. Phosphorus oxychloride (1.8 eq., 0.068 mL) was added to the solution and stored overnight in the freezer. The next morning TLC (10% MeOH in CH2CI2) showed significant starting material, one more equivalent of POCI3 was added. After two hours, TLC still showed starting material. Tributylamine (0.303 mL) and Tributylammonium pyrophosphate (4 eq., 0.734 g) dissolved in 6.3 mL of acetonitrile (added dropwise) were added to the monophosphate solution. The reaction was allowed to warm up to room temperature. After stirring for 15 min, methylamine (10 mL) was added at room temperature and stirring continued for 2 hours. TLC (7:1:2 iPrOH:ΝH4θH:H2θ) showed the appearance of triphosphate material. The solution was concenfrated, dissolved in water and loaded on a DEAE Sephadex A-25 column. The column was washed with a gradient up to 0.6 M TEAB buffer and the product eluted off in fractions 170-179. The fractions were analyzed by ion exchange HPLC. Each fraction showed one triphosphate peak that eluted at ~6.77 minutes. The fractions were combined and pumped down from methanol to remove buffer salt to afford 17 mg of product.
Screening for Novel Enzymatic nucleic acid molecule Motifs Using Modified NTPs (Class I Motif)
Our initial pool contained 3 x IO14 individual sequences of 2'-amino-dCTP/2'- amino-dUTP RNA. We optimized franscription conditions in order to increase the amount of RNA product by inclusion of methanol and lithium chloride. 2'-amino-2'- deoxynucleotides do not interfere with the reverse transcription and amplification steps of selection and confer nuclease resistance. We designed the pool to have two binding arms complementary to the substrate, separated by the random 40 nucleotide region. The 16- mer subsfrate had two domains, 5 and 10 nucleotides long, that bind the pool, separated by an unpaired guanosine. On the 5' end ofthe substrate was a biotin attached by a C18 linker. This enabled us to link the subsfrate to a NeutrAvidin™ resin in a column format.
The desired reaction would be cleavage at the unpaired G upon addition of magnesium cofactor followed by dissociation from the column due to instability ofthe 5 base pair helix. A detailed protocol follows:
Enzymatic nucleic acid molecule Pool Prep: The initial pool DNA was prepared by converting the following template oligonucleotides into double-sfranded DNA by filling in with taq polymerase. (template=5'-ACC CTC ACT AAA GGC CGT (N)40 GGT TGC ACA CCT TTC-3'; primer 1=5'- CAC TTA GCA TTA ACC CTC ACT AAA GGC CGT-3'; primer 2=5'-TAA TAC GAC TCA CTA TAG GAA AGG TGT GCA ACC-3'.) All DNA oligonucleotides were synthesized by Operon technologies. Template oligos were purified by denaturing PAGE and Sep-pak chromatography columns (Waters). RNA subsfrate oligos were using standard solid phase chemistry and purified by denaturing PAGE followed by ethanol precipitation. Substrates for in vitro cleavage assays were 5'- end labeled with gamma-32P-ATP and T4 polynucleotide kinase followed by denaturing PAGE purification and ethanol precipitation. 5 nmole of template, 10 nmole of each primer and 250 U taq polymerase were incubated in a 10 ml volume with IX PCR buffer (10 mM tris-HCl (pH 8.3), 1.5 mM MgCl2, 50 mM KCl) and 0.2 mM each dNTP as follows: 94°C, 4 minutes; (94°C, 1 min; 42°C, 1 min; 72°C, 2 min) through four cycles; and then 72°C, for 10 minutes. The product was analyzed on 2% Separide™ agarose gel for size and then was extracted twice with buffered phenol, then chloroform-isoamyl alcohol, and ethanol precipitated. The initial RNA pool was made by franscription of 500 pmole (3 x IO14 molecules) of this DNA as follows. Template DNA was added to 40 mM tris-HCl (pH 8.0), 12 mM MgCl2, 5 mM dithiothreitol (DTT), 1 mM spermidine, 0.002% triton X-100, 1 mM LiCl, 4% PEG-8000, 10% methanol, 2 mM ATP, 2 mM GTP, 2 mM 2'-amino-dCTP, 2 mM 2'- amino-dUTP, 5 U/ml inorganic pyrophosphatase, and 5 U/μl T7 RNA polymerase at room temperature for a total volume of 1 ml. A separate reaction contained a trace amount of
^7 alpha- P-GTP for detection. Transcriptions were incubated at 37°C for 2 hours followed by addition of equal volume STOP buffer (94% formamide, 20 mM EDTA, 0.05% bromophenol blue). The resulting RNA was purified by 6% denaturing PAGE gel, Sep- pak™ chromatography, and ethanol precipitated.
INITIAL SELECTION: 2 nmole of 16 mer 5 '-biotinylated substrate (5'-biotin-C18 linker-GCC GUG GGU UGC ACA C-3') was linked to 200 μl UltraLink Immobilized
NeutrAvidin™ resin (400 μl slurry, Pierce) in binding buffer (20 mM NaPO4 (pH 7.5), 150 mM NaCl) for 30 minutes at room temperature. The resulting subsfrate column was washed with 2 ml binding buffer followed by 2 ml column buffer (50 mM tris-HCl (pH 8.5), 100 mM NaCl, 50 mM KCl). The flow was capped off and 1000 pmole of initial pool RNA in 200 μl column buffer was added to the column and incubated 30 minutes at room temperature. The column was uncapped and washed with 2 ml column buffer, then capped off. 200 μl elution buffer (=column buffer + 25 mM MgCl2) was added to the column and allowed to incubate 30 minutes at room temperature. The column was uncapped and eluent collected followed by three 200 μl elution buffer washes. The eluent washes were ethanol precipitated using glycogen as carrier and rehydrated in 50 μl sterile H O. The eluted RNA was amplified by standard reverse transcription/PCR amplification techniques. 5-31 μl RNA was incubated with 20 pmol of primer 1 in 14 μl volume 90° for 3 min then placed on ice for 1 minute. The following reagent were added (final concentrations noted): IX PCR buffer, 1 mM each dNTP, 2 U/μl RNase Inhibitor, 10 U/μl Superscript™ IL reverse transcriptase. The reaction was incubated 42° for 1 hour followed by 95° for 5 min in order to inactivate the reverse transcriptase. The volume was then increased to 100 μl by adding water and reagents for PCR: IX PCR buffer, 20 pmol primer 2, and 2.5 U taq DNA polymerase. The reaction was cycled in a Hybaid thermocycler: 94°, 4 min; (94°C, 30 sec; 54°C, 30 sec; 72°C, 1 min) X 25; 72°C, 5 min. Products were analyzed on agarose gel for size and ethanol precipitated. One-third to one- fifth ofthe PCR DNA was used to transcribe the next generation, in 100 μl volume, as described above. Subsequent rounds used 20 pmol RNA for the column with 40 pmol substrate.
TWO COLUMN SELECTION: At generation 8 (G8), the column selection was changed to the two column format. 200 pmoles of 22 mer 5 '-biotinylated subsfrate (5'- biotin-C18 linker-GCC GUG GGU UGC ACA CCU UUC C-C18 linker-thiol modifier C6 S-S-inverted abasic-3') was used in the selection column as described above. Elution was in 200 μl elution buffer followed by a 1 ml elution buffer wash. The 1200 μl eluent was passed through a product trap column by gravity. The product trap column was prepared as follows: 200 pmol 16 mer 5 '-biotinylated "product" (5'-GGU UGC ACA CCU UUC C-C18 linker-biotin-3') was linked to the column as described above and the column was equilibrated in elution buffer. Eluent from the product column was precipitated as
previously described. The products were amplified as above only with 2.5-fold more volume and 100 pmol each primer. 100 μl ofthe PCR reaction was used to do a cycle course; the remaining fraction was amplified the minimal number of cycles needed for product. After 3 rounds (Gl 1), there was visible activity in a single turnover cleavage assay. By generation 13, 45% ofthe substrate was cleaved at 4 hours; kobS ofthe pool was 0.037 min"1 in 25 mM MgCl2. We subcloned and sequenced generation 13; the pool was still very diverse. Since our goal was a enzymatic nucleic acid molecule that would work in a physiological environment, we decided to change selection pressure rather than exhaustively catalog G13. Reselection of the N40 pool was started from Gl 2 DNA. Part of the Gl 2 DNA was subjected to hypermutagenic PCR (Vartanian et al, 1996, Nucleic Acids Research 24, 2627-2631) to introduce a 10% per position mutation frequency and was designated N40H. At round 19, part ofthe DNA was hypermutagenized again, giving N40M and N40HM (a total of 4 parallel pools). The column subsfrates remained the same; buffers were changed and temperature of binding and elution was raised to 37°C. Column buffer was replaced by physiological buffer (50 mM tris-HCl (pH 7.5), 140 mM KCl, 10 mM NaCl) and elution buffer was replaced by 1 mM Mg buffer (physiological buffer + 1 mM MgCl2). Amount of time allowed for the pool to bind the column was eventually reduced to 10 min and elution time was gradually reduced from 30 min to 20 sec. Between rounds 18 and 23, k^s for the N40 pool stayed relatively constant at 0.035-0.04 min"1. Generation 22 from each ofthe 4 pools was cloned and sequenced.
CLONING AND SEQUENCING: Generations 13 and 22 were cloned using Novagen's Perfectly Blunt™ Cloning kit (pT7Blue-3 vector) following the kit protocol. Clones were screened for insert by PCR amplification using vector-specific primers. Positive clones were sequenced using ABI Prism 7700 sequence detection system and vector-specific primer. Sequences were aligned using Mac Vector software; two- dimensional folding was performed using Mulfold software ( Zuker,, 1989, Science 244, 48-52; Jaeger et al, 1989, Biochemistry 86, 7706-7710; Jaeger et al, 1989, R. F. Doolittle ed., Methods in Enzymology, 183, 281-306). Individual clone transcription units were constructed by PCR amplification with 50 pmol each primer 1 and primer 2 in IX PCR buffer, 0.2 mM each dNTP, and 2.5 U of taq polymerase in 100 μl volume cycled as follows: 94°C, 4 min; (94°C, 30 sec; 54°C, 30 sec; 72°C, 1 min) X 20; 72°C, 5 min.
Transcription units were ethanol precipitated, rehydrated in 30 μl H2O, and 10 μl was transcribed in 100 μl volume and purified as previously described.
Thirty-six clones from each pool were sequenced and were found to be variations of the same consensus motif. Unique clones were assayed for activity in 1 mM MgCl2 and physiological conditions; nine clones represented the consensus sequence and were used in subsequent experiments. There were no mutations that significantly increased activity; most ofthe mutations were in regions believed to be duplex, based on the proposed secondary structure. In order to make the motif shorter, we deleted the 3 '-terminal 25 nucleotides necessary to bind the primer for amplification. The measured rates ofthe full length and truncated molecules were both 0.04 min"1 ; thus we were able reduce the size of the motif from 86 to 61 nucleotides. The molecule was shortened even further by truncating base pairs in the stem loop structures as well as the substrate recognition arms to yield a 48 nucleotide molecule. In addition, many ofthe ribonucleotides were replaced with 2-O-methyl modified nucleotides to stabilize the molecule. An example ofthe new motif is given in Figure 13. Those of ordinary skill in the art will recognize that the molecule is not limited to the chemical modifications shown in the figure and that it represents only one possible chemically modified molecule.
Kinetic Analysis: Single turnover kinetics were performed with trace amounts of 5 '-32P-labeled subsfrate and 10-1000 nM pool of enzymatic nucleic acid molecule. 2X substrate in IX buffer and 2X pool enzymatic nucleic acid molecule in IX buffer were incubated separately 90° for 3 min followed by equilibration to 37° for 3 min. Equal volume of 2X subsfrate was added to pool/enzymatic nucleic acid molecule at to and the reaction was incubated at 37°C. Time points were quenched in 1.2 vol STOP buffer on ice. Samples were heated to 90°C for 3 min prior to separation on 15% sequencing gels. Gels were imaged using a Phosphorimager and quantitated using ImageQuant™ software (Molecular Dynamics). Curves were fit to double-exponential decay in most cases, although some of the curves required linear fits. STABILITY: Serum stability assays were performed as previously described
(Beigelman et al., 1995, J. Biol. Chem. 270, 25702-25708). 1 μg of 5'-32P-labeled
synthetic enzymatic nucleic acid molecule was added to 13 μl cold and assayed for decay in human serum. Gels and quantitation were as described in kinetics section.
SUBSTRATE REQUIREMENTS: Table 60 outlines the substrate requirements for Class I motif. Subsfrates maintained Watson-Crick or wobble base pairing with mutant Class I constructs. Activity in single turnover kinetic assay is shown relative to wild type Class I and 22 mer substrate (50 mM Tris-HCL (pH 7.5), 140 mM KCl, 10 mM NaCl, 1 mM MgCl2 , 100 nM ribozyme, 5 nM substrate, 37°C).
RANDOM REGION MUTATION ALIGNMENT: Table 61 outlines the random region alignment of 134 clones from generation 22 (1.x = N40, 2.x = N40M, 3.x = N40H, 4.x = N40HM). The number of copies of each mutant is in parenthesis in the table, deviations from consensus are shown. Mutations that maintain base pair U19:A34 are shown in italic. Activity in single turnover kinetic assay is shown relative to the G22 pool rate (50 mM Tris-HCL pH 7.5, 140 mM KCl, 10 mM NaCl, 1 mM MgCl2 , 100 nM ribozyme, trace subsfrate, 37°C). STEM TRUNCATION AND LOOP REPLACEMENT ANALYSIS: Figure 25 shows a representation of Class I ribozyme stem truncation and loop replacement analysis. The Krej is compared to a 61 mer Class I ribozyme measured as described above. Figure 26 shows examples of Class I ribozymes with truncated stem(s) and/or non-nucleotide linker replaced loop structures.
Inhibition of HCV Using Class I (Amberzyme) Motif
During HCV infection, viral RNA is present as a potential target for enzymatic nucleic acid molecule cleavage at several processes: uncoating, translation, RNA replication and packaging. Target RNA may be more or less accessible to enzymatic nucleic acid molecule cleavage at any one of these steps. Although the association between the HCV initial ribosome entry site (LRES) and the translation apparatus is mimicked in the HCV 5'UTR luciferase reporter system, these other viral processes are not represented in the OST7 system. The resulting RNA/protein complexes associated with the target viral RNA are also absent. Moreover, these processes may be coupled in an HCV-infected cell which could further impact target RNA accessibility. Therefore, we
tested whether enzymatic nucleic acid molecules designed to cleave the HCV 5'UTR could effect a replicating viral system.
Recently, Lu and Wimmer characterized an HCV-poliovirus chimera in which the poliovirus LRES was replaced by the IRES from HCV (Lu & Wimmer, 1996, Proc. Natl. Acad. Sci. USA. 93, 1412-1417). Poliovirus (PV) is a positive strand RNA virus like HCV, but unlike HCV is non-enveloped and replicates efficiently in cell culture. The HCV-PV chimera expresses a stable, small plaque phenotype relative to wild type PV. The capability ofthe new enzymatic nucleic acid molecule motifs to inhibit HCV RNA intracellularly was tested using a dual reporter system that utilizes both firefly and Renilla luciferase (Figure 14). A number of enzymatic nucleic acid molecules having the new class I motif (Amberzyme) were designed and tested (Table 56). The Amberzyme ribozymes were targeted to the 5' HCV UTR region, which when cleaved, would prevent the translation ofthe transcript into luciferase. OST-7 cells were plated at 12,500 cells per well in black walled 96-well plates (Packard) in medium DMEM containing 10%> fetal bovine serum, 1%> pen/strep, and 1% L-glutamine and incubated at 37°C overnight. A plasmid containing T7 promoter expressing 5' HCV UTR and firefly luciferase (T7C1-341 (Wang et al, 1993, J. of Virol. 67, 3338-3344)) was mixed with a pRLSV40 Renilla confrol plasmid (Promega Coφoration) followed by enzymatic nucleic acid molecule, and cationic lipid to make a 5X concentration ofthe reagents (T7C 1-341 (4 μg/ml), pRLSV40 renilla luciferase confrol (6 μg/ml), enzymatic nucleic acid molecule (250 nM), fransfection reagent (28.5 μg/ml). o
The complex mixture was incubated at 37 C for 20 minutes. The media was removed from the cells and 120 μl of Opti-mem media was added to the well followed by 30 μl ofthe 5X complex mixture. 150 μl of Opti-mem was added to the wells holding the untreated cells. The complex mixture was incubated on OST-7 cells for 4 hours, lysed with passive lysis buffer (Promega Coφoration) and luminescent signals were quantified using the Dual Luciferase Assay Kit using the manufacturer's protocol (Promega Coφoration). The data shown in Figure 15 is a dose curve of enzymatic nucleic acid molecule targeting site 146 ofthe HCV RNA and is presented as a ratio between the firefly and Renilla luciferase fluorescence. The enzymatic nucleic acid molecule was able to reduce the quantity of HCV RNA at all enzymatic nucleic acid molecule concentrations
yielding an IC50 of approximately 5 nM. Other sites were also efficacious (Figure 16), in particular enzymatic nucleic acid molecules targeting sites 133, 209, and 273 were also able to reduce HCV RNA compared to the iπelevant (LRR) confrols.
Cleavage of Subsfrates Using Completely Modified class I (Amberzyme) enzymatic nucleic acid molecule
The ability of an enzymatic nucleic acid, which is modified at every 2' position to cleave a target RNA was tested to determine if any ribonucleotide positions are necessary in the Amberzyme motif. Enzymatic nucleic acid molecules were constructed with 2'-O- methyl, and 2'-amino (NH2) nucleotides and included no ribonucleotides (Table 56; gene name: no ribo) and kinetic analysis was performed as described in example 13. 100 nM enzymatic nucleic acid was mixed with trace amounts of substrate in the presence of 1 mM MgCl2 at physiological conditions (37°C). The Amberzyme with no ribonucleotide present in it has a Kreι of 0.13 compared to the enzymatic nucleic acid with a few ribonucleotides present in the molecule shown in Table 56 (ribo). This shows that Amberzyme enzymatic nucleic acid molecule may not require the presence of 2' -OH groups within the molecule for activity.
Substrate Recognition Rules for Class II (zinzyme) enzymatic nucleic acid molecules Class LI (zinzyme) ribozymes were tested for their ability to cleave base-paired subsfrates with all sixteen possible combinations of bases immediately 5' and 3' proximal to the bulged cleavage site G. Ribozymes were identical in all remaining positions of their 7 base pair binding arms. Activity was assessed at two and twenty- four hour time points under standard reaction conditions [20 mM HEPES pH 7.4, 140 mM KCl, 10 mM NaCl, 1 mM MgCl2, 1 mM CaCl2 - 37° C]. Figure 19 shows the results of this study. Base paired subsfrate UGG (not shown in the figure) cleaved as poorly as CGG shown in the figure. The figure shows the cleavage site substrate triplet in the 5'- 3' direction and 2 and 24 hour time points are shown top to bottom respectively. The results indicate the cleavage site triplet is most active with a 5'- Y-G-H -3' (where Y is C or U and H is A, C or U with cleavage between G and H); however, activity is detected particularly with the 24 hour time point for most paired subsfrates. All positions outside ofthe cleavage triplet were found to tolerate any base pairings (data not shown). All possible mispairs immediately 5' and 3' proximal to the bulged cleavage site G were tested to a class II ribozyme designed to cleave a 5'-C-G-C -3'. It was observed the 5' and 3' proximal sites are as active with G:U wobble pairs, in addition, the 5' proximal site will tolerate a mismatch with only a slight reduction in activity [data not shown].
Screening for Novel Enzymatic nucleic acid molecule Motifs (Class LI Motifs)
The selections were initiated with pools of > IO14 modified RNA's ofthe following sequence: S'-GGGAGGAGGAAGUGCCU (N)35 UGCCGCGCUCGCUCCCAGUCC-3'. The RNA was enzymatically generated using the mutant T7 Y639F RNA polymerase prepared by Rui Souza. The following modified NTP's were incoφorated: 2 '-deoxy-2 '-fluoro- adenine triphosphate, 2' -deoxy-2 '-fluoro-uridine triphosphate or 2'-deoxy-2'-fluoro-5-[(N- imidazole-4acetyl)propyl amine] uridine triphosphate, and 2' -deoxy-2 '-amino-cytidine triphosphate; natural guanidine triphosphate was used in all selections so that alpha - P- GTP could be used to label pool RNA's. RNA pools were purified by denaturing gel elecfrophoresus 8% polyacrilamide 7 M Urea. The following target RNA (resin A) was synthesized and coupled to Iodoacetyl
Ulfralink™ resin (Pierce) by the supplier's proceedure:5' -b-L- GGACUGGGAGCGAGCGCGGCGCAGGCACU GAAG-L-S-B-3'; where b is biotin (Glenn
Research cat# 10-1953-nn), L is polyethylene glycol spacer (Glenn Research cat# 10-1918- nn), S is thiol-modifier C6 S-S (Glenn Research cat# 10-1936-nn), B is a standard inverted deoxy abasic.
RNA pools were added to 100 μl of 5 uM Resin A in the buffer A (20 mM HEPES pH 7.4, 140 mM KCL, 10 mM NaCl) and incubated at 22°C for 5 minutes. The temperature was then raised to 37°C for 10 minutes. The resin was washed with 5 ml buffer A. Reaction was triggered by the addition of buffer B(20 mM HEPES pH 7.4, 140 mM KCL, 10 mM NaCl, 1 mM MgCl2, 1 mM CaCl2). Incubation proceeded for 20 minutes in the first generation and was reduced progressively to 1 minute in the final generations; with 13 total generations. The reaction eluent was collected in 5 M NaCl to give a final concenfration of 2 M NaCl. To this was added 100 μl of 50% slurry Ultralink NeufraAvidin™ (Pierce). Binding of cleaved biotin product to the avidin resin was allowed by 20 minute incubation at 22° C. The resin was subsequently washed with 5 ml of 20 mM HEPES pH 7.4, 2 M NaCl. Desired RNA's were removed by a 1.2 ml denaturing wash IM NaCl, 10 M Urea at 94° C over 10 minutes. RNA's were double precipitated in 0.3 M sodium acetate to remove CI" ions inhibitory to reverse franscription. Standard protocols of reverse transcription and PCR amplification were performed. RNA's were again transcribed with the modified NTP's described above. After 13 generations cloning and sequencing provided 14 sequences which were able to cleave the target subsfrate. Six sequences were characterized to determine secondary structure and kinetic cleavage rates. The structures and kinetic data are given in Figure 17. The sequences of eight other enzymatic nucleic acid molecule sequences are given in Table 57. The size, sequence, and chemical compositions of these molecules can be modified as described below or using other techniques well known in the art.
Nucleic Acid Catalyst Engineering
Sequence, chemical and structural variants of Class I and Class LI enzymatic nucleic acid molecule can be engineered and re-engineered using the techniques shown in this application and known in the art. For example, the size of class I and class II enzymatic nucleic acid molecules can, be reduced or increased using the techniques known in the art (Zaug et al, 1986, Nature, 324, 429; Ruffher et al, 1990, Biochem., 29, 10695; Beaudry et
al, 1990, Biochem., 29, 6534; McCall et al, 1992, Proc. Natl. Acad. Sci., USA., 89, 5710; Long et α/., 199A, supra; Hendry et al, 1994, BBA 1219, 405; Benseler et α/., 1993, JACS, 115, 8483; Thompson et al, 1996, Nucl. Acids Res., 24, 4401; Michels et al, 1995, Biochem., 34, 2965; Been et al, 1992, Biochem., 31, 11843; Guo et α/., 1995, EMBO. J, 14, 368; Pan et al, 1994, Biochem., 33, 9561; Cech, 1992, Cwrr. Op. Struc. Bio., 2, 605; Sugiyama et al, 1996, FEBS Lett., 392, 215; Beigelman et α/., 199A, Bioorg. Med. Chem., A, 1715; Santoro et α/., 1997, PN4S 94, 4262; all are incoφorated in their totality by reference herein), to the extent that the overall catalytic activity ofthe ribozyme is not significantly decreased. Further rounds of in vitro selection strategies described herein and variations thereof can be readily used by a person skilled in the art to evolve additional nucleic acid catalysts and such new catalysts are within the scope ofthe instant invention.
Example 16: Activity of Class TJ (zinzyme) nucleic acid catalysts to inhibit HER2 gene expression
Applicant has designed, synthesized and tested several class LI (zinzyme) ribozymes targeted against HER2 RNA (see, for example, Tables 58, 59, and 62) in cell proliferation
RNA reduction assays.
Proliferation assay: The model proliferation assay used in the study can require a cell-plating density of
2000-10000 cells/well in 96-well plates and at least 2 cell doublings over a 5-day freatment period. Cells used in proliferation studies were either human breast or ovarian cancer cells
(SKBR-3 and SKOV-3 cells respectively). To calculate cell density for proliferation assays, the FLPS (fluoro-imaging processing system) method well known in the art was used. This method allows for cell density measurements after nucleic acids are stained with CyQuant® dye, and has the advantage of accurately measuring cell densities over a very wide range 1,000-100,000 cells/well in 96-well format.
Ribozymes (50-200 nM) were delivered in the presence of cationic lipid at 2.0-5.0 μg/mL and inhibition of proliferation was determined on day 5 post-treatment. Two full ribozyme screens were completed resulting in the selection of 14 ribozymes. Class LI
(zinzyme) ribozymes against sites, 314 (RPI No. 18653), 443 (RPI No. 18680), 597 (RPI
No. 18697), 659 (RPI No. 18682), 878 (RPI Nos. 18683 and 18654), 881 (RPI Nos. 18684 and 18685) 934 (RPI No. 18651), 972 (RPI No. 18656, 19292, 19727, 19728, and 19293), 1292 (RPI No. 18726), 1541 (RPI No. 18687), 2116 (RPI No. 18729), 2932 (RPI No. 18678), 2540 (RPI No. 18715), and 3504 (RPI No. 18710) caused inhibition of proliferation ranging from 25-80% as compared to a scrambled control ribozyme. An example of results from a cell culture assay is shown in Figure 20. Referring to Figure 20, Class II ribozymes targeted against HER2 RNA are shown to cause significant inhibition of proliferation of cells. This shows that ribozymes, for instance the Class π (zinzyme) ribozymes are capable of inhibiting HER2 gene expression in mammalian cells.
RNA assay:
RNA was harvested 24 hours post-treatment using the Qiagen RNeasy® 96 procedure. Real time RT-PCR (TaqMan® assay) was performed on purified RNA samples using separate primer/probe sets specific for either target HER2 RNA or confrol actin RNA (to normalize for differences due to cell plating or sample recovery). Results are shown as the average of triplicate determinations of HER2 to actin RNA levels post- treatment. Figure 30 shows class II ribozyme (zinzyme) mediated reduction in HER2 RNA targeting site 972 vs a scrambled attenuated control.
Dose response assays:
Active ribozyme was mixed with binding arm-attenuated control (BAC) ribozyme to a final oligonucleotide concenfration of either 100, 200 or 400 nM and delivered to cells in the presence of cationic lipid at 5.0 μg/mL. Mixing active and BAC in this manner maintains the lipid to ribozyme charge ratio throughout the dose response curve. HER2 RNA reduction was measured 24 hours post-freatment and inhibition of proliferation was determined on day 5 post-freatment. The dose response antiproliferation results are summarized in Figure 31 and the dose-dependent reduction of HER2 RNA results are summarized in Figure 32. Figure 33 shows a combined dose response plot of both anti- proliferation and RNA reduction data for a class LI ribozyme targeting site 972 of HER2 RNA (RPI 19293).
Example 17: Reduction of ribose residues in Class LI (zinzyme) nucleic acid catalysts
Class LI (zinzyme) nucleic acid catalysts were tested for their activity as a function ribonucleotide content. A Zinzyme having no ribonucleotide residue (ie., no 2'-OH group at the 2' position ofthe nucleotide sugar) against the K-Ras site 521 was designed. This molecules were tested utilizing the chemistry shown in Figure 27a. The in vitro catalytic activity zinzyme construct was not significantly effected (the cleavage rate reduced only 10 fold).
The Kras zinzyme shown in Figure 27a was tested in physiological buffer with the divalent concentrations as indicated in the legend (high NaCl is an altered monovalent condition shown) of Figure 28. The 1 mM Ca** condition yielded a rate of 0.005 min"1 while the 1 mM Mg""" condition yielded a rate of 0.002 min"1. The ribose containing wild type yields a rate of 0.05 min while subsfrate in the absence of zinzyme demonstrates less than 2% degradation at the longest time point under reaction conditions shown. This illustrates a well-behaved cleavage reaction done by a non-ribose containing catalyst with only a 10-fold reduced cleavage as compared to ribonucleotide-containing zinzyme and vastly above non-catalyzed degradation.
A more detailed investigation into the role of ribose positions in the Class π (zinzyme) motif was carried out in the context ofthe HER2 site 972 (Applicant has further designed a fully modified Zinzyme as shown in Figure 27b targeting the HER2 RNA site 972). Figure 29 is a diagram ofthe alternate formats tested and their relative rates of catalysis. The effect of substitution of ribose G for the 2'-O-methyl C-2'-O-methyl A in the loop of Zinzyme (see Figure 34) was insignificant when assayed with the Kras target but showed a modest rate enhancement in the HER2 assays. The activity of all Zinzyme motifs, including the fully stabilized "0 ribose" (RPI 19727) are well above background noise level degradation. Zinzyme with only two ribose positions (RPI 19293) are sufficient to restore "wild-type" activity. Motifs containing 3 (RPI 19729), 4 (RPI 19730) or 5 ribose (RPI 19731) positions demonsfrated a greater extent of cleavage and profiles almost identical to the 2 ribose motif. Applicant has thus demonsfrated that a Zinzyme with no ribonucleotides present at any position can catalyze efficient RNA cleavage activity. Thus, Zinzyme enzymatic nucleic acid molecules do not require the presence of 2'-ΟH group within the molecule for catalytic activity.
Example 18: Activity of reduced ribose containing Class II (zinzyme) nucleic acid catalysts to inhibit HER2 gene expression
A cell proliferation assay for testing reduced ribo class LI (zinzyme) nucleic acid catalysts (50-400 nM) targeting HER2 site 972 was performed as described in example 19. The results of this study are summarized in Figure 35. These results indicate significant inhibition of HER2 gene expression using stabilized Class II (zinzyme) motifs, including two ribo (RPI 19293), one ribo (RPI 19728), and non-ribo (RPI 19727) containing nucleic acid catalysts.
Applications
The use of NTP's described in this invention have several research and commercial applications. These modified nucleotide triphosphates can be used for in vitro selection (evolution) of oligonucleotides with novel functions. Examples of in vitro selection protocols are incoφorated herein by reference (Joyce, 1989, Gene, 82, 83-87; Beaudry et al, 1992, Science 257, 635-641; Joyce, 1992, Scientific American 267, 90-97; Breaker et al, 1994, TIBTECH 12, 268; Bartel et α/.,1993, Science 261:1411-1418; Szostak, 1993, TIBS 17, 89-93; Kumar et al, 1995, FASEB , 9, 1183; Breaker, 1996, Curr. Op. Biotech., 7, 442).
Additionally, these modified nucleotide triphosphates can be employed to generate modified oligonucleotide combinatorial chemistry libraries. Several references for this technology exist (Brenner et al, 1992, PNAS 89, 5381-5383, Eaton, 1997, Curr. Opin. Chem. Biol. 1, 10-16), which are all incoφorated herein by reference.
Diagnostic uses Enzymatic nucleic acid molecules of this invention may be used as diagnostic tools to examine genetic drift and mutations within diseased cells or to detect the presence of specific RNA in a cell. The close relationship between enzymatic nucleic acid molecule activity and the structure ofthe target RNA allows the detection of mutations in any region ofthe molecule which alters the base-pairing and three-dimensional structure ofthe target RNA. By using multiple enzymatic nucleic acid molecules described in this invention, one may map nucleotide changes which are important to RNA structure and function in vitro, as well as in cells and tissues. Cleavage of target RNAs with enzymatic nucleic acid
molecules may be used to inhibit gene expression and define the role (essentially) of specified gene products in the progression of disease. In this manner, other genetic targets may be defined as important mediators ofthe disease. These experiments will lead to better treatment ofthe disease progression by affording the possibility of combinational therapies (e.g., multiple enzymatic nucleic acid molecules targeted to different genes, enzymatic nucleic acid molecules coupled with known small molecule inhibitors, radiation or intermittent freatment with combinations of enzymatic nucleic acid molecules and/or other chemical or biological molecules). Other in vitro uses of enzymatic nucleic acid molecules of this invention are well known in the art, and include detection ofthe presence of mRNAs associated with related conditions. Such RNA is detected by determimng the presence of a cleavage product after treatment with a enzymatic nucleic acid molecule using standard methodology.
In a specific example, enzymatic nucleic acid molecules which can cleave only wild- type or mutant forms ofthe target RNA are used for the assay. The first enzymatic nucleic acid molecule is used to identify wild-type RNA present in the sample and the second enzymatic nucleic acid molecule will be used to identify mutant RNA in the sample. As reaction confrols, synthetic subsfrates of both wild-type and mutant RNA will be cleaved by both enzymatic nucleic acid molecules to demonstrate the relative enzymatic nucleic acid molecule efficiencies in the reactions and the absence of cleavage ofthe "non- targeted" RNA species. The cleavage products from the synthetic subsfrates will also serve to generate size markers for the analysis of wild type and mutant RNAs in the sample population. Thus each analysis can involve two enzymatic nucleic acid molecules, two subsfrates and one unknown sample which can be combined into six reactions. The presence of cleavage products can be determined using an RNAse protection assay so that full-length and cleavage fragments of each RNA can be analyzed in one lane of a polyacrylamide gel. It is not absolutely required to quantify the results to gain insight into the expression of mutant RNAs and putative risk ofthe desired phenotypic changes in target cells. The expression of mRNA whose protein product is implicated in the development ofthe phenotype is adequate to establish risk. If probes of comparable specific activity are used for both franscripts, then a qualitative comparison of RNA levels will be adequate and will decrease the cost ofthe initial diagnosis. Higher mutant form to
wild-type ratios will be coπelated with higher risk whether RNA levels are compared qualitatively or quantitatively.
Additional Uses Potential usefulness of sequence-specific enzymatic nucleic acid molecules ofthe instant invention can have many ofthe same applications for the study of RNA that DNA restriction endonucleases have for the study of DNA (Nathans et al, 1975 Ann. Rev. Biochem. 44:273). For example, the pattern of restriction fragments can be used to establish sequence relationships between two related RNAs, and large RNAs could be specifically cleaved to fragments of a size more useful for study. The ability to engineer sequence specificity ofthe enzymatic nucleic acid molecule is ideal for cleavage of RNAs of unknown sequence. Applicant describes the use of nucleic acid molecules to down- regulate gene expression of target genes in bacterial, microbial, fungal, viral, and eukaryotic systems including plant, or mammalian cells. All patents and publications mentioned in the specification are indicative ofthe levels of skill of those skilled in the art to which the invention pertains. All references cited in this disclosure are incoφorated by reference to the same extent as if each reference had been incoφorated by reference in its entirety individually.
One skilled in the art would readily appreciate that the present invention is well adapted to carry out the objects and obtain the ends and advantages mentioned, as well as those inherent therein. The methods and compositions described herein as presently representative of prefeπed embodiments are exemplary and are not intended as limitations on the scope ofthe invention. Changes therein and other uses will occur to those skilled in the art, which are encompassed within the spirit ofthe invention, are defined by the scope ofthe claims.
It will be readily apparent to one skilled in the art that varying substitutions and modifications may be made to the invention disclosed herein without departing from the scope and spirit ofthe invention. Thus, such additional embodiments are within the scope ofthe present invention and the following claims. The invention illustratively described herein suitably may be practiced in the absence of any element or elements, limitation or limitations which is not specifically disclosed herein. Thus, for example, in each instance herein any ofthe terms "comprising",
"consisting essentially of and "consisting of may be replaced with either ofthe other two terms. The terms and expressions which have been employed are used as terms of description and not of limitation, and there is no intention that in the use of such terms and expressions of excluding any equivalents ofthe features shown and described or portions thereof, but it is recognized that various modifications are possible within the scope ofthe invention claimed. Thus, it should be understood that although the present invention has been specifically disclosed by prefeπed embodiments, optional features, modification and variation ofthe concepts herein disclosed may be resorted to by those skilled in the art, and that such modifications and variations are considered to be within the scope of this invention as defined by the description and the appended claims.
In addition, where features or aspects ofthe invention are described in terms of Markush groups or other grouping of alternatives, those skilled in the art will recognize that the invention is also thereby described in terms of any individual member or subgroup of members ofthe Markush group or other group. Thus, additional embodiments are within the scope ofthe invention and within the following claims
Table 1
TABLE 1
Characteristics of naturally occurring ribozymes Group I Introns
Size: -150 to >1000 nucleotides.
• Requires a U in the target sequence immediately 5' of the cleavage site.
• Binds 4-6 nucleotides at the 5'-side of the cleavage site.
• Reaction mechanism: attack by the 3' -OH of guanosine to generate cleavage products with 3'-OH and 5'-guanosine.
• Additional protein cofactors required in some cases to help folding and maintainance of the active structure.
• Over 300 known members of this class. Found as an intervening sequence in Tetrahymena tliermophila rRNA, fungal mitochondria, chloroplasts, phage T4, blue-green algae, and others.
• Major structural features largely established through phylogenetic comparisons, mutagenesis, and biochemical studies [',"].
• Complete kinetic framework established for one ribozyme ["VvΛvi]-
• Studies of ribozyme folding and substrate docking underway [vii,viii,ix]-
• Chemical modification investigation of important residues well established [x,xi].
• The small (4-6 nt) binding site may make this ribozyme too non-specific for targeted RNA cleavage, however, the Tetrahymena group I intron has been used to repair a "defective" β-galactosidase message by the ligation of new β-galactosidase sequences onto the defective message [xii].
RNAse P RNA (M1 RNA)
Size: -290 to 400 nucleotides.
• RNA portion of a ubiquitous ribonucleoprotein enzyme.
• Cleaves tRNA precursors to form mature tRNA [xiii].
2+
• Reaction mechanism: possible attack by M -OH to generate cleavage products with 3'- OH and 5' -phosphate.
• RNAse P is found throughout the prokaryotes and eukaryotes. The RNA subunit has been sequenced from bacteria, yeast, rodents, and primates.
• Recruitment of endogenous RNAse P for therapeutic applications is possible through hybridization of an External Guide Sequence (EGS) to the target RNA [xiv,χv]
• Important phosphate and 2' OH contacts recently identified ["vi viij
Group II Introns
• Size: >1000 nucleotides.
• Trans cleavage of target RNAs recently demonstrated [xvπ^xix].
Table 1
• Sequence requirements not fully determined.
• Reaction mechanism: 2'-OH of an internal adenosine generates cleavage products with 3'- OH and a "lariat" RNA containing a 3' -5' and a 2' -5' branch point.
• Only natural ribozyme with demonstrated participation in DNA cleavage [x ,x i] in addition to RNA cleavage and ligation.
• Major structural features largely established through phylogenetic comparisons [xxϋ].
• Important 2' OH contacts beginning to be identified [xxiii]
• Kinetic framework under development [xxiv]
Neurospora VS RNA
• Size: -144 nucleotides.
• Trans cleavage of hairpin target RNAs recently demonstrated [xxv].
• Sequence requirements not fully determined.
• Reaction mechanism: attack by 2' -OH 5' to the scissile bond to generate cleavage products with 2',3'-cyclic phosphate and 5' -OH ends.
• Binding sites and structural requirements not fully determined.
• Only 1 known member of this class. Found in Neurospora VS RNA.
Hammerhead Ribozyme
(see text for references)
• Size: -13 to 40 nucleotides.
• Requires the target sequence UH immediately 5' of the cleavage site.
• Binds a variable number nucleotides on both sides of the cleavage site.
• Reaction mechanism: attack by 2'-OH 5' to the scissile bond to generate cleavage products with 2',3'-cyclic phosphate and 5'-OH ends.
• 14 known members of this class. Found in a number of plant pathogens (virusoids) that use RNA as the infectious agent.
• Essential structural features largely defined, including 2 crystal structures [xx ^ xvϋj
• Minimal ligation activity demonstrated (for engineering through in vitro selection) [ xviii]
• Complete kinetic framework established for two or more ribozymes [xxix].
• Chemical modification investigation of important residues well established [xxx].
Hairpin Ribozyme
• Size: -50 nucleotides.
• Requires the target sequence GUC immediately 3' of the cleavage site.
• Binds 4-6 nucleotides at the 5'-side of the cleavage site and a variable number to the 3'- side of the cleavage site.
• Reaction mechanism: attack by 2' -OH 5' to the scissile bond to generate cleavage products with 2',3'-cyclic phosphate and 5' -OH ends.
• 3 known members of this class. Found in three plant pathogen (satellite RNAs of the tobacco ringspot virus, arabis mosaic virus and chicory yellow mottle virus) which uses RNA as the infectious agent.
• Essential structural features largely defined [ χi / χχχii / χχχ»i / χχχiv]
Table 1
• Ligation activity (in addition to cleavage activity) makes ribozyme amenable to engineering through in vitro selection [xx v]
• Complete kinetic framework established for one ribozyme [χχχvι].
• Chemical modification investigation of important residues begun [ χχ u xxxvmj
Hepatitis Delta Virus (HDV) Ribozyme
• Size: -60 nucleotides.
• Trans cleavage of target RNAs demonstrated [xχχι ].
• Binding sites and structural requirements not fully determined, although no sequences 5' of cleavage site are required. Folded ribozyme contains a pseudoknot structure [x1].
• Reaction mechanism: attack by 2' -OH 5' to the scissile bond to generate cleavage products with 2',3' -cyclic phosphate and 5'-OH ends.
• Only 2 known members of this class. Found in human HDV.
• Circular form of HDV is active and shows increased nuclease stability [xh]
Michel, Francois; Westhof, Eric. Slippery substrates. Nat. Struct. Biol. (1994), 1(1), 5-7. u . Lisacek, Frederique; Diaz, Yolande; Michel, Francois. Automatic identification of group I intron cores in genomic DNA sequences. J. Mol. Biol. (1994), 235(4), 1206-17. m . Herschlag, Daniel; Cech, Thomas R.. Catalysis of RNA cleavage by the Tetrahymena thermophila ribozyme. 1. Kinetic description of the reaction of an RNA substrate complementary to the active site. Biochemistry (1990), 29(44), 10159-71.
1V . Herschlag, Daniel; Cech, Thomas R.. Catalysis of RNA cleavage by the Tetrahymena thermophila ribozyme. 2. Kinetic description of the reaction of an RNA substrate that forms a mismatch at the active site. Biochemistry (1990), 29(44), 10172-80. v . Knitt, Deborah S.; Herschlag, Daniel. pH Dependencies of the Tetrahymena Ribozyme Reveal an
Unconventional Origin of an Apparent pKa. Biochemistry (1996), 35(5), 1560-70.
V1 . Bevilacqua, Philip C; Sugimoto, Naoki; Turner, Douglas H.. A mechanistic framework for the second step of splicing catalyzed by the Tetrahymena ribozyme. Biochemistry (1996), 35(2), 648-58. vu . Li, Yi; Bevilacqua, Philip C; Mathews, David; Turner, Douglas H.. Thermodynamic and activation parameters for binding of a pyrene-labeled substrate by the Tetrahymena ribozyme: docking is not diffusion-controlled and is driven by a favorable entropy change. Biochemistry (1995), 34(44), 14394-9. vm . Banerjee, Aloke Raj; Turner, Douglas H.. The time dependence of chemical modification reveals slow steps in the folding of a group I ribozyme. Biochemistry (1995), 34(19), 6504-12. ■* . Zarrinkar, Patrick P.; Williamson, James R.. The P9.1-P9.2 peripheral extension helps guide folding of the Tetrahymena ribozyme. Nucleic Acids Res. (1996), 24(5), 854-8.
* . Strobel, Scott A.; Cech, Thomas R.. Minor groove recognition of the conserved G.cntdot.U pair at the Tetrahymena ribozyme reaction site. Science (Washington, D. C.) (1995), 267(5198), 675-9. "• . Strobel, Scott A.; Cech, Thomas R.. Exocyclic Amine of the Conserved G.cntdo U Pair at the
Cleavage Site of the Tetrahymena Ribozyme Contributes to 5'-Splice Site Selection and Transition State Stabilization. Biochemistry (1996), 35(4), 1201-11. xu. Sullenger, Bruce A.; Cech, Thomas R.. Ribozyme-mediated repair of defective mRNA by targeted trans-splicing. Nature (London) (1994), 371(6498), 619-22.
Robertson, H.D.; Altaian, S.; Smith, J.D. J. Biol. Chem., 247, 5243-5251 (1972). *1V. Forster, Anthony C; Altaian, Sidney. External guide sequences for an RNA enzyme. Science
(Washington, D. C, 1883-) (1990), 249(4970), 783-6.
*v. Yuan, Y.; Hwang, E. S.; Altaian, S. Targeted cleavage of mRNA by human RNase P. Proc. Natl.
Acad. Sci. USA (1992) 89, 8006-10. *V1 . Harris, Michael E.; Pace, Norman R.. Identification of phosphates involved in catalysis by the
Table 1
ribozyme RNase P RNA. RNA (1995), 1(2), 210-18. xvu . Pan, Tao; Loria, Andrew; Zhong, Kun. Probing of tertiary interactions in RNA: 2'-hydroxyl-base contacts between the RNase P RNA and pre-tRNA. Proc. Natl. Acad. Sci. U. S. A. (1995), 92(26), 12510-14. xvm Pyle, Anna Marie; Green, Justin B.. Building a Kinetic Framework for Group II Intron Ribozyme
Activity: Quantitation of Interdomain Binding and Reaction Rate. Biochemistry (1994), 33(9), 2716-25. X1X . Michels, William J. Jr.; Pyle, Anna Marie. Conversion of a Group II Intron into a New Multiple-
Turnover Ribozyme that Selectively Cleaves Oligonucleotides: Elucidation of Reaction Mechanism and Structure/ Function Relationships. Biochemistry (1995), 34(9), 2965-77. xx . Zimmerly, Steven; Guo, Huatao; Eskes, Robert; Yang, Jian; Perlman, Philip S.; Lambowitz, Alan
M.. A group II intron RNA is a catalytic component of a DNA endonuclease involved in intron mobility. Cell (Cambridge, Mass.) (1995), 83(4), 529-38.
*X1 . Griffin, Edmund A., Jr.; Qin, Zhifeng; Michels, Williams J., Jr.; Pyle, Anna Marie. Group II intron ribozymes that cleave DNA and RNA linkages with similar efficiency, and lack contacts with substrate 2'- hydroxyl groups. Chem. Biol. (1995), 2(11), 761-70. xu . Michel, Francois; Ferat, Jean Luc. Structure and activities of group II introns. Annu. Rev. Biochem. (1995), 64, 435-61. xx . Abramovitz, Dana L.; Friedman, Richard A.; Pyle, Anna Marie. Catalytic role of 2'-hydroxyl groups within a group II intron active site. Science (Washington, D. C.) (1996), 271(5254), 1410-13. X!αv . Daniels, Danette L.; Michels, William J., Jr.; Pyle, Anna Marie. Two competing pathways for self- splicing by group II introns: a quantitative analysis of in vitro reaction rates and products. J. Mol. Biol. (1996), 256(1), 31-49. XV . Guo, Hans C. T.; Collins, Richard A.. Efficient trans-cleavage of a stem-loop RNA substrate by a ribozyme derived from Neurospora VS RNA. EMBO J. (1995), 14(2), 368-76. XXVI . Scott, W.G., Finch, J.T., Aaron,K. The crystal structure of an all RNA hammerhead ribozyme:Aproposed mechanism for RNA catalytic cleavage. Cell, (1995), 81, 991-1002. xxvu McKay, Structure and function of the hammerhead ribozyme: an unfinished story. RNA, (1996), 2, 395-403. xxvώ . Long, D., Uhlenbeck, O., Hertel, K. Ligation with hammerhead ribozymes. US Patent No. 5,633,133.
XX1X . Hertel, K.J., Herschlag, D., Uhlenbeck, O. A kinetic and thermodynamic framework for the hammerhead ribozyme reaction. Biochemistry, (1994) 33, 3374-3385.Beigelman, L., et al., Chemical modifications of hammerhead ribozymes. J. Biol. Chem., (1995) 270, 25702-25708. xxx Beigelman, L., et al., Chemical modifications of hammerhead ribozymes. J. Biol. Chem., (1995)
270, 25702-25708.
X XI . Hampel, Arnold; Tritz, Richard; Hicks, Margaret; Cruz, Phillip. 'Hairpin' catalytic RNA model: evidence for helixes and sequence requirement for substrate RNA. Nucleic Acids Res. (1990), 18(2), 299- 304. u Chowrira, Bharat M.; Berzal-Herranz, Alfredo; Burke, John M.. Novel guanosine requirement for catalysis by the hairpin ribozyme. Nature (London) (1991), 354(6351), 320-2. xx ui Berzal-Herranz, Alfredo; Joseph, Simpson; Chowrira, Bharat M.; Butcher, Samuel E.; Burke, John M.. Essential nucleotide sequences and secondary structure elements of the hairpin ribozyme. EMBO J. (1993), 12(6), 2567-73. xx iv Joseph, Simpson; Berzal-Herranz, Alfredo; Chowrira, Bharat M.; Butcher, Samuel E.. Substrate selection rules for the hairpin ribozyme determined by in vitro selection, mutation, and analysis of mismatched substrates. Genes Dev. (1993), 7(1), 130-8. x v Berzal-Herranz, Alfredo; Joseph, Simpson; Burke, John M.. In vitro selection of active hairpin ribozymes by sequential RNA-catalyzed cleavage and ligation reactions. Genes Dev. (1992), 6(1), 129-34. xx vi Hegg, Lisa A.; Fedor, Martha J.. Kinetics and Thermodynamics of Intermolecular Catalysis by Hairpin Ribozymes. Biochemistry (1995), 34(48), 15813-28. xxxvu Grasby, Jane A.; Mersmann, Karin; Singh, Mohinder; Gait, Michael J.. Purine Functional Groups in Essential Residues of the Hairpin Ribozyme Required for Catalytic Cleavage of RNA. Biochemistry
Table 1
(1995), 34(12), 4068-76. xxvui Schmidt, Sabine; Beigelman, Leonid; Karpeisky, Alexander; Usman, Nassim; Sorensen, Ulrik S.; Gait, Michael J.. Base and sugar requirements for RNA cleavage of essential nucleoside residues in internal loop B of the hairpin ribozyme: implications for secondary structure. Nucleic Acids Res. (1996), 24(4), 573-81. xx ix Perrotta, Anne T.; Been, Michael D.. Cleavage of oligoribonucleotides by a ribozyme derived from the hepatitis .delta, virus RNA sequence. Biochemistry (1992), 31(1), 16-21. xl . Perrotta, Anne T.; Been, Michael D.. A pseudoknot-like structure required for efficient self- cleavage of hepatitis delta virus RNA. Nature (London) (1991), 350(6317), 434-6. xU . Puttaraju, M.; Perrotta, Anne T.; Been, Michael D.. A circular trans-acting hepatitis delta virus ribozyme. Nucleic Acids Res. (1993), 21(18), 4253-8.
Table 2
Table 2:
A. 2.5 μmol Synthesis Cycle ABI 394 Instrument
Reagent Equivalents Amount Wait Time* DNA Wait Time* 2'- Wait Time' O-methyl RNA
Phosphoramidites 6.5 163 μL 45 sec 2.5 min 7.5 min S-Ethyl Tetrazole 23.8 238 μL 45 sec 2.5 min 7.5 min Acetic Anhydride 100 233 μL 5 sec 5 sec 5 sec Λ/-Methyl Imidazole 186 233 μL 5 sec 5 sec 5 sec TCA 176 2.3 mL 21 sec 21 sec 21 sec Iodine 11.2 1.7 mL 45 sec 45 sec 45 sec Beaucage 12.9 645 μL 100 sec 300 sec 300 sec Acetonitrile NA 6.67 mL NA NA NA
B. 0.2 μmol Synthesis Cycle ABI 394 Instrument
Reagent Equivalents Amount Wait Time* DNA Wait Time* 2'- Wait Time* O-methyl RNA
Phosphoramidites 15 31 μL 45 sec 233 sec 465 sec
S-Ethyl Tetrazole 38.7 31 μL 45 sec 233 min 465 sec
Acetic Anhydride 655 124 μL 5 sec 5 sec 5 sec
Λ/-Methyl Imidazole 1245 124 μL 5 sec 5 sec 5 sec
TCA 700 732 μL 10 sec 10 sec 10 sec
Iodine 20.6 244 μL 15 sec 15 sec 15 sec
Beaucage 71 232 μL 100 sec 300 sec 300 sec
Table 2
Acetonitrile NA 2.64 mL NA NA NA
C. 0.2 μmol Synthesis Cycle 96 well Instrument
Reagent Equivalents Amount Wait Time* Wait Time* 2'-0- Wait Time* Ribo
DNA 2'-0-me thyl/Ribo DNA/2'-0-methyl/Ribo DNA methyl
Phosphoramidites 22/33/66 40/60/120 μL 60 sec 180 sec 360sec
S-Ethyl Tetrazole 70/105/210 40/60/120 μL 60 sec 180 min 360 sec
Acetic Anhydride 265/265/265 50/50/50 μL 10 sec 10 sec 10 sec
Λ/-Methyl Imidazole 502/502/502 50/50/50 μL 10 sec 10 sec 10 sec
TCA 238/475/475 250/500/500 μL 15 sec 15 sec 15 sec
Iodine 6.8/6.8/6.8 80/80/80 μL 30 sec 30 sec 30 sec
Beaucage 34/51/51 80/120/120 100 sec 200 sec 200 sec
Acetonitrile NA 1150/1150/1150 μL NA NA NA
* Wait time does not include contact time during delivery.
Table 3 Table 3: Human PTP-IB Hammerhead Ribozyme and Target Sequence
Nt. Ribozyme Sequence Seq. ID Substrate Sequence Seq. ID Position Nos. Nos.
15 UGCCGCUC CUGAUGAG X CGAA AGGCCGCG 1 CGCGGCCT A GAGCGGCA 529
72 AUCUCCAU CUGAUGAG X CGAA ACGGGCCA 2 TGGCCCGT C ATGGAGAT 530
92 UCUGCUCG CUGAUGAG X CGAA ACUCCUUU 3 AAAGGAGT T CGAGCAGA 531
93 AUCUGCUC CUGAUGAG X CGAA AACUCCUU 4 AAGGAGTT C GAGCAGAT 532
102 GACUUGUC CUGAUGAG X CGAA AUCUGCUC 5 GAGCAGAT C GACAAGTC 533
110 AGCUCCCG CUGAUGAG X CGAA ACUUGUCG 6 CGACAAGT c CGGGAGCT 534
129 UCCUGGUA CUGAUGAG X CGAA AUGGCCGC 7 GCGGCCAT T TACCAGGA 535
130 AUCCUGGU CUGAUGAG X CGAA AAUGGCCG 8 CGGCCATT T ACCAGGAT 536
131 UAUCCUGG CUGAUGAG X CGAA AAAUGGCC 9 GGCCATTT A CCAGGATA 537
139 AUGUCGGA CUGAUGAG X CGAA AUCCUGGU 10 ACCAGGAT A TCCGACAT 538
141 UCAUGUCG CUGAUGAG X CGAA AUAUCCUG 11 CAGGATAT c CGACATGA 539
161 UACAUGGG CUGAUGAG X CGAA AGUCACUG 12 CAGTGACT T CCCATGTA 540
162 CUACAUGG CUGAUGAG X CGAA AAGUCACU 13 AGTGACTT c CCATGTAG 541
169 GGCCACUC CUGAUGAG X CGAA ACAUGGGA 14 TCCCATGT A GAGTGGCC 542
183 UUCUUAGG CUGAUGAG X CGAA AGCUUGGC 15 GCCAAGCT T CCTAAGAA 543
184 GUUCUUAG CUGAUGAG X CGAA AAGCUUGG 16 CCAAGCTT c CTAAGAAC 544
187 UUUGUUCU CUGAUGAG X CGAA AGGAAGCU 17 AGCTTCCT A AGAACAAA 545
205 UCUGUACC CUGAUGAG X CGAA AUUUCGGU 18 ACCGAAAT A GGTACAGA 546
209 CGUCUCUG CUGAUGAG X CGAA ACCUAUUU 19 AAATAGGT A CAGAGACG 547
219 AAGGGACU CUGAUGAG X CGAA ACGUCUCU 20 AGAGACGT C AGTCCCTT 548
223 GUCAAAGG CUGAUGAG X CGAA ACUGACGU 21 ACGTCAGT C CCTTTGAC 549
227 UAUGGUCA CUGAUGAG X CGAA AGGGACUG 22 CAGTCCCT T TGACCATA 550
228 CUAUGGUC CUGAUGAG X CGAA AAGGGACU 23 AGTCCCTT T GACCATAG 551
235 AAUCCGAC CUGAUGAG X CGAA AUGGUCAA 24 TTGACCAT A GTCGGATT 552
238 UUUAAUCC CUGAUGAG X CGAA ACUAUGGU 25 ACCATAGT C GGATTAAA 553
243 UGUAGUUU CUGAUGAG X CGAA AUCCGACU 26 AGTCGGAT T AAACTACA 554
244 AUGUAGUU CUGAUGAG X CGAA AAUCCGAC 27 GTCGGATT A AACTACAT 555
249 UCUUGAUG CUGAUGAG X CGAA AGUUUAAU 28 ATTAAACT A CATCAAGA 556
253 AUCUUCUU CUGAUGAG X CGAA AUGUAGUU 29 AACTACAT C AAGAAGAT 557
262 AUAGUCAU CUGAUGAG X CGAA AUCUUCUU 30 AAGAAGAT A ATGACTAT 558
269 CGUUGAUA CUGAUGAG X CGAA AGUCAUUA 31 TAATGACT A TATCAACG 559
271 AGCGUUGA CUGAUGAG X CGAA AUAGUCAU 32 ATGACTAT A TCAACGCT 560
273 CUAGCGUU CUGAUGAG X CGAA AUAUAGUC 33 GACTATAT C AACGCTAG 561
280 UAUCAAAC CUGAUGAG X CGAA AGCGUUGA 34 TCAACGCT A GTTTGATA 562
283 UUUUAUCA CUGAUGAG X CGAA ACUAGCGU 35 ACGCTAGT T TGATAAAA 563
284 UUUUUAUC CUGAUGAG X CGAA AACUAGCG 36 CGCTAGTT T GATAAAAA 564
288 UCCAUUUU CUGAUGAG X CGAA AUCAAACU 37 AGTTTGAT A AAAATGGA 565
313 AAGAAUGU CUGAUGAG X CGAA ACUCCUUU 38 AAAGGAGT T ACATTCTT 566
314 UAAGAAUG CUGAUGAG X CGAA AACUCCUU 39 AAGGAGTT A CATTCTTA 567
318 UGGGUAAG CUGAUGAG X CGAA AUGUAACU 40 AGTTACAT T CTTACCCA 568
319 CUGGGUAA CUGAUGAG X CGAA AAUGUAAC 41 GTTACATT C TTACCCAG 569
321 CCCUGGGU CUGAUGAG X CGAA AGAAUGUA 42 TACATTCT T ACCCAGGG 570
322 GCCCUGGG CUGAUGAG X CGAA AAGAAUGU 43 ACATTCTT A CCCAGGGC 571
334 GUUAGGCA CUGAUGAG X CGAA AGGGCCCU 44 AGGGCCCT T TGCCTAAC 572
Table 3
335 UGUUAGGC CUGAUGAG X CGAA AAGGGCCC 45 GGGCCCTT T GCCTAACA 573
340 GCAUGUGU CUGAUGAG X CGAA AGGCAAAG 46 CTTTGCCT A ACACATGC 574
352 CCAAAAGU CUGAUGAG X CGAA ACCGCAUG 47 CATGCGGT C ACTTTTGG 575
356 UCUCCCAA CUGAUGAG X CGAA AGUGACCG 48 CGGTCACT T TTGGGAGA 576
357 AUCUCCCA CUGAUGAG X CGAA AAGUGACC 49 GGTCACTT T TGGGAGAT 577
358 CAUCUCCC CUGAUGAG X CGAA AAAGUGAC 50 GTCACTTT T GGGAGATG 578
393 AGCAUGAC CUGAUGAG X CGAA ACACCCCU 51 AGGGGTGT C GTCATGCT 579
396 UUGAGCAU CUGAUGAG X CGAA ACGACACC 52 GGTGTCGT c ATGCTCAA 580
402 ACUCUGUU CUGAUGAG X CGAA AGCAUGAC 53 GTCATGCT c AACAGAGT 581
424 UUUUAACG CUGAUGAG X CGAA ACCUUUCU 54 AGAAAGGT T CGTTAAAA 582
425 AUUUUAAC CUGAUGAG X CGAA AACCUUUC 55 GAAAGGTT C GTTAAAAT 583
428 CGCAUUUU CUGAUGAG X CGAA ACGAACCU 56 AGGTTCGT T AAAATGCG 584
429 GCGCAUUU CUGAUGAG X CGAA AACGAACC 57 GGTTCGTT A AAATGCGC 585
443 GUGGCCAG CUGAUGAG X CGAA AUUGUGCG 58 CGCACAAT A CTGGCCAC 586
474 UCUUCAAA CUGAUGAG X CGAA AUCAUCUC 59 GAGATGAT C TTTGAAGA 587
476 UGUCUUCA CUGAUGAG X CGAA AGAUCAUC 60 GATGATCT T TGAAGACA 588
477 GUGUCUUC CUGAUGAG X CGAA AAGAUCAU 61 ATGATCTT T GAAGACAC 589
490 UAAUUUCA CUGAUGAG X CGAA AUUUGUGU 62 ACACAAAT T TGAAATTA 590
491 UUAAUUUC CUGAUGAG X CGAA AAUUUGUG 63 CACAAATT T GAAATTAA 591
497 UCAAUGUU CUGAUGAG X CGAA AUUUCAAA 64 TTTGAAAT T AACATTGA 592
498 AUCAAUGU CUGAUGAG X CGAA AAUUUCAA 65 TTGAAATT A ACATTGAT 593
503 CAGAGAUC CUGAUGAG X CGAA AUGUUAAU 66 ATTAACAT T GATCTCTG 594
507 UCUUCAGA CUGAUGAG X CGAA AUCAAUGU 67 ACATTGAT c TCTGAAGA 595
509 UAUCUUCA CUGAUGAG X CGAA AGAUCAAU 68 ATTGATCT C TGAAGATA 596
517 UGACUUGA CUGAUGAG X CGAA AUCUUCAG 69 CTGAAGAT A TCAAGTCA 597
519 UAUGACUU CUGAUGAG X CGAA AUAUCUUC 70 GAAGATAT C AAGTCATA 598
524 UAUAAUAU CUGAUGAG X CGAA ACUUGAUA 71 TATCAAGT C ATATTATA 599
527 CUGUAUAA CUGAUGAG X CGAA AUGACUUG 72 CAAGTCAT A TTATACAG 600
529 CACUGUAU CUGAUGAG X CGAA AUAUGACU 73 AGTCATAT T ATACAGTG 601
530 GCACUGUA CUGAUGAG X CGAA AAUAUGAC 74 GTCATATT A TACAGTGC 602
532 UCGCACUG CUGAUGAG X CGAA AUAAUAUG 75 CATATTAT A CAGTGCGA 603
546 UCCAAUUC CUGAUGAG X CGAA AGCUGUCG 76 CGACAGCT A GAATTGGA 604
551 GGUUUUCC CUGAUGAG X CGAA AUUCUAGC 77 GCTAGAAT T GGAAAACC 605
561 UGGGUUGU CUGAUGAG X CGAA AGGUUUUC 78 GAAAACCT T ACAACCCA 606
562 UUGGGUUG CUGAUGAG X CGAA AAGGUUUU 79 AAAACCTT A CAACCCAA 607
577 GAUCUCUC CUGAUGAG X CGAA AGUUUCUU 80 AAGAAACT C GAGAGATC 608
585 AAAUGUAA CUGAUGAG X CGAA AUCUCUCG 81 CGAGAGAT C TTACATTT 609
587 GGAAAUGU CUGAUGAG X CGAA AGAUCUCU 82 AGAGATCT T ACATTTCC 610
588 UGGAAAUG CUGAUGAG X CGAA AAGAUCUC 83 GAGATCTT A CATTTCCA 611
592 AUAGUGGA CUGAUGAG X CGAA AUGUAAGA 84 TCTTACAT T TCCACTAT 612
593 UAUAGUGG CUGAUGAG X CGAA AAUGUAAG 85 CTTACATT T CCACTATA 613
594 GUAUAGUG CUGAUGAG X CGAA AAAUGUAA 86 TTACATTT C CACTATAC 614
599 AUGUGGUA CUGAUGAG X CGAA AGUGGAAA 87 TTTCCACT A TACCACAT 615
601 CCAUGUGG CUGAUGAG X CGAA AUAGUGGA 88 TCCACTAT A CCACATGG 616
617 GGACUCCA CUGAUGAG X CGAA AGUCAGGC 89 GCCTGACT T TGGAGTCC 617
618 GGGACUCC CUGAUGAG X CGAA AAGUCAGG 90 CCTGACTT T GGAGTCCC 618
624 GAUUCAGG CUGAUGAG X CGAA ACUCCAAA 91 TTTGGAGT C CCTGAATC 619
Table 3
632 AGGCUGGU CUGAUGAG X CGAA AUUCAGGG 92 CCCTGAAT C ACCAGCCT 620
641 UCAAGAAU CUGAUGAG X CGAA AGGCUGGU 93 ACCAGCCT C ATTCTTGA 621
644 AGUUCAAG CUGAUGAG X CGAA AUGAGGCU 94 AGCCTCAT T CTTGAACT 622
645 AAGUUCAA CUGAUGAG X CGAA AAUGAGGC 95 GCCTCATT C TTGAACTT 623
647 GAAAGUUC CUGAUGAG X CGAA AGAAUGAG 96 CTCATTCT T GAACTTTC 624
653 UGAAAAGA CUGAUGAG X CGAA AGUUCAAG 97 CTTGAACT T TCTTTTCA 625
654 UUGAAAAG CUGAUGAG X CGAA AAGUUCAA 98 TTGAACTT T CTTTTCAA 626
655 UUUGAAAA CUGAUGAG X CGAA AAAGUUCA 99 TGAACTTT C TTTTCAAA 627
657 ACUUUGAA CUGAUGAG X CGAA AGAAAGUU 100 AACTTTCT T TTCAAAGT 628
658 GACUUUGA CUGAUGAG X CGAA AAGAAAGU 101 ACTTTCTT T TCAAAGTC 629
659 GGACUUUG CUGAUGAG X CGAA AAAGAAAG 102 CTTTCTTT T CAAAGTCC 630
660 CGGACUUU CUGAUGAG X CGAA AAAAGAAA 103 TTTCTTTT C AAAGTCCG 631
666 GACUCUCG CUGAUGAG X CGAA ACUUUGAA 104 TTCAAAGT C CGAGAGTC 632
674 GUGACCCU CUGAUGAG X CGAA ACUCUCGG 105 CCGAGAGT C AGGGTCAC 633
680 GGCUGAGU CUGAUGAG X CGAA ACCCUGAC 106 GTCAGGGT C ACTCAGCC 634
684 UCCGGGCU CUGAUGAG X CGAA AGUGACCC 107 GGGTCACT C AGCCCGGA 635
705 UGCACCAC CUGAUGAG X CGAA ACGGGCCC 108 GGGCCCGT T GTGGTGCA 636
729 GACCUGCC CUGAUGAG X CGAA AUGCCUGC 109 GCAGGCAT C GGCAGGTC 637
737 AGGUUCCA CUGAUGAG X CGAA ACCUGCCG 110 CGGCAGGT C TGGAACCT 638
746 CCAGACAG CUGAUGAG X CGAA AGGUUCCA 111 TGGAACCT T CTGTCTGG 639
747 GCCAGACA CUGAUGAG X CGAA AAGGUUCC 112 GGAACCTT C TGTCTGGC 640
751 AUCAGCCA CUGAUGAG X CGAA ACAGAAGG 113 CCTTCTGT C TGGCTGAT 641
760 GAGGCAGG CUGAUGAG X CGAA AUCAGCCA 114 TGGCTGAT A CCTGCCTC 642
768 AUCAGCAA CUGAUGAG X CGAA AGGCAGGU 115 ACCTGCCT C TTGCTGAT 643
770 CCAUCAGC CUGAUGAG X CGAA AGAGGCAG 116 CTGCCTCT T GCTGATGG 644
796 AACGGAAG CUGAUGAG X CGAA AGGGUCUU 117 AAGACCCT T CTTCCGTT 645
797 CAACGGAA CUGAUGAG X CGAA AAGGGUCU 118 AGACCCTT c TTCCGTTG 646
799 AUCAACGG CUGAUGAG X CGAA AGAAGGGU 119 ACCCTTCT T CCGTTGAT 647
800 UAUCAACG CUGAUGAG X CGAA AAGAAGGG 120 CCCTTCTT c CGTTGATA 648
804 UUGAUAUC CUGAUGAG X CGAA ACGGAAGA 121 TCTTCCGT T GATATCAA 649
808 UUUCUUGA CUGAUGAG X CGAA AUCAACGG 122 CCGTTGAT A TCAAGAAA 650
810 ACUUUCUU CUGAUGAG X CGAA AUAUCAAC 123 GTTGATAT c AAGAAAGT 651
824 UCAUUUCU CUGAUGAG X CGAA ACAGCACU 124 AGTGCTGT T AGAAATGA 652
825 CUCAUUUC CUGAUGAG X CGAA AACAGCAC 125 GTGCTGTT A GAAATGAG 653
839 CCAUCCGA CUGAUGAG X CGAA ACUUCCUC 126 GAGGAAGT T TCGGATGG 654
840 CCCAUCCG CUGAUGAG X CGAA AACUUCCU 127 AGGAAGTT T CGGATGGG 655
841 CCCCAUCC CUGAUGAG X CGAA AAACUUCC 128 GGAAGTTT c GGATGGGG 656
855 GCUGUCUG CUGAUGAG X CGAA AUCAGCCC 129 GGGCTGAT c CAGACAGC 657
878 GGUAGGAG CUGAUGAG X CGAA AGCGCAGC 130 GCTGCGCT T CTCCTACC 658
879 AGGUAGGA CUGAUGAG X CGAA AAGCGCAG 131 CTGCGCTT c TCCTACCT 659
881 CCAGGUAG CUGAUGAG X CGAA AGAAGCGC 132 GCGCTTCT c CTACCTGG 660
884 CAGCCAGG CUGAUGAG X CGAA AGGAGAAG 133 CTTCTCCT A CCTGGCTG 661
897 GCACCUUC CUGAUGAG X CGAA AUCACAGC 134 GCTGTGAT C GAAGGTGC 662
911 CCAUGAUG CUGAUGAG X CGAA AUUUGGCA 135 TGCCAAAT T CATCATGG 663
912 CCCAUGAU CUGAUGAG X CGAA AAUUUGGC 136 GCCAAATT c ATCATGGG 664
915 UCCCCCAU CUGAUGAG X CGAA AUGAAUUU 137 AAATTCAT c ATGGGGGA 665
926 GCACGGAA CUGAUGAG X CGAA AGUCCCCC 138 GGGGGACT c TTCCGTGC 666
Table 3
928 CUGCACGG CUGAUGAG X CGAA AGAGUCCC 139 GGGACTCT T CCGTGCAG 667
929 CCUGCACG CUGAUGAG X CGAA AAGAGUCC 140 GGACTCTT C CGTGCAGG 668
940 CUUCCACU CUGAUGAG X CGAA AUCCUGCA 141 TGCAGGAT C AGTGGAAG 669
954 UCGUGGGA CUGAUGAG X CGAA AGCUCCUU 142 AAGGAGCT T TCCCACGA 670
955 CUCGUGGG CUGAUGAG X CGAA AAGCUCCU 143 AGGAGCTT T CCCACGAG 671
956 CCUCGUGG CUGAUGAG X CGAA AAAGCUCC 144 GGAGCTTT C CCACGAGG 672
988 UGGGGGGA CUGAUGAG X CGAA AUGCUCGG 145 CCGAGCAT A TCCCCCCA 673
990 GGUGGGGG CUGAUGAG X CGAA AUAUGCUC 146 GAGCATAT C CCCCCACC 674
1000 UGGCCGGG CUGAUGAG X CGAA AGGUGGGG 147 CCCCACCT C CCCGGCCA 675
1020 GGCUCCAG CUGAUGAG X CGAA AUUCGUUU 148 AAACGAAT C CTGGAGCC 676
1052 UUGGGAAG CUGAUGAG X CGAA ACUCCCUG 149 CAGGGAGT T CTTCCCAA 677
1053 UUUGGGAA CUGAUGAG X CGAA AACUCCCU 150 AGGGAGTT C TTCCCAAA 678
1055 GAUUUGGG CUGAUGAG X CGAA AGAACUCC 151 GGAGTTCT T CCCAAATC 679
1056 UGAUUUGG CUGAUGAG X CGAA AAGAACUC 152 GAGTTCTT C CCAAATCA 680
1063 CCACUGGU CUGAUGAG X CGAA AUUUGGGA 153 TCCCAAAT C ACCAGTGG 681
1096 GCAGUCUU CUGAUGAG X CGAA AUCCUCCU 154 AGGAGGAT A AAGACTGC 682
1110 UCUUCCUU CUGAUGAG X CGAA AUGGGGCA 155 TGCCCCAT C AAGGAAGA 683
1133 CGGCAUUU CUGAUGAG X CGAA AGGGGCUU 156 AAGCCCCT T AAATGCCG 684
1134 GCGGCAUU CUGAUGAG X CGAA AAGGGGCU 157 AGCCCCTT A AATGCCGC 685
1148 CGAUGCCG CUGAUGAG X CGAA AGGGUGCG 158 CGCACCCT A CGGCATCG 686
1155 AUGCUUUC CUGAUGAG X CGAA AUGCCGUA 159 TACGGCAT C GAAAGCAT 687
1168 AGUGUCUU CUGAUGAG X CGAA ACUCAUGC 160 GCATGAGT C AAGACACT 688
1182 CGACUUCU CUGAUGAG X CGAA ACUUCAGU 161 ACTGAAGT T AGAAGTCG 689
1183 CCGACUUC CUGAUGAG X CGAA AACUUCAG 162 CTGAAGTT A GAAGTCGG 690
1189 CACGACCC CUGAUGAG X CGAA ACUUCUAA 163 TTAGAAGT C GGGTCGTG 691
1194 CCCCCCAC CUGAUGAG X CGAA ACCCGACU 164 AGTCGGGT C GTGGGGGG 692
1207 ACCUCGAA CUGAUGAG X CGAA ACUUCCCC 165 GGGGAAGT C TTCGAGGT 693
1209 GCACCUCG CUGAUGAG X CGAA AGACUUCC 166 GGAAGTCT T CGAGGTGC 694
1210 GGCACCUC CUGAUGAG X CGAA AAGACUUC 167 GAAGTCTT C GAGGTGCC 695
1229 UGGCUGGG CUGAUGAG X CGAA AGGCAGCC 168 GGCTGCCT C CCCAGCCA 696
1250 CGGGCAGU CUGAUGAG X CGAA ACGGCUCC 169 GGAGCCGT C ACTGCCCG 697
1285 CUUCCAGU CUGAUGAG X CGAA ACUCAGUG 170 CACTGAGT T ACTGGAAG 698
1286 GCUUCCAG CUGAUGAG X CGAA AACUCAGU 171 ACTGAGTT A CTGGAAGC 699
1298 UGACCAGG CUGAUGAG X CGAA AGGGCUUC 172 GAAGCCCT T CCTGGTCA 700
1299 UUGACCAG CUGAUGAG X CGAA AAGGGCUU 173 AAGCCCTT C CTGGTCAA 701
1305 CACAUGUU CUGAUGAG X CGAA ACCAGGAA 174 TTCCTGGT C AACATGTG 702
1321 GAGGACCG CUGAUGAG X CGAA AGCCACGC 175 GCGTGGCT A CGGTCCTC 703
1326 GCCGUGAG CUGAUGAG X CGAA ACCGUAGC 176 GCTACGGT C CTCACGGC 704
1329 CCGGCCGU CUGAUGAG X CGAA AGGACCGU 177 ACGGTCCT C ACGGCCGG 705
1342 GCAGAGGU CUGAUGAG X CGAA AGCGCCGG 178 CCGGCGCT T ACCTCTGC 706
1343 AGCAGAGG CUGAUGAG X CGAA AAGCGCCG 179 CGGCGCTT A CCTCTGCT 707
1347 CUGUAGCA CUGAUGAG X CGAA AGGUAAGC 180 GCTTACCT C TGCTACAG 708
1352 GGAACCUG CUGAUGAG X CGAA AGCAGAGG 181 CCTCTGCT A CAGGTTCC 709
1358 UGAACAGG CUGAUGAG X CGAA ACCUGUAG 182 CTACAGGT T CCTGTTCA 710
1359 UUGAACAG CUGAUGAG X CGAA AACCUGUA 183 TACAGGTT C CTGTTCAA 711
1364 UGCUGUUG CUGAUGAG X CGAA ACAGGAAC 184 GTTCCTGT T CAACAGCA 712
1365 UUGCUGUU CUGAUGAG X CGAA AACAGGAA 185 TTCCTGTT C AACAGCAA 713
Table 3
1379 GGUCAGGC CUGAUGAG X CGAA AUGUGUUG 186 CAACACAT A GCCTGACC 714
1390 GAGUGGAG CUGAUGAG X CGAA AGGGUCAG 187 CTGACCCT C CTCCACTC 715
1393 GUGGAGUG CUGAUGAG X CGAA AGGAGGGU 188 ACCCTCCT C CACTCCAC 716
1398 UGGAGGUG CUGAUGAG X CGAA AGUGGAGG 189 CCTCCACT C CACCTCCA 717
1404 AGUGGGUG CUGAUGAG X CGAA AGGUGGAG 190 CTCCACCT C CACCCACT 718
1415 CAGAGGCG CUGAUGAG X CGAA ACAGUGGG 191 CCCACTGT C CGCCTCTG 719
1421 UGCGGGCA CUGAUGAG X CGAA AGGCGGAC 192 GTCCGCCT C TGCCCGCA 720
1446 AUGCCUGC CUGAUGAG X CGAA AGUCGGGC 193 GCCCGACT A GCAGGCAT 721
1463 CCCUUACC CUGAUGAG X CGAA ACCGCGGC 194 GCCGCGGT A GGTAAGGG 722
1467 GCGGCCCU CUGAUGAG X CGAA ACCUACCG 195 CGGTAGGT A AGGGCCGC 723
1486 CGGCUCUC CUGAUGAG X CGAA ACGCGGUC 196 GACCGCGT A GAGAGCCG 724
1511 GCAGAACC CUGAUGAG X CGAA ACGUCCGU 197 ACGGACGT T GGTTCTGC 725
1515 UAGUGCAG CUGAUGAG X CGAA ACCAACGU 198 ACGTTGGT T CTGCACTA 726
1516 UUAGUGCA CUGAUGAG X CGAA AACCAACG 199 CGTTGGTT C TGCACTAA 727
1523 AUGGGUUU CUGAUGAG X CGAA AGUGCAGA 200 TCTGCACT A AAACCCAT 728
1532 CCGGGGAA CUGAUGAG X CGAA AUGGGUUU 201 AAACCCAT C TTCCCCGG 729
1534 AUCCGGGG CUGAUGAG X CGAA AGAUGGGU 202 ACCCATCT T CCCCGGAT 730
1535 CAUCCGGG CUGAUGAG X CGAA AAGAUGGG 203 CCCATCTT c CCCGGATG 731
1549 AGGGGUGA CUGAUGAG X CGAA ACACACAU 204 ATGTGTGT c TCACCCCT 732
1551 UGAGGGGU CUGAUGAG X CGAA AGACACAC 205 GTGTGTCT c ACCCCTCA 733
1558 AAAAGGAU CUGAUGAG X CGAA AGGGGUGA 206 TCACCCCT c ATCCTTTT 734
1561 AGUAAAAG CUGAUGAG X CGAA AUGAGGGG 207 CCCCTCAT c CTTTTACT 735
1564 AAAAGUAA CUGAUGAG X CGAA AGGAUGAG 208 CTCATCCT T TTACTTTT 736
1565 AAAAAGUA CUGAUGAG X CGAA AAGGAUGA 209 TCATCCTT T TACTTTTT 737
1566 CAAAAAGU CUGAUGAG X CGAA AAAGGAUG 210 CATCCTTT T ACTTTTTG 738
1567 GCAAAAAG CUGAUGAG X CGAA AAAAGGAU 211 ATCCTTTT A CTTTTTGC 739
1570 GGGGCAAA CUGAUGAG X CGAA AGUAAAAG 212 CTTTTACT T TTTGCCCC 740
1571 AGGGGCAA CUGAUGAG X CGAA AAGUAAAA 213 TTTTACTT T TTGCCCCT 741
1572 AAGGGGCA CUGAUGAG X CGAA AAAGUAAA 214 TTTACTTT T TGCCCCTT 742
1573 GAAGGGGC CUGAUGAG X CGAA AAAAGUAA 215 TTACTTTT T GCCCCTTC 743
1580 CAAAGUGG CUGAUGAG X CGAA AGGGGCAA 216 TTGCCCCT T CCACTTTG 744
1581 UCAAAGUG CUGAUGAG X CGAA AAGGGGCA 217 TGCCCCTT c CACTTTGA 745
1586 GGUACUCA CUGAUGAG X CGAA AGUGGAAG 218 CTTCCACT T TGAGTACC 746
1587 UGGUACUC CUGAUGAG X CGAA AAGUGGAA 219 TTCCACTT T GAGTACCA 747
1592 GGAUUUGG CUGAUGAG X CGAA ACUCAAAG 220 CTTTGAGT A CCAAATCC 748
1599 GGCUUGUG CUGAUGAG X CGAA AUUUGGUA 221 TACCAAAT C CACAAGCC 749
1610 CCUCAAAA CUGAUGAG X CGAA AUGGCUUG 222 CAAGCCAT T TTTTGAGG 750
1611 UCCUCAAA CUGAUGAG X CGAA AAUGGCUU 223 AAGCCATT T TTTGAGGA 751
1612 CUCCUCAA CUGAUGAG X CGAA AAAUGGCU 224 AGCCATTT T TTGAGGAG 752
1613 UCUCCUCA CUGAUGAG X CGAA AAAAUGGC 225 GCCATTTT T TGAGGAGA 753
1614 CUCUCCUC CUGAUGAG X CGAA AAAAAUGG 226 CCATTTTT T GAGGAGAG 754
1634 CAGCAUGG CUGAUGAG X CGAA ACUCUCUU 227 AAGAGAGT A CCATGCTG 755
1665 GACGGGUG CUGAUGAG X CGAA AGGCCCCU 228 AGGGGCCT A CACCCGTC 756
1673 AGCCCCAA CUGAUGAG X CGAA ACGGGUGU 229 ACACCCGT C TTGGGGCT 757
1675 CGAGCCCC CUGAUGAG X CGAA AGACGGGU 230 ACCCGTCT T GGGGCTCG 758
1682 GGUGGGGC CUGAUGAG X CGAA AGCCCCAA 231 TTGGGGCT C GCCCCACC 759
1698 CCAGGAGG CUGAUGAG X CGAA AGCCCUGG 232 CCAGGGCT C CCTCCTGG 760
Table 3
1702 UGCUCCAG CUGAUGAG X CGAA AGGGAGCC 233 GGCTCCCT C CTGGAGCA 761
1712 CCGCCUGG CUGAUGAG X CGAA AUGCUCCA 234 TGGAGCAT C CCAGGCGG 762
1746 GCAGAUUC CUGAUGAG X CGAA AGGGGGGG 235 CCCCCCCT T GAATCTGC 763
1751 UCCCUGCA CUGAUGAG X CGAA AUUCAAGG 236 CCTTGAAT C TGCAGGGA 764
1766 GGAGUGGA CUGAUGAG X CGAA AGUUGCUC 237 GAGCAACT C TCCACTCC 765
1768 AUGGAGUG CUGAUGAG X CGAA AGAGUUGC 238 GCAACTCT C CACTCCAT 766
1773 UAAAUAUG CUGAUGAG X CGAA AGUGGAGA 239 TCTCCACT C CATATTTA 767
1777 UAAAUAAA CUGAUGAG X CGAA AUGGAGUG 240 CACTCCAT A TTTATTTA 768
1779 UUUAAAUA CUGAUGAG X CGAA AUAUGGAG 241 CTCCATAT T TATTTAAA 769
1780 GUUUAAAU CUGAUGAG X CGAA AAUAUGGA 242 TCCATATT T ATTTAAAC 770
1781 UGUUUAAA CUGAUGAG X CGAA AAAUAUGG 243 CCATATTT A TTTAAACA 771
1783 AUUGUUUA CUGAUGAG X CGAA AUAAAUAU 244 ATATTTAT T TAAACAAT 772
1784 AAUUGUUU CUGAUGAG X CGAA AAUAAAUA 245 TATTTATT T AAACAATT 773
1785 AAAUUGUU CUGAUGAG X CGAA AAAUAAAU 246 ATTTATTT A AACAATTT 774
1792 GGGGAAAA CUGAUGAG X CGAA AUUGUUUA 247 TAAACAAT T TTTTCCCC 775
1793 UGGGGAAA CUGAUGAG X CGAA AAUUGUUU 248 AAACAATT T TTTCCCCA 776
1794 UUGGGGAA CUGAUGAG X CGAA AAAUUGUU 249 AACAATTT T TTCCCCAA 777
1795 UUUGGGGA CUGAUGAG X CGAA AAAAUUGU 250 ACAATTTT T TCCCCAAA 778
1796 CUUUGGGG CUGAUGAG X CGAA AAAAAUUG 251 CAATTTTT T CCCCAAAG 779
1797 CCUUUGGG CUGAUGAG X CGAA AAAAAAUU 252 AATTTTTT C CCCAAAGG 780
1809 GCACUAUG CUGAUGAG X CGAA AUGCCUUU 253 AAAGGCAT C CATAGTGC 781
1813 UAGUGCAC CUGAUGAG X CGAA AUGGAUGC 254 GCATCCAT A GTGCACTA 782
1821 GAAAAUGC CUGAUGAG X CGAA AGUGCACU 255 AGTGCACT A GCATTTTC 783
1826 UUCAAGAA CUGAUGAG X CGAA AUGCUAGU 256 ACTAGCAT T TTCTTGAA 784
1827 GUUCAAGA CUGAUGAG X CGAA AAUGCUAG 257 CTAGCATT T TCTTGAAC 785
1828 GGUUCAAG CUGAUGAG X CGAA AAAUGCUA 258 TAGCATTT T CTTGAACC 786
1829 UGGUUCAA CUGAUGAG X CGAA AAAAUGCU 259 AGCATTTT C TTGAACCA 787
1831 AUUGGUUC CUGAUGAG X CGAA AGAAAAUG 260 CATTTTCT T GAACCAAT 788
1840 UAAUACAU CUGAUGAG X CGAA AUUGGUUC 261 GAACCAAT A ATGTATTA 789
1845 AAUUUUAA CUGAUGAG X CGAA ACAUUAUU 262 AATAATGT A TTAAAATT 790
1847 AAAAUUUU CUGAUGAG X CGAA AUACAUUA 263 TAATGTAT T AAAATTTT 791
1848 AAAAAUUU CUGAUGAG X CGAA AAUACAUU 264 AATGTATT A AAATTTTT 792
1853 CAUCAAAA CUGAUGAG X CGAA AUUUUAAU 265 ATTAAAAT T TTTTGATG 793
1854 ACAUCAAA CUGAUGAG X CGAA AAUUUUAA 266 TTAAAATT T TTTGATGT 794
1855 GACAUCAA CUGAUGAG X CGAA AAAUUUUA 267 TAAAATTT T TTGATGTC 795
1856 UGACAUCA CUGAUGAG X CGAA AAAAUUUU 268 AAAATTTT T TGATGTCA 796
1857 CUGACAUC CUGAUGAG X CGAA AAAAAUUU 269 AAATTTTT T GATGTCAG 797
1863 GCAAGGCU CUGAUGAG X CGAA ACAUCAAA 270 TTTGATGT C AGCCTTGC 798
1869 CUUGAUGC CUGAUGAG X CGAA AGGCUGAC 271 GTCAGCCT T GCATCAAG 799
1874 AAGCCCUU CUGAUGAG X CGAA AUGCAAGG 272 CCTTGCAT c AAGGGCTT 800
1882 UUUUGAUA CUGAUGAG X CGAA AGCCCUUG 273 CAAGGGCT T TATCAAAA 801
1883 UUUUUGAU CUGAUGAG X CGAA AAGCCCUU 274 AAGGGCTT T ATCAAAAA 802
1884 CUUUUUGA CUGAUGAG X CGAA AAAGCCCU 275 AGGGCTTT A TCAAAAAG 803
1886 UACUUUUU CUGAUGAG X CGAA AUAAAGCC 276 GGCTTTAT C AAAAAGTA 804
1894 UAUUAUUG CUGAUGAG X CGAA ACUUUUUG 277 CAAAAAGT A CAATAATA 805
1899 GGAUUUAU CUGAUGAG X CGAA AUUGUACU 278 AGTACAAT A ATAAATCC 806
1902 UGAGGAUU CUGAUGAG X CGAA AUUAUUGU 279 ACAATAAT A AATCCTCA 807
Table 3
1906 UACCUGAG CUGAUGAG X CGAA AUUUAUUA 280 TAATAAAT C CTCAGGTA 808
1909 UACUACCU CUGAUGAG X CGAA AGGAUUUA 281 TAAATCCT C AGGTAGTA 809
1914 CCCAGUAC CUGAUGAG X CGAA ACCUGAGG 282 CCTCAGGT A GTACTGGG 810
1917 AUUCCCAG CUGAUGAG X CGAA ACUACCUG 283 CAGGTAGT A CTGGGAAT 811
1934 CCAUGGCA CUGAUGAG X CGAA AGCCUUCC 284 GGAAGGCT T TGCCATGG 812
1935 CCCAUGGC CUGAUGAG X CGAA AAGCCUUC 285 GAAGGCTT T GCCATGGG 813
1954 ACUGGUCU CUGAUGAG X CGAA ACGCAGCA 286 TGCTGCGT C AGACCAGT 814
1963 CUUCCCAG CUGAUGAG X CGAA ACUGGUCU 287 AGACCAGT A CTGGGAAG 815
1981 CUGCUUAC CUGAUGAG X CGAA ACCGUCCU 288 AGGACGGT T GTAAGCAG 816
1984 CAACUGCU CUGAUGAG X CGAA ACAACCGU 289 ACGGTTGT A AGCAGTTG 817
1991 UAAAUAAC CUGAUGAG X CGAA ACUGCUUA 290 TAAGCAGT T GTTATTTA 818
1994 CACUAAAU CUGAUGAG X CGAA ACAACUGC 291 GCAGTTGT T ATTTAGTG 819
1995 UCACUAAA CUGAUGAG X CGAA AACAACUG 292 CAGTTGTT A TTTAGTGA 820
1997 UAUCACUA CUGAUGAG X CGAA AUAACAAC 293 GTTGTTAT T TAGTGATA 821
1998 AUAUCACU CUGAUGAG X CGAA AAUAACAA 294 TTGTTATT T AGTGATAT 822
1999 AAUAUCAC CUGAUGAG X CGAA AAAUAACA 295 TGTTATTT A GTGATATT 823
2005 ACCCACAA CUGAUGAG X CGAA AUCACUAA 296 TTAGTGAT A TTGTGGGT 824
2007 UUACCCAC CUGAUGAG X CGAA AUAUCACU 297 AGTGATAT T GTGGGTAA 825
2014 UCUCACGU CUGAUGAG X CGAA ACCCACAA 298 TTGTGGGT A ACGTGAGA 826
2027 CAUUGUUC CUGAUGAG X CGAA AUCUUCUC 299 GAGAAGAT A GAACAATG 827
2038 AUAUAUUA CUGAUGAG X CGAA AGCAUUGU 300 ACAATGCT A TAATATAT 828
2040 UUAUAUAU CUGAUGAG X CGAA AUAGCAUU 301 AATGCTAT A ATATATAA 829
2043 UCAUUAUA CUGAUGAG X CGAA AUUAUAGC 302 GCTATAAT A TATAATGA 830
2045 GUUCAUUA CUGAUGAG X CGAA AUAUUAUA 303 TATAATAT A TAATGAAC 831
2047 GUGUUCAU CUGAUGAG X CGAA AUAUAUUA 304 TAATATAT A ATGAACAC 832
2062 UUAUUAAA CUGAUGAG X CGAA ACCCACGU 305 ACGTGGGT A TTTAATAA 833
2064 UCUUAUUA CUGAUGAG X CGAA AUACCCAC 306 GTGGGTAT T TAATAAGA 834
2065 UUCUUAUU CUGAUGAG X CGAA AAUACCCA 307 TGGGTATT T AATAAGAA 835
2066 UUUCUUAU CUGAUGAG X CGAA AAAUACCC 308 GGGTATTT A ATAAGAAA 836
2069 AUGUUUCU CUGAUGAG X CGAA AUUAAAUA 309 TATTTAAT A AGAAACAT 837
2088 GACAAAGU CUGAUGAG X CGAA AUCUCACA 310 TGTGAGAT T ACTTTGTC 838
2089 GGACAAAG CUGAUGAG X CGAA AAUCUCAC 311 GTGAGATT A CTTTGTCC 839
2092 GCGGGACA CUGAUGAG X CGAA AGUAAUCU 312 AGATTACT T TGTCCCGC 840
2093 AGCGGGAC CUGAUGAG X CGAA AAGUAAUC 313 GATTACTT T GTCCCGCT 841
2096 AUAAGCGG CUGAUGAG X CGAA ACAAAGUA 314 TACTTTGT C CCGCTTAT 842
2102 AGCAGAAU CUGAUGAG X CGAA AGCGGGAC 315 GTCCCGCT T ATTCTGCT 843
2103 GAGCAGAA CUGAUGAG X CGAA AAGCGGGA 316 TCCCGCTT A TTCTGCTC 844
2105 GGGAGCAG CUGAUGAG X CGAA AUAAGCGG 317 CCGCTTAT T CTGCTCCC 845
2106 AGGGAGCA CUGAUGAG X CGAA AAUAAGCG 318 CGCTTATT C TGCTCCCT 846
2111 AUAACAGG CUGAUGAG X CGAA AGCAGAAU 319 ATTCTGCT C CCTGTTAT 847
2117 UAGCAGAU CUGAUGAG X CGAA ACAGGGAG 320 CTCCCTGT T ATCTGCTA 848
2118 CUAGCAGA CUGAUGAG X CGAA AACAGGGA 321 TCCCTGTT A TCTGCTAG 849
2120 AUCUAGCA CUGAUGAG X CGAA AUAACAGG 322 CCTGTTAT C TGCTAGAT 850
2125 ACUAGAUC CUGAUGAG X CGAA AGCAGAUA 323 TATCTGCT A GATCTAGT 851
2129 GAGAACUA CUGAUGAG X CGAA AUCUAGCA 324 TGCTAGAT C TAGTTCTC 852
2131 UUGAGAAC CUGAUGAG X CGAA AGAUCUAG 325 CTAGATCT A GTTCTCAA 853
2134 UGAUUGAG CUGAUGAG X CGAA ACUAGAUC 326 GATCTAGT T CTCAATCA 854
Table 3
2135 GUGAUUGA CUGAUGAG X CGAA AACUAGAU 327 ATCTAGTT C TCAATCAC 855
2137 CAGUGAUU CUGAUGAG X CGAA AGAACUAG 328 CTAGTTCT C AATCACTG 856
2141 GGAGCAGU CUGAUGAG X CGAA AUUGAGAA 329 TTCTCAAT C ACTGCTCC 857
2148 ACACGGGG CUGAUGAG X CGAA AGCAGUGA 330 TCACTGCT C CCCCGTGT 858
2159 CAUUCUAA CUGAUGAG X CGAA ACACACGG 331 CCGTGTGT A TTAGAATG 859
2161 UGCAUUCU CUGAUGAG X CGAA AUACACAC 332 GTGTGTAT T AGAATGCA 860
2162 AUGCAUUC CUGAUGAG X CGAA AAUACACA 333 TGTGTATT A GAATGCAT 861
2173 GAAGACCU CUGAUGAG X CGAA ACAUGCAU 334 ATGCATGT A AGGTCTTC 862
2178 CACAAGAA CUGAUGAG X CGAA ACCUUACA 335 TGTAAGGT C TTCTTGTG 863
2180 GACACAAG CUGAUGAG X CGAA AGACCUUA 336 TAAGGTCT T CTTGTGTC 864
2181 GGACACAA CUGAUGAG X CGAA AAGACCUU 337 AAGGTCTT C TTGTGTCC 865
2183 CAGGACAC CUGAUGAG X CGAA AGAAGACC 338 GGTCTTCT T GTGTCCTG 866
2188 UUCAUCAG CUGAUGAG X CGAA ACACAAGA 339 TCTTGTGT C CTGATGAA 867
2201 CAAGCACA CUGAUGAG X CGAA AUUUUUCA 340 TGAAAAAT A TGTGCTTG 868
2208 CUCAUUUC CUGAUGAG X CGAA AGCACAUA 341 TATGTGCT T GAAATGAG 869
2222 AGAGAUCA CUGAUGAG X CGAA AGUUUCUC 342 GAGAAACT T TGATCTCT 870
2223 CAGAGAUC CUGAUGAG X CGAA AAGUUUCU 343 AGAAACTT T GATCTCTG 871
2227 UAAGCAGA CUGAUGAG X CGAA AUCAAAGU 344 ACTTTGAT c TCTGCTTA 872
2229 AGUAAGCA CUGAUGAG X CGAA AGAUCAAA 345 TTTGATCT c TGCTTACT 873
2234 ACAUUAGU CUGAUGAG X CGAA AGCAGAGA 346 TCTCTGCT T ACTAATGT 874
2235 CACAUUAG CUGAUGAG X CGAA AAGCAGAG 347 CTCTGCTT A CTAATGTG 875
2238 GGGCACAU CUGAUGAG X CGAA AGUAAGCA 348 TGCTTACT A ATGTGCCC 876
2252 UGGACUUG CUGAUGAG X CGAA ACAUGGGG 349 CCCCATGT C CAAGTCCA 877
2258 GCAGGUUG CUGAUGAG X CGAA ACUUGGAC 350 GTCCAAGT C CAACCTGC 878
2283 CAUGUAAU CUGAUGAG X CGAA AUCAGGUC 351 GACCTGAT C ATTACATG 879
2286 AGCCAUGU CUGAUGAG X CGAA AUGAUCAG 352 CTGATCAT T ACATGGCT 880
2287 CAGCCAUG CUGAUGAG X CGAA AAUGAUCA 353 TGATCATT A CATGGCTG 881
2300 GGCUUAGG CUGAUGAG X CGAA ACCACAGC 354 GCTGTGGT T CCTAAGCC 882
2301 AGGCUUAG CUGAUGAG X CGAA AACCACAG 355 CTGTGGTT c CTAAGCCT 883
2304 AACAGGCU CUGAUGAG X CGAA AGGAACCA 356 TGGTTCCT A AGCCTGTT 884
2312 ACUUCAGC CUGAUGAG X CGAA ACAGGCUU 357 AAGCCTGT T GCTGAAGT 885
2321 GCGACAAU CUGAUGAG X CGAA ACUUCAGC 358 GCTGAAGT C ATTGTCGC 886
2324 UGAGCGAC CUGAUGAG X CGAA AUGACUUC 359 GAAGTCAT T GTCGCTCA 887
2327 UGCUGAGC CUGAUGAG X CGAA ACAAUGAC 360 GTCATTGT C GCTCAGCA 888
2331 CUAUUGCU CUGAUGAG X CGAA AGCGACAA 361 TTGTCGCT c AGCAATAG 889
2338 CUGCACCC CUGAUGAG X CGAA AUUGCUGA 362 TCAGCAAT A GGGTGCAG 890
2348 UCCUGGAA CUGAUGAG X CGAA ACUGCACC 363 GGTGCAGT T TTCCAGGA 891
2349 UUCCUGGA CUGAUGAG X CGAA AACUGCAC 364 GTGCAGTT T TCCAGGAA 892
2350 AUUCCUGG CUGAUGAG X CGAA AAACUGCA 365 TGCAGTTT T CCAGGAAT 893
2351 UAUUCCUG CUGAUGAG X CGAA AAAACUGC 366 GCAGTTTT c CAGGAATA 894
2359 CAAAUGCC CUGAUGAG X CGAA AUUCCUGG 367 CCAGGAAT A GGCATTTG 895
2365 AUUAGGCA CUGAUGAG X CGAA AUGCCUAU 368 ATAGGCAT T TGCCTAAT 896
2366 AAUUAGGC CUGAUGAG X CGAA AAUGCCUA 369 TAGGCATT T GCCTAATT 897
- 2371 CCAGGAAU CUGAUGAG X CGAA AGGCAAAU 370 ATTTGCCT A ATTCCTGG 898
2374 AUGCCAGG CUGAUGAG X CGAA AUUAGGCA 371 TGCCTAAT T CCTGGCAT 899
2375 CAUGCCAG CUGAUGAG X CGAA AAUUAGGC 372 GCCTAATT C CTGGCATG 900
2389 AGUCACUA CUGAUGAG X CGAA AGUGUCAU 373 ATGACACT C TAGTGACT 901
Table 3
2391 GAAGUCAC CUGAUGAG X CGAA AGAGUGUC 374 GACACTCT A GTGACTTC 902
2398 UCACCAGG CUGAUGAG X CGAA AGUCACUA 375 TAGTGACT T CCTGGTGA 903
2399 CUCACCAG CUGAUGAG X CGAA AAGUCACU 376 AGTGACTT C CTGGTGAG 904
2419 UGUACCAG CUGAUGAG X CGAA ACAGGCUG 377 CAGCCTGT C CTGGTACA 905
2425 CCCUGCUG CUGAUGAG X CGAA ACCAGGAC 378 GTCCTGGT A CAGCAGGG 906
2435 UACAGCAA CUGAUGAG X CGAA ACCCUGCU 379 AGCAGGGT C TTGCTGTA 907
2437 GUUACAGC CUGAUGAG X CGAA AGACCCUG 380 CAGGGTCT T GCTGTAAC 908
2443 GUCUGAGU CUGAUGAG X CGAA ACAGCAAG 381 CTTGCTGT A ACTCAGAC 909
2447 GAAUGUCU CUGAUGAG X CGAA AGUUACAG 382 CTGTAACT C AGACATTC 910
2454 ACCCUUGG CUGAUGAG X CGAA AUGUCUGA 383 TCAGACAT T CCAAGGGT 911
2455 UACCCUUG CUGAUGAG X CGAA AAUGUCUG 384 CAGACATT c CAAGGGTA 912
2463 GCUUCCCA CUGAUGAG X CGAA ACCCUUGG 385 CCAAGGGT A TGGGAAGC 913
2475 GGUGUGAA CUGAUGAG X CGAA AUGGCUUC 386 GAAGCCAT A TTCACACC 914
2477 GAGGUGUG CUGAUGAG X CGAA AUAUGGCU 387 AGCCATAT T CACACCTC 915
2478 UGAGGUGU CUGAUGAG X CGAA AAUAUGGC 388 GCCATATT c ACACCTCA 916
2485 CAGAGCGU CUGAUGAG X CGAA AGGUGUGA 389 TCACACCT c ACGCTCTG 917
2491 CAUGUCCA CUGAUGAG X CGAA AGCGUGAG 390 CTCACGCT C TGGACATG 918
2502 CUUCCCUA CUGAUGAG X CGAA AUCAUGUC 391 GACATGAT T TAGGGAAG 919
2503 GCUUCCCU CUGAUGAG X CGAA AAUCAUGU 392 ACATGATT T AGGGAAGC 920
2504 UGCUUCCC CUGAUGAG X CGAA AAAUCAUG 393 CATGATTT A GGGAAGCA 921
2536 UGAUCCCA CUGAUGAG X CGAA AGGUGGGG 394 CCCCACCT T TGGGATCA 922
2537 CUGAUCCC CUGAUGAG X CGAA AAGGUGGG 395 CCCACCTT T GGGATCAG 923
2543 CGGAGGCU CUGAUGAG X CGAA AUCCCAAA 396 TTTGGGAT c AGCCTCCG 924
2549 GAAUGGCG CUGAUGAG X CGAA AGGCUGAU 397 ATCAGCCT c CGCCATTC 925
2556 CGACUUGG CUGAUGAG X CGAA AUGGCGGA 398 TCCGCCAT T CCAAGTCG 926
2557 UCGACUUG CUGAUGAG X CGAA AAUGGCGG 399 CCGCCATT c CAAGTCGA 927
2563 AGAGUGUC CUGAUGAG X CGAA ACUUGGAA 400 TTCCAAGT c GACACTCT 928
2570 CUCAAGAA CUGAUGAG X CGAA AGUGUCGA 401 TCGACACT c TTCTTGAG 929
2572 UGCUCAAG CUGAUGAG X CGAA AGAGUGUC 402 GACACTCT T CTTGAGCA 930
2573 CUGCUCAA CUGAUGAG X CGAA AAGAGUGU 403 ACACTCTT c TTGAGCAG 931
2575 GUCUGCUC CUGAUGAG X CGAA AGAAGAGU 404 ACTCTTCT T GAGCAGAC 932
2590 CUCUUCCA CUGAUGAG X CGAA AUCACGGU 405 ACCGTGAT T TGGAAGAG 933
2591 UCUCUUCC CUGAUGAG X CGAA AAUCACGG 406 CCGTGATT T GGAAGAGA 934
2622 GUUUCAAG CUGAUGAG X CGAA AGUGUGGU 407 ACCACACT T CTTGAAAC 935
2623 UGUUUCAA CUGAUGAG X CGAA AAGUGUGG 408 CCACACTT c TTGAAACA 936
2625 GCUGUUUC CUGAUGAG X CGAA AGAAGUGU 409 ACACTTCT T GAAACAGC 937
2646 GCCUAAAG CUGAUGAG X CGAA ACCGUCAC 410 GTGACGGT c CTTTAGGC 938
2649 GCUGCCUA CUGAUGAG X CGAA AGGACCGU 411 ACGGTCCT T TAGGCAGC 939
2650 GGCUGCCU CUGAUGAG X CGAA AAGGACCG 412 CGGTCCTT T AGGCAGCC 940
2651 AGGCUGCC CUGAUGAG X CGAA AAAGGACC 413 GGTCCTTT A GGCAGCCT 941
2668 GGGACAGA CUGAUGAG X CGAA ACGGCGGC 414 GCCGCCGT c TCTGTCCC 942
2"670 CCGGGACA CUGAUGAG X CGAA AGACGGCG 415 CGCCGTCT c TGTCCCGG 943
2674 UGAACCGG CUGAUGAG X CGAA ACAGAGAC 416 GTCTCTGT c CCGGTTCA 944
2680 GCAAGGUG CUGAUGAG X CGAA ACCGGGAC 417 GTCCCGGT T CACCTTGC 945
2681 GGCAAGGU CUGAUGAG X CGAA AACCGGGA 418 TCCCGGTT c ACCTTGCC 946
2686 CUCUCGGC CUGAUGAG X CGAA AGGUGAAC 419 GTTCACCT T GCCGAGAG 947
2703 GUGGGGCA CUGAUGAG X CGAA ACGCGCCU 420 AGGCGCGT c TGCCCCAC 948
Table 3
2715 CAGGGUUU CUGAUGAG X CGAA AGGGUGGG 421 CCCACCCT C AAACCCTG 949
2741 AGAGUCGU CUGAUGAG X CGAA AGCACCAU 422 ATGGTGCT C ACGACTCT 950
2748 UGCAGGAA CUGAUGAG X CGAA AGUCGUGA 423 TCACGACT C TTCCTGCA 951
2750 UUUGCAGG CUGAUGAG X CGAA AGAGUCGU 424 ACGACTCT T CCTGCAAA 952
2751 CUUUGCAG CUGAUGAG X CGAA AAGAGUCG 425 CGACTCTT C CTGCAAAG 953
2774 UUAAUGUG CUGAUGAG X CGAA AGGUCUUC 426 GAAGACCT C CACATTAA 954
2780 AGCCACUU CUGAUGAG X CGAA AUGUGGAG 427 CTCCACAT T AAGTGGCT 955
2781 AAGCCACU CUGAUGAG X CGAA AAUGUGGA 428 TCCACATT A AGTGGCTT 956
2789 AUGUUAAA CUGAUGAG X CGAA AGCCACUU 429 AAGTGGCT T TTTAACAT 957
2790 CAUGUUAA CUGAUGAG X CGAA AAGCCACU 430 AGTGGCTT T TTAACATG 958
2791 UCAUGUUA CUGAUGAG X CGAA AAAGCCAC 431 GTGGCTTT T TAACATGA 959
2792 UUCAUGUU CUGAUGAG X CGAA AAAAGCCA 432 TGGCTTTT T AACATGAA 960
2793 UUUCAUGU CUGAUGAG X CGAA AAAAAGCC 433 GGCTTTTT A ACATGAAA 961
2816 UCGGGAGC CUGAUGAG X CGAA ACAGCUGC 434 GCAGCTGT A GCTCCCGA 962
2820 UAGCUCGG CUGAUGAG X CGAA AGCUACAG 435 CTGTAGCT C CCGAGCTA 963
2828 CAAGAGAG CUGAUGAG X CGAA AGCUCGGG 436 CCCGAGCT A CTCTCTTG 964
2831 UGGCAAGA CUGAUGAG X CGAA AGUAGCUC 437 GAGCTACT C TCTTGCCA 965
2833 GCUGGCAA CUGAUGAG X CGAA AGAGUAGC 438 GCTACTCT C TTGCCAGC 966
2835 AUGCUGGC CUGAUGAG X CGAA AGAGAGUA 439 TACTCTCT T GCCAGCAT 967
2844 AAUGUGAA CUGAUGAG X CGAA AUGCUGGC 440 GCCAGCAT T TTCACATT 968
2845 AAAUGUGA CUGAUGAG X CGAA AAUGCUGG 441 CCAGCATT T TCACATTT 969
2846 AAAAUGUG CUGAUGAG X CGAA AAAUGCUG 442 CAGCATTT T CACATTTT 970
2847 CAAAAUGU CUGAUGAG X CGAA AAAAUGCU 443 AGCATTTT C ACATTTTG 971
2852 AAAGGCAA CUGAUGAG X CGAA AUGUGAAA 444 TTTCACAT T TTGCCTTT 972
2853 GAAAGGCA CUGAUGAG X CGAA AAUGUGAA 445 TTCACATT T TGCCTTTC 973
2854 AGAAAGGC CUGAUGAG X CGAA AAAUGUGA 446 TCACATTT T GCCTTTCT 974
2859 CCACGAGA CUGAUGAG X CGAA AGGCAAAA 447 TTTTGCCT T TCTCGTGG 975
2860 ACCACGAG CUGAUGAG X CGAA AAGGCAAA 448 TTTGCCTT T CTCGTGGT 976
2861 UACCACGA CUGAUGAG X CGAA AAAGGCAA 449 TTGCCTTT c TCGTGGTA 977
2863 UCUACCAC CUGAUGAG X CGAA AGAAAGGC 450 GCCTTTCT c GTGGTAGA 978
2869 CUGGCUUC CUGAUGAG X CGAA ACCACGAG 451 CTCGTGGT A GAAGCCAG 979
2879 UUUCUCUG CUGAUGAG X CGAA ACUGGCUU 452 AAGCCAGT A CAGAGAAA 980
2889 CACCACAG CUGAUGAG X CGAA AUUUCUCU 453 AGAGAAAT T CTGTGGTG 981
2890 CCACCACA CUGAUGAG X CGAA AAUUUCUC 454 GAGAAATT C TGTGGTGG 982
2905 ACACCUCG CUGAUGAG X CGAA AUGUUCCC 455 GGGAACAT T CGAGGTGT 983
2906 GACACCUC CUGAUGAG X CGAA AAUGUUCC 456 GGAACATT C GAGGTGTC 984
2914 UGCAGGGU CUGAUGAG X CGAA ACACCUCG 457 CGAGGTGT C ACCCTGCA 985
2928 CCUCACCA CUGAUGAG X CGAA AGCUCUGC 458 GCAGAGCT A TGGTGAGG 986
2944 CUAAGCCU CUGAUGAG X CGAA AUCCACAC 459 GTGTGGAT A AGGCTTAG 987
2950 UGGCACCU CUGAUGAG X CGAA AGCCUUAU 460 ATAAGGCT T AGGTGCCA 988
2951 CUGGCACC CUGAUGAG X CGAA AAGCCUUA 461 TAAGGCTT A GGTGCCAG 989
2965 AGAAUGCU CUGAUGAG X CGAA ACAGCCUG 462 CAGGCTGT A AGCATTCT 990
2971 CAGCUCAG CUGAUGAG X CGAA AUGCUUAC 463 GTAAGCAT T CTGAGCTG 991
2972 CCAGCUCA CUGAUGAG X CGAA AAUGCUUA 464 TAAGCATT C TGAGCTGG 992
2983 AAAACAAC CUGAUGAG X CGAA AGCCAGCU 465 AGCTGGCT T GTTGTTTT 993
2986 UUAAAAAC CUGAUGAG X CGAA ACAAGCCA 466 TGGCTTGT T GTTTTTAA 994
2989 GACUUAAA CUGAUGAG X CGAA ACAACAAG 467 CTTGTTGT T TTTAAGTC 995
Table 3
2990 GGACUUAA CUGAUGAG X CGAA AACAACAA 468 TTGTTGTT T TTAAGTCC 996
2991 AGGACUUA CUGAUGAG X CGAA AAACAACA 469 TGTTGTTT T TAAGTCCT 997
2992 CAGGACUU CUGAUGAG X CGAA AAAACAAC 470 GTTGTTTT T AAGTCCTG 998
2993 ACAGGACU CUGAUGAG X CGAA AAAAACAA 471 TTGTTTTT A AGTCCTGT 999
2997 AUAUACAG CUGAUGAG X CGAA ACUUAAAA 472 TTTTAAGT C CTGTATAT 1000
3002 CAUACAUA CUGAUGAG X CGAA ACAGGACU 473 AGTCCTGT A TATGTATG 1001
3004 UACAUACA CUGAUGAG X CGAA AUACAGGA 474 TCCTGTAT A TGTATGTA 1002
3008 CUACUACA CUGAUGAG X CGAA ACAUAUAC 475 GTATATGT A TGTAGTAG 1003
3012 CAAACUAC CUGAUGAG X CGAA ACAUACAU 476 ATGTATGT A GTAGTTTG 1004
3015 ACCCAAAC CUGAUGAG X CGAA ACUACAUA 477 TATGTAGT A GTTTGGGT 1005
3018 CACACCCA CUGAUGAG X CGAA ACUACUAC 478 GTAGTAGT T TGGGTGTG 1006
3019 ACACACCC CUGAUGAG X CGAA AACUACUA 479 TAGTAGTT T GGGTGTGT 1007
3028 ACUAUAUA CUGAUGAG X CGAA ACACACCC 480 GGGTGTGT A TATATAGT 1008
3030 CUACUAUA CUGAUGAG X CGAA AUACACAC 481 GTGTGTAT A TATAGTAG 1009
3032 UGCUACUA CUGAUGAG X CGAA AUAUACAC 482 GTGTATAT A TAGTAGCA 1010
3034 AAUGCUAC CUGAUGAG X CGAA AUAUAUAC 483 GTATATAT A GTAGCATT 1011
3037 UGAAAUGC CUGAUGAG X CGAA ACUAUAUA 484 TATATAGT A GCATTTCA 1012
3042 CAUUUUGA CUGAUGAG X CGAA AUGCUACU 485 AGTAGCAT T TCAAAATG 1013
3043 CCAUUUUG CUGAUGAG X CGAA AAUGCUAC 486 GTAGCATT T CAAAATGG 1014
3044 UCCAUUUU CUGAUGAG X CGAA AAAUGCUA 487 TAGCATTT c AAAATGGA 1015
3056 UAAACCAG CUGAUGAG X CGAA ACGUCCAU 488 ATGGACGT A CTGGTTTA 1016
3062 GGAGGUUA CUGAUGAG X CGAA ACCAGUAC 489 GTACTGGT T TAACCTCC 1017
3063 AGGAGGUU CUGAUGAG X CGAA AACCAGUA 490 TACTGGTT T AACCTCCT 1018
3064 UAGGAGGU CUGAUGAG X CGAA AAACCAGU 491 ACTGGTTT A ACCTCCTA 1019
3069 AAGGAUAG CUGAUGAG X CGAA AGGUUAAA 492 TTTAACCT C CTATCCTT 1020
3072 UCCAAGGA CUGAUGAG X CGAA AGGAGGUU 493 AACCTCCT A TCCTTGGA 1021
3074 UCUCCAAG CUGAUGAG X CGAA AUAGGAGG 494 CCTCCTAT C CTTGGAGA 1022
3077 UGCUCUCC CUGAUGAG X CGAA AGGAUAGG 495 CCTATCCT T GGAGAGCA 1023
3093 AAGGUGGA CUGAUGAG X CGAA AGCCAGCU 496 AGCTGGCT C TCCACCTT 1024
3095 ACAAGGUG CUGAUGAG X CGAA AGAGCCAG 497 CTGGCTCT C CACCTTGT 1025
3101 UGUGUAAC CUGAUGAG X CGAA AGGUGGAG 498 CTCCACCT T GTTACACA 1026
3104 UAAUGUGU CUGAUGAG X CGAA ACAAGGUG 499 CACCTTGT T ACACATTA 1027
3105 AUAAUGUG CUGAUGAG X CGAA AACAAGGU 500 ACCTTGTT A CACATTAT 1028
3111 UCUAACAU CUGAUGAG X CGAA AUGUGUAA 501 TTACACAT T ATGTTAGA 1029
3112 CUCUAACA CUGAUGAG X CGAA AAUGUGUA 502 TACACATT A TGTTAGAG 1030
3116 ACCUCUCU CUGAUGAG X CGAA ACAUAAUG 503 CATTATGT T AGAGAGGT 1031
3117 UACCUCUC CUGAUGAG X CGAA AACAUAAU 504 ATTATGTT A GAGAGGTA 1032
3125 CAGCUCGC CUGAUGAG X CGAA ACCUCUCU 505 AGAGAGGT A GCGAGCTG 1033
3136 ACAUAGCA CUGAUGAG X CGAA AGCAGCUC 506 GAGCTGCT C TGCTATGT 1034
3141 UAAGGACA CUGAUGAG X CGAA AGCAGAGC 507 GCTCTGCT A TGTCCTTA 1035
3145 GGCUUAAG CUGAUGAG X CGAA ACAUAGCA 508 TGCTATGT C CTTAAGCC 1036
3148 AUUGGCUU CUGAUGAG X CGAA AGGACAUA 509 TATGTCCT T AAGCCAAT 1037
3149 UAUUGGCU CUGAUGAG X CGAA AAGGACAU 510 ATGTCCTT A AGCCAATA 1038
3157 UGAGUAAA CUGAUGAG X CGAA AUUGGCUU 511 AAGCCAAT A TTTACTCA 1039
3159 GAUGAGUA CUGAUGAG X CGAA AUAUUGGC 512 GCCAATAT T TACTCATC 1040
3160 UGAUGAGU CUGAUGAG X CGAA AAUAUUGG 513 CCAATATT T ACTCATCA 1041
3161 CUGAUGAG CUGAUGAG X CGAA AAAUAUUG 514 CAATATTT A CTCATCAG 1042
Table 3
3164 GACCUGAU CUGAUGAG X CGAA AGUAAAUA 515 TATTTACT C ATCAGGTC 1043
3167 AAUGACCU CUGAUGAG X CGAA AUGAGUAA 516 TTACTCAT C AGGTCATT 1044
3172 AAAAUAAU CUGAUGAG X CGAA ACCUGAUG 517 CATCAGGT C ATTATTTT 1045
3175 UAAAAAAU CUGAUGAG X CGAA AUGACCUG 518 CAGGTCAT T ATTTTTTA 1046
3176 GUAAAAAA CUGAUGAG X CGAA AAUGACCU 519 AGGTCATT A TTTTTTAC 1047
3178 UUGUAAAA CUGAUGAG X CGAA AUAAUGAC 520 GTCATTAT T TTTTACAA 1048
3179 AUUGUAAA CUGAUGAG X CGAA AAUAAUGA 521 TCATTATT T TTTACAAT 1049
3180 CAUUGUAA CUGAUGAG X CGAA AAAUAAUG 522 CATTATTT T TTACAATG 1050
3181 CCAUUGUA CUGAUGAG X CGAA AAAAUAAU 523 ATTATTTT T TACAATGG 1051
3182 GCCAUUGU CUGAUGAG X CGAA AAAAAUAA 524 TTATTTTT T ACAATGGC 1052
3183 GGCCAUUG CUGAUGAG X CGAA AAAAAAUA 525 TATTTTTT A CAATGGCC 1053
3199 AAAUGGUU CUGAUGAG X CGAA AUUCCAUG 525 CATGGAAT A AACCATTT 1054
320S UUUGUAAA CUGAUGAG X CGAA AUGGUUUA 527 TAAACCAT T TTTACAAA 1055
3207 UUUUGUAA CUGAUGAG X CGAA AAUGGUUU 528 AAACCATT T TTACAAAA 1056
Input Sequence PTPNl (Homo sapiens protein tyrosine phosphatase, non-receptor type 1 (PTPNl) 3215 bp)
Cut Site UH.
Table 4
Table 4: Human PTP-IB NCH Ribozyme and Target Sequence
Nt. Ribozyme Sequence Seq. ID Substrate Sequence Seq. ID Position Nos. Nos.
13 CCGCUCUA CUGAUGAG X CGAA ICCGCGUC 1057 GACGCGGC C TAGAGCGG 1781
14 GCCGCUCU CUGAUGAG X CGAA IGCCGCGU 1058 ACGCGGCC T AGAGCGGC 1782
23 GCGCCGUC CUGAUGAG X CGAA ICCGCUCU 1059 AGAGCGGC A GACGGCGC 1783
32 CGGCCCAC CUGAUGAG X CGAA ICGCCGUC 1060 GACGGCGC A GTGGGCCG 1784
39 UCCUUCUC CUGAUGAG X CGAA ICCCACUG 1061 CAGTGGGC C GAGAAGGA 1785
53 GCGGCUGC CUGAUGAG X CGAA ICGCCUCC 1062 GGAGGCGC A GCAGCCGC 1786
56 AGGGCGGC CUGAUGAG X CGAA ICUGCGCC 1063 GGCGCAGC A GCCGCCCT 1787
59 GCCAGGGC CUGAUGAG X CGAA ICUGCUGC 1064 GCAGCAGC C GCCCTGGC 1788
62 CGGGCCAG CUGAUGAG X CGAA ICGGCUGC 1065 GCAGCCGC C CTGGCCCG 1789
63 ACGGGCCA CUGAUGAG X CGAA IGCGGCUG 1066 CAGCCGCC C TGGCCCGT 1790
64 GACGGGCC CUGAUGAG X CGAA IGGCGGCU 1067 AGCCGCCC T GGCCCGTC 1791
68 CCAUGACG CUGAUGAG X CGAA ICCAGGGC 1068 GCCCTGGC C CGTCATGG 1792
69 UCCAUGAC CUGAUGAG X CGAA IGCCAGGG 1069 CCCTGGCC C GTCATGGA 1793
73 CAUCUCCA CUGAUGAG X CGAA IACGGGCC 1070 GGCCCGTC A TGGAGATG 1794
98 UGUCGAUC CUGAUGAG X CGAA ICUCGAAC 1071 GTTCGAGC A GATCGACA 1795
106 CCCGGACU CUGAUGAG X CGAA IUCGAUCU 1072 AGATCGAC A AGTCCGGG 1796
111 CAGCUCCC CUGAUGAG X CGAA IACUUGUC 1073 GACAAGTC C GGGAGCTG 1797
118 GGCCGCCC CUGAUGAG X CGAA ICUCCCGG 1074 CCGGGAGC T GGGCGGCC 1798
126 UGGUAAAU CUGAUGAG X CGAA ICCGCCCA 1075 TGGGCGGC C ATTTACCA 1799
127 CUGGUAAA CUGAUGAG X CGAA IGCCGCCC 1076 GGGCGGCC A TTTACCAG 1800
133 GAUAUCCU CUGAUGAG X CGAA IUAAAUGG 1077 CCATTTAC C AGGATATC 1801
134 GGAUAUCC CUGAUGAG X CGAA IGUAAAUG 1078 CATTTACC A GGATATCC 1802
142 UUCAUGUC CUGAUGAG X CGAA IAUAUCCU 1079 AGGATATC C GACATGAA 1803
146 UGGCUUCA CUGAUGAG X CGAA IUCGGAUA 1080 TATCCGAC A TGAAGCCA 1804
153 AAGUCACU CUGAUGAG X CGAA ICUUCAUG 1081 CATGAAGC C AGTGACTT 1805
154 GAAGUCAC CUGAUGAG X CGAA IGCUUCAU 1082 ATGAAGCC A GTGACTTC 1806
160 ACAUGGGA CUGAUGAG X CGAA IUCACUGG 1083 CCAGTGAC T TCCCATGT 1807
163 UCUACAUG CUGAUGAG X CGAA IAAGUCAC 1084 GTGACTTC C CATGTAGA 1808
164 CUCUACAU CUGAUGAG X CGAA IGAAGUCA 1085 TGACTTCC C ATGTAGAG 1809
165 ACUCUACA CUGAUGAG X CGAA IGGAAGUC 1086 GACTTCCC A TGTAGAGT 1810
177 GGAAGCUU CUGAUGAG X CGAA ICCACUCU 1087 AGAGTGGC C AAGCTTCC 1811
178 AGGAAGCU CUGAUGAG X CGAA IGCCACUC 1088 GAGTGGCC A AGCTTCCT 1812
182 UCUUAGGA CUGAUGAG X CGAA ICUUGGCC 1089 GGCCAAGC T TCCTAAGA 1813
185 UGUUCUUA CUGAUGAG X CGAA IAAGCUUG 1090 CAAGCTTC C TAAGAACA 1814
186 UUGUUCUU CUGAUGAG X CGAA IGAAGCUU 1091 AAGCTTCC T AAGAACAA 1815
193 UCGGUUUU CUGAUGAG X CGAA IUUCUUAG 1092 CTAAGAAC A AAAACCGA 1816
199 CCUAUUUC CUGAUGAG X CGAA IUUUUUGU 1093 ACAAAAAC C GAAATAGG 1817
211 GACGUCUC CUGAUGAG X CGAA IUACCUAU 1094 ATAGGTAC A GAGACGTC 1818
220 AAAGGGAC CUGAUGAG X CGAA IACGUCUC 1095 GAGACGTC A GTCCCTTT 1819
224 GGUCAAAG CUGAUGAG X CGAA IACUGACG 1096 CGTCAGTC C CTTTGACC 1820
225 UGGUCAAA CUGAUGAG X CGAA IGACUGAC 1097 GTCAGTCC C TTTGACCA 1821
226 AUGGUCAA CUGAUGAG X CGAA IGGACUGA 1098 TCAGTCCC T TTGACCAT 1822
232 CCGACUAU CUGAUGAG X CGAA IUCAAAGG 1099 CCTTTGAC C ATAGTCGG 1823
Table 4
233 UCCGACUA CUGAUGAG X CGAA IGUCAAAG 1100 CTTTGACC A TAGTCGGA 1824
248 CUUGAUGU CUGAUGAG X CGAA IUUUAAUC 1101 GATTAAAC T ACATCAAG 1825
251 CUUCUUGA CUGAUGAG X CGAA IUAGUUUA 1102 TAAACTAC A TCAAGAAG 1826
254 UAUCUUCU CUGAUGAG X CGAA IAUGUAGU 1103 ACTACATC A AGAAGATA 1827
268 GUUGAUAU CUGAUGAG X CGAA IUCAUUAU 1104 ATAATGAC T ATATCAAC 1828
274 ACUAGCGU CUGAUGAG X CGAA IAUAUAGU 1105 ACTATATC A ACGCTAGT 1829
279 AUCAAACU CUGAUGAG X CGAA ICGUUGAU 1106 ATCAACGC T AGTTTGAT 1830
303 CUCCUUUG CUGAUGAG X CGAA ICUUCUUC 1107 GAAGAAGC C CAAAGGAG 1831
304 ACUCCUUU CUGAUGAG X CGAA IGCUUCUU 1108 AAGAAGCC C AAAGGAGT 1832
305 AACUCCUU CUGAUGAG X CGAA IGGCUUCU 1109 AGAAGCCC A AAGGAGTT 1833
316 GGUAAGAA CUGAUGAG X CGAA IUAACUCC 1110 GGAGTTAC A TTCTTACC 1834
320 CCUGGGUA CUGAUGAG X CGAA IAAUGUAA llll TTACATTC T TACCCAGG 1835
324 GGGCCCUG CUGAUGAG X CGAA IUAAGAAU 1112 ATTCTTAC c CAGGGCCC 1836
325 AGGGCCCU CUGAUGAG X CGAA IGUAAGAA 1113 TTCTTACC c AGGGCCCT 1837
326 AAGGGCCC CUGAUGAG X CGAA IGGUAAGA 1114 TCTTACCC A GGGCCCTT 1838
331 AGGCAAAG CUGAUGAG X CGAA ICCCUGGG 1115 CCCAGGGC C CTTTGCCT 1839
332 UAGGCAAA CUGAUGAG X CGAA IGCCCUGG 1116 CCAGGGCC C TTTGCCTA 1840
333 UUAGGCAA CUGAUGAG X CGAA IGGCCCUG 1117 CAGGGCCC T TTGCCTAA 1841
338 AUGUGUUA CUGAUGAG X CGAA ICAAAGGG 1118 CCCTTTGC c TAACACAT 1842
339 CAUGUGUU CUGAUGAG X CGAA IGCAAAGG 1119 CCTTTGCC T AACACATG 1843
343 ACCGCAUG CUGAUGAG X CGAA IUUAGGCA 1120 TGCCTAAC A CATGCGGT 1844
345 UGACCGCA CUGAUGAG X CGAA IUGUUAGG 1121 CCTAACAC A TGCGGTCA 1845
353 CCCAAAAG CUGAUGAG X CGAA IACCGCAU 1122 ATGCGGTC A CTTTTGGG 1846
355 CUCCCAAA CUGAUGAG X CGAA IUGACCGC 1123 GCGGTCAC T TTTGGGAG 1847
377 UGCUUUUC CUGAUGAG X CGAA ICUCCCAC 1124 GTGGGAGC A GAAAAGCA 1848
385 GACACCCC CUGAUGAG X CGAA ICUUUUCU 1125 AGAAAAGC A GGGGTGTC 1849
397 GUUGAGCA CUGAUGAG X CGAA IACGACAC 1126 GTGTCGTC A TGCTCAAC 1850
401 CUCUGUUG CUGAUGAG X CGAA ICAUGACG 1127 CGTCATGC T CAACAGAG 1851
403 CACUCUGU CUGAUGAG X CGAA IAGCAUGA 1128 TCATGCTC A ACAGAGTG 1852
406 CAUCACUC CUGAUGAG X CGAA IUUGAGCA 1129 TGCTCAAC A GAGTGATG 1853
438 CAGUAUUG CUGAUGAG X CGAA ICGCAUUU 1130 AAATGCGC A CAATACTG 1854
440 GCCAGUAU CUGAUGAG X CGAA IUGCGCAU 1131 ATGCGCAC A ATACTGGC 1855
445 UUGUGGCC CUGAUGAG X CGAA IUAUUGUG 1132 CACAATAC T GGCCACAA 1856
449 CUUUUUGU CUGAUGAG X CGAA ICCAGUAU 1133 ATACTGGC c ACAAAAAG 1857
450 UCUUUUUG CUGAUGAG X CGAA IGCCAGUA 1134 TACTGGCC A CAAAAAGA 1858
452 CUUCUUUU CUGAUGAG X CGAA IUGGCCAG 1135 CTGGCCAC A AAAAGAAG 1859
475 GUCUUCAA CUGAUGAG X CGAA IAUCAUCU 1136 AGATGATC T TTGAAGAC 1860
484 CAAAUUUG CUGAUGAG X CGAA IUCUUCAA 1137 TTGAAGAC A CAAATTTG 1861
486 UUCAAAUU CUGAUGAG X CGAA IUGUCUUC 1138 GAAGACAC A AATTTGAA 1862
501 GAGAUCAA CUGAUGAG X CGAA IUUAAUUU 1139 AAATTAAC A TTGATCTC 1863
508 AUCUUCAG CUGAUGAG X CGAA IAUCAAUG 1140 CATTGATC T CTGAAGAT 1864
510 AUAUCUUC CUGAUGAG X CGAA lAGAUCAA 1141 TTGATCTC T GAAGATAT 1865
520 AUAUGACU CUGAUGAG X CGAA IAUAUCUU 1142 AAGATATC A AGTCATAT 1866
525 GUAUAAUA CUGAUGAG X CGAA IACUUGAU 1143 ATCAAGTC A TATTATAC 1867
534 UGUCGCAC CUGAUGAG X CGAA IUAUAAUA 1144 TATTATAC A GTGCGACA 1868
542 AUUCUAGC CUGAUGAG X CGAA lUCGCACU 1145 AGTGCGAC A GCTAGAAT 1869
545 CCAAUUCU CUGAUGAG X CGAA ICUGUCGC 1146 GCGACAGC T AGAATTGG 1870
Table 4
559 GGUUGUAA CUGAUGAG X CGAA IUUUUCCA 1147 TGGAAAAC C TTACAACC 1871
560 GGGUUGUA CUGAUGAG X CGAA IGUUUUCC 1148 GGAAAACC T TACAACCC 1872
564 UCUUGGGU CUGAUGAG X CGAA IUAAGGUU 1149 AACCTTAC A ACCCAAGA 1873
567 GUUUCUUG CUGAUGAG X CGAA IUUGUAAG 1150 CTTACAAC C CAAGAAAC 1874
568 AGUUUCUU CUGAUGAG X CGAA IGUUGUAA 1151 TTACAACC C AAGAAACT 1875
569 GAGUUUCU CUGAUGAG X CGAA IGGUUGUA 1152 TACAACCC A AGAAACTC 1876
576 AUCUCUCG CUGAUGAG X CGAA IUUUCUUG 1153 CAAGAAAC T CGAGAGAT 1877
586 GAAAUGUA CUGAUGAG X CGAA IAUCUCUC 1154 GAGAGATC T TACATTTC 1878
590 AGUGGAAA CUGAUGAG X CGAA IUAAGAUC 1155 GATCTTAC A TTTCCACT 1879
595 GGUAUAGU CUGAUGAG X CGAA IAAAUGUA 1156 TACATTTC C ACTATACC 1880
596 UGGUAUAG CUGAUGAG X CGAA IGAAAUGU 1157 ACATTTCC A CTATACCA 1881
598 UGUGGUAU CUGAUGAG X CGAA IUGGAAAU 1158 ATTTCCAC T ATACCACA 1882
603 GGCCAUGU CUGAUGAG X CGAA IUAUAGUG 1159 CACTATAC C ACATGGCC 1883
604 AGGCCAUG CUGAUGAG X CGAA IGUAUAGU 1160 ACTATACC A CATGGCCT 1884
606 UCAGGCCA CUGAUGAG X CGAA IUGGUAUA 1161 TATACCAC A TGGCCTGA 1885
611 CAAAGUCA CUGAUGAG X CGAA ICCAUGUG 1162 CACATGGC C TGACTTTG 1886
612 CCAAAGUC CUGAUGAG X CGAA IGCCAUGU 1163 ACATGGCC T GACTTTGG 1887
616 GACUCCAA CUGAUGAG X CGAA IUCAGGCC 1164 GGCCTGAC T TTGGAGTC 1888
625 UGAUUCAG CUGAUGAG X CGAA IACUCCAA 1165 TTGGAGTC C CTGAATCA 1889 -
626 GUGAUUCA CUGAUGAG X CGAA IGACUCCA 1166 TGGAGTCC C TGAATCAC 1890
627 GGUGAUUC CUGAUGAG X CGAA IGGACUCC 1167 GGAGTCCC T GAATCACC 1891
633 GAGGCUGG CUGAUGAG X CGAA IAUUCAGG 1168 CCTGAATC A CCAGCCTC 1892
635 AUGAGGCU CUGAUGAG X CGAA lUGAUUCA 1169 TGAATCAC C AGCCTCAT 1893
636 AAUGAGGC CUGAUGAG X CGAA IGUGAUUC 1170 GAATCACC A GCCTCATT 1894
639 AAGAAUGA CUGAUGAG X CGAA ICUGGUGA 1171 TCACCAGC C TCATTCTT 1895
640 CAAGAAUG CUGAUGAG X CGAA IGCUGGUG 1172 CACCAGCC T CATTCTTG 1896
642 UUCAAGAA CUGAUGAG X CGAA IAGGCUGG 1173 CCAGCCTC A TTCTTGAA 1897
646 AAAGUUCA CUGAUGAG X CGAA IAAUGAGG 1174 CCTCATTC T TGAACTTT 1898
652 GAAAAGAA CUGAUGAG X CGAA IUUCAAGA 1175 TCTTGAAC T TTCTTTTC 1899
656 CUUUGAAA CUGAUGAG X CGAA IAAAGUUC 1176 GAACTTTC T TTTCAAAG 1900
661 UCGGACUU CUGAUGAG X CGAA IAAAAGAA 1177 TTCTTTTC A AAGTCCGA 1901
667 UGACUCUC CUGAUGAG X CGAA IACUUUGA 1178 TCAAAGTC C GAGAGTCA 1902
675 AGUGACCC CUGAUGAG X CGAA IACUCUCG 1179 CGAGAGTC A GGGTCACT 1903
681 GGGCUGAG CUGAUGAG X CGAA IACCCUGA 1180 TCAGGGTC A CTCAGCCC 1904
683 CCGGGCUG CUGAUGAG X CGAA IUGACCCU 1181 AGGGTCAC T CAGCCCGG 1905
685 CUCCGGGC CUGAUGAG X CGAA IAGUGACC 1182 GGTCACTC A GCCCGGAG 1906
688 GUGCUCCG CUGAUGAG X CGAA ICUGAGUG 1183 CACTCAGC C CGGAGCAC 1907
689 CGUGCUCC CUGAUGAG X CGAA IGCUGAGU 1184 ACTCAGCC C GGAGCACG 1908
695 CGGGCCCG CUGAUGAG X CGAA ICUCCGGG 1185 CCCGGAGC A CGGGCCCG 1909
701 CCACAACG CUGAUGAG X CGAA ICCCGUGC 1186 GCACGGGC C CGTTGTGG 1910
702 ACCACAAC CUGAUGAG X CGAA IGCCCGUG 1187 CACGGGCC C GTTGTGGT 1911
713 CACUGCAG CUGAUGAG X CGAA ICACCACA 1188 TGTGGTGC A CTGCAGTG 1912
715 UGCACUGC CUGAUGAG X CGAA lUGCACCA 1189 TGGTGCAC T GCAGTGCA 1913
718 GCCUGCAC CUGAUGAG X CGAA ICAGUGCA 1190 TGCACTGC A GTGCAGGC 1914
723 CCGAUGCC CUGAUGAG X CGAA ICACUGCA 1191 TGCAGTGC A GGCATCGG 1915
727 CCUGCCGA CUGAUGAG X CGAA ICCUGCAC 1192 GTGCAGGC A TCGGCAGG 1916
733 UCCAGACC CUGAUGAG X CGAA ICCGAUGC 1193 GCATCGGC A GGTCTGGA 1917
Table 4
738 AAGGUUCC CUGAUGAG X CGAA IACCUGCC 1194 GGCAGGTC T GGAACCTT 1918
744 AGACAGAA CUGAUGAG X CGAA lUUCCAGA 1195 TCTGGAAC C TTCTGTCT 1919
745 CAGACAGA CUGAUGAG X CGAA IGUUCCAG 1196 CTGGAACC T TCTGTCTG 1920
748 AGCCAGAC CUGAUGAG X CGAA IAAGGUUC 1197 GAACCTTC T GTCTGGCT 1921
752 UAUCAGCC CUGAUGAG X CGAA IACAGAAG 1198 CTTCTGTC T GGCTGATA 1922
756 CAGGUAUC CUGAUGAG X CGAA ICCAGACA 1199 TGTCTGGC T GATACCTG 1923
762 AAGAGGCA CUGAUGAG X CGAA IUAUCAGC 1200 GCTGATAC C TGCCTCTT 1924
763 CAAGAGGC CUGAUGAG X CGAA IGUAUCAG 1201 CTGATACC T GCCTCTTG 1925
766 CAGCAAGA CUGAUGAG X CGAA ICAGGUAU 1202 ATACCTGC C TCTTGCTG 1926
767 UCAGCAAG CUGAUGAG X CGAA IGCAGGUA 1203 TACCTGCC T CTTGCTGA 1927
769 CAUCAGCA CUGAUGAG X CGAA IAGGCAGG 1204 CCTGCCTC T TGCTGATG 1928
773 UGUCCAUC CUGAUGAG X CGAA ICAAGAGG 1205 CCTCTTGC T GATGGACA 1929
781 UUUCCUCU CUGAUGAG X CGAA lUCCAUCA 1206 TGATGGAC A AGAGGAAA 1930
793 GGAAGAAG CUGAUGAG X CGAA IUCUUUCC 1207 GGAAAGAC C CTTCTTCC 1931
794 CGGAAGAA CUGAUGAG X CGAA IGUCUUUC 1208 GAAAGACC C TTCTTCCG 1932
795 ACGGAAGA CUGAUGAG X CGAA IGGUCUUU 1209 AAAGACCC T TCTTCCGT 1933
798 UCAACGGA CUGAUGAG X CGAA IAAGGGUC 1210 GACCCTTC T TCCGTTGA 1934
801 AUAUCAAC CUGAUGAG X CGAA IAAGAAGG 1211 CCTTCTTC C GTTGATAT 1935
811 CACUUUCU CUGAUGAG X CGAA IAUAUCAA 1212 TTGATATC A AGAAAGTG 1936
821 UUUCUAAC CUGAUGAG X CGAA ICACUUUC 1213 GAAAGTGC T GTTAGAAA 1937
851 UCUGGAUC CUGAUGAG X CGAA ICCCCAUC 1214 GATGGGGC T GATCCAGA 1938
856 GGCUGUCU CUGAUGAG X CGAA IAUCAGCC 1215 GGCTGATC C AGACAGCC 1939
857 CGGCUGUC CUGAUGAG X CGAA IGAUCAGC 1216 GCTGATCC A GACAGCCG 1940
861 UGGUCGGC CUGAUGAG X CGAA IUCUGGAU 1217 ATCCAGAC A GCCGACCA 1941
864 AGCUGGUC CUGAUGAG X CGAA ICUGUCUG 1218 CAGACAGC C GACCAGCT 1942
868 GCGCAGCU CUGAUGAG X CGAA IUCGGCUG 1219 CAGCCGAC C AGCTGCGC 1943
869 AGCGCAGC CUGAUGAG X CGAA IGUCGGCU 1220 AGCCGACC A GCTGCGCT 1944
872 AGAAGCGC CUGAUGAG X CGAA ICUGGUCG 1221 CGACCAGC T GCGCTTCT 1945
877 GUAGGAGA CUGAUGAG X CGAA ICGCAGCU 1222 AGCTGCGC T TCTCCTAC 1946
880 CAGGUAGG CUGAUGAG X CGAA IAAGCGCA 1223 TGCGCTTC T CCTACCTG 1947
882 GCCAGGUA CUGAUGAG X CGAA IAGAAGCG 1224 CGCTTCTC C TACCTGGC 1948
883 AGCCAGGU CUGAUGAG X CGAA IGAGAAGC 1225 GCTTCTCC T ACCTGGCT 1949
886 CACAGCCA CUGAUGAG X CGAA IUAGGAGA 1226 TCTCCTAC C TGGCTGTG 1950
887 UCACAGCC CUGAUGAG X CGAA IGUAGGAG 1227 CTCCTACC T GGCTGTGA 1951
891 UCGAUCAC CUGAUGAG X CGAA ICCAGGUA 1228 TACCTGGC T GTGATCGA 1952
906 AUGAAUUU CUGAUGAG X CGAA ICACCUUC 1229 GAAGGTGC C AAATTCAT 1953
907 GAUGAAUU CUGAUGAG X CGAA IGCACCUU 1230 AAGGTGCC A AATTCATC 1954
913 CCCCAUGA CUGAUGAG X CGAA IAAUUUGG 1231 CCAAATTC A TCATGGGG 1955
916 GUCCCCCA CUGAUGAG X CGAA IAUGAAUU 1232 AATTCATC A TGGGGGAC 1956
925 CACGGAAG CUGAUGAG X CGAA IUCCCCCA 1233 TGGGGGAC T CTTCCGTG 1957
927 UGCACGGA CUGAUGAG X CGAA IAGUCCCC 1234 GGGGACTC T TCCGTGCA 1958
930 UCCUGCAC CUGAUGAG X CGAA IAAGAGUC 1235 GACTCTTC C GTGCAGGA 1959
935 ACUGAUCC CUGAUGAG X CGAA ICACGGAA 1236 TTCCGTGC A GGATCAGT 1960
941 CCUUCCAC CUGAUGAG X CGAA IAUCCUGC 1237 GCAGGATC A GTGGAAGG 1961
953 CGUGGGAA CUGAUGAG X CGAA ICUCCUUC 1238 GAAGGAGC T TTCCCACG 1962
957 UCCUCGUG CUGAUGAG X CGAA IAAAGCUC 1239 GAGCTTTC C CACGAGGA 1963
958 GUCCUCGU CUGAUGAG X CGAA IGAAAGCU 1240 AGCTTTCC C ACGAGGAC 1964
Table 4
959 GGUCCUCG CUGAUGAG X CGAA IGGAAAGC 1241 GCTTTCCC A CGAGGACC 1965
967 GGGCUCCA CUGAUGAG X CGAA IUCCUCGU 1242 ACGAGGAC C TGGAGCCC 1966
968 GGGGCUCC CUGAUGAG X CGAA IGUCCUCG 1243 CGAGGACC T GGAGCCCC 1967
974 CGGGUGGG CUGAUGAG X CGAA ICUCCAGG 1244 CCTGGAGC C CCCACCCG 1968
975 UCGGGUGG CUGAUGAG X CGAA IGCUCCAG 1245 CTGGAGCC C CCACCCGA 1969
976 CUCGGGUG CUGAUGAG X CGAA IGGCUCCA 1246 TGGAGCCC C CACCCGAG 1970
977 GCUCGGGU CUGAUGAG X CGAA IGGGCUCC 1247 GGAGCCCC C ACCCGAGC 1971
978 UGCUCGGG CUGAUGAG X CGAA IGGGGCUC 1248 GAGCCCCC A CCCGAGCA 1972
980 UAUGCUCG CUGAUGAG X CGAA IUGGGGGC 1249 GCCCCCAC C CGAGCATA 1973
981 AUAUGCUC CUGAUGAG X CGAA IGUGGGGG 1250 CCCCCACC C GAGCATAT 1974
986 GGGGGAUA CUGAUGAG X CGAA ICUCGGGU 1251 ACCCGAGC A TATCCCCC 1975
991 AGGUGGGG CUGAUGAG X CGAA lAUAUGCU 1252 AGCATATC C CCCCACCT 1976
992 GAGGUGGG CUGAUGAG X CGAA IGAUAUGC 1253 GCATATCC C CCCACCTC 1977
993 GGAGGUGG CUGAUGAG X CGAA IGGAUAUG 1254 CATATCCC C CCACCTCC 1978
994 GGGAGGUG CUGAUGAG X CGAA IGGGAUAU 1255 ATATCCCC C CACCTCCC 1979
995 GGGGAGGU CUGAUGAG X CGAA IGGGGAUA 1256 TATCCCCC C ACCTCCCC 1980
996 CGGGGAGG CUGAUGAG X CGAA IGGGGGAU 1257 ATCCCCCC A CCTCCCCG 1981
998 GCCGGGGA CUGAUGAG X CGAA IUGGGGGG 1258 CCCCCCAC C TCCCCGGC 1982
999 GGCCGGGG CUGAUGAG X CGAA IGUGGGGG 1259 CCCCCACC T CCCCGGCC 1983
1001 GUGGCCGG CUGAUGAG X CGAA IAGGUGGG 1260 CCCACCTC C CCGGCCAC 1984
1002 GGUGGCCG CUGAUGAG X CGAA IGAGGUGG 1261 CCACCTCC C CGGCCACC 1985
1003 GGGUGGCC CUGAUGAG X CGAA IGGAGGUG 1262 CACCTCCC C GGCCACCC 1986
1007 GUUUGGGU CUGAUGAG X CGAA ICCGGGGA 1263 TCCCCGGC C ACCCAAAC 1987
1008 CGUUUGGG CUGAUGAG X CGAA IGCCGGGG 1264 CCCCGGCC A CCCAAACG 1988
1010 UUCGUUUG CUGAUGAG X CGAA IUGGCCGG 1265 CCGGCCAC C CAAACGAA 1989
1011 AUUCGUUU CUGAUGAG X CGAA IGUGGCCG 1266 CGGCCACC C AAACGAAT 1990
1012 GAUUCGUU CUGAUGAG X CGAA IGGUGGCC 1267 GGCCACCC A AACGAATC 1991
1021 UGGCUCCA CUGAUGAG X CGAA IAUUCGUU 1268 AACGAATC C TGGAGCCA 1992
1022 GUGGCUCC CUGAUGAG X CGAA IGAUUCGU 1269 ACGAATCC T GGAGCCAC 1993
1028 CAUUGUGU CUGAUGAG X CGAA ICUCCAGG 1270 CCTGGAGC C ACACAATG 1994
1029 CCAUUGUG CUGAUGAG X CGAA IGCUCCAG 1271 CTGGAGCC A CACAATGG 1995
1031 UCCCAUUG CUGAUGAG X CGAA IUGGCUCC 1272 GGAGCCAC A CAATGGGA 1996
1033 UUUCCCAU CUGAUGAG X CGAA IUGUGGCU 1273 AGCCACAC A ATGGGAAA 1997
1045 GAACUCCC CUGAUGAG X CGAA ICAUUUCC 1274 GGAAATGC A GGGAGTTC 1998
1054 AUUUGGGA CUGAUGAG X CGAA IAACUCCC 1275 GGGAGTTC T TCCCAAAT 1999
1057 GUGAUUUG CUGAUGAG X CGAA IAAGAACU 1276 AGTTCTTC C CAAATCAC 2000
1058 GGUGAUUU CUGAUGAG X CGAA IGAAGAAC 1277 GTTCTTCC C AAATCACC 2001
1059 UGGUGAUU CUGAUGAG X CGAA IGGAAGAA 1278 TTCTTCCC A AATCACCA 2002
1064 CCCACUGG CUGAUGAG X CGAA IAUUUGGG 1279 CCCAAATC A CCAGTGGG 2003
1066 CACCCACU CUGAUGAG X CGAA IUGAUUUG 1280 CAAATCAC C AGTGGGTG 2004
1067 UCACCCAC CUGAUGAG X CGAA IGUGAUUU 1281 AAATCACC A GTGGGTGA 2005
1086 UCCUCCUG CUGAUGAG X CGAA IUCUCUUC 1282 GAAGAGAC C CAGGAGGA 2006
1087 AUCCUCCU CUGAUGAG X CGAA IGUCUCUU 1283 AAGAGACC C AGGAGGAT 2007
1088 UAUCCUCC CUGAUGAG X CGAA IGGUCUCU 1284 AGAGACCC A GGAGGATA 2008
1102 GAUGGGGC CUGAUGAG X CGAA IUCUUUAU 1285 ATAAAGAC T GCCCCATC 2009
1105 CUUGAUGG CUGAUGAG X CGAA ICAGUCUU 1286 AAGACTGC C CCATCAAG 2010
1106 CCUUGAUG CUGAUGAG X CGAA IGCAGUCU 1287 AGACTGCC C CATCAAGG 2011
Table 4
1107 UCCUUGAU CUGAUGAG X CGAA IGGCAGUC 1288 GACTGCCC C ATCAAGGA 2012
1108 UUCCUUGA CUGAUGAG X CGAA IGGGCAGU 1289 ACTGCCCC A TCAAGGAA 2013 llll UUCUUCCU CUGAUGAG X CGAA IAUGGGGC 1290 GCCCCATC A AGGAAGAA 2014
1129 AUUUAAGG CUGAUGAG X CGAA ICUUCCUU 1291 AAGGAAGC C CCTTAAAT 2015
1130 CAUUUAAG CUGAUGAG X CGAA IGCUUCCU 1292 AGGAAGCC C CTTAAATG 2016
1131 GCAUUUAA CUGAUGAG X CGAA IGGCUUCC 1293 GGAAGCCC C TTAAATGC 2017
1132 GGCAUUUA CUGAUGAG X CGAA IGGGCUUC 1294 GAAGCCCC T TAAATGCC 2018
1140 UAGGGUGC CUGAUGAG X CGAA ICAUUUAA 1295 TTAAATGC C GCACCCTA 2019
1143 CCGUAGGG CUGAUGAG X CGAA ICGGCAUU 1296 AATGCCGC A CCCTACGG 2020
1145 UGCCGUAG CUGAUGAG X CGAA IUGCGGCA 1297 TGCCGCAC C CTACGGCA 2021
1146 AUGCCGUA CUGAUGAG X CGAA IGUGCGGC 1298 GCCGCACC c TACGGCAT 2022
1147 GAUGCCGU CUGAUGAG X CGAA IGGUGCGG 1299 CCGCACCC T ACGGCATC 2023
1153 GCUUUCGA CUGAUGAG X CGAA ICCGUAGG 1300 CCTACGGC A TCGAAAGC 2024
1162 UUGACUCA CUGAUGAG X CGAA ICUUUCGA 1301 TCGAAAGC A TGAGTCAA 2025
1169 CAGUGUCU CUGAUGAG X CGAA IACUCAUG 1302 CATGAGTC A AGACACTG 2026
1174 AACUUCAG CUGAUGAG X CGAA IUCUUGAC 1303 GTCAAGAC A CTGAAGTT 2027
1176 CUAACUUC CUGAUGAG X CGAA IUGUCUUG 1304 CAAGACAC T GAAGTTAG 2028
1208 CACCUCGA CUGAUGAG X CGAA IACUUCCC 1305 GGGAAGTC T TCGAGGTG 2029
1218 GCAGCCUG CUGAUGAG X CGAA ICACCUCG 1306 CGAGGTGC c CAGGCTGC 2030
1219 GGCAGCCU CUGAUGAG X CGAA IGCACCUC 1307 GAGGTGCC C AGGCTGCC 2031
1220 AGGCAGCC CUGAUGAG X CGAA IGGCACCU 1308 AGGTGCCC A GGCTGCCT 2032
1224 GGGGAGGC CUGAUGAG X CGAA ICCUGGGC 1309 GCCCAGGC T GCCTCCCC 2033
1227 GCUGGGGA CUGAUGAG X CGAA ICAGCCUG 1310 CAGGCTGC C TCCCCAGC 2034
1228 GGCUGGGG CUGAUGAG X CGAA IGCAGCCU 1311 AGGCTGCC T CCCCAGCC 2035
1230 UUGGCUGG CUGAUGAG X CGAA IAGGCAGC 1312 GCTGCCTC C CCAGCCAA 2036
1231 UUUGGCUG CUGAUGAG X CGAA IGAGGCAG 1313 CTGCCTCC C CAGCCAAA 2037
1232 CUUUGGCU CUGAUGAG X CGAA IGGAGGCA 1314 TGCCTCCC C AGCCAAAG 2038
1233 CCUUUGGC CUGAUGAG X CGAA IGGGAGGC 1315 GCCTCCCC A GCCAAAGG 2039
1236 UCCCCUUU CUGAUGAG X CGAA ICUGGGGA 1316 TCCCCAGC C AAAGGGGA 2040
1237 CUCCCCUU CUGAUGAG X CGAA IGCUGGGG 1317 CCCCAGCC A AAGGGGAG 2041
1247 GCAGUGAC CUGAUGAG X CGAA ICUCCCCU 1318 AGGGGAGC C GTCACTGC 2042
1251 UCGGGCAG CUGAUGAG X CGAA IACGGCUC 1319 GAGCCGTC A CTGCCCGA 2043
1253 UCUCGGGC CUGAUGAG X CGAA IUGACGGC 1320 GCCGTCAC T GCCCGAGA 2044
1256 CCUUCUCG CUGAUGAG X CGAA ICAGUGAC 1321 GTCACTGC C CGAGAAGG 2045
1257 UCCUUCUC CUGAUGAG X CGAA IGCAGUGA 1322 TCACTGCC C GAGAAGGA 2046
1273 CAGUGCAU CUGAUGAG X CGAA IUCCUCGU 1323 ACGAGGAC C ATGCACTG 2047
1274 UCAGUGCA CUGAUGAG X CGAA IGUCCUCG 1324 CGAGGACC A TGCACTGA 2048
1278 UAACUCAG CUGAUGAG X CGAA ICAUGGUC 1325 GACCATGC A CTGAGTTA 2049
1280 AGUAACUC CUGAUGAG X CGAA IUGCAUGG 1326 CCATGCAC T GAGTTACT 2050
1288 GGGCUUCC CUGAUGA rG X CGAA lUAACUCA 1327 TGAGTTAC T GGAAGCCC 2051
1295 CCAGGAAG CUGAUGAG X CGAA ICUUCCAG 1328 CTGGAAGC C CTTCCTGG 2052
1296 ACCAGGAA CUGAUGAG X CGAA IGCUUCCA 1329 TGGAAGCC C TTCCTGGT 2053
1297 GACCAGGA CUGAUGAG X CGAA IGGCUUCC 1330 GGAAGCCC T TCCTGGTC 2054
1300 GUUGACCA CUGAUGAG X CGAA IAAGGGCU 1331 AGCCCTTC C TGGTCAAC 2055
1301 UGUUGACC CUGAUGAG X CGAA IGAAGGGC 1332 GCCCTTCC T GGTCAACA 2056
1306 GCACAUGU CUGAUGAG X CGAA IACCAGGA 1333 TCCTGGTC A ACATGTGC 2057
1309 CACGCACA CUGAUGAG X CGAA IUUGACCA 1334 TGGTCAAC A TGTGCGTG 2058
Table 4
1320 AGGACCGU CUGAUGAG X CGAA ICCACGCA 1335 TGCGTGGC T ACGGTCCT 2059
1327 GGCCGUGA CUGAUGAG X CGAA IACCGUAG 1336 CTACGGTC C TCACGGCC 2060
1328 CGGCCGUG CUGAUGAG X CGAA IGACCGUA 1337 TACGGTCC T CACGGCCG 2061
1330 GCCGGCCG CUGAUGAG X CGAA IAGGACCG 1338 CGGTCCTC A CGGCCGGC 2062
1335 UAAGCGCC CUGAUGAG X CGAA ICCGUGAG 1339 CTCACGGC C GGCGCTTA 2063
1341 CAGAGGUA CUGAUGAG X CGAA ICGCCGGC 1340 GCCGGCGC T TACCTCTG 2064
1345 GUAGCAGA CUGAUGAG X CGAA IUAAGCGC 1341 GCGCTTAC C TCTGCTAC 2065
1346 UGUAGCAG CUGAUGAG X CGAA IGUAAGCG 1342 CGCTTACC T CTGCTACA 2066
1348 CCUGUAGC CUGAUGAG X CGAA IAGGUAAG 1343 CTTACCTC T GCTACAGG 2067
1351 GAACCUGU CUGAUGAG X CGAA ICAGAGGU 1344 ACCTCTGC T ACAGGTTC 2068
1354 CAGGAACC CUGAUGAG X CGAA IUAGCAGA 1345 TCTGCTAC A GGTTCCTG 2069
1360 GUUGAACA CUGAUGAG X CGAA IAACCUGU 1346 ACAGGTTC C TGTTCAAC 2070
1361 UGUUGAAC CUGAUGAG X CGAA IGAACCUG 1347 CAGGTTCC T GTTCAACA 2071
1366 GUUGCUGU CUGAUGAG X CGAA IAACAGGA 1348 TCCTGTTC A ACAGCAAC 2072
1369 UGUGUUGC CUGAUGAG X CGAA IUUGAACA 1349 TGTTCAAC A GCAACACA 2073
1372 CUAUGUGU CUGAUGAG X CGAA ICUGUUGA 1350 TCAACAGC A ACACATAG 2074
1375 AGGCUAUG CUGAUGAG X CGAA lUUGCUGU 1351 ACAGCAAC A CATAGCCT 2075
1377 UCAGGCUA CUGAUGAG X CGAA IUGUUGCU 1352 AGCAACAC A TAGCCTGA 2076
1382 GAGGGUCA CUGAUGAG X CGAA ICUAUGUG 1353 CACATAGC C TGACCCTC 2077
1383 GGAGGGUC CUGAUGAG X CGAA IGCUAUGU 1354 ACATAGCC T GACCCTCC 2078
1387 UGGAGGAG CUGAUGAG X CGAA IUCAGGCU 1355 AGCCTGAC c CTCCTCCA 2079
1388 GUGGAGGA CUGAUGAG X CGAA IGUCAGGC 1356 GCCTGACC c TCCTCCAC 2080
1389 AGUGGAGG CUGAUGAG X CGAA IGGUCAGG 1357 CCTGACCC T CCTCCACT 2081
1391 GGAGUGGA CUGAUGAG X CGAA IAGGGUCA 1358 TGACCCTC c TCCACTCC 2082
1392 UGGAGUGG CUGAUGAG X CGAA IGAGGGUC 1359 GACCCTCC T CCACTCCA 2083
1394 GGUGGAGU CUGAUGAG X CGAA IAGGAGGG 1360 CCCTCCTC c ACTCCACC 2084
1395 AGGUGGAG CUGAUGAG X CGAA IGAGGAGG 1361 CCTCCTCC A CTCCACCT 2085
1397 GGAGGUGG CUGAUGAG X CGAA IUGGAGGA 1362 TCCTCCAC T CCACCTCC 2086
1399 GUGGAGGU CUGAUGAG X CGAA IAGUGGAG 1363 CTCCACTC c ACCTCCAC 2087
1400 GGUGGAGG CUGAUGAG X CGAA IGAGUGGA 1364 TCCACTCC A CCTCCACC 2088
1402 UGGGUGGA CUGAUGAG X CGAA IUGGAGUG 1365 CACTCCAC C TCCACCCA 2089
1403 GUGGGUGG CUGAUGAG X CGAA IGUGGAGU 1366 ACTCCACC T CCACCCAC 2090
1405 CAGUGGGU CUGAUGAG X CGAA IAGGUGGA 1367 TCCACCTC C ACCCACTG 2091
1406 ACAGUGGG CUGAUGAG X CGAA IGAGGUGG 1368 CCACCTCC A CCCACTGT 2092
1408 GGACAGUG CUGAUGAG X CGAA IUGGAGGU 1369 ACCTCCAC C CACTGTCC 2093
1409 CGGACAGU CUGAUGAG X CGAA IGUGGAGG 1370 CCTCCACC C ACTGTCCG 2094
1410 GCGGACAG CUGAUGAG X CGAA IGGUGGAG 1371 CTCCACCC A CTGTCCGC 2095
1412 AGGCGGAC CUGAUGAG X CGAA IUGGGUGG 1372 CCACCCAC T GTCCGCCT 2096
1416 GCAGAGGC CUGAUGAG X CGAA IACAGUGG 1373 CCACTGTC C GCCTCTGC 2097
1419 CGGGCAGA CUGAUGAG X CGAA ICGGACAG 1374 CTGTCCGC C TCTGCCCG 2098
1420 GCGGGCAG CUGAUGAG X CGAA IGCGGACA 1375 TGTCCGCC T CTGCCCGC 2099
1422 CUGCGGGC CUGAUGAG X CGAA IAGGCGGA 1376 TCCGCCTC T GCCCGCAG 2100
1425 GCUCUGCG CUGAUGAG X CGAA ICAGAGGC 1377 GCCTCTGC C CGCAGAGC 2101
1426 GGCUCUGC CUGAUGAG X CGAA IGCAGAGG 1378 CCTCTGCC C GCAGAGCC 2102
1429 GUGGGCUC CUGAUGAG X CGAA ICGGGCAG 1379 CTGCCCGC A GAGCCCAC 2103
1434 CGGGCGUG CUGAUGAG X CGAA ICUCUGCG 1380 CGCAGAGC C CACGCCCG 2104
1435 UCGGGCGU CUGAUGAG X CGAA IGCUCUGC 1381 GCAGAGCC C ACGCCCGA 2105
Table 4
1436 GUCGGGCG CUGAUGAG X CGAA IGGCUCUG 1382 CAGAGCCC A CGCCCGAC 2106
1440 GCUAGUCG CUGAUGAG X CGAA ICGUGGGC 1383 GCCCACGC C CGACTAGC 2107
1441 UGCUAGUC CUGAUGAG X CGAA IGCGUGGG 1384 CCCACGCC C GACTAGCA 2108
1445 UGCCUGCU CUGAUGAG X CGAA IUCGGGCG 1385 CGCCCGAC T AGCAGGCA 2109
1449 GGCAUGCC CUGAUGAG X CGAA ICUAGUCG 1386 CGACTAGC A GGCATGCC 2110
1453 CCGCGGCA CUGAUGAG X CGAA ICCUGCUA 1387 TAGCAGGC A TGCCGCGG 2111
1457 CCUACCGC CUGAUGAG X CGAA ICAUGCCU 1388 AGGCATGC C GCGGTAGG 2112
1473 GGUCCGGC CUGAUGAG X CGAA ICCCUUAC 1389 GTAAGGGC C GCCGGACC 2113
1476 CGCGGUCC CUGAUGAG X CGAA ICGGCCCU 1390 AGGGCCGC C GGACCGCG 2114
1481 CUCUACGC CUGAUGAG X CGAA IUCCGGCG 1391 CGCCGGAC C GCGTAGAG 2115
1493 CGGGGCCC CUGAUGAG X CGAA ICUCUCUA 1392 TAGAGAGC C GGGCCCCG 2116
1498 CCGUCCGG CUGAUGAG X CGAA ICCCGGCU 1393 AGCCGGGC C CCGGACGG 2117
1499 UCCGUCCG CUGAUGAG X CGAA IGCCCGGC 1394 GCCGGGCC C CGGACGGA 2118
1500 GUCCGUCC CUGAUGAG X CGAA IGGCCCGG 1395 CCGGGCCC C GGACGGAC 2119
1517 UUUAGUGC CUGAUGAG X CGAA IAACCAAC 1396 GTTGGTTC T GCACTAAA 2120
1520 GGUUUUAG CUGAUGAG X CGAA ICAGAACC 1397 GGTTCTGC A CTAAAACC 2121
1522 UGGGUUUU CUGAUGAG X CGAA lUGCAGAA 1398 TTCTGCAC T AAAACCCA 2122
1528 GGAAGAUG CUGAUGAG X CGAA IUUUUAGU 1399 ACTAAAAC C CATCTTCC 2123
1529 GGGAAGAU CUGAUGAG X CGAA IGUUUUAG 1400 CTAAAACC C ATCTTCCC 2124
1530 GGGGAAGA CUGAUGAG X CGAA IGGUUUUA 1401 TAAAACCC A TCTTCCCC 2125
1533 UCCGGGGA CUGAUGAG X CGAA IAUGGGUU 1402 AACCCATC T TCCCCGGA 2126
1536 ACAUCCGG CUGAUGAG X CGAA IAAGAUGG 1403 CCATCTTC C CCGGATGT 2127
1537 CACAUCCG CUGAUGAG X CGAA IGAAGAUG 1404 CATCTTCC C CGGATGTG 2128
1538 ACACAUCC CUGAUGAG X CGAA IGGAAGAU 1405 ATCTTCCC C GGATGTGT 2129
1550 GAGGGGUG CUGAUGAG X CGAA IACACACA 1406 TGTGTGTC T CACCCCTC 2130
1552 AUGAGGGG CUGAUGAG X CGAA lAGACACA 1407 TGTGTCTC A CCCCTCAT 2131
1554 GGAUGAGG CUGAUGAG X CGAA IUGAGACA 1408 TGTCTCAC C CCTCATCC 2132
1555 AGGAUGAG CUGAUGAG X CGAA IGUGAGAC 1409 GTCTCACC C CTCATCCT 2133
1556 AAGGAUGA CUGAUGAG X CGAA IGGUGAGA 1410 TCTCACCC C TCATCCTT 2134
1557 AAAGGAUG CUGAUGAG X CGAA IGGGUGAG 1411 CTCACCCC T CATCCTTT 2135
1559 UAAAAGGA CUGAUGAG X CGAA IAGGGGUG 1412 CACCCCTC A TCCTTTTA 2136
1562 AAGUAAAA CUGAUGAG X CGAA IAUGAGGG 1413 CCCTCATC C TTTTACTT 2137
1563 AAAGUAAA CUGAUGAG X CGAA IGAUGAGG 1414 CCTCATCC T TTTACTTT 2138
1569 GGGCAAAA CUGAUGAG X CGAA IUAAAAGG 1415 CCTTTTAC T TTTTGCCC 2139
1576 GUGGAAGG CUGAUGAG X CGAA ICAAAAAG 1416 CTTTTTGC C CCTTCCAC 2140
1577 AGUGGAAG CUGAUGAG X CGAA IGCAAAAA 1417 TTTTTGCC C CTTCCACT 2141
1578 AAGUGGAA CUGAUGAG X CGAA IGGCAAAA 1418 TTTTGCCC C TTCCACTT 2142
1579 AAAGUGGA CUGAUGAG X CGAA IGGGCAAA 1419 TTTGCCCC T TCCACTTT 2143
1582 CUCAAAGU CUGAUGAG X CGAA IAAGGGGC 1420 GCCCCTTC C ACTTTGAG 2144
1583 ACUCAAAG CUGAUGAG X CGAA IGAAGGGG 1421 CCCCTTCC A CTTTGAGT 2145
1585 GUACUCAA CUGAUGAG X CGAA IUGGAAGG 1422 CCTTCCAC T TTGAGTAC 2146
1594 GUGGAUUU CUGAUGAG X CGAA IUACUCAA 1423 TTGAGTAC C AAATCCAC 2147
1595 UGUGGAUU CUGAUGAG X CGAA IGUACUCA 1424 TGAGTACC A AATCCACA 2148
1600 UGGCUUGU CUGAUGAG X CGAA IAUUUGGU 1425 ACCAAATC C ACAAGCCA 2149
1601 AUGGCUUG CUGAUGAG X CGAA IGAUUUGG 1426 CCAAATCC A CAAGCCAT 2150
1603 AAAUGGCU CUGAUGAG X CGAA IUGGAUUU 1427 AAATCCAC A AGCCATTT 2151
1607 CAAAAAAU CUGAUGAG X CGAA ICUUGUGG 1428 CCACAAGC C ATTTTTTG 2152
Table 4
1608 UCAAAAAA CUGAUGAG X CGAA IGCUUGUG 1429 CACAAGCC A TTTTTTGA 2153
1636 GCCAGCAU CUGAUGAG X CGAA IUACUCUC 1430 GAGAGTAC C ATGCTGGC 2154
1637 CGCCAGCA CUGAUGAG X CGAA IGUACUCU 1431 AGAGTACC A TGCTGGCG 2155
1641 GCGCCGCC CUGAUGAG X CGAA ICAUGGUA 1432 TACCATGC T GGCGGCGC 2156
1650 CUUCCCUC CUGAUGAG X CGAA ICGCCGCC 1433 GGCGGCGC A GAGGGAAG 2157
1663 CGGGUGUA CUGAUGAG X CGAA ICCCCUUC 1434 GAAGGGGC C TACACCCG 2158
1664 ACGGGUGU CUGAUGAG X CGAA IGCCCCUU 1435 AAGGGGCC T ACACCCGT 2159
1667 AAGACGGG CUGAUGAG X CGAA IUAGGCCC 1436 GGGCCTAC A CCCGTCTT 2160
1669 CCAAGACG CUGAUGAG X CGAA IUGUAGGC 1437 GCCTACAC C CGTCTTGG 2161
1670 CCCAAGAC CUGAUGAG X CGAA IGUGUAGG 1438 CCTACACC C GTCTTGGG 2162
1674 GAGCCCCA CUGAUGAG X CGAA IACGGGUG 1439 CACCCGTC T TGGGGCTC 2163
1681 GUGGGGCG CUGAUGAG X CGAA ICCCCAAG 1440 CTTGGGGC T CGCCCCAC 2164
1685 CUGGGUGG CUGAUGAG X CGAA ICGAGCCC 1441 GGGCTCGC C CCACCCAG 2165
1686 CCUGGGUG CUGAUGAG X CGAA IGCGAGCC 1442 GGCTCGCC C CACCCAGG 2166
1687 CCCUGGGU CUGAUGAG X CGAA IGGCGAGC 1443 GCTCGCCC C ACCCAGGG 2167
1688 GCCCUGGG CUGAUGAG X CGAA IGGGCGAG 1444 CTCGCCCC A CCCAGGGC 2168
1690 GAGCCCUG CUGAUGAG X CGAA IUGGGGCG 1445 CGCCCCAC C CAGGGCTC 2169
1691 GGAGCCCU CUGAUGAG X CGAA IGUGGGGC 1446 GCCCCACC C AGGGCTCC 2170
1692 GGGAGCCC CUGAUGAG X CGAA IGGUGGGG 1447 CCCCACCC A GGGCTCCC • 2171
1697 CAGGAGGG CUGAUGAG X CGAA ICCCUGGG 1448 CCCAGGGC T CCCTCCTG 2172
1699 UCCAGGAG CUGAUGAG X CGAA IAGCCCUG 1449 CAGGGCTC C CTCCTGGA 2173
1700 CUCCAGGA CUGAUGAG X CGAA IGAGCCCU 1450 AGGGCTCC C TCCTGGAG 2174
1701 GCUCCAGG CUGAUGAG X CGAA IGGAGCCC 1451 GGGCTCCC T CCTGGAGC 2175
1703 AUGCUCCA CUGAUGAG X CGAA IAGGGAGC 1452 GCTCCCTC c TGGAGCAT 2176
1704 GAUGCUCC CUGAUGAG X CGAA IGAGGGAG 1453 CTCCCTCC T GGAGCATC 2177
1710 GCCUGGGA CUGAUGAG X CGAA ICUCCAGG 1454 CCTGGAGC A TCCCAGGC 2178
1713 CCCGCCUG CUGAUGAG X CGAA IAUGCUCC 1455 GGAGCATC C CAGGCGGG 2179
1714 GCCCGCCU CUGAUGAG X CGAA IGAUGCUC 1456 GAGCATCC C AGGCGGGC 2180
1715 CGCCCGCC CUGAUGAG X CGAA IGGAUGCU 1457 AGCATCCC A GGCGGGCG 2181
1726 GUCUGGCG CUGAUGAG X CGAA ICCGCCCG 1458 CGGGCGGC A CGCCAGAC 2182
1730 GGCUGUCU CUGAUGAG X CGAA ICGUGCCG 1459 CGGCACGC C AGACAGCC 2183
1731 GGGCUGUC CUGAUGAG X CGAA IGCGUGCC 1460 GGCACGCC A GACAGCCC 2184
1735 GGGGGGGC CUGAUGAG X CGAA IUCUGGCG 1461 CGCCAGAC A GCCCCCCC 2185
1738 AAGGGGGG CUGAUGAG X CGAA ICUGUCUG 1462 CAGACAGC C CCCCCCTT 2186
1739 CAAGGGGG CUGAUGAG X CGAA IGCUGUCU 1463 AGACAGCC C CCCCCTTG 2187
1740 UCAAGGGG CUGAUGAG X CGAA IGGCUGUC 1464 GACAGCCC C CCCCTTGA 2188
1741 UUCAAGGG CUGAUGAG X CGAA IGGGCUGU 1465 ACAGCCCC C CCCTTGAA 2189
1742 AUUCAAGG CUGAUGAG X CGAA IGGGGCUG 1466 CAGCCCCC C CCTTGAAT 2190
1743 GAUUCAAG CUGAUGAG X CGAA IGGGGGCU 1467 AGCCCCCC C CTTGAATC 2191
1744 AGAUUCAA CUGAUGAG X CGAA IGGGGGGC 1468 GCCCCCCC C TTGAATCT 2192
1745 CAGAUUCA CUGAUGAG X CGAA IGGGGGGG 1469 CCCCCCCC T TGAATCTG 2193
1752 CUCCCUGC CUGAUGAG X CGAA IAUUCAAG 1470 CTTGAATC T GCAGGGAG 2194
1755 UUGCUCCC CUGAUGAG X CGAA ICAGAUUC 1471 GAATCTGC A GGGAGCAA 2195
1762 UGGAGAGU CUGAUGAG X CGAA ICUCCCUG 1472 CAGGGAGC A ACTCTCCA 2196
1765 GAGUGGAG CUGAUGAG X CGAA IUUGCUCC 1473 GGAGCAAC T CTCCACTC 2197
1767 UGGAGUGG CUGAUGAG X CGAA IAGUUGCU 1474 AGCAACTC T CCACTCCA 2198
1769 UAUGGAGU CUGAUGAG X CGAA IAGAGUUG 1475 CAACTCTC C ACTCCATA 2199
Table 4
1770 AUAUGGAG CUGAUGAG X CGAA IGAGAGUU 1476 AACTCTCC A CTCCATAT 2200
1772 AAAUAUGG CUGAUGAG X CGAA IUGGAGAG 1477 CTCTCCAC T CCATATTT 2201
1774 AUAAAUAU CUGAUGAG X CGAA IAGUGGAG 1478 CTCCACTC C ATATTTAT 2202
1775 AAUAAAUA CUGAUGAG X CGAA IGAGUGGA 1479 TCCACTCC A TATTTATT 2203
1789 GAAAAAAU CUGAUGAG X CGAA IUUUAAAU 1480 ATTTAAAC A ATTTTTTC 2204
1798 GCCUUUGG CUGAUGAG X CGAA IAAAAAAU 1481 ATTTTTTC C CCAAAGGC 2205
1799 UGCCUUUG CUGAUGAG X CGAA IGAAAAAA 1482 TTTTTTCC C CAAAGGCA 2206
1800 AUGCCUUU CUGAUGAG X CGAA IGGAAAAA 1483 TTTTTCCC C AAAGGCAT 2207
1801 GAUGCCUU CUGAUGAG X CGAA IGGGAAAA 1484 TTTTCCCC A AAGGCATC 2208
1807 ACUAUGGA CUGAUGAG X CGAA ICCUUUGG 1485 CCAAAGGC A TCCATAGT 2209
1810 UGCACUAU CUGAUGAG X CGAA IAUGCCUU 1486 AAGGCATC C ATAGTGCA 2210
1811 GUGCACUA CUGAUGAG X CGAA IGAUGCCU 1487 AGGCATCC A TAGTGCAC 2211
1818 AAUGCUAG CUGAUGAG X CGAA ICACUAUG 1488 CATAGTGC A CTAGCATT 2212
1820 AAAAUGCU CUGAUGAG X CGAA IUGCACUA 1489 TAGTGCAC T AGCATTTT 2213
1824 CAAGAAAA CUGAUGAG X CGAA ICUAGUGC 1490 GCACTAGC A TTTTCTTG 2214
1830 UUGGUUCA CUGAUGAG X CGAA IAAAAUGC 1491 GCATTTTC T TGAACCAA 2215
1836 ACAUUAUU CUGAUGAG X CGAA IUUCAAGA 1492 TCTTGAAC C AATAATGT 2216
1837 UACAUUAU CUGAUGAG X CGAA IGUUCAAG 1493 CTTGAACC A ATAATGTA 2217
1864 UGCAAGGC CUGAUGAG X CGAA IACAUCAA 1494 TTGATGTC A GCCTTGCA 2218
1867 UGAUGCAA CUGAUGAG X CGAA ICUGACAU 1495 ATGTCAGC C TTGCATCA 2219
1868 UUGAUGCA CUGAUGAG X CGAA IGCUGACA 1496 TGTCAGCC T TGCATCAA 2220
1872 GCCCUUGA CUGAUGAG X CGAA ICAAGGCU 1497 AGCCTTGC A TCAAGGGC 2221
1875 AAAGCCCU CUGAUGAG X CGAA IAUGCAAG 1498 CTTGCATC A AGGGCTTT 2222
1881 UUUGAUAA CUGAUGAG X CGAA ICCCUUGA 1499 TCAAGGGC T TTATCAAA 2223
1887 GUACUUUU CUGAUGAG X CGAA IAUAAAGC 1500 GCTTTATC A AAAAGTAC 2224
1896 UUUAUUAU CUGAUGAG X CGAA IUACUUUU 1501 AAAAGTAC A ATAATAAA 2225
1907 CUACCUGA CUGAUGAG X CGAA IAUUUAUU 1502 AATAAATC C TCAGGTAG 2226
1908 ACUACCUG CUGAUGAG X CGAA IGAUUUAU 1503 ATAAATCC T CAGGTAGT 2227
1910 GUACUACC CUGAUGAG X CGAA IAGGAUUU 1504 AAATCCTC A GGTAGTAC 2228
1919 CCAUUCCC CUGAUGAG X CGAA IUACUACC 1505 GGTAGTAC T GGGAATGG 2229
1933 CAUGGCAA CUGAUGAG X CGAA ICCUUCCA 1506 TGGAAGGC T TTGCCATG 2230
1938 AGGCCCAU CUGAUGAG X CGAA ICAAAGCC 1507 GGCTTTGC C ATGGGCCT 2231
1939 CAGGCCCA CUGAUGAG X CGAA IGCAAAGC 1508 GCTTTGCC A TGGGCCTG 2232
1945 ACGCAGCA CUGAUGAG X CGAA ICCCAUGG 1509 CCATGGGC C TGCTGCGT 2233
1946 GACGCAGC CUGAUGAG X CGAA IGCCCAUG 1510 CATGGGCC T GCTGCGTC 2234
1949 UCUGACGC CUGAUGAG X CGAA ICAGGCCC 1511 GGGCCTGC T GCGTCAGA 2235
1955 UACUGGUC CUGAUGAG X CGAA IACGCAGC 1512 GCTGCGTC A GACCAGTA 2236
1959 CCAGUACU CUGAUGAG X CGAA IUCUGACG 1513 CGTCAGAC C AGTACTGG 2237
1960 CCCAGUAC CUGAUGAG X CGAA IGUCUGAC 1514 GTCAGACC A GTACTGGG 2238
1965 UCCUUCCC CUGAUGAG X CGAA IUACUGGU 1515 ACCAGTAC T GGGAAGGA 2239
1988 AUAACAAC CUGAUGAG X CGAA ICUUACAA 1516 TTGTAAGC A GTTGTTAT 2240
2032 UAUAGCAU CUGAUGAG X CGAA IUUCUAUC 1517 GATAGAAC A ATGCTATA 2241
2037 UAUAUUAU CUGAUGAG X CGAA ICAUUGUU 1518 AACAATGC T ATAATATA 2242
2054 UACCCACG CUGAUGAG X CGAA lUUCAUUA 1519 TAATGAAC A CGTGGGTA 2243
2076 UCACAUCA CUGAUGAG X CGAA IUUUCUUA 1520 TAAGAAAC A TGATGTGA 2244
2091 CGGGACAA CUGAUGAG X CGAA IUAAUCUC 1521 GAGATTAC T TTGTCCCG 2245
2097 AAUAAGCG CUGAUGAG X CGAA lACAAAGU 1522 ACTTTGTC C CGCTTATT 2246
Table 4
2098 GAAUAAGC CUGAUGAG X CGAA IGACAAAG 1523 CTTTGTCC C GCTTATTC 2247
2101 GCAGAAUA CUGAUGAG X CGAA ICGGGACA 1524 TGTCCCGC T TATTCTGC 2248
2107 CAGGGAGC CUGAUGAG X CGAA IAAUAAGC 1525 GCTTATTC T GCTCCCTG 2249
2110 UAACAGGG CUGAUGAG X CGAA ICAGAAUA 1526 TATTCTGC T CCCTGTTA 2250
2112 GAUAACAG CUGAUGAG X CGAA IAGCAGAA 1527 TTCTGCTC C CTGTTATC 2251
2113 AGAUAACA CUGAUGAG X CGAA IGAGCAGA 1528 TCTGCTCC C TGTTATCT 2252
2114 CAGAUAAC CUGAUGAG X CGAA IGGAGCAG 1529 CTGCTCCC T GTTATCTG 2253
2121 GAUCUAGC CUGAUGAG X CGAA IAUAACAG 1530 CTGTTATC T GCTAGATC 2254
2124 CUAGAUCU CUGAUGAG X CGAA ICAGAUAA 1531 TTATCTGC T AGATCTAG 2255
2130 UGAGAACU CUGAUGAG X CGAA IAUCUAGC 1532 GCTAGATC T AGTTCTCA 2256
2136 AGUGAUUG CUGAUGAG X CGAA IAACUAGA 1533 TCTAGTTC T CAATCACT 2257
2138 GCAGUGAU CUGAUGAG X CGAA IAGAACUA 1534 TAGTTCTC A ATCACTGC 2258
2142 GGGAGCAG CUGAUGAG X CGAA IAUUGAGA 1535 TCTCAATC A CTGCTCCC 2259
2144 GGGGGAGC CUGAUGAG X CGAA IUGAUUGA 1536 TCAATCAC T GCTCCCCC 2260
2147 CACGGGGG CUGAUGAG X CGAA ICAGUGAU 1537 ATCACTGC T CCCCCGTG 2261
2149 CACACGGG CUGAUGAG X CGAA IAGCAGUG 1538 CACTGCTC C CCCGTGTG 2262
2150 ACACACGG CUGAUGAG X CGAA IGAGCAGU 1539 ACTGCTCC C CCGTGTGT 2263
2151 UACACACG CUGAUGAG X CGAA IGGAGCAG 1540 CTGCTCCC C CGTGTGTA 2264
2152 AUACACAC CUGAUGAG X CGAA IGGGAGCA 1541 TGCTCCCC C GTGTGTAT 2265
2169 ACCUUACA CUGAUGAG X CGAA ICAUUCUA 1542 TAGAATGC A TGTAAGGT 2266
2179 ACACAAGA CUGAUGAG X CGAA IACCUUAC 1543 GTAAGGTC T TCTTGTGT 2267
2182 AGGACACA CUGAUGAG X CGAA IAAGACCU 1544 AGGTCTTC T TGTGTCCT 2268
2189 UUUCAUCA CUGAUGAG X CGAA IACACAAG 1545 CTTGTGTC c TGATGAAA 2269
2190 UUUUCAUC CUGAUGAG X CGAA IGACACAA 1546 TTGTGTCC T GATGAAAA 2270
2207 UCAUUUCA CUGAUGAG X CGAA ICACAUAU 1547 ATATGTGC T TGAAATGA 2271
2221 GAGAUCAA CUGAUGAG X CGAA IUUUCUCA 1548 TGAGAAAC T TTGATCTC 2272
2228 GUAAGCAG CUGAUGAG X CGAA IAUCAAAG 1549 CTTTGATC T CTGCTTAC 2273
2230 UAGUAAGC CUGAUGAG X CGAA lAGAUCAA 1550 TTGATCTC T GCTTACTA 2274
2233 CAUUAGUA CUGAUGAG X CGAA ICAGAGAU 1551 ATCTCTGC T TACTAATG 2275
2237 GGCACAUU CUGAUGAG X CGAA IUAAGCAG 1552 CTGCTTAC T AATGTGCC 2276
2245 GGACAUGG CUGAUGAG X CGAA ICACAUUA 1553 TAATGTGC c CCATGTCC 2277
2246 UGGACAUG CUGAUGAG X CGAA IGCACAUU 1554 AATGTGCC c CATGTCCA 2278
2247 UUGGACAU CUGAUGAG X CGAA IGGCACAU 1555 ATGTGCCC c ATGTCCAA 2279
2248 CUUGGACA CUGAUGAG X CGAA IGGGCACA 1556 TGTGCCCC A TGTCCAAG 2280
2253 UUGGACUU CUGAUGAG X CGAA IACAUGGG 1557 CCCATGTC c AAGTCCAA 2281
2254 GUUGGACU CUGAUGAG X CGAA IGACAUGG 1558 CCATGTCC A AGTCCAAC 2282
2259 GGCAGGUU CUGAUGAG X CGAA IACUUGGA 1559 TCCAAGTC C AACCTGCC 2283
2260 AGGCAGGU CUGAUGAG X CGAA IGACUUGG 1560 CCAAGTCC A ACCTGCCT 2284
2263 CACAGGCA CUGAUGAG X CGAA IUUGGACU 1561 AGTCCAAC C TGCCTGTG 2285
2264 GCACAGGC CUGAUGAG X CGAA IGUUGGAC 1562 GTCCAACC T GCCTGTGC 2286
2267 CAUGCACA CUGAUGAG X CGAA ICAGGUUG 1563 CAACCTGC C TGTGCATG 2287
2268 UCAUGCAC CUGAUGAG X CGAA IGCAGGUU 1564 AACCTGCC T GTGCATGA 2288
2273 UCAGGUCA CUGAUGAG X CGAA ICACAGGC 1565 GCCTGTGC A TGACCTGA 2289
2278 AAUGAUCA CUGAUGAG X CGAA lUCAUGCA 1566 TGCATGAC C TGATCATT 2290
2279 UAAUGAUC CUGAUGAG X CGAA IGUCAUGC 1567 GCATGACC T GATCATTA 2291
2284 CCAUGUAA CUGAUGAG X CGAA IAUCAGGU 1568 ACCTGATC A TTACATGG 2292
2289 CACAGCCA CUGAUGAG X CGAA IUAAUGAU 1569 ATCATTAC A TGGCTGTG 2293
Table 4
2294 GGAACCAC CUGAUGAG X CGAA ICCAUGUA 1570 TACATGGC T GTGGTTCC 2294
2302 CAGGCUUA CUGAUGAG X CGAA IAACCACA 1571 TGTGGTTC C TAAGCCTG 2295
2303 ACAGGCUU CUGAUGAG X CGAA IGAACCAC 1572 GTGGTTCC T AAGCCTGT 2296
2308 CAGCAACA CUGAUGAG X CGAA ICUUAGGA 1573 TCCTAAGC C TGTTGCTG 2297
2309 UCAGCAAC CUGAUGAG X CGAA IGCUUAGG 1574 CCTAAGCC T GTTGCTGA 2298
2315 AUGACUUC CUGAUGAG X CGAA ICAACAGG 1575 CCTGTTGC T GAAGTCAT 2299
2322 AGCGACAA CUGAUGAG X CGAA IACUUCAG 1576 CTGAAGTC A TTGTCGCT 2300
2330 UAUUGCUG CUGAUGAG X CGAA ICGACAAU 1577 ATTGTCGC T CAGCAATA 2301
2332 CCUAUUGC CUGAUGAG X CGAA IAGCGACA 1578 TGTCGCTC A GCAATAGG 2302
2335 CACCCUAU CUGAUGAG X CGAA ICUGAGCG 1579 CGCTCAGC A ATAGGGTG 2303
2345 UGGAAAAC CUGAUGAG X CGAA ICACCCUA 1580 TAGGGTGC A GTTTTCCA 2304
2352 CUAUUCCU CUGAUGAG X CGAA IAAAACUG 1581 CAGTTTTC C AGGAATAG 2305
2353 CCUAUUCC CUGAUGAG X CGAA IGAAAACU 1582 AGTTTTCC A GGAATAGG 2306
2363 UAGGCAAA CUGAUGAG X CGAA ICCUAUUC 1583 GAATAGGC A TTTGCCTA 2307
2369 AGGAAUUA CUGAUGAG X CGAA ICAAAUGC 1584 GCATTTGC C TAATTCCT 2308
2370 CAGGAAUU CUGAUGAG X CGAA IGCAAAUG 1585 CATTTGCC T AATTCCTG 2309
2376 UCAUGCCA CUGAUGAG X CGAA IAAUUAGG 1586 CCTAATTC C TGGCATGA 2310
2377 GUCAUGCC CUGAUGAG X CGAA IGAAUUAG 1587 CTAATTCC T GGCATGAC 2311
2381 GAGUGUCA CUGAUGAG X CGAA ICCAGGAA 1588 TTCCTGGC A TGACACTC 2312
2386 CACUAGAG CUGAUGAG X CGAA IUCAUGCC 1589 GGCATGAC A CTCTAGTG 2313
2388 GUCACUAG CUGAUGAG X CGAA IUGUCAUG 1590 CATGACAC T CTAGTGAC 2314
2390 AAGUCACU CUGAUGAG X CGAA lAGUGUCA 1591 TGACACTC T AGTGACTT 2315
2397 CACCAGGA CUGAUGAG X CGAA IUCACUAG 1592 CTAGTGAC T TCCTGGTG 2316
2400 CCUCACCA CUGAUGAG X CGAA IAAGUCAC 1593 GTGACTTC C TGGTGAGG 2317
2401 GCCUCACC CUGAUGAG X CGAA IGAAGUCA 1594 TGACTTCC T GGTGAGGC 2318
2410 ACAGGCUG CUGAUGAG X CGAA ICCUCACC 1595 GGTGAGGC C CAGCCTGT 2319
2411 GACAGGCU CUGAUGAG X CGAA IGCCUCAC 1596 GTGAGGCC C AGCCTGTC 2320
2412 GGACAGGC CUGAUGAG X CGAA IGGCCUCA 1597 TGAGGCCC A GCCTGTCC 2321
2415 CCAGGACA CUGAUGAG X CGAA ICUGGGCC 1598 GGCCCAGC C TGTCCTGG 2322
2416 ACCAGGAC CUGAUGAG X CGAA IGCUGGGC 1599 GCCCAGCC T GTCCTGGT 2323
2420 CUGUACCA CUGAUGAG X CGAA IACAGGCU 1600 AGCCTGTC C TGGTACAG 2324
2421 GCUGUACC CUGAUGAG X CGAA IGACAGGC 1601 GCCTGTCC T GGTACAGC 2325
2427 GACCCUGC CUGAUGAG X CGAA IUACCAGG 1602 CCTGGTAC A GCAGGGTC 2326
2430 CAAGACCC CUGAUGAG X CGAA ICUGUACC 1603 GGTACAGC A GGGTCTTG 2327
2436 UUACAGCA CUGAUGAG X CGAA IACCCUGC 1604 GCAGGGTC T TGCTGTAA 2328
2440 UGAGUUAC CUGAUGAG X CGAA ICAAGACC 1605 GGTCTTGC T GTAACTCA 2329
2446 AAUGUCUG CUGAUGAG X CGAA IUUACAGC 1606 GCTGTAAC T CAGACATT 2330
2448 GGAAUGUC CUGAUGAG X CGAA lAGUUACA 1607 TGTAACTC A GACATTCC 2331
2452 CCUUGGAA CUGAUGAG X CGAA IUCUGAGU 1608 ACTCAGAC A TTCCAAGG 2332
2456 AUACCCUU CUGAUGAG X CGAA IAAUGUCU 1609 AGACATTC C AAGGGTAT 2333
2457 CAUACCCU CUGAUGAG X CGAA IGAAUGUC 1610 GACATTCC A AGGGTATG 2334
2472 GUGAAUAU CUGAUGAG X CGAA ICUUCCCA 1611 TGGGAAGC C ATATTCAC 2335
2473 UGUGAAUA CUGAUGAG X CGAA IGCUUCCC 1612 GGGAAGCC A TATTCACA 2336
2479 GUGAGGUG CUGAUGAG X CGAA IAAUAUGG 1613 CCATATTC A CACCTCAC 2337
2481 GCGUGAGG CUGAUGAG X CGAA IUGAAUAU 1614 ATATTCAC A CCTCACGC 2338
2483 GAGCGUGA CUGAUGAG X CGAA IUGUGAAU 1615 ATTCACAC C TCACGCTC 2339
2484 AGAGCGUG CUGAUGAG X CGAA IGUGUGAA 1616 TTCACACC T CACGCTCT 2340
Table 4
2486 CCAGAGCG CUGAUGAG X CGAA IAGGUGUG 1617 CACACCTC A CGCTCTGG 2341
2490 AUGUCCAG CUGAUGAG X CGAA ICGUGAGG 1618 CCTCACGC T CTGGACAT 2342
2492 UCAUGUCC CUGAUGAG X CGAA IAGCGUGA 1619 TCACGCTC T GGACATGA 2343
2497 CUAAAUCA CUGAUGAG X CGAA IUCCAGAG 1620 CTCTGGAC A TGATTTAG 2344
2512 GGUGUCCC CUGAUGAG X CGAA ICUUCCCU 1621 AGGGAAGC A GGGACACC 2345
2518 GCGGGGGG CUGAUGAG X CGAA IUCCCUGC 1622 GCAGGGAC A CCCCCCGC 2346
2520 GGGCGGGG CUGAUGAG X CGAA IUGUCCCU 1623 AGGGACAC C CCCCGCCC 2347
2521 GGGGCGGG CUGAUGAG X CGAA IGUGUCCC 1624 GGGACACC C CCCGCCCC 2348
2522 GGGGGCGG CUGAUGAG X CGAA IGGUGUCC 1625 GGACACCC C CCGCCCCC 2349
2523 GGGGGGCG CUGAUGAG X CGAA IGGGUGUC 1626 GACACCCC C CGCCCCCC 2350
2524 UGGGGGGC CUGAUGAG X CGAA IGGGGUGU 1627 ACACCCCC C GCCCCCCA 2351
2527 AGGUGGGG CUGAUGAG X CGAA ICGGGGGG 1628 CCCCCCGC C CCCCACCT 2352
2528 AAGGUGGG CUGAUGAG X CGAA IGCGGGGG 1629 CCCCCGCC C CCCACCTT 2353
2529 AAAGGUGG CUGAUGAG X CGAA IGGCGGGG 1630 CCCCGCCC C CCACCTTT 2354
2530 CAAAGGUG CUGAUGAG X CGAA IGGGCGGG 1631 CCCGCCCC C CACCTTTG 2355
2531 CCAAAGGU CUGAUGAG X CGAA IGGGGCGG 1632 CCGCCCCC C ACCTTTGG 2356
2532 CCCAAAGG CUGAUGAG X CGAA IGGGGGCG 1633 CGCCCCCC A CCTTTGGG 2357
2534 AUCCCAAA CUGAUGAG X CGAA IUGGGGGG 1634 CCCCCCAC C TTTGGGAT 2358
2535 GAUCCCAA CUGAUGAG X CGAA IGUGGGGG 1635 CCCCCACC T TTGGGATC 2359
2544 GCGGAGGC CUGAUGAG X CGAA IAUCCCAA 1636 TTGGGATC A GCCTCCGC 2360
2547 AUGGCGGA CUGAUGAG X CGAA ICUGAUCC 1637 GGATCAGC C TCCGCCAT 2361
2548 AAUGGCGG CUGAUGAG X CGAA IGCUGAUC 1638 GATCAGCC T CCGCCATT 2362
2550 GGAAUGGC CUGAUGAG X CGAA IAGGCUGA 1639 TCAGCCTC C GCCATTCC 2363
2553 CUUGGAAU CUGAUGAG X CGAA ICGGAGGC 1640 GCCTCCGC C ATTCCAAG 2364
2554 ACUUGGAA CUGAUGAG X CGAA IGCGGAGG 1641 CCTCCGCC A TTCCAAGT 2365
2558 GUCGACUU CUGAUGAG X CGAA IAAUGGCG 1642 CGCCATTC C AAGTCGAC 2366
2559 UGUCGACU CUGAUGAG X CGAA IGAAUGGC 1643 GCCATTCC A AGTCGACA 2367
2567 AAGAAGAG CUGAUGAG X CGAA IUCGACUU 1644 AAGTCGAC A CTCTTCTT 2368
2569 UCAAGAAG CUGAUGAG X CGAA IUGUCGAC 1645 GTCGACAC T CTTCTTGA 2369
2571 GCUCAAGA CUGAUGAG X CGAA IAGUGUCG 1646 CGACACTC T TCTTGAGC 2370
2574 UCUGCUCA CUGAUGAG X CGAA IAAGAGUG 1647 CACTCTTC T TGAGCAGA 2371
2580 UCACGGUC CUGAUGAG X CGAA ICUCAAGA 1648 TCTTGAGC A GACCGTGA 2372
2584 CAAAUCAC CUGAUGAG X CGAA IUCUGCUC 1649 GAGCAGAC C GTGATTTG 2373
2603 CCAGCAGG CUGAUGAG X CGAA ICCUCUCU 1650 AGAGAGGC A CCTGCTGG 2374
2605 UUCCAGCA CUGAUGAG X CGAA IUGCCUCU 1651 AGAGGCAC C TGCTGGAA 2375
2606 UUUCCAGC CUGAUGAG X CGAA IGUGCCUC 1652 GAGGCACC T GCTGGAAA 2376
2609 UGGUUUCC CUGAUGAG X CGAA ICAGGUGC 1653 GCACCTGC T GGAAACCA 2377
2616 AGAAGUGU CUGAUGAG X CGAA IUUUCCAG 1654 CTGGAAAC C ACACTTCT 2378
2617 AAGAAGUG CUGAUGAG X CGAA IGUUUCCA 1655 TGGAAACC A CACTTCTT 2379
2619 UCAAGAAG CUGAUGAG X CGAA IUGGUUUC 1656 GAAACCAC A CTTCTTGA 2380
2621 UUUCAAGA CUGAUGAG X CGAA IUGUGGUU 1657 AACCACAC T TCTTGAAA 2381
2624 CUGUUUCA CUGAUGAG X CGAA IAAGUGUG 1658 CACACTTC T TGAAACAG 2382
2631 ACCCAGGC CUGAUGAG X CGAA IUUUCAAG 1659 CTTGAAAC A GCCTGGGT 2383
2634 GUCACCCA CUGAUGAG X CGAA ICUGUUUC 1660 GAAACAGC C TGGGTGAC 2384
2635 CGUCACCC CUGAUGAG X CGAA IGCUGUUU 1661 AAACAGCC T GGGTGACG 2385
2647 UGCCUAAA CUGAUGAG X CGAA IACCGUCA 1662 TGACGGTC C TTTAGGCA 2386
2648 CUGCCUAA CUGAUGAG X CGAA IGACCGUC 1663 GACGGTCC T TTAGGCAG 2387
Table 4
2655 CGGCAGGC CUGAUGAG X CGAA ICCUAAAG 1664 CTTTAGGC A GCCTGCCG 2388
2658 CGGCGGCA CUGAUGAG X CGAA ICUGCCUA 1665 TAGGCAGC C TGCCGCCG 2389
2659 ACGGCGGC CUGAUGAG X CGAA IGCUGCCU 1666 AGGCAGCC T GCCGCCGT 2390
2662 GAGACGGC CUGAUGAG X CGAA ICAGGCUG 1667 CAGCCTGC C GCCGTCTC 2391
2665 ACAGAGAC CUGAUGAG X CGAA ICGGCAGG 1668 CCTGCCGC C GTCTCTGT 2392
2669 CGGGACAG CUGAUGAG X CGAA IACGGCGG 1669 CCGCCGTC T CTGTCCCG 2393
2671 ACCGGGAC CUGAUGAG X CGAA IAGACGGC 1670 GCCGTCTC T GTCCCGGT 2394
2675 GUGAACCG CUGAUGAG X CGAA IACAGAGA 1671 TCTCTGTC c CGGTTCAC 2395
2676 GGUGAACC CUGAUGAG X CGAA IGACAGAG 1672 CTCTGTCC c GGTTCACC 2396
2682 CGGCAAGG CUGAUGAG X CGAA IAACCGGG 1673 CCCGGTTC A CCTTGCCG 2397
2684 CUCGGCAA CUGAUGAG X CGAA IUGAACCG 1674 CGGTTCAC c TTGCCGAG 2398
2685 UCUCGGCA CUGAUGAG X CGAA IGUGAACC 1675 GGTTCACC T TGCCGAGA 2399
2689 CCUCUCUC CUGAUGAG X CGAA ICAAGGUG 1676 CACCTTGC C GAGAGAGG 2400
2704 GGUGGGGC CUGAUGAG X CGAA IACGCGCC 1677 GGCGCGTC T GCCCCACC 2401
2707 GAGGGUGG CUGAUGAG X CGAA ICAGACGC 1678 GCGTCTGC C CCACCCTC 2402
2708 UGAGGGUG CUGAUGAG X CGAA IGCAGACG 1679 CGTCTGCC C CACCCTCA 2403
2709 UUGAGGGU CUGAUGAG X CGAA IGGCAGAC 1680 GTCTGCCC c ACCCTCAA 2404
2710 UUUGAGGG CUGAUGAG X CGAA IGGGCAGA 1681 TCTGCCCC A CCCTCAAA 2405
2712 GGUUUGAG CUGAUGAG X CGAA IUGGGGCA 1682 TGCCCCAC C CTCAAACC 2406
2713 GGGUUUGA CUGAUGAG X CGAA IGUGGGGC 1683 GCCCCACC C TCAAACCC 2407
2714 AGGGUUUG CUGAUGAG X CGAA IGGUGGGG 1684 CCCCACCC T CAAACCCT 2408
2716 ACAGGGUU CUGAUGAG X CGAA IAGGGUGG 1685 CCACCCTC A AACCCTGT 2409
2720 CCCCACAG CUGAUGAG X CGAA IUUUGAGG 1686 CCTCAAAC C CTGTGGGG 2410
2721 GCCCCACA CUGAUGAG X CGAA IGUUUGAG 1687 CTCAAACC C TGTGGGGC 2411
2722 GGCCCCAC CUGAUGAG X CGAA IGGUUUGA 1688 TCAAACCC T GTGGGGCC 2412
2730 CACCAUCA CUGAUGAG X CGAA ICCCCACA 1689 TGTGGGGC C TGATGGTG 2413
2731 GCACCAUC CUGAUGAG X CGAA IGCCCCAC 1690 GTGGGGCC T GATGGTGC 2414
2740 GAGUCGUG CUGAUGAG X CGAA ICACCAUC 1691 GATGGTGC T CACGACTC 2415
2742 AAGAGUCG CUGAUGAG X CGAA IAGCACCA 1692 TGGTGCTC A CGACTCTT 2416
2747 GCAGGAAG CUGAUGAG X CGAA IUCGUGAG 1693 CTCACGAC T CTTCCTGC 2417
2749 UUGCAGGA CUGAUGAG X CGAA IAGUCGUG 1694 CACGACTC T TCCTGCAA 2418
2752 CCUUUGCA CUGAUGAG X CGAA IAAGAGUC 1695 GACTCTTC C TGCAAAGG 2419
2753 CCCUUUGC CUGAUGAG X CGAA IGAAGAGU 1696 ACTCTTCC T GCAAAGGG 2420
2756 GUUCCCUU CUGAUGAG X CGAA ICAGGAAG 1697 CTTCCTGC A AAGGGAAC 2421
2765 AGGUCUUC CUGAUGAG X CGAA IUUCCCUU 1698 AAGGGAAC T GAAGACCT 2422
2772 AAUGUGGA CUGAUGAG X CGAA IUCUUCAG 1699 CTGAAGAC C TCCACATT 2423
2773 UAAUGUGG CUGAUGAG X CGAA IGUCUUCA 1700 TGAAGACC T CCACATTA 2424
2775 CUUAAUGU CUGAUGAG X CGAA IAGGUCUU 1701 AAGACCTC C ACATTAAG 2425
2776 ACUUAAUG CUGAUGAG X CGAA IGAGGUCU 1702 AGACCTCC A CATTAAGT 2426
2778 CCACUUAA CUGAUGAG X CGAA IUGGAGGU 1703 ACCTCCAC A TTAAGTGG 2427
2788 UGUUAAAA CUGAUGAG X CGAA ICCACUUA 1704 TAAGTGGC T TTTTAACA 2428
2796 GUUUUUCA CUGAUGAG X CGAA IUUAAAAA 1705 TTTTTAAC A TGAAAAAC 2429
2805 AGCUGCCG CUGAUGAG X CGAA IUUUUUCA 1706 TGAAAAAC A CGGCAGCT 2430
2810 GCUACAGC CUGAUGAG X CGAA ICCGUGUU 1707 AACACGGC A GCTGTAGC 2431
2813 GGAGCUAC CUGAUGAG X CGAA ICUGCCGU 1708 ACGGCAGC T GTAGCTCC 2432
2819 AGCUCGGG CUGAUGAG X CGAA ICUACAGC 1709 GCTGTAGC T CCCGAGCT 2433
2821 GUAGCUCG CUGAUGAG X CGAA IAGCUACA 1710 TGTAGCTC C CGAGCTAC 2434
Table 4
2822 AGUAGCUC CUGAUGAG X CGAA IGAGCUAC 1711 GTAGCTCC C GAGCTACT 2435
2827 AAGAGAGU CUGAUGAG X CGAA ICUCGGGA 1712 TCCCGAGC T ACTCTCTT 2436
2830 GGCAAGAG CUGAUGAG X CGAA IUAGCUCG 1713 CGAGCTAC T CTCTTGCC 2437
2832 CUGGCAAG CUGAUGAG X CGAA IAGUAGCU 1714 AGCTACTC T CTTGCCAG 2438
2834 UGCUGGCA CUGAUGAG X CGAA IAGAGUAG 1715 CTACTCTC T TGCCAGCA 2439
2838 AAAAUGCU CUGAUGAG X CGAA ICAAGAGA 1716 TCTCTTGC c AGCATTTT 2440
2839 GAAAAUGC CUGAUGAG X CGAA IGCAAGAG 1717 CTCTTGCC A GCATTTTC 2441
2842 UGUGAAAA CUGAUGAG X CGAA ICUGGCAA 1718 TTGCCAGC A TTTTCACA 2442
2848 GCAAAAUG CUGAUGAG X CGAA IAAAAUGC 1719 GCATTTTC A CATTTTGC 2443
2850 AGGCAAAA CUGAUGAG X CGAA IUGAAAAU 1720 ATTTTCAC A TTTTGCCT 2444
2857 ACGAGAAA CUGAUGAG X CGAA ICAAAAUG 1721 CATTTTGC C TTTCTCGT 2445
2858 CACGAGAA CUGAUGAG X CGAA IGCAAAAU 1722 ATTTTGCC T TTCTCGTG 2446
2862 CUACCACG CUGAUGAG X CGAA IAAAGGCA 1723 TGCCTTTC T CGTGGTAG 2447
2875 UCUGUACU CUGAUGAG X CGAA ICUUCUAC 1724 GTAGAAGC c AGTACAGA 2448
2876 CUCUGUAC CUGAUGAG X CGAA IGCUUCUA 1725 TAGAAGCC A GTACAGAG 2449
2881 AAUUUCUC CUGAUGAG X CGAA IUACUGGC 1726 GCCAGTAC A GAGAAATT 2450
2891 CCCACCAC CUGAUGAG X CGAA IAAUUUCU 1727 AGAAATTC T GTGGTGGG 2451
2903 ACCUCGAA CUGAUGAG X CGAA IUUCCCAC 1728 GTGGGAAC A TTCGAGGT 2452
2915 CUGCAGGG CUGAUGAG X CGAA IACACCUC 1729 GAGGTGTC A CCCTGCAG 2453
2917 CUCUGCAG CUGAUGAG X CGAA IUGACACC 1730 GGTGTCAC c CTGCAGAG 2454
2918 GCUCUGCA CUGAUGAG X CGAA IGUGACAC 1731 GTGTCACC C TGCAGAGC 2455
2919 AGCUCUGC CUGAUGAG X CGAA IGGUGACA 1732 TGTCACCC T GCAGAGCT 2456
2922 CAUAGCUC CUGAUGAG X CGAA ICAGGGUG 1733 CACCCTGC A GAGCTATG 2457
2927 CUCACCAU CUGAUGAG X CGAA ICUCUGCA 1734 TGCAGAGC T ATGGTGAG 2458
2949 GGCACCUA CUGAUGAG X CGAA ICCUUAUC 1735 GATAAGGC T TAGGTGCC 2459
2957 UACAGCCU CUGAUGAG X CGAA ICACCUAA 1736 TTAGGTGC C AGGCTGTA 2460
2958 UUACAGCC CUGAUGAG X CGAA IGCACCUA 1737 TAGGTGCC A GGCTGTAA 2461
2962 AUGCUUAC CUGAUGAG X CGAA ICCUGGCA 1738 TGCCAGGC T GTAAGCAT 2462
2969 GCUCAGAA CUGAUGAG X CGAA ICUUACAG 1739 CTGTAAGC A TTCTGAGC 2463
2973 GCCAGCUC CUGAUGAG X CGAA IAAUGCUU 1740 AAGCATTC T GAGCTGGC 2464
2978 AACAAGCC CUGAUGAG X CGAA ICUCAGAA 1741 TTCTGAGC T GGCTTGTT 2465
2982 AAACAACA CUGAUGAG X CGAA ICCAGCUC 1742 GAGCTGGC T TGTTGTTT 2466
2998 CAUAUACA CUGAUGAG X CGAA IACUUAAA 1743 TTTAAGTC C TGTATATG 2467
2999 ACAUAUAC CUGAUGAG X CGAA IGACUUAA 1744 TTAAGTCC T GTATATGT 2468
3040 UUUUGAAA CUGAUGAG X CGAA ICUACUAU 1745 ATAGTAGC A TTTCAAAA 2469
3045 GUCCAUUU CUGAUGAG X CGAA IAAAUGCU 1746 AGCATTTC A AAATGGAC 2470
3058 GUUAAACC CUGAUGAG X CGAA IUACGUCC 1747 GGACGTAC T GGTTTAAC 2471
3067 GGAUAGGA CUGAUGAG X CGAA IUUAAACC 1748 GGTTTAAC C TCCTATCC 2472
3068 AGGAUAGG CUGAUGAG X CGAA IGUUAAAC 1749 GTTTAACC T CCTATCCT 2473
3070 CAAGGAUA CUGAUGAG X CGAA IAGGUUAA 1750 TTAACCTC C TATCCTTG 2474
3071 CCAAGGAU CUGAUGAG X CGAA IGAGGUUA 1751 TAACCTCC T ATCCTTGG 2475
3075 CUCUCCAA CUGAUGAG X CGAA IAUAGGAG 1752 CTCCTATC C TTGGAGAG 2476
3076 GCUCUCCA CUGAUGAG X CGAA IGAUAGGA 1753 TCCTATCC T TGGAGAGC 2477
3085 GAGCCAGC CUGAUGAG X CGAA ICUCUCCA 1754 TGGAGAGC A GCTGGCTC 2478
3088 GGAGAGCC CUGAUGAG X CGAA ICUGCUCU 1755 AGAGCAGC T GGCTCTCC 2479
3092 AGGUGGAG CUGAUGAG X CGAA ICCAGCUG 1756 CAGCTGGC T CTCCACCT 2480
3094 CAAGGUGG CUGAUGAG X CGAA IAGCCAGC 1757 GCTGGCTC T CCACCTTG 2481
Table 4
3096 AACAAGGU CUGAUGAG X CGAA lAGAGCCA 1758 TGGCTCTC C ACCTTGTT 2482
3097 UAACAAGG CUGAUGAG X CGAA IGAGAGCC 1759 GGCTCTCC A CCTTGTTA 2483
3099 UGUAACAA CUGAUGAG X CGAA IUGGAGAG 1760 CTCTCCAC C TTGTTACA 2484
3100 GUGUAACA CUGAUGAG X CGAA IGUGGAGA 1761 TCTCCACC T TGTTACAC 2485
3107 ACAUAAUG CUGAUGAG X CGAA IUAACAAG 1762 CTTGTTAC A CATTATGT 2486
3109 UAACAUAA CUGAUGAG X CGAA IUGUAACA 1763 TGTTACAC A TTATGTTA 2487
3132 AGCAGAGC CUGAUGAG X CGAA ICUCGCUA . 1764 TAGCGAGC T GCTCTGCT 2488
3135 CAUAGCAG CUGAUGAG X CGAA ICAGCUCG 1765 CGAGCTGC T CTGCTATG 2489
3137 GACAUAGC CUGAUGAG X CGAA IAGCAGCU 1766 AGCTGCTC T GCTATGTC 2490
3140 AAGGACAU CUGAUGAG X CGAA ICAGAGCA 1767 TGCTCTGC T ATGTCCTT 2491
3146 UGGCUUAA CUGAUGAG X CGAA IACAUAGC 1768 GCTATGTC c TTAAGCCA 2492
3147 UUGGCUUA CUGAUGAG X CGAA IGACAUAG 1769 CTATGTCC T TAAGCCAA 2493
3153 UAAAUAUU CUGAUGAG X CGAA ICUUAAGG 1770 CCTTAAGC c AATATTTA 2494
3154 GUAAAUAU CUGAUGAG X CGAA IGCUUAAG 1771 CTTAAGCC A ATATTTAC 2495
3163 ACCUGAUG CUGAUGAG X CGAA IUAAAUAU 1772 ATATTTAC T CATCAGGT 2496
3165 UGACCUGA CUGAUGAG X CGAA IAGUAAAU 1773 ATTTACTC A TCAGGTCA 2497
3168 UAAUGACC CUGAUGAG X CGAA IAUGAGUA 1774 TACTCATC A GGTCATTA 2498
3173 AAAAAUAA CUGAUGAG X CGAA IACCUGAU 1775 ATCAGGTC A TTATTTTT 2499
3185 AUGGCCAU CUGAUGAG X CGAA IUAAAAAA 1776 TTTTTTAC A ATGGCCAT 2500
3191 UAUUCCAU CUGAUGAG X CGAA ICCAUUGU 1777 ACAATGGC c ATGGAATA 2501
3192 UUAUUCCA CUGAUGAG X CGAA IGCCAUUG 1778 CAATGGCC A TGGAATAA 2502
3203 GUAAAAAU CUGAUGAG X CGAA IUUUAUUC 1779 GAATAAAC c ATTTTTAC 2503
3204 UGUAAAAA CUGAUGAG X CGAA IGUUUAUU 1780 AATAAACC A TTTTTACA 2504
Table 5
Table 5: Human PTP-IB G-Cleaver Ribozyme and Target Sequence
Nt. Ribozyme Sequence Seq. ID Substrate Sequence Seq. ID position Nos. Nos.
25 CACUG UGAUG GCAUGCACUAUGC GCG GCCGUCUGCC 2505 GGCAGACGGC G CAGTG 2670
35 CUUCU UGAUG GCAUGCACUAUGC GCG GGCCCACUGC 2506 GCAGTGGGCC G AGAAG 2671
46 UGCUG UGAUG GCAUGCACUAUGC GCG GCCUCCUUCU 2507 AGAAGGAGGC G CAGCA 2672
55 CAGGG UGAUG GCAUGCACUAUGC GCG GGCUGCUGCG 2508 CGCAGCAGCC G CCCTG 2673
89 CUGCU UGAUG GCAUGCACUAUGC GCG GAACUCCUUU 2509 AAAGGAGTTC G AGCAG 2674
98 CUUGU UGAUG GCAUGCACUAUGC GCG GAUCUGCUCG 2510 CGAGCAGATC G ACAAG 2675
138 CAUGU UGAUG GCAUGCACUAUGC GCG GGAUAUCCUG 2511 CAGGATATCC G ACATG 2676
143 GGCUU UGAUG GCAUGCACUAUGC GCG AUGUCGGAUA 2512 TATCCGACAT G AAGCC 2677
152 GAAGU UGAUG GCAUGCACUAUGC GCG ACUGGCUUCA 2513 TGAAGCCAGT G ACTTC 2678
195 UAUUU UGAUG GCAUGCACUAUGC GCG GGUUUUUGUU 2514 AACAAAAACC G AAATA 2679
224 AUGGU UGAUG GCAUGCACUAUGC GCG AAAGGGACUG 2515 CAGTCCCTTT G ACCAT 2680
260 AUAGU UGAUG GCAUGCACUAUGC GCG AUUAUCUUCU 2516 AGAAGATAAT G ACTAT 2681
272 ACUAG UGAUG GCAUGCACUAUGC GCG GUUGAUAUAG 2517 CTATATCAAC G CTAGT 2682
280 UUUAU UGAUG GCAUGCACUAUGC GCG AAACUAGCGU 2518 ACGCTAGTTT G ATAAA 2683
331 UUAGG UGAUG GCAUGCACUAUGC GCG AAAGGGCCCU 2519 AGGGCCCTTT G CCTAA 2684
342 GACCG UGAUG GCAUGCACUAUGC GCG AUGUGUUAGG 2520 CCTAACACAT G CGGTC 2685
394 UUGAG UGAUG GCAUGCACUAUGC GCG AUGACGACAC 2521 GTGTCGTCAT G CTCAA 2686
406 UCCAU UGAUG GCAUGCACUAUGC GCG ACUCUGUUGA 2522 TCAACAGAGT G ATGGA 2687
429 GUGCG UGAUG GCAUGCACUAUGC GCG AUUUUAACGA 2523 TCGTTAAAAT G CGCAC 2688
431 UUGUG UGAUG GCAUGCACUAUGC GCG GCAUUUUAAC 2524 GTTAAAATGC G CACAA 2689
466 AAGAU UGAUG GCAUGCACUAUGC GCG AUCUCUUUUU 2525 AAAAAGAGAT G ATCTT 2690
473 GUCUU UGAUG GCAUGCACUAUGC GCG AAAGAUCAUC 2526 GATGATCTTT G AAGAC 2691
487 AAUUU UGAUG GCAUGCACUAUGC GCG AAAUUUGUGU 2527 ACACAAATTT G AAATT 2692
499 GAGAU UGAUG GCAUGCACUAUGC GCG AAUGUUAAUU 2528 AATTAACATT G ATCTC 2693
506 AUCUU UGAUG GCAUGCACUAUGC GCG AGAGAUCAAU 2529 ATTGATCTCT G AAGAT 2694
532 UGUCG UGAUG GCAUGCACUAUGC GCG ACUGUAUAAU 2530 ATTATACAGT G CGACA 2695
534 GCUGU UGAUG GCAUGCACUAUGC GCG GCACUGUAUA 2531 TATACAGTGC G ACAGC 2696
573 UCUCU UGAUG GCAUGCACUAUGC GCG GAGUUUCUUG 2532 CAAGAAACTC G AGAGA 2697
Table 5
608 AAAGU UGAUG GCAUGCACUAUGC GCG AGGCCAUGUG 2533 CACATGGCCT G ACTTT 2698
623 UGAUU UGAUG GCAUGCACUAUGC GCG AGGGACUCCA 2534 TGGAGTCCCT G AATCA 2699
643 AAGUU UGAUG GCAUGCACUAUGC GCG AAGAAUGAGG 2535 CCTCATTCTT G AACTT 2700
663 ACUCU UGAUG GCAUGCACUAUGC GCG GGACUUUGAA 2536 TTCAAAGTCC G AGAGT 2701
706 CAGUG UGAUG GCAUGCACUAUGC GCG ACCACAACGG 2537 CCGTTGTGGT G CACTG 2702
711 CACUG UGAUG GCAUGCACUAUGC GCG AGUGCACCAC 2538 GTGGTGCACT G CAGTG 2703
716 GCCUG UGAUG GCAUGCACUAUGC GCG ACUGCAGUGC 2539 GCACTGCAGT G CAGGC 2704
752 GGUAU UGAUG GCAUGCACUAUGC GCG AGCCAGACAG 2540 CTGTCTGGCT G ATACC 2705
759 AGAGG UGAUG GCAUGCACUAUGC GCG AGGUAUCAGC 2541 GCTGATACCT G CCTCT 2706
766 AUCAG UGAUG GCAUGCACUAUGC GCG AAGAGGCAGG 2542 CCTGCCTCTT G CTGAT 2707
769 UCCAU UGAUG GCAUGCACUAUGC GCG AGCAAGAGGC 2543 GCCTCTTGCT G ATGGA 2708
800 GAUAU UGAUG GCAUGCACUAUGC GCG AACGGAAGAA 2544 TTCTTCCGTT G ATATC 2709
814 AACAG UGAUG GCAUGCACUAUGC GCG ACUUUCUUGA 2545 TCAAGAAAGT G CTGTT 2710
826 UUCCU UGAUG GCAUGCACUAUGC GCG AUUUCUAACA 2546 TGTTAGAAAT G AGGAA 2711
847 UGGAU UGAUG GCAUGCACUAUGC GCG AGCCCCAUCC 2547 GGATGGGGCT G ATCCA 2712
860 CUGGU UGAUG GCAUGCACUAUGC GCG GGCUGUCUGG 2548 CCAGACAGCC G ACCAG 2713
868 AAGCG UGAUG GCAUGCACUAUGC GCG AGCUGGUCGG 2549 CCGACCAGCT G CGCTT 2714
870 AGAAG UGAUG GCAUGCACUAUGC GCG GCAGCUGGUC 2550 GACCAGCTGC G CTTCT 2715
889 UCGAU UGAUG GCAUGCACUAUGC GCG ACAGCCAGGU 2551 ACCTGGCTGT G ATCGA 2716
893 ACCUU UGAUG GCAUGCACUAUGC GCG GAUCACAGCC 2552 GGCTGTGATC G AAGGT 2717
899 UUUGG UGAUG GCAUGCACUAUGC GCG ACCUUCGAUC 2553 GATCGAAGGT G CCAAA 2718
928 UCCUG UGAUG GCAUGCACUAUGC GCG ACGGAAGAGU 2554 ACTCTTCCGT G CAGGA 2719
956 GUCCU UGAUG GCAUGCACUAUGC GCG GUGGGAAAGC 2555 GCTTTCCCAC G AGGAC 2720
977 AUGCU UGAUG GCAUGCACUAUGC GCG GGGUGGGGGC 2556 GCCCCCACCC G AGCAT 2721
1011 GGAUU UGAUG GCAUGCACUAUGC GCG GUUUGGGUGG 2557 CCACCCAAAC G AATCC 2722
1038 CCCUG UGAUG GCAUGCACUAUGC GCG AUUUCCCAUU 2558 AATGGGAAAT G CAGGG 2723
1069 UCCUU UGAUG GCAUGCACUAUGC GCG ACCCACUGGU 2559 ACCAGTGGGT G AAGGA 2724
1098 UGGGG UGAUG GCAUGCACUAUGC GCG AGUCUUUAUC 2560 GATAAAGACT G CCCCA 2725
1133 UGCGG UGAUG GCAUGCACUAUGC GCG AUUUAAGGGG 2561 CCCCTTAAAT G CCGCA 2726
1136 GGGUG UGAUG GCAUGCACUAUGC GCG GGCAUUUAAG 2562 CTTAAATGCC G CACCC 2727
1151 GCUUU UGAUG GCAUGCACUAUGC GCG GAUGCCGUAG 2563 CTACGGCATC G AAAGC 2728
Table 5
1159 UGACU UGAUG GCAUGCACUAUGC GCG AUGCUUUCGA 2564 TCGAAAGCAT G AGTCA 2729
1172 AACUU UGAUG GCAUGCACUAUGC GCG AGUGUCUUGA 2565 TCAAGACACT G AAGTT 2730
1206 CACCU UGAUG GCAUGCACUAUGC GCG GAAGACUUCC 2566 GGAAGTCTTC G AGGTG 2731
1211 CUGGG UGAUG GCAUGCACUAUGC GCG ACCUCGAAGA 2567 TCTTCGAGGT G CCCAG 2732
1220 GGAGG UGAUG GCAUGCACUAUGC GCG AGCCUGGGCA 2568 TGCCCAGGCT G CCTCC 2733
1249 UCGGG UGAUG GCAUGCACUAUGC GCG AGUGACGGCU 2569 AGCCGTCACT G CCCGA 2734
1253 CUUCU UGAUG GCAUGCACUAUGC GCG GGGCAGUGAC 2570 GTCACTGCCC G AGAAG 2735
1262 GUCCU UGAUG GCAUGCACUAUGC GCG GUCCUUCUCG 2571 CGAGAAGGAC G AGGAC 2736
1271 CAGUG UGAUG GCAUGCACUAUGC GCG AUGGUCCUCG 2572 CGAGGACCAT G CACTG 2737
1276 UAACU UGAUG GCAUGCACUAUGC GCG AGUGCAUGGU 2573 ACCATGCACT G AGTTA 2738
1308 CCACG UGAUG GCAUGCACUAUGC GCG ACAUGUUGAC 2574 GTCAACATGT G CGTGG 2739
1334 GUAAG UGAUG GCAUGCACUAUGC GCG GCCGGCCGUG 2575 CACGGCCGGC G CTTAC 2740
1344 UGUAG UGAUG GCAUGCACUAUGC GCG AGAGGUAAGC 2576 GCTTACCTCT G CTACA 2741
1379 AGGGU UGAUG GCAUGCACUAUGC GCG AGGCUAUGUG 2577 CACATAGCCT G ACCCT 2742
1412 AGAGG UGAUG GCAUGCACUAUGC GCG GGACAGUGGG 2578 CCCACTGTCC G CCTCT 2743
1418 GCGGG UGAUG GCAUGCACUAUGC GCG AGAGGCGGAC 2579 GTCCGCCTCT G CCCGC 2744
1422 CUCUG UGAUG GCAUGCACUAUGC GCG GGGCAGAGGC 2580 GCCTCTGCCC G CAGAG 2745
1433 UCGGG UGAUG GCAUGCACUAUGC GCG GUGGGCUCUG 2581 CAGAGCCCAC G CCCGA 2746
1437 CUAGU UGAUG GCAUGCACUAUGC GCG GGGCGUGGGC 2582 GCCCACGCCC G ACTAG 2747
1450 CGCGG UGAUG GCAUGCACUAUGC GCG AUGCCUGCUA 2583 TAGCAGGCAT G CCGCG 2748
1453 UACCG UGAUG GCAUGCACUAUGC GCG GGCAUGCCUG 2584 CAGGCATGCC G CGGTA 2749
1469 UCCGG UGAUG GCAUGCACUAUGC GCG GGCCCUUACC 2585 GGTAAGGGCC G CCGGA 2750
1477 CUACG UGAUG GCAUGCACUAUGC GCG GGUCCGGCGG 2586 CCGCCGGACC G CGTAG 2751
1513 UAGUG UGAUG GCAUGCACUAUGC GCG AGAACCAACG 2587 CGTTGGTTCT G CACTA 2752
1569 AGGGG UGAUG GCAUGCACUAUGC GCG AAAAAGUAAA 2588 TTTACTTTTT G CCCCT 2753
1583 GUACU UGAUG GCAUGCACUAUGC GCG AAAGUGGAAG 2589 CTTCCACTTT G AGTAC 2754
1610 CUCCU UGAUG GCAUGCACUAUGC GCG AAAAAAUGGC 2590 GCCATTTTTT G AGGAG 2755
1619 UCUUU UGAUG GCAUGCACUAUGC GCG ACUCUCCUCA 2591 TGAGGAGAGT G AAAGA 2756
1634 GCCAG UGAUG GCAUGCACUAUGC GCG AUGGUACUCU 2592 AGAGTACCAT G CTGGC 2757
1643 CUCUG UGAUG GCAUGCACUAUGC GCG GCCGCCAGCA 2593 TGCTGGCGGC G CAGAG 2758
1678 UGGGG UGAUG GCAUGCACUAUGC GCG GAGCCCCAAG 2594 CTTGGGGCTC G CCCCA 2759
Table 5
1723 UCUGG UGAUG GCAUGCACUAUGC GCG GUGCCGCCCG 2595 CGGGCGGCAC G CCAGA 2760
1742 AGAUU , UGAUG GCAUGCACUAUGC GCG AAGGGGGGGG 2596 CCCCCCCCTT G AATCT 2761
1748 CCCUG UGAUG GCAUGCACUAUGC GCG AGAUUCAAGG 2597 CCTTGAATCT G CAGGG 2762
1811 UAGUG UGAUG GCAUGCACUAUGC GCG ACUAUGGAUG 2598 CATCCATAGT G CACTA 2763
1827 UGGUU UGAUG GCAUGCACUAUGC GCG AAGAAAAUGC 2599 GCATTTTCTT G AACCA 2764
1853 GACAU UGAUG GCAUGCACUAUGC GCG AAAAAAUUUU 2600 AAAATTTTTT G ATGTC 2765
1865 UGAUG UGAUG GCAUGCACUAUGC GCG AAGGCUGACA 2601 TGTCAGCCTT G CATCA 2766
1931 CAUGG UGAUG GCAUGCACUAUGC GCG AAAGCCUUCC 2602 GGAAGGCTTT G CCATG 2767
1942 CGCAG UGAUG GCAUGCACUAUGC GCG AGGCCCAUGG 2603 CCATGGGCCT G CTGCG 2768
1945 UGACG UGAUG GCAUGCACUAUGC GCG AGCAGGCCCA 2604 TGGGCCTGCT G CGTCA 2769
1997 AAUAU UGAUG GCAUGCACUAUGC GCG ACUAAAUAAC 2605 GTTATTTAGT G ATATT 2770
2014 CUUCU UGAUG GCAUGCACUAUGC GCG ACGUUACCCA 2606 TGGGTAACGT G AGAAG 2771
2030 UAUAG UGAUG GCAUGCACUAUGC GCG AUUGUUCUAU 2607 ATAGAACAAT G CTATA 2772
2045 GUGUU UGAUG GCAUGCACUAUGC GCG AUUAUAUAUU 2608 AATATATAAT G AACAC 2773
2073 CACAU UGAUG GCAUGCACUAUGC GCG AUGUUUCUUA 2609 TAAGAAACAT G ATGTG 2774
2078 AAUCU UGAUG GCAUGCACUAUGC GCG ACAUCAUGUU 2610 AACATGATGT G AGATT 2775
2094 AUAAG UGAUG GCAUGCACUAUGC GCG GGGACAAAGU 2611 ACTTTGTCCC G CTTAT 2776
2103 GGGAG UGAUG GCAUGCACUAUGC GCG AGAAUAAGCG 2612 CGCTTATTCT G CTCCC 2777
2117 UCUAG UGAUG GCAUGCACUAUGC GCG AGAUAACAGG 2613 CCTGTTATCT G CTAGA 2778
2140 GGGAG UGAUG GCAUGCACUAUGC GCG AGUGAUUGAG 2614 CTCAATCACT G CTCCC 2779
2162 ACAUG UGAUG GCAUGCACUAUGC GCG AUUCUAAUAC 2615 GTATTAGAAT G CATGT 2780
2186 UUCAU UGAUG GCAUGCACUAUGC GCG AGGACACAAG 2616 CTTGTGTCCT G ATGAA 2781
2189 UUUUU UGAUG GCAUGCACUAUGC GCG AUCAGGACAC 2617 GTGTCCTGAT G AAAAA 2782
2200 UCAAG UGAUG GCAUGCACUAUGC GCG ACAUAUUUUU 2618 AAAAATATGT G CTTGA 2783
2204 CAUUU UGAUG GCAUGCACUAUGC GCG AAGCACAUAU 2619 ATATGTGCTT G AAATG 2784
2209 UUUCU UGAUG GCAUGCACUAUGC GCG AUUUCAAGCA 2620 TGCTTGAAAT G AGAAA 2785
2219 GAGAU UGAUG GCAUGCACUAUGC GCG AAAGUUUCUC 2621 GAGAAACTTT G ATCTC 2786
2226 GUAAG UGAUG GCAUGCACUAUGC GCG AGAGAUCAAA 2622 TTTGATCTCT G CTTAC 2787
2238 UGGGG UGAUG GCAUGCACUAUGC GCG ACAUUAGUAA 2623 TTACTAATGT G CCCCA 2788
2260 ACAGG UGAUG GCAUGCACUAUGC GCG AGGUUGGACU 2624 AGTCCAACCT G CCTGT 2789
2266 UCAUG UGAUG GCAUGCACUAUGC GCG ACAGGCAGGU 2625 ACCTGCCTGT G CATGA 2790
Table 5
2270 CAGGU UGAUG GCAUGCACUAUGC GCG AUGCACAGGC 2626 GCCTGTGCAT G ACCTG 2791
2275 AUGAU UGAUG GCAUGCACUAUGC GCG AGGUCAUGCA 2627 TGCATGACCT G ATCAT 2792
2308 UUCAG UGAUG GCAUGCACUAUGC GCG AACAGGCUUA 2628 TAAGCCTGTT G CTGAA 2793
2311 GACUU UGAUG GCAUGCACUAUGC GCG AGCAACAGGC 2629 GCCTGTTGCT G AAGTC 2794
2323 CUGAG UGAUG GCAUGCACUAUGC GCG GACAAUGACU 2630 AGTCATTGTC G CTCAG 2795
2338 AACUG UGAUG GCAUGCACUAUGC GCG ACCCUAUUGC 2631 GCAATAGGGT G CAGTT 2796
2362 UUAGG UGAUG GCAUGCACUAUGC GCG AAAUGCCUAU 2632 ATAGGCATTT G CCTAA 2797
2378 AGUGU UGAUG GCAUGCACUAUGC GCG AUGCCAGGAA 2633 TTCCTGGCAT G ACACT 2798
2389 GAAGU UGAUG GCAUGCACUAUGC GCG ACUAGAGUGU 2634 ACACTCTAGT G ACTTC 2799
2400 GGCCU UGAUG GCAUGCACUAUGC GCG ACCAGGAAGU 2635 ACTTCCTGGT G AGGCC 2800
2433 UACAG UGAUG GCAUGCACUAUGC GCG AAGACCCUGC 2636 GCAGGGTCTT G CTGTA 2801
2483 CAGAG UGAUG GCAUGCACUAUGC GCG GUGAGGUGUG 2637 CACACCTCAC G CTCTG 2802
2494 UAAAU UGAUG GCAUGCACUAUGC GCG AUGUCCAGAG 2638 CTCTGGACAT G ATTTA 2803
2520 GGGGG UGAUG GCAUGCACUAUGC GCG GGGGGGUGUC 2639 GACACCCCCC G CCCCC 2804
2546 AAUGG UGAUG GCAUGCACUAUGC GCG GGAGGCUGAU 2640 ATCAGCCTCC G CCATT 2805
2559 AGUGU UGAUG GCAUGCACUAUGC GCG GACUUGGAAU 2641 ATTCCAAGTC G ACACT 2806
2571 CUGCU UGAUG GCAUGCACUAUGC GCG AAGAAGAGUG 2642 CACTCTTCTT G AGCAG 2807
2582 CAAAU UGAUG GCAUGCACUAUGC GCG ACGGUCUGCU 2643 AGCAGACCGT G ATTTG 2808
2602 UCCAG UGAUG GCAUGCACUAUGC GCG AGGUGCCUCU 2644 AGAGGCACCT G CTGGA 2809
2621 UGUUU UGAUG GCAUGCACUAUGC GCG AAGAAGUGUG 2645 CACACTTCTT G AAACA 2810
2635 ACCGU UGAUG GCAUGCACUAUGC GCG ACCCAGGCUG 2646 CAGCCTGGGT G ACGGT 2811
2655 GGCGG UGAUG GCAUGCACUAUGC GCG AGGCUGCCUA 2647 TAGGCAGCCT G CCGCC 2812
2658 GACGG UGAUG GCAUGCACUAUGC GCG GGCAGGCUGC 2648 GCAGCCTGCC G CCGTC 2813
2682 CUCGG UGAUG GCAUGCACUAUGC GCG AAGGUGAACC 2649 GGTTCACCTT G CCGAG 2814
2685 UCUCU UGAUG GCAUGCACUAUGC GCG GGCAAGGUGA 2650 TCACCTTGCC G AGAGA 2815
2694 AGACG UGAUG GCAUGCACUAUGC GCG GCCUCUCUCG 2651 CGAGAGAGGC G CGTCT 2816
2700 UGGGG UGAUG GCAUGCACUAUGC GCG AGACGCGCCU 2652 AGGCGCGTCT G CCCCA 2817
2727 ACCAU UGAUG GCAUGCACUAUGC GCG AGGCCCCACA 2653 TGTGGGGCCT G ATGGT 2818
2733 GUGAG UGAUG GCAUGCACUAUGC GCG ACCAUCAGGC 2654 GCCTGATGGT G CTCAC 2819
2739 AGAGU UGAUG GCAUGCACUAUGC GCG GUGAGCACCA 2655 TGGTGCTCAC G ACTCT 2820
2749 CUUUG UGAUG GCAUGCACUAUGC GCG AGGAAGAGUC 2656 GACTCTTCCT G CAAAG 2821
Table 5
2761 GUCUU UGAUG GCAUGCACUAUGC GCG AGUUCCCUUU 2657 AAAGGGAACT G AAGAC 2822
2793 UUUUU UGAUG GCAUGCACUAUGC GCG AUGUUAAAAA 2658 TTTTTAACAT G AAAAA 2823
2818 UAGCU UGAUG GCAUGCACUAUGC GCG GGGAGCUACA 2659 TGTAGCTCCC G AGCTA 2824
2831 GCUGG UGAUG GCAUGCACUAUGC GCG AAGAGAGUAG 2660 CTACTCTCTT G CCAGC 2825
2850 AAAGG UGAUG GCAUGCACUAUGC GCG AAAAUGUGAA 2661 TTCACATTTT G CCTTT 2826
2902 CACCU UGAUG GCAUGCACUAUGC GCG GAAUGUUCCC 2662 GGGAACATTC G AGGTG 2827
2915 CUCUG UGAUG GCAUGCACUAUGC GCG AGGGUGACAC 2663 GTGTCACCCT G CAGAG 2828
2928 CACCU UGAUG GCAUGCACUAUGC GCG ACCAUAGCUC 2664 GAGCTATGGT G AGGTG 2829
2950 CCUGG UGAUG GCAUGCACUAUGC GCG ACCUAAGCCU 2665 AGGCTTAGGT G CCAGG 2830
2969 CAGCU UGAUG GCAUGCACUAUGC GCG AGAAUGCUUA 2666 TAAGCATTCT G AGCTG 2831
3123 CAGCU UGAUG GCAUGCACUAUGC GCG GCUACCUCUC 2667 GAGAGGTAGC G AGCTG 2832
3128 CAGAG UGAUG GCAUGCACUAUGC GCG AGCUCGCUAC 2668 GTAGCGAGCT G CTCTG 2833
3133 CAUAG UGAUG GCAUGCACUAUGC GCG AGAGCAGCUC 2669 GAGCTGCTCT G CTATG 2834
Table 6
Table 6: Human PTP-IB DNAzyme and Target Sequence
Table 6
Table 6
275 AACTAGCG GGCTAGCTACAACGA TGATATAG 2894 CUAUAUCA A CGCUAGUU 3604
277 CAAACTAG GGCTAGCTACAACGA GTTGATAT 2895 AUAUCAAC G CUAGUUUG 3605
281 TTATCAAA GGCTAGCTACAACGA TAGCGTTG 2896 CAACGCUA G UUUGAUAA 3606
286 CATTTTTA GGCTAGCTACAACGA CAAACTAG 2897 CUAGUUUG A UAAAAAUG 3607
292 TTCTTCCA GGCTAGCTACAACGA TTTTATCA 2898 UGAUAAAA A UGGAAGAA 3608
301 CCTTTGGG GGCTAGCTACAACGA TTCTTCCA 2899 UGGAAGAA G CCCAAAGG 3609
311 GAATGTAA GGCTAGCTACAACGA TCCTTTGG 2900 CCAAAGGA G UUACAUUC 3610
314 TAAGAATG GGCTAGCTACAACGA AACTCCTT 2901 AAGGAGUU A CAUUCUUA 3611
316 GGTAAGAA GGCTAGCTACAACGA GTAACTCC 2902 GGAGUUAC A UUCUUACC 3612
322 GCCCTGGG GGCTAGCTACAACGA AAGAATGT 2903 ACAUUCUU A CCCAGGGC 3613
329 GCAAAGGG GGCTAGCTACAACGA CCTGGGTA 2904 UACCCAGG G CCCUUUGC 3614
336 GTGTTAGG GGCTAGCTACAACGA AAAGGGCC 2905 GGCCCUUU G CCUAACAC 3615
341 CGCATGTG GGCTAGCTACAACGA TAGGCAAA 2906 UUUGCCUA A CACAUGCG 3616
343 ACCGCATG GGCTAGCTACAACGA GTTAGGCA 2907 UGCCUAAC A CAUGCGGU 3617
345 TGACCGCA GGCTAGCTACAACGA GTGTTAGG 2908 CCUAACAC A UGCGGUCA 3618
347 AGTGACCG GGCTAGCTACAACGA ATGTGTTA 2909 UAACACAU G CGGUCACU 3619
350 AAAAGTGA GGCTAGCTACAACGA CGCATGTG 2910 CACAUGCG G UCACUUUU 3620
353 CCCAAAAG GGCTAGCTACAACGA GACCGCAT 2911 AUGCGGUC A CUUUUGGG 3621
364 CCACACCA GGCTAGCTACAACGA CTCCCAAA 2912 UUUGGGAG A UGGUGUGG 3622
367 CTCCCACA GGCTAGCTACAACGA CATCTCCC 2913 GGGAGAUG G UGUGGGAG 3623
369 TGCTCCCA GGCTAGCTACAACGA ACCATCTC 2914 GAGAUGGU G UGGGAGCA 3624
375 CTTTTCTG GGCTAGCTACAACGA TCCCACAC 2915 GUGUGGGA G CAGAAAAG 3625
383 CACCCCTG GGCTAGCTACAACGA TTTTCTGC 2916 GCAGAAAA G CAGGGGUG 3626
389 TGACGACA GGCTAGCTACAACGA CCCTGCTT 2917 AAGCAGGG G UGUCGUCA 3627
391 CATGACGA GGCTAGCTACAACGA ACCCCTGC 2918 GCAGGGGU G UCGUCAUG 3628
394 GAGCATGA GGCTAGCTACAACGA GACACCCC 2919 GGGGUGUC G UCAUGCUC 3629
397 GTTGAGCA GGCTAGCTACAACGA GACGACAC 2920 GUGUCGUC A UGCUCAAC 3630
399 CTGTTGAG GGCTAGCTACAACGA ATGACGAC 2921 GUCGUCAU G CUCAACAG 3631
404 TCACTCTG GGCTAGCTACAACGA TGAGCATG 2922 CAUGCUCA A CAGAGUGA 3632
409 CTCCATCA GGCTAGCTACAACGA TCTGTTGA 2923 UCAACAGA G UGAUGGAG ' 3633
412 TTTCTCCA GGCTAGCTACAACGA CACTCTGT 2924 ACAGAGUG A UGGAGAAA 3634
Table 6
422 TTAACGAA GGCTAGCTACAACGA CTTTCTCC 2925 GGAGAAAG G UUCGUUAA 3635
426 CATTTTAA GGCTAGCTACAACGA GAACCTTT 2926 AAAGGUUC G UUAAAAUG 3636
432 TGTGCGCA GGCTAGCTACAACGA TTTAACGA 2927 UCGUUAAA A UGCGCACA 3637
434 ATTGTGCG GGCTAGCTACAACGA ATTTTAAC 2928 GUUAAAAU G CGCACAAU 3638
436 GTATTGTG GGCTAGCTACAACGA GCATTTTA 2929 UAAAAUGC G CACAAUAC 3639
438 CAGTATTG GGCTAGCTACAACGA GCGCATTT 2930 AAAUGCGC A CAAUACUG 3640
441 GGCCAGTA GGCTAGCTACAACGA TGTGCGCA 2931 UGCGCACA A UACUGGCC 3641
443 GTGGCCAG GGCTAGCTACAACGA ATTGTGCG 2932 CGCACAAU A CUGGCCAC 3642
447 TTTTGTGG GGCTAGCTACAACGA CAGTATTG 2933 CAAUACUG G CCACAAAA 3643
450 TCTTTTTG GGCTAGCTACAACGA GGCCAGTA 2934 UACUGGCC A CAAAAAGA 3644
469 AAAGATCA GGCTAGCTACAACGA CTCTTTTT 2935 AAAAAGAG A UGAUCUUU 3645
472 TTCAAAGA GGCTAGCTACAACGA CATCTCTT 2936 AAGAGAUG A UCUUUGAA 3646
482 AATTTGTG GGCTAGCTACAACGA CTTCAAAG 2937 CUUUGAAG A CACAAAUU 3647
484 CAAATTTG GGCTAGCTACAACGA GTCTTCAA 2938 UUGAAGAC A CAAAUUUG 3648
488 ATTTCAAA GGCTAGCTACAACGA TTGTGTCT 2939 AGACACAA A UUUGAAAU 3649
495 AATGTTAA GGCTAGCTACAACGA TTCAAATT 2940 AAUUUGAA A UUAACAUU 3650
499 GATCAATG GGCTAGCTACAACGA TAATTTCA 2941 UGAAAUUA A CAUUGAUC 3651
501 GAGATCAA GGCTAGCTACAACGA GTTAATTT 2942 AAAUUAAC A UUGAUCUC 3652
505 TTCAGAGA GGCTAGCTACAACGA CAATGTTA 2943 UAACAUUG A UCUCUGAA 3653
515 ACTTGATA GGCTAGCTACAACGA CTTCAGAG 2944 CUCUGAAG A UAUCAAGU 3654
517 TGACTTGA GGCTAGCTACAACGA ATCTTCAG 2945 CUGAAGAU A UCAAGUCA 3655
522 TAATATGA GGCTAGCTACAACGA TTGATATC 2946 GAUAUCAA G UCAUAUUA 3656
525 GTATAATA GGCTAGCTACAACGA GACTTGAT 2947 AUCAAGUC A UAUUAUAC 3657
527 CTGTATAA GGCTAGCTACAACGA ATGACTTG 2948 CAAGUCAU A UUAUACAG 3658
530 GCACTGTA GGCTAGCTACAACGA AATATGAC 2949 GUCAUAUU A UACAGUGC 3659
532 TCGCACTG GGCTAGCTACAACGA ATAATATG 2950 CAUAUUAU A CAGUGCGA 3660
535 CTGTCGCA GGCTAGCTACAACGA TGTATAAT 2951 AUUAUACA G UGCGACAG 3661
537 AGCTGTCG GGCTAGCTACAACGA ACTGTATA 2952 UAUACAGU G CGACAGCU 3662
540 TCTAGCTG GGCTAGCTACAACGA CGCACTGT 2953 ACAGUGCG A CAGCUAGA 3663
543 AATTCTAG GGCTAGCTACAACGA TGTCGCAC 2954 GUGCGACA G CUAGAAUU 3664
549 TTTTCCAA GGCTAGCTACAACGA TCTAGCTG 2955 CAGCUAGA A UUGGAAAA 3665
Table 6
557 TTGTAAGG GGCTAGCTACAACGA TTTCCAAT 2956 AUUGGAAA A CCUUACAA 3666
562 TTGGGTTG GGCTAGCTACAACGA AAGGTTTT 2957 AAAACCUU A CAACCCAA 3667
565 TTCTTGGG GGCTAGCTACAACGA TGTAAGGT 2958 ACCUUACA A CCCAAGAA 3668
574 CTCTCGAG GGCTAGCTACAACGA TTCTTGGG 2959 CCCAAGAA A CUCGAGAG 3669
583 ATGTAAGA GGCTAGCTACAACGA CTCTCGAG 2960 CUCGAGAG A UCUUACAU 3670
588 TGGAAATG GGCTAGCTACAACGA AAGATCTC 2961 GAGAUCUU A CAUUUCCA 3671
590 AGTGGAAA GGCTAGCTACAACGA GTAAGATC 2962 GAUCUUAC A UUUCCACU 3672
596 TGGTATAG GGCTAGCTACAACGA GGAAATGT 2963 ACAUUUCC A CUAUACCA 3673
599 ATGTGGTA GGCTAGCTACAACGA AGTGGAAA 2964 UUUCCACU A UACCACAU 3674
601 CCATGTGG GGCTAGCTACAACGA ATAGTGGA 2965 UCCACUAU A CCACAUGG 3675
604 AGGCCATG GGCTAGCTACAACGA GGTATAGT 2966 ACUAUACC A CAUGGCCU 3676
606 TCAGGCCA GGCTAGCTACAACGA GTGGTATA 2967 UAUACCAC A UGGCCUGA 3677
609 AAGTCAGG GGCTAGCTACAACGA CATGTGGT 2968 ACCACAUG G CCUGACUU 3678
614 CTCCAAAG GGCTAGCTACAACGA CAGGCCAT 2969 AUGGCCUG A CUUUGGAG 3679
622 TTCAGGGA GGCTAGCTACAACGA TCCAAAGT 2970 ACUUUGGA G UCCCUGAA 3680
630 GCTGGTGA GGCTAGCTACAACGA TCAGGGAC 2971 GUCCCUGA A UCACCAGC 3681
633 GAGGCTGG GGCTAGCTACAACGA GATTCAGG 2972 CCUGAAUC A CCAGCCUC 3682
637 GAATGAGG GGCTAGCTACAACGA TGGTGATT 2973 AAUCACCA G CCUCAUUC 3683
642 TTCAAGAA GGCTAGCTACAACGA GAGGCTGG 2974 CCAGCCUC A UUCUUGAA 3684
650 AAAGAAAG GGCTAGCTACAACGA TCAAGAAT 2975 AUUCUUGA A CUUUCUUU 3685
664 CTCTCGGA GGCTAGCTACAACGA TTTGAAAA 2976 UUUUCAAA G UCCGAGAG 3686
672 GACCCTGA GGCTAGCTACAACGA TCTCGGAC 2977 GUCCGAGA G UCAGGGUC 3687
678 CTGAGTGA GGCTAGCTACAACGA CCTGACTC 2978 GAGUCAGG G UCACUCAG 3688
681 GGGCTGAG GGCTAGCTACAACGA GACCCTGA 2979 UCAGGGUC A CUCAGCCC 3689
686 GCTCCGGG GGCTAGCTACAACGA TGAGTGAC 2980 GUCACUCA G CCCGGAGC 3690
693 GGCCCGTG GGCTAGCTACAACGA TCCGGGCT 2981 AGCCCGGA G CACGGGCC 3691
695 CGGGCCCG GGCTAGCTACAACGA GCTCCGGG 2982 CCCGGAGC A CGGGCCCG 3692
699 ACAACGGG GGCTAGCTACAACGA CCGTGCTC 2983 GAGCACGG G CCCGUUGU 3693
703 CACCACAA GGCTAGCTACAACGA GGGCCCGT 2984 ACGGGCCC G UUGUGGUG 3694
706 GTGCACCA GGCTAGCTACAACGA AACGGGCC 2985 GGCCCGUU G UGGUGCAC 3695
709 GCAGTGCA GGCTAGCTACAACGA CACAACGG 2986 CCGUUGUG G UGCACUGC 3696
Table 6
711 CTGCAGTG GGCTAGCTACAACGA ACCACAAC 2987 GUUGUGGU G CACUGCAG 3697
713 CACTGCAG GGCTAGCTACAACGA GCACCACA 2988 UGUGGUGC A CUGCAGUG 3698
716 CTGCACTG GGCTAGCTACAACGA AGTGCACC 2989 GGUGCACU G CAGUGCAG 3699
719 TGCCTGCA GGCTAGCTACAACGA TGCAGTGC 2990 GCACUGCA G UGCAGGCA 3700
721 GATGCCTG GGCTAGCTACAACGA ACTGCAGT 2991 ACUGCAGU G CAGGCAUC 3701
725 TGCCGATG GGCTAGCTACAACGA CTGCACTG 2992 CAGUGCAG G CAUCGGCA 3702
727 CCTGCCGA GGCTAGCTACAACGA GCCTGCAC 2993 GUGCAGGC A UCGGCAGG 3703
731 CAGACCTG GGCTAGCTACAACGA CGATGCCT 2994 AGGCAUCG G CAGGUCUG 3704
735 GTTCCAGA GGCTAGCTACAACGA CTGCCGAT 2995 AUCGGCAG G UCUGGAAC 3705
742 ACAGAAGG GGCTAGCTACAACGA TCCAGACC 2996 GGUCUGGA A CCUUCUGU 3706
749 CAGCCAGA GGCTAGCTACAACGA AGAAGGTT 2997 AACCUUCU G UCUGGCUG 3707
754 GGTATCAG GGCTAGCTACAACGA CAGACAGA 2998 UCUGUCUG G CUGAUACC 3708
758 GGCAGGTA GGCTAGCTACAACGA CAGCCAGA 2999 UCUGGCUG A UACCUGCC 3709
760 GAGGCAGG GGCTAGCTACAACGA ATCAGCCA 3000 UGGCUGAU A CCUGCCUC 3710
764 GCAAGAGG GGCTAGCTACAACGA AGGTATCA 3001 UGAUACCU G CCUCUUGC 3711
771 TCCATCAG GGCTAGCTACAACGA AAGAGGCA 3002 UGCCUCUU G CUGAUGGA 3712
775 CTTGTCCA GGCTAGCTACAACGA CAGCAAGA 3003 UCUUGCUG A UGGACAAG 3713
779 TCCTCTTG GGCTAGCTACAACGA CCATCAGC 3004 GCUGAUGG A CAAGAGGA 3714
791 AAGAAGGG GGCTAGCTACAACGA CTTTCCTC 3005 GAGGAAAG A CCCUUCUU 3715
802 GATATCAA GGCTAGCTACAACGA GGAAGAAG 3006 CUUCUUCC G UUGAUAUC 3716
806 TCTTGATA GGCTAGCTACAACGA CAACGGAA 3007 UUCCGUUG A UAUCAAGA 3717
808 TTTCTTGA GGCTAGCTACAACGA ATCAACGG 3008 CCGUUGAU A UCAAGAAA 3718
817 TAACAGCA GGCTAGCTACAACGA TTTCTTGA 3009 UCAAGAAA G UGCUGUUA 3719
819 TCTAACAG GGCTAGCTACAACGA ACTTTCTT 3010 AAGAAAGU G CUGUUAGA 3720
822 ATTTCTAA GGCTAGCTACAACGA AGCACTTT 3011 AAAGUGCU G UUAGAAAU 3721
829 CTTCCTCA GGCTAGCTACAACGA TTCTAACA 3012 UGUUAGAA A UGAGGAAG 3722
837 ATCCGAAA GGCTAGCTACAACGA TTCCTCAT 3013 AUGAGGAA G UUUCGGAU 3723
844 CAGCCCCA GGCTAGCTACAACGA CCGAAACT 3014 AGUUUCGG A UGGGGCUG 3724
849 TGGATCAG GGCTAGCTACAACGA CCCATCCG 3015 CGGAUGGG G CUGAUCCA 3725
853 TGTCTGGA GGCTAGCTACAACGA CAGCCCCA 3016- UGGGGCUG A UCCAGACA 3726
859 GTCGGCTG GGCTAGCTACAACGA CTGGATCA 3017 UGAUCCAG A CAGCCGAC 3727
Table 6
Table 6
1018 CTCCAGGA GGCTAGCTACAACGA TCGTTTGG 3049 CCAAACGA A UCCUGGAG 3759
1026 TTGTGTGG GGCTAGCTACAACGA TCCAGGAT 3050 AUCCUGGA G CCACACAA 3760
1029 CCATTGTG GGCTAGCTACAACGA GGCTCCAG 3051 CUGGAGCC A CACAAUGG 3761
1031 TCCCATTG GGCTAGCTACAACGA GTGGCTCC 3052 GGAGCCAC A CAAUGGGA 3762
1034 ATTTCCCA GGCTAGCTACAACGA TGTGTGGC 3053 GCCACACA A UGGGAAAU 3763
1041 TCCCTGCA GGCTAGCTACAACGA TTCCCATT 3054 AAUGGGAA A UGCAGGGA 3764
1043 ACTCCCTG GGCTAGCTACAACGA ATTTCCCA 3055 UGGGAAAU G CAGGGAGU 3765
1050 GGGAAGAA GGCTAGCTACAACGA TCCCTGCA 3056 UGCAGGGA G UUCUUCCC 3766
1061 ACTGGTGA GGCTAGCTACAACGA TTGGGAAG 3057 CUUCCCAA A UCACCAGU 3767
1064 CCCACTGG GGCTAGCTACAACGA GATTTGGG 3058 CCCAAAUC A CCAGUGGG 3768
1068 TTCACCCA GGCTAGCTACAACGA TGGTGATT 3059 AAUCACCA G UGGGUGAA 3769
1072 TTCCTTCA GGCTAGCTACAACGA CCACTGGT 3060 ACCAGUGG G UGAAGGAA 3770
1084 CTCCTGGG GGCTAGCTACAACGA CTCTTCCT 3061 AGGAAGAG A CCCAGGAG 3771
1094 AGTCTTTA GGCTAGCTACAACGA CCTCCTGG 3062 CCAGGAGG A UAAAGACU 3772
1100 TGGGGCAG GGCTAGCTACAACGA CTTTATCC 3063 GGAUAAAG A CUGCCCCA 3773
1103 TGATGGGG GGCTAGCTACAACGA AGTCTTTA 3064 UAAAGACU G CCCCAUCA 3774
1108 TTCCTTGA GGCTAGCTACAACGA GGGGCAGT 3065 ACUGCCCC A UCAAGGAA 3775
1127 TTAAGGGG GGCTAGCTACAACGA TTCCTTTT 3066 AAAAGGAA G CCCCUUAA 3776
1136 GTGCGGCA GGCTAGCTACAACGA TTAAGGGG 3067 CCCCUUAA A UGCCGCAC 3777
1138 GGGTGCGG GGCTAGCTACAACGA ATTTAAGG 3068 CCUUAAAU G CCGCACCC 3778
1141 GTAGGGTG GGCTAGCTACAACGA GGCATTTA 3069 UAAAUGCC G CACCCUAC 3779
1143 CCGTAGGG GGCTAGCTACAACGA GCGGCATT 3070 AAUGCCGC A CCCUACGG 3780
1148 CGATGCCG GGCTAGCTACAACGA AGGGTGCG 3071 CGCACCCU A CGGCAUCG 3781
1151 TTTCGATG GGCTAGCTACAACGA CGTAGGGT 3072 ACCCUACG G CAUCGAAA 3782
1153 GCTTTCGA GGCTAGCTACAACGA GCCGTAGG 3073 CCUACGGC A UCGAAAGC 3783
1160 GACTCATG GGCTAGCTACAACGA TTTCGATG 3074 CAUCGAAA G CAUGAGUC 3784
1162 TTGACTCA GGCTAGCTACAACGA GCTTTCGA 3075 UCGAAAGC A UGAGUCAA 3785
1166 TGTCTTGA GGCTAGCTACAACGA TCATGCTT 3076 AAGCAUGA G UCAAGACA 3786
1172 CTTCAGTG GGCTAGCTACAACGA CTTGACTC 3077 GAGUCAAG A CACUGAAG 3787
1174 AACTTCAG GGCTAGCTACAACGA GTCTTGAC 3078 GUCAAGAC A CUGAAGUU 3788
1180 ACTTCTAA GGCTAGCTACAACGA TTCAGTGT 3079 ACACUGAA G UUAGAAGU 3789
Table 6
1187 CGACCCGA GGCTAGCTACAACGA TTCTAACT 3080 AGUUAGAA G UCGGGUCG 3790
1192 CCCCACGA GGCTAGCTACAACGA CCGACTTC 3081 GAAGUCGG G UCGUGGGG 3791
1195 TCCCCCCA GGCTAGCTACAACGA GACCCGAC 3082 GUCGGGUC G UGGGGGGA 3792
1205 CTCGAAGA GGCTAGCTACAACGA TTCCCCCC 3083 GGGGGGAA G UCUUCGAG 3793
1214 CCTGGGCA GGCTAGCTACAACGA CTCGAAGA 3084 UCUUCGAG G UGCCCAGG 3794
1216 AGCCTGGG GGCTAGCTACAACGA ACCTCGAA 3085 UUCGAGGU G CCCAGGCU 3795
1222 GGAGGCAG GGCTAGCTACAACGA CTGGGCAC 3086 GUGCCCAG G CUGCCUCC 3796
1225 TGGGGAGG GGCTAGCTACAACGA AGCCTGGG 3087 CCCAGGCU G CCUCCCCA 3797
1234 CCCTTTGG GGCTAGCTACAACGA TGGGGAGG 3088 CCUCCCCA G CCAAAGGG 3798
1245 AGTGACGG GGCTAGCTACAACGA TCCCCTTT 3089 AAAGGGGA G CCGUCACU 3799
1248 GGCAGTGA GGCTAGCTACAACGA GGCTCCCC 3090 GGGGAGCC G UCACUGCC 3800
1251 TCGGGCAG GGCTAGCTACAACGA GACGGCTC 3091 GAGCCGUC A CUGCCCGA 3801
1254 TTCTCGGG GGCTAGCTACAACGA AGTGACGG 3092 CCGUCACU G CCCGAGAA 3802
1265 GGTCCTCG GGCTAGCTACAACGA CCTTCTCG 3093 CGAGAAGG A CGAGGACC 3803
1271 GTGCATGG GGCTAGCTACAACGA CCTCGTCC 3094 GGACGAGG A CCAUGCAC 3804
1274 TCAGTGCA GGCTAGCTACAACGA GGTCCTCG 3095 CGAGGACC A UGCACUGA 3805
1276 ACTCAGTG GGCTAGCTACAACGA ATGGTCCT 3096 AGGACCAU G CACUGAGU 3806
1278 TAACTCAG GGCTAGCTACAACGA GCATGGTC 3097 GACCAUGC A CUGAGUUA 3807
1283 TCCAGTAA GGCTAGCTACAACGA TCAGTGCA 3098 UGCACUGA G UUACUGGA 3808
1286 GCTTCCAG GGCTAGCTACAACGA AACTCAGT 3099 ACUGAGUU A CUGGAAGC 3809
1293 AGGAAGGG GGCTAGCTACAACGA TTCCAGTA 3100 UACUGGAA G CCCUUCCU 3810
1303 CATGTTGA GGCTAGCTACAACGA CAGGAAGG 3101 CCUUCCUG G UCAACAUG 3811
1307 CGCACATG GGCTAGCTACAACGA TGACCAGG 3102 CCUGGUCA A CAUGUGCG 3812
1309 CACGCACA GGCTAGCTACAACGA GTTGACCA 3103 UGGUCAAC A UGUGCGUG 3813
1311 GCCACGCA GGCTAGCTACAACGA ATGTTGAC 3104 GUCAACAU G UGCGUGGC 3814
1313 TAGCCACG GGCTAGCTACAACGA ACATGTTG 3105 CAACAUGU G CGUGGCUA 3815
1315 CGTAGCCA GGCTAGCTACAACGA GCACATGT 3106 ACAUGUGC G UGGCUACG 3816
1318 GACCGTAG GGCTAGCTACAACGA CACGCACA 3107 UGUGCGUG G CUACGGUC 3817
1321 GAGGACCG GGCTAGCTACAACGA AGCCACGC 3108 GCGUGGCU A CGGUCCUC 3818
1324 CGTGAGGA GGCTAGCTACAACGA CGTAGCCA 3109 UGGCUACG G UCCUCACG 3819
1330 GCCGGCCG GGCTAGCTACAACGA GAGGACCG 3110 CGGUCCUC A CGGCCGGC 3820
Table 6
1333 AGCGCCGG GGCTAGCTACAACGA CGTGAGGA 3111 UCCUCACG G CCGGCGCU 3821
1337 GGTAAGCG GGCTAGCTACAACGA CGGCCGTG 3112 CACGGCCG G CGCUUACC 3822
1339 GAGGTAAG GGCTAGCTACAACGA GCCGGCCG 3113 CGGCCGGC G CUUACCUC 3823
1343 AGCAGAGG GGCTAGCTACAACGA AAGCGCCG 3114 CGGCGCUU A CCUCUGCU 3824
1349 ACCTGTAG GGCTAGCTACAACGA AGAGGTAA 3115 UUACCUCU G CUACAGGU 3825
1352 GGAACCTG GGCTAGCTACAACGA AGCAGAGG 3116 CCUCUGCU A CAGGUUCC 3826
1356 AACAGGAA GGCTAGCTACAACGA CTGTAGCA 3117 UGCUACAG G UUCCUGUU 3827
1362 CTGTTGAA GGCTAGCTACAACGA AGGAACCT 3118 AGGUUCCU G UUCAACAG 3828
1367 TGTTGCTG GGCTAGCTACAACGA TGAACAGG 3119 CCUGUUCA A CAGCAACA 3829
1370 ATGTGTTG GGCTAGCTACAACGA TGTTGAAC 3120 GUUCAACA G CAACACAU 3830
1373 GCTATGTG GGCTAGCTACAACGA TGCTGTTG 3121 CAACAGCA A CACAUAGC 3831
1375 AGGCTATG GGCTAGCTACAACGA GTTGCTGT 3122 ACAGCAAC A CAUAGCCU 3832
1377 TCAGGCTA GGCTAGCTACAACGA GTGTTGCT 3123 AGCAACAC A UAGCCUGA 3833
1380 GGGTCAGG GGCTAGCTACAACGA TATGTGTT 3124 AACACAUA G CCUGACCC 3834
1385 GAGGAGGG GGCTAGCTACAACGA CAGGCTAT 3125 AUAGCCUG A CCCUCCUC 3835
1395 AGGTGGAG GGCTAGCTACAACGA GGAGGAGG 3126 CCUCCUCC A CUCCACCU 3836
1400 GGTGGAGG GGCTAGCTACAACGA GGAGTGGA 3127 UCCACUCC A CCUCCACC 3837
1406 ACAGTGGG GGCTAGCTACAACGA GGAGGTGG 3128 CCACCUCC A CCCACUGU 3838
1410 GCGGACAG GGCTAGCTACAACGA GGGTGGAG 3129 CUCCACCC A CUGUCCGC 3839
1413 GAGGCGGA GGCTAGCTACAACGA AGTGGGTG 3130 CACCCACU G UCCGCCUC 3840
1417 GGCAGAGG GGCTAGCTACAACGA GGACAGTG 3131 CACUGUCC G CCUCUGCC 3841
1423 TCTGCGGG GGCTAGCTACAACGA AGAGGCGG 3132 CCGCCUCU G CCCGCAGA 3842
1427 GGGCTCTG GGCTAGCTACAACGA GGGCAGAG 3133 CUCUGCCC G CAGAGCCC 3843
1432 GGCGTGGG GGCTAGCTACAACGA TCTGCGGG 3134 CCCGCAGA G CCCACGCC 3844
1436 GTCGGGCG GGCTAGCTACAACGA GGGCTCTG 3135 CAGAGCCC A CGCCCGAC 3845
1438 TAGTCGGG GGCTAGCTACAACGA GTGGGCTC 3136 GAGCCCAC G CCCGACUA 3846
1443 CCTGCTAG' GGCTAGCTACAACGA CGGGCGTG 3137 CACGCCCG A CUAGCAGG 3847
1447 CATGCCTG GGCTAGCTACAACGA TAGTCGGG 3138 CCCGACUA G CAGGCAUG 3848
1451 GCGGCATG GGCTAGCTACAACGA CTGCTAGT 3139 ACUAGCAG G CAUGCCGC 3849
1453 CCGCGGCA GGCTAGCTACAACGA GCCTGCTA 3140 UAGCAGGC A UGCCGCGG 3850
1455 TACCGCGG GGCTAGCTACAACGA ATGCCTGC 3141 GCAGGCAU G CCGCGGUA 3851
Table 6
1458 ACCTACCG GGCTAGCTACAACGA GGCATGCC 3142 GGCAUGCC G CGGUAGGU 3852
1461 CTTACCTA GGCTAGCTACAACGA CGCGGCAT 3143 AUGCCGCG G UAGGUAAG 3853
1465 GGCCCTTA GGCTAGCTACAACGA CTACCGCG 3144 CGCGGUAG G UAAGGGCC 3854
1471 TCCGGCGG GGCTAGCTACAACGA CCTTACCT 3145 AGGUAAGG G CCGCCGGA 3855
1474 CGGTCCGG GGCTAGCTACAACGA GGCCCTTA 3146 UAAGGGCC G CCGGACCG 3856
1479 CTACGCGG GGCTAGCTACAACGA CCGGCGGC 3147 GCCGCCGG A CCGCGUAG 3857
1482 TCTCTACG GGCTAGCTACAACGA GGTCCGGC 3148 GCCGGACC G CGUAGAGA 3858
1484 GCTCTCTA GGCTAGCTACAACGA GCGGTCCG 3149 CGGACCGC G UAGAGAGC 3859
1491 GGGCCCGG GGCTAGCTACAACGA TCTCTACG 3150 CGUAGAGA G CCGGGCCC 3860
1496 GTCCGGGG GGCTAGCTACAACGA CCGGCTCT 3151 AGAGCCGG G CCCCGGAC 3861
1503 AACGTCCG GGCTAGCTACAACGA CCGGGGCC 3152 GGCCCCGG A CGGACGUU 3862
1507 AACCAACG GGCTAGCTACAACGA CCGTCCGG 3153 CCGGACGG A CGUUGGUU 3863
1509 AGAACCAA GGCTAGCTACAACGA GTCCGTCC 3154 GGACGGAC G UUGGUUCU 3864
1513 GTGCAGAA GGCTAGCTACAACGA CAACGTCC 3155 GGACGUUG G UUCUGCAC 3865
1518 TTTTAGTG GGCTAGCTACAACGA AGAACCAA 3156 UUGGUUCU G CACUAAAA 3866
1520 GGTTTTAG GGCTAGCTACAACGA GCAGAACC 3157 GGUUCUGC A CUAAAACC 3867
1526 AAGATGGG GGCTAGCTACAACGA TTTAGTGC 3158 GCACUAAA A CCCAUCUU 3868
1530 GGGGAAGA GGCTAGCTACAACGA GGGTTTTA 3159 UAAAACCC A UCUUCCCC 3869
1541 GACACACA GGCTAGCTACAACGA CCGGGGAA 3160 UUCCCCGG A UGUGUGUC 3870
1543 GAGACACA GGCTAGCTACAACGA ATCCGGGG 3161 CCCCGGAU G UGUGUCUC 3871
1545 GTGAGACA GGCTAGCTACAACGA ACATCCGG 3162 CCGGAUGU G UGUCUCAC 3872
1547 GGGTGAGA GGCTAGCTACAACGA ACACATCC 3163 GGAUGUGU G UCUCACCC 3873
1552 ATGAGGGG GGCTAGCTACAACGA GAGACACA 3164 UGUGUCUC A CCCCUCAU 3874
1559 TAAAAGGA GGCTAGCTACAACGA GAGGGGTG 3165 CACCCCUC A UCCUUUUA 3875
1567 GCAAAAAG GGCTAGCTACAACGA AAAAGGAT 3166 AUCCUUUU A CUUUUUGC 3876
1574 GGAAGGGG GGCTAGCTACAACGA AAAAAGTA 3167 UACUUUUU G CCCCUUCC 3877
1583 ACTCAAAG GGCTAGCTACAACGA GGAAGGGG 3168 CCCCUUCC A CUUUGAGU 3878
1590 ATTTGGTA GGCTAGCTACAACGA TCAAAGTG 3169 CACUUUGA G UACCAAAU 3879
1592 GGATTTGG GGCTAGCTACAACGA ACTCAAAG 3170 CUUUGAGU A CCAAAUCC 3880
1597 CTTGTGGA GGCTAGCTACAACGA TTGGTACT 3171 AGUACCAA A UCCACAAG 3881
1601 ATGGCTTG GGCTAGCTACAACGA GGATTTGG 3172 CCAAAUCC A CAAGCCAU 3882
Table b
1605 AAAAATGG GGCTAGCTACAACGA TTGTGGAT 3173 AUCCACAA G CCAUUUUU 3883
1608 TCAAAAAA GGCTAGCTACAACGA GGCTTGTG 3174 CACAAGCC A UUUUUUGA 3884
1622 CTCTTTCA GGCTAGCTACAACGA TCTCCTCA 3175 UGAGGAGA G UGAAAGAG 3885
1632 GCATGGTA GGCTAGCTACAACGA TCTCTTTC 3176 GAAAGAGA G UACCAUGC 3886
1634 CAGCATGG GGCTAGCTACAACGA ACTCTCTT 3177 AAGAGAGU A CCAUGCUG 3887
1637 CGCCAGCA GGCTAGCTACAACGA GGTACTCT 3178 AGAGUACC A UGCUGGCG 3888
1639 GCCGCCAG GGCTAGCTACAACGA ATGGTACT 3179 AGUACCAU G CUGGCGGC 3889
1643 CTGCGCCG GGCTAGCTACAACGA CAGCATGG 3180 CCAUGCUG G CGGCGCAG 3890
1646 CCTCTGCG GGCTAGCTACAACGA CGCCAGCA 3181 UGCUGGCG G CGCAGAGG 3891
1648 TCCCTCTG GGCTAGCTACAACGA GCCGCCAG 3182 CUGGCGGC G CAGAGGGA 3892
1661 GGTGTAGG GGCTAGCTACAACGA CCCTTCCC 3183 GGGAAGGG G CCUACACC 3893
1665 GACGGGTG GGCTAGCTACAACGA AGGCCCCT 3184 AGGGGCCU A CACCCGUC 3894
1667 AAGACGGG GGCTAGCTACAACGA GTAGGCCC 3185 GGGCCUAC A CCCGUCUU 3895
1671 CCCCAAGA GGCTAGCTACAACGA GGGTGTAG 3186 CUACACCC G UCUUGGGG 3896
1679 GGGGCGAG GGCTAGCTACAACGA CCCAAGAC 3187 GUCUUGGG G CUCGCCCC 3897
1683 GGGTGGGG GGCTAGCTACAACGA GAGCCCCA 3188 UGGGGCUC G CCCCACCC 3898
1688 GCCCTGGG GGCTAGCTACAACGA GGGGCGAG 3189 CUCGCCCC A CCCAGGGC 3899
1695 GGAGGGAG GGCTAGCTACAACGA CCTGGGTG 3190 CACCCAGG G CUCCCUCC 3900
1708 CTGGGATG GGCTAGCTACAACGA TCCAGGAG 3191 CUCCUGGA G CAUCCCAG 3901
1710 GCCTGGGA GGCTAGCTACAACGA GCTCCAGG 3192 CCUGGAGC A UCCCAGGC 3902
1717 GCCGCCCG GGCTAGCTACAACGA CTGGGATG 3193 CAUCCCAG G CGGGCGGC 3903
1721 GCGTGCCG GGCTAGCTACAACGA CCGCCTGG 3194 CCAGGCGG G CGGCACGC 3904
1724 CTGGCGTG GGCTAGCTACAACGA CGCCCGCC 3195 GGCGGGCG G CACGCCAG 3905
1726 GTCTGGCG GGCTAGCTACAACGA GCCGCCCG 3196 CGGGCGGC A CGCCAGAC 3906
1728 CTGTCTGG GGCTAGCTACAACGA GTGCCGCC 3197 GGCGGCAC G CCAGACAG 3907
1733 GGGGGCTG GGCTAGCTACAACGA CTGGCGTG 3198 CACGCCAG A CAGCCCCC 3908
1736 GGGGGGGG GGCTAGCTACAACGA TGTCTGGC 3199 GCCAGACA G CCCCCCCC 3909
1749 CCTGCAGA GGCTAGCTACAACGA TCAAGGGG 3200 CCCCUUGA A UCUGCAGG 3910
1753 GCTCCCTG GGCTAGCTACAACGA AGATTCAA 3201 UUGAAUCU G CAGGGAGC 3911
1760 GAGAGTTG GGCTAGCTACAACGA TCCCTGCA 3202 UGCAGGGA G CAACUCUC 3912
1763 GTGGAGAG GGCTAGCTACAACGA TGCTCCCT 3203 AGGGAGCA A CUCUCCAC 3913
Table 6
1770 ATATGGAG GGCTAGCTACAACGA GGAGAGTT 3204 AACUCUCC A CUCCAUAU 3914
1775. AATAAATA GGCTAGCTACAACGA GGAGTGGA 3205 UCCACUCC A UAUUUAUU 3915
1777 TAAATAAA GGCTAGCTACAACGA ATGGAGTG 3206 CACUCCAU A UUUAUUUA 3916
1781 TGTTTAAA GGCTAGCTACAACGA AAATATGG 3207 CCAUAUUU A UUUAAACA 3917
1787 AAAAATTG GGCTAGCTACAACGA TTAAATAA 3208 UUAUUUAA A CAAUUUUU 3918
1790 GGAAAAAA GGCTAGCTACAACGA TGTTTAAA 3209 UUUAAACA A UUUUUUCC 3919
1805 TATGGATG GGCTAGCTACAACGA CTTTGGGG 3210 CCCCAAAG G CAUCCAUA 3920
1807 ACTATGGA GGCTAGCTACAACGA GCCTTTGG 3211 CCAAAGGC A UCCAUAGU 3921
1811 GTGCACTA GGCTAGCTACAACGA GGATGCCT 3212 AGGCAUCC A UAGUGCAC 3922
1814 CTAGTGCA GGCTAGCTACAACGA TATGGATG 3213 CAUCCAUA G UGCACUAG 3923
1816 TGCTAGTG GGCTAGCTACAACGA ACTATGGA 3214 UCCAUAGU G CACUAGCA 3924
1818 AATGCTAG GGCTAGCTACAACGA GCACTATG 3215 CAUAGUGC A CUAGCAUU 3925
1822 AGAAAATG GGCTAGCTACAACGA TAGTGCAC 3216 GUGCACUA G CAUUUUCU 3926
1824 CAAGAAAA GGCTAGCTACAACGA GCTAGTGC 3217 GCACUAGC A UUUUCUUG 3927
1834 ATTATTGG GGCTAGCTACAACGA TCAAGAAA 3218 UUUCUUGA A CCAAUAAU 3928
1838 ATACATTA GGCTAGCTACAACGA TGGTTCAA 3219 UUGAACCA A UAAUGUAU 3929
1841 TTAATACA GGCTAGCTACAACGA TATTGGTT 3220 AACCAAUA A UGUAUUAA 3930
1843 TTTTAATA GGCTAGCTACAACGA ATTATTGG 3221 CCAAUAAU G UAUUAAAA 3931
1845 AATTTTAA GGCTAGCTACAACGA ACATTATT 3222 AAUAAUGU A UUAAAAUU 3932
1851 TCAAAAAA GGCTAGCTACAACGA TTTAATAC 3223 GUAUUAAA A UUUUUUGA 3933
1859 GGCTGACA GGCTAGCTACAACGA CAAAAAAT 3224 AUUUUUUG A UGUCAGCC 3934
1861 AAGGCTGA GGCTAGCTACAACGA ATCAAAAA 3225 UUUUUGAU G UCAGCCUU 3935
1865 ATGCAAGG GGCTAGCTACAACGA TGACATCA 3226 UGAUGUCA G CCUUGCAU 3936
1870 CCTTGATG GGCTAGCTACAACGA AAGGCTGA 3227 UCAGCCUU G CAUCAAGG 3937
1872 GCCCTTGA GGCTAGCTACAACGA GCAAGGCT 3228 AGCCUUGC A UCAAGGGC 3938
1879 TGATAAAG GGCTAGCTACAACGA CCTTGATG 3229 CAUCAAGG G CUUUAUCA 3939
1884 CTTTTTGA GGCTAGCTACAACGA AAAGCCCT 3230 AGGGCUUU A UCAAAAAG 3940
1892 TTATTGTA GGCTAGCTACAACGA TTTTTGAT 3231 AUCAAAAA G UACAAUAA 3941
1894 TATTATTG GGCTAGCTACAACGA ACTTTTTG 3232 CAAAAAGU A CAAUAAUA 3942
1897 ATTTATTA GGCTAGCTACAACGA TGTACTTT 3233 AAAGUACA A UAAUAAAU 3943
1900 AGGATTTA GGCTAGCTACAACGA TATTGTAC 3234 GUACAAUA A UAAAUCCU 3944
Table 6
1904 CCTGAGGA GGCTAGCTACAACGA TTATTATT 3235 AAUAAUAA A UCCUCAGG 3945
1912 CAGTACTA GGCTAGCTACAACGA CTGAGGAT 3236 AUCCUCAG G UAGUACUG 3946
1915 TCCCAGTA GGCTAGCTACAACGA TACCTGAG 3237 CUCAGGUA G UACUGGGA 3947
1917 ATTCCCAG GGCTAGCTACAACGA ACTACCTG 3238 CAGGUAGU A CUGGGAAU 3948
1924 GCCTTCCA GGCTAGCTACAACGA TCCCAGTA 3239 UACUGGGA A UGGAAGGC 3949
1931 TGGCAAAG GGCTAGCTACAACGA CTTCCATT 3240 AAUGGAAG G CUUUGCCA 3950
1936 GCCCATGG GGCTAGCTACAACGA AAAGCCTT 3241 AAGGCUUU G CCAUGGGC 3951
1939 CAGGCCCA GGCTAGCTACAACGA GGCAAAGC 3242 GCUUUGCC A UGGGCCUG 3952
1943 GCAGCAGG GGCTAGCTACAACGA CCATGGCA 3243 UGCCAUGG G CCUGCUGC 3953
1947 TGACGCAG GGCTAGCTACAACGA AGGCCCAT 3244 AUGGGCCU G CUGCGUCA 3954
1950 GTCTGACG GGCTAGCTACAACGA AGCAGGCC 3245 GGCCUGCU G CGUCAGAC 3955
1952 TGGTCTGA GGCTAGCTACAACGA GCAGCAGG 3246 CCUGCUGC G UCAGACCA 3956
1957 AGTACTGG GGCTAGCTACAACGA CTGACGCA 3247 UGCGUCAG A CCAGUACU 3957
1961 TCCCAGTA GGCTAGCTACAACGA TGGTCTGA 3248 UCAGACCA G UACUGGGA 3958
1963 CTTCCCAG GGCTAGCTACAACGA ACTGGTCT 3249 AGACCAGU A CUGGGAAG 3959
1976 TACAACCG GGCTAGCTACAACGA CCTCCTTC 3250 GAAGGAGG A CGGUUGUA 3960
1979 GCTTACAA GGCTAGCTACAACGA CGTCCTCC 3251 GGAGGACG G UUGUAAGC 3961
1982 ACTGCTTA GGCTAGCTACAACGA AACCGTCC 3252 GGACGGUU G UAAGCAGU 3962
1986 AACAACTG GGCTAGCTACAACGA TTACAACC 3253 GGUUGUAA G CAGUUGUU 3963
1989 AATAACAA GGCTAGCTACAACGA TGCTTACA 3254 UGUAAGCA G UUGUUAUU 3964
1992 CTAAATAA GGCTAGCTACAACGA AACTGCTT 3255 AAGCAGUU G UUAUUUAG 3965
1995 TCACTAAA GGCTAGCTACAACGA AACAACTG 3256 CAGUUGUU A UUUAGUGA 3966
2000 CAATATCA GGCTAGCTACAACGA TAAATAAC 3257 GUUAUUUA G UGAUAUUG 3967
2003 CCACAATA GGCTAGCTACAACGA CACTAAAT 3258 AUUUAGUG A UAUUGUGG 3968
2005 ACCCACAA GGCTAGCTACAACGA ATCACTAA 3259 UUAGUGAU A UUGUGGGU 3969
2008 GTTACCCA GGCTAGCTACAACGA AATATCAC 3260 GUGAUAUU G UGGGUAAC 3970
2012 TCACGTTA GGCTAGCTACAACGA CCACAATA 3261 UAUUGUGG G UAACGUGA 3971
2015 TTCTCACG GGCTAGCTACAACGA TACCCACA 3262 UGUGGGUA A CGUGAGAA 3972
2017 TCTTCTCA GGCTAGCTACAACGA GTTACCCA 3263 UGGGUAAC G UGAGAAGA 3973
2025 TTGTTCTA GGCTAGCTACAACGA CTTCTCAC 3264 GUGAGAAG A UAGAACAA 3974
2030 TAGCATTG GGCTAGCTACAACGA TCTATCTT 3265 AAGAUAGA A CAAUGCUA 3975
Table 6
2033 TTATAGCA GGCTAGCTACAACGA TGTTCTAT 3266 AUAGAACA A UGCUAUAA 3976
2035 TATTATAG GGCTAGCTACAACGA ATTGTTCT 3267 AGAACAAU G CUAUAAUA 3977
2038 ATATATTA GGCTAGCTACAACGA AGCATTGT 3268 ACAAUGCU A UAAUAUAU 3978
2041 ATTATATA GGCTAGCTACAACGA TATAGCAT 3269 AUGCUAUA A UAUAUAAU 3979
2043 TCATTATA GGCTAGCTACAACGA ATTATAGC 3270 GCUAUAAU A UAUAAUGA 3980
2045 GTTCATTA GGCTAGCTACAACGA ATATTATA 3271 UAUAAUAU A UAAUGAAC 3981
2048 CGTGTTCA GGCTAGCTACAACGA TATATATT 3272 AAUAUAUA A UGAACACG 3982
2052 CCCACGTG GGCTAGCTACAACGA TCATTATA 3273 UAUAAUGA A CACGUGGG 3983
2054 TACCCACG GGCTAGCTACAACGA GTTCATTA 3274 UAAUGAAC A CGUGGGUA 3984
2056 AATACCCA GGCTAGCTACAACGA GTGTTCAT 3275 AUGAACAC G UGGGUAUU 3985
2060 ATTAAATA GGCTAGCTACAACGA CCACGTGT 3276 ACACGUGG G UAUUUAAU 3986
2062 TTATTAAA GGCTAGCTACAACGA ACCCACGT 3277 ACGUGGGU A UUUAAUAA 3987
2067 GTTTCTTA GGCTAGCTACAACGA TAAATACC 3278 GGUAUUUA A UAAGAAAC 3988
2074 ACATCATG GGCTAGCTACAACGA TTCTTATT 3279 AAUAAGAA A CAUGAUGU 3989
2076 TCACATCA GGCTAGCTACAACGA GTTTCTTA 3280 UAAGAAAC A UGAUGUGA 3990
2079 ATCTCACA GGCTAGCTACAACGA CATGTTTC 3281 GAAACAUG A UGUGAGAU 3991
2081 TAATCTCA GGCTAGCTACAACGA ATCATGTT 3282 AACAUGAU G UGAGAUUA 3992
2086 CAAAGTAA GGCTAGCTACAACGA CTCACATC 3283 GAUGUGAG A UUACUUUG 3993
2089 GGACAAAG GGCTAGCTACAACGA AATCTCAC • 3284 GUGAGAUU A CUUUGUCC 3994
2094 AAGCGGGA GGCTAGCTACAACGA AAAGTAAT 3285 AUUACUUU G UCCCGCUU 3995
2099 AGAATAAG GGCTAGCTACAACGA GGGACAAA 3286 UUUGUCCC G CUUAUUCU 3996
2103 GAGCAGAA GGCTAGCTACAACGA AAGCGGGA 3287 UCCCGCUU A UUCUGCUC 3997
2108 ACAGGGAG GGCTAGCTACAACGA AGAATAAG 3288 CUUAUUCU G CUCCCUGU 3998
2115 GCAGATAA GGCTAGCTACAACGA AGGGAGCA 3289 UGCUCCCU G UUAUCUGC 3999
2118 CTAGCAGA GGCTAGCTACAACGA AACAGGGA 3290 UCCCUGUU A UCUGCUAG 4000
2122 AGATCTAG GGCTAGCTACAACGA AGATAACA 3291 UGUUAUCU G CUAGAUCU 4001
2127 GAACTAGA GGCTAGCTACAACGA CTAGCAGA 3292 UCUGCUAG A UCUAGUUC 4002
2132 ATTGAGAA GGCTAGCTACAACGA TAGATCTA 3293 UAGAUCUA G UUCUCAAU 4003
2139 AGCAGTGA GGCTAGCTACAACGA TGAGAACT 3294 AGUUCUCA A UCACUGCU 4004
2142 GGGAGCAG GGCTAGCTACAACGA GATTGAGA 3295 UCUCAAUC A CUGCUCCC 4005
2145 CGGGGGAG GGCTAGCTACAACGA AGTGATTG 3296 CAAUCACU G CUCCCCCG 4006
Table 6
2153 AATACACA GGCTAGCTACAACGA GGGGGAGC 3297 GCUCCCCC G UGUGUAUU 4007
2155 CTAATACA GGCTAGCTACAACGA ACGGGGGA 3298 UCCCCCGU G UGUAUUAG 4008
2157 TTCTAATA GGCTAGCTACAACGA ACACGGGG 3299 CCCCGUGU G UAUUAGAA 4009
2159 CATTCTAA GGCTAGCTACAACGA ACACACGG 3300. CCGUGUGU A UUAGAAUG 4010
2165 TACATGCA GGCTAGCTACAACGA TCTAATAC 3301 GUAUUAGA A UGCAUGUA 4011
2167 CTTACATG GGCTAGCTACAACGA ATTCTAAT 3302 AUUAGAAU G CAUGUAAG 4012
2169 ACCTTACA GGCTAGCTACAACGA GCATTCTA 3303 UAGAAUGC A UGUAAGGU 4013
2171 AGACCTTA GGCTAGCTACAACGA ATGCATTC 3304 GAAUGCAU G UAAGGUCU 4014
2176 CAAGAAGA GGCTAGCTACAACGA CTTACATG 3305 CAUGUAAG G UCUUCUUG 4015
2184 TCAGGACA GGCTAGCTACAACGA AAGAAGAC 3306 GUCUUCUU G UGUCCUGA 4016
2186 CATCAGGA GGCTAGCTACAACGA ACAAGAAG 3307 CUUCUUGU G UCCUGAUG 4017
2192 ATTTTTCA GGCTAGCTACAACGA CAGGACAC 3308 GUGUCCUG A UGAAAAAU 4018
2199 AGCACATA GGCTAGCTACAACGA TTTTCATC 3309 GAUGAAAA A UAUGUGCU 4019
2201 CAAGCACA GGCTAGCTACAACGA ATTTTTCA 3310 UGAAAAAU A UGUGCUUG 4020
2203 TTCAAGCA GGCTAGCTACAACGA ATATTTTT 3311 AAAAAUAU G UGCUUGAA 4021
2205 ATTTCAAG GGCTAGCTACAACGA ACATATTT 3312 AAAUAUGU G CUUGAAAU 4022
2212 GTTTCTCA GGCTAGCTACAACGA TTCAAGCA 3313 UGCUUGAA A UGAGAAAC 4023
2219 GATCAAAG GGCTAGCTACAACGA TTCTCATT 3314 AAUGAGAA A CUUUGAUC 4024
2225 AGCAGAGA GGCTAGCTACAACGA CAAAGTTT 3315 AAACUUUG A UCUCUGCU 4025
2231 TTAGTAAG GGCTAGCTACAACGA AGAGATCA 3316 UGAUCUCU G CUUACUAA 4026
2235 CACATTAG GGCTAGCTACAACGA AAGCAGAG 3317 CUCUGCUU A CUAAUGUG 4027
2239 GGGGCACA GGCTAGCTACAACGA TAGTAAGC 3318 GCUUACUA A UGUGCCCC 4028
2241 ATGGGGCA GGCTAGCTACAACGA ATTAGTAA 3319 UUACUAAU G UGCCCCAU 4029
2243 ACATGGGG GGCTAGCTACAACGA ACATTAGT 3320 ACUAAUGU G CCCCAUGU 4030
2248 CTTGGACA GGCTAGCTACAACGA GGGGCACA 3321 UGUGCCCC A UGUCCAAG 4031
2250 GACTTGGA GGCTAGCTACAACGA ATGGGGCA 3322 UGCCCCAU G UCCAAGUC 4032
2256 AGGTTGGA GGCTAGCTACAACGA TTGGACAT 3323 AUGUCCAA G UCCAACCU 4033
2261 CAGGCAGG GGCTAGCTACAACGA TGGACTTG 3324 CAAGUCCA A CCUGCCUG 4034
2265 TGCACAGG GGCTAGCTACAACGA AGGTTGGA 3325 UCCAACCU G CCUGUGCA 4035
2269 GTCATGCA GGCTAGCTACAACGA AGGCAGGT 3326 ACCUGCCU G UGCAUGAC 4036
2271 AGGTCATG GGCTAGCTACAACGA ACAGGCAG 3327 CUGCCUGU G CAUGACCU 4037
Table 6
2273 TCAGGTCA GGCTAGCTACAACGA GCACAGGC 3328 GCCUGUGC A UGACCUGA 4038
2276 TGATCAGG GGCTAGCTACAACGA CATGCACA 3329 UGUGCAUG A CCUGAUCA 4039
2281 TGTAATGA GGCTAGCTACAACGA CAGGTCAT 3330 AUGACCUG A UCAUUACA 4040
2284 CCATGTAA GGCTAGCTACAACGA GATCAGGT 3331 ACCUGAUC A UUACAUGG 4041
2287 CAGCCATG GGCTAGCTACAACGA AATGATCA 3332 UGAUCAUU A CAUGGCUG 4042
2289 CACAGCCA GGCTAGCTACAACGA GTAATGAT 3333 AUCAUUAC A UGGCUGUG 4043
2292 AACCACAG GGCTAGCTACAACGA CATGTAAT 3334 AUUACAUG G CUGUGGUU 4044
2295 AGGAACCA GGCTAGCTACAACGA AGCCATGT 3335 ACAUGGCU G UGGUUCCU 4045
2298 CTTAGGAA GGCTAGCTACAACGA CACAGCCA 3336 UGGCUGUG G UUCCUAAG 4046
2306 GCAACAGG GGCTAGCTACAACGA TTAGGAAC 3337 GUUCCUAA G CCUGUUGC 4047
2310 TTCAGCAA GGCTAGCTACAACGA AGGCTTAG 3338 CUAAGCCU G UUGCUGAA 4048
2313 GACTTCAG GGCTAGCTACAACGA AACAGGCT 3339 AGCCUGUU G CUGAAGUC 4049
2319 GACAATGA GGCTAGCTACAACGA TTCAGCAA 3340 UUGCUGAA G UCAUUGUC 4050
2322 AGCGACAA GGCTAGCTACAACGA GACTTCAG 3341 CUGAAGUC A UUGUCGCU 4051
2325 CTGAGCGA GGCTAGCTACAACGA AATGACTT 3342 AAGUCAUU G UCGCUCAG 4052
2328 TTGCTGAG GGCTAGCTACAACGA GACAATGA 3343 UCAUUGUC G CUCAGCAA 4053
2333 CCCTATTG GGCTAGCTACAACGA TGAGCGAC 3344 GUCGCUCA G CAAUAGGG 4054
2336 GCACCCTA GGCTAGCTACAACGA TGCTGAGC 3345 GCUCAGCA A UAGGGUGC 4055
2341 AAACTGCA GGCTAGCTACAACGA CCTATTGC 3346 GCAAUAGG G UGCAGUUU 4056
2343 GAAAACTG GGCTAGCTACAACGA ACCCTATT 3347 AAUAGGGU G CAGUUUUC 4057
2346 CTGGAAAA GGCTAGCTACAACGA TGCACCCT 3348 AGGGUGCA G UUUUCCAG 4058
2357 AATGCCTA GGCTAGCTACAACGA TCCTGGAA 3349 UUCCAGGA A UAGGCAUU 4059
2361 GGCAAATG GGCTAGCTACAACGA CTATTCCT 3350 AGGAAUAG G CAUUUGCC 4060
2363 TAGGCAAA GGCTAGCTACAACGA GCCTATTC 3351 GAAUAGGC A UUUGCCUA 4061
2367 GAATTAGG GGCTAGCTACAACGA AAATGCCT 3352 AGGCAUUU G CCUAAUUC 4062
2372 GCCAGGAA GGCTAGCTACAACGA TAGGCAAA 3353 UUUGCCUA A UUCCUGGC 4063
2379 GTGTCATG GGCTAGCTACAACGA CAGGAATT 3354 AAUUCCUG G CAUGACAC 4064
2381 GAGTGTCA GGCTAGCTACAACGA GCCAGGAA 3355 UUCCUGGC A UGACACUC 4065
2384 CTAGAGTG GGCTAGCTACAACGA CATGCCAG 3356 CUGGCAUG A CACUCUAG 4066
2386 CACTAGAG GGCTAGCTACAACGA GTCATGCC 3357 GGCAUGAC A CUCUAGUG 4067
2392 GGAAGTCA GGCTAGCTACAACGA TAGAGTGT 3358 ACACUCUA G UGACUUCC 4068
Table 6
Table 6
2541 GAGGCTGA GGCTAGCTACAACGA CCCAAAGG 3390 CCUUUGGG A UCAGCCUC 4100
2545 GGCGGAGG GGCTAGCTACAACGA TGATCCCA 3391 UGGGAUCA G CCUCCGCC 4101
2551 TGGAATGG GGCTAGCTACAACGA GGAGGCTG 3392 CAGCCUCC G CCAUUCCA 4102
2554 ACTTGGAA GGCTAGCTACAACGA GGCGGAGG 3393 CCUCCGCC A UUCCAAGU 4103
2561 AGTGTCGA GGCTAGCTACAACGA TTGGAATG 3394 CAUUCCAA G UCGACACU 4104
2565 GAAGAGTG GGCTAGCTACAACGA CGACTTGG 3395 CCAAGUCG A CACUCUUC 4105
2567 AAGAAGAG GGCTAGCTACAACGA GTCGACTT 3396 AAGUCGAC A CUCUUCUU 4106
2578 ACGGTCTG GGCTAGCTACAACGA TCAAGAAG 3397 CUUCUUGA G CAGACCGU 4107
2582 AATCACGG GGCTAGCTACAACGA CTGCTCAA 3398 UUGAGCAG A CCGUGAUU 4108
2585 CCAAATCA GGCTAGCTACAACGA GGTCTGCT 3399 AGCAGACC G UGAUUUGG 4109
2588 CTTCCAAA GGCTAGCTACAACGA CACGGTCT 3400 AGACCGUG A UUUGGAAG 4110
2601 AGCAGGTG GGCTAGCTACAACGA CTCTCTTC 3401 GAAGAGAG G CACCUGCU 4111
2603 CCAGCAGG GGCTAGCTACAACGA GCCTCTCT 3402 AGAGAGGC A CCUGCUGG 4112
2607 GTTTCCAG GGCTAGCTACAACGA AGGTGCCT 3403 AGGCACCU G CUGGAAAC 4113
2614 AAGTGTGG GGCTAGCTACAACGA TTCCAGCA 3404 UGCUGGAA A CCACACUU 4114
2617 AAGAAGTG GGCTAGCTACAACGA GGTTTCCA 3405 UGGAAACC A CACUUCUU 4115
2619 TCAAGAAG GGCTAGCTACAACGA GTGGTTTC 3406 GAAACCAC A CUUCUUGA 4116
2629 CCAGGCTG GGCTAGCTACAACGA TTCAAGAA 3407 UUCUUGAA A CAGCCUGG 4117
2632 CACCCAGG GGCTAGCTACAACGA TGTTTCAA 3408 UUGAAACA G CCUGGGUG 4118
2638 GACCGTCA GGCTAGCTACAACGA CCAGGCTG 3409 CAGCCUGG G UGACGGUC 4119
2641 AAGGACCG GGCTAGCTACAACGA CACCCAGG 3410 CCUGGGUG A CGGUCCUU 4120
2644 CTAAAGGA GGCTAGCTACAACGA CGTCACCC 3411 GGGUGACG G UCCUUUAG 4121
2653 GCAGGCTG GGCTAGCTACAACGA CTAAAGGA 3412 UCCUUUAG G CAGCCUGC 4122
2656 GCGGCAGG GGCTAGCTACAACGA TGCCTAAA 3413 UUUAGGCA G CCUGCCGC 4123
2660 GACGGCGG GGCTAGCTACAACGA AGGCTGCC 3414 GGCAGCCU G CCGCCGUC 4124
2663 AGAGACGG GGCTAGCTACAACGA GGCAGGCT 3415 AGCCUGCC G CCGUCUCU 4125
2666 GACAGAGA GGCTAGCTACAACGA GGCGGCAG 3416 CUGCCGCC G UCUCUGUC 4126
2672 AACCGGGA GGCTAGCTACAACGA AGAGACGG 3417 CCGUCUCU G UCCCGGUU 4127
2678 AAGGTGAA GGCTAGCTACAACGA CGGGACAG 3418 CUGUCCCG G UUCACCUU 4128
2682 CGGCAAGG GGCTAGCTACAACGA GAACCGGG 3419 CCCGGUUC A CCUUGCCG 4129
2687 TCTCTCGG GGCTAGCTACAACGA AAGGTGAA 3420 UUCACCUU G CCGAGAGA 4130
Table 6
2697 CAGACGCG GGCTAGCTACAACGA CTCTCTCG 3421 CGAGAGAG G CGCGUCUG 4131
2699 GGCAGACG GGCTAGCTACAACGA GCCTCTCT 3422 AGAGAGGC G CGUCUGCC 4132
2701 GGGGCAGA GGCTAGCTACAACGA GCGCCTCT 3423 AGAGGCGC G UCUGCCCC 4133
2705 GGGTGGGG GGCTAGCTACAACGA AGACGCGC 3424 GCGCGUCU G CCCCACCC 4134
2710 TTTGAGGG GGCTAGCTACAACGA GGGGCAGA 3425 UCUGCCCC A CCCUCAAA 4135
2718 CCACAGGG GGCTAGCTACAACGA TTGAGGGT 3426 ACCCUCAA A CCCUGUGG 4136
2723 AGGCCCCA GGCTAGCTACAACGA AGGGTTTG 3427 CAAACCCU G UGGGGCCU 4137
2728 CCATCAGG GGCTAGCTACAACGA CCCACAGG 3428 CCUGUGGG G CCUGAUGG 4138
2733 GAGCACCA GGCTAGCTACAACGA CAGGCCCC 3429 GGGGCCUG A UGGUGCUC 4139
2736 CGTGAGCA GGCTAGCTACAACGA CATCAGGC 3430 GCCUGAUG G UGCUCACG 4140
2738 GTCGTGAG GGCTAGCTACAACGA ACCATCAG 3431 CUGAUGGU G CUCACGAC 4141
2742 AAGAGTCG GGCTAGCTACAACGA GAGCACCA 3432 UGGUGCUC A CGACUCUU 4142
2745 AGGAAGAG GGCTAGCTACAACGA CGTGAGCA 3433 UGCUCACG A CUCUUCCU 4143
2754 TCCCTTTG GGCTAGCTACAACGA AGGAAGAG 3434 ■ CUCUUCCU G CAAAGGGA 4144
2763 GTCTTCAG GGCTAGCTACAACGA TCCCTTTG 3435 CAAAGGGA A CUGAAGAC 4145
2770 TGTGGAGG GGCTAGCTACAACGA CTTCAGTT 3436 AACUGAAG A CCUCCACA 4146
2776 ACTTAATG GGCTAGCTACAACGA GGAGGTCT 3437 AGACCUCC A CAUUAAGU 4147
2778 CCACTTAA GGCTAGCTACAACGA GTGGAGGT 3438 ACCUCCAC A UUAAGUGG 4148
2783 AAAAGCCA GGCTAGCTACAACGA TTAATGTG 3439 CACAUUAA G UGGCUUUU 4149
2786 TTAAAAAG GGCTAGCTACAACGA CACTTAAT 3440 AUUAAGUG G CUUUUUAA 4150
2794 TTTTCATG GGCTAGCTACAACGA TAAAAAGC 3441 GCUUUUUA A CAUGAAAA 4151
2796 GTTTTTCA GGCTAGCTACAACGA GTTAAAAA 3442 UUUUUAAC A UGAAAAAC 4152
2803 CTGCCGTG GGCTAGCTACAACGA TTTTCATG 3443 CAUGAAAA A CACGGCAG 4153
2805 AGCTGCCG GGCTAGCTACAACGA GTTTTTCA 3444 UGAAAAAC A CGGCAGCU 4154
2808 TACAGCTG GGCTAGCTACAACGA CGTGTTTT 3445 AAAACACG G CAGCUGUA 4155
2811 AGCTACAG GGCTAGCTACAACGA TGCCGTGT 3446 ACACGGCA G CUGUAGCU 4156
2814 GGGAGCTA GGCTAGCTACAACGA AGCTGCCG 3447 CGGCAGCU G UAGCUCCC 4157
2817 CTCGGGAG GGCTAGCTACAACGA TACAGCTG 3448 CAGCUGUA G CUCCCGAG 4158
2825 GAGAGTAG GGCTAGCTACAACGA TCGGGAGC 3449 GCUCCCGA G CUACUCUC 4159
2828 CAAGAGAG GGCTAGCTACAACGA AGCTCGGG 3450 CCCGAGCU A CUCUCUUG 4160
2836 AATGCTGG GGCTAGCTACAACGA AAGAGAGT 3451 ACUCUCUU G CCAGCAUU 4161
Table 6
2840 TGAAAATG GGCTAGCTACAACGA TGGCAAGA 3452 UCUUGCCA G CAUUUUCA 4162
2842 TGTGAAAA GGCTAGCTACAACGA GCTGGCAA 3453 UUGCCAGC A UUUUCACA 4163
2848 GCAAAATG GGCTAGCTACAACGA GAAAATGC 3454 GCAUUUUC A CAUUUUGC 4164
2850 AGGCAAAA GGCTAGCTACAACGA GTGAAAAT 3455 AUUUUCAC A UUUUGCCU 4165
2855 GAGAAAGG GGCTAGCTACAACGA AAAATGTG 3456 CACAUUUU G CCUUUCUC 4166
2864 TTCTACCA GGCTAGCTACAACGA GAGAAAGG 3457 CCUUUCUC G UGGUAGAA 4167
2867 GGCTTCTA GGCTAGCTACAACGA CACGAGAA 3458 UUCUCGUG G UAGAAGCC 4168
2873 TGTACTGG GGCTAGCTACAACGA TTCTACCA 3459 UGGUAGAA G CCAGUACA 4169
2877 TCTCTGTA GGCTAGCTACAACGA TGGCTTCT 3460 AGAAGCCA G UACAGAGA 4170
2879 TTTCTCTG GGCTAGCTACAACGA ACTGGCTT 3461 AAGCCAGU A CAGAGAAA 4171
2887 CCACAGAA GGCTAGCTACAACGA TTCTCTGT 3462 ACAGAGAA A UUCUGUGG 4172
2892 TCCCACCA GGCTAGCTACAACGA AGAATTTC 3463 GAAAUUCU G UGGUGGGA 4173
2895 TGTTCCCA GGCTAGCTACAACGA CACAGAAT 3464 AUUCUGUG G UGGGAACA 4174
2901 CTCGAATG GGCTAGCTACAACGA TCCCACCA 3465 UGGUGGGA A CAUUCGAG 4175
2903 ACCTCGAA GGCTAGCTACAACGA GTTCCCAC 3466 GUGGGAAC A UUCGAGGU 4176
2910 GGGTGACA GGCTAGCTACAACGA CTCGAATG 3467 CAUUCGAG G UGUCACCC 4177
2912 CAGGGTGA GGCTAGCTACAACGA ACCTCGAA 3468 UUCGAGGU G UCACCCUG 4178
2915 CTGCAGGG GGCTAGCTACAACGA GACACCTC 3469 GAGGUGUC A CCCUGCAG 4179
2920 TAGCTCTG GGCTAGCTACAACGA AGGGTGAC 3470 GUCACCCU G CAGAGCUA 4180
2925 CACCATAG GGCTAGCTACAACGA TCTGCAGG 3471 CCUGCAGA G CUAUGGUG 4181
2928 CCTCACCA GGCTAGCTACAACGA AGCTCTGC 3472 GCAGAGCU A UGGUGAGG 4182
2931 ACACCTCA GGCTAGCTACAACGA CATAGCTC 3473 GAGCUAUG G UGAGGUGU 4183
2936 TATCCACA GGCTAGCTACAACGA CTCACCAT 3474 AUGGUGAG G UGUGGAUA 4184
2938 CTTATCCA GGCTAGCTACAACGA ACCTCACC 3475 GGUGAGGU G UGGAUAAG 4185
2942 AAGCCTTA GGCTAGCTACAACGA CCACACCT 3476 AGGUGUGG A UAAGGCUU 4186
2947 CACCTAAG GGCTAGCTACAACGA CTTATCCA 3477 UGGAUAAG G CUUAGGUG 4187
2953 GCCTGGCA GGCTAGCTACAACGA CTAAGCCT 3478 AGGCUUAG G UGCCAGGC 4188
2955 CAGCCTGG GGCTAGCTACAACGA ACCTAAGC 3479 GCUUAGGU G CCAGGCUG 4189
2960 GCTTACAG GGCTAGCTACAACGA CTGGCACC 3480 GGUGCCAG G CUGUAAGC 4190
2963 AATGCTTA GGCTAGCTACAACGA AGCCTGGC 3481 GCCAGGCU G UAAGCAUU 4191
2967 TCAGAATG GGCTAGCTACAACGA TTACAGCC 3482 GGCUGUAA G CAUUCUGA 4192
Table 6
2969 GCTCAGAA GGCTAGCTACAACGA GCTTACAG 3483 CUGUAAGC A UUCUGAGC 4193
2976 CAAGCCAG GGCTAGCTACAACGA TCAGAATG 3484 CAUUCUGA G CUGGCUUG 4194
2980 ACAACAAG GGCTAGCTACAACGA CAGCTCAG 3485 CUGAGCUG G CUUGUUGU 4195
2984 AAAAACAA GGCTAGCTACAACGA AAGCCAGC 3486 GCUGGCUU G UUGUUUUU 4196
2987 CTTAAAAA GGCTAGCTACAACGA AACAAGCC 3487 GGCUUGUU G UUUUUAAG 4197
2995 ATACAGGA GGCTAGCTACAACGA TTAAAAAC 3488 GUUUUUAA G UCCUGUAU 4198
3000 TACATATA GGCTAGCTACAACGA AGGACTTA 3489 UAAGUCCU G UAUAUGUA 4199
3002 CATACATA GGCTAGCTACAACGA ACAGGACT 3490 AGUCCUGU A UAUGUAUG 4200
3004 TACATACA GGCTAGCTACAACGA ATACAGGA 3491 UCCUGUAU A UGUAUGUA 4201
3006 ACTACATA GGCTAGCTACAACGA ATATACAG 3492 CUGUAUAU G UAUGUAGU 4202
3008 CTACTACA GGCTAGCTACAACGA ACATATAC 3493 GUAUAUGU A UGUAGUAG 4203
3010 AACTACTA GGCTAGCTACAACGA ATACATAT 3494 AUAUGUAU G UAGUAGUU 4204
3013 CCAAACTA GGCTAGCTACAACGA TACATACA 3495 UGUAUGUA G UAGUUUGG 4205
3016 CACCCAAA GGCTAGCTACAACGA TACTACAT 3496 AUGUAGUA G UUUGGGUG 4206
3022 TATACACA GGCTAGCTACAACGA CCAAACTA 3497 UAGUUUGG G UGUGUAUA 4207
3024 TATATACA GGCTAGCTACAACGA ACCCAAAC 3498 GUUUGGGU G UGUAUAUA 4208
3026 TATATATA GGCTAGCTACAACGA ACACCCAA 3499 UUGGGUGU G UAUAUAUA 4209
3028 ACTATATA GGCTAGCTACAACGA ACACACCC 3500 GGGUGUGU A UAUAUAGU 4210
3030 CTACTATA GGCTAGCTACAACGA ATACACAC 3501 GUGUGUAU A UAUAGUAG 4211
3032 TGCTACTA GGCTAGCTACAACGA ATATACAC 3502 GUGUAUAU A UAGUAGCA 4212
3035 AAATGCTA GGCTAGCTACAACGA TATATATA 3503 UAUAUAUA G UAGCAUUU 4213
3038 TTGAAATG GGCTAGCTACAACGA TACTATAT 3504 AUAUAGUA G CAUUUCAA 4214
3040 TTTTGAAA GGCTAGCTACAACGA GCTACTAT 3505 AUAGUAGC A UUUCAAAA 4215
3048 TACGTCCA GGCTAGCTACAACGA TTTGAAAT 3506 AUUUCAAA A UGGACGUA 4216
3052 CCAGTACG GGCTAGCTACAACGA CCATTTTG 3507 CAAAAUGG A CGUACUGG 4217
3054 AACCAGTA GGCTAGCTACAACGA GTCCATTT 3508 AAAUGGAC G UACUGGUU 4218
3056 TAAACCAG GGCTAGCTACAACGA ACGTCCAT 3509 AUGGACGU A CUGGUUUA 4219
3060 AGGTTAAA GGCTAGCTACAACGA CAGTACGT 3510 ACGUACUG G UUUAACCU 4220
3065 ATAGGAGG GGCTAGCTACAACGA TAAACCAG 3511 CUGGUUUA A CCUCCUAU 4221
3072 TCCAAGGA GGCTAGCTACAACGA AGGAGGTT 3512 AACCUCCU A UCCUUGGA 4222
3083 GCCAGCTG GGCTAGCTACAACGA TCTCCAAG 3513 CUUGGAGA G CAGCUGGC 4223
Table 6
3086 AGAGCCAG GGCTAGCTACAACGA TGCTCTCC 3514 GGAGAGCA G CUGGCUCU 4224
3090 GTGGAGAG GGCTAGCTACAACGA CAGCTGCT 3515 AGCAGCUG G CUCUCCAC 4225
3097 TAACAAGG GGCTAGCTACAACGA GGAGAGCC 3516 GGCUCUCC A CCUUGUUA 4226
3102 ATGTGTAA GGCTAGCTACAACGA AAGGTGGA 3517 UCCACCUU G UUACACAU 4227
3105 ATAATGTG GGCTAGCTACAACGA AACAAGGT 3518 ACCUUGUU A CACAUUAU 4228
3107 ACATAATG GGCTAGCTACAACGA GTAACAAG 3519 CUUGUUAC A CAUUAUGU 4229
3109 TAACATAA GGCTAGCTACAACGA GTGTAACA 3520 UGUUACAC A UUAUGUUA 4230
3112 CTCTAACA GGCTAGCTACAACGA AATGTGTA 3521 UACACAUU A UGUUAGAG 4231
3114 CTCTCTAA GGCTAGCTACAACGA ATAATGTG 3522 CACAUUAU G UUAGAGAG 4232
3123 GCTCGCTA GGCTAGCTACAACGA CTCTCTAA 3523 UUAGAGAG G UAGCGAGC 4233
3126 GCAGCTCG GGCTAGCTACAACGA TACCTCTC 3524 GAGAGGUA G CGAGCUGC 4234
3130 CAGAGCAG GGCTAGCTACAACGA TCGCTACC 3525 GGUAGCGA G CUGCUCUG 4235
3133 TAGCAGAG GGCTAGCTACAACGA AGCTCGCT 3526 AGCGAGCU G CUCUGCUA 4236
3138 GGACATAG GGCTAGCTACAACGA AGAGCAGC 3527 GCUGCUCU G CUAUGUCC 4237
3141 TAAGGACA GGCTAGCTACAACGA AGCAGAGC 3528 GCUCUGCU A UGUCCUUA 4238
3143 CTTAAGGA GGCTAGCTACAACGA ATAGCAGA 3529 UCUGCUAU G UCCUUAAG 4239
3151 AATATTGG GGCTAGCTACAACGA TTAAGGAC 3530 GUCCUUAA G CCAAUAUU 4240
3155 AGTAAATA GGCTAGCTACAACGA TGGCTTAA 3531 UUAAGCCA A UAUUUACU 4241
3157 TGAGTAAA GGCTAGCTACAACGA ATTGGCTT 3532 AAGCCAAU A UUUACUCA 4242
3161 CTGATGAG GGCTAGCTACAACGA AAATATTG 3533 CAAUAUUU A CUCAUCAG 4243
3165 TGACCTGA GGCTAGCTACAACGA GAGTAAAT 3534 AUUUACUC A UCAGGUCA 4244
3170 AATAATGA GGCTAGCTACAACGA CTGATGAG 3535 CUCAUCAG G UCAUUAUU 4245
3173 AAAAATAA GGCTAGCTACAACGA GACCTGAT 3536 AUCAGGUC A UUAUUUUU 4246
3176 GTAAAAAA GGCTAGCTACAACGA AATGACCT 3537 AGGUCAUU A UUUUUUAC 4247
3183 GGCCATTG GGCTAGCTACAACGA AAAAAATA 3538 UAUUUUUU A CAAUGGCC 4248
3186 CATGGCCA GGCTAGCTACAACGA TGTAAAAA 3539 UUUUUACA A UGGCCAUG 4249
3189 TTCCATGG GGCTAGCTACAACGA CATTGTAA 3540 UUACAAUG G CCAUGGAA 4250
3192 TTATTCCA GGCTAGCTACAACGA GGCCATTG 3541 CAAUGGCC A UGGAAUAA 4251
3197 ATGGTTTA GGCTAGCTACAACGA TCCATGGC 3542 GCCAUGGA A UAAACCAU 4252
3201 AAAAATGG GGCTAGCTACAACGA TTATTCCA 3543 UGGAAUAA A CCAUUUUU 4253
3204 TGTAAAAA GGCTAGCTACAACGA GGTTTATT 3544 AAUAAACC A UUUUUACA 4254
Table 7
Table 7: Human PTP-IB Hairpin Ribozyme and Target Sequence
Nt. Ribozyme sequence Seq. ID Substrate Sequence Seq. ID Position Nos. Nos.
10 GCUCUA AGAA GCGU ACCAGAGAAACA X GUACAUUACCUGGUA 4255 ACGC GGCC TAGAGC 4331
23 UGCGCC AGAA GCCG ACCAGAGAAACA X GUACAUUACCUGGUA 4256 CGGC AGAC GGCGCA 4332
56 CAGGGC AGAA GCUG ACCAGAGAAACA X GUACAUUACCUGGUA 4257 CAGC AGCC GCCCTG 4333
59 GGCCAG AGAA GCUG ACCAGAGAAACA X GUACAUUACCUGGUA 4258 CAGC CGCC CTGGCC 4334
98 UUGUCG AGAA GCUC ACCAGAGAAACA X GUACAUUACCUGGUA 4259 GAGC AGAT CGACAA 4335
220 CAAAGG AGAA GACG ACCAGAGAAACA X GUACAUUACCUGGUA 4260 CGTC AGTC CCTTTG 4336
239 AGUUUA AGAA GACU ACCAGAGAAACA X GUACAUUACCUGGUA 4261 AGTC GGAT TAAACT 4337
612 UCCAAA AGAA GGCC ACCAGAGAAACA X GUACAUUACCUGGUA 4262 GGCC TGAC TTTGGA 4338
636 GAAUGA AGAA GGUG ACCAGAGAAACA X GUACAUUACCUGGUA 4263 CACC AGCC TCATTC 4339
685 GCUCCG AGAA GAGU ACCAGAGAAACA X GUACAUUACCUGGUA 4264 ACTC AGCC CGGAGC 4340
702 CACCAC AGAA GGCC ACCAGAGAAACA X GUACAUUACCUGGUA 4265 GGCC CGTT GTGGTG 4341
748 CAGCCA AGAA GAAG ACCAGAGAAACA X GUACAUUACCUGGUA 4266 CTTC TGTC TGGCTG 4342
763 GCAAGA AGAA GGUA ACCAGAGAAACA X GUACAUUACCUGGUA 4267 TACC TGCC TCTTGC 4343
773 UUGUCC AGAA GCAA ACCAGAGAAACA X GUACAUUACCUGGUA 4268 TTGC TGAT GGACAA 4344
801 GAUAUC AGAA GAAG ACCAGAGAAACA X GUACAUUACCUGGUA 4269 CTTC CGTT GATATC 4345
842 AGCCCC AGAA GAAA ACCAGAGAAACA X GUACAUUACCUGGUA 4270 TTTC GGAT GGGGCT 4346
851 GUCUGG AGAA GCCC ACCAGAGAAACA X GUACAUUACCUGGUA 4271 GGGC TGAT CCAGAC 4347
861 CUGGUC AGAA GUCU ACCAGAGAAACA X GUACAUUACCUGGUA 4272 AGAC AGCC GACCAG 4348
864 CAGCUG AGAA GCUG ACCAGAGAAACA X GUACAUUACCUGGUA 4273 CAGC CGAC CAGCTG 4349
869 AAGCGC AGAA GGUC ACCAGAGAAACA X GUACAUUACCUGGUA 4274 GACC AGCT GCGCTT 4350
1102 UGAUGG AGAA GUCU ACCAGAGAAACA X GUACAUUACCUGGUA 4275 AGAC TGCC CCATCA 4351
1224 UGGGGA AGAA GCCU ACCAGAGAAACA X GUACAUUACCUGGUA 4276 AGGC TGCC TCCCCA 4352
1253 UUCUCG AGAA GUGA ACCAGAGAAACA X GUACAUUACCUGGUA 4277 TCAC TGCC CGAGAA 4353
1323 CGUGAG AGAA GUAG ACCAGAGAAACA X GUACAUUACCUGGUA 4278 CTAC GGTC CTCACG 4354
1332 AGCGCC AGAA GUGA ACCAGAGAAACA X GUACAUUACCUGGUA 4279 TCAC GGCC GGCGCT 4355
1361 CUGUUG AGAA GGAA ACCAGAGAAACA X GUACAUUACCUGGUA 4280 TTCC TGTT CAACAG 4356
1383 AGGAGG AGAA GGCU ACCAGAGAAACA X GUACAUUACCUGGUA 4281 AGCC TGAC CCTCCT 4357
1412 GAGGCG AGAA GUGG ACCAGAGAAACA X GUACAUUACCUGGUA 4282 CCAC TGTC CGCCTC 4358
Table 7
1416 GGCAGA AGAA GACA ACCAGAGAAACA X GUACAUUACCUGGUA 4283 TGTC CGCC TCTGCC 4359
1422 UCUGCG AGAA GAGG ACCAGAGAAACA X GUACAUUACCUGGUA 4284 CCTC TGCC CGCAGA 4360
1441 CUGCUA AGAA GGCG ACCAGAGAAACA X GUACAUUACCUGGUA 4285 CGCC CGAC TAGCAG 4361
1460 CUUACC AGAA GCGG ACCAGAGAAACA X GUACAUUACCUGGUA 4286 CCGC GGTA GGTAAG 4362
1473 CGGUCC AGAA GCCC ACCAGAGAAACA X GUACAUUACCUGGUA 4287 GGGC CGCC GGACCG 4363
1477 UACGCG AGAA GGCG ACCAGAGAAACA X GUACAUUACCUGGUA 4288 CGCC GGAC CGCGTA 4364
1501 ACGUCC AGAA GGGG ACCAGAGAAACA X GUACAUUACCUGGUA 4289 CCCC GGAC GGACGT 4365
1505 ACCAAC AGAA GUCC ACCAGAGAAACA X GUACAUUACCUGGUA 4290 GGAC GGAC GTTGGT 4366
1539 ACACAC AGAA GGGG ACCAGAGAAACA X GUACAUUACCUGGUA 4291 CCCC GGAT GTGTGT 4367
1670 CCCCAA AGAA GGUG ACCAGAGAAACA X GUACAUUACCUGGUA 4292 CACC CGTC TTGGGG 4368
1735 GGGGGG AGAA GUCU ACCAGAGAAACA X GUACAUUACCUGGUA 4293 AGAC AGCC CCCCCC 4369
1864 AUGCAA AGAA GACA ACCAGAGAAACA X GUACAUUACCUGGUA 4294 TGTC AGCC TTGCAT 4370
1946 UGACGC AGAA GGCC ACCAGAGAAACA X GUACAUUACCUGGUA 4295 GGCC TGCT GCGTCA 4371
1955 GUACUG AGAA GACG ACCAGAGAAACA X GUACAUUACCUGGUA 4296 CGTC AGAC CAGTAC 4372
1960 UCCCAG AGAA GGUC ACCAGAGAAACA X GUACAUUACCUGGUA 4297 GACC AGTA CTGGGA 4373
1978 GCUUAC AGAA GUCC ACCAGAGAAACA X GUACAUUACCUGGUA 4298 GGAC GGTT GTAAGC 4374
1988 AAUAAC AGAA GCUU ACCAGAGAAACA X GUACAUUACCUGGUA 4299 AAGC AGTT GTTATT 4375
2098 AGAAUA AGAA GGAC ACCAGAGAAACA X GUACAUUACCUGGUA 4300 GTCC CGCT TATTCT 4376
2107 ACAGGG AGAA GAAU ACCAGAGAAACA X GUACAUUACCUGGUA 4301 ATTC TGCT CCCTGT 4377
2144 CGGGGG AGAA GUGA ACCAGAGAAACA X GUACAUUACCUGGUA 4302 TCAC TGCT CCCCCG 4378
2190 UUUUUC AGAA GGAC ACCAGAGAAACA X GUACAUUACCUGGUA 4303 GTCC TGAT GAAAAA 4379
2230 UUAGUA AGAA GAGA ACCAGAGAAACA X GUACAUUACCUGGUA 4304 TCTC TGCT TACTAA 4380
2264 UGCACA AGAA GGUU ACCAGAGAAACA X GUACAUUACCUGGUA 4305 AACC TGCC TGTGCA 4381
2279 GUAAUG AGAA GGUC ACCAGAGAAACA X GUACAUUACCUGGUA 4306 GACC TGAT CATTAC 4382
2309 UUCAGC AGAA GGCU ACCAGAGAAACA X GUACAUUACCUGGUA 4307 AGCC TGTT GCTGAA 4383
2345 CUGGAA AGAA GCAC ACCAGAGAAACA X GUACAUUACCUGGUA 4308 GTGC AGTT TTCCAG 4384
2412 AGGACA AGAA GGGC ACCAGAGAAACA X GUACAUUACCUGGUA 4309 GCCC AGCC TGTCCT 4385
2416 UACCAG AGAA GGCU ACCAGAGAAACA X GUACAUUACCUGGUA 4310 AGCC TGTC CTGGTA - 4386
2524 GUGGGG AGAA GGGG ACCAGAGAAACA X GUACAUUACCUGGUA 4311 CCCC CGCC CCCCAC 4387
2544 GGCGGA AGAA GAUC ACCAGAGAAACA X GUACAUUACCUGGUA 4312 GATC AGCC TCCGCC 4388
2580 AUCACG AGAA GCUC ACCAGAGAAACA X GUACAUUACCUGGUA 4313 GAGC AGAC CGTGAT 4389
Table 7
2606 GUUUCC AGAA GGUG ACCAGAGAAACA X GUACAUUACCUGGUA 4314 CACC TGCT GGAAAC 4390
2631 CACCCA AGAA GUUU ACCAGAGAAACA X GUACAUUACCUGGUA 4315 AAAC AGCC TGGGTG 4391
2643 CUAAAG AGAA GUCA ACCAGAGAAACA X GUACAUUACCUGGUA 4316 TGAC GGTC CTTTAG 4392
2655 GCGGCA AGAA GCCU ACCAGAGAAACA X GUACAUUACCUGGUA 4317 AGGC AGCC TGCCGC 4393
2659 GACGGC AGAA GGCU ACCAGAGAAACA X GUACAUUACCUGGUA 4318 AGCC TGCC GCCGTC 4394
2662 AGAGAC AGAA GCAG ACCAGAGAAACA X GUACAUUACCUGGUA 4319 CTGC CGCC GTCTCT 4395
2665 GACAGA AGAA GCGG ACCAGAGAAACA X GUACAUUACCUGGUA 4320 CCGC CGTC TCTGTC 4396
2671 AACCGG AGAA GAGA ACCAGAGAAACA X GUACAUUACCUGGUA 4321 TCTC TGTC CCGGTT 4397
2677 AAGGUG AGAA GGGA ACCAGAGAAACA X GUACAUUACCUGGUA 4322 TCCC GGTT CACCTT 4398
2704 GGGUGG AGAA GACG ACCAGAGAAACA X GUACAUUACCUGGUA 4323 CGTC TGCC CCACCC 4399
2731 AGCACC AGAA GGCC ACCAGAGAAACA X GUACAUUACCUGGUA 4324 GGCC TGAT GGTGCT 4400
2810 AGCUAC AGAA GCCG ACCAGAGAAACA X GUACAUUACCUGGUA 4325 CGGC AGCT GTAGCT 4401
2813 GGGAGC AGAA GCUG ACCAGAGAAACA X GUACAUUACCUGGUA 4326 CAGC TGTA GCTCCC 4402
2876 UCUCUG AGAA GGCU ACCAGAGAAACA X GUACAUUACCUGGUA 4327 AGCC AGTA CAGAGA 4403
2999 UACAUA AGAA GGAC ACCAGAGAAACA X GUACAUUACCUGGUA 4328 GTCC TGTA TATGTA 4404
3085 AGAGCC AGAA GCUC ACCAGAGAAACA X GUACAUUACCUGGUA 4329 GAGC AGCT GGCTCT 4405
3132 UAGCAG AGAA GCUC ACCAGAGAAACA X GUACAUUACCUGGUA 4330 GAGC TGCT CTGCTA 4406
Table 8
Table 8: Anti Human PTP-IB HH, NCH, and G Cleaver Ribozymes
Table 9
Table 9: Human methionine aminopeptidase type 2 (Met AP-2) Hammerhead Ribozyme and Target Sequence
528 CUUCUCGA CUGAUGAG X CGAA AAUCAUUC 43 GAATGATT T TCGAGAAG 455
529 GCUUCUCG CUGAUGAG X CGAA AAAUCAUU 44 AATGATTT T CGAGAAGC 456
530 AGCUUCUC CUGAUGAG X CGAA AAAAUCAU 45 ATGATTTT c GAGAAGCT 457
551 AACUUGUC CUGAUGAG X CGAA AUGUGCUU 46 AAGCACAT c GACAAGTT 458
559 UAUUUUCU CUGAUGAG X CGAA ACUUGUCG 47 CGACAAGT T AGAAAATA 459
560 GUAUUUUC CUGAUGAG X CGAA AACUUGUC 48 GACAAGTT A GAAAATAC 460
567 UCAUUACG CUGAUGAG X CGAA AUUUUCUA 49 TAGAAAAT A CGTAATGA 461
571 CAGCUCAU CUGAUGAG X CGAA ACGUAUUU 50 AAATACGT A ATGAGCTG 462
583 CCAGGCUU CUGAUGAG X CGAA AUCCAGCU 51 AGCTGGAT C AAGCCTGG 463
604 CAGAUUUC CUGAUGAG X CGAA AUCAUUGU 52 ACAATGAT A GAAATCTG 464
610 UUUUCACA CUGAUGAG X CGAA AUUUCUAU 53 ATAGAAAT C TGTGAAAA 465
621 AGUCUUCC CUGAUGAG X CGAA ACUUUUCA 54 TGAAAAGT T GGAAGACT 466
632 CUUGCGUG CUGAUGAG X CGAA ACAGUCUU 55 AAGACTGT T CACGCAAG 467
633 ACUUGCGU CUGAUGAG X CGAA AACAGUCU 56 AGACTGTT C ACGCAAGT 468
642 CUUUUAUU CUGAUGAG X CGAA ACUUGCGU 57 ACGCAAGT T AATAAAAG 469
643 UCUUUUAU CUGAUGAG X CGAA AACUUGCG 58 CGCAAGTT A ATAAAAGA 470
646 UUCUCUUU CUGAUGAG X CGAA AUUAACUU 59 AAGTTAAT A AAAGAGAA 471
660 CUGCAUUU CUGAUGAG X CGAA AUCCAUUC 60 GAATGGAT T AAATGCAG 472
661 CCUGCAUU CUGAUGAG X CGAA AAUCCAUU 61 AATGGATT A AATGCAGG 473
678 CAGUAGGA CUGAUGAG X CGAA AUGCCAGG 62 CCTGGCAT T TCCTACTG 474
679 CCAGUAGG CUGAUGAG X CGAA AAUGCCAG 63 CTGGCATT T CCTACTGG 475
680 UCCAGUAG CUGAUGAG X CGAA AAAUGCCA 64 TGGCATTT c CTACTGGA 476
683 ACAUCCAG CUGAUGAG X CGAA AGGAAAUG 65 CATTTCCT A CTGGATGT 477
692 AUUGAGAG CUGAUGAG X CGAA ACAUCCAG 66 CTGGATGT T CTCTCAAT 478
693 UAUUGAGA CUGAUGAG X CGAA AACAUCCA 67 TGGATGTT C TCTCAATA 479
695 AUUAUUGA CUGAUGAG X CGAA AGAACAUC 68 GATGTTCT C TCAATAAT 480
697 CAAUUAUU CUGAUGAG X CGAA AGAGAACA 69 TGTTCTCT C AATAATTG 481
701 AGCACAAU CUGAUGAG X CGAA AUUGAGAG 70 CTCTCAAT A ATTGTGCT 482
704 GGCAGCAC CUGAUGAG X CGAA AUUAUUGA 71 TCAATAAT T GTGCTGCC 483
716 GGGAGUAU CUGAUGAG X CGAA AUGGGCAG 72 CTGCCCAT T ATACTCCC 484
717 UGGGAGUA CUGAUGAG X CGAA AAUGGGCA 73 TGCCCATT A TACTCCCA 485
719 AUUGGGAG CUGAUGAG X CGAA AUAAUGGG 74 CCCATTAT A CTCCCAAT 486
722 GGCAUUGG CUGAUGAG X CGAA AGUAUAAU 75 ATTATACT C CCAATGCC 487
745 UACUGUAA CUGAUGAG X CGAA ACUGUUGU 76 ACAACAGT A TTACAGTA 488
747 CAUACUGU CUGAUGAG X CGAA AUACUGUU 77 AACAGTAT T ACAGTATG 489
748 UCAUACUG CUGAUGAG X CGAA AAUACUGU 78 ACAGTATT A CAGTATGA 490
753 UGUCAUCA CUGAUGAG X CGAA ACUGUAAU 79 ATTACAGT A TGATGACA 491
763 AUUUUACA CUGAUGAG X CGAA AUGUCAUC 80 GATGACAT C TGTAAAAT 492
767 GUCUAUUU CUGAUGAG X CGAA ACAGAUGU 81 ACATCTGT A AAATAGAC 493
772 CCAAAGUC CUGAUGAG X CGAA AUUUUACA 82 TGTAAAAT A GACTTTGG 494
777 GUGUUCCA CUGAUGAG X CGAA AGUCUAUU 83 AATAGACT T TGGAACAC 495
778 UGUGUUCC CUGAUGAG X CGAA AAGUCUAU 84 ATAGACTT T GGAACACA 496
788 ACCACUUA CUGAUGAG X CGAA AUGUGUUC 85 GAACACAT A TAAGTGGT 497
790 CUACCACU CUGAUGAG X CGAA AUAUGUGU 86 ACACATAT A AGTGGTAG 498
797 AAUAAUCC CUGAUGAG X CGAA ACCACUUA 87 TAAGTGGT A GGATTATT 499
802 CAGUCAAU CUGAUGAG X CGAA AUCCUACC 88 GGTAGGAT T ATTGACTG 500
803 ACAGUCAA CUGAUGAG X CGAA AAUCCUAC 89 GTAGGATT A TTGACTGT 501
Table 9
805 GCACAGUC CUGAUGAG X CGAA AUAAUCCU 90 AGGATTAT T GACTGTGC 502
815 GACAGUAA CUGAUGAG X CGAA AGCACAGU 91 ACTGTGCT T TTACTGTC 503
816 UGACAGUA CUGAUGAG X CGAA AAGCACAG 92 CTGTGCTT T TACTGTCA 504
817 GUGACAGU CUGAUGAG X CGAA AAAGCACA 93 TGTGCTTT T ACTGTCAC 505
818 AGUGACAG CUGAUGAG X CGAA AAAAGCAC 94 GTGCTTTT A CTGTCACT 506
823 UUAAAAGU CUGAUGAG X CGAA ACAGUAAA 95 TTTACTGT C ACTTTTAA 507
827 GGGAUUAA CUGAUGAG X CGAA AGUGACAG 96 CTGTCACT T TTAATCCC 508
828 UGGGAUUA CUGAUGAG X CGAA AAGUGACA 97 TGTCACTT T TAATCCCA 509
829 UUGGGAUU CUGAUGAG X CGAA AAAGUGAC 98 GTCACTTT T AATCCCAA 510
830 UUUGGGAU CUGAUGAG X CGAA AAAAGUGA 99 TCACTTTT A ATCCCAAA 511
833 AUAUUUGG CUGAUGAG X CGAA AUUAAAAG 100 CTTTTAAT C CCAAATAT 512
840 ACGUAUCA CUGAUGAG X CGAA AUUUGGGA 101 TCCCAAAT A TGATACGT 513
845 UAAUAACG CUGAUGAG X CGAA AUCAUAUU 102 AATATGAT A CGTTATTA 514
849 CUUUUAAU CUGAUGAG X CGAA ACGUAUCA 103 TGATACGT T ATTAAAAG 515
850 GCUUUUAA CUGAUGAG X CGAA AACGUAUC 104 GATACGTT A TTAAAAGC 516
852 CAGCUUUU CUGAUGAG X CGAA AUAACGUA 105 TACGTTAT T AAAAGCTG 517
853 ACAGCUUU CUGAUGAG X CGAA AAUAACGU 106 ACGTTATT A AAAGCTGT 518
862 GCAUCUUU CUGAUGAG X CGAA ACAGCUUU 107 AAAGCTGT A AAAGATGC 519
872 AGUGUUAG CUGAUGAG X CGAA AGCAUCUU 108 AAGATGCT A CTAACACT 520
875 UCCAGUGU CUGAUGAG X CGAA AGUAGCAU 109 ATGCTACT A ACACTGGA 521
886 GCACACUU CUGAUGAG X CGAA AUUCCAGU 110 ACTGGAAT A AAGTGTGC 522
901 CGAACAUC CUGAUGAG X CGAA AUUCCAGC 111 GCTGGAAT T GATGTTCG 523
907 CACAGACG CUGAUGAG X CGAA ACAUCAAU 112 ATTGATGT T CGTCTGTG 524
908 ACACAGAC CUGAUGAG X CGAA AACAUCAA 113 TTGATGTT C GTCTGTGT 525
911 AUCACACA CUGAUGAG X CGAA ACGAACAU 114 ATGTTCGT C TGTGTGAT 526
922 GCCUCACC CUGAUGAG X CGAA ACAUCACA 115 TGTGATGT T GGTGAGGC 527
934 ACUUCUUG CUGAUGAG X CGAA AUGGCCUC 116 GAGGCCAT C CAAGAAGT 528
943 GACUCCAU CUGAUGAG X CGAA ACUUCUUG 117 CAAGAAGT T ATGGAGTC 529
944 GGACUCCA CUGAUGAG X CGAA AACUUCUU 118 AAGAAGTT A TGGAGTCC 530
951 CUUCAUAG CUGAUGAG X CGAA ACUCCAUA 119 TATGGAGT C CTATGAAG 531
954 CAACUUCA CUGAUGAG X CGAA AGGACUCC 120 GGAGTCCT A TGAAGTTG 532
961 UCUAUUUC CUGAUGAG X CGAA ACUUCAUA 121 TATGAAGT T GAAATAGA 533
967 UUCCCAUC CUGAUGAG X CGAA AUUUCAAC 122 GTTGAAAT A GATGGGAA 534
981 UCACUUGA CUGAUGAG X CGAA AUGUCUUC 123 GAAGACAT A TCAAGTGA 535
983 UUUCACUU CUGAUGAG X CGAA AUAUGUCU 124 AGACATAT C AAGTGAAA 536
997 AGAUUACG CUGAUGAG X CGAA AUUGGUUU 125 AAACCAAT C CGTAATCT 537
1001 AUUUAGAU CUGAUGAG X CGAA ACGGAUUG 126 CAATCCGT A ATCTAAAT 538
1004 UCCAUUUA CUGAUGAG X CGAA AUUACGGA 127 TCCGTAAT C TAAATGGA 539
1006 UGUCCAUU CUGAUGAG X CGAA AGAUUACG 128 CGTAATCT A AATGGACA 540
1016 CCCAAUUG CUGAUGAG X CGAA AUGUCCAU 129 ATGGACAT T CAATTGGG 541
1017 GCCCAAUU CUGAUGAG X CGAA AAUGUCCA 130 TGGACATT C AATTGGGC 542
1021 UAUUGCCC CUGAUGAG X CGAA AUUGAAUG 131 CATTCAAT T GGGCAATA 543
1029 GUAUUCUA CUGAUGAG X CGAA AUUGCCCA 132 TGGGCAAT A TAGAATAC 544
1031 AUGUAUUC CUGAUGAG X CGAA AUAUUGCC 133 GGCAATAT A GAATACAT 545
1036 CCAGCAUG CUGAUGAG X CGAA AUUCUAUA 134 TATAGAAT A CATGCTGG 546
1060 CCUUUCAC CUGAUGAG X CGAA AUCGGCAC 135 GTGCCGAT T GTGAAAGG 547
1102 AUUGCAUA CUGAUGAG X CGAA ACUUCUCC 136 GGAGAAGT A TATGCAAT 548
Table 9
1104 CAAUUGCA CUGAUGAG X CGAA AUACUUCU 137 AGAAGTAT A TGCAATTG 549 llll AAGGUUUC CUGAUGAG X CGAA AUUGCAUA 138 TATGCAAT T GAAACCTT 550
1119 UACUACCA CUGAUGAG X CGAA AGGUUUCA 139 TGAAACCT T TGGTAGTA 551
1120 GUACUACC CUGAUGAG X CGAA AAGGUUUC 140 GAAACCTT T GGTAGTAC 552
1124 UCCUGUAC CUGAUGAG X CGAA ACCAAAGG 141 CCTTTGGT A GTACAGGA 553
1127 UUUUCCUG CUGAUGAG X CGAA ACUACCAA 142 TTGGTAGT A CAGGAAAA 554
1141 UCAUGAAC CUGAUGAG X CGAA ACACCUUU 143 AAAGGTGT T GTTCATGA 555
1144 UCAUCAUG CUGAUGAG X CGAA ACAACACC 144 GGTGTTGT T CATGATGA 556
1145 AUCAUCAU CUGAUGAG X CGAA AACAACAC 145 GTGTTGTT C ATGATGAT 557
1154 ACAUUCCA CUGAUGAG X CGAA AUCAUCAU 146 ATGATGAT A TGGAATGT 558
1163 GUAAUGUG CUGAUGAG X CGAA ACAUUCCA 147 TGGAATGT T CACATTAC 559
1164 UGUAAUGU CUGAUGAG X CGAA AACAUUCC 148 GGAATGTT C ACATTACA 560
1169 UUUCAUGU CUGAUGAG X CGAA AUGUGAAC 149 GTTCACAT T ACATGAAA 561
1170 UUUUCAUG CUGAUGAG X CGAA AAUGUGAA 150 TTCACATT A CATGAAAA 562
1181 AACAUCAA CUGAUGAG X CGAA AUUUUUCA 151 TGAAAAAT T TTGATGTT 563
1182 CAACAUCA CUGAUGAG X CGAA AAUUUUUC 152 GAAAAATT T TGATGTTG 564
1183 CCAACAUC CUGAUGAG X CGAA AAAUUUUU 153 AAAAATTT T GATGTTGG 565
1189 ACAUGUCC CUGAUGAG X CGAA ACAUCAAA 154 TTTGATGT T GGACATGT 566
1204 GGAAGCCU CUGAUGAG X CGAA AUUGGCAC 155 GTGCCAAT A AGGCTTCC 567
1210 GUUCUUGG CUGAUGAG X CGAA AGCCUUAU 156 ATAAGGCT T CCAAGAAC 568
1211 UGUUCUUG CUGAUGAG X CGAA AAGCCUUA 157 TAAGGCTT C CAAGAACA 569
1227 CAUUUAAC CUGAUGAG X CGAA AGUGUUUU 158 AAAACACT T GTTAAATG 570
1230 UGACAUUU CUGAUGAG X CGAA ACAAGUGU 159 ACACTTGT T AAATGTCA 571
1231 AUGACAUU CUGAUGAG X CGAA AACAAGUG 160 CACTTGTT A AATGTCAT 572
1237 UCAUUGAU CUGAUGAG X CGAA ACAUUUAA 161 TTAAATGT C ATCAATGA 573
1240 UUUUCAUU CUGAUGAG X CGAA AUGACAUU 162 AATGTCAT C AATGAAAA 574
1251 GGGUUCCA CUGAUGAG X CGAA AGUUUUCA 163 TGAAAACT T TGGAACCC 575
1252 AGGGUUCC CUGAUGAG X CGAA AAGUUUUC 164 GAAAACTT T GGAACCCT 576
1261 CAGAAGGC CUGAUGAG X CGAA AGGGUUCC 165 GGAACCCT T GCCTTCTG 577
1266 UGCGGCAG CUGAUGAG X CGAA AGGCAAGG 166 CCTTGCCT T CTGCCGCA 578
1267 CUGCGGCA CUGAUGAG X CGAA AAGGCAAG 167 CTTGCCTT c TGCCGCAG 579
1286 UCCCAAGC CUGAUGAG X CGAA AUCCAGCC 168 GGCTGGAT c GCTTGGGA 580
1290 UUUCUCCC CUGAUGAG X CGAA AGCGAUCC 169 GGATCGCT T GGGAGAAA 581
1301 CAAGUAUU CUGAUGAG X CGAA ACUUUCUC 170 GAGAAAGT A AATACTTG 582
1305 CCAUCAAG CUGAUGAG X CGAA AUUUACUU 171 AAGTAAAT A CTTGATGG 583
1308 GAGCCAUC CUGAUGAG X CGAA AGUAUUUA 172 TAAATACT T GATGGCTC 584
1316 AUUCUUCA CUGAUGAG X CGAA AGCCAUCA 173 TGATGGCT c TGAAGAAT 585
1325 GUCACACA CUGAUGAG X CGAA AUUCUUCA 174 TGAAGAAT c TGTGTGAC 586
1335 CAAUGCCC CUGAUGAG X CGAA AGUCACAC 175 GTGTGACT T GGGCATTG 587
1342 GGAUCUAC CUGAUGAG X CGAA AUGCCCAA 176 TTGGGCAT T GTAGATCC 588
1345 UAUGGAUC CUGAUGAG X CGAA ACAAUGCC 177 GGCATTGT A GATCCATA 589
1349 UGGAUAUG CUGAUGAG X CGAA AUCUACAA 178 TTGTAGAT C CATATCCA 590
1353 AUGGUGGA CUGAUGAG X CGAA AUGGAUCU 179 AGATCCAT A TCCACCAT 591
1355 UAAUGGUG CUGAUGAG X CGAA AUAUGGAU 180 ATCCATAT C CACCATTA 592
1362 UGUCACAU CUGAUGAG X CGAA AUGGUGGA 181 TCCACCAT T ATGTGACA 593
1363 AUGUCACA CUGAUGAG X CGAA AAUGGUGG 182 CCACCATT A TGTGACAT 594
1372 GAUCCUUU CUGAUGAG X CGAA AUGUCACA 183 TGTGACAT T AAAGGATC 595
Table 9
1373 UGAUCCUU CUGAUGAG X CGAA AAUGUCAC 184 GTGACATT A AAGGATCA 596
1380 CUGUAUAU CUGAUGAG X CGAA AUCCUUUA 185 TAAAGGAT C ATATACAG 597
1383 GCGCUGUA CUGAUGAG X CGAA AUGAUCCU 186 AGGATCAT A TACAGCGC 598
1385 UUGCGCUG CUGAUGAG X CGAA AUAUGAUC 187 GATCATAT A CAGCGCAA 599
1395 UAUGUUCA CUGAUGAG X CGAA AUUGCGCU 188 AGCGCAAT T TGAACATA 600
1396 GUAUGUUC CUGAUGAG X CGAA AAUUGCGC 189 GCGCAATT T GAACATAC 601
1403 CAGGAUGG CUGAUGAG X CGAA AUGUUCAA 190 TTGAACAT A CCATCCTG 602
1408 CGCAACAG CUGAUGAG X CGAA AUGGUAUG 191 CATACCAT C CTGTTGCG 603
1413 UUGGACGC CUGAUGAG X CGAA ACAGGAUG 192 CATCCTGT T GCGTCCAA 604
1418 ACAUGUUG CUGAUGAG X CGAA ACGCAACA 193 TGTTGCGT C CAACATGT 605
1427 AACUUCUU CUGAUGAG X CGAA ACAUGUUG 194 CAACATGT A AAGAAGTT 606
1435 CUGCUGAC CUGAUGAG X CGAA ACUUCUUU 195 AAAGAAGT T GTCAGCAG 607
1438 CCUCUGCU CUGAUGAG X CGAA ACAACUUC 196 GAAGTTGT C AGCAGAGG 608
1455 AAGUUUAA CUGAUGAG X CGAA AGUCAUCU 197 AGATGACT A TTAAACTT 609
1457 CUAAGUUU CUGAUGAG X CGAA AUAGUCAU 198 ATGACTAT T AAACTTAG 610
1458 ACUAAGUU CUGAUGAG X CGAA AAUAGUCA 199 TGACTATT A AACTTAGT 611
1463 UUUGGACU CUGAUGAG X CGAA AGUUUAAU 200 ATTAAACT T AGTCCAAA 612
1464 CUUUGGAC CUGAUGAG X CGAA AAGUUUAA 201 TTAAACTT A GTCCAAAG 613
1467 UGGCUUUG CUGAUGAG X CGAA ACUAAGUU 202 AACTTAGT C CAAAGCCA 614
1479 AAGGUGUU CUGAUGAG X CGAA AGGUGGCU 203 AGCCACCT C AACACCTT 615
1487 AGAAAAUA CUGAUGAG X CGAA AGGUGUUG 204 CAACACCT T TATTTTCT 616
1488 CAGAAAAU CUGAUGAG X CGAA AAGGUGUU 205 AACACCTT T ATTTTCTG 617
1489 UCAGAAAA CUGAUGAG X CGAA AAAGGUGU 206 ACACCTTT A TTTTCTGA 618
1491 GCUCAGAA CUGAUGAG X CGAA AUAAAGGU 207 ACCTTTAT T TTCTGAGC 619
1492 AGCUCAGA CUGAUGAG X CGAA AAUAAAGG 208 CCTTTATT T TCTGAGCT 620
1493 AAGCUCAG CUGAUGAG X CGAA AAAUAAAG 209 CTTTATTT T CTGAGCTT 621
1494 AAAGCUCA CUGAUGAG X CGAA AAAAUAAA 210 TTTATTTT C TGAGCTTT 622
1501 UUCCAACA CUGAUGAG X CGAA AGCUCAGA 211 TCTGAGCT T TGTTGGAA 623
1502 UUUCCAAC CUGAUGAG X CGAA AAGCUCAG 212 CTGAGCTT T GTTGGAAA 624
1505 UGUUUUCC CUGAUGAG X CGAA ACAAAGCU 213 AGCTTTGT T GGAAAACA 625
1518 AAUUCUGG CUGAUGAG X CGAA AUCAUGUU 214 AACATGAT A CCAGAATT 626
1526 GGCAAAUU CUGAUGAG X CGAA AUUCUGGU 215 ACCAGAAT T AATTTGCC 627
1527 UGGCAAAU CUGAUGAG X CGAA AAUUCUGG 216 CCAGAATT A ATTTGCCA 628
1530 AUGUGGCA CUGAUGAG X CGAA AUUAAUUC 217 GAATTAAT T TGCCACAT 629
1531 CAUGUGGC CUGAUGAG X CGAA AAUUAAUU 218 AATTAATT T GCCACATG 630
1541 AAACAGAC CUGAUGAG X CGAA ACAUGUGG 219. CCACATGT T GTCTGTTT 631
1544 UUAAAACA CUGAUGAG X CGAA ACAACAUG 220 CATGTTGT c TGTTTTAA 632
1548 ACUGUUAA CUGAUGAG X CGAA ACAGACAA 221 TTGTCTGT T TTAACAGT 633
1549 CACUGUUA CUGAUGAG X CGAA AACAGACA 222 TGTCTGTT T TAACAGTG 634
1550 CCACUGUU CUGAUGAG X CGAA AAACAGAC 223 GTCTGTTT T AACAGTGG 635
1551 UCCACUGU CUGAUGAG X CGAA AAAACAGA 224 TCTGTTTT A ACAGTGGA 636
1567 AAAAGUAU CUGAUGAG X CGAA ACAUGGGU 225 ACCCATGT A ATACTTTT 637
1570 GAUAAAAG CUGAUGAG X CGAA AUUACAUG 226 CATGTAAT A CTTTTATC 638
1573 AUGGAUAA CUGAUGAG X CGAA AGUAUUAC 227 GTAATACT T TTATCCAT 639
1574 CAUGGAUA CUGAUGAG X CGAA AAGUAUUA 228 TAATACTT T TATCCATG 640
1575 ACAUGGAU CUGAUGAG X CGAA AAAGUAUU 229 AATACTTT T ATCCATGT 641
1576 AACAUGGA CUGAUGAG X CGAA AAAAGUAU 230 ATACTTTT A TCCATGTT 642
Table 9
1578 UAAACAUG CUGAUGAG X CGAA AUAAAAGU 231 ACTTTTAT C CATGTTTA 643
1584 CUUUUUUA CUGAUGAG X CGAA ACAUGGAU 232 ATCCATGT T TAAAAAAG 644
1585 UCUUUUUU CUGAUGAG X CGAA AACAUGGA 233 TCCATGTT T AAAAAAGA 645
1586 UUCUUUUU CUGAUGAG X CGAA AAACAUGG 234 CCATGTTT A AAAAAGAA 646
1600 UUUGUCCA CUGAUGAG X CGAA AUUCCUUC 235 GAAGGAAT T TGGACAAA 647
1601 CUUUGUCC CUGAUGAG X CGAA AAUUCCUU 236 AAGGAATT T GGACAAAG 648
1619 UUACAUUA CUGAUGAG X CGAA ACGGUUUG 237 CAAACCGT C TAATGTAA 649
1621 AAUUACAU CUGAUGAG X CGAA AGACGGUU 238 AACCGTCT A ATGTAATT 650
1626 UGGUUAAU CUGAUGAG X CGAA ACAUUAGA 239 TCTAATGT A ATTAACCA 651
1629 CGUUGGUU CUGAUGAG X CGAA AUUACAUU 240 AATGTAAT T AACCAACG 652
1630 UCGUUGGU CUGAUGAG X CGAA AAUUACAU 241 ATGTAATT A ACCAACGA 653
1646 AGUCCGGA CUGAUGAG X CGAA AGCUUUUU 242 AAAAAGCT T TCCGGACT 654
1647 AAGUCCGG CUGAUGAG X CGAA AAGCUUUU 243 AAAAGCTT T CCGGACTT 655
, 1648 AAAGUCCG CUGAUGAG X CGAA AAAGCUUU 244 AAAGCTTT c CGGACTTT 656
1655 GCAUUUAA CUGAUGAG X CGAA AGUCCGGA 245 TCCGGACT T TTAAATGC 657
1656 AGCAUUUA CUGAUGAG X CGAA AAGUCCGG 246 CCGGACTT T TAAATGCT 658
1657 UAGCAUUU CUGAUGAG X CGAA AAAGUCCG 247 CGGACTTT T AAATGCTA 659
1658 UUAGCAUU CUGAUGAG X CGAA AAAAGUCC 248 GGACTTTT A AATGCTAA 660
1665 AAAACAGU CUGAUGAG X CGAA AGCAUUUA 249 TAAATGCT A ACTGTTTT 661
1671 AGGGGAAA CUGAUGAG X CGAA ACAGUUAG 250 CTAACTGT T TTTCCCCT 662
1672 AAGGGGAA CUGAUGAG X CGAA AACAGUUA 251 TAACTGTT T TTCCCCTT 663
1673 GAAGGGGA CUGAUGAG X CGAA AAACAGUU 252 AACTGTTT T TCCCCTTC 664
1674 GGAAGGGG CUGAUGAG X CGAA AAAACAGU 253 ACTGTTTT T CCCCTTCC 665
1675 AGGAAGGG CUGAUGAG X CGAA AAAAACAG 254 CTGTTTTT c CCCTTCCT 666
1680 UAGACAGG CUGAUGAG X CGAA AGGGGAAA 255 TTTCCCCT T CCTGTCTA 667
1681 CUAGACAG CUGAUGAG X CGAA AAGGGGAA 256 TTCCCCTT c CTGTCTAG 668
1686 UUUUCCUA CUGAUGAG X CGAA ACAGGAAG 257 CTTCCTGT c TAGGAAAA 669
1688 CAUUUUCC CUGAUGAG X CGAA AGACAGGA 258 TCCTGTCT A GGAAAATG 670
1699 GAGCUUUA CUGAUGAG X CGAA AGCAUUUU 259 AAAATGCT A TAAAGCTC 671
1701 UUGAGCUU CUGAUGAG X CGAA AUAGCAUU 260 AATGCTAT A AAGCTCAA 672
1707 ACUAAUUU CUGAUGAG X CGAA AGCUUUAU 261 ATAAAGCT C AAATTAGT 673
1712 UCCUAACU CUGAUGAG X CGAA AUUUGAGC 262 GCTCAAAT T AGTTAGGA 674
1713 UUCCUAAC CUGAUGAG X CGAA AAUUUGAG 263 CTCAAATT A GTTAGGAA 675
1716 UCAUUCCU CUGAUGAG X CGAA ACUAAUUU 264 AAATTAGT T AGGAATGA 676
1717. GUCAUUCC CUGAUGAG X CGAA AACUAAUU 265 AATTAGTT A GGAATGAC 677
1727 AAACGUAU CUGAUGAG X CGAA AGUCAUUC 266 GAATGACT T ATACGTTT 678
1728 AAAACGUA CUGAUGAG X CGAA AAGUCAUU 267 AATGACTT A TACGTTTT 679
1730 ACAAAACG CUGAUGAG X CGAA AUAAGUCA 268 TGACTTAT A CGTTTTGT 680
1734 CAAAACAA CUGAUGAG X CGAA ACGUAUAA 269 TTATACGT T TTGTTTTG 681
1735 UCAAAACA CUGAUGAG X CGAA AACGUAUA 270 TATACGTT T TGTTTTGA 682
1736 UUCAAAAC CUGAUGAG X CGAA AAACGUAU 271 "~ ATACGTTT T GTTTTGAA 683
1739 GUAUUCAA CUGAUGAG X CGAA ACAAAACG 272 CGTTTTGT T TTGAATAC 684
1740 GGUAUUCA CUGAUGAG X CGAA AACAAAAC 273 GTTTTGTT T TGAATACC 685
1741 AGGUAUUC CUGAUGAG X CGAA AAACAAAA 274 TTTTGTTT T GAATACCT 686
1746 CUCUUAGG CUGAUGAG X CGAA AUUCAAAA 275 TTTTGAAT A CCTAAGAG 687
1750 GUAUCUCU CUGAUGAG X CGAA AGGUAUUC 276 GAATACCT A AGAGATAC 688
1757 CCAAAAAG CUGAUGAG X CGAA AUCUCUUA 277 TAAGAGAT A CTTTTTGG 689
Table 9
1760 UAUCCAAA CUGAUGAG X CGAA AGUAUCUC 278 GAGATACT T TTTGGATA 690
1761 AUAUCCAA CUGAUGAG X CGAA AAGUAUCU 279 AGATACTT T TTGGATAT 691
1762 AAUAUCCA CUGAUGAG X CGAA AAAGUAUC 280 GATACTTT T TGGATATT 692
1763 AAAUAUCC CUGAUGAG X CGAA AAAAGUAU 281 ATACTTTT T GGATATTT 693
1768 AAUAUAAA CUGAUGAG X CGAA AUCCAAAA 282 TTTTGGAT A TTTATATT 694
1770 GCAAUAUA CUGAUGAG X CGAA AUAUCCAA 283 TTGGATAT T TATATTGC 695
1771 GGCAAUAU CUGAUGAG X CGAA AAUAUCCA 284 TGGATATT T ATATTGCC 696
1772 UGGCAAUA CUGAUGAG X CGAA AAAUAUCC 285 GGATATTT A TATTGCCA 697
1774 UAUGGCAA CUGAUGAG X CGAA AUAAAUAU 286 ATATTTAT A TTGCCATA 698
1776 AAUAUGGC CUGAUGAG X CGAA AUAUAAAU 287 ATTTATAT T GCCATATT 699
1782 AGUAAGAA CUGAUGAG X CGAA AUGGCAAU 288 ATTGCCAT A TTCTTACT 700
1784 CAAGUAAG CUGAUGAG X CGAA AUAUGGCA 289 TGCCATAT T CTTACTTG 701
1785 UCAAGUAA CUGAUGAG X CGAA AAUAUGGC 290 GCCATATT C TTACTTGA 702
1787 AUUCAAGU CUGAUGAG X CGAA AGAAUAUG 291 CATATTCT T ACTTGAAT 703
1788 CAUUCAAG CUGAUGAG X CGAA AAGAAUAU 292 ATATTCTT A CTTGAATG 704
1791 AAGCAUUC CUGAUGAG X CGAA AGUAAGAA 293 TTCTTACT T GAATGCTT 705
1799 GUCAUUCA CUGAUGAG X CGAA AGCAUUCA 294 TGAATGCT T TGAATGAC 706
1800 AGUCAUUC CUGAUGAG X CGAA AAGCAUUC 295 GAATGCTT T GAATGACT 707
1809 ACUGGAUG CUGAUGAG X CGAA AGUCAUUC 296 GAATGACT A CATCCAGT 708
1813 CAGAACUG CUGAUGAG X CGAA AUGUAGUC 297 GACTACAT C CAGTTCTG 709
1818 AGGUGCAG CUGAUGAG X CGAA ACUGGAUG 298 CATCCAGT T CTGCACCT 710
1819 UAGGUGCA CUGAUGAG X CGAA AACUGGAU 299 ATCCAGTT C TGCACCTA 711
1827 AGAGGGUA CUGAUGAG X CGAA AGGUGCAG 300 CTGCACCT A TACCCTCT 712
1829 CCAGAGGG CUGAUGAG X CGAA AUAGGUGC 301 GCACCTAT A CCCTCTGG 713
1834 CAACACCA CUGAUGAG X CGAA AGGGUAUA 302 TATACCCT C TGGTGTTG 714
1841 UAAAAAGC CUGAUGAG X CGAA ACACCAGA 303 TCTGGTGT T GCTTTTTA 715
1845 AGGUUAAA CUGAUGAG X CGAA AGCAACAC 304 GTGTTGCT T TTTAACCT 716
1846 AAGGUUAA CUGAUGAG X CGAA AAGCAACA 305 TGTTGCTT T TTAACCTT 717
1847 GAAGGUUA CUGAUGAG X CGAA AAAGCAAC 306 GTTGCTTT T TAACCTTC 718
1848 GGAAGGUU CUGAUGAG X CGAA AAAAGCAA 307 TTGCTTTT T AACCTTCC 719
1849 AGGAAGGU CUGAUGAG X CGAA AAAAAGCA 308 TGCTTTTT A ACCTTCCT 720
1854 AUUCCAGG CUGAUGAG X CGAA AGGUUAAA 309 TTTAACCT T CCTGGAAT 721
1855 GAUUCCAG CUGAUGAG X CGAA AAGGUUAA 310 TTAACCTT c CTGGAATC 722
1863 AGAAAAUG CUGAUGAG X CGAA AUUCCAGG 311 CCTGGAAT c CATTTTCT 723
1867 UUUUAGAA CUGAUGAG X CGAA AUGGAUUC 312 GAATCCAT T TTCTAAAA 724
1868 UUUUUAGA CUGAUGAG X CGAA AAUGGAUU 313 AATCCATT T TCTAAAAA 725
1869 UUUUUUAG CUGAUGAG X CGAA AAAUGGAU 314 ATCCATTT T CTAAAAAA 726
1870 AUUUUUUA CUGAUGAG X CGAA AAAAUGGA 315 TCCATTTT c TAAAAAAT 727
1872 UUAUUUUU CUGAUGAG X CGAA AGAAAAUG 316 CATTTTCT A AAAAATAA 728
1879 UGUGUCUU CUGAUGAG X CGAA AUUUUUUA 317 TAAAAAAT A AAGACACA 729
1889 CUGAGAAG CUGAUGAG X CGAA AUGUGUCU 318 AGACACAT T CTTCTCAG 730
1890 GCUGAGAA CUGAUGAG X CGAA AAUGUGUC 319 GACACATT C TTCTCAGC 731
1892 GUGCUGAG CUGAUGAG X CGAA AGAAUGUG 320 CACATTCT T CTCAGCAC 732
1893 GGUGCUGA CUGAUGAG X CGAA AAGAAUGU 321 ACATTCTT C TCAGCACC 733
1895 GUGGUGCU CUGAUGAG X CGAA AGAAGAAU 322 ATTCTTCT C AGCACCAC 734
1913 UUUUGGAA CUGAUGAG X CGAA AGGUGUUG 323 CAACACCT A TTCCAAAA 735
1915 GAUUUUGG CUGAUGAG X CGAA AUAGGUGU 324 ACACCTAT T CCAAAATC 736
Table 9
1916 CGAUUUUG CUGAUGAG X CGAA AAUAGGUG 325 CACCTATT C CAAAATCG 737
1923 AUGUGGUC CUGAUGAG X CGAA AUUUUGGA 326 TCCAAAAT C GACCACAT 738
1932 CUUCCAAA CUGAUGAG X CGAA AUGUGGUC 327 GACCACAT A TTTGGAAG 739
1934 UACUUCCA CUGAUGAG X CGAA AUAUGUGG 328 CCACATAT T TGGAAGTA 740
1935 UUACUUCC CUGAUGAG X CGAA AAUAUGUG 329 CACATATT T GGAAGTAA 741
1942 GAGAGCUU CUGAUGAG X CGAA ACUUCCAA 330 TTGGAAGT A AAGCTCTC 742
1948 GCUGAGGA CUGAUGAG X CGAA AGCUUUAC 331 GTAAAGCT C TCCTCAGC 743
1950 UUGCUGAG CUGAUGAG X CGAA AGAGCUUU 332 AAAGCTCT C CTCAGCAA 744
1953 CAUUUGCU CUGAUGAG X CGAA AGGAGAGC 333 GCTCTCCT C AGCAAATG 745
1963 UGUUCUUU CUGAUGAG X CGAA ACAUUUGC 334 GCAAATGT A AAAGAACA 746
1977 UUUGUUAU CUGAUGAG X CGAA AUUUCUGU 335 ACAGAAAT T ATAACAAA 747
1978 GUUUGUUA CUGAUGAG X CGAA AAUUUCUG 336 CAGAAATT A TAACAAAC 748
1980 CAGUUUGU CUGAUGAG X CGAA AUAAUUUC 337 GAAATTAT A ACAAACTG 749
1990 GUCUGAGA CUGAUGAG X CGAA ACAGUUUG 338 CAAACTGT C TCTCAGAC 750
1992 UGGUCUGA CUGAUGAG X CGAA AGACAGUU 339 ' AACTGTCT C TCAGACCA 751
1994 UGUGGUCU CUGAUGAG X CGAA AGAGACAG 340 CTGTCTCT C AGACCACA 752
2005 UUUGGUUA CUGAUGAG X CGAA ACUGUGGU 341 ACCACAGT A TAACCAAA 753
2007 AGUUUGGU CUGAUGAG X CGAA AUACUGUG 342 CACAGTAT A ACCAAACT 754
2016 CUGAGUUC CUGAUGAG X CGAA AGUUUGGU 343 ACCAAACT A GAACTCAG 755
2022 UUAAUCCU CUGAUGAG X CGAA AGUUCUAG 344 CTAGAACT C AGGATTAA 756
2028 AGUUUCUU CUGAUGAG X CGAA AUCCUGAG 345 CTCAGGAT T AAGAAACT 757
2029 GAGUUUCU CUGAUGAG X CGAA AAUCCUGA 346 TCAGGATT A AGAAACTC 758
2037 UUUUGAGU CUGAUGAG X CGAA AGUUUCUU 347 AAGAAACT C ACTCAAAA 759
2041 GUGGUUUU CUGAUGAG X CGAA AGUGAGUU 348 AACTCACT C AAAACCAC 760
2056 UUUCCAUG CUGAUGAG X CGAA AGUUGUGU 349 ACACAACT A CATGGAAA 761
2079 UCAUUCAG CUGAUGAG X CGAA AGCAGGUU 350 AACCTGCT C CTGAATGA 762
2090 GUAUCCAG CUGAUGAG X CGAA AGUCAUUC 351 GAATGACT A CTGGATAC 763
2097 UUGUUAUG CUGAUGAG X CGAA AUCCAGUA 352 TACTGGAT A CATAACAA 764
2101 CAUUUUGU CUGAUGAG X CGAA AUGUAUCC 353 GGATACAT A ACAAAATG 765
2121 AACAUCUU CUGAUGAG X CGAA AUUUCUGC 354 GCAGAAAT A AAGATGTT 766
2129 UUUUAAAG CUGAUGAG X CGAA ACAUCUUU 355 AAAGATGT T CTTTAAAA 767
2130 GUUUUAAA CUGAUGAG X CGAA AACAUCUU 356 AAGATGTT C TTTAAAAC 768
2132 UGGUUUUA CUGAUGAG X CGAA AGAACAUC 357 GATGTTCT T TAAAACCA 769
2133 UUGGUUUU CUGAUGAG X CGAA AAGAACAU 358 ATGTTCTT T AAAACCAA 770
2134 AUUGGUUU CUGAUGAG X CGAA AAAGAACA 359 TGTTCTTT A AAACCAAT 771
2162 GAUUCUGG CUGAUGAG X CGAA AUGUUGUG 360 CACAACAT A CCAGAATC 772
2170 GUCCCAGA CUGAUGAG X CGAA AUUCUGGU 361 ACCAGAAT C TCTGGGAC 773
2172 GUGUCCCA CUGAUGAG X CGAA AGAUUCUG 362 CAGAATCT C TGGGACAC 774
2183 CUGCUUUG CUGAUGAG X CGAA AUGUGUCC 363 GGACACAT T CAAAGCAG 775
2184 ACUGCUUU CUGAUGAG X CGAA AAUGUGUC 364 GACACATT C AAAGCAGT 776
2197 UUUCCCUC CUGAUGAG X CGAA ACACACUG 365 CAGTGTGT A GAGGGAAA 777
2207 GUGCUAUA CUGAUGAG X CGAA AUUUCCCU 366 AGGGAAAT T TATAGCAC 778
2208 AGUGCUAU CUGAUGAG X CGAA AAUUUCCC 367 GGGAAATT T ATAGCACT 779
2209 UAGUGCUA CUGAUGAG X CGAA AAAUUUCC 368 GGAAATTT A TAGCACTA 780
2211 UUUAGUGC CUGAUGAG X CGAA AUAAAUUU 369 AAATTTAT A GCACTAAA 781
2217 UGGGCAUU CUGAUGAG X CGAA AGUGCUAU 370 ATAGCACT A AATGCCCA 782
2244 AUUUUAGA CUGAUGAG X CGAA AUUUCCUG 371 CAGGAAAT A TCTAAAAT 783
Table 9
2246 CAAUUUUA CUGAUGAG X CGAA AUAUUUCC 372 GGAAATAT C TAAAATTG 784
2248 GUCAAUUU CUGAUGAG X CGAA AGAUAUUU 373 AAATATCT A AAATTGAC 785
2253 AGGGUGUC CUGAUGAG X CGAA AUUUUAGA 374 TCTAAAAT T GACACCCT 786
2262 UGUGAUGU CUGAUGAG X CGAA AGGGUGUC 375 GACACCCT A ACATCACA 787
2267 UUAAUUGU CUGAUGAG X CGAA AUGUUAGG 376 CCTAACAT C ACAATTAA 788
2273 GUUCUUUU CUGAUGAG X CGAA AUUGUGAU 377 ATCACAAT T AAAAGAAC 789
2274 AGUUCUUU CUGAUGAG X CGAA AAUUGUGA 378 TCACAATT A AAAGAACT 790
2283 UGCUUCUC CUGAUGAG X CGAA AGUUCUUU 379 AAAGAACT A GAGAAGCA 791
2305 AGCUUUUC CUGAUGAG X CGAA AUGUGUUU 380 AAACACAT T GAAAAGCT 792
2314 CCUUCUCU CUGAUGAG X CGAA AGCUUUUC 381 GAAAAGCT A AGAGAAGG 793
2331 AUCUUAGU CUGAUGAG X CGAA AUUUCUUG 382 CAAGAAAT A ACTAAGAT 794
2335 UCUGAUCU CUGAUGAG X CGAA AGUUAUUU 383 AAATAACT A AGATCAGA 795
2340 UCUGCUCU CUGAUGAG X CGAA AUCUUAGU 384 ACTAAGAT C AGAGCAGA 796
2361 UGUGUCUC CUGAUGAG X CGAA AUUUCCUU 385 AAGGAAAT A GAGACACA 797
2377 UUUUUGAA CUGAUGAG X CGAA AGUUUUUU 386 AAAAAACT C TTCAAAAA 798
2379 AUUUUUUG CUGAUGAG X CGAA AGAGUUUU 387 AAAACTCT T CAAAAAAT 799
2380 GAUUUUUU CUGAUGAG X CGAA AAGAGUUU 388 AAACTCTT C AAAAAATC 800
2388 GAUUCAUU CUGAUGAG X CGAA AUUUUUUG 389 CAAAAAAT C AATGAATC 801
2396 AGCUCCUG CUGAUGAG X CGAA AUUCAUUG 390 CAATGAAT C CAGGAGCT 802
2408 UUUCAAAA CUGAUGAG X CGAA ACCAGCUC 391 GAGCTGGT T TTTTGAAA 803
2409 GUUUCAAA CUGAUGAG X CGAA AACCAGCU 392 AGCTGGTT T TTTGAAAC 804
2410 CGUUUCAA CUGAUGAG X CGAA AAACCAGC 393 GCTGGTTT T TTGAAACG 805
2411 UCGUUUCA CUGAUGAG X CGAA AAAACCAG 394 CTGGTTTT T TGAAACGA 806
2412 AUCGUUUC CUGAUGAG X CGAA AAAAACCA 395 TGGTTTTT T GAAACGAT 807
2421 AUUUUGUU CUGAUGAG X CGAA AUCGUUUC 396 GAAACGAT C AACAAAAT 808
2430 UGUCUAUC CUGAUGAG X CGAA AUUUUGUU 397 AACAAAAT T GATAGACA 809
2434 CUAGUGUC CUGAUGAG X CGAA AUCAAUUU 398 AAATTGAT A GACACTAG 810
2441 AGUCUUGC CUGAUGAG X CGAA AGUGUCUA 399 TAGACACT A GCAAGACT 811
2450 . UUCUUUAU CUGAUGAG X CGAA AGUCUUGC 400 GCAAGACT A ATAAAGAA 812
2453 UUCUUCUU CUGAUGAG X CGAA AUUAGUCU 401 AGACTAAT A AAGAAGAA 813
2475 UUCUAUUU CUGAUGAG X CGAA AUUCUUCU 402 AGAAGAAT C AAATAGAA 814
2480 AUUGCUUC CUGAUGAG X CGAA AUUUGAUU 403 AATCAAAT A GAAGCAAT 815
2489 UCAUUUUU CUGAUGAG X CGAA AUUGCUUC 404 GAAGCAAT A AAAAATGA 816
2499 AUCCCCUU CUGAUGAG X CGAA AUCAUUUU 405 AAAATGAT A AAGGGGAT 817
2508 GGUGGUGA CUGAUGAG X CGAA AUCCCCUU 406 AAGGGGAT A TCACCACC 818
2510 UUGGUGGU CUGAUGAG X CGAA AUAUCCCC 407 GGGGATAT C ACCACCAA 819
2520 UUCUGUGG CUGAUGAG X CGAA AUUGGUGG 408 CCACCAAT C CCACAGAA 820
2531 UGGUGGUU CUGAUGAG X CGAA AUUUCUGU 409 ACAGAAAT A AACCACCA 821
2541 UAUUCUCU CUGAUGAG X CGAA AUGGUGGU 410 ACCACCAT C AGAGAATA 822
2549 GUUUGUAG CUGAUGAG X CGAA AUUCUCUG 411 CAGAGAAT A CTACAAAC 823
2552 GGUGUUUG CUGAUGAG X CGAA AGUAUUCU 412 AGAATACT A CAAACACC 824
Input Sequence = HSU29607. Cut Site = UH/.
Stem Length = 8 . Core Sequence = CUGAUGAG X CGAA (X = GCCGUUAGGC or other stem II)
Seql = HSU29607 (Human methionine aminopeptidase mRNA, complete eds., 2569 bp)
Table 10
Table 10: Human methionine aminopeptidase type 2 (MetAP-2) NCH Ribozyme and Target Sequence
Nt. Ribozyme Sequence Seq. ID Substrate Sequence Seq. ID Nos. position Nos.
10 CCCGAGAG CUGAUGAG X CGAA IACGAGGG 825 CCCTCGTC T CTCTCGGG 1255
12 UGCCCGAG CUGAUGAG X CGAA IAGACGAG 826 CTCGTCTC T CTCGGGCA 1256
14 GUUGCCCG CUGAUGAG X CGAA IAGAGACG 827 CGTCTCTC T CGGGCAAC 1257
20 CGCCAUGU CUGAUGAG X CGAA ICCCGAGA 828 TCTCGGGC A ACATGGCG 1258
23 GCCCGCCA CUGAUGAG X CGAA IUUGCCCG 829 CGGGCAAC A TGGCGGGC 1259
49 CUCCCGGA CUGAUGAG X CGAA ICCGCUAC 830 GTAGCGGC C TCCGGGAG 1260
50 GCUCCCGG CUGAUGAG X CGAA IGCCGCUA 831 TAGCGGCC T CCGGGAGC 1261
52 UGGCUCCC CUGAUGAG X CGAA IAGGCCGC 832 GCGGCCTC C GGGAGCCA 1262
59 AUUCAGGU CUGAUGAG X CGAA ICUCCCGG 833 CCGGGAGC C ACCTGAAT 1263
60 CAUUCAGG CUGAUGAG X CGAA IGCUCCCG 834 CGGGAGCC A CCTGAATG 1264
62 GCCAUUCA CUGAUGAG X CGAA IUGGCUCC 835 GGAGCCAC C TGAATGGC 1265
63 CGCCAUUC CUGAUGAG X CGAA IGUGGCUC 836 GAGCCACC T GAATGGCG 1266
74 UGGAUCCA CUGAUGAG X CGAA IUCGCCAU 837 ATGGCGAC C TGGATCCA 1267
75 CUGGAUCC CUGAUGAG X CGAA IGUCGCCA 838 TGGCGACC T GGATCCAG 1268
81 UGUCGUCU CUGAUGAG X CGAA IAUCCAGG 839 CCTGGATC C AGACGACA 1269
82 CUGUCGUC CUGAUGAG X CGAA IGAUCCAG 840 CTGGATCC A GACGACAG 1270
89 UUCUUCCC CUGAUGAG X CGAA IUCGUCUG 841 CAGACGAC A GGGAAGAA 1271
103 GUAGAGGC CUGAUGAG X CGAA ICUCCUUC 842 GAAGGAGC T GCCTCTAC 1272
106 GCCGUAGA CUGAUGAG X CGAA ICAGCUCC 843 GGAGCTGC C TCTACGGC 1273
107 AGCCGUAG CUGAUGAG X CGAA IGCAGCUC 844 GAGCTGCC T CTACGGCT 1274
109 UCAGCCGU CUGAUGAG X CGAA IAGGCAGC 845 GCTGCCTC T ACGGCTGA 1275
115 GCUUCCUC CUGAUGAG X CGAA ICCGUAGA 846 TCTACGGC T GAGGAAGC 1276
124 UUCUUGGC CUGAUGAG X CGAA ICUUCCUC 847 GAGGAAGC A GCCAAGAA 1277
127 UUUUUCUU CUGAUGAG X CGAA ICUGCUUC 848 GAAGCAGC C AAGAAAAA 1278
128 UUUUUUCU CUGAUGAG X CGAA IGCUGCUU 849 AAGCAGCC A AGAAAAAA 1279
158 AGGCCCUU CUGAUGAG X CGAA ICUCUUCU 850 AGAAGAGC A AAGGGCCT 1280
165 CUGCAGAA CUGAUGAG X CGAA ICCCUUUG 851 CAAAGGGC C TTCTGCAG 1281
166 GCUGCAGA CUGAUGAG X CGAA IGCCCUUU 852 AAAGGGCC T TCTGCAGC 1282
169 CCUGCUGC CUGAUGAG X CGAA IAAGGCCC 853 GGGCCTTC T GCAGCAGG 1283
172 UCCCCUGC CUGAUGAG X CGAA ICAGAAGG 854 CCTTCTGC A GCAGGGGA 1284
175 UGUUCCCC CUGAUGAG X CGAA ICUGCAGA 855 TCTGCAGC A GGGGAACA 1285
183 CAGGUUCC CUGAUGAG X CGAA IUUCCCCU 856 AGGGGAAC A GGAACCTG 1286
189 CUUUAUCA CUGAUGAG X CGAA IUUCCUGU 857 ACAGGAAC C TGATAAAG 1287
190 UCUUUAUC CUGAUGAG X CGAA IGUUCCUG 858 CAGGAACC T GATAAAGA 1288
202 GAGGCUCC CUGAUGAG X CGAA IAUUCUUU 859 AAAGAATC A GGAGCCTC 1289
208 UCCACUGA CUGAUGAG X CGAA ICUCCUGA 860 TCAGGAGC C TCAGTGGA 1290
209 AUCCACUG CUGAUGAG X CGAA IGCUCCUG 861 CAGGAGCC T CAGTGGAT 1291
211 UCAUCCAC CUGAUGAG X CGAA IAGGCUCC 862 GGAGCCTC A GTGGATGA 1292
226 AACUGUCU CUGAUGAG X CGAA ICUACUUC 863 GAAGTAGC A AGACAGTT 1293
231 UUUCCAAC CUGAUGAG X CGAA IUCUUGCU 864 AGCAAGAC A GTTGGAAA 1294
244 UCCAAUGC CUGAUGAG X CGAA IAUCUUUC 865 GAAAGATC A GCATTGGA 1295
247 UCUUCCAA CUGAUGAG X CGAA ICUGAUCU 866 AGATCAGC A TTGGAAGA 1296
Table 10
307 UUUCCAGU CUGAUGAG X CGAA ICUCCAUC 867 GATGGAGC A ACTGGAAA 1297
310 UUCUUUCC CUGAUGAG X CGAA IUUGCUCC 868 GGAGCAAC T GGAAAGAA 1298
348 GAACUUUU CUGAUGAG X CGAA IUCCUCUC 869 GAGAGGAC C AAAAGTTC 1299
349 UGAACUUU CUGAUGAG X CGAA IGUCCUCU 870 AGAGGACC A AAAGTTCA 1300
357 GGUCUGUU CUGAUGAG X CGAA IAACUUUU 871 AAAAGTTC A AACAGACC 1301
361 GGAGGGUC CUGAUGAG X CGAA IUUUGAAC 872 GTTCAAAC A GACCCTCC 1302
365 UGAGGGAG CUGAUGAG X CGAA IUCUGUUU 873 AAACAGAC C CTCCCTCA 1303
366 CUGAGGGA CUGAUGAG X CGAA IGUCUGUU 874 AACAGACC C TCCCTCAG 1304
367 ACUGAGGG CUGAUGAG X CGAA IGGUCUGU 875 ACAGACCC T CCCTCAGT 1305
369 GAACUGAG CUGAUGAG X CGAA IAGGGUCU 876 AGACCCTC C CTCAGTTC 1306
370 GGAACUGA CUGAUGAG X CGAA IGAGGGUC 877 GACCCTCC C TCAGTTCC 1307
371 UGGAACUG CUGAUGAG X CGAA IGGAGGGU 878 ACCCTCCC T CAGTTCCA 1308
373 AUUGGAAC CUGAUGAG X CGAA IAGGGAGG 879 CCTCCCTC A GTTCCAAT 1309
378 CACAUAUU CUGAUGAG X CGAA IAACUGAG 880 CTCAGTTC C AATATGTG 1310
379 UCACAUAU CUGAUGAG X CGAA IGAACUGA 881 TCAGTTCC A ATATGTGA 1311
389 AGGAUACA CUGAUGAG X CGAA lUCACAUA 882 TATGTGAC C TGTATCCT 1312
390 UAGGAUAC CUGAUGAG X CGAA IGUCACAU 883 ATGTGACC T GTATCCTA 1313
396 CACCAUUA CUGAUGAG X CGAA IAUACAGG 884 CCTGTATC C TAATGGTG 1314
397 ACACCAUU CUGAUGAG X CGAA IGAUACAG 885 CTGTATCC T AATGGTGT 1315
411 GUCCUUUG CUGAUGAG X CGAA lAAAUACA 886 TGTATTTC C CAAAGGAC 1316
412 UGUCCUUU CUGAUGAG X CGAA IGAAAUAC 887 GTATTTCC C AAAGGACA 1317
413 UUGUCCUU CUGAUGAG X CGAA IGGAAAUA 888 TATTTCCC A AAGGACAA 1318
420 CGCAUUCU CUGAUGAG X CGAA IUCCUUUG 889 CAAAGGAC A AGAATGCG 1319
434 UGUGGGUG CUGAUGAG X CGAA IUAUUCGC 890 GCGAATAC C CACCCACA 1320
435 GUGUGGGU CUGAUGAG X CGAA IGUAUUCG 891 CGAATACC C ACCCACAC 1321
436 UGUGUGGG CUGAUGAG X CGAA IGGUAUUC 892 GAATACCC A CCCACACA 1322
438 CUUGUGUG CUGAUGAG X CGAA IUGGGUAU 893 ATACCCAC C CACACAAG 1323
439 UCUUGUGU CUGAUGAG X CGAA IGUGGGUA 894 TACCCACC C ACACAAGA 1324
440 AUCUUGUG CUGAUGAG X CGAA IGGUGGGU 895 ACCCACCC A CACAAGAT 1325
442 CCAUCUUG CUGAUGAG X CGAA IUGGGUGG 896 CCACCCAC A CAAGATGG 1326
444 GCCCAUCU CUGAUGAG X CGAA IUGUGGGU 897 ACCCACAC A AGATGGGC 1327
457 CAAGCAGC CUGAUGAG X CGAA IUUCGCCC 898 GGGCGAAC A GCTGCTTG 1328
460 CUCCAAGC CUGAUGAG X CGAA ICUGUUCG 899 CGAACAGC T GCTTGGAG 1329
463 GUUCUCCA CUGAUGAG X CGAA ICAGCUGU 900 ACAGCTGC T TGGAGAAC 1330
472 UCACUUGU CUGAUGAG X CGAA lUUCUCCA 901 TGGAGAAC T ACAAGTGA 1331
475 UCUUCACU CUGAUGAG X CGAA IUAGUUCU 902 AGAACTAC A AGTGAAGA 1332
493 UGAUCUAA CUGAUGAG X CGAA ICUUUCUU 903 AAGAAAGC A TTAGATCA 1333
501 CACUUGCC CUGAUGAG X CGAA IAUCUAAU 904 ATTAGATC A GGCAAGTG 1334
505 UCUUCACU CUGAUGAG X CGAA ICCUGAUC 905 GATCAGGC A AGTGAAGA 1335
538 GCUUCUGC CUGAUGAG X CGAA ICUUCUCG 906 CGAGAAGC T GCAGAAGC 1336
541 UGUGCUUC CUGAUGAG X CGAA ICAGCUUC 907 GAAGCTGC A GAAGCACA 1337
547 UGUCGAUG CUGAUGAG X CGAA ICUUCUGC 908 GCAGAAGC A CATCGACA 1338
549 CUUGUCGA CUGAUGAG X CGAA IUGCUUCU 909 AGAAGCAC A TCGACAAG 1339
555 UUCUAACU CUGAUGAG X CGAA IUCGAUGU 910 ACATCGAC A AGTTAGAA 1340
578 CUUGAUCC CUGAUGAG X CGAA ICUCAUUA 911 TAATGAGC T GGATCAAG 1341
584 CCCAGGCU CUGAUGAG X CGAA IAUCCAGC 912 GCTGGATC A AGCCTGGG 1342
588 UCAUCCCA CUGAUGAG X CGAA ICUUGAUC 913 GATCAAGC C TGGGATGA 1343
Table 10
589 GUCAUCCC CUGAUGAG X CGAA IGCUUGAU 914 ATCAAGCC T GGGATGAC 1344
598 UCUAUCAU CUGAUGAG X CGAA IUCAUCCC 915 GGGATGAC A ATGATAGA 1345
611 CUUUUCAC CUGAUGAG X CGAA IAUUUCUA 916 TAGAAATC T GTGAAAAG 1346
629 GCGUGAAC CUGAUGAG X CGAA IUCUUCCA 917 TGGAAGAC T GTTCACGC 1347
634 AACUUGCG CUGAUGAG X CGAA IAACAGUC 918 GACTGTTC A CGCAAGTT 1348
638 UAUUAACU CUGAUGAG X CGAA ICGUGAAC 919 GTTCACGC A AGTTAATA 1349
667 GCCAGGCC CUGAUGAG X CGAA ICAUUUAA 920 TTAAATGC A GGCCTGGC 1350
671 AAAUGCCA CUGAUGAG X CGAA ICCUGCAU 921 ATGCAGGC C TGGCATTT 1351
672 GAAAUGCC CUGAUGAG X CGAA IGCCUGCA 922 TGCAGGCC T GGCATTTC 1352
676 GUAGGAAA CUGAUGAG X CGAA ICCAGGCC 923 GGCCTGGC A TTTCCTAC 1353
681 AUCCAGUA CUGAUGAG X CGAA IAAAUGCC 924 GGCATTTC C TACTGGAT 1354
682 CAUCCAGU CUGAUGAG X CGAA IGAAAUGC 925 GCATTTCC T ACTGGATG 1355
685 GAACAUCC CUGAUGAG X CGAA IUAGGAAA 926 TTTCCTAC T GGATGTTC 1356
694 UUAUUGAG CUGAUGAG X CGAA IAACAUCC 927 GGATGTTC T CTCAATAA 1357
696 AAUUAUUG CUGAUGAG X CGAA IAGAACAU 928 ATGTTCTC T CAATAATT 1358
698 ACAAUUAU CUGAUGAG X CGAA IAGAGAAC 929 GTTCTCTC A ATAATTGT 1359
709 UAAUGGGC CUGAUGAG X CGAA ICACAAUU 930 AATTGTGC T GCCCATTA 1360
712 GUAUAAUG CUGAUGAG X CGAA ICAGCACA 931 TGTGCTGC C CATTATAC 1361
713 AGUAUAAU CUGAUGAG X CGAA IGCAGCAC 932 GTGCTGCC C ATTATACT 1362
714 GAGUAUAA CUGAUGAG X CGAA IGGCAGCA 933 TGCTGCCC A TTATACTC 1363
721 GCAUUGGG CUGAUGAG X CGAA IUAUAAUG 934 CATTATAC T CCCAATGC 1364
723 CGGCAUUG CUGAUGAG X CGAA IAGUAUAA 935 TTATACTC C CAATGCCG 1365
724 CCGGCAUU CUGAUGAG X CGAA IGAGUAUA 936 TATACTCC C AATGCCGG 1366
725 ACCGGCAU CUGAUGAG X CGAA IGGAGUAU 937 ATACTCCC A ATGCCGGT 1367
730 GUGUCACC CUGAUGAG X CGAA ICAUUGGG 938 CCCAATGC C GGTGACAC 1368
737 UACUGUUG CUGAUGAG X CGAA IUCACCGG 939 CCGGTGAC A CAACAGTA 1369
739 AAUACUGU CUGAUGAG X CGAA IUGUCACC 940 GGTGACAC A ACAGTATT 1370
742 UGUAAUAC CUGAUGAG X CGAA IUUGUGUC 941 GACACAAC A GTATTACA 1371
750 CAUCAUAC CUGAUGAG X CGAA IUAAUACU 942 AGTATTAC A GTATGATG 1372
761 UUUACAGA CUGAUGAG X CGAA IUCAUCAU 943 ATGATGAC A TCTGTAAA 1373
764 UAUUUUAC CUGAUGAG X CGAA IAUGUCAU 944 ATGACATC T GTAAAATA 1374
776 UGUUCCAA CUGAUGAG X CGAA IUCUAUUU 945 AAATAGAC T TTGGAACA 1375
784 CUUAUAUG CUGAUGAG X CGAA IUUCCAAA 946 TTTGGAAC A CATATAAG 1376
786 CACUUAUA CUGAUGAG X CGAA IUGUUCCA 947 TGGAACAC A TATAAGTG 1377
809 AAAAGCAC CUGAUGAG X CGAA lUCAAUAA 948 TTATTGAC T GTGCTTTT 1378
814 ACAGUAAA CUGAUGAG X CGAA ICACAGUC 949 GACTGTGC T TTTACTGT 1379
820 AAAGUGAC CUGAUGAG X CGAA lUAAAAGC 950 GCTTTTAC T GTCACTTT 1380
824 AUUAAAAG CUGAUGAG X CGAA IACAGUAA 951 TTACTGTC A CTTTTAAT 1381
• 826 GGAUUAAA CUGAUGAG X CGAA IUGACAGU 952 ACTGTCAC T TTTAATCC 1382
834 CAUAUUUG CUGAUGAG X CGAA IAUUAAAA 953 TTTTAATC C CAAATATG 1383
835 UCAUAUUU CUGAUGAG X CGAA IGAUUAAA 954 TTTAATCC C AAATATGA 1384
836 AUCAUAUU CUGAUGAG X CGAA IGGAUUAA 955 TTAATCCC A AATATGAT 1385
859 UCUUUUAC CUGAUGAG X CGAA ICUUUUAA 956 TTAAAAGC T GTAAAAGA 1386
871 GUGUUAGU CUGAUGAG X CGAA ICAUCUUU 957 AAAGATGC T ACTAACAC 1387
874 CCAGUGUU CUGAUGAG X CGAA IUAGCAUC 958 GATGCTAC T AACACTGG 1388
878 UAUUCCAG CUGAUGAG X CGAA IUUAGUAG 959 CTACTAAC A CTGGAATA 1389
880 UUUAUUCC CUGAUGAG X CGAA IUGUUAGU 960 ACTAACAC T GGAATAAA 1390
Table 10
895 UCAAUUCC CUGAUGAG X CGAA ICACACUU 961 AAGTGTGC T GGAATTGA 1391
912 CAUCACAC CUGAUGAG X CGAA IACGAACA 962 TGTTCGTC T GTGTGATG " 1392
931 UCUUGGAU CUGAUGAG X CGAA ICCUCACC 963 GGTGAGGC c ATCCAAGA 1393
932 UUCUUGGA CUGAUGAG X CGAA IGCCUCAC 964 GTGAGGCC A TCCAAGAA 1394
935 AACUUCUU CUGAUGAG X CGAA IAUGGCCU 965 AGGCCATC C AAGAAGTT 1395
936 UAACUUCU CUGAUGAG X CGAA IGAUGGCC 966 GGCCATCC A AGAAGTTA 1396
952 ACUUCAUA CUGAUGAG X CGAA IACUCCAU 967 ATGGAGTC C TATGAAGT 1397
953 AACUUCAU CUGAUGAG X CGAA IGACUCCA 968 TGGAGTCC T ATGAAGTT 1398
979 ACUUGAUA CUGAUGAG X CGAA IUCUUCCC 969 GGGAAGAC A TATCAAGT 1399
984 GUUUCACU CUGAUGAG X CGAA IAUAUGUC 970 GACATATC A AGTGAAAC 1400
993 UACGGAUU CUGAUGAG X CGAA IUUUCACU 971 AGTGAAAC C AATCCGTA 1401
994 UUACGGAU CUGAUGAG X CGAA IGUUUCAC 972 GTGAAACC A ATCCGTAA 1402
998 UAGAUUAC CUGAUGAG X CGAA IAUUGGUU 973 AACCAATC C GTAATCTA 1403
1005 GUCCAUUU CUGAUGAG X CGAA IAUUACGG 974 CCGTAATC T AAATGGAC 1404
1014 CAAUUGAA CUGAUGAG X CGAA IUCCAUUU 975 AAATGGAC A TTCAATTG 1405
1018 UGCCCAAU CUGAUGAG X CGAA IAAUGUCC 976 GGACATTC A ATTGGGCA 1406
1026 UUCUAUAU CUGAUGAG X CGAA ICCCAAUU 977 AATTGGGC A ATATAGAA 1407
1038 UUCCAGCA CUGAUGAG X CGAA IUAUUCUA 978 TAGAATAC A TGCTGGAA 1408
1042 GUUUUUCC CUGAUGAG X CGAA ICAUGUAU 979 ATACATGC T GGAAAAAC 1409
1051 AUCGGCAC CUGAUGAG X CGAA IUUUUUCC 980 GGAAAAAC A GTGCCGAT 1410
1056 UCACAAUC CUGAUGAG X CGAA ICACUGUU 981 AACAGTGC C GATTGTGA 1411
1078 AUUCUUGU CUGAUGAG X CGAA ICCUCCCC 982 GGGGAGGC A ACAAGAAT 1412
1081 UCCAUUCU CUGAUGAG X CGAA IUUGCCUC 983 GAGGCAAC A AGAATGGA 1413
1108 GUUUCAAU CUGAUGAG X CGAA ICAUAUAC 984 GTATATGC A ATTGAAAC 1414
1117 CUACCAAA CUGAUGAG X CGAA IUUUCAAU 985 ATTGAAAC C TTTGGTAG 1415
1118 ACUACCAA CUGAUGAG X CGAA IGUUUCAA 986 TTGAAACC T TTGGTAGT 1416
1129 CCUUUUCC CUGAUGAG X CGAA IUACUACC 987 GGTAGTAC A GGAAAAGG 1417
1146 UAUCAUCA CUGAUGAG X CGAA lAACAACA 988 TGTTGTTC A TGATGATA 1418
1165 AUGUAAUG CUGAUGAG X CGAA IAACAUUC 989 GAATGTTC A CATTACAT 1419
1167 UCAUGUAA CUGAUGAG X CGAA IUGAACAU 990 ATGTTCAC A TTACATGA 1420
1172 AUUUUUCA CUGAUGAG X CGAA IUAAUGUG 991 CACATTAC A TGAAAAAT 1421
1194 UUGGCACA CUGAUGAG X CGAA IUCCAACA 992 TGTTGGAC A TGTGCCAA 1422
1200 GCCUUAUU CUGAUGAG X CGAA ICACAUGU 993 ACATGTGC C AATAAGGC 1423
1201 AGCCUUAU CUGAUGAG X CGAA IGCACAUG 994 CATGTGCC A ATAAGGCT 1424
1209 UUCUUGGA CUGAUGAG X CGAA ICCUUAUU 995 AATAAGGC T TCCAAGAA 1425
1212 UUGUUCUU CUGAUGAG X CGAA IAAGCCUU 996 AAGGCTTC C AAGAACAA 1426
1213 UUUGUUCU CUGAUGAG X CGAA IGAAGCCU 997 AGGCTTCC A AGAACAAA 1427
1219 AAGUGUUU CUGAUGAG X CGAA IUUCUUGG 998 CCAAGAAC A AAACACTT 1428
1224 UUAACAAG CUGAUGAG X CGAA IUUUUGUU 999 AACAAAAC A CTTGTTAA 1429
1226 AUUUAACA CUGAUGAG X CGAA IUGUUUUG 1000 CAAAACAC T TGTTAAAT 1430
1238 UUCAUUGA CUGAUGAG X CGAA IACAUUUA 1001 TAAATGTC A TCAATGAA 1431
1241 GUUUUCAU CUGAUGAG X CGAA IAUGACAU 1002 ATGTCATC A ATGAAAAC 1432
1250 GGUUCCAA CUGAUGAG X CGAA IUUUUCAU 1003 ATGAAAAC T TTGGAACC 1433
1258 AAGGCAAG CUGAUGAG X CGAA IUUCCAAA 1004 TTTGGAAC C CTTGCCTT 1434
1259 GAAGGCAA CUGAUGAG X CGAA IGUUCCAA 1005 TTGGAACC C TTGCCTTC 1435
1260 AGAAGGCA CUGAUGAG X CGAA IGGUUCCA 1006 TGGAACCC T TGCCTTCT 1436
1264 CGGCAGAA CUGAUGAG X CGAA ICAAGGGU 1007 ACCCTTGC C TTCTGCCG 1437
Table 10
1265 GCGGCAGA CUGAUGAG X CGAA IGCAAGGG 1008 CCCTTGCC T TCTGCCGC 1438
1268 UCUGCGGC CUGAUGAG X CGAA IAAGGCAA 1009 TTGCCTTC T GCCGCAGA 1439
1271 CCAUCUGC CUGAUGAG X CGAA ICAGAAGG 1010 CCTTCTGC C GCAGATGG 1440
1274 CAGCCAUC CUGAUGAG X CGAA ICGGCAGA 1011 TCTGCCGC A GATGGCTG 1441
1281 AGCGAUCC CUGAUGAG X CGAA ICCAUCUG 1012 CAGATGGC T GGATCGCT 1442
1289 UUCUCCCA CUGAUGAG X CGAA ICGAUCCA 1013 TGGATCGC T TGGGAGAA 1443
1307 AGCCAUCA CUGAUGAG X CGAA IUAUUUAC 1014 GTAAATAC T TGATGGCT 1444
1315 UUCUUCAG CUGAUGAG X CGAA ICCAUCAA 1015 TTGATGGC T CTGAAGAA 1445
1317 GAUUCUUC CUGAUGAG X CGAA IAGCCAUC 1016 GATGGCTC T GAAGAATC 1446
1326 AGUCACAC CUGAUGAG X CGAA IAUUCUUC 1017 GAAGAATC T GTGTGACT 1447
1334 AAUGCCCA CUGAUGAG X CGAA IUCACACA 1018 TGTGTGAC T TGGGCATT 1448
1340 AUCUACAA CUGAUGAG X CGAA ICCCAAGU 1019 ACTTGGGC A TTGTAGAT 1449
1350 GUGGAUAU CUGAUGAG X CGAA IAUCUACA 1020 TGTAGATC C ATATCCAC 1450
1351 GGUGGAUA CUGAUGAG X CGAA IGAUCUAC 1021 GTAGATCC A TATCCACC 1451
1356 AUAAUGGU CUGAUGAG X CGAA IAUAUGGA 1022 TCCATATC C ACCATTAT 1452
1357 CAUAAUGG CUGAUGAG X CGAA IGAUAUGG 1023 CCATATCC A CCATTATG 1453
1359 CACAUAAU CUGAUGAG X CGAA IUGGAUAU 1024 ATATCCAC C ATTATGTG 1454
1360 UCACAUAA CUGAUGAG X CGAA IGUGGAUA 1025 TATCCACC A TTATGTGA 1455
1370 UCCUUUAA CUGAUGAG X CGAA lUCACAUA 1026 TATGTGAC A TTAAAGGA 1456
1381 GCUGUAUA CUGAUGAG X CGAA IAUCCUUU 1027 AAAGGATC A TATACAGC 1457
1387 AAUUGCGC CUGAUGAG X CGAA IUAUAUGA 1028 TCATATAC A GCGCAATT 1458
1392 GUUCAAAU CUGAUGAG X CGAA ICGCUGUA 1029 TACAGCGC A ATTTGAAC 1459
1401 GGAUGGUA CUGAUGAG X CGAA IUUCAAAU 1030 ATTTGAAC A TACCATCC 1460
1405 AACAGGAU CUGAUGAG X CGAA IUAUGUUC 1031 GAACATAC C ATCCTGTT 1461
1406 CAACAGGA CUGAUGAG X CGAA IGUAUGUU 1032 AACATACC A TCCTGTTG 1462
1409 ACGCAACA CUGAUGAG X CGAA IAUGGUAU 1033 ATACCATC C TGTTGCGT 1463
1410 GACGCAAC CUGAUGAG X CGAA IGAUGGUA 1034 TACCATCC T GTTGCGTC 1464
1419 UACAUGUU CUGAUGAG X CGAA IACGCAAC 1035 GTTGCGTC C AACATGTA 1465
1420 UUACAUGU CUGAUGAG X CGAA IGACGCAA 1036 TTGCGTCC A ACATGTAA 1466
1423 UCUUUACA CUGAUGAG X CGAA IUUGGACG 1037 CGTCCAAC A TGTAAAGA 1467
1439 UCCUCUGC CUGAUGAG X CGAA IACAACUU 1038 AAGTTGTC A GCAGAGGA 1468
1442 AUCUCCUC CUGAUGAG X CGAA ICUGACAA 1039 TTGTCAGC A GAGGAGAT 1469
1454 AGUUUAAU CUGAUGAG X CGAA IUCAUCUC 1040 GAGATGAC T ATTAAACT 1470
1462 UUGGACUA CUGAUGAG X CGAA IUUUAAUA 1041 TATTAAAC T TAGTCCAA 1471
1468 GUGGCUUU CUGAUGAG X CGAA IACUAAGU 1042 ACTTAGTC C AAAGCCAC 1472
1469 GGUGGCUU CUGAUGAG X CGAA IGACUAAG 1043 CTTAGTCC A AAGCCACC 1473
1474 GUUGAGGU CUGAUGAG X CGAA ICUUUGGA 1044 TCCAAAGC C ACCTCAAC 1474
1475 UGUUGAGG CUGAUGAG X CGAA IGCUUUGG 1045 CCAAAGCC A CCTCAACA 1475
1477 GGUGUUGA CUGAUGAG X CGAA IUGGCUUU 1046 AAAGCCAC C TCAACACC 1476
1478 AGGUGUUG CUGAUGAG X CGAA IGUGGCUU 1047 AAGCCACC T CAACACCT 1477
1480 AAAGGUGU CUGAUGAG X CGAA IAGGUGGC 1048 GCCACCTC A ACACCTTT 1478
1483 AAUAAAGG CUGAUGAG X CGAA IUUGAGGU 1049 ACCTCAAC A CCTTTATT 1479
1485 AAAAUAAA CUGAUGAG X CGAA lUGUUGAG 1050 CTCAACAC C TTTATTTT 1480
1486 GAAAAUAA CUGAUGAG X CGAA IGUGUUGA 1051 TCAACACC T TTATTTTC 1481
1495 CAAAGCUC CUGAUGAG X CGAA IAAAAUAA 1052 TTATTTTC T GAGCTTTG 1482
1500 UCCAACAA CUGAUGAG X CGAA ICUCAGAA 1053 TTCTGAGC T TTGTTGGA 1483
1513 UGGUAUCA CUGAUGAG X CGAA lUUUUCCA 1054 TGGAAAAC A TGATACCA 1484
Table 10
1520 UUAAUUCU CUGAUGAG X CGAA IUAUCAUG 1055 CATGATAC C AGAATTAA 1485
1521 AUUAAUUC CUGAUGAG X CGAA IGUAUCAU 1056 ATGATACC A GAATTAAT 1486
1534 CAACAUGU CUGAUGAG X CGAA ICAAAUUA 1057 TAATTTGC C ACATGTTG 1487
1535 ACAACAUG CUGAUGAG X CGAA IGCAAAUU 1058 AATTTGCC A CATGTTGT 1488
1537 AGACAACA CUGAUGAG X CGAA IUGGCAAA 1059 TTTGCCAC A TGTTGTCT 1489
1545 GUUAAAAC CUGAUGAG X CGAA IACAACAU 1060 ATGTTGTC T GTTTTAAC 1490
1554 GGGUCCAC CUGAUGAG X CGAA IUUAAAAC 1061 GTTTTAAC A GTGGACCC 1491
1561 AUUACAUG CUGAUGAG X CGAA IUCCACUG 1062 CAGTGGAC C CATGTAAT 1492
1562 UAUUACAU CUGAUGAG X CGAA IGUCCACU 1063 AGTGGACC C ATGTAATA 1493
1563 GUAUUACA CUGAUGAG X CGAA IGGUCCAC 1064 GTGGACCC A TGTAATAC 1494
1572 UGGAUAAA CUGAUGAG X CGAA IUAUUACA 1065 TGTAATAC T TTTATCCA 1495
1579 UUAAACAU CUGAUGAG X CGAA IAUAAAAG 1066 CTTTTATC C ATGTTTAA 1496
1580 UUUAAACA CUGAUGAG X CGAA IGAUAAAA 1067 TTTTATCC A TGTTTAAA 1497
1606 UUUGCCUU CUGAUGAG X CGAA IUCCAAAU 1068 ATTTGGAC A AAGGCAAA 1498
1612 AGACGGUU CUGAUGAG X CGAA ICCUUUGU 1069 ACAAAGGC A AACCGTCT 1499
1616 CAUUAGAC CUGAUGAG X CGAA IUUUGCCU 1070 AGGCAAAC C GTCTAATG 1500
1620 AUUACAUU CUGAUGAG X CGAA IACGGUUU 1071 AAACCGTC T AATGTAAT 1501
1633 UUUUCGUU CUGAUGAG X CGAA IUUAAUUA 1072 TAATTAAC C AACGAAAA 1502
1634 UUUUUCGU CUGAUGAG X CGAA IGUUAAUU 1073 AATTAACC A ACGAAAAA 1503
1645 GUCCGGAA CUGAUGAG X CGAA ICUUUUUC 1074 GAAAAAGC T TTCCGGAC 1504
1649 AAAAGUCC CUGAUGAG X CGAA IAAAGCUU 1075 AAGCTTTC C GGACTTTT 1505
1654 CAUUUAAA CUGAUGAG X CGAA IUCCGGAA 1076 TTCCGGAC T TTTAAATG 1506
1664 AAACAGUU CUGAUGAG X CGAA ICAUUUAA 1077 TTAAATGC T AACTGTTT 1507
1668 GGAAAAAC CUGAUGAG X CGAA IUUAGCAU 1078 ATGCTAAC T GTTTTTCC 1508
1676 CAGGAAGG CUGAUGAG X CGAA IAAAAACA 1079 TGTTTTTC c CCTTCCTG 1509
1677 ACAGGAAG CUGAUGAG X CGAA IGAAAAAC 1080 GTTTTTCC c CTTCCTGT 1510
1678 GACAGGAA CUGAUGAG X CGAA IGGAAAAA 1081 TTTTTCCC c TTCCTGTC 1511
1679 AGACAGGA CUGAUGAG X CGAA IGGGAAAA 1082 TTTTCCCC T TCCTGTCT 1512
1682 CCUAGACA CUGAUGAG X CGAA IAAGGGGA 1083 TCCCCTTC c TGTCTAGG 1513
1683 UCCUAGAC CUGAUGAG X CGAA IGAAGGGG 1084 CCCCTTCC T GTCTAGGA 1514
1687 AUUUUCCU CUGAUGAG X CGAA lACAGGAA 1085 TTCCTGTC T AGGAAAAT 1515
1698 AGCUUUAU CUGAUGAG X CGAA ICAUUUUC 1086 GAAAATGC T ATAAAGCT 1516
1706 CUAAUUUG CUGAUGAG X CGAA ICUUUAUA 1087 TATAAAGC T CAAATTAG 1517
1708 AACUAAUU CUGAUGAG X CGAA IAGCUUUA 1088 TAAAGCTC A AATTAGTT 1518
1726 AACGUAUA CUGAUGAG X CGAA IUCAUUCC 1089 GGAATGAC T TATACGTT 1519
1748 AUCUCUUA CUGAUGAG X CGAA IUAUUCAA 1090 TTGAATAC c TAAGAGAT 1520
1749 UAUCUCUU CUGAUGAG X CGAA IGUAUUCA 1091 TGAATACC T AAGAGATA 1521
1759 AUCCAAAA CUGAUGAG X CGAA IUAUCUCU 1092 AGAGATAC T TTTTGGAT 1522
1779 AAGAAUAU CUGAUGAG X CGAA ICAAUAUA 1093 TATATTGC c ATATTCTT 1523
1780 UAAGAAUA CUGAUGAG X CGAA IGCAAUAU 1094 ATATTGCC A TATTCTTA 1524
1786 UUCAAGUA CUGAUGAG X CGAA IAAUAUGG 1095 CCATATTC T TACTTGAA 1525
1790 AGCAUUCA CUGAUGAG X CGAA IUAAGAAU 1096 ATTCTTAC T TGAATGCT 1526
1798 UCAUUCAA CUGAUGAG X CGAA ICAUUCAA 1097 TTGAATGC T TTGAATGA 1527
1808 CUGGAUGU CUGAUGAG X CGAA IUCAUUCA 1098 TGAATGAC T ACATCCAG 1528
1811 GAACUGGA CUGAUGAG X CGAA IUAGUCAU 1099 ATGACTAC A TCCAGTTC 1529
1814 GCAGAACU CUGAUGAG X CGAA IAUGUAGU 1100 ACTACATC C AGTTCTGC 1530
1815 UGCAGAAC CUGAUGAG X CGAA IGAUGUAG 1101 CTACATCC A GTTCTGCA 1531
Table 10
1820 AUAGGUGC CUGAUGAG X CGAA lAACUGGA 1102 TCCAGTTC T GCACCTAT 1532
1823 GGUAUAGG CUGAUGAG X CGAA ICAGAACU 1103 AGTTCTGC A CCTATACC 1533
1825 AGGGUAUA CUGAUGAG X CGAA lUGCAGAA 1104 TTCTGCAC C TATACCCT 1534
1826 GAGGGUAU CUGAUGAG X CGAA IGUGCAGA 1105 TCTGCACC T ATACCCTC 1535
1831 CACCAGAG CUGAUGAG X CGAA IUAUAGGU 1106 ACCTATAC C CTCTGGTG 1536
1832 ACACCAGA CUGAUGAG X CGAA IGUAUAGG 1107 CCTATACC C TCTGGTGT 1537
1833 AACACCAG CUGAUGAG X CGAA IGGUAUAG 1108 CTATACCC T CTGGTGTT 1538
1835 GCAACACC CUGAUGAG X CGAA IAGGGUAU 1109 ATACCCTC T GGTGTTGC 1539
1844 GGUUAAAA CUGAUGAG X CGAA ICAACACC 1110 GGTGTTGC T TTTTAACC 1540
1852 UCCAGGAA CUGAUGAG X CGAA IUUAAAAA llll TTTTTAAC C TTCCTGGA 1541
1853 UUCCAGGA CUGAUGAG X CGAA IGUUAAAA 1112 TTTTAACC T TCCTGGAA 1542
1856 GGAUUCCA CUGAUGAG X CGAA IAAGGUUA 1113 TAACCTTC C TGGAATCC 1543
1857 UGGAUUCC CUGAUGAG X CGAA IGAAGGUU 1114 AACCTTCC T GGAATCCA 1544
1864 UAGAAAAU CUGAUGAG X CGAA IAUUCCAG 1115 CTGGAATC C ATTTTCTA 1545
1865 UUAGAAAA CUGAUGAG X CGAA IGAUUCCA 1116 TGGAATCC A TTTTCTAA 1546
1871 UAUUUUUU CUGAUGAG X CGAA IAAAAUGG 1117 CCATTTTC T AAAAAATA 1547
1885 GAAGAAUG CUGAUGAG X CGAA IUCUUUAU 1118 ATAAAGAC A CATTCTTC 1548
1887 GAGAAGAA CUGAUGAG X CGAA IUGUCUUU 1119 AAAGACAC A TTCTTCTC 1549
1891 UGCUGAGA CUGAUGAG X CGAA IAAUGUGU 1120 ACACATTC T TCTCAGCA 1550
1894 UGGUGCUG CUGAUGAG X CGAA IAAGAAUG 1121 CATTCTTC T CAGCACCA 1551
1896 UGUGGUGC CUGAUGAG X CGAA IAGAAGAA 1122 TTCTTCTC A GCACCACA 1552
1899 UUGUGUGG CUGAUGAG X CGAA ICUGAGAA 1123 TTCTCAGC A CCACACAA 1553
1901 UGUUGUGU CUGAUGAG X CGAA IUGCUGAG 1124 CTCAGCAC C ACACAACA 1554
1902 GUGUUGUG CUGAUGAG X CGAA IGUGCUGA 1125 TCAGCACC A CACAACAC 1555
1904 AGGUGUUG CUGAUGAG X CGAA IUGGUGCU 1126 AGCACCAC A CAACACCT 1556
1906 AUAGGUGU CUGAUGAG X CGAA IUGUGGUG 1127 CACCACAC A ACACCTAT 1557
1909 GGAAUAGG CUGAUGAG X CGAA IUUGUGUG 1128 CACACAAC A CCTATTCC 1558
1911 UUGGAAUA CUGAUGAG X CGAA IUGUUGUG 1129 CACAACAC C TATTCCAA 1559
1912 UUUGGAAU CUGAUGAG X CGAA IGUGUUGU 1130 ACAACACC T ATTCCAAA 1560
1917 UCGAUUUU CUGAUGAG X CGAA IAAUAGGU 1131 ACCTATTC C AAAATCGA 1561
1918 GUCGAUUU CUGAUGAG X CGAA IGAAUAGG 1132 CCTATTCC A AAATCGAC 1562
1927 AAAUAUGU CUGAUGAG X CGAA IUCGAUUU 1133 AAATCGAC C ACATATTT 1563
1928 CAAAUAUG CUGAUGAG X CGAA IGUCGAUU 1134 AATCGACC A CATATTTG 1564
1930 UCCAAAUA CUGAUGAG X CGAA IUGGUCGA 1135 TCGACCAC A TATTTGGA 1565
1947 CUGAGGAG CUGAUGAG X CGAA ICUUUACU 1136 AGTAAAGC T CTCCTCAG 1566
1949 UGCUGAGG CUGAUGAG X CGAA IAGCUUUA 1137 TAAAGCTC T CCTCAGCA 1567
1951 UUUGCUGA CUGAUGAG X CGAA IAGAGCUU 1138 AAGCTCTC C TCAGCAAA 1568
1952 AUUUGCUG CUGAUGAG X CGAA IGAGAGCU 1139 AGCTCTCC T CAGCAAAT 1569
1954 ACAUUUGC CUGAUGAG X CGAA IAGGAGAG 1140 CTCTCCTC A GCAAATGT 1570
1957 UUUACAUU CUGAUGAG X CGAA ICUGAGGA 1141 TCCTCAGC A AATGTAAA 1571
1971 AUAAUUUC CUGAUGAG X CGAA IUUCUUUU 1142 AAAAGAAC A GAAATTAT 1572
1983 AGACAGUU CUGAUGAG X CGAA IUUAUAAU 1143 ATTATAAC A AACTGTCT 1573
1987 UGAGAGAC CUGAUGAG X CGAA IUUUGUUA 1144 TAACAAAC T GTCTCTCA 1574
1991 GGUCUGAG CUGAUGAG X CGAA IACAGUUU 1145 AAACTGTC T CTCAGACC 1575
1993 GUGGUCUG CUGAUGAG X CGAA IAGACAGU 1146 ACTGTCTC T CAGACCAC 1576
1995 CUGUGGUC CUGAUGAG X CGAA IAGAGACA 1147 TGTCTCTC A GACCACAG 1577
1999 UAUACUGU CUGAUGAG X CGAA IUCUGAGA 1148 TCTCAGAC C ACAGTATA 1578
Table 10
2000 UUAUACUG CUGAUGAG X CGAA IGUCUGAG 1149 CTCAGACC A CAGTATAA 1579
2002 GGUUAUAC CUGAUGAG X CGAA IUGGUCUG 1150 CAGACCAC A GTATAACC 1580
2010 UCUAGUUU CUGAUGAG X CGAA lUUAUACU 1151 AGTATAAC C AAACTAGA 1581
2011 UUCUAGUU CUGAUGAG X CGAA IGUUAUAC 1152 GTATAACC A AACTAGAA 1582
2015 UGAGUUCU CUGAUGAG X CGAA lUUUGGUU 1153 AACCAAAC T AGAACTCA 1583
2021 UAAUCCUG CUGAUGAG X CGAA IUUCUAGU 1154 ACTAGAAC T CAGGATTA 1584
2023 CUUAAUCC CUGAUGAG X CGAA IAGUUCUA 1155 TAGAACTC A GGATTAAG 1585
2036 UUUGAGUG CUGAUGAG X CGAA IUUUCUUA 1156 TAAGAAAC T CACTCAAA 1586
2038 GUUUUGAG CUGAUGAG X CGAA IAGUUUCU 1157 AGAAACTC A CTCAAAAC 1587
2040 UGGUUUUG CUGAUGAG X CGAA IUGAGUUU 1158 AAACTCAC T CAAAACCA 1588
2042 UGUGGUUU CUGAUGAG X CGAA IAGUGAGU 1159 ACTCACTC A AAACCACA 1589
2047 AGUUGUGU CUGAUGAG X CGAA IUUUUGAG 1160 CTCAAAAC C ACACAACT 1590
2048 UAGUUGUG CUGAUGAG X CGAA IGUUUUGA 1161 TCAAAACC A CACAACTA 1591
2050 UGUAGUUG CUGAUGAG X CGAA IUGGUUUU 1162 AAAACCAC A CAACTACA 1592
2052 CAUGUAGU CUGAUGAG X CGAA IUGUGGUU 1163 AACCACAC A ACTACATG 1593
2055 UUCCAUGU CUGAUGAG X CGAA IUUGUGUG 1164 CACACAAC T ACATGGAA 1594
2058 AGUUUCCA CUGAUGAG X CGAA lUAGUUGU 1165 ACAACTAC A TGGAAACT 1595
2066 GGUUGUUC CUGAUGAG X CGAA lUUUCCAU 1166 ATGGAAAC T GAACAACC 1596
2071 GAGCAGGU CUGAUGAG X CGAA IUUCAGUU 1167 AACTGAAC A ACCTGCTC 1597
2074 CAGGAGCA CUGAUGAG X CGAA IUUGUUCA 1168 TGAACAAC C TGCTCCTG 1598
2075 UCAGGAGC CUGAUGAG X CGAA IGUUGUUC 1169 GAACAACC T GCTCCTGA 1599
2078 CAUUCAGG CUGAUGAG X CGAA ICAGGUUG 1170 CAACCTGC T CCTGAATG 1600
2080 GUCAUUCA CUGAUGAG X CGAA IAGCAGGU 1171 ACCTGCTC c TGAATGAC 1601
2081 AGUCAUUC CUGAUGAG X CGAA IGAGCAGG 1172 CCTGCTCC T GAATGACT 1602
2089 UAUCCAGU CUGAUGAG X CGAA IUCAUUCA 1173 TGAATGAC T ACTGGATA 1603
2092 AUGUAUCC CUGAUGAG X CGAA IUAGUCAU 1174 ATGACTAC T GGATACAT 1604
2099 UUUUGUUA CUGAUGAG X CGAA IUAUCCAG 1175 CTGGATAC A TAACAAAA 1605
2104 CUUCAUUU CUGAUGAG X CGAA IUUAUGUA 1176 TACATAAC A AAATGAAG 1606
2115 UUUAUUUC CUGAUGAG X CGAA ICCUUCAU 1177 ATGAAGGC A GAAATAAA 1607
2131 GGUUUUAA CUGAUGAG X CGAA IAACAUCU 1178 AGATGTTC T TTAAAACC 1608
2139 UUCUCAUU CUGAUGAG X CGAA IUUUUAAA 1179 TTTAAAAC C AATGAGAA 1609
2140 GUUCUCAU CUGAUGAG X CGAA IGUUUUAA 1180 TTAAAACC A ATGAGAAC 1610
2149 UGUGUCUU CUGAUGAG X CGAA IUUCUCAU 1181 ATGAGAAC A AAGACACA 1611
2155 GUAUGUUG CUGAUGAG X CGAA IUCUUUGU 1182 ACAAAGAC A CAACATAC 1612
2157 UGGUAUGU CUGAUGAG X CGAA IUGUCUUU 1183 AAAGACAC A ACATACCA 1613
2160 UUCUGGUA CUGAUGAG X CGAA IUUGUGUC 1184 GACACAAC A TACCAGAA 1614
2164 GAGAUUCU CUGAUGAG X CGAA IUAUGUUG 1185 CAACATAC C AGAATCTC 1615
2165 AGAGAUUC CUGAUGAG X CGAA IGUAUGUU 1186 AACATACC A GAATCTCT 1616
2171 UGUCCCAG CUGAUGAG X CGAA IAUUCUGG 1187 CCAGAATC T CTGGGACA 1617
2173 UGUGUCCC CUGAUGAG X CGAA IAGAUUCU 1188 AGAATCTC T GGGACACA 1618
2179 UUUGAAUG CUGAUGAG X CGAA IUCCCAGA 1189 TCTGGGAC A CATTCAAA 1619
2181 GCUUUGAA CUGAUGAG X CGAA IUGUCCCA 1190 TGGGACAC A TTCAAAGC 1620
2185 CACUGCUU CUGAUGAG X CGAA IAAUGUGU 1191 ACACATTC A AAGCAGTG 1621
2190 CUACACAC CUGAUGAG X CGAA ICUUUGAA 1192 TTCAAAGC A GTGTGTAG 1622
2214 GCAUUUAG CUGAUGAG X CGAA ICUAUAAA 1193 TTTATAGC A CTAAATGC 1623
2216 GGGCAUUU CUGAUGAG X CGAA IUGCUAUA 1194 TATAGCAC T AAATGCCC 1624
2223 CUCUUGUG CUGAUGAG X CGAA ICAUUUAG 1195 CTAAATGC C CACAAGAG 1625
Table 10
2224 UCUCUUGU CUGAUGAG X CGAA IGCAUUUA 1196 TAAATGCC C ACAAGAGA 1626
2225 UUCUCUUG CUGAUGAG X CGAA IGGCAUUU 1197 AAATGCCC A CAAGAGAA 1627
2227 CUUUCUCU CUGAUGAG X CGAA IUGGGCAU 1198 ATGCCCAC A AGAGAAAG 1628
2237 AUAUUUCC CUGAUGAG X CGAA ICUUUCUC 1199 GAGAAAGC A GGAAATAT 1629
2247 UCAAUUUU CUGAUGAG X CGAA IAUAUUUC 1200 GAAATATC T AAAATTGA 1630
2257 UGUUAGGG CUGAUGAG X CGAA IUCAAUUU 1201 AAATTGAC A CCCTAACA 1631
2259 GAUGUUAG CUGAUGAG X CGAA IUGUCAAU 1202 ATTGACAC C CTAACATC 1632
2260 UGAUGUUA CUGAUGAG X CGAA IGUGUCAA 1203 TTGACACC C TAACATCA 1633
2261 GUGAUGUU CUGAUGAG X CGAA IGGUGUCA 1204 TGACACCC T AACATCAC 1634
2265 AAUUGUGA CUGAUGAG X CGAA IUUAGGGU 1205 ACCCTAAC A TCACAATT 1635
2268 UUUAAUUG CUGAUGAG X CGAA IAUGUUAG 1206 CTAACATC A CAATTAAA 1636
2270 CUUUUAAU CUGAUGAG X CGAA IUGAUGUU 1207 AACATCAC A ATTAAAAG 1637
2282 GCUUCUCU CUGAUGAG X CGAA IUUCUUUU 1208 AAAAGAAC T AGAGAAGC 1638
2291 UUUGCUCU CUGAUGAG X CGAA ICUUCUCU 1209 AGAGAAGC A AGAGCAAA 1639
2297 AAUGUGUU CUGAUGAG X CGAA ICUCUUGC 1210 GCAAGAGC A AACACATT 1640
2301 UUUCAAUG CUGAUGAG X CGAA IUUUGCUC 1211 GAGCAAAC A CATTGAAA 1641
2303 CUUUUCAA CUGAUGAG X CGAA IUGUUUGC 1212 GCAAACAC A TTGAAAAG 1642
2313 CUUCUCUU CUGAUGAG X CGAA ICUUUUCA 1213 TGAAAAGC T AAGAGAAG 1643
2324 UUAUUUCU CUGAUGAG X CGAA ICCUUCUC 1214 GAGAAGGC A AGAAATAA 1644
2334 CUGAUCUU CUGAUGAG X CGAA IUUAUUUC 1215 GAAATAAC T AAGATCAG 1645
2341 UUCUGCUC CUGAUGAG X CGAA IAUCUUAG 1216 CTAAGATC A GAGCAGAA 1646
2346 UUCAGUUC CUGAUGAG X CGAA ICUCUGAU 1217 ATCAGAGC A GAACTGAA 1647
2351 UUUCCUUC CUGAUGAG X CGAA IUUCUGCU 1218 AGCAGAAC T GAAGGAAA 1648
2367 GUUUUUUG CUGAUGAG X CGAA IUCUCUAU 1219 ATAGAGAC A CAAAAAAC 1649
2369 GAGUUUUU CUGAUGAG X CGAA IUGUCUCU 1220 AGAGACAC A AAAAACTC 1650
2376 UUUUGAAG CUGAUGAG X CGAA IUUUUUUG 1221 CAAAAAAC T CTTCAAAA 1651
2378 UUUUUUGA CUGAUGAG X CGAA IAGUUUUU 1222 AAAAACTC T TCAAAAAA 1652
2381 UGAUUUUU CUGAUGAG X CGAA IAAGAGUU 1223 AACTCTTC A AAAAATCA 1653
2389 GGAUUCAU CUGAUGAG X CGAA IAUUUUUU 1224 AAAAAATC A ATGAATCC 1654
2397 CAGCUCCU CUGAUGAG X CGAA IAUUCAUU 1225 AATGAATC C AGGAGCTG 1655
2398 CCAGCUCC CUGAUGAG X CGAA IGAUUCAU 1226 ATGAATCC A GGAGCTGG 1656
2404 AAAAAACC CUGAUGAG X CGAA ICUCCUGG 1227 CCAGGAGC T GGTTTTTT 1657
2422 AAUUUUGU CUGAUGAG X CGAA IAUCGUUU 1228 AAACGATC A ACAAAATT 1658
2425 AUCAAUUU CUGAUGAG X CGAA IUUGAUCG 1229 CGATCAAC A AAATTGAT 1659
2438 CUUGCUAG CUGAUGAG X CGAA IUCUAUCA 1230 TGATAGAC A CTAGCAAG 1660
2440 GUCUUGCU CUGAUGAG X CGAA IUGUCUAU 1231 ATAGACAC T AGCAAGAC 1661
2444 AUUAGUCU CUGAUGAG X CGAA ICUAGUGU 1232 ACACTAGC A AGACTAAT 1662
2449 UCUUUAUU CUGAUGAG X CGAA IUCUUGCU 1233 AGCAAGAC T AATAAAGA 1663
2476 CUUCUAUU CUGAUGAG X CGAA IAUUCUUC 1234 GAAGAATC A AATAGAAG 1664
2486 UUUUUUAU CUGAUGAG X CGAA ICUUCUAU 1235 ATAGAAGC A ATAAAAAA 1665
2511 AUUGGUGG CUGAUGAG X CGAA IAUAUCCC 1236 GGGATATC A CCACCAAT 1666
2513 GGAUUGGU CUGAUGAG X CGAA IUGAUAUC 1237 GATATCAC C ACCAATCC 1667
2514 GGGAUUGG CUGAUGAG X CGAA IGUGAUAU 1238 ATATCACC A CCAATCCC 1668
2516 GUGGGAUU CUGAUGAG X CGAA lUGGUGAU 1239 ATCACCAC C AATCCCAC 1669
2517 UGUGGGAU CUGAUGAG X CGAA IGUGGUGA 1240 TCACCACC A ATCCCACA 1670
2521 UUUCUGUG CUGAUGAG X CGAA IAUUGGUG 1241 CACCAATC C CACAGAAA 1671
2522 AUUUCUGU CUGAUGAG X CGAA IGAUUGGU 1242 ACCAATCC C ACAGAAAT 1672
Table 10
2523 UAUUUCUG CUGAUGAG X CGAA IGGAUUGG 1243 CCAATCCC A CAGAAATA 1S73
2525 UUUAUUUC CUGAUGAG X CGAA IUGGGAUU 1244 AATCCCAC A GAAATAAA 1674
2535 CUGAUGGU CUGAUGAG X CGAA IUUUAUUU 1245 AAATAAAC C ACCATCAG 1675
2536 UCUGAUGG CUGAUGAG X CGAA IGUUUAUU 124S AATAAACC A CCATCAGA 1676
2538 UCUCUGAU CUGAUGAG X CGAA IUGGUUUA 1247 TAAACCAC C ATCAGAGA 1677
2539 UUCUCUGA CUGAUGAG X CGAA IGUGGUUU 1248 AAACCACC A TCAGAGAA 1678
2542 GUAUUCUC CUGAUGAG X CGAA IAUGGUGG 1249 CCACCATC A GAGAATAC 1679
2551 GUGUUUGU CUGAUGAG X CGAA IUAUUCUC 1250 GAGAATAC T ACAAACAC 1680
2554 GAGGUGUU CUGAUGAG X CGAA I AGUAUU 1251 AATACTAC A AACACCTC 1681
2558 CGUAGAGG CUGAUGAG X CGAA IUUUGUAG 1252 CTACAAAC A CCTCTACG 1682
2560 UGCGUAGA CUGAUGAG X CGAA IUGUUUGU 1253 ACAAACAC C TCTACGCA 1683
2561 UUGCGUAG CUGAUGAG X CGAA IGUGUUUG 1254 CAAACACC T CTACGCAA 1684
Input Sequence = HSU29607 Cut Site = CH/
Stem Length = 8 Core Sequence = CUGAUGAG X CGAA (X = GCCGUUAGGC or other stem II)
Seq 1 = HSU2960 (Human methionine aminopeptidase mRNA, complete eds , 2569 bp)
Table 11
Table 11: Human Methionine Aminopeptidase type 2 (MetAP-2) G-cleaver Ribozyme and Target Sequence
Nt. Substrate Sequence Seq. ID Ribozyme Sequence Seq. ID position Nos. Nos.
64 GGAGCCACCU G AAUGG 1685 CCAUU UGAUGGCAUGCACUAUGCGCG AGGUGGCUCC 1834
71 CCUGAAUGGC G ACCUG 1686 CAGGU UGAUGGCAUGCACUAUGCGCG GCCAUUCAGG 1835
86 GGAUCCAGAC G ACAGG 1687 CCUGU UGAUGGCAUGCACUAUGCGCG GUCUGGAUCC 1836
104 AGAAGGAGCU G CCUCU 1688 AGAGG UGAUGGCAUGCACUAUGCGCG AGCUCCUUCU 1837
116 CUCUACGGCU G AGGAA 1689 UUCCU UGAUGGCAUGCACUAUGCGCG AGCCGUAGAG 1838
141 AAAAAAAGAC G AAAGA 1690 UCUUU UGAUGGCAUGCACUAUGCGCG GUCUUUUUUU 1839
170 AGGGCCUUCU G CAGCA 1691 UGCUG UGAUGGCAUGCACUAUGCGCG AGAAGGCCCU 1840
191 ACAGGAACCU G AUAAA 1692 UUUAU UGAUGGCAUGCACUAUGCGCG AGGUUCCUGU 1841
218 CUCAGUGGAU G AAGUA 1693 UACUU UGAUGGCAUGCACUAUGCGCG AUCCACUGAG 1842
269 AGAAAGAGAU G AAGAU 1694 AUCUU UGAUGGCAUGCACUAUGCGCG AUCUCUUUCU 1843
275 AGAUGAAGAU G AUGAA 1695 UUCAU UGAUGGCAUGCACUAUGCGCG AUCUUCAUCU 1844
278 UGAAGAUGAU G AAGAU 1696 AUCUU UGAUGGCAUGCACUAUGCGCG AUCAUCUUCA 1845
293 UGGAGAUGGC G AUGGA 1697 UCCAU UGAUGGCAUGCACUAUGCGCG GCCAUCUCCA 1846
384 GUUCCAAUAU G UGACC 1698 GGUCA UGAUGGCAUGCACUAUGCGCG AUAUUGGAAC 1847
386 UCCAAUAUGU G ACCUG 1699 CAGGU UGAUGGCAUGCACUAUGCGCG ACAUAUUGGA 1848
391 UAUGUGACCU G UAUCC 1700 GGAUA UGAUGGCAUGCACUAUGCGCG AGGUCACAUA . 1849
404 UCCUAAUGGU G UAUUU 1701 AAAUA UGAUGGCAUGCACUAUGCGCG ACCAUUAGGA 1850
426 GGACAAGAAU G CGAAU 1702 AUUCG UGAUGGCAUGCACUAUGCGCG AUUCUUGUCC 1851
428 ACAAGAAUGC G AAUAC 1703 GUAUU UGAUGGCAUGCACUAUGCGCG GCAUUCUUGU 1852
453 CAAGAUGGGC G AACAG 1704 CUGUU UGAUGGCAUGCACUAUGCGCG GCCCAUCUUG 1853
461 GCGAACAGCU G CUUGG 1705 CCAAG UGAUGGCAUGCACUAUGCGCG AGCUGUUCGC 1854
479 AACUACAAGU G AAGAA 1706 UUCUU UGAUGGCAUGCACUAUGCGCG ACUUGUAGUU 1855
509 UCAGGCAAGU G AAGAG 1707 CUCUU UGAUGGCAUGCACUAUGCGCG ACUUGCCUGA 1856
524 GAUUUGGAAU G AUUUU 1708 AAAAU UGAUGGCAUGCACUAUGCGCG AUUCCAAAUC 1857
531 AAUGAUUUUC G AGAAG 1709 CUUCU UGAUGGCAUGCACUAUGCGCG GAAAAUCAUU 1858
539 UCGAGAAGCU G CAGAA 1710 UUCUG UGAUGGCAUGCACUAUGCGCG AGCUUCUCGA 1859
552 GAAGCACAUC G ACAAG 1711 CUUGU UGAUGGCAUGCACUAUGCGCG GAUGUGCUUC 1860
574 AAUACGUAAU G AGCUG 1712 CAGCU UGAUGGCAUGCACUAUGCGCG AUUACGUAUU 1861
Table 11
595 AGCCUGGGAU G ACAAU 1713 AUUGU UGAUGGCAUGCACUAUGCGCG AUCCCAGGCU 1862
601 GGAUGACAAU G AUAGA 1714 UCUAU UGAUGGCAUGCACUAUGCGCG AUUGUCAUCC 1863
612 AUAGAAAUCU G UGAAA 1715 UUUCA UGAUGGCAUGCACUAUGCGCG AGAUUUCUAU 1864
614 AGAAAUCUGU G AAAAG 1716 CUUUU UGAUGGCAUGCACUAUGCGCG ACAGAUUUCU 1865
630 UUGGAAGACU G UUCAC 1717 GUGAA UGAUGGCAUGCACUAUGCGCG AGUCUUCCAA 1866
636 GACUGUUCAC G CAAGU 1718 ACUUG UGAUGGCAUGCACUAUGCGCG GUGAACAGUC 1867
665 UGGAUUAAAU G CAGGC 1719 GCCUG UGAUGGCAUGCACUAUGCGCG AUUUAAUCCA 1868
690 CCUACUGGAU G UUCUC 1720 GAGAA UGAUGGCAUGCACUAUGCGCG AUCCAGUAGG 1869
705 CUCAAUAAUU G UGCUG 1721 CAGCA UGAUGGCAUGCACUAUGCGCG AAUUAUUGAG 1870
707 CAAUAAUUGU G CUGCC 1722 GGCAG UGAUGGCAUGCACUAUGCGCG ACAAUUAUUG 1871
710 UAAUUGUGCU G CCCAU 1723 AUGGG UGAUGGCAUGCACUAUGCGCG AGCACAAUUA 1872
728 UACUCCCAAU G CCGGU 1724 ACCGG UGAUGGCAUGCACUAUGCGCG AUUGGGAGUA 1873
734 CAAUGCCGGU G ACACA 1725 UGUGU UGAUGGCAUGCACUAUGCGCG ACCGGCAUUG 1874
755 AUUACAGUAU G AUGAC 1726 GUCAU UGAUGGCAUGCACUAUGCGCG AUACUGUAAU 1875
758 ACAGUAUGAU G ACAUC 1727 GAUGU UGAUGGCAUGCACUAUGCGCG AUCAUACUGU 1876
765 GAUGACAUCU G UAAAA 1728 UUUUA UGAUGGCAUGCACUAUGCGCG AGAUGUCAUC 1877
806 UAGGAUUAUU G ACUGU 1729 ACAGU UGAUGGCAUGCACUAUGCGCG AAUAAUCCUA 1878
810 AUUAUUGACU G UGCUU 1730 AAGCA UGAUGGCAUGCACUAUGCGCG AGUCAAUAAU 1879
812 UAUUGACUGU G CUUUU 1731 AAAAG UGAUGGCAUGCACUAUGCGCG ACAGUCAAUA 1880
821 UGCUUUUACU G UCACU 1732 AGUGA UGAUGGCAUGCACUAUGCGCG AGUAAAAGCA 1881
842 UCCCAAAUAU G AUACG 1733 CGUAU UGAUGGCAUGCACUAUGCGCG AUAUUUGGGA 1882
860 AUUAAAAGCU G UAAAA 1734 UUUUA UGAUGGCAUGCACUAUGCGCG AGCUUUUAAU 1883
869 UGUAAAAGAU G CUACU 1735 AGUAG UGAUGGCAUGCACUAUGCGCG AUCUUUUACA 1884
891 GGAAUAAAGU G UGCUG 1736 CAGCA UGAUGGCAUGCACUAUGCGCG ACUUUAUUCC 1885
893 AAUAAAGUGU G CUGGA 1737 UCCAG UGAUGGCAUGCACUAUGCGCG ACACUUUAUU 1886
902 UGCUGGAAUU G AUGUU 1738 AACAU UGAUGGCAUGCACUAUGCGCG AAUUCCAGCA 1887
905 UGGAAUUGAU G UUCGU 1739 ACGAA UGAUGGCAUGCACUAUGCGCG AUCAAUUCCA 1888
913 AUGUUCGUCU G UGUGA 1740 UCACA UGAUGGCAUGCACUAUGCGCG AGACGAACAU 1889
915 GUUCGUCUGU G UGAUG 1741 CAUCA UGAUGGCAUGCACUAUGCGCG ACAGACGAAC 1890
917 UCGUCUGUGU G AUGUU 1742 AACAU UGAUGGCAUGCACUAUGCGCG ACACAGACGA 1891
920 UCUGUGUGAU G UUGGU 1743 ACCAA UGAUGGCAUGCACUAUGCGCG AUCACACAGA 1892
Table 11
926 UGAUGUUGGU G AGGCC 1744 GGCCU UGAUGGCAUGCACUAUGCGCG ACCAACAUCA 1893
956 GGAGUCCUAU G AAGUU 1745 AACUU UGAUGGCAUGCACUAUGCGCG AUAGGACUCC 1894
962 CUAUGAAGUU G AAAUA 1746 UAUUU UGAUGGCAUGCACUAUGCGCG AACUUCAUAG 1895
988 CAUAUCAAGU G AAACC 1747 GGUUU UGAUGGCAUGCACUAUGCGCG ACUUGAUAUG 1896
1040 UAGAAUACAU G CUGGA 1748 UCCAG UGAUGGCAUGCACUAUGCGCG AUGUAUUCUA 1897
1054 GAAAAACAGU G CCGAU 1749 AUCGG UGAUGGCAUGCACUAUGCGCG ACUGUUUUUC 1898
1057 AAACAGUGCC G AUUGU 1750 ACAAU UGAUGGCAUGCACUAUGCGCG GGCACUGUUU 1899
1061 AGUGCCGAUU G UGAAA 1751 UUUCA UGAUGGCAUGCACUAUGCGCG AAUCGGCACU 1900
1063 UGCCGAUUGU G AAAGG 1752 CCUUU UGAUGGCAUGCACUAUGCGCG ACAAUCGGCA 1901
1106 AGAAGUAUAU G CAAUU 1753 AAUUG UGAUGGCAUGCACUAUGCGCG AUAUACUUCU 1902
1112 AUAUGCAAUU G AAACC 1754 GGUUU UGAUGGCAUGCACUAUGCGCG AAUUGCAUAU 1903
1139 AGGAAAAGGU G UUGUU 1755 AACAA UGAUGGCAUGCACUAUGCGCG ACCUUUUCCU 1904
1142 AAAAGGUGUU G UUCAU 1756 AUGAA UGAUGGCAUGCACUAUGCGCG AACACCUUUU 1905
1148 UGUUGUUCAU G AUGAU 1757 AUCAU UGAUGGCAUGCACUAUGCGCG AUGAACAACA 1906
1151 UGUUCAUGAU G AUAUG 1758 CAUAU UGAUGGCAUGCACUAUGCGCG AUCAUGAACA 1907
1161 GAUAUGGAAU G UUCAC 1759 GUGAA UGAUGGCAUGCACUAUGCGCG AUUCCAUAUC 1908
1174 CACAUUACAU G AAAAA 1760 UUUUU UGAUGGCAUGCACUAUGCGCG AUGUAAUGUG 1909
1184 GAAAAAUUUU G AUGUU 1761 AACAU UGAUGGCAUGCACUAUGCGCG AAAAUUUUUC 1910
1187 AAAUUUUGAU G UUGGA 1762 UCCAA UGAUGGCAUGCACUAUGCGCG AUCAAAAUUU 1911
1196 UGUUGGACAU G UGCCA 1763 UGGCA UGAUGGCAUGCACUAUGCGCG AUGUCCAACA 1912
1198 UUGGACAUGU G CCAAU 1764 AUUGG UGAUGGCAUGCACUAUGCGCG ACAUGUCCAA 1913
1228 CAAAACACUU G UUAAA 1765 UUUAA UGAUGGCAUGCACUAUGCGCG AAGUGUUUUG 1914
1235 CUUGUUAAAU G UCAUC 1766 GAUGA UGAUGGCAUGCACUAUGCGCG AUUUAACAAG 1915
1244 UGUCAUCAAU G AAAAC 1767 GUUUU UGAUGGCAUGCACUAUGCGCG AUUGAUGACA 1916
1262 UGGAACCCUU σ CCUUC 1768 GAAGG UGAUGGCAUGCACUAUGCGCG AAGGGUUCCA 1917
1269 CUUGCCUUCU G CCGCA 1769 UGCGG UGAUGGCAUGCACUAUGCGCG AGAAGGCAAG 1918
1272 GCCUUCUGCC G CAGAU 1770 AUCUG UGAUGGCAUGCACUAUGCGCG GGCAGAAGGC 1919
1287 UGGCUGGAUC G CUUGG 1771 CCAAG UGAUGGCAUGCACUAUGCGCG GAUCCAGCCA 1920
1309 GUAAAUACUU G AUGGC 1772 GCCAU UGAUGGCAUGCACUAUGCGCG AAGUAUUUAC 1921
1318 UGAUGGCUCU G AAGAA 1773 UUCUU UGAUGGCAUGCACUAUGCGCG AGAGCCAUCA 1922
1327 UGAAGAAUCU G UGUGA 1774 UCACA UGAUGGCAUGCACUAUGCGCG AGAUUCUUCA 1923
Table 11
1329 AAGAAUCUGU G UGACU 1775 AGUCA UGAUGGCAUGCACUAUGCGCG ACAGAUUCUU 1924
1331 GAAUCUGUGU G ACUUG 1776 CAAGU UGAUGGCAUGCACUAUGCGCG ACACAGAUUC 1925
1343 CUUGGGCAUU G UAGAU 1777 AUCUA UGAUGGCAUGCACUAUGCGCG AAUGCCCAAG 1926
1365 CCACCAUUAU G UGACA 1778 UGUCA UGAUGGCAUGCACUAUGCGCG AUAAUGGUGG 1927
1367 ACCAUUAUGU G ACAUU 1779 AAUGU UGAUGGCAUGCACUAUGCGCG ACAUAAUGGU 1928
1390 CAUAUACAGC G CAAUU 1780 AAUUG UGAUGGCAUGCACUAUGCGCG GCUGUAUAUG 1929
1397 AGCGCAAUUU G AACAU 1781 AUGUU UGAUGGCAUGCACUAUGCGCG AAAUUGCGCU 1930
1411 AUACCAUCCU G UUGCG 1782 CGCAA UGAUGGCAUGCACUAUGCGCG AGGAUGGUAU 1931
1414 CCAUCCUGUU G CGUCC 1783 GGACG UGAUGGCAUGCACUAUGCGCG AACAGGAUGG 1932
1425 CGUCCAACAU G UAAAG 1784 CUUUA UGAUGGCAUGCACUAUGCGCG AUGUUGGACG 1933
1436 UAAAGAAGUU G UCAGC 1785 GCUGA UGAUGGCAUGCACUAUGCGCG AACUUCUUUA 1934
1451 CAGAGGAGAU G ACUAU 1786 AUAGU UGAUGGCAUGCACUAUGCGCG AUCUCCUCUG 1935
1496 UUUAUUUUCU G AGCUU 1787 AAGCU UGAUGGCAUGCACUAUGCGCG AGAAAAUAAA 1936
1503 UCUGAGCUUU G UUGGA 1788 UCCAA UGAUGGCAUGCACUAUGCGCG AAAGCUCAGA 1937
1515 UGGAAAACAU G AUACC 1789 GGUAU UGAUGGCAUGCACUAUGCGCG AUGUUUUCCA 1938
1532 GAAUUAAUUU G CCACA 1790 UGUGG UGAUGGCAUGCACUAUGCGCG AAAUUAAUUC 1939
1539 UUUGCCACAU G UUGUC 1791 GACAA UGAUGGCAUGCACUAUGCGCG AUGUGGCAAA 1940
1542 GCCACAUGUU G UCUGU 1792 ACAGA UGAUGGCAUGCACUAUGCGCG AACAUGUGGC 1941
1546 CAUGUUGUCU G UUUUA 1793 UAAAA UGAUGGCAUGCACUAUGCGCG AGACAACAUG 1942
1565 GUGGACCCAU G UAAUA 1794 UAUUA UGAUGGCAUGCACUAUGCGCG AUGGGUCCAC 1943
1582 UUUUAUCCAU G UUUAA 1795 UUAAA UGAUGGCAUGCACUAUGCGCG AUGGAUAAAA 1944
1624 ACCGUCUAAU G UAAUU 1796 AAUUA UGAUGGCAUGCACUAUGCGCG AUUAGACGGU 1945
1637 AUUAACCAAC G AAAAA 1797 UUUUU UGAUGGCAUGCACUAUGCGCG GUUGGUUAAU 1946
1662 ACUUUUAAAU G CUAAC 1798 GUUAG UGAUGGCAUGCACUAUGCGCG AUUUAAAAGU 1947
1669 AAUGCUAACU G UUUUU 1799 AAAAA UGAUGGCAUGCACUAUGCGCG AGUUAGCAUU 1948
1684 UCCCCUUCCU G UCUAG 1800 CUAGA UGAUGGCAUGCACUAUGCGCG AGGAAGGGGA 1949
1696 CUAGGAAAAU G CUAUA 1801 UAUAG UGAUGGCAUGCACUAUGCGCG AUUUUCCUAG 1950
1723 AGUUAGGAAU G ACUUA 1802 UAAGU UGAUGGCAUGCACUAUGCGCG AUUCCUAACU 1951
1737 UAUACGUUUU G UUUUG 1803 CAAAA UGAUGGCAUGCACUAUGCGCG AAAACGUAUA 1952
1742 GUUUUGUUUU G AAUAC 1804 GUAUU UGAUGGCAUGCACUAUGCGCG AAAACAAAAC 1953
1777 UAUUUAUAUU G CCAUA 1805 UAUGG UGAUGGCAUGCACUAUGCGCG AAUAUAAAUA 1954
1792 AUUCUUACUU G AAUGC 1806 GCAUU UGAUGGCAUGCACUAUGCGCG AAGUAAGAAU 1955
1796 UUACUUGAAU G CUUUG 1807 CAAAG UGAUGGCAUGCACUAUGCGCG AUUCAAGUAA 1956
1801 UGAAUGCUUU G AAUGA 1808 UCAUU UGAUGGCAUGCACUAUGCGCG AAAGCAUUCA 1957
1805 UGCUUUGAAU G ACUAC 1809 GUAGU UGAUGGCAUGCACUAUGCGCG AUUCAAAGCA 1958
1821 AUCCAGUUCU G CACCU 1810 AGGUG UGAUGGCAUGCACUAUGCGCG AGAACUGGAU 1959
1839 ACCCUCUGGU G UUGCU 1811 AGCAA UGAUGGCAUGCACUAUGCGCG ACCAGAGGGU 1960
1842 CUCUGGUGUU G CUUUU 1812 AAAAG UGAUGGCAUGCACUAUGCGCG AACACCAGAG 1961
1924 UUCCAAAAUC G ACCAC 1813 GUGGU UGAUGGCAUGCACUAUGCGCG GAUUUUGGAA 1962
1961 CUCAGCAAAU G UAAAA 1814 UUUUA UGAUGGCAUGCACUAUGCGCG AUUUGCUGAG 1963
1988 AUAACAAACU G UCUCU 1815 AGAGA UGAUGGCAUGCACUAUGCGCG AGUUUGUUAU 1964
2067 CAUGGAAACU G AACAA 1816 UUGUU UGAUGGCAUGCACUAUGCGCG AGUUUCCAUG 1965
2076 UGAACAACCU G CUCCU 1817 AGGAG UGAUGGCAUGCACUAUGCGCG AGGUUGUUCA 1966
2082 ACCUGCUCCU G AAUGA 1818 UCAUU UGAUGGCAUGCACUAUGCGCG AGGAGCAGGU 1967
2086 GCUCCUGAAU G ACUAC 1819 GUAGU UGAUGGCAUGCACUAUGCGCG AUUCAGGAGC 1968
2109 AUAACAAAAU G AAGGC 1820 GCCUU UGAUGGCAUGCACUAUGCGCG AUUUUGUUAU 1969
2127 AAAUAAAGAU G UUCUU 1821 AAGAA UGAUGGCAUGCACUAUGCGCG AUCUUUAUUU 1970
2143 UAAAACCAAU G AGAAC 1822 GUUCU UGAUGGCAUGCACUAUGCGCG AUUGGUUUUA 1971
2193 UCAAAGCAGU G UGUAG 1823 CUACA UGAUGGCAUGCACUAUGCGCG ACUGCUUUGA 1972
2195 AAAGCAGUGU G UAGAG 1824 CUCUA UGAUGGCAUGCACUAUGCGCG ACACUGCUUU 1973
2221 AGCACUAAAU G CCCAC 1825 GUGGG UGAUGGCAUGCACUAUGCGCG AUUUAGUGCU 1974
2254 AUCUAAAAUU G ACACC 1826 GGUGU UGAUGGCAUGCACUAUGCGCG AAUUUUAGAU 1975
2306 CAAACACAUU G AAAAG 1827 CUUUU UGAUGGCAUGCACUAUGCGCG AAUGUGUUUG 1976
2352 GAGCAGAACU G AAGGA 1828 UCCUU UGAUGGCAUGCACUAUGCGCG AGUUCUGCUC 1977
2392 AAAAAUCAAU G AAUCC 1829 GGAUU UGAUGGCAUGCACUAUGCGCG AUUGAUUUUU 1978
2413 CUGGUUUUUU G AAACG 1830 CGUUU UGAUGGCAUGCACUAUGCGCG AAAAAACCAG 1979
2418 UUUUUGAAAC G AUCAA 1831 UUGAU UGAUGGCAUGCACUAUGCGCG GUUUCAAAAA 1980
2431 CAACAAAAUU G AUAGA 1832 UCUAU UGAUGGCAUGCACUAUGCGCG AAUUUUGUUG 1981
2496 AAUAAAAAAU G AUAAA 1833 UUUAU UGAUGGCAUGCACUAUGCGCG AUUUUUUAUU 1982
Input Sequence = HSU29607. Cut Site = YG/ or UG/U.
Stem Length 5/10. Core Sequence = UGAUG GCAUGCACUAUGC GCG
Table 11
Table 12
Table 12: Anti Human MetAP-2 HH, NCH, and G-Cleaver Ribozymes
Table 13
Table 13: Human telomerase reverse transcriptase (TERT) Hammerhead Ribozyme and Target Sequence nt. Ribozyme Sequence Seq ID Substrate Sequence Seq ID Position Nos. Nos.
13 CGCAGCAG CUGAUGAG X CGAA ACGCAGCG CGCTGCGT C CTGCTGCG
68 GCAGCGGG CUGAUGAG X CGAA AGCGCGCG CGCGCGCT C CCCGCTGC
90 GCAGCAGG CUGAUGAG X CGAA AGCGCACG CGTGCGCT C CCTGCTGC
108 CCUCGCGG CUGAUGAG X CGAA AGUGGCUG CAGCCACT A CCGCGAGG
135 GCCGCACG CUGAUGAG X CGAA ACGUGGCC GGCCACGT T CGTGCGGC
136 CGCCGCAC CUGAUGAG X CGAA AACGUGGC GCCACGTT C GTGCGGCG
194 CGCGCGGA CUGAUGAG X CGAA AGCCGCCG CGGCGGCT T TCCGCGCG
195 GCGCGCGG CUGAUGAG X CGAA AAGCCGCC GGCGGCTT T CCGCGCGC
196 AGCGCGCG CUGAUGAG X CGAA AAAGCCGC GCGGCTTT C CGCGCGCT
264 GGCGGAAG CUGAUGAG X CGAA AGGGGGCG CGCCCCCT C CTTCCGCC
267 CCUGGCGG CUGAUGAG X CGAA AGGAGGGG CCCCTCCT T CCGCCAGG
268 ACCUGGCG CUGAUGAG X CGAA AAGGAGGG CCCTCCTT C CGCCAGGT
279 UCAGGCAG CUGAUGAG X CGAA ACACCUGG CCAGGTGT C CTGCCTGA
351 CGAAGCCG CUGAUGAG X CGAA AGGCCAGC GCTGGCCT T CGGCTTCG
352 GCGAAGCC CUGAUGAG X CGAA AAGGCCAG CTGGCCTT c GGCTTCGC
357 GCAGCGCG CUGAUGAG X CGAA AGCCGAAG CTTCGGCT T CGCGCTGC
358 AGCAGCGC CUGAUGAG X CGAA AAGCCGAA TTCGGCTT c GCGCTGCT
399 UGGUGGUG CUGAUGAG X CGAA AGGCCUCG CGAGGCCT T CACCACCA
400 CUGGUGGU CUGAUGAG X CGAA AAGGCCUC GAGGCCTT c ACCACCAG
420 UGGGCAGG CUGAUGAG X CGAA AGCUGCGC GCGCAGCT A CCTGCCCA .
505 AGCAGGUG CUGAUGAG X CGAA ACCAGCAC GTGCTGGT T CACCTGCT
506 CAGCAGGU CUGAUGAG X CGAA AACCAGCA TGCTGGTT C ACCTGCTG
529 AGCACAAA CUGAUGAG X CGAA AGCGCGCA TGCGCGCT C TTTGTGCT
531 CCAGCACA CUGAUGAG X CGAA AGAGCGCG CGCGCTCT T TGTGCTGG
532 ACCAGCAC CUGAUGAG X CGAA AAGAGCGC GCGCTCTT τ GTGCTGGT
545 GCAGCUGG CUGAUGAG X CGAA AGCCACCA TGGTGGCT c CCAGCTGC
558 ACACCUGG CUGAUGAG X CGAA AGGCGCAG CTGCGCCT A CCAGGTGT
582 CGAGCUGG CUGAUGAG X CGAA ACAGCGGC GCCGCTGT A CCAGCTCG
589 GCAGCGCC CUGAUGAG X CGAA AGCUGGUA TACCAGCT c GGCGCTGC
602 CCGGGCCU CUGAUGAG X CGAA AGUGGCAG CTGCCACT c AGGCCCGG
626 GGGUCCAC CUGAUGAG X CGAA AGCGUGUG CACACGCT A GTGGACCC
644 GCAUCCCA CUGAUGAG X CGAA ACGCCUUC GAAGGCGT c TGGGATGC
671 CCUGACGC CUGAUGAG X CGAA AUGGUUCC . GGAACCAT A GCGTCAGG
676 GCCUCCCU CUGAUGAG X CGAA ACGCUAUG CATAGCGT C AGGGAGGC
691 CCCAGGGG CUGAUGAG X CGAA ACCCCGGC GCCGGGGT C CCCCTGGG
749 CAACGGCA CUGAUGAG X CGAA ACUUCGGC GCCGAAGT c TGCCGTTG
756 UCUUGGGC CUGAUGAG X CGAA ACGGCAGA TCTGCCGT T GCCCAAGA
808 CCCUGCCC CUGAUGAG X CGAA ACGGGCGU ACGCCCGT T GGGCAGGG
819 GGGCCCAG CUGAUGAG X CGAA ACCCCUGC GCAGGGGT c CTGGGCCC
863 CACACAGA CUGAUGAG X CGAA ACCACGGU ACCGTGGT T TCTGTGTG
864 CCACACAG CUGAUGAG X CGAA AACCACGG CCGTGGTT T CTGTGTGG
865 ACCACACA CUGAUGAG X CGAA AAACCACG CGTGGTTT c TGTGTGGT
876 UGGCAGGU CUGAUGAG X CGAA ACACCACA TGTGGTGT c ACCTGCCA
Table 13
906 CCUCCAAA CUGAUGAG X CGAA AGGUGGCU AGCCACCT C TTTGGAGG
908 ACCCUCCA CUGAUGAG X CGAA AGAGGUGG CCACCTCT T TGGAGGGT
909 CACCCUCC CUGAUGAG X CGAA AAGAGGUG CACCTCTT T GGAGGGTG
922 GUGCCAGA CUGAUGAG X CGAA AGCGCACC GGTGCGCT C TCTGGCAC
924 GCGUGCCA CUGAUGAG X CGAA AGAGCGCA TGCGCTCT C TGGCACGC
939 AUGGGUGG CUGAUGAG X CGAA AGUGGCGC GCGCCACT C CCACCCAT
948 GGCCCACG CUGAUGAG X CGAA AUGGGUGG CCACCCAT C CGTGGGCC
981 GCGAUGUG CUGAUGAG X CGAA AUGGGGGG CCCCCCAT C CACATCGC
987 GUGGCCGC CUGAUGAG X CGAA AUGUGGAU ATCCACAT C GCGGCCAC
1001 GUCCCAGG CUGAUGAG X CGAA ACGUGGUG CACCACGT C CCTGGGAC
1016 CGGGGGAC CUGAUGAG X CGAA AGGCGUGU ACACGCCT T GTCCCCCG
1019 CACCGGGG CUGAUGAG X CGAA ACAAGGCG CGCCTTGT C CCCCGGTG
1029 UCUCGGCG CUGAUGAG X CGAA ACACCGGG CCCGGTGT A CGCCGAGA
1047 AGUAGAGG CUGAUGAG X CGAA AGUGCUUG CAAGCACT T CCTCTACT
1048 GAGUAGAG CUGAUGAG X CGAA AAGUGCUU AAGCACTT C CTCTACTC
1051 GAGGAGUA CUGAUGAG X CGAA AGGAAGUG CACTTCCT C TACTCCTC
1053 CUGAGGAG CUGAUGAG X CGAA AGAGGAAG CTTCCTCT A CTCCTCAG
1056 CGCCUGAG CUGAUGAG X CGAA AGUAGAGG CCTCTACT C CTCAGGCG
1059 UGUCGCCU CUGAUGAG X CGAA AGGAGUAG CTACTCCT C AGGCGACA
1086 GUAGGAAG CUGAUGAG X CGAA AGGGCCGC GCGGCCCT C CTTCCTAC
1089 UGAGUAGG CUGAUGAG X CGAA AGGAGGGC GCCCTCCT T CCTACTCA
1090 CUGAGUAG CUGAUGAG X CGAA AAGGAGGG CCCTCCTT C CTACTCAG
1093 GAGCUGAG CUGAUGAG X CGAA AGGAAGGA TCCTTCCT A CTCAGCTC
1096 AGAGAGCU CUGAUGAG X CGAA AGUAGGAA TTCCTACT C AGCTCTCT
1101 GCCUCAGA CUGAUGAG X CGAA AGCUGAGU ACTCAGCT C TCTGAGGC
1103 GGGCCUCA CUGAUGAG X CGAA AGAGCUGA TCAGCTCT C TGAGGCCC
1127 GAGCCUCC CUGAUGAG X CGAA AGCGCCAG CTGGCGCT C GGAGGCTC
1135 GUCUCCAC CUGAUGAG X CGAA AGCCUCCG CGGAGGCT C GTGGAGAC
1147 CCCAGAAA CUGAUGAG X CGAA AUGGUCUC GAGACCAT C TTTCTGGG
1149 AACCCAGA CUGAUGAG X CGAA AGAUGGUC GACCATCT T TCTGGGTT
1150 GAACCCAG CUGAUGAG X CGAA AAGAUGGU ACCATCTT T CTGGGTTC
1151 GGAACCCA CUGAUGAG X CGAA AAAGAUGG CCATCTTT c TGGGTTCC
1157 GGGCCUGG CUGAUGAG X CGAA ACCCAGAA TTCTGGGT T CCAGGCCC
1158 AGGGCCUG CUGAUGAG X CGAA AACCCAGA TCTGGGTT c CAGGCCCT
1181 CCUGCGGG CUGAUGAG X CGAA AGUCCCUG CAGGGACT c CCCGCAGG
1191 GGCGGGGC CUGAUGAG X CGAA ACCUGCGG CCGCAGGT T GCCCCGCC
1212 UUUGCCAG CUGAUGAG X CGAA AGCGCUGG CCAGCGCT A CTGGCAAA
1233 GCUCCAGA CUGAUGAG X CGAA ACAGGGGC GCCCCTGT T TCTGGAGC
1234 AGCUCCAG CUGAUGAG X CGAA AACAGGGG CCCCTGTT T CTGGAGCT
1235 CAGCUCCA CUGAUGAG X CGAA AAACAGGG CCCTGTTT C TGGAGCTG
1246 UGGUUCCC CUGAUGAG X CGAA AGCAGCUC GAGCTGCT T GGGAACCA
1269 GCACCCCG CUGAUGAG X CGAA AGGGGCAC GTGCCCCT A CGGGGTGC
1279 GUCUUGAG CUGAUGAG X CGAA AGCACCCC GGGGTGCT C CTCAAGAC
1282 UGCGUCUU CUGAUGAG X CGAA AGGAGCAC GTGCTCCT C AAGACGCA
1312 GCUGGGGU CUGAUGAG X CGAA ACCGCAGC GCTGCGGT C ACCCCAGC
1330 CGGGCACA CUGAUGAG X CGAA ACACCGGC GCCGGTGT C TGTGCCCG
1356 CCGCCACA CUGAUGAG X CGAA AGCCCUGG CCAGGGCT C TGTGGCGG
Table 13
1394 CACCAGGC CUGAUGAG X CGAA ACGGGGGU ACCCCCGT c GCCTGGTG
1411 UGCUGGCG CUGAUGAG X CGAA AGCAGCUG CAGCTGCT c CGCCAGCA
1440 CGAAGCCG CUGAUGAG X CGAA ACACCUGC GCAGGTGT A CGGCTTCG
1446 CCCGCACG CUGAUGAG X CGAA AGCCGUAC GTACGGCT T CGTGCGGG
1447 GCCCGCAC CUGAUGAG X CGAA AAGCCGUA TACGGCTT C GTGCGGGC
1486 GAGCCCCA CUGAUGAG X CGAA AGGCCUGG CCAGGCCT C TGGGGCTC
1494 UGUGCCUG CUGAUGAG X CGAA AGCCCCAG CTGGGGCT c CAGGCACA
1515 UCCUGAGG CUGAUGAG X CGAA AGCGGCGU ACGCCGCT T CCTCAGGA
1516 UUCCUGAG CUGAUGAG X CGAA AAGCGGCG CGCCGCTT c CTCAGGAA
1519 GUGUUCCU CUGAUGAG X CGAA AGGAAGCG CGCTTCCT c AGGAACAC
1536 GGGAGAUG CUGAUGAG X CGAA ACUUCUUG CAAGAAGT T CATCTCCC
1537 AGGGAGAU CUGAUGAG X CGAA AACUUCUU AAGAAGTT c ATCTCCCT
1540 CCCAGGGA CUGAUGAG X CGAA AUGAACUU AAGTTCAT c TCCCTGGG
1542 UCCCCAGG CUGAUGAG X CGAA AGAUGAAC GTTCATCT c CCTGGGGA
1564 UGCAGCGA CUGAUGAG X CGAA AGCUUGGC GCCAAGCT c TCGCTGCA
1566 CCUGCAGC CUGAUGAG X CGAA AGAGCUUG CAAGCTCT c GCTGCAGG
1610 GCGCAGCC CUGAUGAG X CGAA AGCGCAGU ACTGCGCT T GGCTGCGC
1633 ACACAGCC CUGAUGAG X CGAA ACCCCUGG CCAGGGGT T GGCTGTGT
1642 GCGGCCGG CUGAUGAG X CGAA ACACAGCC GGCTGTGT T CCGGCCGC
1643 UGCGGCCG CUGAUGAG X CGAA AACACAGC GCTGTGTT c CGGCCGCA
1661 CUCACGCA CUGAUGAG X CGAA ACGGUGCU AGCACCGT c TGCGTGAG
1675 UUGGCCAG CUGAUGAG X CGAA AUCUCCUC GAGGAGAT c CTGGCCAA
1686 AGUGCAGG CUGAUGAG X CGAA ACUUGGCC GGCCAAGT T CCTGCACT
1687 CAGUGCAG CUGAUGAG X CGAA AACUUGGC GCCAAGTT c CTGCACTG
1710 CGACGACG CUGAUGAG X CGAA ACACACUC GAGTGTGT A CGTCGTCG
1714 AGCUCGAC CUGAUGAG X CGAA ACGUACAC GTGTACGT c GTCGAGCT
1717 AGCAGCUC CUGAUGAG X CGAA ACGACGUA TACGTCGT c GAGCTGCT
1726 AAAGACCU CUGAUGAG X CGAA AGCAGCUC GAGCTGCT c AGGTCTTT
1731 AAAAGAAA CUGAUGAG X CGAA ACCUGAGC GCTCAGGT c TTTCTTTT
1733 AUAAAAGA CUGAUGAG X CGAA AGACCUGA TCAGGTCT T TCTTTTAT
1734 CAUAAAAG CUGAUGAG X CGAA AAGACCUG CAGGTCTT T CTTTTATG
1735 ACAUAAAA CUGAUGAG X CGAA AAAGACCU AGGTCTTT c TTTTATGT
1737 UGACAUAA CUGAUGAG X CGAA AGAAAGAC GTCTTTCT T TTATGTCA
1738 GUGACAUA CUGAUGAG X CGAA AAGAAAGA TCTTTCTT T TATGTCAC
1739 CGUGACAU CUGAUGAG X CGAA AAAGAAAG CTTTCTTT T ATGTCACG
1740 CCGUGACA CUGAUGAG X CGAA AAAAGAAA TTTCTTTT A TGTCACGG
1744 GUCUCCGU CUGAUGAG X CGAA ACAUAAAA TTTTATGT c ACGGAGAC
1758 UCUUUUGA CUGAUGAG X CGAA ACGUGGUC GACCACGT T TCAAAAGA
1759 UUCUUUUG CUGAUGAG X CGAA AACGUGGU ACCACGTT T CAAAAGAA
1760 GUUCUUUU CUGAUGAG X CGAA AAACGUGG CCACGTTT c AAAAGAAC
1774 UAGAAAAA CUGAUGAG X CGAA AGCCUGUU AACAGGCT c TTTTTCTA
1776 GGUAGAAA CUGAUGAG X CGAA AGAGCCUG CAGGCTCT T TTTCTACC
1777 CGGUAGAA CUGAUGAG X CGAA AAGAGCCU AGGCTCTT T TTCTACCG
1778 CCGGUAGA CUGAUGAG X CGAA AAAGAGCC GGCTCTTT T TCTACCGG
1779 UCCGGUAG CUGAUGAG X CGAA AAAAGAGC GCTCTTTT T CTACCGGA
1780 UUCCGGUA CUGAUGAG X CGAA AAAAAGAG CTCTTTTT c TACCGGAA
1782 UCUUCCGG CUGAUGAG X CGAA AGAAAAAG CTTTTTCT A CCGGAAGA
Table 13
1795 UUGCUCCA CUGAUGAG X CGAA ACACUCUU AAGAGTGT C TGGAGCAA
1806 UGCUUUGC CUGAUGAG X CGAA ACUUGCUC GAGCAAGT T GCAAAGCA
1816 CUGAUUCC CUGAUGAG X CGAA AUGCUUUG CAAAGCAT T GGAATCAG
1822 UGCUGUCU CUGAUGAG X CGAA AUUCCAAU ATTGGAAT c AGACAGCA
1833 CCCUCUUC CUGAUGAG X CGAA AGUGCUGU ACAGCACT T GAAGAGGG
1860 CUGCUUCC CUGAUGAG X CGAA ACAGCUCC GGAGCTGT c GGAAGCAG
1873 UGCUGCCU CUGAUGAG X CGAA ACCUCUGC GCAGAGGT c AGGCAGCA
1883 GGCUUCCC CUGAUGAG X CGAA AUGCUGCC GGCAGCAT c GGGAAGCC
1911 GGAGUCUG CUGAUGAG X CGAA ACGUCAGC GCTGACGT c CAGACTCC
1918 AUGAAGCG CUGAUGAG X CGAA AGUCUGGA TCCAGACT c CGCTTCAT
1923 UGGGGAUG CUGAUGAG X CGAA AGCGGAGU ACTCCGCT T CATCCCCA
1924 UUGGGGAU CUGAUGAG X CGAA AAGCGGAG CTCCGCTT c ATCCCCAA
1927 GGCUUGGG CUGAUGAG X CGAA AUGAAGCG CGCTTCAT c CCCAAGCC
1954 AUGUUCAC CUGAUGAG X CGAA AUCGGCCG CGGCCGAT T GTGAACAT
1968 CCACGACG CUGAUGAG X CGAA AGUCCAUG CATGGACT A CGTCGTGG
1972 GCUCCCAC CUGAUGAG X CGAA ACGUAGUC GACTACGT c GTGGGAGC
1989 CUCUGCGG CUGAUGAG X CGAA ACGUUCUG CAGAACGT T CCGCAGAG
1990 UCUCUGCG CUGAUGAG X CGAA AACGUUCU AGAACGTT c CGCAGAGA
2015 CGAGGUGA CUGAUGAG X CGAA ACGCUCGG CCGAGCGT c TCACCTCG
2017 CUCGAGGU CUGAUGAG X CGAA AGACGCUC GAGCGTCT c ACCTCGAG
2022 UCACCCUC CUGAUGAG X CGAA AGGUGAGA TCTCACCT c GAGGGTGA
2040 GCACGCUG CUGAUGAG X CGAA ACAGUGCC GGCACTGT T CAGCGTGC
2041 AGCACGCU CUGAUGAG X CGAA AACAGUGC GCACTGTT c AGCGTGCT
2050 UCGUAGUU CUGAUGAG X CGAA AGCACGCU AGCGTGCT c AACTACGA
2055 CCCGCUCG CUGAUGAG X CGAA AGUUGAGC GCTCAACT A CGAGCGGG
2080 GCGCCCAG CUGAUGAG X CGAA AGGCCGGG CCCGGCCT c CTGGGCGC
2091 CCAGCACA CUGAUGAG X CGAA AGGCGCCC GGGCGCCT c TGTGCTGG
2111 CCUGUGGA CUGAUGAG X CGAA AUCGUCCA TGGACGAT A TCCACAGG
2113 GCCCUGUG CUGAUGAG X CGAA AUAUCGUC GACGATAT C CACAGGGC
2133 GCAGCACG CUGAUGAG X CGAA AGGUGCGC GCGCACCT T CGTGCTGC
2134 CGCAGCAC CUGAUGAG X CGAA AAGGUGCG CGCACCTT c GTGCTGCG
2175 UGACAAAG CUGAUGAG X CGAA ACAGCUCA TGAGCTGT A CTTTGTCA
2178 CCUUGACA CUGAUGAG X CGAA AGUACAGC GCTGTACT T TGTCAAGG
2179 ACCUUGAC CUGAUGAG X CGAA AAGUACAG CTGTACTT T GTCAAGGT
2182 UCCACCUU CUGAUGAG X CGAA ACAAAGUA TACTTTGT C AAGGTGGA
2205 UGGUGUCG CUGAUGAG X CGAA ACGCGCCC GGGCGCGT A CGACACCA
2215 UCCUGGGG CUGAUGAG X CGAA AUGGUGUC GACACCAT C CCCCAGGA
2230 ACCUCCGU CUGAUGAG X CGAA AGCCUGUC GACAGGCT c ACGGAGGT
2239 CUGGCGAU CUGAUGAG X CGAA ACCUCCGU ACGGAGGT c ATCGCCAG
2242 AUGCUGGC CUGAUGAG X CGAA AUGACCUC GAGGTCAT c GCCAGCAT
2251 GGUUUGAU CUGAUGAG X CGAA AUGCUGGC GCCAGCAT c ATCAAACC
2254 UGGGGUUU CUGAUGAG X CGAA AUGAUGCU AGCATCAT c AAACCCCA
2271 GCACGCAG CUGAUGAG X CGAA ACGUGUUC GAACACGT A CTGCGTGC
2282 GGCAUACC CUGAUGAG X CGAA ACGCACGC GCGTGCGT C GGTATGCC
2286 CCACGGCA CUGAUGAG X CGAA ACCGACGC GCGTCGGT A TGCCGTGG
2296 GCCUUCUG CUGAUGAG X CGAA ACCACGGC GCCGTGGT C CAGAAGGC
2320 GCCUUGCG CUGAUGAG X CGAA ACGUGCCC GGGCACGT C CGCAAGGC
Table 13
2331 GGCUCUUG CUGAUGAG X CGAA AGGCCUUG CAAGGCCT T CAAGAGCC
2332 UGGCUCUU CUGAUGAG X CGAA AAGGCCUU AAGGCCTT C AAGAGCCA
2344 AAGGUAGA CUGAUGAG X CGAA ACGUGGCU AGCCACGT C TCTACCTT
2346 UCAAGGUA CUGAUGAG X CGAA AGACGUGG CCACGTCT C TACCTTGA
2348 UGUCAAGG CUGAUGAG X CGAA AGAGACGU ACGTCTCT A CCTTGACA
2352 GGUCUGUC CUGAUGAG X CGAA AGGUAGAG CTCTACCT T GACAGACC
2362 UACGGCUG CUGAUGAG X CGAA AGGUCUGU ACAGACCT C CAGCCGTA
2370 GUCGCAUG CUGAUGAG X CGAA ACGGCUGG CCAGCCGT A CATGCGAC
2382 GAGCCACG CUGAUGAG X CGAA ACUGUCGC GCGACAGT T CGTGGCTC
2383 UGAGCCAC CUGAUGAG X CGAA AACUGUCG CGACAGTT C GTGGCTCA
2390 CUGCAGGU CUGAUGAG X CGAA AGCCACGA TCGTGGCT C ACCTGCAG
2425 UCGAUGAC CUGAUGAG X CGAA ACGGCAUC GATGCCGT C GTCATCGA
2428 UGCUCGAU CUGAUGAG X CGAA ACGACGGC GCCGTCGT C ATCGAGCA
2431 CUCUGCUC CUGAUGAG X CGAA AUGACGAC GTCGTCAT C GAGCAGAG
2442 UCAGGGAG CUGAUGAG X CGAA AGCUCUGC GCAGAGCT C CTCCCTGA
2445 CAUUCAGG CUGAUGAG X CGAA AGGAGCUC GAGCTCCT C CCTGAATG
2470 ACGUCGAA CUGAUGAG X CGAA AGGCCACU AGTGGCCT C TTCGACGT
2472 AGACGUCG CUGAUGAG X CGAA AGAGGCCA TGGCCTCT T CGACGTCT
2473 AAGACGUC CUGAUGAG X CGAA AAGAGGCC GGCCTCTT C GACGTCTT
2479 CGUAGGAA CUGAUGAG X CGAA ACGUCGAA TTCGACGT c TTCCTACG
2481 AGCGUAGG CUGAUGAG X CGAA AGACGUCG CGACGTCT T CCTACGCT
2482 AAGCGUAG CUGAUGAG X CGAA AAGACGUC GACGTCTT c CTACGCTT
2485 AUGAAGCG CUGAUGAG X CGAA AGGAAGAC GTCTTCCT A CGCTTCAT
2490 GGCACAUG CUGAUGAG X CGAA AGCGUAGG CCTACGCT T CATGTGCC
2491 UGGCACAU CUGAUGAG X CGAA AAGCGUAG CTACGCTT C ATGTGCCA
2515 UUGCCCCU CUGAUGAG X CGAA AUGCGCAC GTGCGCAT C AGGGGCAA
2526 GGACGUAG CUGAUGAG X CGAA ACUUGCCC GGGCAAGT C CTACGTCC
2529 ACUGGACG CUGAUGAG X CGAA AGGACUUG CAAGTCCT A CGTCCAGT
2533 UGGCACUG CUGAUGAG X CGAA ACGUAGGA TCCTACGT C CAGTGCCA
2548 CCCUGCGG CUGAUGAG X CGAA AUCCCCUG CAGGGGAT C CCGCAGGG
2559 AGAGGAUG CUGAUGAG X CGAA AGCCCUGC GCAGGGCT C CATCCTCT
2563 GUGGAGAG CUGAUGAG X CGAA AUGGAGCC GGCTCCAT C CTCTCCAC
2566 AGCGUGGA CUGAUGAG X CGAA AGGAUGGA TCCATCCT C TCCACGCT
2568 GCAGCGUG CUGAUGAG X CGAA AGAGGAUG CATCCTCT c CACGCTGC
2578 AGGCUGCA CUGAUGAG X CGAA AGCAGCGU ACGCTGCT c TGCAGCCT
2592 UGUCGCCG CUGAUGAG X CGAA AGCACAGG CCTGTGCT A CGGCGACA
2616 UCCCCGCA CUGAUGAG X CGAA AGAGCUUG CAAGCTGT T TGCGGGGA
2617 AUCCCCGC CUGAUGAG X CGAA AACAGCUU AAGCTGTT T GCGGGGAT
2626 UCCCGCCG CUGAUGAG X CGAA AUCCCCGC GCGGGGAT T CGGCGGGA
2627 GUCCCGCC CUGAUGAG X CGAA AAUCCCCG CGGGGATT C GGCGGGAC
2644 AAACGCAG CUGAUGAG X CGAA AGCAGCCC GGGCTGCT C CTGCGTTT
2651 AUCCACCA CUGAUGAG X CGAA ACGCAGGA TCCTGCGT T TGGTGGAT
2652 CAUCCACC CUGAUGAG X CGAA AACGCAGG CCTGCGTT T GGTGGATG
2663 CAACAAGA CUGAUGAG X CGAA AUCAUCCA TGGATGAT T TCTTGTTG
2664 CCAACAAG CUGAUGAG X CGAA AAUCAUCC GGATGATT T CTTGTTGG
2665 ACCAACAA CUGAUGAG X CGAA AAAUCAUC GATGATTT c TTGTTGGT
2667 UCACCAAC CUGAUGAG X CGAA AGAAAUCA TGATTTCT T GTTGGTGA
Table 13
2670 GUGUCACC CUGAUGAG X CGAA ACAAGAAA TTTCTTGT T GGTGACAC
2681 GGUGAGGU CUGAUGAG X CGAA AGGUGUCA TGACACCT C ACCTCACC
2686 GCGUGGGU CUGAUGAG X CGAA AGGUGAGG CCTCACCT C ACCCACGC
2703 UCCUGAGG CUGAUGAG X CGAA AGGUUUUC GAAAACCT T CCTCAGGA
2704 GUCCUGAG CUGAUGAG X CGAA AAGGUUUU AAAACCTT C CTCAGGAC
2707 AGGGUCCU CUGAUGAG X CGAA AGGAAGGU ACCTTCCT C AGGACCCT
2719 ACACCUCG CUGAUGAG X CGAA ACCAGGGU ACCCTGGT C CGAGGTGT
2728 UACUCAGG CUGAUGAG X CGAA ACACCUCG CGAGGTGT c CCTGAGTA
2736 CGCAGCCA CUGAUGAG X CGAA ACUCAGGG CCCTGAGT A TGGCTGCG
2754 UCUUCCGC CUGAUGAG X CGAA AGUUCACC GGTGAACT T GCGGAAGA
2775 CUACAGGG CUGAUGAG X CGAA AGUUCACC GGTGAACT T CCCTGTAG
2776 UCUACAGG CUGAUGAG X CGAA AAGUUCAC GTGAACTT c CCTGTAGA
2782 UCGUCUUC CUGAUGAG X CGAA ACAGGGAA TTCCCTGT A GAAGACGA
2810 CUGAACAA CUGAUGAG X CGAA AGCCGUGC GCACGGCT T TTGTTCAG
2811 UCUGAACA CUGAUGAG X CGAA AAGCCGUG CACGGCTT T TGTTCAGA
2812 AUCUGAAC CUGAUGAG X CGAA AAAGCCGU ACGGCTTT T GTTCAGAT
2815 GGCAUCUG CUGAUGAG X CGAA ACAAAAGC GCTTTTGT T CAGATGCC
2816 CGGCAUCU CUGAUGAG X CGAA AACAAAAG CTTTTGTT c AGATGCCG
2836 CAGGGGAA CUGAUGAG X CGAA AGGCCGUG CACGGCCT A TTCCCCTG
2838 ACCAGGGG CUGAUGAG X CGAA AUAGGCCG CGGCCTAT T CCCCTGGT
2839 CACCAGGG CUGAUGAG X CGAA AAUAGGCC GGCCTATT c CCCTGGTG
2864 GGUCCGGG CUGAUGAG X CGAA AUCCAGCA TGCTGGAT A CCCGGACC
2892 AGCUGGAG CUGAUGAG X CGAA AGUCGCUC GAGCGACT A CTCCAGCT
2895 CAUAGCUG CUGAUGAG X CGAA AGUAGUCG CGACTACT C CAGCTATG
2901 UCCGGGCA CUGAUGAG X CGAA AGCUGGAG CTCCAGCT A TGCCCGGA
2913 CUCUGAUG CUGAUGAG X CGAA AGGUCCGG CCGGACCT C CATCAGAG
2917 CUGGCUCU CUGAUGAG X CGAA AUGGAGGU ACCTCCAT C AGAGCCAG
2927 GAAGGUGA CUGAUGAG X CGAA ACUGGCUC GAGCCAGT C TCACCTTC
2929 UUGAAGGU CUGAUGAG X CGAA AGACUGGC GCCAGTCT C ACCTTCAA
2934 CGCGGUUG CUGAUGAG X CGAA AGGUGAGA TCTCACCT T CAACCGCG
2935 CCGCGGUU CUGAUGAG X CGAA AAGGUGAG CTCACCTT C AACCGCGG
2946 CAGCCUUG CUGAUGAG X CGAA AGCCGCGG CCGCGGCT T CAAGGCTG
2947 CCAGCCUU CUGAUGAG X CGAA AAGCCGCG CGCGGCTT C AAGGCTGG
2969 GAGUUUGC CUGAUGAG X CGAA ACGCAUGU ACATGCGT C GCAAACTC
2977 ACCCCAAA CUGAUGAG X CGAA AGUUUGCG CGCAAACT C TTTGGGGT
2979 AGACCCCA CUGAUGAG X CGAA AGAGUUUG CAAACTCT T TGGGGTCT
2980 AAGACCCC CUGAUGAG X CGAA AAGAGUUU AAACTCTT T GGGGTCTT
2986 AGCCGCAA CUGAUGAG X CGAA ACCCCAAA TTTGGGGT c TTGCGGCT
2988 UCAGCCGC CUGAUGAG X CGAA AGACCCCA TGGGGTCT T GCGGCTGA
3002 CAGGCUGU CUGAUGAG X CGAA ACACUUCA TGAAGTGT c ACAGCCTG
3012 AAUCCAGA CUGAUGAG X CGAA ACAGGCUG CAGCCTGT T TCTGGATT
3013 AAAUCCAG CUGAUGAG X CGAA AACAGGCU AGCCTGTT T CTGGATTT
3014 CAAAUCCA CUGAUGAG X CGAA AAACAGGC GCCTGTTT c TGGATTTG
3020 CACCUGCA CUGAUGAG X CGAA AUCCAGAA TTCTGGAT T TGCAGGTG
3021 UCACCUGC CUGAUGAG X CGAA AAUCCAGA TCTGGATT T GCAGGTGA
3037 ACCGUCUG CUGAUGAG X CGAA AGGCUGUU AACAGCCT c CAGACGGT
3058 AUCUUGUA CUGAUGAG X CGAA AUGUUGGU ACCAACAT c TACAAGAT
Table 13
3060 GGAUCUUG CUGAUGAG X CGAA AGAUGUUG CAACATCT A CAAGATCC
3067 AGCAGGAG CUGAUGAG X CGAA AUCUUGUA TACAAGAT C CTCCTGCT
3070 UGCAGCAG CUGAUGAG X CGAA AGGAUCUU AAGATCCT C CTGCTGCA
3084 GAAACCUG CUGAUGAG X CGAA ACGCCUGC GCAGGCGT A CAGGTTTC
3090 AUGCGUGA CUGAUGAG X CGAA ACCUGUAC GTACAGGT T TCACGCAT
3091 CAUGCGUG CUGAUGAG X CGAA AACCUGUA TACAGGTT T CACGCATG
3092 ACAUGCGU CUGAUGAG X CGAA AAACCUGU ACAGGTTT C ACGCATGT
3112 UGAAAUGG CUGAUGAG X CGAA AGCUGCAG CTGCAGCT C CCATTTCA
3117 GCUGAUGA CUGAUGAG X CGAA AUGGGAGC GCTCCCAT T TCATCAGC
3118 UGCUGAUG CUGAUGAG X CGAA AAUGGGAG CTCCCATT T CATCAGCA
3119 UUGCUGAU CUGAUGAG X CGAA AAAUGGGA TCCCATTT C ATCAGCAA
3122 AACUUGCU CUGAUGAG X CGAA AUGAAAUG CATTTCAT C AGCAAGTT
3130 UUCUUCCA CUGAUGAG X CGAA ACUUGCUG CAGCAAGT T TGGAAGAA
3131 GUUCUUCC CUGAUGAG X CGAA AACUUGCU AGCAAGTT T GGAAGAAC
3147 GCAGGAAA CUGAUGAG X CGAA AUGUGOGG CCCCACAT T TTTCCTGC
3148 CGCAGGAA CUGAUGAG X CGAA AAUGUGGG CCCACATT T TTCCTGCG
3149 GCGCAGGA CUGAUGAG X CGAA AAAUGUGG CCACATTT T TCCTGCGC
3150 CGCGCAGG CUGAUGAG X CGAA AAAAUGUG CACATTTT T CCTGCGCG
3151 ACGCGCAG CUGAUGAG X CGAA AAAAAUGU ACATTTTT c CTGCGCGT
3160 UCAGAGAU CUGAUGAG X CGAA ACGCGCAG CTGCGCGT c ATCTCTGA
3163 GUGUCAGA CUGAUGAG X CGAA AUGACGCG CGCGTCAT c TCTGACAC
3165 CCGUGUCA CUGAUGAG X CGAA AGAUGACG CGTCATCT c TGACACGG
3177 AGCAGAGG CUGAUGAG X CGAA AGGCCGUG CACGGCCT c CCTCTGCT
3181 GAGUAGCA CUGAUGAG X CGAA AGGGAGGC GCCTCCCT c TGCTACTC
3186 GGAUGGAG CUGAUGAG X CGAA AGCAGAGG CCTCTGCT A CTCCATCC
3189 UCAGGAUG CUGAUGAG X CGAA AGUAGCAG CTGCTACT c CATCCTGA
3193 GCUUUCAG CUGAUGAG X CGAA AUGGAGUA TACTCCAT c CTGAAAGC
3219 CCCCCAGC CUGAUGAG X CGAA ACAUCCCU AGGGATGT c GCTGGGGG
3248 GGAGGGCA CUGAUGAG X CGAA AGGGCCGG CCGGCCCT c TGCCCTCC
3255 CGGCCUCG CUGAUGAG X CGAA AGGGCAGA TCTGCCCT c CGAGGCCG
3288 UGAGCAGG CUGAUGAG X CGAA AUGCUUGG CCAAGCAT T CCTGCTCA
3289 UUGAGCAG CUGAUGAG X CGAA AAUGCUUG CAAGCATT c CTGCTCAA
3295 GUCAGCUU CUGAUGAG X CGAA AGCAGGAA TTCCTGCT c AAGCTGAC
3305 ACGGUGUC CUGAUGAG X CGAA AGUCAGCU AGCTGACT c GACACCGT
3316 ACGUAGGU CUGAUGAG X CGAA ACACGGUG CACCGTGT c ACCTACGT
3321 GUGGCACG CUGAUGAG X CGAA AGGUGACA TGTCACCT A CGTGCCAC
3331 GACCCCAG CUGAUGAG X CGAA AGUGGCAC GTGCCACT c CTGGGGTC
3339 UCCUGAGU CUGAUGAG X CGAA ACCCCAGG CCTGGGGT c ACTCAGGA
3343 GCUGUCCU CUGAUGAG X CGAA AGUGACCC GGGTCACT c AGGACAGC
3368 GAGCUUCC CUGAUGAG X CGAA ACUCAGCU AGCTGAGT c GGAAGCTC
3376 GUCCCCGG CUGAUGAG X CGAA AGCUUCCG CGGAAGCT c CCGGGGAC
3429 UGAAGUCU CUGAUGAG X CGAA AGGGCAGU ACTGCCCT c AGACTTCA
3435 UGGUCUUG CUGAUGAG X CGAA AGUCUGAG CTCAGACT T CAAGACCA
3436 AUGGUCUU CUGAUGAG X CGAA AAGUCUGA TCAGACTT c AAGACCAT
3445 CAGUCCAG CUGAUGAG X CGAA AUGGUCUU AAGACCAT c CTGGACTG
3503 CCCGGCGU CUGAUGAG X CGAA ACAGGGCU AGCCCTGT c ACGCCGGG
3514 GGGACGUA CUGAUGAG X CGAA AGCCCGGC GCCGGGCT c TACGTCCC
Table 13
3516 CUGGGACG CUGAUGAG X CGAA AGAGCCCG CGGGCTCT A CGTCCCAG
3520 CUCCCUGG CUGAUGAG X CGAA ACGUAGAG CTCTACGT C CCAGGGAG
3568 AGGCCUCA CUGAUGAG X CGAA ACUCCCAG CTGGGAGT C TGAGGCCT
3587 CUCGGCCA CUGAUGAG X CGAA ACACUCAC GTGAGTGT T TGGCCGAG
3588 CCUCGGCC CUGAUGAG X CGAA AACACUCA TGAGTGTT T GGCCGAGG
3606 UUCAGCCG CUGAUGAG X CGAA ACAUGCAG CTGCATGT C CGGCTGAA
3625 CUCAGCCG CUGAUGAG X CGAA ACACUCAG CTGAGTGT C CGGCTGAG
3648 CUUGGCUG CUGAUGAG X CGAA ACACUCGC GCGAGTGT C CAGCCAAG
3667 GUGUGCUG CUGAUGAG X CGAA ACACUCAG CTGAGTGT C CAGCACAC
3683 GAACUGAA CUGAUGAG X CGAA ACGGCAGG CCTGCCGT C TTCACTTC
3685 GGGAAGUG CUGAUGAG X CGAA AGACGGCA TGCCGTCT T CACTTCCC
3686 GGGGAAGU CUGAUGAG X CGAA AAGACGGC GCCGTCTT C ACTTCCCC
3690 CUGUGGGG CUGAUGAG X CGAA AGUGAAGA TCTTCACT T CCCCACAG
3691 CCUGUGGG CUGAUGAG X CGAA AAGUGAAG CTTCACTT C CCCACAGG
3708 GUGGAGCC CUGAUGAG X CGAA AGCGCCAG CTGGCGCT C GGCTCCAC
3713 CUGGGGUG CUGAUGAG X CGAA AGCCGAGC GCTCGGCT C CACCCCAG
3730 GUGAGGAA CUGAUGAG X CGAA AGCUGGCC GGCCAGCT T TTCCTCAC
3731 GGUGAGGA CUGAUGAG X CGAA AAGCUGGC GCCAGCTT T TCCTCACC
3732 UGGUGAGG CUGAUGAG X CGAA AAAGCUGG CCAGCTTT T CCTCACCA
3733 CUGGUGAG CUGAUGAG X CGAA AAAAGCUG CAGCTTTT C CTCACCAG
3736 CUCCUGGU CUGAUGAG X CGAA AGGAAAAG CTTTTCCT C ACCAGGAG
3752 GGGAGUGG CUGAUGAG X CGAA AGCCGGOC GCCCGGCT T CCACTCCC
3753 GGGGAGUG CUGAUGAG X CGAA AAGCCGGG CCCGGCTT c CACTCCCC
3758 UAUGUGGG CUGAUGAG X CGAA AGUGGAAG CTTCCACT c CCCACATA
3766 ACUAUUCC CUGAUGAG X CGAA AUGUGGGG CCCCACAT A GGAATAGT
3772 GGAUGGAC CUGAUGAG X CGAA AUUCCUAU ATAGGAAT A GTCCATCC
3775 UGGGGAUG CUGAUGAG X CGAA ACUAUUCC GGAATAGT C CATCCCCA
3779 AAUCUGGG CUGAUGAG X CGAA AUGGACUA TAGTCCAT C CCCAGATT
3787 CAAUGGCG CUGAUGAG X CGAA AUCUGGGG CCCCAGAT T CGCCATTG
3788 ACAAUGGC CUGAUGAG X CGAA AAUCUGGG CCCAGATT C GCCATTGT
3794 GGGUGAAC CUGAUGAG X CGAA AUGGCGAA TTCGCCAT T GTTCACCC
3797 GAGGGGUG CUGAUGAG X CGAA ACAAUGGC GCCATTGT T CACCCCTC
3798 CGAGGGGU CUGAUGAG X CGAA AACAAUGG CCATTGTT c ACCCCTCG
3805 GGCAGGGC CUGAUGAG X CGAA AGGGGUGA TCACCCCT c GCCCTGCC
3816 AGGCAAAG CUGAUGAG X CGAA AGGGCAGG CCTGCCCT c CTTTGCCT
3819 GGAAGGCA CUGAUGAG X CGAA AGGAGGGC GCCCTCCT T TGCCTTCC
3820 UGGAAGGC CUGAUGAG X CGAA AAGGAGGG CCCTCCTT T GCCTTCCA
3825 GGGGGUGG CUGAUGAG X CGAA AGGCAAAG CTTTGCCT T CCACCCCC
3826 UGGGGGUG CUGAUGAG X CGAA AAGGCAAA TTTGCCTT c CACCCCCA
3839 UCCACCUG CUGAUGAG X CGAA AUGGUGGG CCCACCAT c CAGGTGGA
3873 AAUUCCCA CUGAUGAG X CGAA AGCUCCCA TGGGAGCT c TGGGAATT
3881 UCACUCCA CUGAUGAG X CGAA AUUCCCAG CTGGGAAT T TGGAGTGA
3882 GUCACUCC CUGAUGAG X CGAA AAUUCCCA TGGGAATT T GGAGTGAC
3907 CGCCUGUG CUGAUGAG X CGAA ACAGGGCA TGCCCTGT A CACAGGCG
3940 CCCACAGG CUGAUGAG X CGAA ACCCCCAU ATGGGGGT c CCTGTGGG
3950 CCCAAUUU CUGAUGAG X CGAA ACCCACAG CTGTGGGT c AAATTGGG
3955 CUCCCCCC CUGAUGAG X CGAA AUUUGACC GGTCAAAT T GGOGGGAG
Table 13
3977 CAGUAUUU CUGAUGAG X CGAA ACUCCCAC GTGGGAGT A AAATACTG
3982 AUAUUCAG CUGAUGAG X CGAA AUUUUACU AGTAAAAT A CTGAATAT
3989 AACUCAUA CUGAUGAG X CGAA AUUCAGUA TACTGAAT A TATGAGTT
3991 AAAACUCA CUGAUGAG X CGAA AUAUUCAG CTGAATAT A TGAGTTTT
3997 AACUGAAA CUGAUGAG X CGAA ACUCAUAU ATATGAGT T TTTCAGTT
3998 AAACUGAA CUGAUGAG X CGAA AACUCAUA TATGAGTT T TTCAGTTT
3999 AAAACUGA CUGAUGAG X CGAA AAACUCAU ATGAGTTT T TCAGTTTT
4000 CAAAACUG CUGAUGAG X CGAA AAAACUCA TGAGTTTT T CAGTTTTG
4001 UCAAAACU CUGAUGAG X CGAA AAAAACUC GAGTTTTT c AGTTTTGA
4005 UUUUUCAA CUGAUGAG X CGAA ACUGAAAA TTTTCAGT T TTGAAAAA
4006 UUUUUUCA CUGAUGAG X CGAA AACUGAAA TTTCAGTT T TGAAAAAA
4007 UUUUUUUC CUGAUGAG X CGAA AAACUGAA TTCAGTTT T GAAAAAAA
Stem Length = 8 . Core Sequence = CUGAUGAG X CGAA (X = GCCGUUAGGC or other stem II sequence and length (greater than or equal to 2 base-pairs))
Seql = TERT (Homo sapiens telomerase reverse transcriptase (TERT) mRNA, 4015 bp); Nakamura et al., Science 277 (5328), 955-959 ( 1997)
Table 14
Table 14: Human telomerase reverse transcriptase (TERT) NCH Ribozyme and Target Sequence
155 CCAGCCCU CUGAUGAG X CGAA IGGCCCCA TGGGGCCC C AGGGCTGG
156 GCCAGCCC CUGAUGAG X CGAA IGGGCCCC GGGGCCCC A GGGCTGGC
161 CAGCCGCC CUGAUGAG X CGAA ICCCUGGG CCCAGGGC T GGCGGCTG
168 GCUGCACC CUGAUGAG X CGAA ICCGCCAG CTGGCGGC T GGTGCAGC
174 CCCCGCGC CUGAUGAG X CGAA ICACCAGC GCTGGTGC A GCGCGGGG
185 AGCCGCCG CUGAUGAG X CGAA IUCCCCGC GCGGGGAC C CGGCGGCT
186 AAGCCGCC CUGAUGAG X CGAA IGUCCCCG CGGGGACC C GGCGGCTT
193 GCGCGGAA CUGAUGAG X CGAA ICCGCCGG CCGGCGGC T TTCCGCGC
197 CAGCGCGC CUGAUGAG X CGAA IAAAGCCG CGGCTTTC C GCGCGCTG
204 GGGCCACC CUGAUGAG X CGAA ICGCGCGG CCGCGCGC T GGTGGCCC
211 AGGCACUG CUGAUGAG X CGAA ICCACCAG CTGGTGGC c CAGTGCCT
212 CAGGCACU CUGAUGAG X CGAA IGCCACCA TGGTGGCC c AGTGCCTG
213 CCAGGCAC CUGAUGAG X CGAA IGGCCACC GGTGGCCC A GTGCCTGG
218 GCACACCA CUGAUGAG X CGAA ICACUGGG CCCAGTGC C TGGTGTGC
219 CGCACACC CUGAUGAG X CGAA IGCACUGG CCAGTGCC T GGTGTGCG
231 CGUCCCAG CUGAUGAG X CGAA ICACGCAC GTGCGTGC C CTGGGACG
232 GCGUCCCA CUGAUGAG X CGAA IGCACGCA TGCGTGCC C TGGGACGC
233 UGCGUCCC CUGAUGAG X CGAA IGGCACGC GCGTGCCC T GGGACGCA
241 GGCGGCCG CUGAUGAG X CGAA ICGUCCCA TGGGACGC A CGGCCGCC
246 CGGGGGGC CUGAUGAG X CGAA ICCGUGCG CGCACGGC C GCCCCCCG
249 CGGCGGGG CUGAUGAG X CGAA ICGGCCGU ACGGCCGC C CCCCGCCG
250 GCGGCGGG CUGAUGAG X CGAA IGCGGCCG CGGCCGCC C CCCGCCGC
251 GGCGGCGG CUGAUGAG X CGAA IGGCGGCC GGCCGCCC C CCGCCGCC
252 GGGCGGCG CUGAUGAG X CGAA IGGGCGGC GCCGCCCC C CGCCGCCC
253 GGGGCGGC CUGAUGAG X CGAA IGGGGCGG CCGCCCCC c GCCGCCCC
256 GAGGGGGC CUGAUGAG X CGAA ICGGGGGG CCCCCCGC c GCCCCCTC
259 AAGGAGGG CUGAUGAG X CGAA ICGGCGGG CCCGCCGC c CCCTCCTT
260 GAAGGAGG CUGAUGAG X CGAA IGCGGCGG CCGCCGCC c CCTCCTTC
261 GGAAGGAG CUGAUGAG X CGAA IGGCGGCG CGCCGCCC c CTCCTTCC
262 CGGAAGGA CUGAUGAG X CGAA IGGGCGGC GCCGCCCC c TCCTTCCG
263 GCGGAAGG CUGAUGAG X CGAA IGGGGCGG CCGCCCCC T CCTTCCGC
265 UGGCGGAA CUGAUGAG X CGAA IAGGGGGC GCCCCCTC c TTCCGCCA
266 CUGGCGGA CUGAUGAG X CGAA IGAGGGGG CCCCCTCC T TCCGCCAG
269 CACCUGGC CUGAUGAG X CGAA IAAGGAGG CCTCCTTC c GCCAGGTG
272 GGACACCU CUGAUGAG X CGAA ICGGAAGG CCTTCCGC c AGGTGTCC
273 AGGACACC CUGAUGAG X CGAA IGCGGAAG CTTCCGCC A GGTGTCCT
280 UUCAGGCA CUGAUGAG X CGAA IACACCUG CAGGTGTC c TGCCTGAA
281 CUUCAGGC CUGAUGAG X CGAA IGACACCU AGGTGTCC T GCCTGAAG
284 CUCCUUCA CUGAUGAG X CGAA ICAGGACA TGTCCTGC c TGAAGGAG
285 GCUCCUUC CUGAUGAG X CGAA IGCAGGAC GTCCTGCC T GAAGGAGC
294 GGGCCACC CUGAUGAG X CGAA ICUCCUUC GAAGGAGC T GGTGGCCC
301 AGCACUCG CUGAUGAG X CGAA ICCACCAG CTGGTGGC c CGAGTGCT
302 CAGCACUC CUGAUGAG X CGAA IGCCACCA TGGTGGCC c GAGTGCTG
309 GCCUCUGC CUGAUGAG X CGAA ICACUCGG CCGAGTGC T GCAGAGGC
312 ACAGCCUC CUGAUGAG X CGAA ICAGCACU AGTGCTGC A GAGGCTGT
318 GCUCGCAC CUGAUGAG X CGAA ICCUCUGC GCAGAGGC T GTGCGAGC
345 CGAAGGCC CUGAUGAG X CGAA ICACGUUC GAACGTGC T GGCCTTCG
Table 14
349 AAGCCGAA CUGAUGAG X CGAA ICCAGCAC GTGCTGGC C TTCGGCTT
350 GAAGCCGA CUGAUGAG X CGAA IGCCAGCA TGCTGGCC T TCGGCTTC
356 CAGCGCGA CUGAUGAG X CGAA ICCGAAGG CCTTCGGC T TCGCGCTG
363 CGUCCAGC CUGAUGAG X CGAA ICGCGAAG CTTCGCGC T GCTGGACG
366 CCCCGUCC CUGAUGAG X CGAA ICAGCGCG CGCGCTGC T GGACGGGG
376 CCCCCGCG CUGAUGAG X CGAA ICCCCGUC GACGGGGC C CGCGGGGG
377 GCCCCCGC CUGAUGAG X CGAA IGCCCCGU ACGGGGCC c GCGGGGGC
386 CUCGGGGG CUGAUGAG X CGAA ICCCCCGC GCGGGGGC c CCCCCGAG
387 CCUCGGGG CUGAUGAG X CGAA IGCCCCCG CGGGGGCC c CCCCGAGG
388 GCCUCGGG CUGAUGAG X CGAA IGGCCCCC GGGGGCCC c CCCGAGGC
389 GGCCUCGG CUGAUGAG X CGAA IGGGCCCC GGGGCCCC c CCGAGGCC
390 AGGCCUCG CUGAUGAG X CGAA IGGGGCCC GGGCCCCC c CGAGGCCT
391 AAGGCCUC CUGAUGAG X CGAA IGGGGGCC GGCCCCCC c GAGGCCTT
397 GUGGUGAA CUGAUGAG X CGAA ICCUCGGG CCCGAGGC c TTCACCAC
398 GGUGGUGA CUGAUGAG X CGAA IGCCUCGG CCGAGGCC T TCACCACC
401 GCUGGUGG CUGAUGAG X CGAA IAAGGCCU AGGCCTTC A CCACCAGC
403 ACGCUGGU CUGAUGAG X CGAA lUGAAGGC GCCTTCAC c ACCAGCGT
404 CACGCUGG CUGAUGAG X CGAA IGUGAAGG CCTTCACC A CCAGCGTG
406 CGCACGCU CUGAUGAG X CGAA IUGGUGAA TTCACCAC C AGCGTGCG
407 GCGCACGC CUGAUGAG X CGAA IGUGGUGA TCACCACC A GCGTGCGC
416 CAGGUAGC CUGAUGAG X CGAA ICGCACGC GCGTGCGC A GCTACCTG
419 GGGCAGGU CUGAUGAG X CGAA ICUGCGCA TGCGCAGC T ACCTGCCC
422 GUUGGGCA CUGAUGAG X CGAA IUAGCUGC GCAGCTAC C TGCCCAAC
423 UGUUGGGC CUGAUGAG X CGAA IGUAGCUG CAGCTACC T GCCCAACA
426 CCGUGUUG CUGAUGAG X CGAA ICAGGUAG CTACCTGC C CAACACGG
427 ACCGUGUU CUGAUGAG X CGAA IGCAGGUA TACCTGCC C AACACGGT
428 CACCGUGU CUGAUGAG X CGAA IGGCAGGU ACCTGCCC A ACACGGTG
431 GGUCACCG CUGAUGAG X CGAA IUUGGGCA TGCCCAAC A CGGTGACC
439 AGUGCGUC CUGAUGAG X CGAA lUCACCGU ACGGTGAC C GACGCACT
445 CCCCGCAG CUGAUGAG X CGAA ICGUCGGU ACCGACGC A CTGCGGGG
447 UCCCCCGC CUGAUGAG X CGAA IUGCGUCG CGACGCAC T GCGGGGGA
471 GCAGCAGC CUGAUGAG X CGAA ICCCCCAC GTGGGGGC T GCTGCTGC
474 GGCGCAGC CUGAUGAG X CGAA ICAGCCCC GGGGCTGC T GCTGCGCC
477 CGCGGCGC CUGAUGAG X CGAA ICAGCAGC GCTGCTGC T GCGCCGCG
482 GCCCACGC CUGAUGAG X CGAA ICGCAGCA TGCTGCGC C GCGTGGGC
501 GGUGAACC CUGAUGAG X CGAA ICACGUCG CGACGTGC T GGTTCACC
507 CCAGCAGG CUGAUGAG X CGAA IAACCAGC GCTGGTTC A CCTGCTGG
509 UGCCAGCA CUGAUGAG X CGAA IUGAACCA TGGTTCAC C TGCTGGCA
510 GUGCCAGC CUGAUGAG X CGAA IGUGAACC GGTTCACC T GCTGGCAC
513 AGCGUGCC CUGAUGAG X CGAA ICAGGUGA TCACCTGC T GGCACGCT
517 GCGCAGCG CUGAUGAG X CGAA ICCAGCAG CTGCTGGC A CGCTGCGC
521 GAGCGCGC CUGAUGAG X CGAA ICGUGCCA TGGCACGC T GCGCGCTC
528 GCACAAAG CUGAUGAG X CGAA ICGCGCAG CTGCGCGC T CTTTGTGC
530 CAGCACAA CUGAUGAG X CGAA IAGCGCGC GCGCGCTC T TTGTGCTG
537 GAGCCACC CUGAUGAG X CGAA ICACAAAG CTTTGTGC T GGTGGCTC
544 CAGCUGGG CUGAUGAG X CGAA ICCACCAG CTGGTGGC T CCCAGCTG
546 CGCAGCUG CUGAUGAG X CGAA IAGCCACC GGTGGCTC C CAGCTGCG
Table 14
547 GCGCAGCU CUGAUGAG X CGAA IGAGCCAC GTGGCTCC C AGCTGCGC
548 GGCGCAGC CUGAUGAG X CGAA IGGAGCCA TGGCTCCC A GCTGCGCC
551 GUAGGCGC CUGAUGAG X CGAA ICUGGGAG CTCCCAGC T GCGCCTAC
556 ACCUGGUA CUGAUGAG X CGAA ICGCAGCU AGCTGCGC c TACCAGGT
557 CACCUGGU CUGAUGAG X CGAA IGCGCAGC GCTGCGCC T ACCAGGTG
560 GCACACCU CUGAUGAG X CGAA IUAGGCGC GCGCCTAC c AGGTGTGC
561 CGCACACC CUGAUGAG X CGAA IGUAGGCG CGCCTACC A GGTGTGCG
573 ACAGCGGC CUGAUGAG X CGAA ICCCGCAC GTGCGGGC C GCCGCTGT
576 GGUACAGC CUGAUGAG X CGAA ICGGCCCG CGGGCCGC C GCTGTACC
579 GCUGGUAC CUGAUGAG X CGAA ICGGCGGC GCCGCCGC T GTACCAGC
584 GCCGAGCU CUGAUGAG X CGAA IUACAGCG CGCTGTAC C AGCTCGGC
585 CGCCGAGC CUGAUGAG X CGAA IGUACAGC GCTGTACC A GCTCGGCG
588 CAGCGCCG CUGAUGAG X CGAA ICUGGUAC GTACCAGC T CGGCGCTG
595 UGAGUGGC CUGAUGAG X CGAA ICGCCGAG CTCGGCGC T GCCACTCA
598 GCCUGAGU CUGAUGAG X CGAA ICAGCGCC GGCGCTGC c ACTCAGGC
599 GGCCUGAG CUGAUGAG X CGAA IGCAGCGC GCGCTGCC A CTCAGGCC
601 CGGGCCUG CUGAUGAG X CGAA IUGGCAGC GCTGCCAC T CAGGCCCG
603 GCCGGGCC CUGAUGAG X CGAA lAGUGGCA TGCCACTC A GGCCCGGC
607 GGGGGCCG CUGAUGAG X CGAA ICCUGAGU ACTCAGGC C CGGCCCCC
608 CGGGGGCC CUGAUGAG X CGAA IGCCUGAG CTCAGGCC C GGCCCCCG
612 GUGGCGGG CUGAUGAG X CGAA ICCGGGCC GGCCCGGC C CCCGCCAC
613 UGUGGCGG CUGAUGAG X CGAA IGCCGGGC GCCCGGCC C CCGCCACA
614 GUGUGGCG CUGAUGAG X CGAA IGGCCGGG CCCGGCCC C CGCCACAC
615 CGUGUGGC CUGAUGAG X CGAA IGGGCCGG CCGGCCCC C GCCACACG
618 UAGCGUGU CUGAUGAG X CGAA ICGGGGGC GCCCCCGC C ACACGCTA
619 CUAGCGUG CUGAUGAG X CGAA IGCGGGGG CCCCCGCC A CACGCTAG
621 CACUAGCG CUGAUGAG X CGAA IUGGCGGG CCCGCCAC A CGCTAGTG
625 GGUCCACU CUGAUGAG X CGAA ICGUGUGG CCACACGC T AGTGGACC
633 GCCUUCGG CUGAUGAG X CGAA IUCCACUA TAGTGGAC C CCGAAGGC
634 CGCCUUCG CUGAUGAG X CGAA IGUCCACU AGTGGACC C CGAAGGCG
635 ACGCCUUC CUGAUGAG X CGAA IGGUCCAC GTGGACCC C GAAGGCGT
645 CGCAUCCC CUGAUGAG X CGAA IACGCCUU AAGGCGTC T GGGATGCG
661 UGGUUCCA CUGAUGAG X CGAA ICCCGUUC GAACGGGC c TGGAACCA
662 AUGGUUCC CUGAUGAG X CGAA IGCCCGUU AACGGGCC T GGAACCAT
668 GACGCUAU CUGAUGAG X CGAA IUUCCAGG CCTGGAAC c ATAGCGTC
669 UGACGCUA CUGAUGAG X CGAA IGUUCCAG CTGGAACC A TAGCGTCA
677 GGCCUCCC CUGAUGAG X CGAA IACGCUAU ATAGCGTC A GGGAGGCC
685 GGGACCCC CUGAUGAG X CGAA ICCUCCCU AGGGAGGC C GGGGTCCC
692 GCCCAGGG CUGAUGAG X CGAA IACCCCGG CCGGGGTC C CCCTGGGC
693 GGCCCAGG CUGAUGAG X CGAA IGACCCCG CGGGGTCC C CCTGGGCC
694 AGGCCCAG CUGAUGAG X CGAA IGGACCCC GGGGTCCC C CTGGGCCT
695 CAGGCCCA CUGAUGAG X CGAA IGGGACCC GGGTCCCC C TGGGCCTG
696 GCAGGCCC CUGAUGAG X CGAA IGGGGACC GGTCCCCC T GGGCCTGC
701 GGCUGGCA CUGAUGAG X CGAA ICCCAGGG CCCTGGGC C TGCCAGCC
702 GGGCUGGC CUGAUGAG X CGAA IGCCCAGG CCTGGGCC T GCCAGCCC
705 CCGGGGCU CUGAUGAG X CGAA ICAGGCCC GGGCCTGC C AGCCCCGG
706 CCCGGGGC CUGAUGAG X CGAA IGCAGGCC GGCCTGCC A GCCCCGGG
Table 14
709 GCACCCGG CUGAUGAG X CGAA ICUGGCAG CTGCCAGC C CCGGGTGC
710 CGCACCCG CUGAUGAG X CGAA IGCUGGCA TGCCAGCC C CGGGTGCG
711 UCGCACCC CUGAUGAG X CGAA IGGCUGGC GCCAGCCC C GGGTGCGA
734 GCUGGCAC CUGAUGAG X CGAA ICCCCCGC GCGGGGGC A GTGCCAGC
739 CUUCGGCU CUGAUGAG X CGAA ICACUGCC GGCAGTGC C AGCCGAAG
740 ACUUCGGC CUGAUGAG X CGAA IGCACUGC GCAGTGCC A GCCGAAGT
743 CAGACUUC CUGAUGAG X CGAA ICUGGCAC GTGCCAGC C GAAGTCTG
750 GCAACGGC CUGAUGAG X CGAA IACUUCGG CCGAAGTC T GCCGTTGC
753 UGGGCAAC CUGAUGAG X CGAA ICAGACUU AAGTCTGC C GTTGCCCA
759 GCCUCUUG CUGAUGAG X CGAA ICAACGGC GCCGTTGC C CAAGAGGC
760 GGCCUCUU CUGAUGAG X CGAA IGCAACGG CCGTTGCC C AAGAGGCC
761 GGGCCUCU CUGAUGAG X CGAA IGGCAACG CGTTGCCC A AGAGGCCC
768 CACGCCUG CUGAUGAG X CGAA ICCUCUUG CAAGAGGC C CAGGCGTG
769 CCACGCCU CUGAUGAG X CGAA iσccucuu AAGAGGCC C AGGCGTGG
770 GCCACGCC CUGAUGAG X CGAA IGGCCUCU AGAGGCCC A GGCGTGGC
781 UCAGGGGC CUGAUGAG X CGAA ICGCCACG CGTGGCGC T GCCCCTGA
784 GGCUCAGG CUGAUGAG X CGAA ICAGCGCC GGCGCTGC c CCTGAGCC
785 CGGCUCAG CUGAUGAG X CGAA IGCAGCGC GCGCTGCC c CTGAGCCG
786 CCGGCUCA CUGAUGAG X CGAA IGGCAGCG CGCTGCCC c TGAGCCGG
787 UCCGGCUC CUGAUGAG X CGAA IGGGCAGC GCTGCCCC T GAGCCGGA
792 UCCGCUCC CUGAUGAG X CGAA ICUCAGGG CCCTGAGC c GGAGCGGA
804 GCCCAACG CUGAUGAG X CGAA ICGUCCGC GCGGACGC c CGTTGGGC
805 UGCCCAAC CUGAUGAG X CGAA IGCGUCCG CGGACGCC c GTTGGGCA
813 AGGACCCC CUGAUGAG X CGAA ICCCAACG CGTTGGGC A GGGGTCCT
820 UGGGCCCA CUGAUGAG X CGAA IACCCCUG CAGGGGTC C TGGGCCCA
821 GUGGGCCC CUGAUGAG X CGAA IGACCCCU AGGGGTCC T GGGCCCAC
826 CCCGGGUG CUGAUGAG X CGAA ICCCAGGA TCCTGGGC c CACCCGGG
827 GCCCGGGU CUGAUGAG X CGAA IGCCCAGG CCTGGGCC c ACCCGGGC
828 UGCCCGGG CUGAUGAG X CGAA IGGCCCAG CTGGGCCC A CCCGGGCA
830 CCUGCCCG CUGAUGAG X CGAA IUGGGCCC GGGCCCAC C CGGGCAGG
831 UCCUGCCC CUGAUGAG X CGAA IGUGGGCC GGCCCACC C GGGCAGGA
836 ACGCGUCC CUGAUGAG X CGAA ICCCGGGU ACCCGGGC A GGACGCGT
849 GGUCACUC CUGAUGAG X CGAA IUCCACGC GCGTGGAC C GAGTGACC
857 GAAACCAC CUGAUGAG X CGAA IUCACUCG CGAGTGAC C GTGGTTTC
866 CACCACAC CUGAUGAG X CGAA IAAACCAC GTGGTTTC T GTGTGGTG
877 CUGGCAGG CUGAUGAG X CGAA IACACCAC GTGGTGTC A CCTGCCAG
879 GUCUGGCA CUGAUGAG X CGAA IUGACACC GGTGTCAC C TGCCAGAC
880 GGUCUGGC CUGAUGAG X CGAA IGUGACAC GTGTCACC T GCCAGACC
883 GCGGGUCU CUGAUGAG X CGAA ICAGGUGA TCACCTGC C AGACCCGC
884 GGCGGGUC CUGAUGAG X CGAA IGCAGGUG CACCTGCC A GACCCGCC
888 CUUCGGCG CUGAUGAG X CGAA IUCUGGCA TGCCAGAC C CGCCGAAG
889 UCUUCGGC CUGAUGAG X CGAA IGUCUGGC GCCAGACC C GCCGAAGA
892 GCUUCUUC CUGAUGAG X CGAA ICGGGUCU AGACCCGC C GAAGAAGC
901 AAAGAGGU CUGAUGAG X CGAA ICUUCUUC GAAGAAGC C ACCTCTTT
902 CAAAGAGG CUGAUGAG X CGAA IGCUUCUU AAGAAGCC A CCTCTTTG
904 UCCAAAGA CUGAUGAG X CGAA IUGGCUUC GAAGCCAC C TCTTTGGA
905 CUCCAAAG CUGAUGAG X CGAA IGUGGCUU AAGCCACC T CTTTGGAG
Table 14
907 CCCUCCAA CUGAUGAG X CGAA IAGGUGGC GCCACCTC T TTGGAGGG
921 UGCCAGAG CUGAUGAG X CGAA ICGCACCC GGGTGCGC T CTCTGGCA
923 CGUGCCAG CUGAUGAG X CGAA IAGCGCAC GTGCGCTC T CTGGCACG
925 CGCGUGCC CUGAUGAG X CGAA IAGAGCGC GCGCTCTC T GGCACGCG
929 GUGGCGCG CUGAUGAG X CGAA ICCAGAGA TCTCTGGC A CGCGCCAC
935 GUGGGAGU CUGAUGAG X CGAA ICGCGUGC GCACGCGC C ACTCCCAC
936 GGUGGGAG CUGAUGAG X CGAA IGCGCGUG CACGCGCC A CTCCCACC
938 UGGGUGGG CUGAUGAG X CGAA IUGGCGCG CGCGCCAC T CCCACCCA
940 GAUGGGUG CUGAUGAG X CGAA IAGUGGCG CGCCACTC c CACCCATC
941 GGAUGGGU CUGAUGAG X CGAA IGAGUGGC GCCACTCC c ACCCATCC
942 CGGAUGGG CUGAUGAG X CGAA IGGAGUGG CCACTCCC A CCCATCCG
944 CACGGAUG CUGAUGAG X CGAA IUGGGAGU ACTCCCAC C CATCCGTG
945 CCACGGAU CUGAUGAG X CGAA IGUGGGAG CTCCCACC C ATCCGTGG
946 CCCACGGA CUGAUGAG X CGAA IGGUGGGA TCCCACCC A TCCGTGGG
949 CGGCCCAC CUGAUGAG X CGAA IAUGGGUG CACCCATC C GTGGGCCG
956 GUGCUGGC CUGAUGAG X CGAA ICCCACGG CCGTGGGC C GCCAGCAC
959 GUGGUGCU CUGAUGAG X CGAA ICGGCCCA TGGGCCGC C AGCACCAC
960 CGUGGUGC CUGAUGAG X CGAA IGCGGCCC GGGCCGCC A GCACCACG
963 CCGCGUGG CUGAUGAG X CGAA ICUGGCGG CCGCCAGC A CCACGCGG
965 GCCCGCGU CUGAUGAG X CGAA IUGCUGGC GCCAGCAC C ACGCGGGC
966 GGCCCGCG CUGAUGAG X CGAA IGUGCUGG CCAGCACC A CGCGGGCC
974 GGAUGGGG CUGAUGAG X CGAA ICCCGCGU ACGCGGGC C CCCCATCC
975 UGGAUGGG CUGAUGAG X CGAA IGCCCGCG CGCGGGCC C CCCATCCA
976 GUGGAUGG CUGAUGAG X CGAA IGGCCCGC GCGGGCCC C CCATCCAC
977 UGUGGAUG CUGAUGAG X CGAA IGGGCCCG CGGGCCCC C CATCCACA
978 AUGUGGAU CUGAUGAG X CGAA IGGGGCCC GGGCCCCC c ATCCACAT
979 GAUGUGGA CUGAUGAG X CGAA IGGGGGCC GGCCCCCC A TCCACATC
982 CGCGAUGU CUGAUGAG X CGAA IAUGGGGG CCCCCATC C ACATCGCG
983 CCGCGAUG CUGAUGAG X CGAA IGAUGGGG CCCCATCC A CATCGCGG
985 GGCCGCGA CUGAUGAG X CGAA IUGGAUGG CCATCCAC A TCGCGGCC
993 GACGUGGU CUGAUGAG X CGAA ICCGCGAU ATCGCGGC C ACCACGTC
994 GGACGUGG CUGAUGAG X CGAA IGCCGCGA TCGCGGCC A CCACGTCC
996 AGGGACGU CUGAUGAG X CGAA IUGGCCGC GCGGCCAC C ACGTCCCT
997 CAGGGACG CUGAUGAG X CGAA IGUGGCCG CGGCCACC A CGTCCCTG
1002 UGUCCCAG CUGAUGAG X CGAA lACGUGGU ACCACGTC C CTGGGACA
1003 GUGUCCCA CUGAUGAG X CGAA IGACGUGG CCACGTCC C TGGGACAC
1004 CGUGUCCC CUGAUGAG X CGAA IGGACGUG CACGTCCC T GGGACACG
1010 ACAAGGCG CUGAUGAG X CGAA IUCCCAGG CCTGGGAC A CGCCTTGT
1014 GGGGACAA CUGAUGAG X CGAA ICGUGUCC GGACACGC C TTGTCCCC
1015 GGGGGACA CUGAUGAG X CGAA IGCGUGUC GACACGCC T TGTCCCCC
1020 ACACCGGG CUGAUGAG X CGAA IACAAGGC GCCTTGTC C CCCGGTGT
1021 UACACCGG CUGAUGAG X CGAA IGACAAGG CCTTGTCC C CCGGTGTA
1022 GUACACCG CUGAUGAG X CGAA IGGACAAG CTTGTCCC C CGGTGTAC
1023 CGUACACC CUGAUGAG X CGAA IGGGACAA TTGTCCCC C GGTGTACG
1033 UUGGUCUC CUGAUGAG X CGAA ICGUACAC GTGTACGC C GAGACCAA
1039 AAGUGCUU CUGAUGAG X CGAA IUCUCGGC GCCGAGAC C AAGCACTT
1040 GAAGUGCU CUGAUGAG X CGAA IGUCUCGG CCGAGACC A AGCACTTC
Table 14
1044 AGAGGAAG CUGAUGAG X CGAA ICUUGGUC GACCAAGC A CTTCCTCT
1046 GUAGAGGA CUGAUGAG X CGAA IUGCUUGG CCAAGCAC T TCCTCTAC
1049 GGAGUAGA CUGAUGAG X CGAA IAAGUGCU AGCACTTC c TCTACTCC
1050 AGGAGUAG CUGAUGAG X CGAA IGAAGUGC GCACTTCC T CTACTCCT
1052 UGAGGAGU CUGAUGAG X CGAA IAGGAAGU ACTTCCTC T ACTCCTCA
1055 GCCUGAGG CUGAUGAG X CGAA lUAGAGGA TCCTCTAC T CCTCAGGC
1057 UCGCCUGA CUGAUGAG X CGAA IAGUAGAG CTCTACTC c TCAGGCGA
1058 GUCGCCUG CUGAUGAG X CGAA IGAGUAGA TCTACTCC T CAGGCGAC
1060 UUGUCGCC CUGAUGAG X CGAA lAGGAGUA TACTCCTC A GGCGACAA
1067 CUGCUCCU CUGAUGAG X CGAA IUCGCCUG CAGGCGAC A AGGAGCAG
1074 GCCGCAGC CUGAUGAG X CGAA ICUCCUUG CAAGGAGC A GCTGCGGC
1077 AGGGCCGC CUGAUGAG X CGAA ICUGCUCC GGAGCAGC T GCGGCCCT
1083 GGAAGGAG CUGAUGAG X CGAA ICCGCAGC GCTGCGGC C CTCCTTCC
1084 AGGAAGGA CUGAUGAG X CGAA IGCCGCAG CTGCGGCC C TCCTTCCT
1085 UAGGAAGG CUGAUGAG X CGAA IGGCCGCA TGCGGCCC T CCTTCCTA
1087 AGUAGGAA CUGAUGAG X CGAA IAGGGCCG CGGCCCTC C TTCCTACT
1088 GAGUAGGA CUGAUGAG X CGAA IGAGGGCC GGCCCTCC T TCCTACTC
1091 GCUGAGUA CUGAUGAG X CGAA IAAGGAGG CCTCCTTC C TACTCAGC
1092 AGCUGAGU CUGAUGAG X CGAA IGAAGGAG CTCCTTCC T ACTCAGCT
1095 GAGAGCUG CUGAUGAG X CGAA IUAGGAAG CTTCCTAC T CAGCTCTC
1097 CAGAGAGC CUGAUGAG X CGAA IAGUAGGA TCCTACTC A GCTCTCTG
1100 CCUCAGAG CUGAUGAG X CGAA ICUGAGUA TACTCAGC T CTCTGAGG
1102 GGCCUCAG CUGAUGAG X CGAA IAGCUGAG CTCAGCTC T CTGAGGCC
1104 UGGGCCUC CUGAUGAG X CGAA IAGAGCUG CAGCTCTC T GAGGCCCA
1110 UCAGGCUG CUGAUGAG X CGAA ICCUCAGA TCTGAGGC c CAGCCTGA llll GUCAGGCU CUGAUGAG X CGAA IGCCUCAG CTGAGGCC c AGCCTGAC
1112 AGUCAGGC CUGAUGAG X CGAA IGGCCUCA TGAGGCCC A GCCTGACT
1115 GCCAGUCA CUGAUGAG X CGAA ICUGGGCC GGCCCAGC C TGACTGGC
1116 CGCCAGUC CUGAUGAG X CGAA IGCUGGGC GCCCAGCC T GACTGGCG
1120 CGAGCGCC CUGAUGAG X CGAA IUCAGGCU AGCCTGAC T GGCGCTCG
1126 AGCCUCCG CUGAUGAG X CGAA ICGCCAGU ACTGGCGC T CGGAGGCT
1134 UCUCCACG CUGAUGAG X CGAA ICCUCCGA TCGGAGGC T CGTGGAGA
1144 AGAAAGAU CUGAUGAG X CGAA IUCUCCAC GTGGAGAC c ATCTTTCT
1145 CAGAAAGA CUGAUGAG X CGAA IGUCUCCA TGGAGACC A TCTTTCTG
1148 ACCCAGAA CUGAUGAG X CGAA IAUGGUCU AGACCATC T TTCTGGGT
1152 UGGAACCC CUGAUGAG X CGAA IAAAGAUG CATCTTTC T GGGTTCCA
1159 CAGGGCCU CUGAUGAG X CGAA IAACCCAG CTGGGTTC c AGGCCCTG
1160 CCAGGGCC CUGAUGAG X CGAA IGAACCCA TGGGTTCC A GGCCCTGG
1164 GCAUCCAG CUGAUGAG X CGAA ICCUGGAA TTCCAGGC C CTGGATGC
1165 GGCAUCCA CUGAUGAG X CGAA IGCCUGGA TCCAGGCC C TGGATGCC
1166 UGGCAUCC CUGAUGAG X CGAA IGGCCUGG CCAGGCCC T GGATGCCA
1173 GAGUCCCU CUGAUGAG X CGAA ICAUCCAG CTGGATGC c AGGGACTC
1174 GGAGUCCC CUGAUGAG X CGAA IGCAUCCA TGGATGCC A GGGACTCC
1180 CUGCGGGG CUGAUGAG X CGAA IUCCCUGG CCAGGGAC T CCCCGCAG
1182 ACCUGCGG CUGAUGAG X CGAA IAGUCCCU AGGGACTC C CCGCAGGT
1183 AACCUGCG CUGAUGAG X CGAA IGAGUCCC GGGACTCC C CGCAGGTT
1184 CAACCUGC CUGAUGAG X CGAA IGGAGUCC GGACTCCC C GCAGGTTG
Table 14
1187 GGGCAACC CUGAUGAG X CGAA ICGGGGAG CTCCCCGC A GGTTGCCC
1194 GCAGGCGG CUGAUGAG X CGAA ICAACCUG CAGGTTGC C CCGCCTGC
1195 GGCAGGCG CUGAUGAG X CGAA IGCAACCU AGGTTGCC C CGCCTGCC
1196 GGGCAGGC CUGAUGAG X CGAA IGGCAACC GGTTGCCC C GCCTGCCC
1199 CUGGGGCA CUGAUGAG X CGAA ICGGGGCA TGCCCCGC C TGCCCCAG
1200 GCUGGGGC CUGAUGAG X CGAA IGCGGGGC GCCCCGCC T GCCCCAGC
1203 AGCGCUGG CUGAUGAG X CGAA ICAGGCGG CCGCCTGC C CCAGCGCT
1204 UAGCGCUG CUGAUGAG X CGAA IGCAGGCG CGCCTGCC C CAGCGCTA
1205 GUAGCGCU CUGAUGAG X CGAA IGGCAGGC GCCTGCCC C AGCGCTAC
1206 AGUAGCGC CUGAUGAG X CGAA IGGGCAGG CCTGCCCC A GCGCTACT
1211 UUGCCAGU CUGAUGAG X CGAA ICGCUGGG CCCAGCGC T ACTGGCAA
1214 CAUUUGCC CUGAUGAG X CGAA IUAGCGCU AGCGCTAC T GGCAAATG
1218 GCCGCAUU CUGAUGAG X CGAA ICCAGUAG CTACTGGC A AATGCGGC
1227 GAAACAGG CUGAUGAG X CGAA ICCGCAUU AATGCGGC C CCTGTTTC
1228 AGAAACAG CUGAUGAG X CGAA IGCCGCAU ATGCGGCC C CTGTTTCT
1229 CAGAAACA CUGAUGAG X CGAA IGGCCGCA TGCGGCCC C TGTTTCTG
1230 CCAGAAAC CUGAUGAG X CGAA IGGGCCGC GCGGCCCC T GTTTCTGG
1236 GCAGCUCC CUGAUGAG X CGAA IAAACAGG CCTGTTTC T GGAGCTGC
1242 UCCCAAGC CUGAUGAG X CGAA ICUCCAGA TCTGGAGC T GCTTGGGA
1245 GGUUCCCA CUGAUGAG X CGAA ICAGCUCC GGAGCTGC T TGGGAACC
1253 CUGCGCGU CUGAUGAG X CGAA IUUCCCAA TTGGGAAC c ACGCGCAG
1254 ACUGCGCG CUGAUGAG X CGAA IGUUCCCA TGGGAACC A CGCGCAGT
1260 AGGGGCAC CUGAUGAG X CGAA ICGCGUGG CCACGCGC A GTGCCCCT
1265 CCCGUAGG CUGAUGAG X CGAA ICACUGCG CGCAGTGC C CCTACGGG
1266 CCCCGUAG CUGAUGAG X CGAA IGCACUGC GCAGTGCC C CTACGGGG
1267 ACCCCGUA CUGAUGAG X CGAA IGGCACUG CAGTGCCC C TACGGGGT
1268 CACCCCGU CUGAUGAG X CGAA IGGGCACU AGTGCCCC T ACGGGGTG
1278 UCUUGAGG CUGAUGAG X CGAA ICACCCCG CGGGGTGC T CCTCAAGA
1280 CGUCUUGA CUGAUGAG X CGAA IAGCACCC GGGTGCTC c TCAAGACG
1281 GCGUCUUG CUGAUGAG X CGAA IGAGCACC GGTGCTCC T CAAGACGC
1283 GUGCGUCU CUGAUGAG X CGAA lAGGAGCA TGCTCCTC A AGACGCAC
1290 GCGGGCAG CUGAUGAG X CGAA ICGUCUUG CAAGACGC A CTGCCCGC
1292 CAGCGGGC CUGAUGAG X CGAA IUGCGUCU AGACGCAC T GCCCGCTG
1295 UCGCAGCG CUGAUGAG X CGAA ICAGUGCG CGCACTGC C CGCTGCGA
1296 CUCGCAGC CUGAUGAG X CGAA IGCAGUGC GCACTGCC C GCTGCGAG
1299 CAGCUCGC CUGAUGAG X CGAA ICGGGCAG CTGCCCGC T GCGAGCTG
1306 GUGACCGC CUGAUGAG X CGAA ICUCGCAG CTGCGAGC T GCGGTCAC
1313 UGCUGGGG CUGAUGAG X CGAA IACCGCAG CTGCGGTC A CCCCAGCA
1315 GCUGCUGG CUGAUGAG X CGAA IUGACCGC GCGGTCAC C CCAGCAGC
1316 GGCUGCUG CUGAUGAG X CGAA IGUGACCG CGGTCACC C CAGCAGCC
1317 CGGCUGCU CUGAUGAG X CGAA IGGUGACC GGTCACCC C AGCAGCCG
1318 CCGGCUGC CUGAUGAG X CGAA IGGGUGAC GTCACCCC A GCAGCCGG
1321 ACACCGGC CUGAUGAG X CGAA ICUGGGGU ACCCCAGC A GCCGGTGT
1324 CAGACACC CUGAUGAG X CGAA ICUGCUGG CCAGCAGC C GGTGTCTG
1331 CCGGGCAC CUGAUGAG X CGAA IACACCGG CCGGTGTC T GTGCCCGG
1336 UUCUCCCG CUGAUGAG X CGAA ICACAGAC GTCTGTGC c CGGGAGAA
1337 CUUCUCCC CUGAUGAG X CGAA IGCACAGA TCTGTGCC c GGGAGAAG
Table 14
1347 AGCCCUGG CUGAUGAG X CGAA ICUUCUCC GGAGAAGC C CCAGGGCT
1348 GAGCCCUG CUGAUGAG X CGAA IGCUUCUC GAGAAGCC C CAGGGCTC
1349 AGAGCCCU CUGAUGAG X CGAA IGGCUUCU AGAAGCCC C AGGGCTCT
1350 CAGAGCCC CUGAUGAG X CGAA IGGGCUUC GAAGCCCC A GGGCTCTG
1355 CGCCACAG CUGAUGAG X CGAA ICCCUGGG CCCAGGGC T CTGTGGCG
1357 GCCGCCAC CUGAUGAG X CGAA IAGCCCUG CAGGGCTC T GTGGCGGC
1366 UCCUCGGG CUGAUGAG X CGAA ICCGCCAC GTGGCGGC C CCCGAGGA
1367 CUCCUCGG CUGAUGAG X CGAA IGCCGCCA TGGCGGCC C CCGAGGAG
1368 CCUCCUCG CUGAUGAG X CGAA IGGCCGCC GGCGGCCC C CGAGGAGG
1369 UCCUCCUC CUGAUGAG X CGAA IGGGCCGC GCGGCCCC C GAGGAGGA
1382 GGGGUCUG CUGAUGAG X CGAA IUCCUCCU AGGAGGAC A CAGACCCC
1384 CGGGGGUC CUGAUGAG X CGAA IUGUCCUC GAGGACAC A GACCCCCG
1388 GCGACGGG CUGAUGAG X CGAA IUCUGUGU ACACAGAC C CCCGTCGC
1389 GGCGACGG CUGAUGAG X CGAA IGUCUGUG CACAGACC C CCGTCGCC
1390 AGGCGACG CUGAUGAG X CGAA IGGUCUGU ACAGACCC C CGTCGCCT
1391 CAGGCGAC CUGAUGAG X CGAA IGGGUCUG CAGACCCC C GTCGCCTG
1397 CUGCACCA CUGAUGAG X CGAA ICGACGGG CCCGTCGC C TGGTGCAG
1398 GCUGCACC CUGAUGAG X CGAA IGCGACGG CCGTCGCC T GGTGCAGC
1404 GGAGCAGC CUGAUGAG X CGAA ICACCAGG CCTGGTGC A GCTGCTCC
1407 GGCGGAGC CUGAUGAG X CGAA ICUGCACC GGTGCAGC T GCTCCGCC
1410 GCUGGCGG CUGAUGAG X CGAA ICAGCUGC GCAGCTGC T CCGCCAGC
1412 GUGCUGGC CUGAUGAG X CGAA IAGCAGCU AGCTGCTC C GCCAGCAC
1415 GCUGUGCU CUGAUGAG X CGAA ICGGAGCA TGCTCCGC C AGCACAGC
1416 UGCUGUGC CUGAUGAG X CGAA IGCGGAGC GCTCCGCC A GCACAGCA
1419 GGCUGCUG CUGAUGAG X CGAA ICUGGCGG CCGCCAGC A CAGCAGCC
1421 GGGGCUGC CUGAUGAG X CGAA IUGCUGGC GCCAGCAC A GCAGCCCC
1424 CCAGGGGC CUGAUGAG X CGAA ICUGUGCU AGCACAGC A GCCCCTGG
1427 CUGCCAGG CUGAUGAG X CGAA ICUGCUGU ACAGCAGC C CCTGGCAG
1428 CCUGCCAG CUGAUGAG X CGAA IGCUGCUG CAGCAGCC C CTGGCAGG
1429 ACCUGCCA CUGAUGAG X CGAA IGGCUGCU AGCAGCCC C TGGCAGGT
1430 CACCUGCC CUGAUGAG X CGAA IGGGCUGC GCAGCCCC T GGCAGGTG
1434 CGUACACC CUGAUGAG X CGAA ICCAGGGG CCCCTGGC A GGTGTACG
1445 CCGCACGA CUGAUGAG X CGAA ICCGUACA TGTACGGC T TCGTGCGG
1456 CGCAGGCA CUGAUGAG X CGAA ICCCGCAC GTGCGGGC C TGCCTGCG
1457 GCGCAGGC CUGAUGAG X CGAA IGCCCGCA TGCGGGCC T GCCTGCGC
1460 CCGGCGCA CUGAUGAG X CGAA ICAGGCCC GGGCCTGC c TGCGCCGG
1461 GCCGGCGC CUGAUGAG X CGAA IGCAGGCC GGCCTGCC T GCGCCGGC
1466 CACCAGCC CUGAUGAG X CGAA ICGCAGGC GCCTGCGC c GGCTGGTG
1470 GGGGCACC CUGAUGAG X CGAA ICCGGCGC GCGCCGGC T GGTGCCCC
1476 GGCCUGGG CUGAUGAG X CGAA ICACCAGC GCTGGTGC c CCCAGGCC
1477 AGGCCUGG CUGAUGAG X CGAA IGCACCAG CTGGTGCC c CCAGGCCT
1478 GAGGCCUG CUGAUGAG X CGAA IGGCACCA TGGTGCCC c CAGGCCTC
1479 AGAGGCCU CUGAUGAG X CGAA IGGGCACC GGTGCCCC c AGGCCTCT
1480 CAGAGGCC CUGAUGAG X CGAA IGGGGCAC GTGCCCCC A GGCCTCTG
1484 GCCCCAGA CUGAUGAG X CGAA ICCUGGGG CCCCAGGC C TCTGGGGC
1485 AGCCCCAG CUGAUGAG X CGAA IGCCUGGG CCCAGGCC T CTGGGGCT
1487 GGAGCCCC CUGAUGAG X CGAA IAGGCCUG CAGGCCTC T GGGGCTCC
Table 14
1493 GUGCCUGG CUGAUGAG X CGAA ICCCCAGA TCTGGGGC T CCAGGCAC
1495 UUGUGCCU CUGAUGAG X CGAA IAGCCCCA TGGGGCTC C AGGCACAA
1496 GUUGUGCC CUGAUGAG X CGAA IGAGCCCC GGGGCTCC A GGCACAAC
1500 GUUCGUUG CUGAUGAG X CGAA ICCUGGAG CTCCAGGC A CAACGAAC
1502 GCGUUCGU CUGAUGAG X CGAA IUGCCUGG CCAGGCAC A ACGAACGC
1511 GAGGAAGC CUGAUGAG X CGAA ICGUUCGU ACGAACGC C GCTTCCTC
1514 CCUGAGGA CUGAUGAG X CGAA ICGGCGUU AACGCCGC T TCCTCAGG
1517 GUUCCUGA CUGAUGAG X CGAA IAAGCGGC GCCGCTTC c TCAGGAAC
1518 UGUUCCUG CUGAUGAG X CGAA IGAAGCGG CCGCTTCC T CAGGAACA
1520 GGUGUUCC CUGAUGAG X CGAA IAGGAAGC GCTTCCTC A GGAACACC
1526 CUUCUUGG CUGAUGAG X CGAA IUUCCUGA TCAGGAAC A CCAAGAAG
1528 AACUUCUU CUGAUGAG X CGAA lUGUUCCU AGGAACAC C AAGAAGTT
1529 GAACUUCU CUGAUGAG X CGAA IGUGUUCC GGAACACC A AGAAGTTC
1538 CAGGGAGA CUGAUGAG X CGAA IAACUUCU AGAAGTTC A TCTCCCTG
1541 CCCCAGGG CUGAUGAG X CGAA IAUGAACU AGTTCATC T CCCTGGGG
1543 UUCCCCAG CUGAUGAG X CGAA IAGAUGAA TTCATCTC C CTGGGGAA
1544 CUUCCCCA CUGAUGAG X CGAA IGAGAUGA TCATCTCC C TGGGGAAG
1545 GCUUCCCC CUGAUGAG X CGAA IGGAGAUG CATCTCCC T GGGGAAGC
1554 GCUUGGCA CUGAUGAG X CGAA ICUUCCCC GGGGAAGC A TGCCAAGC
1558 GAGAGCUU CUGAUGAG X CGAA ICAUGCUU AAGCATGC C AAGCTCTC
1559 CGAGAGCU CUGAUGAG X CGAA IGCAUGCU AGCATGCC A AGCTCTCG
1563 GCAGCGAG CUGAUGAG X CGAA ICUUGGCA TGCCAAGC T CTCGCTGC
1565 CUGCAGCG CUGAUGAG X CGAA IAGCUUGG CCAAGCTC T CGCTGCAG
1569 GCUCCUGC CUGAUGAG X CGAA ICGAGAGC GCTCTCGC T GCAGGAGC
1572 UCAGCUCC CUGAUGAG X CGAA ICAGCGAG CTCGCTGC A GGAGCTGA
1578 UCCACGUC CUGAUGAG X CGAA ICUCCUGC GCAGGAGC T GACGTGGA
1604 CCAAGCGC CUGAUGAG X CGAA IUCCCGCA TGCGGGAC T GCGCTTGG
1609 CGCAGCCA CUGAUGAG X CGAA ICGCAGUC GACTGCGC T TGGCTGCG
1614 UCCUGCGC CUGAUGAG X CGAA ICCAAGCG CGCTTGGC T GCGCAGGA
1619 UGGGCUCC CUGAUGAG X CGAA ICGCAGCC GGCTGCGC A GGAGCCCA
1625 AACCCCUG CUGAUGAG X CGAA ICUCCUGC GCAGGAGC C CAGGGGTT
1626 CAACCCCU CUGAUGAG X CGAA IGCUCCUG CAGGAGCC C AGGGGTTG
1627 CCAACCCC CUGAUGAG X CGAA IGGCUCCU AGGAGCCC A GGGGTTGG
1637 CGGAACAC CUGAUGAG X CGAA ICCAACCC GGGTTGGC T GTGTTCCG
1644 CUGCGGCC CUGAUGAG X CGAA IAACACAG CTGTGTTC c GGCCGCAG
1648 UGCUCUGC CUGAUGAG X CGAA ICCGGAAC GTTCCGGC c GCAGAGCA
1651 CGGUGCUC CUGAUGAG X CGAA ICGGCCGG CCGGCCGC A GAGCACCG
1656 GCAGACGG CUGAUGAG X CGAA ICUCUGCG CGCAGAGC A CCGTCTGC
1658 ACGCAGAC CUGAUGAG X CGAA lUGCUCUG CAGAGCAC C GTCTGCGT
1662 CCUCACGC CUGAUGAG X CGAA IACGGUGC GCACCGTC T GCGTGAGG
1676 CUUGGCCA CUGAUGAG X CGAA IAUCUCCU AGGAGATC C TGGCCAAG
1677 ACUUGGCC CUGAUGAG X CGAA IGAUCUCC GGAGATCC T GGCCAAGT
1681 AGGAACUU CUGAUGAG X CGAA ICCAGGAU ATCCTGGC C AAGTTCCT
1682 CAGGAACU CUGAUGAG X CGAA IGCCAGGA TCCTGGCC A AGTTCCTG
1688 CCAGUGCA CUGAUGAG X CGAA IAACUUGG CCAAGTTC C TGCACTGG
1689 GCCAGUGC CUGAUGAG X CGAA IGAACUUG CAAGTTCC T GCACTGGC
1692 UCAGCCAG CUGAUGAG X CGAA ICAGGAAC GTTCCTGC A CTGGCTGA
Table 14
1694 CAUCAGCC CUGAUGAG X CGAA IUGCAGGA TCCTGCAC T GGCTGATG
1698 CACUCAUC CUGAUGAG X CGAA ICCAGUGC GCACTGGC T GATGAGTG
1722 ACCUGAGC CUGAUGAG X CGAA ICUCGACG CGTCGAGC T GCTCAGGT
1725 AAGACCUG CUGAUGAG X CGAA ICAGCUCG CGAGCTGC T CAGGTCTT
1727 GAAAGACC CUGAUGAG X CGAA IAGCAGCU AGCTGCTC A GGTCTTTC
1732 UAAAAGAA CUGAUGAG X CGAA IACCUGAG CTCAGGTC T TTCTTTTA
1736 GACAUAAA CUGAUGAG X CGAA IAAAGACC GGTCTTTC T TTTATGTC
1745 GGUCUCCG CUGAUGAG X CGAA IACAUAAA TTTATGTC A CGGAGACC
1753 UGAAACGU CUGAUGAG X CGAA IUCUCCGU ACGGAGAC C ACGTTTCA
1754 UUGAAACG CUGAUGAG X CGAA IGUCUCCG CGGAGACC A CGTTTCAA
1761 UGUUCUUU CUGAUGAG X CGAA IAAACGUG CACGTTTC A AAAGAACA
1769 AAAGAGCC CUGAUGAG X CGAA IUUCUUUU AAAAGAAC A GGCTCTTT
1773 AGAAAAAG CUGAUGAG X CGAA ICCUGUUC GAACAGGC T CTTTTTCT
1775 GUAGAAAA CUGAUGAG X CGAA IAGCCUGU ACAGGCTC T TTTTCTAC
1781 CUUCCGGU CUGAUGAG X CGAA IAAAAAGA TCTTTTTC T ACCGGAAG
1784 ACUCUUCC CUGAUGAG X CGAA lUAGAAAA TTTTCTAC C GGAAGAGT
1796 CUUGCUCC CUGAUGAG X CGAA IACACUCU AGAGTGTC T GGAGCAAG
1802 UUGCAACU CUGAUGAG X CGAA ICUCCAGA TCTGGAGC A AGTTGCAA
1809 CAAUGCUU CUGAUGAG X CGAA ICAACUUG CAAGTTGC A AAGCATTG
1814 GAUUCCAA CUGAUGAG X CGAA ICUUUGCA TGCAAAGC A TTGGAATC
1823 GUGCUGUC CUGAUGAG X CGAA IAUUCCAA TTGGAATC A GACAGCAC
1827 UCAAGUGC CUGAUGAG X CGAA IUC'UGAUU AATCAGAC A GCACTTGA
1830 UCUUCAAG CUGAUGAG X CGAA ICUGUCUG CAGACAGC A CTTGAAGA
1832 CCUCUUCA CUGAUGAG X CGAA IUGCUGUC GACAGCAC T TGAAGAGG
1845 CCCGCAGC CUGAUGAG X CGAA ICACCCUC GAGGGTGC A GCTGCGGG
1848 GCUCCCGC CUGAUGAG X CGAA ICUGCACC GGTGCAGC T. GCGGGAGC
1857 CUUCCGAC CUGAUGAG X CGAA ICUCCCGC GCGGGAGC T GTCGGAAG
1867 CUGACCUC CUGAUGAG X CGAA ICUUCCGA TCGGAAGC A GAGGTCAG
1874 AUGCUGCC CUGAUGAG X CGAA IACCUCUG CAGAGGTC A GGCAGCAT
1878 CCCGAUGC CUGAUGAG X CGAA ICCUGACC GGTCAGGC A GCATCGGG
1881 CUUCCCGA CUGAUGAG X CGAA ICUGCCUG CAGGCAGC A TCGGGAAG
1891 GCGGGCCU CUGAUGAG X CGAA ICUUCCCG CGGGAAGC C AGGCCCGC
1892 GGCGGGCC CUGAUGAG X CGAA IGCUUCCC GGGAAGCC A GGCCCGCC
1896 GCAGGGCG CUGAUGAG X CGAA ICCUGGCU AGCCAGGC C CGCCCTGC
1897 AGCAGGGC CUGAUGAG X CGAA IGCCUGGC GCCAGGCC C GCCCTGCT
1900 GUCAGCAG CUGAUGAG X CGAA ICGGGCCU AGGCCCGC C CTGCTGAC
1901 CGUCAGCA CUGAUGAG X CGAA IGCGGGCC GGCCCGCC C TGCTGACG
1902 ACGUCAGC CUGAUGAG X CGAA IGGCGGGC GCCCGCCC T GCTGACGT
1905 UGGACGUC CUGAUGAG X CGAA ICAGGGCG CGCCCTGC T GACGTCCA
1912 CGGAGUCU CUGAUGAG X CGAA IACGUCAG CTGACGTC C AGACTCCG
1913 GCGGAGUC CUGAUGAG X CGAA IGACGUCA TGACGTCC A GACTCCGC
1917 UGAAGCGG CUGAUGAG X CGAA IUCUGGAC GTCCAGAC T CCGCTTCA
1919 GAUGAAGC CUGAUGAG X CGAA IAGUCUGG CCAGACTC C GCTTCATC
1922 GGGGAUGA CUGAUGAG X CGAA ICGGAGUC GACTCCGC T TCATCCCC
1925 CUUGGGGA CUGAUGAG X CGAA IAAGCGGA TCCGCTTC A TCCCCAAG
1928 AGGCUUGG CUGAUGAG X CGAA IAUGAAGC GCTTCATC C CCAAGCCT
1929 CAGGCUUG CUGAUGAG X CGAA IGAUGAAG CTTCATCC C CAAGCCTG
Table 14
1930 UCAGGCUU CUGAUGAG X CGAA IGGAUGAA TTCATCCC C AAGCCTGA
1931 GUCAGGCU CUGAUGAG X CGAA IGGGAUGA TCATCCCC A AGCCTGAC
1935 GCCCGUCA CUGAUGAG X CGAA ICUUGGGG CCCCAAGC C TGACGGGC
1936 AGCCCGUC CUGAUGAG X CGAA IGCUUGGG CCCAAGCC T GACGGGCT
1944 UCGGCCGC CUGAUGAG X CGAA ICCCGUCA TGACGGGC T GCGGCCGA
1950 UCACAAUC CUGAUGAG X CGAA ICCGCAGC GCTGCGGC C GATTGTGA
1961 GUAGUCCA CUGAUGAG X CGAA lUUCACAA TTGTGAAC A TGGACTAC
1967 CACGACGU CUGAUGAG X CGAA IUCCAUGU ACATGGAC T ACGTCGTG
1981 AACGUUCU CUGAUGAG X CGAA ICUCCCAC GTGGGAGC C AGAACGTT
1982 GAACGUUC CUGAUGAG X CGAA IGCUCCCA TGGGAGCC A GAACGTTC
1991 UUCUCUGC CUGAUGAG X CGAA IAACGUUC GAACGTTC C GCAGAGAA
1994 CUUUUCUC CUGAUGAG X CGAA ICGGAACG CGTTCCGC A GAGAAAAG
2008 AGACGCUC CUGAUGAG X CGAA ICCCUCUU AAGAGGGC C GAGCGTCT
2016 UCGAGGUG CUGAUGAG X CGAA IACGCUCG CGAGCGTC T CACCTCGA
2018 CCUCGAGG CUGAUGAG X CGAA IAGACGCU AGCGTCTC A CCTCGAGG
2020 ACCCUCGA CUGAUGAG X CGAA IUGAGACG CGTCTCAC C TCGAGGGT
2021 CACCCUCG CUGAUGAG X CGAA IGUGAGAC GTCTCACC T CGAGGGTG
2035 CUGAACAG CUGAUGAG X CGAA ICCUUCAC GTGAAGGC A CTGTTCAG
2037 CGCUGAAC CUGAUGAG X CGAA IUGCCUUC GAAGGCAC T GTTCAGCG
2042 GAGCACGC CUGAUGAG X CGAA IAACAGUG CACTGTTC A GCGTGCTC
2049 CGUAGUUG CUGAUGAG X CGAA ICACGCUG CAGCGTGC T CAACTACG
2051 CUCGUAGU CUGAUGAG X CGAA IAGCACGC GCGTGCTC A ACTACGAG
2054 CCGCUCGU CUGAUGAG X CGAA IUUGAGCA TGCTCAAC T ACGAGCGG
2072 GAGGCCGG CUGAUGAG X CGAA ICGCCGCG CGCGGCGC C CCGGCCTC
2073 GGAGGCCG CUGAUGAG X CGAA IGCGCCGC GCGGCGCC C CGGCCTCC
2074 AGGAGGCC CUGAUGAG X CGAA IGGCGCCG CGGCGCCC C GGCCTCCT
2078 GCCCAGGA CUGAUGAG X CGAA ICCGGGGC GCCCCGGC C TCCTGGGC
2079 CGCCCAGG CUGAUGAG X CGAA IGCCGGGG CCCCGGCC T CCTGGGCG
2081 GGCGCCCA CUGAUGAG X CGAA IAGGCCGG CCGGCCTC C TGGGCGCC
2082 AGGCGCCC CUGAUGAG X CGAA IGAGGCCG CGGCCTCC T GGGCGCCT
2089 AGCACAGA CUGAUGAG X CGAA ICGCCCAG CTGGGCGC C TCTGTGCT
2090 CAGCACAG CUGAUGAG X CGAA IGCGCCCA TGGGCGCC T CTGTGCTG
2092 CCCAGCAC CUGAUGAG X CGAA IAGGCGCC GGCGCCTC T GTGCTGGG
2097 CCAGGCCC CUGAUGAG X CGAA ICACAGAG CTCTGTGC T GGGCCTGG
2102 AUCGUCCA CUGAUGAG X CGAA ICCCAGCA TGCTGGGC c TGGACGAT
2103 UAUCGUCC CUGAUGAG X CGAA IGCCCAGC GCTGGGCC T GGACGATA
2114 GGCCCUGU CUGAUGAG X CGAA IAUAUCGU ACGATATC c ACAGGGCC
2115 AGGCCCUG CUGAUGAG X CGAA IGAUAUCG CGATATCC A CAGGGCCT
2117 CCAGGCCC CUGAUGAG X CGAA IUGGAUAU ATATCCAC A GGGCCTGG
2122 GUGCGCCA CUGAUGAG X CGAA ICCCUGUG CACAGGGC C TGGCGCAC
2123 GGUGCGCC CUGAUGAG X CGAA IGCCCUGU ACAGGGCC T GGCGCACC
2129 CACGAAGG CUGAUGAG X CGAA ICGCCAGG CCTGGCGC A CCTTCGTG
2131 AGCACGAA CUGAUGAG X CGAA lUGCGCCA TGGCGCAC C TTCGTGCT
2132 CAGCACGA CUGAUGAG X CGAA IGUGCGCC GGCGCACC T TCGTGCTG
2139 GCACACGC CUGAUGAG X CGAA ICACGAAG CTTCGTGC T GCGTGTGC
2152 GGGUCCUG CUGAUGAG X CGAA ICCCGCAC GTGCGGGC C CAGGACCC
2153 CGGGUCCU CUGAUGAG X CGAA IGCCCGCA TGCGGGCC C AGGACCCG
Table 14
2154 GCGGGUCC CUGAUGAG X CGAA IGGCCCGC GCGGGCCC A GGACCCGC
2159 AGGCGGCG CUGAUGAG X CGAA IUCCUGGG CCCAGGAC C CGCCGCCT
2160 CAGGCGGC CUGAUGAG X CGAA IGUCCUGG CCAGGACC C GCCGCCTG
2163 GCUCAGGC CUGAUGAG X CGAA ICGGGUCC GGACCCGC C GCCTGAGC
2166 ACAGCUCA CUGAUGAG X CGAA ICGGCGGG CCCGCCGC C TGAGCTGT
2167 UACAGCUC CUGAUGAG X CGAA IGCGGCGG CCGCCGCC T GAGCTGTA
2172 CAAAGUAC CUGAUGAG X CGAA ICUCAGGC GCCTGAGC T GTACTTTG
2177 CUUGACAA CUGAUGAG X CGAA IUACAGCU AGCTGTAC T TTGTCAAG
2183 AUCCACCU CUGAUGAG X CGAA lACAAAGU ACTTTGTC A AGGTGGAT
2210 GGGGAUGG CUGAUGAG X CGAA IUCGUACG CGTACGAC A CCATCCCC
2212 UGGGGGAU CUGAUGAG X CGAA lUGUCGUA TACGACAC C ATCCCCCA
2213 CUGGGGGA CUGAUGAG X CGAA IGUGUCGU ACGACACC A TCCCCCAG
2216 GUCCUGGG CUGAUGAG X CGAA IAUGGUGU ACACCATC C CCCAGGAC
2217 UGUCCUGG CUGAUGAG X CGAA IGAUGGUG CACCATCC C CCAGGACA
2218 CUGUCCUG CUGAUGAG X CGAA IGGAUGGU ACCATCCC C CAGGACAG
2219 CCUGUCCU CUGAUGAG X CGAA IGGGAUGG CCATCCCC C AGGACAGG
2220 GCCUGUCC CUGAUGAG X CGAA IGGGGAUG CATCCCCC A GGACAGGC
2225 CGUGAGCC CUGAUGAG X CGAA IUCCUGGG CCCAGGAC A GGCTCACG
2229 CCUCCGUG CUGAUGAG X CGAA ICCUGUCC GGACAGGC T CACGGAGG
2231 GACCUCCG CUGAUGAG X CGAA IAGCCUGU ACAGGCTC A CGGAGGTC
2240 GCUGGCGA CUGAUGAG X CGAA IACCUCCG CGGAGGTC A TCGCCAGC
2245 AUGAUGCU CUGAUGAG X CGAA ICGAUGAC GTCATCGC C AGCATCAT
2246 GAUGAUGC CUGAUGAG X CGAA IGCGAUGA TCATCGCC A GCATCATC
2249 UUUGAUGA CUGAUGAG X CGAA ICUGGCGA TCGCCAGC A TCATCAAA
2252 GGGUUUGA CUGAUGAG X CGAA IAUGCUGG CCAGCATC A TCAAACCC
2255 CUGGGGUU CUGAUGAG X CGAA IAUGAUGC GCATCATC A AACCCCAG
2259 UGUUCUGG CUGAUGAG X CGAA IUUUGAUG CATCAAAC C CCAGAACA
2260 GUGUUCUG CUGAUGAG X CGAA IGUUUGAU ATCAAACC C CAGAACAC
2261 CGUGUUCU CUGAUGAG X CGAA IGGUUUGA TCAAACCC C AGAACACG
2262 ACGUGUUC CUGAUGAG X CGAA IGGGUUUG CAAACCCC A GAACACGT
2267 GCAGUACG CUGAUGAG X CGAA IUUCUGGG CCCAGAAC A CGTACTGC
2273 ACGCACGC CUGAUGAG X CGAA IUACGUGU ACACGTAC T GCGTGCGT
2290 UGGACCAC CUGAUGAG X CGAA ICAUACCG CGGTATGC C GTGGTCCA
2297 GGCCUUCU CUGAUGAG X CGAA IACCACGG CCGTGGTC C AGAAGGCC
2298 CGGCCUUC CUGAUGAG X CGAA IGACCACG CGTGGTCC A GAAGGCCG
2305 CCAUGGGC CUGAUGAG X CGAA ICCUUCUG CAGAAGGC C GCCCATGG
2308 UGCCCAUG CUGAUGAG X CGAA ICGGCCUU AAGGCCGC C CATGGGCA
2309 GUGCCCAU CUGAUGAG X CGAA IGCGGCCU AGGCCGCC C ATGGGCAC
2310 CGUGCCCA CUGAUGAG X CGAA IGGCGGCC GGCCGCCC A TGGGCACG
2316 UGCGGACG CUGAUGAG X CGAA ICCCAUGG CCATGGGC A CGTCCGCA
2321 GGCCUUGC CUGAUGAG X CGAA IACGUGCC GGCACGTC C GCAAGGCC
2324 GAAGGCCU CUGAUGAG X CGAA ICGGACGU ACGTCCGC A AGGCCTTC
2329 CUCUUGAA CUGAUGAG X CGAA ICCUUGCG CGCAAGGC C TTCAAGAG
2330 GCUCUUGA CUGAUGAG X CGAA IGCCUUGC GCAAGGCC T TCAAGAGC
2333 GUGGCUCU CUGAUGAG X CGAA IAAGGCCU AGGCCTTC A AGAGCCAC
2339 AGAGACGU CUGAUGAG X CGAA ICUCUUGA TCAAGAGC C ACGTCTCT
2340 UAGAGACG CUGAUGAG X CGAA IGCUCUUG CAAGAGCC A CGTCTCTA
Table 14
2345 CAAGGUAG CUGAUGAG X CGAA IACGUGGC GCCACGTC T CTACCTTG
2347 GUCAAGGU CUGAUGAG X CGAA IAGACGUG CACGTCTC T ACCTTGAC
2350 UCUGUCAA CUGAUGAG X CGAA IUAGAGAC GTCTCTAC C TTGACAGA
2351 GUCUGUCA CUGAUGAG X CGAA IGUAGAGA TCTCTACC T TGACAGAC
2356 UGGAGGUC CUGAUGAG X CGAA IUCAAGGU ACCTTGAC A GACCTCCA
2360 CGGCUGGA CUGAUGAG X CGAA IUCUGUCA TGACAGAC C TCCAGCCG
2361 ACGGCUGG CUGAUGAG X CGAA IGUCUGUC GACAGACC T CCAGCCGT
2363 GUACGGCU CUGAUGAG X CGAA IAGGUCUG CAGACCTC C AGCCGTAC
2364 UGUACGGC CUGAUGAG X CGAA IGAGGUCU AGACCTCC A GCCGTACA
2367 GCAUGUAC CUGAUGAG X CGAA ICUGGAGG CCTCCAGC C GTACATGC
2372 CUGUCGCA CUGAUGAG X CGAA IUACGGCU AGCCGTAC A TGCGACAG
2379 CCACGAAC CUGAUGAG X CGAA IUCGCAUG CATGCGAC A GTTCGTGG
2389 UGCAGGUG CUGAUGAG X CGAA ICCACGAA TTCGTGGC T CACCTGCA
2391 CCUGCAGG CUGAUGAG X CGAA IAGCCACG CGTGGCTC A CCTGCAGG
2393 CUCCUGCA CUGAUGAG X CGAA IUGAGCCA TGGCTCAC C TGCAGGAG
2394 UCUCCUGC CUGAUGAG X CGAA IGUGAGCC GGCTCACC T GCAGGAGA
2397 UGGUCUCC CUGAUGAG X CGAA ICAGGUGA TCACCTGC A GGAGACCA
2404 AGCGGGCU CUGAUGAG X CGAA IUCUCCUG CAGGAGAC C AGCCCGCT
2405 CAGCGGGC CUGAUGAG X CGAA IGUCUCCU AGGAGACC A GCCCGCTG
2408 CCUCAGCG CUGAUGAG X CGAA ICUGGUCU AGACCAGC C CGCTGAGG
2409 CCCUCAGC CUGAUGAG X CGAA IGCUGGUC GACCAGCC C GCTGAGGG
2412 CAUCCCUC CUGAUGAG X CGAA ICGGGCUG CAGCCCGC T GAGGGATG
2422 AUGACGAC CUGAUGAG X CGAA ICAUCCCU AGGGATGC C GTCGTCAT
2429 CUGCUCGA CUGAUGAG X CGAA IACGACGG CCGTCGTC A TCGAGCAG
2436 AGGAGCUC CUGAUGAG X CGAA ICUCGAUG CATCGAGC A GAGCTCCT
2441 CAGGGAGG CUGAUGAG X CGAA ICUCUGCU AGCAGAGC T CCTCCCTG
2443 UUCAGGGA CUGAUGAG X CGAA IAGCUCUG CAGAGCTC c TCCCTGAA
2444 AUUCAGGG CUGAUGAG X CGAA IGAGCUCU AGAGCTCC T CCCTGAAT
2446 UCAUUCAG CUGAUGAG X CGAA IAGGAGCU AGCTCCTC c CTGAATGA
2447 CUCAUUCA CUGAUGAG X CGAA IGAGGAGC GCTCCTCC c TGAATGAG
2448 CCUCAUUC CUGAUGAG X CGAA IGGAGGAG CTCCTCCC T GAATGAGG
2458 CCACUGCU CUGAUGAG X CGAA ICCUCAUU AATGAGGC c AGCAGTGG
2459 GCCACUGC CUGAUGAG X CGAA IGCCUCAU ATGAGGCC A GCAGTGGC
2462 GAGGCCAC CUGAUGAG X CGAA ICUGGCCU AGGCCAGC A GTGGCCTC
2468 GUCGAAGA CUGAUGAG X CGAA ICCACUGC GCAGTGGC C TCTTCGAC
2469 CGUCGAAG CUGAUGAG X CGAA IGCCACUG CAGTGGCC T CTTCGACG
2471 GACGUCGA CUGAUGAG X CGAA IAGGCCAC GTGGCCTC T TCGACGTC
2480 GCGUAGGA CUGAUGAG X CGAA IACGUCGA TCGACGTC T TCCTACGC
2483 GAAGCGUA CUGAUGAG X CGAA IAAGACGU ACGTCTTC C TACGCTTC
2484 UGAAGCGU CUGAUGAG X CGAA IGAAGACG CGTCTTCC T ACGCTTCA
2489 GCACAUGA CUGAUGAG X CGAA ICGUAGGA TCCTACGC T TCATGTGC
2492 GUGGCACA CUGAUGAG X CGAA IAAGCGUA TACGCTTC A TGTGCCAC
2498 GGCGUGGU CUGAUGAG X CGAA ICACAUGA TCATGTGC C ACCACGCC
2499 CGGCGUGG CUGAUGAG X CGAA IGCACAUG CATGTGCC A CCACGCCG
2501 CACGGCGU CUGAUGAG X CGAA IUGGCACA TGTGCCAC C ACGCCGTG
2502 GCACGGCG CUGAUGAG X CGAA IGUGGCAC GTGCCACC A CGCCGTGC
2506 AUGCGCAC CUGAUGAG X CGAA ICGUGGUG CACCACGC C GTGCGCAT
Table 14
2513 GCCCCUGA CUGAUGAG X CGAA ICGCACGG CCGTGCGC A TCAGGGGC
2516 CUUGCCCC CUGAUGAG X CGAA IAUGCGCA TGCGCATC A GGGGCAAG
2522 GUAGGACU CUGAUGAG X CGAA ICCCCUGA TCAGGGGC A AGTCCTAC
2527 UGGACGUA CUGAUGAG X CGAA IACUUGCC GGCAAGTC C TACGTCCA
2528 CUGGACGU CUGAUGAG X CGAA IGACUUGC GCAAGTCC T ACGTCCAG
2534 CUGGCACU CUGAUGAG X CGAA IACGUAGG CCTACGTC C AGTGCCAG
2535 CCUGGCAC CUGAUGAG X CGAA IGACGUAG CTACGTCC A GTGCCAGG
2540 GAUCCCCU CUGAUGAG X CGAA ICACUGGA TCCAGTGC C AGGGGATC
2541 GGAUCCCC CUGAUGAG X CGAA IGCACUGG CCAGTGCC A GGGGATCC
2549 GCCCUGCG CUGAUGAG X CGAA IAUCCCCU AGGGGATC C CGCAGGGC
2550 AGCCCUGC CUGAUGAG X CGAA IGAUCCCC GGGGATCC C GCAGGGCT
2553 UGGAGCCC CUGAUGAG X CGAA ICGGGAUC GATCCCGC A GGGCTCCA
2558 GAGGAUGG CUGAUGAG X CGAA ICCCUGCG CGCAGGGC T CCATCCTC
2560 GAGAGGAU CUGAUGAG X CGAA IAGCCCUG CAGGGCTC c ATCCTCTC
2561 GGAGAGGA CUGAUGAG X CGAA IGAGCCCU AGGGCTCC A TCCTCTCC
2564 CGUGGAGA CUGAUGAG X CGAA IAUGGAGC GCTCCATC C TCTCCACG
2565 GCGUGGAG CUGAUGAG X CGAA IGAUGGAG CTCCATCC T CTCCACGC
2567 CAGCGUGG CUGAUGAG X CGAA IAGGAUGG CCATCCTC T CCACGCTG
2569 AGCAGCGU CUGAUGAG X CGAA IAGAGGAU ATCCTCTC c ACGCTGCT
2570 GAGCAGCG CUGAUGAG X CGAA IGAGAGGA TCCTCTCC A CGCTGCTC
2574 UGCAGAGC CUGAUGAG X CGAA ICGUGGAG CTCCACGC T GCTCTGCA
2577 GGCUGCAG CUGAUGAG X CGAA ICAGCGUG CACGCTGC T CTGCAGCC
2579 CAGGCUGC CUGAUGAG X CGAA IAGCAGCG CGCTGCTC T GCAGCCTG
2582 GCACAGGC CUGAUGAG X CGAA ICAGAGCA TGCTCTGC A GCCTGTGC
2585 GUAGCACA CUGAUGAG X CGAA ICUGCAGA TCTGCAGC C TGTGCTAC
2586 CGUAGCAC CUGAUGAG X CGAA IGCUGCAG CTGCAGCC T GTGCTACG
2591 GUCGCCGU CUGAUGAG X CGAA ICACAGGC GCCTGTGC T ACGGCGAC
2600 GUUCUCCA CUGAUGAG X CGAA IUCGCCGU ACGGCGAC A TGGAGAAC
2609 AAACAGCU CUGAUGAG X CGAA lUUCUCCA TGGAGAAC A AGCTGTTT
2613 CCGCAAAC CUGAUGAG X CGAA ICUUGUUC GAACAAGC T GTTTGCGG
2640 GCAGGAGC CUGAUGAG X CGAA ICCCGUCC GGACGGGC T GCTCCTGC
2643 AACGCAGG CUGAUGAG X CGAA ICAGCCCG CGGGCTGC T CCTGCGTT
2645 CAAACGCA CUGAUGAG X CGAA IAGCAGCC GGCTGCTC C TGCGTTTG
2646 CCAAACGC CUGAUGAG X CGAA IGAGCAGC GCTGCTCC T GCGTTTGG
2666 CACCAACA CUGAUGAG X CGAA IAAAUCAU ATGATTTC T TGTTGGTG
2677 AGGUGAGG CUGAUGAG X CGAA IUCACCAA TTGGTGAC A CCTCACCT
2679 UGAGGUGA CUGAUGAG X CGAA IUGUCACC GGTGACAC C TCACCTCA
2680 GUGAGGUG CUGAUGAG X CGAA IGUGUCAC GTGACACC T CACCTCAC
2682 GGGUGAGG CUGAUGAG X CGAA IAGGUGUC GACACCTC A CCTCACCC
2684 GUGGGUGA CUGAUGAG X CGAA IUGAGGUG CACCTCAC C TCACCCAC
2685 CGUGGGUG CUGAUGAG X CGAA IGUGAGGU ACCTCACC T CACCCACG
2687 CGCGUGGG CUGAUGAG X CGAA IAGGUGAG CTCACCTC A CCCACGCG
2689 UUCGCGUG CUGAUGAG X CGAA IUGAGGUG CACCTCAC C CACGCGAA
2690 UUUCGCGU CUGAUGAG X CGAA IGUGAGGU ACCTCACC C ACGCGAAA
2691 UUUUCGCG CUGAUGAG X CGAA IGGUGAGG CCTCACCC A CGCGAAAA
2701 CUGAGGAA CUGAUGAG X CGAA IUUUUCGC GCGAAAAC C TTCCTCAG
2702 CCUGAGGA CUGAUGAG X CGAA IGUUUUCG CGAAAACC T TCCTCAGG
Table 14
2705 GGUCCUGA CUGAUGAG X CGAA IAAGGUUU AAACCTTC C TCAGGACC
2706 GGGUCCUG CUGAUGAG X CGAA IGAAGGUU AACCTTCC T CAGGACCC
2708 CAGGGUCC CUGAUGAG X CGAA IAGGAAGG CCTTCCTC A GGACCCTG
2713 CGGACCAG CUGAUGAG X CGAA IUCCUGAG CTCAGGAC C CTGGTCCG
2714 UCGGACCA CUGAUGAG X CGAA IGUCCUGA TCAGGACC C TGGTCCGA
2715 CUCGGACC CUGAUGAG X CGAA IGGUCCUG CAGGACCC T GGTCCGAG
2720 GACACCUC CUGAUGAG X CGAA IACCAGGG CCCTGGTC c GAGGTGTC
2729 AUACUCAG CUGAUGAG X CGAA IACACCUC GAGGTGTC c CTGAGTAT
2730 CAUACUCA CUGAUGAG X CGAA IGACACCU AGGTGTCC c TGAGTATG
2731 CCAUACUC CUGAUGAG X CGAA IGGACACC GGTGTCCC T GAGTATGG
2741 CACCACGC CUGAUGAG X CGAA ICCAUACU AGTATGGC T GCGTGGTG
2753 CUUCCGCA CUGAUGAG X CGAA IUUCACCA TGGTGAAC T TGCGGAAG
2764 UUCACCAC CUGAUGAG X CGAA IUCUUCCG CGGAAGAC A GTGGTGAA
2774 UACAGGGA CUGAUGAG X CGAA IUUCACCA TGGTGAAC T TCCCTGTA
2777 UUCUACAG CUGAUGAG X CGAA lAAGUUCA TGAACTTC C CTGTAGAA
2778 CUUCUACA CUGAUGAG X CGAA IGAAGUUC GAACTTCC C TGTAGAAG
2779 UCUUCUAC CUGAUGAG X CGAA IGGAAGUU AACTTCCC T GTAGAAGA
2794 CCACCCAG CUGAUGAG X CGAA ICCUCGUC GACGAGGC c CTGGGTGG
2795 GCCACCCA CUGAUGAG X CGAA IGCCUCGU ACGAGGCC c TGGGTGGC
2796 UGCCACCC CUGAUGAG X CGAA IGGCCUCG CGAGGCCC T GGGTGGCA
2804 AAAAGCCG CUGAUGAG X CGAA ICCACCCA TGGGTGGC A CGGCTTTT
2809 UGAACAAA CUGAUGAG X CGAA ICCGUGCC GGCACGGC T TTTGTTCA
2817 CCGGCAUC CUGAUGAG X CGAA IAACAAAA TTTTGTTC A GATGCCGG
2823 CGUGGGCC CUGAUGAG X CGAA ICAUCUGA TCAGATGC C GGCCCACG
2827 AGGCCGUG CUGAUGAG X CGAA ICCGGCAU ATGCCGGC C CACGGCCT
2828 UAGGCCGU CUGAUGAG X CGAA IGCCGGCA TGCCGGCC C ACGGCCTA
2829 AUAGGCCG CUGAUGAG X CGAA IGGCCGGC GCCGGCCC A CGGCCTAT
2834 GGGGAAUA CUGAUGAG X CGAA ICCGUGGG CCCACGGC C TATTCCCC
2835 AGGGGAAU CUGAUGAG X CGAA IGCCGUGG CCACGGCC T ATTCCCCT
2840 GCACCAGG CUGAUGAG X CGAA IAAUAGGC GCCTATTC C CCTGGTGC
2841 CGCACCAG CUGAUGAG X CGAA IGAAUAGG CCTATTCC C CTGGTGCG
2842 CCGCACCA CUGAUGAG X CGAA IGGAAUAG CTATTCCC C TGGTGCGG
2843 GCCGCACC CUGAUGAG X CGAA IGGGAAUA TATTCCCC T GGTGCGGC
2852 CAGCAGCA CUGAUGAG X CGAA ICCGCACC GGTGCGGC c TGCTGCTG
2853 CCAGCAGC CUGAUGAG X CGAA IGCCGCAC GTGCGGCC T GCTGCTGG
2856 UAUCCAGC CUGAUGAG X CGAA ICAGGCCG CGGCCTGC T GCTGGATA
2859 GGGUAUCC CUGAUGAG X CGAA ICAGCAGG CCTGCTGC T GGATACCC
2866 AGGGUCCG CUGAUGAG X CGAA IUAUCCAG CTGGATAC c CGGACCCT
2867 CAGGGUCC CUGAUGAG X CGAA IGUAUCCA TGGATACC c GGACCCTG
2872 ACCUCCAG CUGAUGAG X CGAA IUCCGGGU ACCCGGAC c CTGGAGGT
2873 CACCUCCA CUGAUGAG X CGAA IGUCCGGG CCCGGACC c TGGAGGTG
2874 GCACCUCC CUGAUGAG X CGAA IGGUCCGG CCGGACCC T GGAGGTGC
2883 AGUCGCUC CUGAUGAG X CGAA ICACCUCC GGAGGTGC A GAGCGACT
2891 GCUGGAGU CUGAUGAG X CGAA IUCGCUCU AGAGCGAC T ACTCCAGC
2894 AUAGCUGG CUGAUGAG X CGAA IUAGUCGC GCGACTAC T CCAGCTAT
2896 GCAUAGCU CUGAUGAG X CGAA IAGUAGUC GACTACTC c AGCTATGC
2897 GGCAUAGC CUGAUGAG X CGAA IGAGUAGU ACTACTCC A GCTATGCC
Table 14
2900 CCGGGCAU CUGAUGAG X CGAA ICUGGAGU ACTCCAGC T ATGCCCGG
2905 GAGGUCCG CUGAUGAG X CGAA ICAUAGCU AGCTATGC C CGGACCTC
2906 GGAGGUCC CUGAUGAG X CGAA IGCAUAGC GCTATGCC C GGACCTCC
2911 CUGAUGGA CUGAUGAG X CGAA IUCCGGGC GCCCGGAC C TCCATCAG
2912 UCUGAUGG CUGAUGAG X CGAA IGUCCGGG CCCGGACC T CCATCAGA
2914 GCUCUGAU CUGAUGAG X CGAA IAGGUCCG CGGACCTC C ATCAGAGC
2915 GGCUCUGA CUGAUGAG X CGAA IGAGGUCC GGACCTCC A TCAGAGCC
2918 ACUGGCUC CUGAUGAG X CGAA lAUGGAGG CCTCCATC A GAGCCAGT
2923 GUGAGACU CUGAUGAG X CGAA ICUCUGAU ATCAGAGC C AGTCTCAC
2924 GGUGAGAC CUGAUGAG X CGAA IGCUCUGA TCAGAGCC A GTCTCACC
2928 UGAAGGUG CUGAUGAG X CGAA IACUGGCU AGCCAGTC T CACCTTCA
2930 GUUGAAGG CUGAUGAG X CGAA IAGACUGG CCAGTCTC A CCTTCAAC
2932 CGGUUGAA CUGAUGAG X CGAA IUGAGACU AGTCTCAC C TTCAACCG
2933 GCGGUUGA CUGAUGAG X CGAA IGUGAGAC GTCTCACC T TCAACCGC
2936 GCCGCGGU CUGAUGAG X CGAA IAAGGUGA TCACCTTC A ACCGCGGC
2939 GAAGCCGC CUGAUGAG X CGAA IUUGAAGG CCTTCAAC C GCGGCTTC
2945 AGCCUUGA CUGAUGAG X CGAA ICCGCGGU ACCGCGGC T TCAAGGCT
2948 CCCAGCCU CUGAUGAG X CGAA IAAGCCGC GCGGCTTC A AGGCTGGG
2953 UUCCUCCC CUGAUGAG X CGAA ICCUUGAA TTCAAGGC T GGGAGGAA
2963 GCGACGCA CUGAUGAG X CGAA IUUCCUCC GGAGGAAC A TGCGTCGC
2972 AAAGAGUU CUGAUGAG X CGAA ICGACGCA TGCGTCGC A AACTCTTT
2976 CCCCAAAG CUGAUGAG X CGAA IUUUGCGA TCGCAAAC T CTTTGGGG
2978 GACCCCAA CUGAUGAG X CGAA IAGUUUGC GCAAACTC T TTGGGGTC
2987 CAGCCGCA CUGAUGAG X CGAA IACCCCAA TTGGGGTC T TGCGGCTG
2994 GACACUUC CUGAUGAG X CGAA ICCGCAAG CTTGCGGC T GAAGTGTC
3003 ACAGGCUG CUGAUGAG X CGAA IACACUUC GAAGTGTC A CAGCCTGT
3005 AAACAGGC CUGAUGAG X CGAA IUGACACU AGTGTCAC A GCCTGTTT
3008 CAGAAACA CUGAUGAG X CGAA ICUGUGAC GTCACAGC C TGTTTCTG
3009 CCAGAAAC CUGAUGAG X CGAA IGCUGUGA TCACAGCC T GTTTCTGG
3015 GCAAAUCC CUGAUGAG X CGAA IAAACAGG CCTGTTTC T GGATTTGC
-3024 UGUUCACC CUGAUGAG X CGAA ICAAAUCC GGATTTGC A GGTGAACA
3032 CUGGAGGC CUGAUGAG X CGAA IUUCACCU AGGTGAAC A GCCTCCAG
3035 CGUCUGGA CUGAUGAG X CGAA ICUGUUCA TGAACAGC C TCCAGACG
3036 CCGUCUGG CUGAUGAG X CGAA IGCUGUUC GAACAGCC T CCAGACGG
3038 CACCGUCU CUGAUGAG X CGAA IAGGCUGU ACAGCCTC C AGACGGTG
3039 ACACCGUC CUGAUGAG X CGAA IGAGGCUG CAGCCTCC A GACGGTGT
3050 GAUGUUGG CUGAUGAG X CGAA ICACACCG CGGTGTGC A CCAACATC
3052 UAGAUGUU CUGAUGAG X CGAA IUGCACAC GTGTGCAC C AACATCTA
3053 GUAGAUGU CUGAUGAG X CGAA IGUGCACA TGTGCACC A ACATCTAC
3056 CUUGUAGA CUGAUGAG X CGAA IUUGGUGC GCACCAAC A TCTACAAG
3059 GAUCUUGU CUGAUGAG X CGAA IAUGUUGG CCAACATC T ACAAGATC
3062 GAGGAUCU CUGAUGAG X CGAA IUAGAUGU ACATCTAC A AGATCCTC
3068 CAGCAGGA CUGAUGAG X CGAA IAUCUUGU ACAAGATC C TCCTGCTG
3069 GCAGCAGG CUGAUGAG X CGAA IGAUCUUG CAAGATCC T CCTGCTGC
3071 CUGCAGCA CUGAUGAG X CGAA IAGGAUCU AGATCCTC C TGCTGCAG
3072 CCUGCAGC CUGAUGAG X CGAA IGAGGAUC GATCCTCC T GCTGCAGG
3075 ACGCCUGC CUGAUGAG X CGAA ICAGGAGG CCTCCTGC T GCAGGCGT
Table 14
3078 UGUACGCC CUGAUGAG X CGAA ICAGCAGG CCTGCTGC A GGCGTACA
3086 GUGAAACC CUGAUGAG X CGAA IUACGCCU AGGCGTAC A GGTTTCAC
3093 CACAUGCG CUGAUGAG X CGAA IAAACCUG CAGGTTTC A CGCATGTG
3097 AGCACACA CUGAUGAG X CGAA ICGUGAAA TTTCACGC A TGTGTGCT
3105 GGAGCUGC CUGAUGAG X CGAA ICACACAU ATGTGTGC T GCAGCTCC
3108 AUGGGAGC CUGAUGAG X CGAA ICAGCACA TGTGCTGC A GCTCCCAT
3111 GAAAUGGG CUGAUGAG X CGAA ICUGCAGC GCTGCAGC T CCCATTTC
3113 AUGAAAUG CUGAUGAG X CGAA IAGCUGCA TGCAGCTC C CATTTCAT
3114 GAUGAAAU CUGAUGAG X CGAA IGAGCUGC GCAGCTCC c ATTTCATC
3115 UGAUGAAA CUGAUGAG X CGAA IGGAGCUG CAGCTCCC A TTTCATCA
3120 CUUGCUGA CUGAUGAG X CGAA IAAAUGGG CCCATTTC A TCAGCAAG
3123 AAACUUGC CUGAUGAG X CGAA IAUGAAAU ATTTCATC A GCAAGTTT
3126 UCCAAACU CUGAUGAG X CGAA ICUGAUGA TCATCAGC A AGTTTGGA
3140 AAAUGUGG CUGAUGAG X CGAA IUUCUUCC GGAAGAAC c CCACATTT
3141 AAAAUGUG CUGAUGAG X CGAA IGUUCUUC GAAGAACC c CACATTTT
3142 AAAAAUGU CUGAUGAG X CGAA IGGUUCUU AAGAACCC c ACATTTTT
3143 GAAAAAUG CUGAUGAG X CGAA IGGGUUCU AGAACCCC A CATTTTTC
3145 AGGAAAAA CUGAUGAG X CGAA lUGGGGUU AACCCCAC A TTTTTCCT
3152 GACGCGCA CUGAUGAG X CGAA IAAAAAUG CATTTTTC C TGCGCGTC
3153 UGACGCGC CUGAUGAG X CGAA IGAAAAAU ATTTTTCC T GCGCGTCA
3161 GUCAGAGA CUGAUGAG X CGAA IACGCGCA TGCGCGTC A TCTCTGAC
3164 CGUGUCAG CUGAUGAG X CGAA IAUGACGC GCGTCATC T CTGACACG
3166 GCCGUGUC CUGAUGAG X CGAA IAGAUGAC GTCATCTC T GACACGGC
3170 GGAGGCCG CUGAUGAG X CGAA IUCAGAGA TCTCTGAC A CGGCCTCC
3175 CAGAGGGA CUGAUGAG X CGAA ICCGUGUC GACACGGC C TCCCTCTG
3176 GCAGAGGG CUGAUGAG X CGAA IGCCGUGU ACACGGCC T CCCTCTGC
3178 UAGCAGAG CUGAUGAG X CGAA IAGGCCGU ACGGCCTC C CTCTGCTA
3179 GUAGCAGA CUGAUGAG X CGAA IGAGGCCG CGGCCTCC C TCTGCTAC
3180 AGUAGCAG CUGAUGAG X CGAA IGGAGGCC GGCCTCCC T CTGCTACT
3182 GGAGUAGC CUGAUGAG X CGAA IAGGGAGG CCTCCCTC T GCTACTCC
3185 GAUGGAGU CUGAUGAG X CGAA ICAGAGGG CCCTCTGC T ACTCCATC
3188 CAGGAUGG CUGAUGAG X CGAA IUAGCAGA TCTGCTAC T CCATCCTG
3190 UUCAGGAU CUGAUGAG X CGAA IAGUAGCA TGCTACTC C ATCCTGAA
3191 UUUCAGGA CUGAUGAG X CGAA IGAGUAGC GCTACTCC A TCCTGAAA
3194 GGCUUUCA CUGAUGAG X CGAA lAUGGAGU ACTCCATC C TGAAAGCC
3195 UGGCUUUC CUGAUGAG X CGAA IGAUGGAG CTCCATCC T GAAAGCCA
3202 GCGUUCUU CUGAUGAG X CGAA ICUUUCAG CTGAAAGC C AAGAACGC
3203 UGCGUUCU CUGAUGAG X CGAA IGCUUUCA TGAAAGCC A AGAACGCA
3211 GACAUCCC CUGAUGAG X CGAA ICGUUCUU AAGAACGC A GGGATGTC
3222 UGGCCCCC CUGAUGAG X CGAA ICGACAUC GATGTCGC T GGGGGCCA
3229 σcGcccuu CUGAUGAG X CGAA ICCCCCAG CTGGGGGC C AAGGGCGC
3230 GGCGCCCU CUGAUGAG X CGAA IGCCCCCA TGGGGGCC A AGGGCGCC
3238 GGGCCGGC CUGAUGAG X CGAA ICGCCCUU AAGGGCGC C GCCGGCCC
3241 AGAGGGCC CUGAUGAG X CGAA ICGGCGCC GGCGCCGC C GGCCCTCT
3245 GGGCAGAG CUGAUGAG X CGAA ICCGGCGG CCGCCGGC C CTCTGCCC
3246 AGGGCAGA CUGAUGAG X CGAA IGCCGGCG CGCCGGCC C TCTGCCCT
3247 GAGGGCAG CUGAUGAG X CGAA IGGCCGGC GCCGGCCC T CTGCCCTC
Table 14
3249 CGGAGGGC CUGAUGAG X CGAA IAGGGCCG CGGCCCTC T GCCCTCCG
3252 CCUCGGAG CUGAUGAG X CGAA ICAGAGGG CCCTCTGC C CTCCGAGG
3253 GCCUCGGA CUGAUGAG X CGAA IGCAGAGG CCTCTGCC C TCCGAGGC
3254 GGCCUCGG CUGAUGAG X CGAA IGGCAGAG CTCTGCCC T CCGAGGCC
3256 ACGGCCUC CUGAUGAG X CGAA IAGGGCAG CTGCCCTC C GAGGCCGT
3262 CACUGCAC CUGAUGAG X CGAA ICCUCGGA TCCGAGGC C GTGCAGTG
3267 ACAGCCAC CUGAUGAG X CGAA ICACGGCC GGCCGTGC A GTGGCTGT
3273 GGUGGCAC CUGAUGAG X CGAA ICCACUGC GCAGTGGC T GTGCCACC
3278 UGCUUGGU CUGAUGAG X CGAA ICACAGCC GGCTGTGC C ACCAAGCA
3279 AUGCUUGG CUGAUGAG X CGAA IGCACAGC GCTGTGCC A CCAAGCAT
3281 GAAUGCUU CUGAUGAG X CGAA IUGGCACA TGTGCCAC C AAGCATTC
3282 GGAAUGCU CUGAUGAG X CGAA IGUGGCAC GTGCCACC A AGCATTCC
3286 AGCAGGAA CUGAUGAG X CGAA ICUUGGUG CACCAAGC A TTCCTGCT
3290 CUUGAGCA CUGAUGAG X CGAA IAAUGCUU AAGCATTC C TGCTCAAG
3291 GCUUGAGC CUGAUGAG X CGAA IGAAUGCU AGCATTCC T GCTCAAGC
3294 UCAGCUUG CUGAUGAG X CGAA ICAGGAAU ATTCCTGC T CAAGCTGA
3296 AGUCAGCU CUGAUGAG X CGAA IAGCAGGA TCCTGCTC A AGCTGACT
3300 GUCGAGUC CUGAUGAG X CGAA ICUUGAGC GCTCAAGC T GACTCGAC
3304 CGGUGUCG CUGAUGAG X CGAA IUCAGCUU AAGCTGAC T CGACACCG
3309 UGACACGG CUGAUGAG X CGAA IUCGAGUC GACTCGAC A CCGTGTCA
3311 GGUGACAC CUGAUGAG X CGAA IUGUCGAG CTCGACAC C GTGTCACC
3317 CACGUAGG CUGAUGAG X CGAA IACACGGU ACCGTGTC A CCTACGTG
3319 GGCACGUA CUGAUGAG X CGAA IUGACACG CGTGTCAC C TACGTGCC
3320 UGGCACGU CUGAUGAG X CGAA IGUGACAC GTGTCACC T ACGTGCCA
3327 CCAGGAGU CUGAUGAG X CGAA ICACGUAG CTACGTGC C ACTCCTGG
3328 CCCAGGAG CUGAUGAG X CGAA IGCACGUA TACGTGCC A CTCCTGGG
3330 ACCCCAGG CUGAUGAG X CGAA IUGGCACG CGTGCCAC T CCTGGGGT
3332 UGACCCCA CUGAUGAG X CGAA lAGUGGCA TGCCACTC C TGGGGTCA
3333 GUGACCCC CUGAUGAG X CGAA IGAGUGGC GCCACTCC T GGGGTCAC
3340 GUCCUGAG CUGAUGAG X CGAA IACCCCAG CTGGGGTC A CTCAGGAC
3342 CUGUCCUG CUGAUGAG X CGAA IUGACCCC GGGGTCAC T CAGGACAG
3344 GGCUGUCC CUGAUGAG X CGAA IAGUGACC GGTCACTC A GGACAGCC
3349 GUCUGGGC CUGAUGAG X CGAA IUCCUGAG CTCAGGAC A GCCCAGAC
3352 UGCGUCUG CUGAUGAG X CGAA ICUGUCCU AGGACAGC C CAGACGCA
3353 CUGCGUCU CUGAUGAG X CGAA IGCUGUCC GGACAGCC C AGACGCAG
3354 GCUGCGUC CUGAUGAG X CGAA IGGCUGUC GACAGCCC A GACGCAGC
3360 GACUCAGC CUGAUGAG X CGAA ICGUCUGG CCAGACGC A GCTGAGTC
3363 UCCGACUC CUGAUGAG X CGAA ICUGCGUC GACGCAGC T GAGTCGGA
3375 UCCCCGGG CUGAUGAG X CGAA ICUUCCGA TCGGAAGC T CCCGGGGA
3377 CGUCCCCG CUGAUGAG X CGAA IAGCUUCC GGAAGCTC C CGGGGACG
3378 UCGUCCCC CUGAUGAG X CGAA IGAGCUUC GAAGCTCC C GGGGACGA
3390 GGGCAGUC CUGAUGAG X CGAA ICGUCGUC GACGACGC T GACTGCCC
3394 UCCAGGGC CUGAUGAG X CGAA IUCAGCGU ACGCTGAC T GCCCTGGA
3397 GCCUCCAG CUGAUGAG X CGAA ICAGUCAG CTGACTGC c CTGGAGGC
3398 GGCCUCCA CUGAUGAG X CGAA IGCAGUCA TGACTGCC c TGGAGGCC
3399 CGGCCUCC CUGAUGAG X CGAA IGGCAGUC GACTGCCC T GGAGGCCG
3406 UUGGCUGC CUGAUGAG X CGAA ICCUCCAG CTGGAGGC c GCAGCCAA
Table 14
3409 GGGUUGGC CUGAUGAG X CGAA ICGGCCUC GAGGCCGC A GCCAACCC
3412 GCCGGGUU CUGAUGAG X CGAA ICUGCGGC GCCGCAGC C AACCCGGC
3413 UGCCGGGU CUGAUGAG X CGAA IGCUGCGG CCGCAGCC A ACCCGGCA
3416 CAGUGCCG CUGAUGAG X CGAA IUUGGCUG CAGCCAAC C CGGCACTG
3417 GCAGUGCC CUGAUGAG X CGAA IGUUGGCU AGCCAACC C GGCACTGC
3421 GAGGGCAG CUGAUGAG X CGAA ICCGGGUU AACCCGGC A CTGCCCTC
3423 CUGAGGGC CUGAUGAG X CGAA IUGCCGGG CCCGGCAC T GCCCTCAG
3426 AGUCUGAG CUGAUGAG X CGAA ICAGUGCC GGCACTGC C CTCAGACT
3427 AAGUCUGA CUGAUGAG X CGAA IGCAGUGC GCACTGCC C TCAGACTT
3428 GAAGUCUG CUGAUGAG X CGAA IGGCAGUG CACTGCCC T CAGACTTC
3430 UUGAAGUC CUGAUGAG X CGAA IAGGGCAG CTGCCCTC A GACTTCAA
3434 GGUCUUGA CUGAUGAG X CGAA IUCUGAGG CCTCAGAC T TCAAGACC
3437 GAUGGUCU CUGAUGAG X CGAA IAAGUCUG CAGACTTC A AGACCATC
3442 UCCAGGAU CUGAUGAG X CGAA IUCUUGAA TTCAAGAC C ATCCTGGA
3443 GUCCAGGA CUGAUGAG X CGAA IGUCUUGA TCAAGACC A TCCTGGAC
3446 UCAGUCCA CUGAUGAG X CGAA IAUGGUCU AGACCATC C TGGACTGA
3447 AUCAGUCC CUGAUGAG X CGAA IGAUGGUC GACCATCC T GGACTGAT
3452 UGGCCAUC CUGAUGAG X CGAA IUCCAGGA TCCTGGAC T GATGGCCA
3459 GGGCGGGU CUGAUGAG X CGAA ICCAUCAG CTGATGGC c ACCCGCCC
3460 UGGGCGGG CUGAUGAG X CGAA IGCCAUCA TGATGGCC A CCCGCCCA
3462 UGUGGGCG CUGAUGAG X CGAA IUGGCCAU ATGGCCAC C CGCCCACA
3463 CUGUGGGC CUGAUGAG X CGAA IGUGGCCA TGGCCACC C GCCCACAG
3466 UGGCUGUG CUGAUGAG X CGAA ICGGGUGG CCACCCGC C CACAGCCA
3467 CUGGCUGU CUGAUGAG X CGAA IGCGGGUG CACCCGCC C ACAGCCAG
3468 CCUGGCUG CUGAUGAG X CGAA IGGCGGGU ACCCGCCC A CAGCCAGG
3470 GGCCUGGC CUGAUGAG X CGAA IUGGGCGG CCGCCCAC A GCCAGGCC
3473 CUCGGCCU CUGAUGAG X CGAA ICUGUGGG CCCACAGC C AGGCCGAG
3474 UCUCGGCC CUGAUGAG X CGAA IGCUGUGG CCACAGCC A GGCCGAGA
3478 CUGCUCUC CUGAUGAG X CGAA ICCUGGCU AGCCAGGC C GAGAGCAG
3485 CUGGUGUC CUGAUGAG X CGAA ICUCUCGG CCGAGAGC A GACACCAG
3489 GCUGCUGG CUGAUGAG X CGAA IUCUGCUC GAGCAGAC A CCAGCAGC
3491 GGGCUGCU CUGAUGAG X CGAA IUGUCUGC GCAGACAC C AGCAGCCC
3492 AGGGCUGC CUGAUGAG X CGAA IGUGUCUG CAGACACC A GCAGCCCT
3495 GACAGGGC CUGAUGAG X CGAA ICUGGUGU ACACCAGC A GCCCTGTC
3498 CGUGACAG CUGAUGAG X CGAA ICUGCUGG CCAGCAGC C CTGTCACG
3499 GCGUGACA CUGAUGAG X CGAA IGCUGCUG CAGCAGCC C TGTCACGC
3500 GGCGUGAC CUGAUGAG X CGAA IGGCUGCU AGCAGCCC T GTCACGCC
3504 GCCCGGCG CUGAUGAG X CGAA IACAGGGC GCCCTGTC A CGCCGGGC
3508 UAGAGCCC CUGAUGAG X CGAA ICGUGACA TGTCACGC C GGGCTCTA
3513 GGACGUAG CUGAUGAG X CGAA ICCCGGCG CGCCGGGC T CTACGTCC
3515 UGGGACGU CUGAUGAG X CGAA IAGCCCGG CCGGGCTC T ACGTCCCA
3521 CCUCCCUG CUGAUGAG X CGAA IACGUAGA TCTACGTC c CAGGGAGG
3522 CCCUCCCU CUGAUGAG X CGAA IGACGUAG CTACGTCC c AGGGAGGG
3523 UCCCUCCC CUGAUGAG X CGAA IGGACGUA TACGTCCC A GGGAGGGA
3540 UGGGUGUG CUGAUGAG X CGAA ICCGCCCC GGGGCGGC C CACACCCA
3541 CUGGGUGU CUGAUGAG X CGAA IGCCGCCC GGGCGGCC C ACACCCAG
3542 CCUGGGUG CUGAUGAG X CGAA IGGCCGCC GGCGGCCC A CACCCAGG
Table 14
3544 GGCCUGGG CUGAUGAG X CGAA IUGGGCCG CGGCCCAC A CCCAGGCC
3546 CGGGCCUG CUGAUGAG X CGAA IUGUGGGC GCCCACAC C CAGGCCCG
3547 GCGGGCCU CUGAUGAG X CGAA IGUGUGGG CCCACACC C AGGCCCGC
3548 UGCGGGCC CUGAUGAG X CGAA IGGUGUGG CCACACCC A GGCCCGCA
3552 GCGGUGCG CUGAUGAG X CGAA ICCUGGGU ACCCAGGC C CGCACCGC
3553 AGCGGUGC CUGAUGAG X CGAA IGCCUGGG CCCAGGCC C GCACCGCT
3556 CCCAGCGG CUGAUGAG X CGAA ICGGGCCU AGGCCCGC A CCGCTGGG
3558 CUCCCAGC CUGAUGAG X CGAA IUGCGGGC GCCCGCAC C GCTGGGAG
3561 AGACUCCC CUGAUGAG X CGAA ICGGUGCG CGCACCGC T GGGAGTCT
3569 CAGGCCUC CUGAUGAG X CGAA IACUCCCA TGGGAGTC T GAGGCCTG
3575 CUCACUCA CUGAUGAG X CGAA ICCUCAGA TCTGAGGC C TGAGTGAG
3576 ACUCACUC CUGAUGAG X CGAA IGCCUCAG CTGAGGCC T GAGTGAGT
3592 CAGGCCUC CUGAUGAG X CGAA ICCAAACA TGTTTGGC C GAGGCCTG
3598 GACAUGCA CUGAUGAG X CGAA ICCUCGGC GCCGAGGC C TGCATGTC
3599 GGACAUGC CUGAUGAG X CGAA IGCCUCGG CCGAGGCC T GCATGTCC
3602 GCCGGACA CUGAUGAG X CGAA ICAGGCCU AGGCCTGC A TGTCCGGC
3607 CUUCAGCC CUGAUGAG X CGAA IACAUGCA TGCATGTC C GGCTGAAG
3611 CAGCCUUC CUGAUGAG X CGAA ICCGGACA TGTCCGGC T GAAGGCTG
3618 GGACACUC CUGAUGAG X CGAA ICCUUCAG CTGAAGGC T GAGTGTCC
3626 CCUCAGCC CUGAUGAG X CGAA IACACUCA TGAGTGTC c GGCTGAGG
3630 CAGGCCUC CUGAUGAG X CGAA ICCGGACA TGTCCGGC T GAGGCCTG
3636 CUCGCUCA CUGAUGAG X CGAA ICCUCAGC GCTGAGOC c TGAGCGAG
3637 ACUCGCUC CUGAUGAG X CGAA IGCCUCAG CTGAGGCC T GAGCGAGT
3649 CCUUGGCU CUGAUGAG X CGAA IACACUCG CGAGTGTC c AGCCAAGG
3650 CCCUUGGC CUGAUGAG X CGAA IGACACUC GAGTGTCC A GCCAAGGG
3653 CAGCCCUU CUGAUGAG X CGAA ICUGGACA TGTCCAGC C AAGGGCTG
3654 UCAGCCCU CUGAUGAG X CGAA IGCUGGAC GTCCAGCC A AGGGCTGA
3660 GGACACUC CUGAUGAG X CGAA ICCCUUGG CCAAGGGC T GAGTGTCC
3668 GGUGUGCU CUGAUGAG X CGAA IACACUCA TGAGTGTC C AGCACACC
3669 AGGUGUGC CUGAUGAG X CGAA IGACACUC GAGTGTCC A GCACACCT
3672 GGCAGGUG CUGAUGAG X CGAA ICUGGACA TGTCCAGC A CACCTGCC
3674 ACGGCAGG CUGAUGAG X CGAA IUGCUGGA TCCAGCAC A CCTGCCGT
3676 AGACGGCA CUGAUGAG X CGAA IUGUGCUG CAGCACAC C TGCCGTCT
3677 AAGACGGC CUGAUGAG X CGAA IGUGUGCU AGCACACC T GCCGTCTT
3680 GUGAAGAC CUGAUGAG X CGAA ICAGGUGU ACACCTGC C GTCTTCAC
3684 GGAAGUGA CUGAUGAG X CGAA IACGGCAG CTGCCGTC T TCACTTCC
3687 UGGGGAAG CUGAUGAG X CGAA IAAGACGG CCGTCTTC A CTTCCCCA
3689 UGUGGGGA CUGAUGAG X CGAA IUGAAGAC GTCTTCAC T TCCCCACA
3692 GCCUGUGG CUGAUGAG X CGAA IAAGUGAA TTCACTTC C CCACAGGC
3693 AGCCUGUG CUGAUGAG X CGAA IGAAGUGA TCACTTCC C CACAGGCT
3694 CAGCCUGU CUGAUGAG X CGAA IGGAAGUG CACTTCCC C ACAGGCTG
3695 CCAGCCUG CUGAUGAG X CGAA IGGGAAGU ACTTCCCC A CAGGCTGG
3697 CGCCAGCC CUGAUGAG X CGAA IUGGGGAA TTCCCCAC A GGCTGGCG
3701 CGAGCGCC CUGAUGAG X CGAA ICCUGUGG CCACAGGC T GGCGCTCG
3707 UGGAGCCG CUGAUGAG X CGAA ICGCCAGC GCTGGCGC T CGGCTCCA
3712 UGGGGUGG CUGAUGAG X CGAA ICCGAGCG CGCTCGGC T CCACCCCA
3714 CCUGGGGU CUGAUGAG X CGAA IAGCCGAG CTCGGCTC C ACCCCAGG
Table 14
3715 CCCUGGGG CUGAUGAG X CGAA IGAGCCGA TCGGCTCC A CCCCAGGG
3717 GGCCCUGG CUGAUGAG X CGAA IUGGAGCC GGCTCCAC C CCAGGGCC
3718 UGGCCCUG CUGAUGAG X CGAA IGUGGAGC GCTCCACC C CAGGGCCA
3719 CUGGCCCU CUGAUGAG X CGAA IGGUGGAG CTCCACCC C AGGGCCAG
3720 GCUGGCCC CUGAUGAG X CGAA IGGGUGGA TCCACCCC A GGGCCAGC
3725 GAAAAGCU CUGAUGAG X CGAA ICCCUGGG CCCAGGGC C AGCTTTTC
3726 GGAAAAGC CUGAUGAG X CGAA IGCCCUGG CCAGGGCC A GCTTTTCC
3729 UGAGGAAA CUGAUGAG X CGAA ICUGGCCC GGGCCAGC T TTTCCTCA
3734 CCUGGUGA CUGAUGAG X CGAA IAAAAGCU AGCTTTTC C TCACCAGG
3735 UCCUGGUG CUGAUGAG X CGAA IGAAAAGC GCTTTTCC T CACCAGGA
3737 GCUCCUGG CUGAUGAG X CGAA lAGGAAAA TTTTCCTC A CCAGGAGC
3739 GGGCUCCU CUGAUGAG X CGAA IUGAGGAA TTCCTCAC C AGGAGCCC
3740 CGGGCUCC CUGAUGAG X CGAA IGUGAGGA TCCTCACC A GGAGCCCG
3746 GGAAGCCG CUGAUGAG X CGAA ICUCCUGG CCAGGAGC C CGGCTTCC
3747 UGGAAGCC CUGAUGAG X CGAA IGCUCCUG CAGGAGCC C GGCTTCCA
3751 GGAGUGGA CUGAUGAG X CGAA ICCGGGCU AGCCCGGC T TCCACTCC
3754 UGGGGAGU CUGAUGAG X CGAA IAAGCCGG CCGGCTTC C ACTCCCCA
3755 GUGGGGAG CUGAUGAG X CGAA IGAAGCCG CGGCTTCC A CTCCCCAC
3757 AUGUGGGG CUGAUGAG X CGAA IUGGAAGC GCTTCCAC T CCCCACAT
3759 CUAUGUGG CUGAUGAG X CGAA IAGUGGAA TTCCACTC C CCACATAG
3760 CCUAUGUG CUGAUGAG X CGAA IGAGUGGA TCCACTCC C CACATAGG
3761 UCCUAUGU CUGAUGAG X CGAA IGGAGUGG CCACTCCC C ACATAGGA
3762 UUCCUAUG CUGAUGAG X CGAA IGGGAGUG CACTCCCC A CATAGGAA
3764 UAUUCCUA CUGAUGAG X CGAA IUGGGGAG CTCCCCAC A TAGGAATA
3776 CUGGGGAU CUGAUGAG X CGAA IACUAUUC GAATAGTC C ATCCCCAG
3777 UCUGGGGA CUGAUGAG X CGAA IGACUAUU AATAGTCC A TCCCCAGA
3780 GAAUCUGG CUGAUGAG X CGAA IAUGGACU AGTCCATC C CCAGATTC
3781 CGAAUCUG CUGAUGAG X CGAA IGAUGGAC GTCCATCC C CAGATTCG
3782 GCGAAUCU CUGAUGAG X CGAA IGGAUGGA TCCATCCC C AGATTCGC
3783 GGCGAAUC CUGAUGAG X CGAA IGGGAUGG CCATCCCC A GATTCGCC
3791 UGAACAAU CUGAUGAG X CGAA ICGAAUCU AGATTCGC C ATTGTTCA'
3792 GUGAACAA CUGAUGAG X CGAA IGCGAAUC GATTCGCC A TTGTTCAC
3799 GCGAGGGG' CUGAUGAG X CGAA IAACAAUG CATTGTTC A CCCCTCGC
3801 GGGCGAGG CUGAUGAG X CGAA IUGAACAA TTGTTCAC C CCTCGCCC
3802 AGGGCGAG CUGAUGAG X CGAA IGUGAACA TGTTCACC C cTcσcccT
3803 CAGGGCGA CUGAUGAG X CGAA IGGUGAAC GTTCACCC C TCGCCCTG
3804 GCAGGGCG CUGAUGAG X CGAA IGGGUGAA TTCACCCC T CGCCCTGC
3808 GAGGGCAG CUGAUGAG X CGAA ICGAGGGG CCCCTCGC C CTGCCCTC
3809 GGAGGGCA CUGAUGAG X CGAA IGCGAGGG CCCTCGCC C TGCCCTCC
3810 AGGAGGGC CUGAUGAG X CGAA IGGCGAGG CCTCGCCC T GCCCTCCT
3813 CAAAGGAG CUGAUGAG X CGAA ICAGGGCG CGCCCTGC C CTCCTTTG
3814 GCAAAGGA CUGAUGAG X CGAA IGCAGGGC GCCCTGCC C TCCTTTGC
3815 GGCAAAGG CUGAUGAG X CGAA IGGCAGGG CCCTGCCC T CCTTTGCC
3817 AAGGCAAA CUGAUGAG X CGAA IAGGGCAG CTGCCCTC c TTTGCCTT
3818 GAAGGCAA CUGAUGAG X CGAA IGAGGGCA TGCCCTCC T TTGCCTTC
3823 GGGUGGAA CUGAUGAG X CGAA ICAAAGGA TCCTTTGC c TTCCACCC
3824 GGGGUGGA CUGAUGAG X CGAA IGCAAAGG CCTTTGCC T TCCACCCC
Table 14
3827 GUGGGGGU CUGAUGAG X CGAA IAAGGCAA TTGCCTTC C ACCCCCAC
3828 GGUGGGGG CUGAUGAG X CGAA IGAAGGCA TGCCTTCC A CCCCCACC
3830 AUGGUGGG CUGAUGAG X CGAA IUGGAAGG CCTTCCAC C CCCACCAT
3831 GAUGGUGG CUGAUGAG X CGAA IGUGGAAG CTTCCACC C CCACCATC
3832 GGAUGGUG CUGAUGAG X CGAA IGGUGGAA TTCCACCC C CACCATCC
3833 UGGAUGGU CUGAUGAG X CGAA IGGGUGGA TCCACCCC C ACCATCCA
3834 CUGGAUGG CUGAUGAG X CGAA IGGGGUGG CCACCCCC A CCATCCAG
3836 ACCUGGAU CUGAUGAG X CGAA IUGGGGGU ACCCCCAC C ATCCAGGT
3837 CACCUGGA CUGAUGAG X CGAA IGUGGGGG CCCCCACC A TCCAGGTG
3840 CUCCACCU CUGAUGAG X CGAA IAUGGUGG CCACCATC C AGGTGGAG
3841 UCUCCACC CUGAUGAG X CGAA IGAUGGUG CACCATCC A GGTGGAGA
3851 CUUCUCAG CUGAUGAG X CGAA IUCUCCAC GTGGAGAC C CTGAGAAG
3852 CCUUCUCA CUGAUGAG X CGAA IGUCUCCA TGGAGACC C TGAGAAGG
3853 UCCUUCUC CUGAUGAG X CGAA IGGUCUCC GGAGACCC T GAGAAGGA
3863 GCUCCCAG CUGAUGAG X CGAA IUCCUUCU AGAAGGAC C CTGGGAGC
3864 AGCUCCCA CUGAUGAG X CGAA IGUCCUUC GAAGGACC C TGGGAGCT
3865 GAGCUCCC CUGAUGAG X CGAA IGGUCCUU AAGGACCC T GGGAGCTC
3872 AUUCCCAG CUGAUGAG X CGAA ICUCCCAG CTGGGAGC T CTGGGAAT
3874 AAAUUCCC CUGAUGAG X CGAA IAGCUCCC GGGAGCTC T GGGAATTT
3891 ACACCUUU CUGAUGAG X CGAA IUCACUCC GGAGTGAC c AAAGGTGT
3892 CACACCUU CUGAUGAG X CGAA IGUCACUC GAGTGACC A AAGGTGTG
3902 GUGUACAG CUGAUGAG X CGAA ICACACCU AGGTGTGC C CTGTACAC
3903 UGUGUACA CUGAUGAG X CGAA IGCACACC GGTGTGCC C TGTACACA
3904 CUGUGUAC CUGAUGAG X CGAA IGGCACAC GTGTGCCC T GTACACAG
3909 CUCGCCUG CUGAUGAG X CGAA IUACAGGG CCCTGTAC A CAGGCGAG
3911 UCCUCGCC CUGAUGAG X CGAA IUGUACAG CTGTACAC A GGCGAGGA
3921 AGGUGCAG CUGAUGAG X CGAA IUCCUCGC GCGAGGAC C CTGCACCT
3922 CAGGUGCA CUGAUGAG X CGAA IGUCCUCG CGAGGACC C TGCACCTG
3923 CCAGGUGC CUGAUGAG X CGAA IGGUCCUC GAGGACCC T GCACCTGG
3926 CAUCCAGG CUGAUGAG X CGAA ICAGGGUC GACCCTGC A CCTGGATG
3928 CCCAUCCA CUGAUGAG X CGAA IUGCAGGG CCCTGCAC C TGGATGGG
3929 CCCCAUCC CUGAUGAG X CGAA IGUGCAGG CCTGCACC T GGATGGGG
3941 ACCCACAG CUGAUGAG X CGAA IACCCCCA TGGGGGTC C CTGTGGGT
3942 GACCCACA CUGAUGAG X CGAA IGACCCCC GGGGGTCC C TGTGGGTC
3943 UGACCCAC CUGAUGAG X CGAA IGGACCCC GGGGTCCC T GTGGGTCA
3951 CCCCAAUU CUGAUGAG X CGAA IACCCACA TGTGGGTC A AATTGGGG
3968 ACUCCCAC CUGAUGAG X CGAA ICACCUCC GGAGGTGC T GTGGGAGT
3984 AUAUAUUC CUGAUGAG X CGAA IUAUUUUA TAAAATAC T GAATATAT
4002 UUCAAAAC CUGAUGAG X CGAA IAAAAACU AGTTTTTC A GTTTTGAA
Stem Length = 8 . Core Sequence = CUGAUGAG X CGAA (X = GCCGUUAGGC or other stem II sequence and length (greater than or equal to 2 base-pairs)). I = Inosine nucleotide
Seql = TERT (Homo sapiens telomerase reverse transcriptase (TERT) mRNA, 4015 bp); Nakamura et al.. Science 277 (5328), 955-959 (1997)
Table 15
Table 15: Human telomerase reverse transcriptase (TERT) G-Cleaver Ribozyme and Target Sequence
Table 15
202 CUUUCCGCGC G CUGGU ACCAG UGAUGGCAUGCACUAUGCGCG GCGCGGAAAG
216 GUGGCCCAGU G CCUGG CCAGG UGAUGGCAUGCACUAUGCGCG ACUGGGCCAC
223 AGUGCCUGGU G UGCGU ACGCA UGAUGGCAUGCACUAUGCGCG ACCAGGCACU
225 UGCCUGGUGU G CGUGC GCACG UGAUGGCAUGCACUAUGCGCG ACACCAGGCA
229 UGGUGUGCGU G CCCUG CAGGG UGAUGGCAUGCACUAUGCGCG ACGCACACCA
239 GCCCUGGGAC G CACGG CCGUG UGAUGGCAUGCACUAUGCGCG GUCCCAGGGC
247 ACGCACGGCC G CCCCC GGGGG UGAUGGCAUGCACUAUGCGCG GGCCGUGCGU
254 GCCGCCCCCC G CCGCC GGCGG UGAUGGCAUGCACUAUGCGCG GGGGGGCGGC
257 GCCCCCCGCC G CCCCC GGGGG UGAUGGCAUGCACUAUGCGCG GGCGGGGGGC
270 CCCUCCUUCC G CCAGG CCUGG UGAUGGCAUGCACUAUGCGCG GGAAGGAGGG
277 UCCGCCAGGU G UCCUG CAGGA UGAUGGCAUGCACUAUGCGCG ACCUGGCGGA
282 CAGGUGUCCU G CCUGA UCAGG UGAUGGCAUGCACUAUGCGCG AGGACACCUG
286 UGUCCUGCCU G AAGGA UCCUU UGAUGGCAUGCACUAUGCGCG AGGCAGGACA
303 CUGGUGGCCC G AGUGC GCACU UGAUGGCAUGCACUAUGCGCG GGGCCACCAG
307 UGGCCCGAGU G CUGCA UGCAG UGAUGGCAUGCACUAUGCGCG ACUCGGGCCA
310 CCCGAGUGCU G CAGAG CUCUG UGAUGGCAUGCACUAUGCGCG AGCACUCGGG
319 UGCAGAGGCU G UGCGA UCGCA UGAUGGCAUGCACUAUGCGCG AGCCUCUGCA
321 CAGAGGCUGU G CGAGC GCUCG UGAUGGCAUGCACUAUGCGCG ACAGCCUCUG
323 GAGGCUGUGC G AGCGC GCGCU UGAUGGCAUGCACUAUGCGCG GCACAGCCUC
327 CUGUGCGAGC G CGGCG CGCCG UGAUGGCAUGCACUAUGCGCG GCUCGCACAG
332 CGAGCGCGGC G CGAAG CUUCG UGAUGGCAUGCACUAUGCGCG GCCGCGCUCG
334 AGCGCGGCGC G AAGAA UUCUU UGAUGGCAUGCACUAUGCGCG GCGCCGCGCU
343 CGAAGAACGU G CUGGC GCCAG UGAUGGCAUGCACUAUGCGCG ACGUUCUUCG
359 CUUCGGCUUC G CGCUG CAGCG UGAUGGCAUGCACUAUGCGCG GAAGCCGAAG
361 UCGGCUUCGC G CUGCU AGCAG UGAUGGCAUGCACUAUGCGCG GCGAAGCCGA
364 GCUUCGCGCU G CUGGA UCCAG UGAUGGCAUGCACUAUGCGCG AGCGCGAAGC
378 GACGGGGCCC G CGGGG CCCCG UGAUGGCAUGCACUAUGCGCG GGGCCCCGUC
392 GGGCCCCCCC G AGGCC GGCCU UGAUGGCAUGCACUAUGCGCG GGGGGGGCCC
412 CCACCAGCGU G CGCAG CUGCG UGAUGGCAUGCACUAUGCGCG ACGCUGGUGG
414 ACCAGCGUGC G CAGCU AGCUG UGAUGGCAUGCACUAUGCGCG GCACGCUGGU
424 GCAGCUACCU G CCCAA UUGGG UGAUGGCAUGCACUAUGCGCG AGGUAGCUGC
Table 15
436 CCAACACGGU G ACCGA UCGGU UGAUGGCAUGCACUAUGCGCG ACCGUGUUGG
440 CACGGUGACC G ACGCA UGCGU UGAUGGCAUGCACUAUGCGCG GGUCACCGUG
443 GGUGACCGAC G CACUG CAGUG UGAUGGCAUGCACUAUGCGCG GUCGGUCACC
448 CCGACGCACU G CGGGG CCCCG UGAUGGCAUGCACUAUGCGCG AGUGCGUCGG
472 CGUGGGGGCU G CUGCU AGCAG UGAUGGCAUGCACUAUGCGCG AGCCCCCACG
475 GGGGGCUGCU G CUGCG CGCAG UGAUGGCAUGCACUAUGCGCG AGCAGCCCCC
478 GGCUGCUGCU G CGCCG CGGCG UGAUGGCAUGCACUAUGCGCG AGCAGCAGCC
480 CUGCUGCUGC G CCGCG CGCGG UGAUGGCAUGCACUAUGCGCG GCAGCAGCAG
483 CUGCUGCGCC G CGUGG CCACG UGAUGGCAUGCACUAUGCGCG GGCGCAGCAG
491 CCGCGUGGGC G ACGAC GUCGU UGAUGGCAUGCACUAUGCGCG GCCCACGCGG
494 CGUGGGCGAC G ACGUG CACGU UGAUGGCAUGCACUAUGCGCG GUCGCCCACG
499 GCGACGACGU G CUGGU ACCAG UGAUGGCAUGCACUAUGCGCG ACGUCGUCGC
511 UGGUUCACCU G CUGGC GCCAG UGAUGGCAUGCACUAUGCGCG AGGUGAACCA
519 CUGCUGGCAC G CUGCG CGCAG UGAUGGCAUGCACUAUGCGCG GUGCCAGCAG
522 CUGGCACGCU G CGCGC GCGCG UGAUGGCAUGCACUAUGCGCG AGCGUGCCAG
524 GGCACGCUGC G CGCUC GAGCG UGAUGGCAUGCACUAUGCGCG GCAGCGUGCC
526 CACGCUGCGC G CUCUU AAGAG UGAUGGCAUGCACUAUGCGCG GCGCAGCGUG
533 CGCGCUCUUU G UGCUG CAGCA UGAUGGCAUGCACUAUGCGCG AAAGAGCGCG
535 CGCUCUUUGU G CUGGU ACCAG UGAUGGCAUGCACUAUGCGCG ACAAAGAGCG
552 GCUCCCAGCU G CGCCU AGGCG UGAUGGCAUGCACUAUGCGCG AGCUGGGAGC
554 UCCCAGCUGC G CCUAC GUAGG UGAUGGCAUGCACUAUGCGCG GCAGCUGGGA
565 CCUACCAGGU G UGCGG CCGCA UGAUGGCAUGCACUAUGCGCG ACCUGGUAGG
567 UACCAGGUGU G CGGGC GCCCG UGAUGGCAUGCACUAUGCGCG ACACCUGGUA
574 UGUGCGGGCC G CCGCU AGCGG UGAUGGCAUGCACUAUGCGCG GGCCCGCACA
577 GCGGGCCGCC G CUGUA UACAG UGAUGGCAUGCACUAUGCGCG GGCGGCCCGC
580 GGCCGCCGCU G UACCA UGGUA UGAUGGCAUGCACUAUGCGCG AGCGGCGGCC
593 CCAGCUCGGC G CUGCC GGCAG UGAUGGCAUGCACUAUGCGCG GCCGAGCUGG
596 GCUCGGCGCU G CCACU AGUGG UGAUGGCAUGCACUAUGCGCG AGCGCCGAGC
616 CCCGGCCCCC G CCACA UGUGG UGAUGGCAUGCACUAUGCGCG GGGGGCCGGG
623 CCCGCCACAC G CUAGU ACUAG UGAUGGCAUGCACUAUGCGCG GUGUGGCGGG
636 AGUGGACCCC G AAGGC GCCUU UGAUGGCAUGCACUAUGCGCG GGGGUCCACU
1 able 15
651 CGUCUGGGAU G CGAAC GUUCG UGAUGGCAUGCACUAUGCGCG AUCCCAGACG
653 UCUGGGAUGC G AACGG CCGUU UGAUGGCAUGCACUAUGCGCG GCAUCCCAGA
703 CCCUGGGCCU G CCAGC GCUGG UGAUGGCAUGCACUAUGCGCG AGGCCCAGGG
716 AGCCCCGGGU G CGAGG CCUCG UGAUGGCAUGCACUAUGCGCG ACCCGGGGCU
718 CCCCGGGUGC G AGGAG CUCCU UGAUGGCAUGCACUAUGCGCG GCACCCGGGG
726 GCGAGGAGGC G CGGGG CCCCG UGAUGGCAUGCACUAUGCGCG GCCUCCUCGC
737 CGGGGGCAGU G CCAGC GCUGG UGAUGGCAUGCACUAUGCGCG ACUGCCCCCG
744 AGUGCCAGCC G AAGUC GACUU UGAUGGCAUGCACUAUGCGCG GGCUGGCACU
751 GCCGAAGUCU G CCGUU AACGG UGAUGGCAUGCACUAUGCGCG AGACUUCGGC
757 GUCUGCCGUU G CCCAA UUGGG UGAUGGCAUGCACUAUGCGCG AACGGCAGAC
779 CAGGCGUGGC G CUGCC GGCAG UGAUGGCAUGCACUAUGCGCG GCCACGCCUG
782 GCGUGGCGCU G CCCCU AGGGG UGAUGGCAUGCACUAUGCGCG AGCGCCACGC
788 CGCUGCCCCU G AGCCG CGGCU UGAUGGCAUGCACUAUGCGCG AGGGGCAGCG
802 CGGAGCGGAC G CCCGU ACGGG UGAUGGCAUGCACUAUGCGCG GUCCGCUCCG
841 CGGGCAGGAC G CGUGG CCACG UGAUGGCAUGCACUAUGCGCG GUCCUGCCCG
850 CGCGUGGACC G AGUGA UCACU UGAUGGCAUGCACUAUGCGCG GGUCCACGCG
854 UGGACCGAGU G ACCGU ACGGU UGAUGGCAUGCACUAUGCGCG ACUCGGUCCA
867 CGUGGUUUCU G UGUGG CCACA UGAUGGCAUGCACUAUGCGCG AGAAACCACG
869 UGGUUUCUGU G UGGUG CACCA UGAUGGCAUGCACUAUGCGCG ACAGAAACCA
874 UCUGUGUGGU G UCACC GGUGA UGAUGGCAUGCACUAUGCGCG ACCACACAGA
881 GGUGUCACCU G CCAGA UCUGG UGAUGGCAUGCACUAUGCGCG AGGUGACACC
890 UGCCAGACCC G CCGAA UUCGG UGAUGGCAUGCACUAUGCGCG GGGUCUGGCA
893 CAGACCCGCC G AAGAA UUCUU UGAUGGCAUGCACUAUGCGCG GGCGGGUCUG
917 UUUGGAGGGU G CGCUC GAGCG UGAUGGCAUGCACUAUGCGCG ACCCUCCAAA
919 UGGAGGGUGC G CUCUC GAGAG UGAUGGCAUGCACUAUGCGCG GCACCCUCCA
931 UCUCUGGCAC G CGCCA UGGCG UGAUGGCAUGCACUAUGCGCG GUGCCAGAGA
933 UCUGGCACGC G CCACU AGUGG UGAUGGCAUGCACUAUGCGCG GCGUGCCAGA
957 UCCGUGGGCC G CCAGC GCUGG UGAUGGCAUGCACUAUGCGCG GGCCCACGGA
968 CCAGCACCAC G CGGGC GCCCG UGAUGGCAUGCACUAUGCGCG GUGGUGCUGG
988 CAUCCACAUC G CGGCC GGCCG UGAUGGCAUGCACUAUGCGCG GAUGUGGAUG
1012 CCUGGGACAC G CCUUG CAAGG UGAUGGCAUGCACUAUGCGCG GUGUCCCAGG
Table 15
1017 GACACGCCUU G UCCCC GGGGA UGAUGGCAUGCACUAUGCGCG AAGGCGUGUC
1027 GUCCCCCGGU G UACGC GCGUA UGAUGGCAUGCACUAUGCGCG ACCGGGGGAC
1031 CCCGGUGUAC G CCGAG CUCGG UGAUGGCAUGCACUAUGCGCG GUACACCGGG
1034 GGUGUACGCC G AGACC GGUCU UGAUGGCAUGCACUAUGCGCG GGCGUACACC
1064 CUCCUCAGGC G ACAAG CUUGU UGAUGGCAUGCACUAUGCGCG GCCUGAGGAG
1078 AGGAGCAGCU G CGGCC GGCCG UGAUGGCAUGCACUAUGCGCG AGCUGCUCCU
1105 UCAGCUCUCU G AGGCC GGCCU UGAUGGCAUGCACUAUGCGCG AGAGAGCUGA
1117 GGCCCAGCCU G ACUGG CCAGU UGAUGGCAUGCACUAUGCGCG AGGCUGGGCC
1124 CCUGACUGGC G CUCGG CCGAG UGAUGGCAUGCACUAUGCGCG GCCAGUCAGG
1171 GGCCCUGGAU G CCAGG CCUGG UGAUGGCAUGCACUAUGCGCG AUCCAGGGCC
1185 GGGACUCCCC G CAGGU ACCUG UGAUGGCAUGCACUAUGCGCG GGGGAGUCCC
1192 CCCGCAGGUU G CCCCG CGGGG UGAUGGCAUGCACUAUGCGCG AACCUGCGGG
1197 AGGUUGCCCC G CCUGC GCAGG UGAUGGCAUGCACUAUGCGCG GGGGCAACCU
1201 UGCCCCGCCU G CCCCA UGGGG UGAUGGCAUGCACUAUGCGCG AGGCGGGGCA
1209 CUGCCCCAGC G CUACU AGUAG UGAUGGCAUGCACUAUGCGCG GCUGGGGCAG
1222 ACUGGCAAAU G CGGCC GGCCG UGAUGGCAUGCACUAUGCGCG AUUUGCCAGU
1231 UGCGGCCCCU G UUUCU AGAAA UGAUGGCAUGCACUAUGCGCG AGGGGCCGCA
1243 UUCUGGAGCU G CUUGG CCAAG UGAUGGCAUGCACUAUGCGCG AGCUCCAGAA
1256 UGGGAACCAC G CGCAG CUGCG UGAUGGCAUGCACUAUGCGCG GUGGUUCCCA
1258 GGAACCACGC G CAGUG CACUG UGAUGGCAUGCACUAUGCGCG GCGUGGUUCC
1263 CACGCGCAGU G CCCCU AGGGG UGAUGGCAUGCACUAUGCGCG ACUGCGCGUG
1276 CCUACGGGGU G CUCCU AGGAG UGAUGGCAUGCACUAUGCGCG ACCCCGUAGG
1288 UCCUCAAGAC G CACUG CAGUG UGAUGGCAUGCACUAUGCGCG GUCUUGAGGA
1293 AAGACGCACU G CCCGC GCGGG UGAUGGCAUGCACUAUGCGCG AGUGCGUCUU
1297 CGCACUGCCC G CUGCG CGCAG UGAUGGCAUGCACUAUGCGCG GGGCAGUGCG
1300 ACUGCCCGCU G CGAGC GCUCG UGAUGGCAUGCACUAUGCGCG AGCGGGCAGU
1302 UGCCCGCUGC G AGCUG CAGCU UGAUGGCAUGCACUAUGCGCG GCAGCGGGCA
1307 GCUGCGAGCU G CGGUC GACCG UGAUGGCAUGCACUAUGCGCG AGCUCGCAGC
1328 AGCAGCCGGU G UCUGU ACAGA UGAUGGCAUGCACUAUGCGCG ACCGGCUGCU
1332 GCCGGUGUCU G UGCCC GGGCA UGAUGGCAUGCACUAUGCGCG AGACACCGGC
1334 CGGUGUCUGU G CCCGG CCGGG UGAUGGCAUGCACUAUGCGCG ACAGACACCG
Table 15
1358 CCAGGGCUCU G UGGCG CGCCA UGAUGGCAUGCACUAUGCGCG AGAGCCCUGG
1370 GGCGGCCCCC G AGGAG CUCCU UGAUGGCAUGCACUAUGCGCG GGGGGCCGCC
1395 GACCCCCGUC G CCUGG CCAGG UGAUGGCAUGCACUAUGCGCG GACGGGGGUC
1402 GUCGCCUGGU G CAGCU AGCUG UGAUGGCAUGCACUAUGCGCG ACCAGGCGAC
1408 UGGUGCAGCU G CUCCG CGGAG UGAUGGCAUGCACUAUGCGCG AGCUGCACCA
1413 CAGCUGCUCC G CCAGC GCUGG UGAUGGCAUGCACUAUGCGCG GGAGCAGCUG
1438 CCUGGCAGGU G UACGG CCGUA UGAUGGCAUGCACUAUGCGCG ACCUGCCAGG
1450 ACGGCUUCGU G CGGGC GCCCG UGAUGGCAUGCACUAUGCGCG ACGAAGCCGU
1458 GUGCGGGCCU G CCUGC GCAGG UGAUGGCAUGCACUAUGCGCG AGGCCCGCAC
1462 GGGCCUGCCU G CGCCG CGGCG UGAUGGCAUGCACUAUGCGCG AGGCAGGCCC
1464 GCCUGCCUGC G CCGGC GCCGG UGAUGGCAUGCACUAUGCGCG GCAGGCAGGC
1474 GCCGGCUGGU G CCCCC GGGGG UGAUGGCAUGCACUAUGCGCG ACCAGCCGGC
1505 CAGGCACAAC G AACGC GCGUU UGAUGGCAUGCACUAUGCGCG GUUGUGCCUG
1509 CACAACGAAC G CCGCU AGCGG UGAUGGCAUGCACUAUGCGCG GUUCGUUGUG
1512 AACGAACGCC G CUUCC GGAAG UGAUGGCAUGCACUAUGCGCG GGCGUUCGUU
1556 GGGGAAGCAU G CCAAG CUUGG UGAUGGCAUGCACUAUGCGCG AUGCUUCCCC
1567 CCAAGCUCUC G CUGCA UGCAG UGAUGGCAUGCACUAUGCGCG GAGAGCUUGG
1570 AGCUCUCGCU G CAGGA UCCUG UGAUGGCAUGCACUAUGCGCG AGCGAGAGCU
1579 UGCAGGAGCU G ACGUG CACGU UGAUGGCAUGCACUAUGCGCG AGCUCCUGCA
1591 CGUGGAAGAU G AGCGU ACGCU UGAUGGCAUGCACUAUGCGCG AUCUUCCACG
1597 AGAUGAGCGU G CGGGA UCCCG UGAUGGCAUGCACUAUGCGCG ACGCUCAUCU
1605 GUGCGGGACU G CGCUU AAGCG UGAUGGCAUGCACUAUGCGCG AGUCCCGCAC
1607 GCGGGACUGC G CUUGG CCAAG UGAUGGCAUGCACUAUGCGCG GCAGUCCCGC
1615 GCGCUUGGCU G CGCAG CUGCG UGAUGGCAUGCACUAUGCGCG AGCCAAGCGC
1617 GCUUGGCUGC G CAGGA UCCUG UGAUGGCAUGCACUAUGCGCG GCAGCCAAGC
1638 GGGGUUGGCU G UGUUC GAACA UGAUGGCAUGCACUAUGCGCG AGCCAACCCC
1640 GGUUGGCUGU G UUCCG CGGAA UGAUGGCAUGCACUAUGCGCG ACAGCCAACC
1649 UGUUCCGGCC G CAGAG CUCUG UGAUGGCAUGCACUAUGCGCG GGCCGGAACA
1663 AGCACCGUCU G CGUGA UCACG UGAUGGCAUGCACUAUGCGCG AGACGGUGCU
1667 CCGUCUGCGU G AGGAG CUCCU UGAUGGCAUGCACUAUGCGCG ACGCAGACGG
1690 CCAAGUUCCU G CACUG CAGUG UGAUGGCAUGCACUAUGCGCG AGGAACUUGG
Table 15
1699 UGCACUGGCU G AUGAG CUCAU UGAUGGCAUGCACUAUGCGCG AGCCAGUGCA
1702 ACUGGCUGAU G AGUGU ACACU UGAUGGCAUGCACUAUGCGCG AUCAGCCAGU
1706 GCUGAUGAGU G UGUAC GUACA UGAUGGCAUGCACUAUGCGCG ACUCAUCAGC
1708 UGAUGAGUGU G UACGU ACGUA UGAUGGCAUGCACUAUGCGCG ACACUCAUCA
1718 GUACGUCGUC G AGCUG CAGCU UGAUGGCAUGCACUAUGCGCG GACGACGUAC
1723 UCGUCGAGCU G CUCAG CUGAG UGAUGGCAUGCACUAUGCGCG AGCUCGACGA
1742 UUUCUUUUAU G UCACG CGUGA UGAUGGCAUGCACUAUGCGCG AUAAAAGAAA
1793 CCGGAAGAGU G UCUGG CCAGA UGAUGGCAUGCACUAUGCGCG ACUCUUCCGG
1807 GGAGCAAGUU G CAAAG CUUUG UGAUGGCAUGCACUAUGCGCG AACUUGCUCC
1834 GACAGCACUU G AAGAG CUCUU UGAUGGCAUGCACUAUGCGCG AAGUGCUGUC
1843 UGAAGAGGGU G CAGCU AGCUG UGAUGGCAUGCACUAUGCGCG ACCCUCUUCA
1849 GGGUGCAGCU G CGGGA UCCCG UGAUGGCAUGCACUAUGCGCG AGCUGCACCC
1858 UGCGGGAGCU G UCGGA UCCGA UGAUGGCAUGCACUAUGCGCG AGCUCCCGCA
1898 AGCCAGGCCC G CCCUG CAGGG UGAUGGCAUGCACUAUGCGCG GGGCCUGGCU
1903 GGCCCGCCCU G CUGAC GUCAG UGAUGGCAUGCACUAUGCGCG AGGGCGGGCC
1906 CCGCCCUGCU G ACGUC GACGU UGAUGGCAUGCACUAUGCGCG AGCAGGGCGG
1920 UCCAGACUCC G CUUCA UGAAG UGAUGGCAUGCACUAUGCGCG GGAGUCUGGA
1937 CCCCAAGCCU G ACGGG CCCGU UGAUGGCAUGCACUAUGCGCG AGGCUUGGGG
1945 CUGACGGGCU G CGGCC GGCCG UGAUGGCAUGCACUAUGCGCG AGCCCGUCAG
1951 GGCUGCGGCC G AUUGU ACAAU UGAUGGCAUGCACUAUGCGCG GGCCGCAGCC
1955 GCGGCCGAUU G UGAAC GUUCA UGAUGGCAUGCACUAUGCGCG AAUCGGCCGC
1957 GGCCGAUUGU G AACAU AUGUU UGAUGGCAUGCACUAUGCGCG ACAAUCGGCC
1992 AGAACGUUCC G CAGAG CUCUG UGAUGGCAUGCACUAUGCGCG GGAACGUUCU
2009 AAAGAGGGCC G AGCGU ACGCU UGAUGGCAUGCACUAUGCGCG GGCCCUCUUU
2023 GUCUCACCUC G AGGGU ACCCU UGAUGGCAUGCACUAUGCGCG GAGGUGAGAC
2029 CCUCGAGGGU G AAGGC GCCUU UGAUGGCAUGCACUAUGCGCG ACCCUCGAGG
2038 UGAAGGCACU G UUCAG CUGAA UGAUGGCAUGCACUAUGCGCG AGUGCCUUCA
2047 UGUUCAGCGU G CUCAA UUGAG UGAUGGCAUGCACUAUGCGCG ACGCUGAACA
2057 GCUCAACUAC G AGCGG CCGCU UGAUGGCAUGCACUAUGCGCG GUAGUUGAGC
2065 ACGAGCGGGC G CGGCG CGCCG UGAUGGCAUGCACUAUGCGCG GCCCGCUCGU
2070 CGGGCGCGGC G CCCCG CGGGG UGAUGGCAUGCACUAUGCGCG GCCGCGCCCG
Table 15
2087 CCUCCUGGGC G CCUCU AGAGG UGAUGGCAUGCACUAUGCGCG GCCCAGGAGG
2093 GGGCGCCUCU G UGCUG CAGCA UGAUGGCAUGCACUAUGCGCG AGAGGCGCCC
2095 GCGCCUCUGU G CUGGG CCCAG UGAUGGCAUGCACUAUGCGCG ACAGAGGCGC
2108 GGGCCUGGAC G AUAUC GAUAU UGAUGGCAUGCACUAUGCGCG GUCCAGGCCC
2127 AGGGCCUGGC G CACCU AGGUG UGAUGGCAUGCACUAUGCGCG GCCAGGCCCU
2137 GCACCUUCGU G CUGCG CGCAG UGAUGGCAUGCACUAUGCGCG ACGAAGGUGC
2140 CCUUCGUGCU G CGUGU ACACG UGAUGGCAUGCACUAUGCGCG AGCACGAAGG
2144 CGUGCUGCGU G UGCGG CCGCA UGAUGGCAUGCACUAUGCGCG ACGCAGCACG
2146 UGCUGCGUGU G CGGGC GCCCG UGAUGGCAUGCACUAUGCGCG ACACGCAGCA
2161 CCCAGGACCC G CCGCC GGCGG UGAUGGCAUGCACUAUGCGCG GGGUCCUGGG
2164 AGGACCCGCC G CCUGA UCAGG UGAUGGCAUGCACUAUGCGCG GGCGGGUCCU
2168 CCCGCCGCCU G AGCUG CAGCU UGAUGGCAUGCACUAUGCGCG AGGCGGCGGG
2173 CGCCUGAGCU G UACUU AAGUA UGAUGGCAUGCACUAUGCGCG AGCUCAGGCG
2180 GCUGUACUUU G UCAAG CUUGA UGAUGGCAUGCACUAUGCGCG AAAGUACAGC
2192 CAAGGUGGAU G UGACG CGUCA UGAUGGCAUGCACUAUGCGCG AUCCACCUUG
2194 AGGUGGAUGU G ACGGG CCCGU UGAUGGCAUGCACUAUGCGCG ACAUCCACCU
2201 UGUGACGGGC G CGUAC GUACG UGAUGGCAUGCACUAUGCGCG GCCCGUCACA
2207 GGGCGCGUAC G ACACC GGUGU UGAUGGCAUGCACUAUGCGCG GUACGCGCCC
2243 GGAGGUCAUC G CCAGC GCUGG UGAUGGCAUGCACUAUGCGCG GAUGACCUCC
2274 AACACGUACU G CGUGC GCACG UGAUGGCAUGCACUAUGCGCG AGUACGUGUU
2278 CGUACUGCGU G CGUCG CGACG UGAUGGCAUGCACUAUGCGCG ACGCAGUACG
2288 GCGUCGGUAU G CCGUG CACGG UGAUGGCAUGCACUAUGCGCG AUACCGACGC
2306 CCAGAAGGCC G CCCAU AUGGG UGAUGGCAUGCACUAUGCGCG GGCCUUCUGG
2322 GGGCACGUCC G CAAGG CCUUG UGAUGGCAUGCACUAUGCGCG GGACGUGCCC
2353 UCUCUACCUU G ACAGA UCUGU UGAUGGCAUGCACUAUGCGCG AAGGUAGAGA
2374 AGCCGUACAU G CGACA UGUCG UGAUGGCAUGCACUAUGCGCG AUGUACGGCU
2376 CCGUACAUGC G ACAGU ACUGU UGAUGGCAUGCACUAUGCGCG GCAUGUACGG
2395 UGGCUCACCU G CAGGA UCCUG UGAUGGCAUGCACUAUGCGCG AGGUGAGCCA
2410 AGACCAGCCC G CUGAG CUCAG UGAUGGCAUGCACUAUGCGCG GGGCUGGUCU
2413 CCAGCCCGCU G AGGGA UCCCU UGAUGGCAUGCACUAUGCGCG AGCGGGCUGG
2420 GCUGAGGGAU G CCGUC GACGG UGAUGGCAUGCACUAUGCGCG AUCCCUCAGC
Table 15
2432 CGUCGUCAUC G AGCAG CUGCU UGAUGGCAUGCACUAUGCGCG GAUGACGACG
2449 GCUCCUCCCU G AAUGA UCAUU UGAUGGCAUGCACUAUGCGCG AGGGAGGAGC
2453 CUCCCUGAAU G AGGCC GGCCU UGAUGGCAUGCACUAUGCGCG AUUCAGGGAG
2474 UGGCCUCUUC G ACGUC GACGU UGAUGGCAUGCACUAUGCGCG GAAGAGGCCA
2487 GUCUUCCUAC G CUUCA UGAAG UGAUGGCAUGCACUAUGCGCG GUAGGAAGAC
2494 UACGCUUCAU G UGCCA UGGCA UGAUGGCAUGCACUAUGCGCG AUGAAGGGUA
2496 CGCUUCAUGU G CCACC GGUGG UGAUGGCAUGCACUAUGCGCG ACAUGAAGCG
2504 GUGCCACCAC G CCGUG CACGG UGAUGGCAUGCACUAUGCGCG GUGGUGGCAC
2509 ACCACGCCGU G CGCAU AUGCG UGAUGGCAUGCACUAUGCGCG ACGGCGUGGU
2511 CACGCCGUGC G CAUCA UGAUG UGAUGGCAUGCACUAUGCGCG GCACGGCGUG
2538 UACGUCCAGU G CCAGG CCUGG UGAUGGCAUGCACUAUGCGCG ACUGGACGUA
2551 AGGGGAUCCC G CAGGG CCCUG UGAUGGCAUGCACUAUGCGCG GGGAUCCCCU
2572 UCCUCUCCAC G CUGCU AGCAG UGAUGGCAUGCACUAUGCGCG GUGGAGAGGA
2575 UCUCCACGCU G CUCUG CAGAG UGAUGGCAUGCACUAUGCGCG AGCGUGGAGA
2580 ACGCUGCUCU G CAGCC GGCUG UGAUGGCAUGCACUAUGCGCG AGAGCAGCGU
2587 UCUGCAGCCU G UGCUA UAGCA UGAUGGCAUGCACUAUGCGCG AGGCUGCAGA
2589 UGCAGCCUGU G CUACG CGUAG UGAUGGCAUGCACUAUGCGCG ACAGGCUGCA
2597 GUGCUACGGC G ACAUG CAUGU UGAUGGCAUGCACUAUGCGCG GCCGUAGCAC
2614 AGAACAAGCU G UUUGC GCAAA UGAUGGCAUGCACUAUGCGCG AGCUUGUUCU
2618 CAAGCUGUUU G CGGGG CCCCG UGAUGGCAUGCACUAUGCGCG AAACAGCUUG
2641 GGGACGGGCU G CUCCU AGGAG UGAUGGCAUGCACUAUGCGCG AGCCCGUCCC
2647 GGCUGCUCCU G CGUUU AAACG UGAUGGCAUGCACUAUGCGCG AGGAGCAGCC
2660 UUUGGUGGAU G AUUUC GAAAU UGAUGGCAUGCACUAUGCGCG AUCCACCAAA
2668 AUGAUUUCUU G UUGGU ACCAA UGAUGGCAUGCACUAUGCGCG AAGAAAUCAU
2674 UCUUGUUGGU G ACACC GGUGU UGAUGGCAUGCACUAUGCGCG ACCAACAAGA
2693 CCUCACCCAC G CGAAA UUUCG UGAUGGCAUGCACUAUGCGCG GUGGGUGAGG
2695 UCACCCACGC G AAAAC GUUUU UGAUGGCAUGCACUAUGCGCG GCGUGGGUGA
2721 ACCCUGGUCC G AGGUG CACCU UGAUGGCAUGCACUAUGCGCG GGACCAGGGU
2726 GGUCCGAGGU G UCCCU AGGGA UGAUGGCAUGCACUAUGCGCG ACCUCGGACC
2732 AGGUGUCCCU G AGUAU AUACU UGAUGGCAUGCACUAUGCGCG AGGGACACCU
2742 GAGUAUGGCU G CGUGG CCACG UGAUGGCAUGCACUAUGCGCG AGCCAUACUC
Table 15
2749 GCUGCGUGGU G AACUU AAGUU UGAUGGCAUGCACUAUGCGCG ACCACGCAGC
2755 UGGUGAACUU G CGGAA UUCCG UGAUGGCAUGCACUAUGCGCG AAGUUCACCA
2770 AGACAGUGGU G AACUU AAGUU UGAUGGCAUGCACUAUGCGCG ACCACUGUCU
2780 GAACUUCCCU G UAGAA UUCUA UGAUGGCAUGCACUAUGCGCG AGGGAAGUUC
2789 UGUAGAAGAC G AGGCC GGCCU UGAUGGCAUGCACUAUGCGCG GUCUUCUACA
2813 CACGGCUUUU G UUCAG CUGAA UGAUGGCAUGCACUAUGCGCG AAAAGCCGUG
2821 UUGUUCAGAU G CCGGC GCCGG UGAUGGCAUGCACUAUGCGCG AUCUGAACAA
2847 UUCCCCUGGU G CGGCC GGCCG UGAUGGCAUGCACUAUGCGCG ACCAGGGGAA
2854 GGUGCGGCCU G CUGCU AGCAG UGAUGGCAUGCACUAUGCGCG AGGCCGCACC
2857 GCGGCCUGCU G CUGGA UCCAG UGAUGGCAUGCACUAUGCGCG AGCAGGCCGC
2881 CCCUGGAGGU G CAGAG CUCUG UGAUGGCAUGCACUAUGCGCG ACCUCCAGGG
2888 GGUGCAGAGC G ACUAC GUAGU UGAUGGCAUGCACUAUGCGCG GCUCUGCACC
2903 CUCCAGCUAU G CCCGG CCGGG UGAUGGCAUGCACUAUGCGCG AUAGCUGGAG
2940 ACCUUCAACC G CGGCU AGCCG UGAUGGCAUGCACUAUGCGCG GGUUGAAGGU
2965 GGAGGAACAU G CGUCG CGACG UGAUGGCAUGCACUAUGCGCG AUGUUCCUCC
2970 AACAUGCGUC G CAAAC GUUUG UGAUGGCAUGCACUAUGCGCG GACGCAUGUU
2989 UUGGGGUCUU G CGGCU AGCCG UGAUGGCAUGCACUAUGCGCG AAGACCCCAA
2995 UCUUGCGGCU G AAGUG CACUU UGAUGGCAUGCACUAUGCGCG AGCCGCAAGA
3000 CGGCUGAAGU G UCACA UGUGA UGAUGGCAUGCACUAUGCGCG ACUUCAGCCG
3010 GUCACAGCCU G UUUCU AGAAA UGAUGGCAUGCACUAUGCGCG AGGCUGUGAC
3022 UUCUGGAUUU G CAGGU ACCUG UGAUGGCAUGCACUAUGCGCG AAAUCCAGAA
3028 AUUUGCAGGU G AACAG CUGUU UGAUGGCAUGCACUAUGCGCG ACCUGCAAAU
3046 UCCAGACGGU G UGCAC GUGCA UGAUGGCAUGCACUAUGCGCG ACCGUCUGGA
3048 CAGACGGUGU G CACCA UGGUG UGAUGGCAUGCACUAUGCGCG ACACCGUCUG
3073 AGAUCCUCCU G CUGCA UGCAG UGAUGGCAUGCACUAUGCGCG AGGAGGAUCU
3076 UCCUCCUGCU G CAGGC GCCUG UGAUGGCAUGCACUAUGCGCG AGCAGGAGGA
3095 CAGGUUUCAC G CAUGU ACAUG UGAUGGCAUGCACUAUGCGCG GUGAAACCUG
3099 UUUCACGCAU G UGUGC GCACA UGAUGGCAUGCACUAUGCGCG AUGCGUGAAA
3101 UCACGCAUGU G UGCUG CAGCA UGAUGGCAUGCACUAUGCGCG ACAUGCGUGA
3103 ACGCAUGUGU G CUGCA UGCAG UGAUGGCAUGCACUAUGCGCG ACACAUGCGU
3106 CAUGUGUGCU G CAGCU AGCUG UGAUGGCAUGCACUAUGCGCG AGCACACAUG
Table 15
3154 CAUUUUUCCU G CGCGU ACGCG UGAUGGCAUGCACUAUGCGCG AGGAAAAAUG
3156 UUUUUCCUGC G CGUCA UGACG UGAUGGCAUGCACUAUGCGCG GCAGGAAAAA
3167 CGUCAUCUCU G ACACG CGUGU UGAUGGCAUGCACUAUGCGCG AGAGAUGACG
3183 GCCUCCCUCU G CUACU AGUAG UGAUGGCAUGCACUAUGCGCG AGAGGGAGGC
3196 ACUCCAUCCU G AAAGC GCUUU UGAUGGCAUGCACUAUGCGCG AGGAUGGAGU
3209 AGCCAAGAAC G CAGGG CCCUG UGAUGGCAUGCACUAUGCGCG GUUCUUGGCU
3217 ACGCAGGGAU G UCGCU AGCGA UGAUGGCAUGCACUAUGCGCG AUCCCUGCGU
3220 CAGGGAUGUC G CUGGG CCCAG UGAUGGCAUGCACUAUGCGCG GACAUCCCUG
3236 GGCCAAGGGC G CCGCC GGCGG UGAUGGCAUGCACUAUGCGCG GCCCUUGGCC
3239 CAAGGGCGCC G CCGGC GCCGG UGAUGGCAUGCACUAUGCGCG GGCGCCCUUG
3250 CCGGCCCUCU G CCCUC GAGGG UGAUGGCAUGCACUAUGCGCG AGAGGGCCGG
3257 UCUGCCCUCC G AGGCC GGCCU UGAUGGCAUGCACUAUGCGCG GGAGGGCAGA
3265 CCGAGGCCGU G CAGUG CACUG UGAUGGCAUGCACUAUGCGCG ACGGCCUCGG
3274 UGCAGUGGCU G UGCCA UGGCA UGAUGGCAUGCACUAUGCGCG AGCCACUGCA
3276 CAGUGGCUGU G CCACC GGUGG UGAUGGCAUGCACUAUGCGCG ACAGCCACUG
3292 AAGCAUUCCU G CUCAA UUGAG UGAUGGCAUGCACUAUGCGCG AGGAAUGCUU
3301 UGCUCAAGCU G ACUCG CGAGU UGAUGGCAUGCACUAUGCGCG AGCUUGAGCA
3306 AAGCUGACUC G ACACC GGUGU UGAUGGCAUGCACUAUGCGCG GAGUCAGCUU
3314 UCGACACCGU G UCACC GGUGA UGAUGGCAUGCACUAUGCGCG ACGGUGUCGA
3325 UCACCUACGU G CCACU AGUGG UGAUGGCAUGCACUAUGCGCG ACGUAGGUGA
3358 CAGCCCAGAC G CAGCU AGCUG UGAUGGCAUGCACUAUGCGCG GUCUGGGCUG
3364 AGACGCAGCU G AGUCG CGACU UGAUGGCAUGCACUAUGCGCG AGCUGCGUCU
3385 UCCCGGGGAC G ACGCU AGCGU UGAUGGCAUGCACUAUGCGCG GUCCCCGGGA
3388 CGGGGACGAC G CUGAC GUCAG UGAUGGCAUGCACUAUGCGCG GUCGUCCCCG
3391 GGACGACGCU G ACUGC GCAGU UGAUGGCAUGCACUAUGCGCG AGCGUCGUCC
3395 GACGCUGACU G CCCUG CAGGG UGAUGGCAUGCACUAUGCGCG AGUCAGCGUC
3407 CCUGGAGGCC G CAGCC GGCUG UGAUGGCAUGCACUAUGCGCG GGCCUCCAGG
3424 ACCCGGCACU G CCCUC GAGGG UGAUGGCAUGCACUAUGCGCG AGUGCCGGGU
3453 AUCCUGGACU G AUGGC GCCAU UGAUGGCAUGCACUAUGCGCG AGUCCAGGAU
3464 AUGGCCACCC G CCCAC GUGGG UGAUGGCAUGCACUAUGCGCG GGGUGGCCAU
3479 CAGCCAGGCC G AGAGC GCUCU UGAUGGCAUGCACUAUGCGCG GGCCUGGCUG
Table 15
3501 CAGCAGCCCU G UCACG CGUGA UGAUGGCAUGCACUAUGCGCG AGGGCUGCUG
3506 GCCCUGUCAC G CCGGG CCCGG UGAUGGCAUGCACUAUGCGCG GUGACAGGGC
3554 ACCCAGGCCC G CACCG CGGUG UGAUGGCAUGCACUAUGCGCG GGGCCUGGGU
3559 GGCCCGCACC G CUGGG CCCAG UGAUGGCAUGCACUAUGCGCG GGUGCGGGCC
3570 CUGGGAGUCU G AGGCC GGCCU UGAUGGCAUGCACUAUGCGCG AGACUCCCAG
3577 UCUGAGGCCU G AGUGA UCACU UGAUGGCAUGCACUAUGCGCG AGGCCUCAGA
3581 AGGCCUGAGU G AGUGU ACACU UGAUGGCAUGCACUAUGCGCG ACUCAGGCCU
3585 CUGAGUGAGU G UUUGG CCAAA UGAUGGCAUGCACUAUGCGCG ACUCACUCAG
3593 GUGUUUGGCC G AGGCC GGCCU UGAUGGCAUGCACUAUGCGCG GGCCAAACAC
3600 GCCGAGGCCU G CAUGU ACAUG UGAUGGCAUGCACUAUGCGCG AGGCCUCGGC
3604 AGGCCUGCAU G UCCGG CCGGA UGAUGGCAUGCACUAUGCGCG AUGCAGGCCU
3612 AUGUCCGGCU G AAGGC GCCUU UGAUGGCAUGCACUAUGCGCG AGCCGGACAU
3619 GCUGAAGGCU G AGUGU ACACU UGAUGGCAUGCACUAUGCGCG AGCCUUCAGC
3623 AAGGCUGAGU G UCCGG CCGGA UGAUGGCAUGCACUAUGCGCG ACUCAGCCUU
3631 GUGUCCGGCU G AGGCC GGCCU UGAUGGCAUGCACUAUGCGCG AGCCGGACAC
3638 GCUGAGGCCU G AGCGA UCGCU UGAUGGCAUGCACUAUGCGCG AGGCCUCAGC
3642 AGGCCUGAGC G AGUGU ACACU UGAUGGCAUGCACUAUGCGCG GCUCAGGCCU
3646 CUGAGCGAGU G UCCAG CUGGA UGAUGGCAUGCACUAUGCGCG ACUCGCUCAG
3661 GCCAAGGGCU G AGUGU ACACU UGAUGGCAUGCACUAUGCGCG AGCCCUUGGC
3665 AGGGCUGAGU G UCCAG CUGGA UGAUGGCAUGCACUAUGCGCG ACUCAGCCCU
3678 CAGCACACCU G CCGUC GACGG UGAUGGCAUGCACUAUGCGCG AGGUGUGCUG
3705 ACAGGCUGGC G CUCGG CCGAG UGAUGGCAUGCACUAUGCGCG GCCAGCCUGU
3789 CCCCAGAUUC G CCAUU AAUGG UGAUGGCAUGCACUAUGCGCG GAAUCUGGGG
3795 AUUCGCCAUU G UUCAC GUGAA UGAUGGCAUGCACUAUGCGCG AAUGGCGAAU
3806 UUCACCCCUC G CCCUG CAGGG UGAUGGCAUGCACUAUGCGCG GAGGGGUGAA
3811 CCCUCGCCCU G CCCUC GAGGG UGAUGGCAUGCACUAUGCGCG AGGGCGAGGG
3821 GCCCUCCUUU G CCUUC GAAGG UGAUGGCAUGCACUAUGCGCG AAAGGAGGGC
3854 UGGAGACCCU G AGAAG CUUCU UGAUGGCAUGCACUAUGCGCG AGGGUCUCCA
3888 AAUUUGGAGU G ACCAA UUGGU UGAUGGCAUGCACUAUGCGCG ACUCCAAAUU
3898 GACCAAAGGU G UGCCC GGGCA UGAUGGCAUGCACUAUGCGCG ACCUUUGGUC
3900 CCAAAGGUGU G CCCUG CAGGG UGAUGGCAUGCACUAUGCGCG ACACCUUUGG
Table 15
3905 GGUGUGCCCU G UACAC GUGUA UGAUGGCAUGCACUAUGCGCG AGGGCACACC
3915 GUACACAGGC G AGGAC GUCCU UGAUGGCAUGCACUAUGCGCG GCCUGUGUAC
3924 CGAGGACCCU G CACCU AGGUG UGAUGGCAUGCACUAUGCGCG AGGGUCCUCG
3944 GGGGGUCCCU G UGGGU ACCCA UGAUGGCAUGCACUAUGCGCG AGGGACCCCC
3966 GGGGGGAGGU G CUGUG CACAG UGAUGGCAUGCACUAUGCGCG ACCUCCCCCC
3969 GGGAGGUGCU G UGGGA UCCCA UGAUGGCAUGCACUAUGCGCG AGCACCUCCC
3985 GUAAAAUACU G AAUAU AUAUU UGAUGGCAUGCACUAUGCGCG AGUAUUUUAC
3993 CUGAAUAUAU G AGUUU AAACU UGAUGGCAUGCACUAUGCGCG AUAUAUUCAG
4008 UUUCAGUUUU G AAAAA UUUUU UGAUGGCAUGCACUAUGCGCG AAAACUGAAA
Seq l = TERT (Homo sapiens telomerase reverse transcriptase (TERT) mRNA, 4015 bp); Nakamura et al., Science 277 (5328), 955-959 (1997)
Input Sequence = TERT. Cut Site = YG/M or UG/U.
Stem Length = 5/10. Core Sequence = UGAUG GCAUGCACUAUGC GCG
Table 16
Table 16: Human telomerase reverse transcriptase (TERT) DNAzyme and Target Sequence
Table 16
105 GCGGTAGT GGCTAGCTACAACGA GGCTGCGC GCGCAGCC A ACTACCGC
108 CTCGCGGT GGCTAGCTACAACGA AGTGGCTG CAGCCACT A ACCGCGAG
111 CACCTCGC GGCTAGCTACAACGA GGTAGTGG CCACTACC G GCGAGGTG
116 GGCAGCAC GGCTAGCTACAACGA CTCGCGGT ACCGCGAG G GTGCTGCC
118 GCGGCAGC GGCTAGCTACAACGA ACCTCGCG CGCGAGGT G GCTGCCGC
121 CCAGCGGC GGCTAGCTACAACGA AGCACCTC GAGGTGCT G GCCGCTGG
124 TGGCCAGC GGCTAGCTACAACGA GGCAGCAC GTGCTGCC G GCTGGCCA
128 AACGTGGC GGCTAGCTACAACGA CAGCGGCA TGCCGCTG G GCCACGTT
131 ACGAACGT GGCTAGCTACAACGA GGCCAGCG CGCTGGCC A ACGTTCGT
133 GCACGAAC GGCTAGCTACAACGA GTGGCCAG CTGGCCAC G GTTCGTGC
137 CGCCGCAC GGCTAGCTACAACGA GAACGTGG CCACGTTC G GTGCGGCG
139 GGCGCCGC GGCTAGCTACAACGA ACGAACGT ACGTTCGT G GCGGCGCC
142 CCAGGCGC GGCTAGCTACAACGA CGCACGAA TTCGTGCG G GCGCCTGG
144 CCCCAGGC GGCTAGCTACAACGA GCCGCACG CGTGCGGC G GCCTGGGG
151 CCTGGGGC GGCTAGCTACAACGA CCCAGGCG CGCCTGGG G GCCCCAGG
159 CCGCCAGC GGCTAGCTACAACGA CCTGGGGC GCCCCAGG G GCTGGCGG
163 CCAGCCGC GGCTAGCTACAACGA CAGCCCTG CAGGGCTG G GCGGCTGG
166 GCACCAGC GGCTAGCTACAACGA CGCCAGCC GGCTGGCG G GCTGGTGC
170 CGCTGCAC GGCTAGCTACAACGA CAGCCGCC GGCGGCTG G GTGCAGCG
172 CGCGCTGC GGCTAGCTACAACGA ACCAGCCG CGGCTGGT G GCAGCGCG
175 CCCCGCGC GGCTAGCTACAACGA TGCACCAG CTGGTGCA G GCGCGGGG
177 GTCCCCGC GGCTAGCTACAACGA GCTGCACC GGTGCAGC G GCGGGGAC
183 CGCCGGGT GGCTAGCTACAACGA CCCCGCGC GCGCGGGG A ACCCGGCG
188 AAAGCCGC GGCTAGCTACAACGA CGGGTCCC GGGACCCG G GCGGCTTT
191 CGGAAAGC GGCTAGCTACAACGA CGCCGGGT ACCCGGCG G GCTTTCCG
198 CAGCGCGC GGCTAGCTACAACGA GGAAAGCC GGCTTTCC G GCGCGCTG
200 ACCAGCGC GGCTAGCTACAACGA GCGGAAAG CTTTCCGC G GCGCTGGT
202 CCACCAGC GGCTAGCTACAACGA GCGCGGAA TTCCGCGC G GCTGGTGG
206 TGGGCCAC GGCTAGCTACAACGA CAGCGCGC GCGCGCTG G GTGGCCCA
209 CACTGGGC GGCTAGCTACAACGA CACCAGCG CGCTGGTG G GCCCAGTG
214 CCAGGCAC GGCTAGCTACAACGA TGGGCCAC GTGGCCCA G GTGCCTGG
Table 16
Table 16
341 GCCAGCAC GGCTAGCTACAACGA GTTCTTCG CGAAGAAC G GTGCTGGC
343 AGGCCAGC GGCTAGCTACAACGA ACGTTCTT AAGAACGT G GCTGGCCT
347 CCGAAGGC GGCTAGCTACAACGA CAGCACGT ACGTGCTG G GCCTTCGG
354 CGCGAAGC GGCTAGCTACAACGA CGAAGGCC GGCCTTCG G GCTTCGCG
359 AGCAGCGC GGCTAGCTACAACGA GAAGCCGA TCGGCTTC G GCGCTGCT
361 CCAGCAGC GGCTAGCTACAACGA GCGAAGCC GGCTTCGC G GCTGCTGG
364 CGTCCAGC GGCTAGCTACAACGA AGCGCGAA TTCGCGCT G GCTGGACG
369 GGCCCCGT GGCTAGCTACAACGA CCAGCAGC GCTGCTGG A ACGGGGCC
374 CCGCGGGC GGCTAGCTACAACGA CCCGTCCA TGGACGGG G GCCCGCGG
378 GCCCCCGC GGCTAGCTACAACGA GGGCCCCG CGGGGCCC G GCGGGGGC
384 GGGGGGGC GGCTAGCTACAACGA CCCCGCGG CCGCGGGG G GCCCCCCC
395 GTGAAGGC GGCTAGCTACAACGA CTCGGGGG CCCCCGAG G GCCTTCAC
401 CTGGTGGT GGCTAGCTACAACGA GAAGGCCT AGGCCTTC A ACCACCAG
404 ACGCTGGT GGCTAGCTACAACGA GGTGAAGG CCTTCACC A ACCAGCGT
408 GCGCACGC GGCTAGCTACAACGA TGGTGGTG CACCACCA G GCGTGCGC
410 CTGCGCAC GGCTAGCTACAACGA GCTGGTGG CCACCAGC G GTGCGCAG
412 AGCTGCGC GGCTAGCTACAACGA ACGCTGGT ACCAGCGT G GCGCAGCT
414 GTAGCTGC GGCTAGCTACAACGA GCACGCTG CAGCGTGC G GCAGCTAC
417 CAGGTAGC GGCTAGCTACAACGA TGCGCACG CGTGCGCA G GCTACCTG
420 GGGCAGGT GGCTAGCTACAACGA AGCTGCGC GCGCAGCT A ACCTGCCC
424 TGTTGGGC GGCTAGCTACAACGA AGGTAGCT AGCTACCT G GCCCAACA
429 CACCGTGT GGCTAGCTACAACGA TGGGCAGG CCTGCCCA A ACACGGTG
431 GTCACCGT GGCTAGCTACAACGA GTTGGGCA TGCCCAAC A ACGGTGAC
434 TCGGTCAC GGCTAGCTACAACGA CGTGTTGG CCAACACG G GTGACCGA
437 GCGTCGGT GGCTAGCTACAACGA CACCGTGT ACACGGTG A ACCGACGC
441 CAGTGCGT GGCTAGCTACAACGA CGGTCACC GGTGACCG A ACGCACTG
443 CGCAGTGC GGCTAGCTACAACGA GTCGGTCA TGACCGAC G GCACTGCG
445 CCCGCAGT GGCTAGCTACAACGA GCGTCGGT ACCGACGC A ACTGCGGG
448 TCCCCCGC GGCTAGCTACAACGA AGTGCGTC GACGCACT G GCGGGGGA
456 CGCCCCGC GGCTAGCTACAACGA TCCCCCGC GCGGGGGA G GCGGGGCG
461 CCCCACGC GGCTAGCTACAACGA CCCGCTCC GGAGCGGG G GCGTGGGG
Table 16
Table 16
Table 16
689 AGGGGGAC GGCTAGCTACAACGA CCCGGCCT AGGCCGGG G GTCCCCCT
699 TGGCAGGC GGCTAGCTACAACGA CCAGGGGG CCCCCTGG G GCCTGCCA
703 GGGCTGGC GGCTAGCTACAACGA AGGCCCAG CTGGGCCT G GCCAGCCC
707 CCCGGGGC GGCTAGCTACAACGA TGGCAGGC GCCTGCCA G GCCCCGGG
714 CCTCGCAC GGCTAGCTACAACGA CCGGGGCT AGCCCCGG G GTGCGAGG
716 CTCCTCGC GGCTAGCTACAACGA ACCCGGGG CCCCGGGT G GCGAGGAG
724 CCCCGCGC GGCTAGCTACAACGA CTCCTCGC GCGAGGAG G GCGCGGGG
726 GCCCCCGC GGCTAGCTACAACGA GCCTCCTC GAGGAGGC G GCGGGGGC
732 GGCACTGC GGCTAGCTACAACGA CCCCGCGC GCGCGGGG G GCAGTGCC
735 GCTGGCAC GGCTAGCTACAACGA TGCCCCCG CGGGGGCA G GTGCCAGC
737 CGGCTGGC GGCTAGCTACAACGA ACTGCCCC GGGGCAGT G GCCAGCCG
741 ACTTCGGC GGCTAGCTACAACGA TGGCACTG CAGTGCCA G GCCGAAGT
747 CGGCAGAC GGCTAGCTACAACGA TTCGGCTG CAGCCGAA G GTCTGCCG
751 GCAACGGC GGCTAGCTACAACGA AGACTTCG CGAAGTCT G GCCGTTGC
754 TGGGCAAC GGCTAGCTACAACGA GGCAGACT AGTCTGCC G GTTGCCCA
757 TCTTGGGC GGCTAGCTACAACGA AACGGCAG CTGCCGTT G GCCCAAGA
766 GCCTGGGC GGCTAGCTACAACGA CTCTTGGG CCCAAGAG G GCCCAGGC
772 CGCCACGC GGCTAGCTACAACGA CTGGGCCT AGGCCCAG G GCGTGGCG
774 AGCGCCAC GGCTAGCTACAACGA GCCTGGGC GCCCAGGC G GTGGCGCT
777 GGCAGCGC GGCTAGCTACAACGA CACGCCTG CAGGCGTG G GCGCTGCC
779 GGGGCAGC GGCTAGCTACAACGA GCCACGCC GGCGTGGC G GCTGCCCC
782 TCAGGGGC GGCTAGCTACAACGA AGCGCCAC GTGGCGCT G GCCCCTGA
790 GCTCCGGC GGCTAGCTACAACGA TCAGGGGC GCCCCTGA G GCCGGAGC
796 GCGTCCGC GGCTAGCTACAACGA TCCGGCTC GAGCCGGA G GCGGACGC
800 ACGGGCGT GGCTAGCTACAACGA CCGCTCCG CGGAGCGG A ACGCCCGT
802 CAACGGGC GGCTAGCTACAACGA GTCCGCTC GAGCGGAC G GCCCGTTG
806 TGCCCAAC GGCTAGCTACAACGA GGGCGTCC GGACGCCC G GTTGGGCA
811 ACCCCTGC GGCTAGCTACAACGA CCAACGGG CCCGTTGG G GCAGGGGT
817 CCCAGGAC GGCTAGCTACAACGA CCCTGCCC GGGCAGGG G GTCCTGGG
824 GGGTGGGC GGCTAGCTACAACGA CCAGGACC GGTCCTGG G GCCCACCC
828 GCCCGGGT GGCTAGCTACAACGA GGGCCCAG CTGGGCCC A ACCCGGGC
Table 16
834 CGTCCTGC GGCTAGCTACAACGA CCGGGTGG CCACCCGG G GCAGGACG
839 CCACGCGT GGCTAGCTACAACGA CCTGCCCG CGGGCAGG A ACGCGTGG
841 GTCCACGC GGCTAGCTACAACGA GTCCTGCC GGCAGGAC G GCGTGGAC
843 CGGTCCAC GGCTAGCTACAACGA GCGTCCTG CAGGACGC G GTGGACCG
847 CACTCGGT GGCTAGCTACAACGA CCACGCGT ACGCGTGG A ACCGAGTG
852 .ACGGTCAC GGCTAGCTACAACGA TCGGTCCA TGGACCGA G GTGACCGT
855 ACCACGGT GGCTAGCTACAACGA CACTCGGT ACCGAGTG A ACCGTGGT
858 GAAACCAC GGCTAGCTACAACGA GGTCACTC GAGTGACC G GTGGTTTC
861 ACAGAAAC GGCTAGCTACAACGA CACGGTCA TGACCGTG G GTTTCTGT
867 CACCACAC GGCTAGCTACAACGA AGAAACCA TGGTTTCT G GTGTGGTG
869 GACACCAC GGCTAGCTACAACGA ACAGAAAC GTTTCTGT G GTGGTGTC
872 GGTGACAC GGCTAGCTACAACGA CACACAGA TCTGTGTG G GTGTCACC
874 CAGGTGAC GGCTAGCTACAACGA ACCACACA TGTGTGGT G GTCACCTG
877 TGGCAGGT GGCTAGCTACAACGA GACACCAC GTGGTGTC A ACCTGCCA
881 GGTCTGGC GGCTAGCTACAACGA AGGTGACA TGTCACCT G GCCAGACC
886 CGGCGGGT GGCTAGCTACAACGA CTGGCAGG CCTGCCAG A ACCCGCCG
890 TCTTCGGC GGCTAGCTACAACGA GGGTCTGG CCAGACCC G GCCGAAGA
899 GAGGTGGC GGCTAGCTACAACGA TTCTTCGG CCGAAGAA G GCCACCTC
902 AAAGAGGT GGCTAGCTACAACGA GGCTTCTT AAGAAGCC A ACCTCTTT
915 GAGCGCAC GGCTAGCTACAACGA CCTCCAAA TTTGGAGG G GTGCGCTC
917 GAGAGCGC GGCTAGCTACAACGA ACCCTCCA TGGAGGGT G GCGCTCTC
919 CAGAGAGC GGCTAGCTACAACGA GCACCCTC GAGGGTGC G GCTCTCTG
927 GCGCGTGC GGCTAGCTACAACGA CAGAGAGC GCTCTCTG G GCACGCGC
929 TGGCGCGT GGCTAGCTACAACGA GCCAGAGA TCTCTGGC A ACGCGCCA
931 AGTGGCGC GGCTAGCTACAACGA GTGCCAGA TCTGGCAC G GCGCCACT
933 GGAGTGGC GGCTAGCTACAACGA GCGTGCCA TGGCACGC G GCCACTCC
936 GTGGGAGT GGCTAGCTACAACGA GGCGCGTG CACGCGCC A ACTCCCAC
942 GGATGGGT GGCTAGCTACAACGA GGGAGTGG CCACTCCC A ACCCATCC
946 CCACGGAT GGCTAGCTACAACGA GGGTGGGA TCCCACCC A ATCCGTGG
950 CGGCCCAC GGCTAGCTACAACGA GGATGGGT ACCCATCC G GTGGGCCG
954 CTGGCGGC GGCTAGCTACAACGA CCACGGAT ATCCGTGG G GCCGCCAG
Table 16
957 GTGCTGGC GGCTAGCTACAACGA GGCCCACG CGTGGGCC G GCCAGCAC
961 CGTGGTGC GGCTAGCTACAACGA TGGCGGCC GGCCGCCA G GCACCACG
963 CGCGTGGT GGCTAGCTACAACGA GCTGGCGG CCGCCAGC A ACCACGCG
966 GCCCGCGT GGCTAGCTACAACGA GGTGCTGG CCAGCACC A ACGCGGGC
968 GGGCCCGC GGCTAGCTACAACGA GTGGTGCT AGCACCAC G GCGGGCCC
972 TGGGGGGC GGCTAGCTACAACGA CCGCGTGG CCACGCGG G GCCCCCCA
979 ATGTGGAT GGCTAGCTACAACGA GGGGGGCC GGCCCCCC A ATCCACAT
983 CGCGATGT GGCTAGCTACAACGA GGATGGGG CCCCATCC A ACATCGCG
985 GCCGCGAT GGCTAGCTACAACGA GTGGATGG CCATCCAC A ATCGCGGC
988 GTGGCCGC GGCTAGCTACAACGA GATGTGGA TCCACATC G GCGGCCAC
991 GTGGTGGC GGCTAGCTACAACGA CGCGATGT ACATCGCG G GCCACCAC
994 GACGTGGT GGCTAGCTACAACGA GGCCGCGA TCGCGGCC A ACCACGTC
997 AGGGACGT GGCTAGCTACAACGA GGTGGCCG CGGCCACC A ACGTCCCT
999 CCAGGGAC GGCTAGCTACAACGA GTGGTGGC GCCACCAC G GTCCCTGG
1008 AGGCGTGT GGCTAGCTACAACGA CCCAGGGA TCCCTGGG A ACACGCCT
1010 CAAGGCGT GGCTAGCTACAACGA GTCCCAGG CCTGGGAC A ACGCCTTG
1012 GACAAGGC GGCTAGCTACAACGA GTGTCCCA TGGGACAC G GCCTTGTC
1017 CGGGGGAC GGCTAGCTACAACGA AAGGCGTG CACGCCTT G GTCCCCCG
1025 GCGTACAC GGCTAGCTACAACGA CGGGGGAC GTCCCCCG G GTGTACGC
1027 CGGCGTAC GGCTAGCTACAACGA ACCGGGGG CCCCCGGT G GTACGCCG
1029 CTCGGCGT GGCTAGCTACAACGA ACACCGGG CCCGGTGT A ACGCCGAG
1031 GTCTCGGC GGCTAGCTACAACGA GTACACCG CGGTGTAC G GCCGAGAC
1037 TGCTTGGT GGCTAGCTACAACGA CTCGGCGT ACGCCGAG A ACCAAGCA
1042 GGAAGTGC GGCTAGCTACAACGA TTGGTCTC GAGACCAA G GCACTTCC
1044 GAGGAAGT GGCTAGCTACAACGA GCTTGGTC GACCAAGC A ACTTCCTC
1053 TGAGGAGT GGCTAGCTACAACGA AGAGGAAG CTTCCTCT A ACTCCTCA
1062 CTTGTCGC GGCTAGCTACAACGA CTGAGGAG CTCCTCAG G GCGACAAG
1065 CTCCTTGT GGCTAGCTACAACGA CGCCTGAG CTCAGGCG A ACAAGGAG
1072 GCAGCTGC GGCTAGCTACAACGA TCCTTGTC GACAAGGA G GCAGCTGC
1075 GCCGCAGC GGCTAGCTACAACGA TGCTCCTT AAGGAGCA G GCTGCGGC
1078 AGGGCCGC GGCTAGCTACAACGA AGCTGCTC GAGCAGCT G GCGGCCCT
Table 16
1081 AGGAGGGC GGCTAGCTACAACGA CGCAGCTG CAGCTGCG G GCCCTCCT
1093 AGCTGAGT GGCTAGCTACAACGA AGGAAGGA TCCTTCCT A ACTCAGCT
1098 CAGAGAGC GGCTAGCTACAACGA TGAGTAGG CCTACTCA G GCTCTCTG
1108 GGCTGGGC GGCTAGCTACAACGA CTCAGAGA TCTCTGAG G GCCCAGCC
1113 AGTCAGGC GGCTAGCTACAACGA TGGGCCTC GAGGCCCA G GCCTGACT
1118 GCGCCAGT GGCTAGCTACAACGA CAGGCTGG CCAGCCTG A ACTGGCGC
1122 CCGAGCGC GGCTAGCTACAACGA CAGTCAGG CCTGACTG G GCGCTCGG
1124 CTCCGAGC GGCTAGCTACAACGA GCCAGTCA TGACTGGC G GCTCGGAG
1132 CCACGAGC GGCTAGCTACAACGA CTCCGAGC GCTCGGAG G GCTCGTGG
1136 GTCTCCAC GGCTAGCTACAACGA GAGCCTCC GGAGGCTC G GTGGAGAC
1142 AAGATGGT GGCTAGCTACAACGA CTCCACGA TCGTGGAG A ACCATCTT
1145 AGAAAGAT GGCTAGCTACAACGA GGTCTCCA TGGAGACC A ATCTTTCT
1155 CCTGGAAC GGCTAGCTACAACGA CCAGAAAG CTTTCTGG G GTTCCAGG
1162 TCCAGGGC GGCTAGCTACAACGA CTGGAACC GGTTCCAG G GCCCTGGA
1169 CCTGGCAT GGCTAGCTACAACGA CCAGGGCC GGCCCTGG A ATGCCAGG
1171 TCCCTGGC GGCTAGCTACAACGA ATCCAGGG CCCTGGAT G GCCAGGGA
1178 CGGGGAGT GGCTAGCTACAACGA CCCTGGCA TGCCAGGG A ACTCCCCG
1185 CAACCTGC GGCTAGCTACAACGA GGGGAGTC GACTCCCC G GCAGGTTG
1189 GGGGCAAC GGCTAGCTACAACGA CTGCGGGG CCCCGCAG G GTTGCCCC
1192 GGCGGGGC GGCTAGCTACAACGA AACCTGCG CGCAGGTT G GCCCCGCC
1197 GGGCAGGC GGCTAGCTACAACGA GGGGCAAC GTTGCCCC G GCCTGCCC
1201 GCTGGGGC GGCTAGCTACAACGA AGGCGGGG CCCCGCCT G GCCCCAGC
1207 AGTAGCGC GGCTAGCTACAACGA TGGGGCAG CTGCCCCA G GCGCTACT
1209 CCAGTAGC GGCTAGCTACAACGA GCTGGGGC GCCCCAGC G GCTACTGG
1212 TTGCCAGT GGCTAGCTACAACGA AGCGCTGG CCAGCGCT A ACTGGCAA
1216 GCATTTGC GGCTAGCTACAACGA CAGTAGCG CGCTACTG G GCAAATGC
1220 GGCCGCAT GGCTAGCTACAACGA TTGCCAGT ACTGGCAA A ATGCGGCC
1222 GGGGCCGC GGCTAGCTACAACGA ATTTGCCA TGGCAAAT G GCGGCCCC
1225 ACAGGGGC GGCTAGCTACAACGA CGCATTTG CAAATGCG G GCCCCTGT
1231 CCAGAAAC GGCTAGCTACAACGA AGGGGCCG CGGCCCCT G GTTTCTGG
1240 CAAGCAGC GGCTAGCTACAACGA TCCAGAAA TTTCTGGA G GCTGCTTG
Table 16
1243 TCCCAAGC GGCTAGCTACAACGA AGCTCCAG CTGGAGCT G GCTTGGGA
1251 CGCGTGGT GGCTAGCTACAACGA TCCCAAGC GCTTGGGA A ACCACGCG
1254 CTGCGCGT GGCTAGCTACAACGA GGTTCCCA TGGGAACC A ACGCGCAG
1256 CACTGCGC GGCTAGCTACAACGA GTGGTTCC GGAACCAC G GCGCAGTG
1258 GGCACTGC GGCTAGCTACAACGA GCGTGGTT AACCACGC G GCAGTGCC
1261 AGGGGCAC GGCTAGCTACAACGA TGCGCGTG CACGCGCA G GTGCCCCT
1263 GTAGGGGC GGCTAGCTACAACGA ACTGCGCG CGCGCAGT G GCCCCTAC
1269 CACCCCGT GGCTAGCTACAACGA AGGGGCAC GTGCCCCT A ACGGGGTG
1274 AGGAGCAC GGCTAGCTACAACGA CCCGTAGG CCTACGGG G GTGCTCCT
1276 TGAGGAGC GGCTAGCTACAACGA ACCCCGTA TACGGGGT G GCTCCTCA
1286 CAGTGCGT GGCTAGCTACAACGA CTTGAGGA TCCTCAAG A ACGCACTG
1288 GGCAGTGC GGCTAGCTACAACGA GTCTTGAG CTCAAGAC G GCACTGCC
1290 CGGGCAGT GGCTAGCTACAACGA GCGTCTTG CAAGACGC A ACTGCCCG
1293 CAGCGGGC GGCTAGCTACAACGA AGTGCGTC GACGCACT G GCCCGCTG
1297 CTCGCAGC GGCTAGCTACAACGA GGGCAGTG CACTGCCC G GCTGCGAG
1300 CAGCTCGC GGCTAGCTACAACGA AGCGGGCA TGCCCGCT G GCGAGCTG
1304 ACCGCAGC GGCTAGCTACAACGA TCGCAGCG CGCTGCGA G GCTGCGGT
1307 GTGACCGC GGCTAGCTACAACGA AGCTCGCA TGCGAGCT G GCGGTCAC
1310 GGGGTGAC GGCTAGCTACAACGA CGCAGCTC GAGCTGCG G GTCACCCC
1313 GCTGGGGT GGCTAGCTACAACGA GACCGCAG CTGCGGTC A ACCCCAGC
1319 CCGGCTGC GGCTAGCTACAACGA TGGGGTGA TCACCCCA G GCAGCCGG
1322 ACACCGGC GGCTAGCTACAACGA TGCTGGGG CCCCAGCA G GCCGGTGT
1326 ACAGACAC GGCTAGCTACAACGA CGGCTGCT AGCAGCCG G GTGTCTGT
1328 GCACAGAC GGCTAGCTACAACGA ACCGGCTG CAGCCGGT G GTCTGTGC
1332 CCGGGCAC GGCTAGCTACAACGA AGACACCG CGGTGTCT G GTGCCCGG
1334 TCCCGGGC GGCTAGCTACAACGA ACAGACAC GTGTCTGT G GCCCGGGA
1345 CCTGGGGC GGCTAGCTACAACGA TTCTCCCG CGGGAGAA G GCCCCAGG
1353 CACAGAGC GGCTAGCTACAACGA CCTGGGGC GCCCCAGG G GCTCTGTG
1358 GCCGCCAC GGCTAGCTACAACGA AGAGCCCT AGGGCTCT G GTGGCGGC
1361 GGGGCCGC GGCTAGCTACAACGA CACAGAGC GCTCTGTG G GCGGCCCC
1364 TCGGGGGC GGCTAGCTACAACGA CGCCACAG CTGTGGCG G GCCCCCGA
Table 16
1380 GTCTGTGT GGCTAGCTACAACGA CCTCCTCC GGAGGAGG A ACACAGAC
1382 GGGTCTGT GGCTAGCTACAACGA GTCCTCCT AGGAGGAC A ACAGACCC
1386 ACGGGGGT GGCTAGCTACAACGA CTGTGTCC GGACACAG A ACCCCCGT
1392 CAGGCGAC GGCTAGCTACAACGA GGGGGTCT AGACCCCC G GTCGCCTG
1395 CACCAGGC GGCTAGCTACAACGA GACGGGGG CCCCCGTC G GCCTGGTG
1400 AGCTGCAC GGCTAGCTACAACGA CAGGCGAC GTCGCCTG G GTGCAGCT
1402 GCAGCTGC GGCTAGCTACAACGA ACCAGGCG CGCCTGGT G GCAGCTGC
1405 GGAGCAGC GGCTAGCTACAACGA TGCACCAG CTGGTGCA G GCTGCTCC
1408 GGCGGAGC GGCTAGCTACAACGA AGCTGCAC GTGCAGCT G GCTCCGCC
1413 GTGCTGGC GGCTAGCTACAACGA GGAGCAGC GCTGCTCC G GCCAGCAC
1417 TGCTGTGC GGCTAGCTACAACGA TGGCGGAG CTCCGCCA G GCACAGCA
1419 GCTGCTGT GGCTAGCTACAACGA GCTGGCGG CCGCCAGC A ACAGCAGC
1422 GGGGCTGC GGCTAGCTACAACGA TGTGCTGG CCAGCACA G GCAGCCCC
1425 CCAGGGGC GGCTAGCTACAACGA TGCTGTGC GCACAGCA G GCCCCTGG
1432 ACACCTGC GGCTAGCTACAACGA CAGGGGCT AGCCCCTG G GCAGGTGT
1436 CCGTACAC GGCTAGCTACAACGA CTGCCAGG CCTGGCAG G GTGTACGG
1438 AGCCGTAC GGCTAGCTACAACGA ACCTGCCA TGGCAGGT G GTACGGCT
1440 GAAGCCGT GGCTAGCTACAACGA ACACCTGC GCAGGTGT A ACGGCTTC
1443 CACGAAGC GGCTAGCTACAACGA CGTACACC GGTGTACG G GCTTCGTG
1448 GCCCGCAC GGCTAGCTACAACGA GAAGCCGT ACGGCTTC G GTGCGGGC
1450 AGGCCCGC GGCTAGCTACAACGA ACGAAGCC GGCTTCGT G GCGGGCCT
1454 AGGCAGGC GGCTAGCTACAACGA CCGCACGA TCGTGCGG G GCCTGCCT
1458 GCGCAGGC GGCTAGCTACAACGA AGGCCCGC GCGGGCCT G GCCTGCGC
1462 GCCGGCGC GGCTAGCTACAACGA AGGCAGGC GCCTGCCT G GCGCCGGC
1464 CAGCCGGC GGCTAGCTACAACGA GCAGGCAG CTGCCTGC G GCCGGCTG
1468 GCACCAGC GGCTAGCTACAACGA CGGCGCAG CTGCGCCG G GCTGGTGC
1472 GGGGGCAC GGCTAGCTACAACGA CAGCCGGC GCCGGCTG G GTGCCCCC
1474 CTGGGGGC GGCTAGCTACAACGA ACCAGCCG CGGCTGGT G GCCCCCAG
1482 CCAGAGGC GGCTAGCTACAACGA CTGGGGGC GCCCCCAG G GCCTCTGG
1491 CCTGGAGC GGCTAGCTACAACGA CCCAGAGG CCTCTGGG G GCTCCAGG
1498 CGTTGTGC GGCTAGCTACAACGA CTGGAGCC GGCTCCAG G GCACAACG
Table 16
Table 16
1638 CGGAACAC GGCTAGCTACAACGA AGCCAACC GGTTGGCT G GTGTTCCG
1640 GCCGGAAC GGCTAGCTACAACGA ACAGCCAA TTGGCTGT G GTTCCGGC
1646 TCTGCGGC GGCTAGCTACAACGA CGGAACAC GTGTTCCG G GCCGCAGA
1649 TGCTCTGC GGCTAGCTACAACGA GGCCGGAA TTCCGGCC G GCAGAGCA
1654 GACGGTGC GGCTAGCTACAACGA TCTGCGGC GCCGCAGA G GCACCGTC
1656 CAGACGGT GGCTAGCTACAACGA GCTCTGCG CGCAGAGC A ACCGTCTG
1659 ACGCAGAC GGCTAGCTACAACGA GGTGCTCT AGAGCACC G GTCTGCGT
1663 CCTCACGC GGCTAGCTACAACGA AGACGGTG CACCGTCT G GCGTGAGG
1665 CTCCTCAC GGCTAGCTACAACGA GCAGACGG CCGTCTGC G GTGAGGAG
1673 GCCAGGAT GGCTAGCTACAACGA CTCCTCAC GTGAGGAG A ATCCTGGC
1679 AACTTGGC GGCTAGCTACAACGA CAGGATCT AGATCCTG G GCCAAGTT
1684 GCAGGAAC GGCTAGCTACAACGA TTGGCCAG CTGGCCAA G GTTCCTGC
1690 GCCAGTGC GGCTAGCTACAACGA AGGAACTT AAGTTCCT G GCACTGGC
1692 CAGCCAGT GGCTAGCTACAACGA GCAGGAAC GTTCCTGC A ACTGGCTG
1696 TCATCAGC GGCTAGCTACAACGA CAGTGCAG CTGCACTG G GCTGATGA
1700 ACACTCAT GGCTAGCTACAACGA CAGCCAGT ACTGGCTG A ATGAGTGT
1704 GTACACAC GGCTAGCTACAACGA TCATCAGC GCTGATGA G GTGTGTAC
1706 ACGTACAC GGCTAGCTACAACGA ACTCATCA TGATGAGT G GTGTACGT
1708 CGACGTAC GGCTAGCTACAACGA ACACTCAT ATGAGTGT G GTACGTCG
1710 GACGACGT GGCTAGCTACAACGA ACACACTC GAGTGTGT A ACGTCGTC
1712 TCGACGAC GGCTAGCTACAACGA GTACACAC GTGTGTAC G GTCGTCGA
1715 AGCTCGAC GGCTAGCTACAACGA GACGTACA TGTACGTC G GTCGAGCT
1720 TGAGCAGC GGCTAGCTACAACGA TCGACGAC GTCGTCGA G GCTGCTCA
1723 ACCTGAGC GGCTAGCTACAACGA AGCTCGAC GTCGAGCT G GCTCAGGT
1729 AGAAAGAC GGCTAGCTACAACGA CTGAGCAG CTGCTCAG G GTCTTTCT
1740 CGTGACAT GGCTAGCTACAACGA AAAAGAAA TTTCTTTT A ATGTCACG
1742 TCCGTGAC GGCTAGCTACAACGA ATAAAAGA TCTTTTAT G GTCACGGA
1745 GTCTCCGT GGCTAGCTACAACGA GACATAAA TTTATGTC A ACGGAGAC
1751 AACGTGGT GGCTAGCTACAACGA CTCCGTGA TCACGGAG A ACCACGTT
1754 TGAAACGT GGCTAGCTACAACGA GGTCTCCG CGGAGACC A ACGTTTCA
1756 TTTGAAAC GGCTAGCTACAACGA GTGGTCTC GAGACCAC G GTTTCAAA
Table 16
1767 GAGCCTGT GGCTAGCTACAACGA TCTTTTGA TCAAAAGA A ACAGGCTC
1771 AAAAGAGC GGCTAGCTACAACGA CTGTTCTT AAGAACAG G GCTCTTTT
1782 CTTCCGGT GGCTAGCTACAACGA AGAAAAAG CTTTTTCT A ACCGGAAG
1791 CCAGACAC GGCTAGCTACAACGA TCTTCCGG CCGGAAGA G GTGTCTGG
1793 CTCCAGAC GGCTAGCTACAACGA ACTCTTCC GGAAGAGT G GTCTGGAG
1800 CAACTTGC GGCTAGCTACAACGA TCCAGACA TGTCTGGA G GCAAGTTG
1804 TTTGCAAC GGCTAGCTACAACGA TTGCTCCA TGGAGCAA G GTTGCAAA
1807 TGCTTTGC GGCTAGCTACAACGA AACTTGCT AGCAAGTT G GCAAAGCA
1812 TCCAATGC GGCTAGCTACAACGA TTTGCAAC GTTGCAAA G GCATTGGA
1814 ATTCCAAT GGCTAGCTACAACGA GCTTTGCA TGCAAAGC A ATTGGAAT
1820 TGTCTGAT GGCTAGCTACAACGA TCCAATGC GCATTGGA A ATCAGACA
1825 AGTGCTGT GGCTAGCTACAACGA CTGATTCC GGAATCAG A ACAGCACT
1828 TCAAGTGC GGCTAGCTACAACGA TGTCTGAT ATCAGACA G GCACTTGA
1830 CTTCAAGT GGCTAGCTACAACGA GCTGTCTG CAGACAGC A ACTTGAAG 1
1841 AGCTGCAC GGCTAGCTACAACGA CCTCTTCA TGAAGAGG G GTGCAGCT
1843 GCAGCTGC GGCTAGCTACAACGA ACCCTCTT AAGAGGGT G GCAGCTGC
1846 CCCGCAGC GGCTAGCTACAACGA TGCACCCT AGGGTGCA G GCTGCGGG
1849 GCTCCCGC GGCTAGCTACAACGA AGCTGCAC GTGCAGCT G GCGGGAGC
1855 CCGACAGC GGCTAGCTACAACGA TCCCGCAG CTGCGGGA G GCTGTCGG
1858 CTTCCGAC GGCTAGCTACAACGA AGCTCCCG CGGGAGCT G GTCGGAAG
1865 ACCTCTGC GGCTAGCTACAACGA TTCCGACA TGTCGGAA G GCAGAGGT
1871 TGCCTGAC GGCTAGCTACAACGA CTCTGCTT AAGCAGAG G GTCAGGCA
1876 GATGCTGC GGCTAGCTACAACGA CTGACCTC GAGGTCAG G GCAGCATC
1879 CCCGATGC GGCTAGCTACAACGA TGCCTGAC GTCAGGCA G GCATCGGG
1881 TTCCCGAT GGCTAGCTACAACGA GCTGCCTG CAGGCAGC A ATCGGGAA
1889 GGCCTGGC GGCTAGCTACAACGA TTCCCGAT ATCGGGAA G GCCAGGCC
1894 GGGCGGGC GGCTAGCTACAACGA CTGGCTTC GAAGCCAG G GCCCGCCC
1898 AGCAGGGC GGCTAGCTACAACGA GGGCCTGG CCAGGCCC G GCCCTGCT
1903 ACGTCAGC GGCTAGCTACAACGA AGGGCGGG CCCGCCCT G GCTGACGT
1907 CTGGACGT GGCTAGCTACAACGA CAGCAGGG CCCTGCTG A ACGTCCAG
1909 GTCTGGAC GGCTAGCTACAACGA GTCAGCAG CTGCTGAC G GTCCAGAC
Table 16
1915 AGCGGAGT GGCTAGCTACAACGA CTGGACGT ACGTCCAG A ACTCCGCT
1920 GATGAAGC GGCTAGCTACAACGA GGAGTCTG CAGACTCC G GCTTCATC
1925 TTGGGGAT GGCTAGCTACAACGA GAAGCGGA TCCGCTTC A ATCCCCAA
1933 CGTCAGGC GGCTAGCTACAACGA TTGGGGAT ATCCCCAA G GCCTGACG
1938 CAGCCCGT GGCTAGCTACAACGA CAGGCTTG CAAGCCTG A ACGGGCTG
1942 GCCGCAGC GGCTAGCTACAACGA CCGTCAGG CCTGACGG G GCTGCGGC
1945 TCGGCCGC GGCTAGCTACAACGA AGCCCGTC GACGGGCT G GCGGCCGA
1948 CAATCGGC GGCTAGCTACAACGA CGCAGCCC GGGCTGCG G GCCGATTG
1952 TTCACAAT GGCTAGCTACAACGA CGGCCGCA TGCGGCCG A ATTGTGAA
1955 ATGTTCAC GGCTAGCTACAACGA AATCGGCC GGCCGATT G GTGAACAT
1959 GTCCATGT GGCTAGCTACAACGA TCACAATC GATTGTGA A ACATGGAC
1961 TAGTCCAT GGCTAGCTACAACGA GTTCACAA TTGTGAAC A ATGGACTA
1965 GACGTAGT GGCTAGCTACAACGA CCATGTTC GAACATGG A ACTACGTC
1968 CACGACGT GGCTAGCTACAACGA AGTCCATG CATGGACT A ACGTCGTG
1970 CCCACGAC GGCTAGCTACAACGA GTAGTCCA TGGACTAC G GTCGTGGG
1973 GCTCCCAC GGCTAGCTACAACGA GACGTAGT ACTACGTC G GTGGGAGC
1979 GTTCTGGC GGCTAGCTACAACGA TCCCACGA TCGTGGGA G GCCAGAAC
1985 CGGAACGT GGCTAGCTACAACGA TCTGGCTC GAGCCAGA A ACGTTCCG
1987 TGCGGAAC GGCTAGCTACAACGA GTTCTGGC GCCAGAAC G GTTCCGCA
1992 TTCTCTGC GGCTAGCTACAACGA GGAACGTT AACGTTCC G GCAGAGAA
2006 CGCTCGGC GGCTAGCTACAACGA CCTCTTTT AAAAGAGG G GCCGAGCG
2011 TGAGACGC GGCTAGCTACAACGA TCGGCCCT AGGGCCGA G GCGTCTCA
2013 GGTGAGAC GGCTAGCTACAACGA GCTCGGCC GGCCGAGC G GTCTCACC
2018 CTCGAGGT GGCTAGCTACAACGA GAGACGCT AGCGTCTC A ACCTCGAG
2027 GCCTTCAC GGCTAGCTACAACGA CCTCGAGG CCTCGAGG G GTGAAGGC
2033 AACAGTGC GGCTAGCTACAACGA CTTCACCC GGGTGAAG G GCACTGTT
2035 TGAACAGT GGCTAGCTACAACGA GCCTTCAC GTGAAGGC A ACTGTTCA
2038 CGCTGAAC GGCTAGCTACAACGA AGTGCCTT AAGGCACT G GTTCAGCG
2043 GAGCACGC GGCTAGCTACAACGA TGAACAGT ACTGTTCA G GCGTGCTC
2045 TTGAGCAC GGCTAGCTACAACGA GCTGAACA TGTTCAGC G GTGCTCAA
2047 AGTTGAGC GGCTAGCTACAACGA ACGCTGAA TTCAGCGT G GCTCAACT
Table 16
2052 CTCGTAGT GGCTAGCTACAACGA TGAGCACG CGTGCTCA A ACTACGAG
2055 CCGCTCGT GGCTAGCTACAACGA AGTTGAGC GCTCAACT A ACGAGCGG
2059 GCGCCCGC GGCTAGCTACAACGA TCGTAGTT AACTACGA G GCGGGCGC
2063 CGCCGCGC GGCTAGCTACAACGA CCGCTCGT ACGAGCGG G GCGCGGCG
2065 GGCGCCGC GGCTAGCTACAACGA GCCCGCTC GAGCGGGC G GCGGCGCC
2068 CGGGGCGC GGCTAGCTACAACGA CGCGCCCG CGGGCGCG G GCGCCCCG
2070 GCCGGGGC GGCTAGCTACAACGA GCCGCGCC GGCGCGGC G GCCCCGGC
2076 CAGGAGGC GGCTAGCTACAACGA CGGGGCGC GCGCCCCG G GCCTCCTG
2085 AGAGGCGC GGCTAGCTACAACGA CCAGGAGG CCTCCTGG G GCGCCTCT
2087 ACAGAGGC GGCTAGCTACAACGA GCCCAGGA TCCTGGGC G GCCTCTGT
2093 CCCAGCAC GGCTAGCTACAACGA AGAGGCGC GCGCCTCT G GTGCTGGG
2095 GGCCCAGC GGCTAGCTACAACGA ACAGAGGC GCCTCTGT G GCTGGGCC
2100 GTCCAGGC GGCTAGCTACAACGA CCAGCACA TGTGCTGG G GCCTGGAC
2106 GATATCGT GGCTAGCTACAACGA CCAGGCCC GGGCCTGG A ACGATATC
2109 GTGGATAT GGCTAGCTACAACGA CGTCCAGG CCTGGACG A ATATCCAC
2111 CTGTGGAT GGCTAGCTACAACGA ATCGTCCA TGGACGAT A ATCCACAG
2115 GGCCCTGT GGCTAGCTACAACGA GGATATCG ' CGATATCC A ACAGGGCC
2120 CGCCAGGC GGCTAGCTACAACGA CCTGTGGA TCCACAGG G GCCTGGCG
2125 AGGTGCGC GGCTAGCTACAACGA CAGGCCCT AGGGCCTG G GCGCACCT
2127 GAAGGTGC GGCTAGCTACAACGA GCCAGGCC GGCCTGGC G GCACCTTC
2129 ACGAAGGT GGCTAGCTACAACGA GCGCCAGG CCTGGCGC A ACCTTCGT
2135 CGCAGCAC GGCTAGCTACAACGA GAAGGTGC GCACCTTC G GTGCTGCG
2137 CACGCAGC GGCTAGCTACAACGA ACGAAGGT ACCTTCGT G GCTGCGTG
2140 GCACACGC GGCTAGCTACAACGA AGCACGAA TTCGTGCT G GCGTGTGC
2142 CCGCACAC GGCTAGCTACAACGA GCAGCACG CGTGCTGC G GTGTGCGG
2144 GCCCGCAC GGCTAGCTACAACGA ACGCAGCA TGCTGCGT G GTGCGGGC
2146 GGGCCCGC GGCTAGCTACAACGA ACACGCAG CTGCGTGT G GCGGGCCC
2150 TCCTGGGC GGCTAGCTACAACGA CCGCACAC GTGTGCGG G GCCCAGGA
2157 CGGCGGGT GGCTAGCTACAACGA CCTGGGCC GGCCCAGG A ACCCGCCG
2161 CAGGCGGC GGCTAGCTACAACGA GGGTCCTG CAGGACCC G GCCGCCTG
2164 GCTCAGGC GGCTAGCTACAACGA GGCGGGTC GACCCGCC G GCCTGAGC
Table 16
Table 16
2278 ACCGACGC GGCTAGCTACAACGA ACGCAGTA TACTGCGT G GCGTCGGT
2280 ATACCGAC GGCTAGCTACAACGA GCACGCAG CTGCGTGC G GTCGGTAT
2284 CGGCATAC GGCTAGCTACAACGA CGACGCAC GTGCGTCG G GTATGCCG
2286 CACGGCAT GGCTAGCTACAACGA ACCGACGC GCGTCGGT A ATGCCGTG
2288 ACCACGGC GGCTAGCTACAACGA ATACCGAC GTCGGTAT G GCCGTGGT
2291 TGGACCAC GGCTAGCTACAACGA GGCATACC GGTATGCC G GTGGTCCA
2294 TTCTGGAC GGCTAGCTACAACGA CACGGCAT ATGCCGTG G GTCCAGAA
2303 TGGGCGGC GGCTAGCTACAACGA CTTCTGGA TCCAGAAG G GCCGCCCA
2306 CCATGGGC GGCTAGCTACAACGA GGCCTTCT AGAAGGCC G GCCCATGG
2310 GTGCCCAT GGCTAGCTACAACGA GGGCGGCC GGCCGCCC A ATGGGCAC
2314 GGACGTGC GGCTAGCTACAACGA CCATGGGC GCCCATGG G GCACGTCC
2316 GCGGACGT GGCTAGCTACAACGA GCCCATGG CCATGGGC A ACGTCCGC
2318 TTGCGGAC GGCTAGCTACAACGA GTGCCCAT ATGGGCAC G GTCCGCAA
2322 GGCCTTGC GGCTAGCTACAACGA GGACGTGC GCACGTCC G GCAAGGCC
2327 TTGAAGGC GGCTAGCTACAACGA CTTGCGGA TCCGCAAG G GCCTTCAA
2337 GACGTGGC GGCTAGCTACAACGA TCTTGAAG CTTCAAGA G GCCACGTC
2340 AGAGACGT GGCTAGCTACAACGA GGCTCTTG CAAGAGCC A ACGTCTCT
2342 GTAGAGAC GGCTAGCTACAACGA GTGGCTCT AGAGCCAC G GTCTCTAC
2348 GTCAAGGT GGCTAGCTACAACGA AGAGACGT ACGTCTCT A ACCTTGAC
2354 AGGTCTGT GGCTAGCTACAACGA CAAGGTAG CTACCTTG A ACAGACCT
2358 CTGGAGGT GGCTAGCTACAACGA CTGTCAAG CTTGACAG A ACCTCCAG
2365 TGTACGGC GGCTAGCTACAACGA TGGAGGTC GACCTCCA G GCCGTACA
2368 GCATGTAC GGCTAGCTACAACGA GGCTGGAG CTCCAGCC G GTACATGC
2370 TCGCATGT GGCTAGCTACAACGA ACGGCTGG CCAGCCGT A ACATGCGA
2372 TGTCGCAT GGCTAGCTACAACGA GTACGGCT AGCCGTAC A ATGCGACA
2374 ACTGTCGC GGCTAGCTACAACGA ATGTACGG CCGTACAT G GCGACAGT
2377 CGAACTGT GGCTAGCTACAACGA CGCATGTA TACATGCG A ACAGTTCG
2380 CCACGAAC GGCTAGCTACAACGA TGTCGCAT ATGCGACA G GTTCGTGG
2384 TGAGCCAC GGCTAGCTACAACGA GAACTGTC GACAGTTC G GTGGCTCA
2387 AGGTGAGC GGCTAGCTACAACGA CACGAACT AGTTCGTG G GCTCACCT
2391 CTGCAGGT GGCTAGCTACAACGA GAGCCACG CGTGGCTC A ACCTGCAG
Table 16
2395 TCTCCTGC GGCTAGCTACAACGA AGGTGAGC GCTCACCT G GCAGGAGA
2402 GGGCTGGT GGCTAGCTACAACGA CTCCTGCA TGCAGGAG A ACCAGCCC
2406 CAGCGGGC GGCTAGCTACAACGA TGGTCTCC GGAGACCA G GCCCGCTG
2410 CCCTCAGC GGCTAGCTACAACGA GGGCTGGT ACCAGCCC G GCTGAGGG
2418 GACGGCAT GGCTAGCTACAACGA CCCTCAGC GCTGAGGG A ATGCCGTC
2420 ACGACGGC GGCTAGCTACAACGA ATCCCTCA TGAGGGAT G GCCGTCGT
2423 ATGACGAC GGCTAGCTACAACGA GGCATCCC GGGATGCC G GTCGTCAT
2426 TCGATGAC GGCTAGCTACAACGA GACGGCAT ATGCCGTC G GTCATCGA
2429 TGCTCGAT GGCTAGCTACAACGA GACGACGG CCGTCGTC A ATCGAGCA
2434 AGCTCTGC GGCTAGCTACAACGA TCGATGAC GTCATCGA G GCAGAGCT
2439 GGAGGAGC GGCTAGCTACAACGA TCTGCTCG CGAGCAGA G GCTCCTCC
2451 GGCCTCAT GGCTAGCTACAACGA TCAGGGAG CTCCCTGA A ATGAGGCC
2456 CTGCTGGC GGCTAGCTACAACGA CTCATTCA TGAATGAG G GCCAGCAG
2460 GCCACTGC GGCTAGCTACAACGA TGGCCTCA TGAGGCCA G GCAGTGGC
2463 GAGGCCAC GGCTAGCTACAACGA TGCTGGCC GGCCAGCA G GTGGCCTC
2466 GAAGAGGC GGCTAGCTACAACGA CACTGCTG CAGCAGTG G GCCTCTTC
2475 GAAGACGT GGCTAGCTACAACGA CGAAGAGG CCTCTTCG A ACGTCTTC
2477 AGGAAGAC GGCTAGCTACAACGA GTCGAAGA TCTTCGAC G GTCTTCCT
2485 TGAAGCGT GGCTAGCTACAACGA AGGAAGAC GTCTTCCT A ACGCTTCA
2487 CATGAAGC GGCTAGCTACAACGA GTAGGAAG CTTCCTAC G GCTTCATG
2492 TGGCACAT GGCTAGCTACAACGA GAAGCGTA TACGCTTC A ATGTGCCA
2494 GGTGGCAC GGCTAGCTACAACGA ATGAAGCG CGCTTCAT G GTGCCACC
2496 GTGGTGGC GGCTAGCTACAACGA ACATGAAG CTTCATGT G GCCACCAC
2499 GGCGTGGT GGCTAGCTACAACGA GGCACATG CATGTGCC A ACCACGCC
2502 CACGGCGT GGCTAGCTACAACGA GGTGGCAC GTGCCACC A ACGCCGTG
2504 CGCACGGC GGCTAGCTACAACGA GTGGTGGC GCCACCAC G GCCGTGCG
2507 ATGCGCAC GGCTAGCTACAACGA GGCGTGGT ACCACGCC G GTGCGCAT
2509 TGATGCGC GGCTAGCTACAACGA ACGGCGTG CACGCCGT G GCGCATCA
2511 CCTGATGC GGCTAGCTACAACGA GCACGGCG CGCCGTGC G GCATCAGG
2513 CCCCTGAT GGCTAGCTACAACGA GCGCACGG CCGTGCGC A ATCAGGGG
2520 GGACTTGC GGCTAGCTACAACGA CCCTGATG CATCAGGG G GCAAGTCC
Table 16
2524 CGTAGGAC GGCTAGCTACAACGA TTGCCCCT AGGGGCAA G GTCCTACG
2529 CTGGACGT GGCTAGCTACAACGA AGGACTTG CAAGTCCT A ACGTCCAG
2531 CACTGGAC GGCTAGCTACAACGA GTAGGACT AGTCCTAC G GTCCAGTG
2536 CCTGGCAC GGCTAGCTACAACGA TGGACGTA TACGTCCA G GTGCCAGG
2538 CCCCTGGC GGCTAGCTACAACGA ACTGGACG CGTCCAGT G GCCAGGGG
2546 TGCGGGAT GGCTAGCTACAACGA CCCCTGGC GCCAGGGG A ATCCCGCA
2551 AGCCCTGC GGCTAGCTACAACGA GGGATCCC GGGATCCC G GCAGGGCT
2556 GATGGAGC GGCTAGCTACAACGA CCTGCGGG CCCGCAGG G GCTCCATC
2561 GAGAGGAT GGCTAGCTACAACGA GGAGCCCT AGGGCTCC A ATCCTCTC
2570 AGCAGCGT GGCTAGCTACAACGA GGAGAGGA TCCTCTCC A ACGCTGCT
2572 AGAGCAGC GGCTAGCTACAACGA GTGGAGAG CTCTCCAC G GCTGCTCT
2575 TGCAGAGC GGCTAGCTACAACGA AGCGTGGA TCCACGCT G GCTCTGCA
2580 CAGGCTGC GGCTAGCTACAACGA AGAGCAGC GCTGCTCT G GCAGCCTG
2583 GCACAGGC GGCTAGCTACAACGA TGCAGAGC GCTCTGCA G GCCTGTGC
2587 CGTAGCAC GGCTAGCTACAACGA AGGCTGCA TGCAGCCT G GTGCTACG
2589 GCCGTAGC GGCTAGCTACAACGA ACAGGCTG CAGCCTGT G GCTACGGC
2592 GTCGCCGT GGCTAGCTACAACGA AGCACAGG CCTGTGCT A ACGGCGAC
2595 CATGTCGC GGCTAGCTACAACGA CGTAGCAC GTGCTACG G GCGACATG
2598 CTCCATGT GGCTAGCTACAACGA CGCCGTAG CTACGGCG A ACATGGAG
2600 TTCTCCAT GGCTAGCTACAACGA GTCGCCGT ACGGCGAC A ATGGAGAA
2607 CAGCTTGT GGCTAGCTACAACGA TCTCCATG CATGGAGA A ACAAGCTG
2611 CAAACAGC GGCTAGCTACAACGA TTGTTCTC GAGAACAA G GCTGTTTG
2614 CCGCAAAC GGCTAGCTACAACGA AGCTTGTT AACAAGCT G GTTTGCGG
2618 ATCCCCGC GGCTAGCTACAACGA AAACAGCT AGCTGTTT G GCGGGGAT
2624 CGCCGAAT GGCTAGCTACAACGA CCCCGCAA TTGCGGGG A ATTCGGCG
2629 CGTCCCGC GGCTAGCTACAACGA CGAATCCC GGGATTCG G GCGGGACG
2634 CAGCCCGT GGCTAGCTACAACGA CCCGCCGA TCGGCGGG A ACGGGCTG
2638 GGAGCAGC GGCTAGCTACAACGA CCGTCCCG CGGGACGG G GCTGCTCC
2641 GCAGGAGC GGCTAGCTACAACGA AGCCCGTC GACGGGCT G GCTCCTGC
2647 CCAAACGC GGCTAGCTACAACGA AGGAGCAG CTGCTCCT G GCGTTTGG
2649 CACCAAAC GGCTAGCTACAACGA GCAGGAGC GCTCCTGC G GTTTGGTG
Table 16
2654 TCATCCAC GGCTAGCTACAACGA CAAACGCA TGCGTTTG G GTGGATGA
2658 GAAATCAT GGCTAGCTACAACGA CCACCAAA TTTGGTGG A ATGATTTC
2661 CAAGAAAT GGCTAGCTACAACGA CATCCACC GGTGGATG A ATTTCTTG
2668 TCACCAAC GGCTAGCTACAACGA AAGAAATC GATTTCTT G GTTGGTGA
2672 GGTGTCAC GGCTAGCTACAACGA CAACAAGA TCTTGTTG G GTGACACC
2675 TGAGGTGT GGCTAGCTACAACGA CACCAACA TGTTGGTG A ACACCTCA
2677 GGTGAGGT GGCTAGCTACAACGA GTCACCAA TTGGTGAC A ACCTCACC
2682 GGTGAGGT GGCTAGCTACAACGA GAGGTGTC GACACCTC A ACCTCACC
2687 GCGTGGGT GGCTAGCTACAACGA GAGGTGAG CTCACCTC A ACCCACGC
2691 TTTCGCGT GGCTAGCTACAACGA GGGTGAGG CCTCACCC A ACGCGAAA
2693 GTTTTCGC GGCTAGCTACAACGA GTGGGTGA TCACCCAC G GCGAAAAC
2699 AGGAAGGT GGCTAGCTACAACGA TTTCGCGT ACGCGAAA A ACCTTCCT
2711 ACCAGGGT GGCTAGCTACAACGA CCTGAGGA TCCTCAGG A ACCCTGGT
2717 CCTCGGAC GGCTAGCTACAACGA CAGGGTCC GGACCCTG G GTCCGAGG
2724 AGGGACAC GGCTAGCTACAACGA CTCGGACC GGTCCGAG G GTGTCCCT
2726 TCAGGGAC GGCTAGCTACAACGA ACCTCGGA TCCGAGGT G GTCCCTGA
2734 AGCCATAC GGCTAGCTACAACGA TCAGGGAC GTCCCTGA G GTATGGCT
2736 GCAGCCAT GGCTAGCTACAACGA ACTCAGGG CCCTGAGT A ATGGCTGC
2739 CACGCAGC GGCTAGCTACAACGA CATACTCA TGAGTATG G GCTGCGTG
2742 CACCACGC GGCTAGCTACAACGA AGCCATAC GTATGGCT G GCGTGGTG
2744 TTCACCAC GGCTAGCTACAACGA GCAGCCAT ATGGCTGC G GTGGTGAA
2747 AAGTTCAC GGCTAGCTACAACGA CACGCAGC GCTGCGTG G GTGAACTT
2751 CCGCAAGT GGCTAGCTACAACGA TCACCACG CGTGGTGA A ACTTGCGG
2755 TCTTCCGC GGCTAGCTACAACGA AAGTTCAC GTGAACTT G GCGGAAGA
2762 ACCACTGT GGCTAGCTACAACGA CTTCCGCA TGCGGAAG A ACAGTGGT
2765 TTCACCAC GGCTAGCTACAACGA TGTCTTCC GGAAGACA G GTGGTGAA
2768 AAGTTCAC GGCTAGCTACAACGA CACTGTCT AGACAGTG G GTGAACTT
2772 AGGGAAGT GGCTAGCTACAACGA TCACCACT AGTGGTGA A ACTTCCCT
2780 TCTTCTAC GGCTAGCTACAACGA AGGGAAGT ACTTCCCT G GTAGAAGA
2787 GGCCTCGT GGCTAGCTACAACGA CTTCTACA TGTAGAAG A ACGAGGCC
2792 CCCAGGGC GGCTAGCTACAACGA CTCGTCTT AAGACGAG G GCCCTGGG
Table 16
2799 CGTGCCAC GGCTAGCTACAACGA CCAGGGCC GGCCCTGG G GTGGCACG
2802 AGCCGTGC GGCTAGCTACAACGA CACCCAGG CCTGGGTG G GCACGGCT
2804 AAAGCCGT GGCTAGCTACAACGA GCCACCCA TGGGTGGC A ACGGCTTT
2807 ACAAAAGC GGCTAGCTACAACGA CGTGCCAC GTGGCACG G GCTTTTGT
2813 ATCTGAAC GGCTAGCTACAACGA AAAAGCCG CGGCTTTT G GTTCAGAT
2819 GCCGGCAT GGCTAGCTACAACGA CTGAACAA TTGTTCAG A ATGCCGGC
2821 GGGCCGGC GGCTAGCTACAACGA ATCTGAAC GTTCAGAT G GCCGGCCC
2825 CCGTGGGC GGCTAGCTACAACGA CGGCATCT AGATGCCG G GCCCACGG
2829 TAGGCCGT GGCTAGCTACAACGA GGGCCGGC GCCGGCCC A ACGGCCTA
2832 GAATAGGC GGCTAGCTACAACGA CGTGGGCC GGCCCACG G GCCTATTC
2836 AGGGGAAT GGCTAGCTACAACGA AGGCCGTG CACGGCCT A ATTCCCCT
2845 GGCCGCAC GGCTAGCTACAACGA CAGGGGAA TTCCCCTG G GTGCGGCC
2847 CAGGCCGC GGCTAGCTACAACGA ACCAGGGG CCCCTGGT G GCGGCCTG
2850 CAGCAGGC GGCTAGCTACAACGA CGCACCAG CTGGTGCG G GCCTGCTG
2854 CCAGCAGC GGCTAGCTACAACGA AGGCCGCA TGCGGCCT G GCTGCTGG
2857 TATCCAGC GGCTAGCTACAACGA AGCAGGCC GGCCTGCT G GCTGGATA
2862 CCGGGTAT GGCTAGCTACAACGA CCAGCAGC GCTGCTGG A ATACCCGG
2864 GTCCGGGT GGCTAGCTACAACGA ATCCAGCA TGCTGGAT A ACCCGGAC
2870 TCCAGGGT GGCTAGCTACAACGA CCGGGTAT ATACCCGG A ACCCTGGA
2879 CTCTGCAC GGCTAGCTACAACGA CTCCAGGG CCCTGGAG G GTGCAGAG
2881 CGCTCTGC GGCTAGCTACAACGA ACCTCCAG CTGGAGGT G GCAGAGCG
2886 GTAGTCGC GGCTAGCTACAACGA TCTGCACC GGTGCAGA G GCGACTAC
2889 GGAGTAGT GGCTAGCTACAACGA CGCTCTGC GCAGAGCG A ACTACTCC
2892 GCTGGAGT GGCTAGCTACAACGA AGTCGCTC GAGCGACT A ACTCCAGC
2898 GGCATAGC GGCTAGCTACAACGA TGGAGTAG CTACTCCA G GCTATGCC
2901 CCGGGCAT GGCTAGCTACAACGA AGCTGGAG CTCCAGCT A ATGCCCGG
2903 GTCCGGGC GGCTAGCTACAACGA ATAGCTGG CCAGCTAT G GCCCGGAC
2909 ATGGAGGT GGCTAGCTACAACGA CCGGGCAT ATGCCCGG A ACCTCCAT
2915 GCTCTGAT GGCTAGCTACAACGA GGAGGTCC GGACCTCC A ATCAGAGC
2921 AGACTGGC GGCTAGCTACAACGA TCTGATGG CCATCAGA G GCCAGTCT
2925 GGTGAGAC GGCTAGCTACAACGA TGGCTCTG CAGAGCCA G GTCTCACC
Table 16
2930 TTGAAGGT GGCTAGCTACAACGA GAGACTGG CCAGTCTC A ACCTTCAA
2937 GCCGCGGT GGCTAGCTACAACGA TGAAGGTG CACCTTCA A ACCGCGGC
2940 GAAGCCGC GGCTAGCTACAACGA GGTTGAAG CTTCAACC G GCGGCTTC
2943 CTTGAAGC GGCTAGCTACAACGA CGCGGTTG CAACCGCG G GCTTCAAG
2951 CTCCCAGC GGCTAGCTACAACGA CTTGAAGC GCTTCAAG G GCTGGGAG
2961 ACGCATGT GGCTAGCTACAACGA TCCTCCCA TGGGAGGA A ACATGCGT
2963 CGACGCAT GGCTAGCTACAACGA GTTCCTCC GGAGGAAC A ATGCGTCG
2965 TGCGACGC GGCTAGCTACAACGA ATGTTCCT AGGAACAT G GCGTCGCA
2967 TTTGCGAC GGCTAGCTACAACGA GCATGTTC ' GAACATGC G GTCGCAAA
2970 GAGTTTGC GGCTAGCTACAACGA GACGCATG CATGCGTC G GCAAACTC
2974 CAAAGAGT GGCTAGCTACAACGA TTGCGACG CGTCGCAA A ACTCTTTG
2984 CGCAAGAC GGCTAGCTACAACGA CCCAAAGA TCTTTGGG G GTCTTGCG
2989 TCAGCCGC GGCTAGCTACAACGA AAGACCCC GGGGTCTT G GCGGCTGA
2992 ACTTCAGC GGCTAGCTACAACGA CGCAAGAC GTCTTGCG G GCTGAAGT
2998 TGTGACAC GGCTAGCTACAACGA TTCAGCCG CGGCTGAA G GTGTCACA
3000 GCTGTGAC GGCTAGCTACAACGA ACTTCAGC GCTGAAGT G GTCACAGC
3003 CAGGCTGT GGCTAGCTACAACGA GACACTTC GAAGTGTC A ACAGCCTG
3006 AAACAGGC GGCTAGCTACAACGA TGTGACAC GTGTCACA G GCCTGTTT
3010 CCAGAAAC GGCTAGCTACAACGA AGGCTGTG CACAGCCT G GTTTCTGG
3018 CTGCAAAT GGCTAGCTACAACGA CCAGAAAC GTTTCTGG A ATTTGCAG
3022 TCACCTGC GGCTAGCTACAACGA AAATCCAG CTGGATTT G GCAGGTGA
3026 CTGTTCAC GGCTAGCTACAACGA CTGCAAAT ATTTGCAG G GTGAACAG
3030 GAGGCTGT GGCTAGCTACAACGA TCACCTGC GCAGGTGA A ACAGCCTC
3033 CTGGAGGC GGCTAGCTACAACGA TGTTCACC GGTGAACA G GCCTCCAG
3041 CACACCGT GGCTAGCTACAACGA CTGGAGGC GCCTCCAG A ACGGTGTG
3044 GTGCACAC GGCTAGCTACAACGA CGTCTGGA TCCAGACG G GTGTGCAC
3046 TGGTGCAC GGCTAGCTACAACGA ACCGTCTG CAGACGGT G GTGCACCA
3048 GTTGGTGC GGCTAGCTACAACGA ACACCGTC GACGGTGT G GCACCAAC
3050 ATGTTGGT GGCTAGCTACAACGA GCACACCG CGGTGTGC A ACCAACAT
3054 GTAGATGT GGCTAGCTACAACGA TGGTGCAC GTGCACCA A ACATCTAC
3056 TTGTAGAT GGCTAGCTACAACGA GTTGGTGC GCACCAAC A ATCTACAA
Table 16
3060 GATCTTGT GGCTAGCTACAACGA AGATGTTG CAACATCT A ACAAGATC
3065 AGGAGGAT GGCTAGCTACAACGA CTTGTAGA TCTACAAG A ATCCTCCT
3073 CCTGCAGC GGCTAGCTACAACGA AGGAGGAT ATCCTCCT G GCTGCAGG
3076 ACGCCTGC GGCTAGCTACAACGA AGCAGGAG CTCCTGCT G GCAGGCGT
3080 CTGTACGC GGCTAGCTACAACGA CTGCAGCA TGCTGCAG G GCGTACAG
3082 ACCTGTAC GGCTAGCTACAACGA GCCTGCAG CTGCAGGC G GTACAGGT
3084 AAACCTGT GGCTAGCTACAACGA ACGCCTGC GCAGGCGT A ACAGGTTT
3088 CGTGAAAC GGCTAGCTACAACGA CTGTACGC GCGTACAG G GTTTCACG
3093 ACATGCGT GGCTAGCTACAACGA GAAACCTG CAGGTTTC A ACGCATGT
3095 ACACATGC GGCTAGCTACAACGA GTGAAACC GGTTTCAC G GCATGTGT
3097 GCACACAT GGCTAGCTACAACGA GCGTGAAA TTTCACGC A ATGTGTGC
3099 CAGCACAC GGCTAGCTACAACGA ATGCGTGA TCACGCAT G GTGTGCTG
3101 TGCAGCAC GGCTAGCTACAACGA ACATGCGT ACGCATGT G GTGCTGCA
3103 GCTGCAGC GGCTAGCTACAACGA ACACATGC GCATGTGT G GCTGCAGC
3106 GGAGCTGC GGCTAGCTACAACGA AGCACACA TGTGTGCT G GCAGCTCC
3109 ATGGGAGC GGCTAGCTACAACGA TGCAGCAC GTGCTGCA G GCTCCCAT
3115 GATGAAAT GGCTAGCTACAACGA GGGAGCTG CAGCTCCC A ATTTCATC
3120 TTGCTGAT GGCTAGCTACAACGA GAAATGGG CCCATTTC A ATCAGCAA
3124 AAACTTGC GGCTAGCTACAACGA TGATGAAA TTTCATCA G GCAAGTTT
3128 TTCCAAAC GGCTAGCTACAACGA TTGCTGAT ATCAGCAA G GTTTGGAA
3138 TGTGGGGT GGCTAGCTACAACGA TCTTCCAA TTGGAAGA A ACCCCACA
3143 AAAAATGT GGCTAGCTACAACGA GGGGTTCT AGAACCCC A ACATTTTT
3145 GGAAAAAT GGCTAGCTACAACGA GTGGGGTT AACCCCAC A ATTTTTCC
3154 TGACGCGC GGCTAGCTACAACGA AGGAAAAA TTTTTCCT G GCGCGTCA
3156 GATGACGC GGCTAGCTACAACGA GCAGGAAA TTTCCTGC G GCGTCATC
3158 GAGATGAC GGCTAGCTACAACGA GCGCAGGA TCCTGCGC G GTCATCTC
3161 TCAGAGAT GGCTAGCTACAACGA GACGCGCA TGCGCGTC A ATCTCTGA
3168 GGCCGTGT GGCTAGCTACAACGA CAGAGATG CATCTCTG A ACACGGCC
3170 GAGGCCGT GGCTAGCTACAACGA GTCAGAGA TCTCTGAC A ACGGCCTC
3173 AGGGAGGC GGCTAGCTACAACGA CGTGTCAG CTGACACG G GCCTCCCT
3183 GGAGTAGC GGCTAGCTACAACGA AGAGGGAG CTCCCTCT G GCTACTCC
Table 16
Table 16
3317 ACGTAGGT GGCTAGCTACAACGA GACACGGT ACCGTGTC A ACCTACGT
3321 TGGCACGT GGCTAGCTACAACGA AGGTGACA TGTCACCT A ACGTGCCA
3323 AGTGGCAC GGCTAGCTACAACGA GTAGGTGA TCACCTAC G GTGCCACT
3325 GGAGTGGC GGCTAGCTACAACGA ACGTAGGT ACCTACGT G GCCACTCC
3328 CCAGGAGT GGCTAGCTACAACGA GGCACGTA TACGTGCC A ACTCCTGG
3337 TGAGTGAC GGCTAGCTACAACGA CCCAGGAG CTCCTGGG G GTCACTCA
3340 TCCTGAGT GGCTAGCTACAACGA GACCCCAG CTGGGGTC A ACTCAGGA
3347 TGGGCTGT GGCTAGCTACAACGA CCTGAGTG CACTCAGG A ACAGCCCA
3350 GTCTGGGC GGCTAGCTACAACGA TGTCCTGA TCAGGACA G GCCCAGAC
3356 AGCTGCGT GGCTAGCTACAACGA CTGGGCTG CAGCCCAG A ACGCAGCT
3358 TCAGCTGC GGCTAGCTACAACGA GTCTGGGC GCCCAGAC G GCAGCTGA
3361 GACTCAGC GGCTAGCTACAACGA TGCGTCTG CAGACGCA G GCTGAGTC
3366 CTTCCGAC GGCTAGCTACAACGA TCAGCTGC GCAGCTGA G GTCGGAAG
3373 CCGGGAGC GGCTAGCTACAACGA TTCCGACT AGTCGGAA G GCTCCCGG
3383 AGCGTCGT GGCTAGCTACAACGA CCCCGGGA TCCCGGGG A ACGACGCT
3386 GTCAGCGT GGCTAGCTACAACGA CGTCCCCG CGGGGACG A ACGCTGAC
3388 CAGTCAGC GGCTAGCTACAACGA GTCGTCCC GGGACGAC G GCTGACTG
3392 AGGGCAGT GGCTAGCTACAACGA CAGCGTCG CGACGCTG A ACTGCCCT
3395 TCCAGGGC GGCTAGCTACAACGA AGTCAGCG CGCTGACT G GCCCTGGA
3404 GCTGCGGC GGCTAGCTACAACGA CTCCAGGG CCCTGGAG G GCCGCAGC
3407 TTGGCTGC GGCTAGCTACAACGA GGCCTCCA TGGAGGCC G GCAGCCAA
3410 GGGTTGGC GGCTAGCTACAACGA TGCGGCCT AGGCCGCA G GCCAACCC
3414 TGCCGGGT GGCTAGCTACAACGA TGGCTGCG CGCAGCCA A ACCCGGCA
3419 GGCAGTGC GGCTAGCTACAACGA CGGGTTGG CCAACCCG G GCACTGCC
3421 AGGGCAGT GGCTAGCTACAACGA GCCGGGTT AACCCGGC A ACTGCCCT
3424 CTGAGGGC GGCTAGCTACAACGA AGTGCCGG CCGGCACT G GCCCTCAG
3432 CTTGAAGT GGCTAGCTACAACGA CTGAGGGC GCCCTCAG A ACTTCAAG
3440 AGGATGGT GGCTAGCTACAACGA CTTGAAGT ACTTCAAG A ACCATCCT
3443 TCCAGGAT GGCTAGCTACAACGA GGTCTTGA TCAAGACC A ATCCTGGA
3450 CCATCAGT GGCTAGCTACAACGA CCAGGATG CATCCTGG A ACTGATGG
3454 GTGGCCAT GGCTAGCTACAACGA CAGTCCAG CTGGACTG A ATGGCCAC
Table 16
3457 CGGGTGGC GGCTAGCTACAACGA CATCAGTC GACTGATG G GCCACCCG
3460 GGGCGGGT GGCTAGCTACAACGA GGCCATCA TGATGGCC A ACCCGCCC
3464 CTGTGGGC GGCTAGCTACAACGA GGGTGGCC GGCCACCC G GCCCACAG
3468 CTGGCTGT GGCTAGCTACAACGA GGGCGGGT ACCCGCCC A ACAGCCAG
3471 GGCCTGGC GGCTAGCTACAACGA TGTGGGCG CGCCCACA G GCCAGGCC
3476 CTCTCGGC GGCTAGCTACAACGA CTGGCTGT ACAGCCAG G GCCGAGAG
3483 GTGTCTGC GGCTAGCTACAACGA TCTCGGCC GGCCGAGA G GCAGACAC
3487 GCTGGTGT GGCTAGCTACAACGA CTGCTCTC GAGAGCAG A ACACCAGC
3489 CTGCTGGT GGCTAGCTACAACGA GTCTGCTC GAGCAGAC A ACCAGCAG
3493 AGGGCTGC GGCTAGCTACAACGA TGGTGTCT AGACACCA G GCAGCCCT
3496 GACAGGGC GGCTAGCTACAACGA TGCTGGTG CACCAGCA G GCCCTGTC
3501 GGCGTGAC GGCTAGCTACAACGA AGGGCTGC GCAGCCCT G GTCACGCC
3504 CCCGGCGT GGCTAGCTACAACGA GACAGGGC GCCCTGTC A ACGCCGGG
3506 AGCCCGGC GGCTAGCTACAACGA GTGACAGG CCTGTCAC G GCCGGGCT
3511 CGTAGAGC GGCTAGCTACAACGA CCGGCGTG CACGCCGG G GCTCTACG
3516 TGGGACGT GGCTAGCTACAACGA AGAGCCCG CGGGCTCT A ACGTCCCA
3518 CCTGGGAC GGCTAGCTACAACGA GTAGAGCC GGCTCTAC G GTCCCAGG
3535 TGGGCCGC GGCTAGCTACAACGA CCCTCCCT AGGGAGGG G GCGGCCCA
3538 GTGTGGGC GGCTAGCTACAACGA CGCCCCTC GAGGGGCG G GCCCACAC
3542 CTGGGTGT GGCTAGCTACAACGA GGGCCGCC GGCGGCCC A ACACCCAG
3544 GCCTGGGT GGCTAGCTACAACGA GTGGGCCG CGGCCCAC A ACCCAGGC
3550 GTGCGGGC GGCTAGCTACAACGA CTGGGTGT ACACCCAG G GCCCGCAC
3554 AGCGGTGC GGCTAGCTACAACGA GGGCCTGG CCAGGCCC G GCACCGCT
3556 CCAGCGGT GGCTAGCTACAACGA GCGGGCCT AGGCCCGC A ACCGCTGG
3559 CTCCCAGC GGCTAGCTACAACGA GGTGCGGG CCCGCACC G GCTGGGAG
3566 CCTCAGAC GGCTAGCTACAACGA TCCCAGCG CGCTGGGA G GTCTGAGG
3573 ACTCAGGC GGCTAGCTACAACGA CTCAGACT AGTCTGAG G GCCTGAGT
3579 ACACTCAC GGCTAGCTACAACGA TCAGGCCT AGGCCTGA G GTGAGTGT
3583 CCAAACAC GGCTAGCTACAACGA TCACTCAG CTGAGTGA G GTGTTTGG
3585 GGCCAAAC GGCTAGCTACAACGA ACTCACTC GAGTGAGT G GTTTGGCC
3590 GCCTCGGC GGCTAGCTACAACGA CAAACACT AGTGTTTG G GCCGAGGC
Table 16
3596 ATGCAGGC GGCTAGCTACAACGA CTCGGCCA TGGCCGAG G GCCTGCAT
3600 GGACATGC GGCTAGCTACAACGA AGGCCTCG ,CGAGGCCT G GCATGTCC
3602 CCGGACAT GGCTAGCTACAACGA GCAGGCCT AGGCCTGC A ATGTCCGG
3604 AGCCGGAC GGCTAGCTACAACGA ATGCAGGC GCCTGCAT G GTCCGGCT
3609 CCTTCAGC GGCTAGCTACAACGA CGGACATG CATGTCCG G GCTGAAGG
3616 CACTCAGC GGCTAGCTACAACGA CTTCAGCC GGCTGAAG G GCTGAGTG
3621 CCGGACAC GGCTAGCTACAACGA TCAGCCTT AAGGCTGA G GTGTCCGG
3623 AGCCGGAC GGCTAGCTACAACGA ACTCAGCC GGCTGAGT G GTCCGGCT
3628 GCCTCAGC GGCTAGCTACAACGA CGGACACT AGTGTCCG G GCTGAGGC
3634 GCTCAGGC GGCTAGCTACAACGA CTCAGCCG CGGCTGAG G GCCTGAGC
3640 ACACTCGC GGCTAGCTACAACGA TCAGGCCT AGGCCTGA G GCGAGTGT
3644 CTGGACAC GGCTAGCTACAACGA TCGCTCAG CTGAGCGA G GTGTCCAG
3646 GGCTGGAC GGCTAGCTACAACGA ACTCGCTC GAGCGAGT G GTCCAGCC
3651 CCCTTGGC GGCTAGCTACAACGA TGGACACT AGTGTCCA G GCCAAGGG
3658 CACTCAGC GGCTAGCTACAACGA CCTTGGCT AGCCAAGG G GCTGAGTG
3663 CTGGACAC GGCTAGCTACAACGA TCAGCCCT AGGGCTGA G GTGTCCAG
3665 TGCTGGAC GGCTAGCTACAACGA ACTCAGCC GGCTGAGT G GTCCAGCA
3670 AGGTGTGC GGCTAGCTACAACGA TGGACACT AGTGTCCA G GCACACCT
3672 GCAGGTGT GGCTAGCTACAACGA GCTGGACA TGTCCAGC A ACACCTGC
3674 CGGCAGGT GGCTAGCTACAACGA GTGCTGGA TCCAGCAC A ACCTGCCG
3678 AAGACGGC GGCTAGCTACAACGA AGGTGTGC GCACACCT G GCCGTCTT
3681 GTGAAGAC GGCTAGCTACAACGA GGCAGGTG CACCTGCC G GTCTTCAC
3687 GGGGAAGT GGCTAGCTACAACGA GAAGACGG CCGTCTTC A ACTTCCCC
3695 CAGCCTGT GGCTAGCTACAACGA GGGGAAGT ACTTCCCC A ACAGGCTG
3699 GCGCCAGC GGCTAGCTACAACGA CTGTGGGG CCCCACAG G GCTGGCGC
3703 CCGAGCGC GGCTAGCTACAACGA CAGCCTGT ACAGGCTG G GCGCTCGG
3705 AGCCGAGC GGCTAGCTACAACGA GCCAGCCT AGGCTGGC G GCTCGGCT
3710 GGTGGAGC GGCTAGCTACAACGA CGAGCGCC GGCGCTCG G GCTCCACC
3715 CCTGGGGT GGCTAGCTACAACGA GGAGCCGA TCGGCTCC A ACCCCAGG
3723 AAGCTGGC GGCTAGCTACAACGA CCTGGGGT ACCCCAGG G GCCAGCTT
3727 GGAAAAGC GGCTAGCTACAACGA TGGCCCTG CAGGGCCA G GCTTTTCC
Table 16
3737 CTCCTGGT GGCTAGCTACAACGA GAGGAAAA TTTTCCTC A ACCAGGAG
3744 AGCCGGGC GGCTAGCTACAACGA TCCTGGTG CACCAGGA G GCCCGGCT
3749 GTGGAAGC GGCTAGCTACAACGA CGGGCTCC GGAGCCCG G GCTTCCAC
3755 TGGGGAGT GGCTAGCTACAACGA GGAAGCCG CGGCTTCC A ACTCCCCA
3762 TCCTATGT GGCTAGCTACAACGA GGGGAGTG CACTCCCC A ACATAGGA
3764 ATTCCTAT GGCTAGCTACAACGA GTGGGGAG CTCCCCAC A ATAGGAAT
3770 TGGACTAT GGCTAGCTACAACGA TCCTATGT ACATAGGA A ATAGTCCA
3773 GGATGGAC GGCTAGCTACAACGA TATTCCTA TAGGAATA G GTCCATCC
3777 CTGGGGAT GGCTAGCTACAACGA GGACTATT AATAGTCC A ATCCCCAG
3785 TGGCGAAT GGCTAGCTACAACGA CTGGGGAT ATCCCCAG A ATTCGCCA
3789 ACAATGGC GGCTAGCTACAACGA GAATCTGG CCAGATTC G GCCATTGT
3792 TGAACAAT GGCTAGCTACAACGA GGCGAATC GATTCGCC A ATTGTTCA
3795 GGGTGAAC GGCTAGCTACAACGA AATGGCGA TCGCCATT G GTTCACCC
3799 CGAGGGGT GGCTAGCTACAACGA GAACAATG CATTGTTC A ACCCCTCG
3806 GGCAGGGC GGCTAGCTACAACGA GAGGGGTG CACCCCTC G GCCCTGCC
3811 AGGAGGGC GGCTAGCTACAACGA AGGGCGAG CTCGCCCT G GCCCTCCT
3821 TGGAAGGC GGCTAGCTACAACGA AAAGGAGG CCTCCTTT G GCCTTCCA
3828 GTGGGGGT GGCTAGCTACAACGA GGAAGGCA TGCCTTCC A ACCCCCAC
3834 TGGATGGT GGCTAGCTACAACGA GGGGGTGG CCACCCCC A ACCATCCA
3837 ACCTGGAT GGCTAGCTACAACGA GGTGGGGG CCCCCACC A ATCCAGGT
3843 GTCTCCAC GGCTAGCTACAACGA CTGGATGG CCATCCAG G GTGGAGAC
3849 CTCAGGGT GGCTAGCTACAACGA CTCCACCT AGGTGGAG A ACCCTGAG
3861 CCCAGGGT GGCTAGCTACAACGA CCTTCTCA TGAGAAGG A ACCCTGGG
3870 CCCAGAGC GGCTAGCTACAACGA TCCCAGGG CCCTGGGA G GCTCTGGG
3879 CTCCAAAT GGCTAGCTACAACGA TCCCAGAG CTCTGGGA A ATTTGGAG
3886 TTGGTCAC GGCTAGCTACAACGA TCCAAATT AATTTGGA G GTGACCAA
3889 CCTTTGGT GGCTAGCTACAACGA CACTCCAA TTGGAGTG A ACCAAAGG
3896 GGGCACAC GGCTAGCTACAACGA CTTTGGTC GACCAAAG G GTGTGCCC
3898 CAGGGCAC GGCTAGCTACAACGA ACCTTTGG CCAAAGGT G GTGCCCTG
3900 TACAGGGC GGCTAGCTACAACGA ACACCTTT AAAGGTGT G GCCCTGTA
3905 CTGTGTAC GGCTAGCTACAACGA AGGGCACA TGTGCCCT G GTACACAG
Table 16
3907 GCCTGTGT GGCTAGCTACAACGA ACAGGGCA TGCCCTGT A ACACAGGC
3909 TCGCCTGT GGCTAGCTACAACGA GTACAGGG CCCTGTAC A ACAGGCGA
3913 GTCCTCGC GGCTAGCTACAACGA CTGTGTAC GTACACAG G GCGAGGAC
3919 TGCAGGGT GGCTAGCTACAACGA CCTCGCCT AGGCGAGG A ACCCTGCA
3924 CCAGGTGC GGCTAGCTACAACGA AGGGTCCT AGGACCCT G GCACCTGG
3926 ATCCAGGT GGCTAGCTACAACGA GCAGGGTC GACCCTGC A ACCTGGAT
3932 ACCCCCAT GGCTAGCTACAACGA CCAGGTGC GCACCTGG A ATGGGGGT
3938 ACAGGGAC GGCTAGCTACAACGA CCCCATCC GGATGGGG G GTCCCTGT
3944 TGACCCAC GGCTAGCTACAACGA AGGGACCC GGGTCCCT G GTGGGTCA
3948 AATTTGAC GGCTAGCTACAACGA CCACAGGG CCCTGTGG G GTCAAATT
3953 CCCCCAAT GGCTAGCTACAACGA TTGACCCA TGGGTCAA A ATTGGGGG
3964 CACAGCAC GGCTAGCTACAACGA CTCCCCCC GGGGGGAG G GTGCTGTG
3966 CCCACAGC GGCTAGCTACAACGA ACCTCCCC GGGGAGGT G GCTGTGGG
3969 ACTCCCAC GGCTAGCTACAACGA AGCACCTC GAGGTGCT G GTGGGAGT
3975 TATTTTAC GGCTAGCTACAACGA TCCCACAG CTGTGGGA G GTAAAATA
3980 TTCAGTAT GGCTAGCTACAACGA TTTACTCC GGAGTAAA A ATACTGAA
3982 TATTCAGT GGCTAGCTACAACGA ATTTTACT AGTAAAAT A ACTGAATA
3987 TCATATAT GGCTAGCTACAACGA TCAGTATT AATACTGA A ATATATGA
3989 ACTCATAT GGCTAGCTACAACGA ATTCAGTA TACTGAAT A ATATGAGT
3991 AAACTCAT GGCTAGCTACAACGA ATATTCAG CTGAATAT A ATGAGTTT
3995 TGAAAAAC GGCTAGCTACAACGA TCATATAT ATATATGA G GTTTTTCA
4003 TTCAAAAC GGCTAGCTACAACGA TGAAAAAC GTTTTTCA G GTTTTGAA
Seql = TERT (Homo sapiens telomerase reverse transcriptase (TERT) mRNA, 4015 bp); Nakamura et al, Science 277 (5328), 955-959 (1997) Cut Site = R/Y (Purine/Pyrimidine)
Stem Length = 8 . Core Sequence = GGCTAGCTACAACGA
Table 17
Table 17: Anti-TERT HH and G-Cleaver Ribozymes
Table 18 Table 18: Human BACE Hammerhead Ribozyme and Target Sequence
Pos Substrate Seq ID Ribozyme Rz Seq ID
9 CCACGCGU C CGCAGCCC 1 GGGCUGCG CUGAUGAG X CGAA ACGCGUGG 1776
47 AGCUGGAU U AUGGUGGC 2 GCCACCAU CUGAUGAG X CGAA AUCCAGCU 1777
48 GCUGGAUU A UGGUGGCC 3 GGCCACCA CUGAUGAG X CGAA AAUCCAGC 1778
93 GGAGCCCU U GCCCCUGC 4 GCAGGGGC CUGAUGAG X CGAA AGGGCUCC 1779
163 CCGCCCCU C CCAGCCCC 5 GGGGCUGG CUGAUGAG X CGAA AGGGGCGG 1780
221 GCCGAUGU A GCGGGCUC 6 GAGCCCGC CUGAUGAG X CGAA ACAUCGGC 1781
229 AGCGGGCU C CGGAUCCC 7 GGGAUCCG CUGAUGAG X CGAA AGCCCGCU 1782
235 CUCCGGAU C CCAGCCUC 8 GAGGCUGG CUGAUGAG X CGAA AUCCGGAG 1783
243 CCCAGCCU C UCCCCUGC 9 GCAGGGGA CUGAUGAG X CGAA AGGCUGGG 1784
245 CAGCCUCU C CCCUGCUC 10 GAGCAGGG CUGAUGAG X CGAA AGAGGCUG 1785
253 CCCCUGCU C CCGUGCUC 11 GAGCACGG CUGAUGAG X CGAA AGCAGGGG 1786
261 CCCGUGCU C UGCGGAUC 12 GAUCCGCA CUGAUGAG X CGAA AGCACGGG 1787
269 CUGCGGAU C UCCCCUGA 13 UCAGGGGA CUGAUGAG X CGAA AUCCGCAG 1788
271 GCGGAUCU C CCCUGACC 14 GGUCAGGG CUGAUGAG X CGAA AGAUCCGC 1789
283 UGACCGCU C UCCACAGC 15 GCUGUGGA CUGAUGAG X CGAA AGCGGUCA 1790
285 ACCGCUCU c CACAGCCC 16 GGGCUGUG CUGAUGAG X CGAA AGAGCGGU 1791
334 CCUGGCGU c CUGAUGCC 17 GGCAUCAG CUGAUGAG X CGAA ACGCCAGG 1792
351 CCCAAGCU c CCUCUCCU 18 AGGAGAGG CUGAUGAG X CGAA AGCUUGGG 1793
355 AGCUCCCU c UCCUGAGA 19 UCUCAGGA CUGAUGAG X CGAA AGGGAGCU 1794
357 CUCCCUCU c CUGAGAAG 20 CUUCUCAG CUGAUGAG X CGAA AGAGGGAG 1795
386 CCCAGACU U GGGGGCAG 21 CUGCCCCC CUGAUGAG X CGAA AGUCUGGG 1796
477 CCCUGGCU c CUGCUGUG 22 CACAGCAG CUGAUGAG X CGAA AGCCAGGG 1797
531 CACGGCAU c CGGCUGCC 23 GGCAGCCG CUGAUGAG X CGAA AUGCCGUG 1798
632 GGGCAGCU u UGUGGAGA 24 UCUCCACA CUGAUGAG X CGAA AGCUGCCC 1799
633 GGCAGCUU u GUGGAGAU 25 AUCUCCAC CUGAUGAG X CGAA AAGCUGCC 1800
665 GGGCAAGU c GGGGCAGG 26 CCUGCCCC CUGAUGAG X CGAA ACUUGCCC 1801
677 GCAGGGCU A CUACGUGG 27 CCACGUAG CUGAUGAG X CGAA AGCCCUGC 1802
680 GGGCUACU A CGUGGAGA 28 UCUCCACG CUGAUGAG X CGAA AGUAGCCC 1803
717 CAGACGCU C AACAUCCU 29 AGGAUGUU CUGAUGAG X CGAA AGCGUCUG 1804
723 CUCAACAU C CUGGUGGA 30 UCCACCAG CUGAUGAG X CGAA AUGUUGAG 1805
733 UGGUGGAU A CAGGCAGC 31 GCUGCCUG CUGAUGAG X CGAA AUCCACCA 1806
745 GCAGCAGU A ACUUUGCA 32 UGCAAAGU CUGAUGAG X CGAA ACUGCUGC 1807
749 CAGUAACU U UGCAGUGG 33 CCACUGCA CUGAUGAG X CGAA AGUUACUG 1808
750 AGUAACUU U GCAGUGGG 34 CCCACUGC CUGAUGAG X CGAA AAGUUACU 1809
776 CCACCCCU U CCUGCAUC 35 GAUGCAGG CUGAUGAG X CGAA AGGGGUGG 1810
777 CACCCCUU C CUGCAUCG 36 CGAUGCAG CUGAUGAG X CGAA AAGGGGUG 1811
784 UCCUGCAU C GCUACUAC 37 GUAGUAGC CUGAUGAG X CGAA AUGCAGGA 1812
788 GCAUCGCU A CUACCAGA 38 UCUGGUAG CUGAUGAG X CGAA AGCGAUGC 1813
791 UCGCUACU A CCAGAGGC 39 GCCUCUGG CUGAUGAG X CGAA AGUAGCGA 1814
806 GCAGCUGU C CAGCACAU 40 AUGUGCUG CUGAUGAG X CGAA ACAGCUGC 1815
815 CAGCACAU A CCGGGACC 41 GGUCCCGG CUGAUGAG X CGAA AUGUGCUG 1816
825 CGGGACCU C CGGAAGGG 42 CCCUUCCG CUGAUGAG X CGAA AGGUCCCG 1817
839 GGGUGUGU A UGUGCCCU 43 AGGGCACA CUGAUGAG X CGAA ACACACCC 1818
848 UGUGCCCU A CACCCAGG 44 CCUGGGUG CUGAUGAG X CGAA AGGGCACA 1819
891 GACCUGGU A AGCAUCCC 45 GGGAUGCU CUGAUGAG X CGAA ACCAGGUC 1820
897 GUAAGCAU C CCCCAUGG 46 CCAUGGGG CUGAUGAG X CGAA AUGCUUAC 1821
915 CCCAACGU C ACUGUGCG 47 CGCACAGU CUGAUGAG X CGAA ACGUUGGG 1822
Table 18
933 GCCAACAU U GCUGCCAU 48 AUGGCAGC CUGAUGAG X CGAA AUGUUGGC 1823
942 GCUGCCAU C ACUGAAUC 49 GAUUCAGU CUGAUGAG X CGAA AUGGCAGC 1824
950 CACUGAAU C AGACAAGU 50 ACUUGUCU CUGAUGAG X CGAA AUUCAGUG 1825
959 AGACAAGU U CUUCAUCA 51 UGAUGAAG CUGAUGAG X CGAA ACUUGUCU 1826
960 GACAAGUU C UUCAUCAA 52 UUGAUGAA CUGAUGAG X CGAA AACUUGUC 1827
962 CAAGUUCU U CAUCAACG 53 CGUUGAUG CUGAUGAG X CGAA AGAACUUG 1828
963 AAGUUCUU C AUCAACGG 54 CCGUUGAU CUGAUGAG X CGAA AAGAACUU 1829
966 UUCUUCAU C AACGGCUC 55 GAGCCGUU CUGAUGAG X CGAA AUGAAGAA 1830
974 CAACGGCU C CAACUGGG 56 CCCAGUUG CUGAUGAG X CGAA AGCCGUUG 1831
990 GAAGGCAU C CUGGGGCU 57 AGCCCCAG CUGAUGAG X CGAA AUGCCUUC 1832
1004 GCUGGCCU A UGCUGAGA 58 UCUCAGCA CUGAUGAG X CGAA AGGCCAGC 1833
1014 GCUGAGAU U GCCAGGCC 59 GGCCUGGC CUGAUGAG X CGAA AUCUCAGC 1834
1031 UGACGACU C CCUGGAGC 60 GCUCCAGG CUGAUGAG X CGAA AGUCGUCA 1835
1042 UGGAGCCU U UCUUUGAC 61 GUCAAAGA CUGAUGAG X CGAA AGGCUCCA 1836
1043 GGAGCCUU U CUUUGACU 62 AGUCAAAG CUGAUGAG X CGAA AAGGCUCC 1837
1044 GAGCCUUU C UUUGACUC 63 GAGUCAAA CUGAUGAG X CGAA AAAGGCUC 1838
1046 GCCUUUCU U UGACUCUC 64 GAGAGUCA CUGAUGAG X CGAA AGAAAGGC 1839
1047 CCUUUCUU U GACUCUCU 65 AGAGAGUC CUGAUGAG X CGAA AAGAAAGG 1840
1052 CUUUGACU C UCUGGUAA 66 UUACCAGA CUGAUGAG X CGAA AGUCAAAG 1841
1054 UUGACUCU C UGGUAAAG 67 CUUUACCA CUGAUGAG X CGAA AGAGUCAA 1842
1059 UCUCUGGU A AAGCAGAC 68 GUCUGCUU CUGAUGAG X CGAA ACCAGAGA 1843
1074 ACCCACGU U CCCAACCU 69 AGGUUGGG CUGAUGAG X CGAA ACGUGGGU 1844
1075 CCCACGUU C CCAACCUC 70 GAGGUUGG CUGAUGAG X CGAA AACGUGGG 1845
1083 CCCAACCU C UUCUCCCU 71 AGGGAGAA CUGAUGAG X CGAA AGGUUGGG 1846
1085 CAACCUCU U CUCCCUGC 72 GCAGGGAG CUGAUGAG X CGAA AGAGGUUG 1847
1086 AACCUCUU C UCCCUGCA 73 UGCAGGGA CUGAUGAG X CGAA AAGAGGUU 1848
1088 CCUCUUCU C CCUGCAGC 74 GCUGCAGG CUGAUGAG X CGAA AGAAGAGG 1849
1098 CUGCAGCU U UGUGGUGC 75 GCACCACA CUGAUGAG X CGAA AGCUGCAG 1850
1099 UGCAGCUU U GUGGUGCU 76 AGCACCAC CUGAUGAG X CGAA AAGCUGCA 1851
1112 UGCUGGCU U CCCCCUCA 77 UGAGGGGG CUGAUGAG X CGAA AGCCAGCA 1852
1113 GCUGGCUU C CCCCUCAA 78 UUGAGGGG CUGAUGAG X CGAA AAGCCAGC 1853
1119 UUCCCCCU C AACCAGUC 79 GACUGGUU CUGAUGAG X CGAA AGGGGGAA 1854
1127 CAACCAGU c UGAAGUGC 80 GCACUUCA CUGAUGAG X CGAA ACUGGUUG 1855
1142 GCUGGCCU c UGUCGGAG 81 CUCCGACA CUGAUGAG X CGAA AGGCCAGC 1856
1146 GCCUCUGU c GGAGGGAG 82 CUCCCUCC CUGAUGAG X CGAA ACAGAGGC 1857
1161 AGCAUGAU c AUUGGAGG 83 CCUCCAAU CUGAUGAG X CGAA AUCAUGCU 1858
1164 AUGAUCAU u GGAGGUAU 84 AUACCUCC CUGAUGAG X CGAA AUGAUCAU 1859
1171 UUGGAGGU A UCGACCAC 85 GUGGUCGA CUGAUGAG X CGAA ACCUCCAA 1860
1173 GGAGGUAU C GACCACUC 86 GAGUGGUC CUGAUGAG X CGAA AUACCUCC 1861
1181 CGACCACU C GCUGUACA 87 UGUACAGC CUGAUGAG X CGAA AGUGGUCG 1862
1187 CUCGCUGU A CACAGGCA 88 UGCCUGUG CUGAUGAG X CGAA ACAGCGAG 1863
1198 CAGGCAGU C UCUGGUAU 89 AUACCAGA CUGAUGAG X CGAA ACUGCCUG 1864
1200 GGCAGUCU C UGGUAUAC 90 GUAUACCA CUGAUGAG X CGAA AGACUGCC 1865
1205 UCUCUGGU A UACACCCA 91 UGGGUGUA CUGAUGAG X CGAA ACCAGAGA 1866
1207 UCUGGUAU A CACCCAUC 92 GAUGGGUG CUGAUGAG X CGAA AUACCAGA 1867
1215 ACACCCAU C CGGCGGGA 93 UCCCGCCG CUGAUGAG X CGAA AUGGGUGU 1868
1229 GGAGUGGU A UUAUGAGG 94 CCUCAUAA CUGAUGAG X CGAA ACCACUCC 1869
1231 AGUGGUAU U AUGAGGUG 95 CACCUCAU CUGAUGAG X CGAA AUACCACU 1870
1232 GUGGUAUU A UGAGGUGA 96 UCACCUCA CUGAUGAG X CGAA AAUACCAC 1871
1242 GAGGUGAU C AUUGUGCG 97 CGCACAAU CUGAUGAG X CGAA AUCACCUC 1872
1245 GUGAUCAU U GUGCGGGU 98 ACCCGCAC CUGAUGAG X CGAA AUGAUCAC 1873
Table 18
1260 GUGGAGAU C AAUGGACA 99 UGUCCAUU CUGAUGAG X CGAA AUCUCCAC 1874
1273 GACAGGAU C UGAAAAUG 100 CAUUUUCA CUGAUGAG X CGAA AUCCUGUC 1875
1295 CAAGGAGU A CAACUAUG 101 CAUAGUUG CUGAUGAG X CGAA ACUCCUUG 1876
1301 GUACAACU A UGACAAGA 102 UCUUGUCA CUGAUGAG X CGAA AGUUGUAC 1877
1314 AAGAGCAU U GUGGACAG 103 CUGUCCAC CUGAUGAG X CGAA AUGCUCUU 1878
1338 ACCAACCU U CGUUUGCC 104 GGCAAACG CUGAUGAG X CGAA AGGUUGGU 1879
1339 CCAACCUU C GUUUGCCC 105 GGGCAAAC CUGAUGAG X CGAA AAGGUUGG 1880
1342 ACCUUCGU u UGCCCAAG 106 CUUGGGCA CUGAUGAG X CGAA ACGAAGGU 1881
1343 CCUUCGUU u GCCCAAGA 107 UCUUGGGC CUGAUGAG X CGAA AACGAAGG 1882
1358 GAAAGUGU u UGAAGCUG 108 CAGCUUCA CUGAUGAG X CGAA ACACUUUC 1883
1359 AAAGUGUU u GAAGCUGC 109 GCAGCUUC CUGAUGAG X CGAA AACACUUU 1884
1371 GCUGCAGU c AAAUCCAU 110 AUGGAUUU CUGAUGAG X CGAA ACUGCAGC 1885
1376 AGUCAAAU c CAUCAAGG 111 CCUUGAUG CUGAUGAG X CGAA AUUUGACU 1886
1380 AAAUCCAU c AAGGCAGC 112 GCUGCCUU CUGAUGAG X CGAA AUGGAUUU 1887
1391 GGCAGCCU c CUCCACGG 113 CCGUGGAG CUGAUGAG X CGAA AGGCUGCC 1888
1394 AGCCUCCU c CACGGAGA 114 UCUCCGUG CUGAUGAG X CGAA AGGAGGCU 1889
1406 GGAGAAGU u CCCUGAUG 115 CAUCAGGG CUGAUGAG X CGAA ACUUCUCC 1890
1407 GAGAAGUU c CCUGAUGG 116 CCAUCAGG CUGAUGAG X CGAA AACUUCUC 1891
1417 CUGAUGGU u UCUGGCUA 117 UAGCCAGA CUGAUGAG X CGAA ACCAUCAG 1892
1418 UGAUGGUU u CUGGCUAG 118 CUAGCCAG CUGAUGAG X CGAA AACCAUCA 1893
1419 GAUGGUUU c UGGCUAGG 119 CCUAGCCA CUGAUGAG X CGAA AAACCAUC 1894
1425 UUCUGGCU A GGAGAGCA 120 UGCUCUCC CUGAUGAG X CGAA AGCCAGAA 1895
1465 CCACCCCU u GGAACAUU 121 AAUGUUCC CUGAUGAG X CGAA AGGGGUGG 1896
1473 UGGAACAU u UUCCCAGU 122 ACUGGGAA CUGAUGAG X CGAA AUGUUCCA 1897
1474 GGAACAUU u UCCCAGUC 123 GACUGGGA CUGAUGAG X CGAA AAUGUUCC 1898
1475 GAACAUUU u CCCAGUCA 124 UGACUGGG CUGAUGAG X CGAA AAAUGUUC 1899
1476 AACAUUUU c CCAGUCAU 125 AUGACUGG CUGAUGAG X CGAA AAAAUGUU 1900
1482 UUCCCAGU c AUCUCACU 126 AGUGAGAU CUGAUGAG X CGAA ACUGGGAA 1901
1485 CCAGUCAU c UCACUCUA 127 UAGAGUGA CUGAUGAG X CGAA AUGACUGG 1902
1487 AGUCAUCU c ACUCUACC 128 GGUAGAGU CUGAUGAG X CGAA AGAUGACU 1903
1491 AUCUCACU c UACCUAAU 129 AUUAGGUA CUGAUGAG X CGAA AGUGAGAU 1904
1493 CUCACUCU A CCUAAUGG 130 CCAUUAGG CUGAUGAG X CGAA AGAGUGAG 1905
1497 CUCUACCU A AUGGGUGA 131 UCACCCAU CUGAUGAG X CGAA AGGUAGAG 1906
1509 GGUGAGGU U ACCAACCA 132 UGGUUGGU CUGAUGAG X CGAA ACCUCACC 1907
1510 GUGAGGUU A CCAACCAG 133 CUGGUUGG CUGAUGAG X CGAA AACCUCAC 1908
1520 CAACCAGU C CUUCCGCA 134 UGCGGAAG CUGAUGAG X CGAA ACUGGUUG 1909
1523 CCAGUCCU U CCGCAUCA 135 UGAUGCGG CUGAUGAG X CGAA AGGACUGG 1910
1524 CAGUCCUU C CGCAUCAC 136 GUGAUGCG CUGAUGAG X CGAA AAGGACUG 1911
1530 UUCCGCAU C ACCAUCCU 137 AGGAUGGU CUGAUGAG X CGAA AUGCGGAA 1912
1536 AUCACCAU C CUUCCGCA 138 UGCGGAAG CUGAUGAG X CGAA AUGGUGAU 1913
1539 ACCAUCCU U CCGCAGCA 139 UGCUGCGG CUGAUGAG X CGAA AGGAUGGU 1914
1540 CCAUCCUU C CGCAGCAA 140 UUGCUGCG CUGAUGAG X CGAA AAGGAUGG 1915
1550 GCAGCAAU A CCUGCGGC 141 GCCGCAGG CUGAUGAG X CGAA AUUGCUGC 1916
1580 GGCCACGU C CCAAGACG 142 CGUCUUGG CUGAUGAG X CGAA ACGUGGCC 1917
1594 ACGACUGU U ACAAGUUU 143 AAACUUGU CUGAUGAG X CGAA ACAGUCGU 1918
1595 CGACUGUU A CAAGUUUG 144 CAAACUUG CUGAUGAG X CGAA AACAGUCG 1919
1601 UUACAAGU U UGCCAUCU 145 AGAUGGCA CUGAUGAG X CGAA ACUUGUAA 1920
1602 UACAAGUU U GCCAUCUC 146 GAGAUGGC CUGAUGAG X CGAA AACUUGUA 1921
1608 UUUGCCAU C UCACAGUC 147 GACUGUGA CUGAUGAG X CGAA AUGGCAAA 1922
1610 UGCCAUCU C ACAGUCAU 148 AUGACUGU CUGAUGAG X CGAA AGAUGGCA 1923
1616 CUCACAGU C AUCCACGG 149 CCGUGGAU CUGAUGAG X CGAA ACUGUGAG 1924
Table 18
1619 ACAGUCAU C CACGGGCA 150 UGCCCGUG CUGAUGAG X CGAA AUGACUGU 1925
1632 GGCACUGU U AUGGGAGC 151 GCUCCCAU CUGAUGAG X CGAA ACAGUGCC 1926
1633 GCACUGUU A UGGGAGCU 152 AGCUCCCA CUGAUGAG X CGAA AACAGUGC 1927
1644 GGAGCUGU U AUCAUGGA 153 UCCAUGAU CUGAUGAG X CGAA ACAGCUCC 1928
1645 GAGCUGUU A UCAUGGAG 154 CUCCAUGA CUGAUGAG X CGAA AACAGCUC 1929
1647 GCUGUUAU C AUGGAGGG 155 CCCUCCAU CUGAUGAG X CGAA AUAACAGC 1930
1658 GGAGGGCU U CUACGUUG 156 CAACGUAG CUGAUGAG X CGAA AGCCCUCC 1931
1659 GAGGGCUU C UACGUUGU 157 ACAACGUA CUGAUGAG X CGAA AAGCCCUC 1932
1661 GGGCUUCU A CGUUGUCU 158 AGACAACG CUGAUGAG X CGAA AGAAGCCC 1933
1665 UUCUACGU U GUCUUUGA 159 UCAAAGAC CUGAUGAG X CGAA ACGUAGAA 1934
1668 UACGUUGU C UUUGAUCG 160 CGAUCAAA CUGAUGAG X CGAA ACAACGUA 1935
1670 CGUUGUCU U UGAUCGGG 161 CCCGAUCA CUGAUGAG X CGAA AGACAACG 1936
1671 GUUGUCUU U GAUCGGGC 162 GCCCGAUC CUGAUGAG X CGAA AAGACAAC 1937
1675 UCUUUGAU C GGGCCCGA 163 UCGGGCCC CUGAUGAG X CGAA AUCAAAGA 1938
1692 AAACGAAU U GGCUUUGC 164 GCAAAGCC CUGAUGAG X CGAA AUUCGUUU 1939
1697 AAUUGGCU U UGCUGUCA 165 UGACAGCA CUGAUGAG X CGAA AGCCAAUU 1940
1698 AUUGGCUU U GCUGUCAG 166 CUGACAGC CUGAUGAG X CGAA AAGCCAAU 1941
1704 UUUGCUGU C AGCGCUUG 167 CAAGCGCU CUGAUGAG X CGAA ACAGCAAA 1942
1711 UCAGCGCU U GCCAUGUG 168 CACAUGGC CUGAUGAG X CGAA AGCGCUGA 1943
1730 CGAUGAGU U CAGGACGG 169 CCGUCCUG CUGAUGAG X CGAA ACUCAUCG 1944
1731 GAUGAGUU C AGGACGGC 170 GCCGUCCU CUGAUGAG X CGAA AACUCAUC 1945
1756 AAGGCCCU U UUGUCACC 171 GGUGACAA CUGAUGAG X CGAA AGGGCCUU 1946
1757 AGGCCCUU U UGUCACCU 172 AGGUGACA CUGAUGAG X CGAA AAGGGCCU 1947
1758 GGCCCUUU U GUCACCUU 173 AAGGUGAC CUGAUGAG X CGAA AAAGGGCC 1948
1761 CCUUUUGU C ACCUUGGA 174 UCCAAGGU CUGAUGAG X CGAA ACAAAAGG 1949
1766 UGUCACCU U GGACAUGG 175 CCAUGUCC CUGAUGAG X CGAA AGGUGACA 1950
1787 CUGUGGCU A CAACAUUC 176 GAAUGUUG CUGAUGAG X CGAA AGCCACAG 1951
1794 UACAACAU U CCACAGAC 177 GUCUGUGG CUGAUGAG X CGAA AUGUUGUA 1952
1795 ACAACAUU C CACAGACA 178 UGUCUGUG CUGAUGAG X CGAA AAUGUUGU 1953
1811 AGAUGAGU C AACCCUCA , 179 UGAGGGUU CUGAUGAG X CGAA ACUCAUCU 1954
1818 UCAACCCU C AUGACCAU 180 AUGGUCAU CUGAUGAG X CGAA AGGGUUGA 1955
1827 AUGACCAU A GCCUAUGU 181 ACAUAGGC CUGAUGAG X CGAA AUGGUCAU 1956
1832 CAUAGCCU A UGUCAUGG 182 CCAUGACA CUGAUGAG X CGAA AGGCUAUG 1957
1836 GCCUAUGU C AUGGCUGC 183 GCAGCCAU CUGAUGAG X CGAA ACAUAGGC 1958
1848 GCUGCCAU C UGCGCCCU 184 AGGGCGCA CUGAUGAG X CGAA AUGGCAGC 1959
1857 UGCGCCCU C UUCAUGCU 185 AGCAUGAA CUGAUGAG X CGAA AGGGCGCA 1960
1859 CGCCCUCU U CAUGCUGC 186 GCAGCAUG CUGAUGAG X CGAA AGAGGGCG 1961
1860 GCCCUCUU C AUGCUGCC 187 GGCAGCAU CUGAUGAG X CGAA AAGAGGGC 1962
1872 CUGCCACU C UGCCUCAU 188 AUGAGGCA CUGAUGAG X CGAA AGUGGCAG 1963
1878 CUCUGCCU C AUGGUGUG 189 CACACCAU CUGAUGAG X CGAA AGGCAGAG 1964
1888 UGGUGUGU C AGUGGCGC 190 GCGCCACU CUGAUGAG X CGAA ACACACCA 1965
1902 CGCUGCCU C CGCUGCCU 191 AGGCAGCG CUGAUGAG X CGAA AGGCAGCG 1966
1931 UGAUGACU U UGCUGAUG 192 CAUCAGCA CUGAUGAG X CGAA AGUCAUCA 1967
1932 GAUGACUU U GCUGAUGA 193 UCAUCAGC CUGAUGAG X CGAA AAGUCAUC 1968
1944 GAUGACAU C UCCCUGCU 194 AGCAGGGA CUGAUGAG X CGAA AUGUCAUC 1969
1946 UGACAUCU C CCUGCUGA 195 UCAGCAGG CUGAUGAG X CGAA AGAUGUCA 1970
1981 CAGAAGAU A GAGAUUCC 196 GGAAUCUC CUGAUGAG X CGAA AUCUUCUG 1971
1987 AUAGAGAU U CCCCUGGA 197 UCCAGGGG CUGAUGAG X CGAA AUCUCUAU 1972
1988 UAGAGAUU C CCCUGGAC 198 GUCCAGGG CUGAUGAG X CGAA AAUCUCUA 1973
2004 CCACACCU C CGUGGUUC 199 GAACCACG CUGAUGAG X CGAA AGGUGUGG 1974
2011 UCCGUGGU U CACUUUGG 200 CCAAAGUG CUGAUGAG X CGAA ACCACGGA 1975
Table 18
2012 CCGUGGUU c ACUUUGGU 201 ACCAAAGU CUGAUGAG X CGAA AACCACGG 1976
2016 GGUUCACU u UGGUCACA 202 UGUGACCA CUGAUGAG X CGAA AGUGAACC 1977
2017 GUUCACUU u GGUCACAA 203 UUGUGACC CUGAUGAG X CGAA AAGUGAAC 1978
2021 ACUUUGGU c ACAAGUAG 204 CUACUUGU CUGAUGAG X CGAA ACCAAAGU 1979
2028 UCACAAGU A GGAGACAC 205 GUGUCUCC CUGAUGAG X CGAA ACUUGUGA 1980
2063 GAGCACCU c AGGACCCU 206 AGGGUCCU CUGAUGAG X CGAA AGGUGCUC 1981
2072 AGGACCCU c CCCACCCA 207 UGGGUGGG CUGAUGAG X CGAA AGGGUCCU 1982
2091 AAAUGCCU c UGCCUUGA 208 UCAAGGCA CUGAUGAG X CGAA AGGCAUUU 1983
2097 CUCUGCCU u GAUGGAGA 209 UCUCCAUC CUGAUGAG X CGAA AGGCAGAG 1984
2129 AGGUGGGU u CCAGGGAC 210 GUCCCUGG CUGAUGAG X CGAA ACCCACCU 1985
2130 GGUGGGUU c CAGGGACU 211 AGUCCCUG CUGAUGAG X CGAA AACCCACC 1986
2141 GGGACUGU A CCUGUAGG 212 CCUACAGG CUGAUGAG X CGAA ACAGUCCC 1987
2147 GUACCUGU A GGAAACAG 213 CUGUUUCC CUGAUGAG X CGAA ACAGGUAC 1988
2177 GAAGCACU C UGCUGGCG 214 CGCCAGCA CUGAUGAG X CGAA AGUGCUUC 1989
2191 GCGGGAAU A CUCUUGGU 215 ACCAAGAG CUGAUGAG X CGAA AUUCCCGC 1990
2194 GGAAUACU C UUGGUCAC 216 GUGACCAA CUGAUGAG X CGAA AGUAUUCC 1991
2196 AAUACUCU u GGUCACCU 217 AGGUGACC CUGAUGAG X CGAA AGAGUAUU 1992
2200 CUCUUGGU C ACCUCAAA 218 UUUGAGGU CUGAUGAG X CGAA ACCAAGAG 1993
2205 GGUCACCU c AAAUUUAA 219 UUAAAUUU CUGAUGAG X CGAA AGGUGACC 1994
2210 CCUCAAAU u UAAGUCGG 220 CCGACUUA CUGAUGAG X CGAA AUUUGAGG 1995
2211 CUCAAAUU u AAGUCGGG 221 CCCGACUU CUGAUGAG X CGAA AAUUUGAG 1996
2212 UCAAAUUU A AGUCGGGA 222 UCCCGACU CUGAUGAG X CGAA AAAUUUGA 1997
2216 AUUUAAGU C GGGAAAUU 223 AAUUUCCC CUGAUGAG X CGAA ACUUAAAU 1998
2224 CGGGAAAU U CUGCUGCU 224 AGCAGCAG CUGAUGAG X CGAA AUUUCCCG 1999
2225 GGGAAAUU C UGCUGCUU 225 AAGCAGCA CUGAUGAG X CGAA AAUUUCCC 2000
2233 CUGCUGCU U GAAACUUC 226 GAAGUUUC CUGAUGAG X CGAA AGCAGCAG 2001
2240 UUGAAACU u CAGCCCUG 227 CAGGGCUG CUGAUGAG X CGAA AGUUUCAA 2002
2241 UGAAACUU c AGCCCUGA 228 UCAGGGCU CUGAUGAG X CGAA AAGUUUCA 2003
2254 CUGAACCU u UGUCCACC 229 GGUGGACA CUGAUGAG X CGAA AGGUUCAG 2004
2255 UGAACCUU u GUCCACCA 230 UGGUGGAC CUGAUGAG X CGAA AAGGUUCA 2005
2258 ACCUUUGU c CACCAUUC 231 GAAUGGUG CUGAUGAG X CGAA ACAAAGGU 2006
2265 UCCACCAU u CCUUUAAA 232 UUUAAAGG CUGAUGAG X CGAA AUGGUGGA 2007
2266 CCACCAUU c CUUUAAAU 233 AUUUAAAG CUGAUGAG X CGAA AAUGGUGG 2008
2269 CCAUUCCU u UAAAUUCU 234 AGAAUUUA CUGAUGAG X CGAA AGGAAUGG 2009
2270 CAUUCCUU u AAAUUCUC 235 GAGAAUUU CUGAUGAG X CGAA AAGGAAUG 2010
2271 AUUCCUUU A AAUUCUCC 236 GGAGAAUU CUGAUGAG X CGAA AAAGGAAU 2011
2275 CUUUAAAU U CUCCAACC 237 GGUUGGAG CUGAUGAG X CGAA AUUUAAAG 2012
2276 UUUAAAUU C UCCAACCC 238 GGGUUGGA CUGAUGAG X CGAA AAUUUAAA 2013
2278 UAAAUUCU c CAACCCAA 239 UUGGGUUG CUGAUGAG X CGAA AGAAUUUA 2014
2290 CCCAAAGU A UUCUUCUU 240 AAGAAGAA CUGAUGAG X CGAA ACUUUGGG 2015
2292 CAAAGUAU U CUUCUUUU 241 AAAAGAAG CUGAUGAG X CGAA AUACUUUG 2016
2293 AAAGUAUU C UUCUUUUC 242 GAAAAGAA CUGAUGAG X CGAA AAUACUUU 2017
2295 AGUAUUCU U CUUUUCUU 243 AAGAAAAG CUGAUGAG X CGAA AGAAUACU 2018
2296 GUAUUCUU C UUUUCUUA 244 UAAGAAAA CUGAUGAG X CGAA AAGAAUAC 2019
2298 AUUCUUCU U UUCUUAGU 245 ACUAAGAA CUGAUGAG X CGAA AGAAGAAU 2020
2299 UUCUUCUU U UCUUAGUU 246 AACUAAGA CUGAUGAG X CGAA AAGAAGAA 2021
2300 UCUUCUUU U CUUAGUUU 247 AAACUAAG CUGAUGAG X CGAA AAAGAAGA 2022
2301 cuucuuuu c UUAGUUUC 248 GAAACUAA CUGAUGAG X CGAA AAAAGAAG 2023
2303 ucuuuucu u AGUUUCAG 249 CUGAAACU CUGAUGAG X CGAA AGAAAAGA 2024
2304 cuuuucuu A GUUUCAGA 250 UCUGAAAC CUGAUGAG X CGAA AAGAAAAG 2025
2307 UUCUUAGU U UCAGAAGU 251 ACUUCUGA CUGAUGAG X CGAA ACUAAGAA 2026
Table 18
2308 UCUUAGUU U CAGAAGUA 252 UACUUCUG CUGAUGAG X CGAA AACUAAGA 2027
2309 CUUAGUUU C AGAAGUAC 253 GUACUUCU CUGAUGAG X CGAA AAACUAAG 2028
2316 UCAGAAGU A CUGGCAUC 254 GAUGCCAG CUGAUGAG X CGAA ACUUCUGA 2029
2324 ACUGGCAU C ACACGCAG 255 CUGCGUGU CUGAUGAG X CGAA AUGCCAGU 2030
2335 ACGCAGGU U ACCUUGGC 256 GCCAAGGU CUGAUGAG X CGAA ACCUGCGU 2031
2336 CGCAGGUU A CCUUGGCG 257 CGCCAAGG CUGAUGAG X CGAA AACCUGCG 2032
2340 GGUUACCU U GGCGUGUG 258 CACACGCC CUGAUGAG X CGAA AGGUAACC 2033
2350 GCGUGUGU C CCUGUGGU 259 ACCACAGG CUGAUGAG X CGAA ACACACGC 2034
2359 CCUGUGGU A CCCUGGCA 260 UGCCAGGG CUGAUGAG X CGAA ACCACAGG 2035
2384 ACCAAGCU U GUUUCCCU 261 AGGGAAAC CUGAUGAG X CGAA AGCUUGGU 2036
2387 AAGCUUGU U UCCCUGCU 262 AGCAGGGA CUGAUGAG X CGAA ACAAGCUU 2037
2388 AGCUUGUU U CCCUGCUG 263 CAGCAGGG CUGAUGAG X CGAA AACAAGCU 2038
2389 GCUUGUUU C CCUGCUGG 264 CCAGCAGG CUGAUGAG X CGAA AAACAAGC 2039
2405 GCCAAAGU C AGUAGGAG 265 CUCCUACU CUGAUGAG X CGAA ACUUUGGC 2040
2409 AAGUCAGU A GGAGAGGA 266 UCCUCUCC CUGAUGAG X CGAA ACUGACUU 2041
2426 UGCACAGU U UGCUAUUU 267 AAAUAGCA CUGAUGAG X CGAA ACUGUGCA 2042
2427 GCACAGUU u GCUAUUUG 268 CAAAUAGC CUGAUGAG X CGAA AACUGUGC 2043
2431 AGUUUGCU A UUUGCUUU 269 AAAGCAAA CUGAUGAG X CGAA AGCAAACU 2044
2433 UUUGCUAU U UGCUUUAG 270 CUAAAGCA CUGAUGAG X CGAA AUAGCAAA 2045
2434 UUGCUAUU U GCUUUAGA 271 UCUAAAGC CUGAUGAG X CGAA AAUAGCAA 2046
2438 UAUUUGCU U UAGAGACA 272 UGUCUCUA CUGAUGAG X CGAA AGCAAAUA 2047
2439 AUUUGCUU U AGAGACAG 273 CUGUCUCU CUGAUGAG X CGAA AAGCAAAU 2048
2440 UUUGCUUU A GAGACAGG 274 ccuσucuc CUGAUGAG X CGAA AAAGCAAA 2049
2455 GGGACUGU A UAAACAAG 275 CUUGUUUA CUGAUGAG X CGAA ACAGUCCC 2050
2457 GACUGUAU A AACAAGCC 276 GGCUUGUU CUGAUGAG X CGAA AUACAGUC 2051
2467 ACAAGCCU A ACAUUGGU , 277 ACCAAUGU CUGAUGAG X CGAA AGGCUUGU 2052
2472 CCUAACAU U GGUGCAAA 278 UUUGCACC CUGAUGAG X CGAA AUGUUAGG 2053
2484 GCAAAGAU U GCCUCUUG 279 CAAGAGGC CUGAUGAG X CGAA AUCUUUGC 2054
2489 GAUUGCCU C UUGAAUUA 280 UAAUUCAA CUGAUGAG X CGAA AGGCAAUC 2055
2491 UUGCCUCU U GAAUUAAA 281 UUUAAUUC CUGAUGAG X CGAA AGAGGCAA 2056
2496 UCUUGAAU U AAAAAAAA 282 UUUUUUUU CUGAUGAG X CGAA AUUCAAGA 2057
2497 CUUGAAUU A AAAAAAAA 283 UUUUUUUU CUGAUGAG X CGAA AAUUCAAG 2058
2510 AAAAAACU A GAAAAAAA 284 uuuuuuuc CUGAUGAG X CGAA AGUUUUUU 2059
Input Sequence = AF190725. Cut Site = G/ .
Stem Length = 8 . Core Sequence = CUGAUGAG X CGAA (X = GCCGUUAGGC or other stem II)
AF190725 (Homo sapiens beta- site APP cleaving enzyme (BACE) mRNA; 2526 bp)
Table 19 Table 19: Human BACE NCH Ribozyme and Target Sequence
Pos Substrate Seq ID Ribozyme Rz Seq ID
10 CACGCGUC C GCAGCCCG 285 CGGGCUGC CUGAUGAG X CGAA IACGCGUG 2060
13 GCGUCCGC A GCCCGCCC 286 J GGGCGGGC CUGAUGAG X CGAA ICGGACGC 2061
16 UCCGCAGC C CGCCCGGG 287 CCCGGGCG CUGAUGAG X CGAA ICUGCGGA 2062
17 CCGCAGCC C GCCCGGGA 288 UCCCGGGC CUGAUGAG X CGAA IGCUGCGG 2063
20 CAGCCCGC C CGGGAGCU 289 AGCUCCCG CUGAUGAG X CGAA ICGGGCUG 2064
21 AGCCCGCC C GGGAGCUG 290 CAGCUCCC CUGAUGAG X CGAA IGCGGGCU 2065
28 CCGGGAGC U GCGAGCCG 291 CGGCUCGC CUGAUGAG X CGAA ICUCCCGG 2066
35 CUGCGAGC C GCGAGCUG 292 CAGCUCGC CUGAUGAG X CGAA ICUCGCAG 2067
42 CCGCGAGC U GGAUUAUG 293 CAUAAUCC CUGAUGAG X CGAA ICUCGCGG 2068
56 AUGGUGGC C UGAGCAGC 294 GCUGCUCA CUGAUGAG X CGAA ICCACCAU 2069
57 UGGUGGCC U GAGCAGCC 295 GGCUGCUC CUGAUGAG X CGAA IGCCACCA 2070
62 GCCUGAGC A GCCAACGC 296 GCGUUGGC CUGAUGAG X CGAA ICUCAGGC 2071
65 UGAGCAGC C AACGCAGC 297 GCUGCGUU CUGAUGAG X CGAA ICUGCUCA 2072
66 GAGCAGCC A ACGCAGCC 298 GGCUGCGU CUGAUGAG X CGAA IGCUGCUC 2073
71 GCCAACGC A GCCGCAGG 299 CCUGCGGC CUGAUGAG X CGAA ICGUUGGC 2074
74 AACGCAGC C GCAGGAGC 300 GCUCCUGC CUGAUGAG X CGAA ICUGCGUU 2075
77 GCAGCCGC A GGAGCCCG 301 CGGGCUCC CUGAUGAG X CGAA ICGGCUGC 2076
83 GCAGGAGC C CGGAGCCC 302 GGGCUCCG CUGAUGAG X CGAA ICUCCUGC 2077
84 CAGGAGCC C GGAGCCCU 303 AGGGCUCC CUGAUGAG X CGAA IGCUCCUG 2078
90 CCCGGAGC C CUUGCCCC 304 GGGGCAAG CUGAUGAG X CGAA ICUCCGGG 2079
91 CCGGAGCC C UUGCCCCU 305 AGGGGCAA CUGAUGAG X CGAA IGCUCCGG 2080
92 CGGAGCCC U UGCCCCUG 306 CAGGGGCA CUGAUGAG X CGAA IGGCUCCG 2081
96 GCCCUUGC C CCUGCCCG 307 CGGGCAGG CUGAUGAG X CGAA ICAAGGGC 2082
97 CCCUUGCC C CUGCCCGC 308 GCGGGCAG CUGAUGAG X CGAA IGCAAGGG 2083
98 CCUUGCCC C UGCCCGCG 309 CGCGGGCA CUGAUGAG X CGAA IGGCAAGG 2084
99 CUUGCCCC U GCCCGCGC 310 GCGCGGGC CUGAUGAG X CGAA IGGGCAAG 2085
102 GCCCCUGC C CGCGCCGC 311 GCGGCGCG CUGAUGAG X CGAA ICAGGGGC 2086
103 CCCCUGCC C GCGCCGCC 312 GGCGGCGC CUGAUGAG X CGAA IGCAGGGG 2087
108 GCCCGCGC C GCCGCCCG 313 CGGGCGGC CUGAUGAG X CGAA ICGCGGGC 2088
111 CGCGCCGC C GCCCGCCG 314 CGGCGGGC CUGAUGAG X CGAA ICGGCGCG 2089
114 GCCGCCGC C CGCCGGGG 315 CCCCGGCG CUGAUGAG X CGAA ICGGCGGC 2090
115 CCGCCGCC C GCCGGGGG 316 CCCCCGGC CUGAUGAG X CGAA IGCGGCGG 2091
118 CCGCCCGC C GGGGGGAC 317 GUCCCCCC CUGAUGAG X CGAA ICGGGCGG 2092
127 GGGGGGAC C AGGGAAGC 318 GCUUCCCU CUGAUGAG X CGAA IUCCCCCC 2093
128 GGGGGACC A GGGAAGCC 319 GGCUUCCC CUGAUGAG X CGAA IGUCCCCC 2094
136 AGGGAAGC C GCCACCGG 320 CCGGUGGC CUGAUGAG X CGAA ICUUCCCU 2095
139 GAAGCCGC C ACCGGCCC 321 GGGCCGGU CUGAUGAG X CGAA ICGGCUUC 2096
140 AAGCCGCC A CCGGCCCG 322 CGGGCCGG CUGAUGAG X CGAA IGCGGCUU 2097
142 GCCGCCAC C GGCCCGCC 323 GGCGGGCC CUGAUGAG X CGAA IUGGCGGC 2098
146 CCACCGGC C CGCCAUGC 324 GCAUGGCG CUGAUGAG X CGAA ICCGGUGG 2099
147 CACCGGCC C GCCAUGCC 325 GGCAUGGC CUGAUGAG X CGAA IGCCGGUG 2100
150 CGGCCCGC C AUGCCCGC 326 GCGGGCAU CUGAUGAG X CGAA ICGGGCCG 2101
151 GGCCCGCC A UGCCCGCC 327 GGCGGGCA CUGAUGAG X CGAA IGCGGGCC 2102
155 CGCCAUGC C CGCCCCUC 328 GAGGGGCG CUGAUGAG X CGAA ICAUGGCG 2103
156 GCCAUGCC C GCCCCUCC 329 GGAGGGGC CUGAUGAG X CGAA IGCAUGGC 2104
159 AUGCCCGC C CCUCCCAG 330 CUGGGAGG CUGAUGAG X CGAA ICGGGCAU 2105
160 UGCCCGCC C CUCCCAGC 331 GCUGGGAG CUGAUGAG X CGAA IGCGGGCA 2106
Table 19
161 GCCCGCCC C UCCCAGCC 332 GGCUGGGA CUGAUGAG X CGAA IGGCGGGC 2107
162 CCCGCCCC U CCCAGCCC 333 GGGCUGGG CUGAUGAG X CGAA IGGGCGGG 2108
164 CGCCCCUC C CAGCCCCG 334 CGGGGCUG CUGAUGAG X CGAA IAGGGGCG 2109
165 GCCCCUCC C AGCCCCGC 335 GCGGGGCU CUGAUGAG X CGAA IGAGGGGC 2110
166 CCCCUCCC A GCCCCGCC 336 GGCGGGGC CUGAUGAG X CGAA IGGAGGGG 2111
169 CUCCCAGC C CCGCCGGG 337 CCCGGCGG CUGAUGAG X CGAA ICUGGGAG 2112
170 UCCCAGCC C CGCCGGGA 338 UCCCGGCG CUGAUGAG X CGAA IGCUGGGA 2113
171 CCCAGCCC C GCCGGGAG 339 CUCCCGGC CUGAUGAG X CGAA IGGCUGGG 2114
174 AGCCCCGC C GGGAGCCC 340 GGGCUCCC CUGAUGAG X CGAA ICGGGGCU 2115
181 CCGGGAGC C CGCGCCCG 341 CGGGCGCG CUGAUGAG X CGAA ICUCCCGG 2116
182 CGGGAGCC C GCGCCCGC 342 GCGGGCGC CUGAUGAG X CGAA IGCUCCCG 2117
187 GCCCGCGC C CGCUGCCC 343 GGGCAGCG CUGAUGAG X CGAA ICGCGGGC 2118
188 CCCGCGCC C GCUGCCCA 344 UGGGCAGC CUGAUGAG X CGAA IGCGCGGG 2119
191 GCGCCCGC U GCCCAGGC 345 GCCUGGGC CUGAUGAG X CGAA ICGGGCGC 2120
194 CCCGCUGC C CAGGCUGG 346 CCAGCCUG CUGAUGAG X CGAA ICAGCGGG 2121
195 CCGCUGCC C AGGCUGGC 347 GCCAGCCU CUGAUGAG X CGAA IGCAGCGG 2122
196 CGCUGCCC A GGCUGGCC 348 GGCCAGCC CUGAUGAG X CGAA IGGCAGCG 2123
200 GCCCAGGC U GGCCGCCG 349 CGGCGGCC CUGAUGAG X CGAA ICCUGGGC 2124
204 AGGCUGGC C GCCGCCGU 350 ACGGCGGC CUGAUGAG X CGAA ICCAGCCU 2125
207 CUGGCCGC C GCCGUGCC 351 GGCACGGC CUGAUGAG X CGAA ICGGCCAG 2126
210 GCCGCCGC C GUGCCGAU 352 AUCGGCAC CUGAUGAG X CGAA ICGGCGGC 2127
215 CGCCGUGC C GAUGUAGC 353 GCUACAUC CUGAUGAG X CGAA ICACGGCG 2128
228 UAGCGGGC U CCGGAUCC 354 GGAUCCGG CUGAUGAG X CGAA ICCCGCUA 2129
230 GCGGGCUC C GGAUCCCA 355 UGGGAUCC CUGAUGAG X CGAA IAGCCCGC 2130
236 UCCGGAUC C CAGCCUCU 356 AGAGGCUG CUGAUGAG X CGAA IAUCCGGA 2131
237 CCGGAUCC C AGCCUCUC 357 GAGAGGCU CUGAUGAG X CGAA IGAUCCGG 2132
238 CGGAUCCC A GCCUCUCC 358 GGAGAGGC CUGAUGAG X CGAA IGGAUCCG 2133
241 AUCCCAGC C UCUCCCCU 359 AGGGGAGA CUGAUGAG X CGAA ICUGGGAU 2134
242 UCCCAGCC U CUCCCCUG 360 CAGGGGAG CUGAUGAG X CGAA IGCUGGGA 2135
244 CCAGCCUC U CCCCUGCU 361 AGCAGGGG CUGAUGAG X CGAA lAGGCUGG 2136
246 AGCCUCUC C CCUGCUCC 362 GGAGCAGG CUGAUGAG X CGAA IAGAGGCU 2137
247 GCCUCUCC C CUGCUCCC 363 GGGAGCAG CUGAUGAG X CGAA IGAGAGGC 2138
248 CCUCUCCC C UGCUCCCG 364 CGGGAGCA CUGAUGAG X CGAA IGGAGAGG 2139
249 CUCUCCCC U GCUCCCGU 365 ACGGGAGC CUGAUGAG X CGAA IGGGAGAG 2140
252 UCCCCUGC u CCCGUGCU 366 AGCACGGG CUGAUGAG X CGAA ICAGGGGA 2141
254 CCCUGCUC c CGUGCUCU 367 AGAGCACG CUGAUGAG X CGAA IAGCAGGG 2142
255 CCUGCUCC c GUGCUCUG 368 CAGAGCAC CUGAUGAG X CGAA IGAGCAGG 2143
260 UCCCGUGC u CUGCGGAU 369 AUCCGCAG CUGAUGAG X' CGAA ICACGGGA 2144
262 CCGUGCUC u GCGGAUCU 370 AGAUCCGC CUGAUGAG X CGAA IAGCACGG 2145
270 UGCGGAUC u CCCCUGAC 371 GUCAGGGG CUGAUGAG X CGAA IAUCCGCA 2146
272 CGGAUCUC c CCUGACCG 372 CGGUCAGG CUGAUGAG X CGAA IAGAUCCG 2147
273 GGAUCUCC c CUGACCGC 373 GCGGUCAG CUGAUGAG X CGAA IGAGAUCC 2148
274 GAUCUCCC c UGACCGCU 374 AGCGGUCA CUGAUGAG X CGAA IGGAGAUC 2149
275 AUCUCCCC u GACCGCUC 375 GAGCGGUC CUGAUGAG X CGAA IGGGAGAU 2150
279 CCCCUGAC c GCUCUCCA 376 UGGAGAGC CUGAUGAG X CGAA IUCAGGGG 2151
282 CUGACCGC u CUCCACAG 377 CUGUGGAG CUGAUGAG X CGAA ICGGUCAG 2152
284 GACCGCUC u CCACAGCC 378 GGCUGUGG CUGAUGAG X CGAA IAGCGGUC 2153
286 CCGCUCUC c ACAGCCCG 379 CGGGCUGU CUGAUGAG X CGAA IAGAGCGG 2154
287 CGCUCUCC A CAGCCCGG 380 CCGGGCUG CUGAUGAG X CGAA IGAGAGCG 2155
289 CUCUCCAC A GCCCGGAC 381 GUCCGGGC CUGAUGAG X CGAA IUGGAGAG 2156
292 UCCACAGC C CGGACCCG 382 CGGGUCCG CUGAUGAG X CGAA ICUGUGGA 2157
Table 19
293 CCACAGCC C GGACCCGG 383 CCGGGUCC CUGAUGAG X CGAA IGCUGUGG 2158
298 GCCCGGAC C CGGGGGCU 384 AGCCCCCG CUGAUGAG X CGAA IUCCGGGC 2159
299 CCCGGACC C GGGGGCUG 385 CAGCCCCC CUGAUGAG X CGAA IGUCCGGG 2160
306 CCGGGGGC U GGCCCAGG 386 CCUGGGCC CUGAUGAG X CGAA ICCCCCGG 2161
310 GGGCUGGC C CAGGGCCC 387 GGGCCCUG CUGAUGAG X CGAA ICCAGCCC 2162
311 GGCUGGCC C AGGGCCCU 388 AGGGCCCU CUGAUGAG X CGAA IGCCAGCC 2163
312 GCUGGCCC A GGGCCCUG 389 CAGGGCCC CUGAUGAG X CGAA IGGCCAGC 2164
317 CCCAGGGC C CUGCAGGC 390 GCCUGCAG CUGAUGAG X CGAA ICCCUGGG 2165
318 CCAGGGCC C UGCAGGCC 391 GGCCUGCA CUGAUGAG X CGAA IGCCCUGG 2166
319 CAGGGCCC U GCAGGCCC 392 GGGCCUGC CUGAUGAG X CGAA IGGCCCUG 2167
322 GGCCCUGC A GGCCCUGG 393 CCAGGGCC CUGAUGAG X CGAA ICAGGGCC 2168
326 CUGCAGGC C CUGGCGUC 394 GACGCCAG CUGAUGAG X CGAA ICCUGCAG 2169
327 UGCAGGCC C UGGCGUCC 395 GGACGCCA CUGAUGAG X CGAA IGCCUGCA 2170
328 GCAGGCCC U GGCGUCCU 396 AGGACGCC CUGAUGAG X CGAA IGGCCUGC 2171
335 CUGGCGUC C UGAUGCCC 397 GGGCAUCA CUGAUGAG X CGAA IACGCCAG 2172
336 UGGCGUCC u GAUGCCCC 398 GGGGCAUC CUGAUGAG X CGAA IGACGCCA 2173
342 CCUGAUGC c CCCAAGCU 399 AGCUUGGG CUGAUGAG X CGAA ICAUCAGG 2174
343 CUGAUGCC c CCAAGCUC 400 GAGCUUGG CUGAUGAG X CGAA IGCAUCAG 2175
344 UGAUGCCC c CAAGCUCC 401 GGAGCUUG CUGAUGAG X CGAA IGGCAUCA 2176
345 GAUGCCCC c AAGCUCCC 402 GGGAGCUU CUGAUGAG X CGAA IGGGCAUC 2177
346 AUGCCCCC A AGCUCCCU 403 AGGGAGCU CUGAUGAG X CGAA IGGGGCAU 2178
350 CCCCAAGC U CCCUCUCC 404 GGAGAGGG CUGAUGAG X CGAA ICUUGGGG 2179
352 CCAAGCUC C CUCUCCUG 405 CAGGAGAG CUGAUGAG X CGAA IAGCUUGG 2180
353 CAAGCUCC C UCUCCUGA 406 UCAGGAGA CUGAUGAG X CGAA IGAGCUUG 2181
354 AAGCUCCC u CUCCUGAG 407 CUCAGGAG CUGAUGAG X CGAA IGGAGCUU 2182
356 GCUCCCUC u CCUGAGAA 408 UUCUCAGG CUGAUGAG X CGAA IAGGGAGC 2183
358 UCCCUCUC c UGAGAAGC 409 GCUUCUCA CUGAUGAG X CGAA IAGAGGGA 2184
359 CCCUCUCC u GAGAAGCC 410 GGCUUCUC CUGAUGAG X CGAA IGAGAGGG 2185
367 UGAGAAGC c ACCAGCAC 411 GUGCUGGU CUGAUGAG X CGAA ICUUCUCA 2186
368 GAGAAGCC A CCAGCACC 412 GGUGCUGG CUGAUGAG X CGAA IGCUUCUC 2187
370 GAAGCCAC C AGCACCAC 413 GUGGUGCU CUGAUGAG X CGAA IUGGCUUC 2188
371 AAGCCACC A GCACCACC 414 GGUGGUGC CUGAUGAG X CGAA IGUGGCUU 2189
374 CCACCAGC A CCACCCAG 415 CUGGGUGG CUGAUGAG X CGAA ICUGGUGG 2190
376 ACCAGCAC C ACCCAGAC 416 GUCUGGGU CUGAUGAG X CGAA IUGCUGGU 2191
377 CCAGCACC A CCCAGACU 417 AGUCUGGG CUGAUGAG X CGAA IGUGCUGG 2192
379 AGCACCAC C CAGACUUG 418 CAAGUCUG CUGAUGAG X CGAA IUGGUGCU 2193
380 GCACCACC C AGACUUGG 419 CCAAGUCU CUGAUGAG X CGAA IGUGGUGC 2194
381 CACCACCC A GACUUGGG 420 CCCAAGUC CUGAUGAG X CGAA IGGUGGUG 2195
385 ACCCAGAC u UGGGGGCA 421 UGCCCCCA CUGAUGAG X CGAA lUCUGGGU 2196
393 UUGGGGGC A GGCGCCAG 422 CUGGCGCC CUGAUGAG X CGAA ICCCCCAA 2197
399 GCAGGCGC C AGGGACGG 423 CCGUCCCU CUGAUGAG X CGAA ICGCCUGC 2198
400 CAGGCGCC A GGGACGGA 424 UCCGUCCC CUGAUGAG X CGAA IGCGCCUG 2199
416 ACGUGGGC C AGUGCGAG 425 CUCGCACU CUGAUGAG X CGAA ICCCACGU 2200
417 CGUGGGCC A GUGCGAGC 426 GCUCGCAC CUGAUGAG X CGAA IGCCCACG 2201
426 GUGCGAGC C CAGAGGGC 427 GCCCUCUG CUGAUGAG X CGAA ICUCGCAC 2202
427 UGCGAGCC C AGAGGGCC 428 GGCCCUCU CUGAUGAG X CGAA IGCUCGCA 2203
428 GCGAGCCC A GAGGGCCC 429 GGGCCCUC CUGAUGAG X CGAA IGGCUCGC 2204
435 CAGAGGGC C CGAAGGCC 430 GGCCUUCG CUGAUGAG X CGAA ICCCUCUG 2205
436 AGAGGGCC C GAAGGCCG 431 CGGCCUUC CUGAUGAG X CGAA IGCCCUCU 2206
443 CCGAAGGC C GGGGCCCA 432 UGGGCCCC CUGAUGAG X CGAA ICCUUCGG 2207
449 GCCGGGGC C CACCAUGG 433 CCAUGGUG CUGAUGAG X CGAA ICCCCGGC 2208
Table 19
450 CCGGGGCC C ACCAUGGC 434 GCCAUGGU CUGAUGAG X CGAA IGCCCCGG 2209
451 CGGGGCCC A CCAUGGCC 435 GGCCAUGG CUGAUGAG X CGAA IGGCCCCG 2210
453 GGGCCCAC C AUGGCCCA 436 UGGGCCAU CUGAUGAG X CGAA IUGGGCCC 2211
454 GGCCCACC A UGGCCCAA 437 UUGGGCCA CUGAUGAG X CGAA IGUGGGCC 2212
459 ACCAUGGC C CAAGCCCU 438 AGGGCUUG CUGAUGAG X CGAA ICCAUGGU 2213
460 CCAUGGCC C AAGCCCUG 439 CAGGGCUU CUGAUGAG X CGAA IGCCAUGG 2214
461 CAUGGCCC A AGCCCUGC 440 GCAGGGCU CUGAUGAG X CGAA IGGCCAUG 2215
465 GCCCAAGC C CUGCCCUG 441 CAGGGCAG CUGAUGAG X CGAA ICUUGGGC 2216
466 CCCAAGCC C UGCCCUGG 442 CCAGGGCA CUGAUGAG X CGAA IGCUUGGG 2217
467 CCAAGCCC U GCCCUGGC 443 GCCAGGGC CUGAUGAG X CGAA IGGCUUGG 2218
470 AGCCCUGC c CUGGCUCC 444 GGAGCCAG CUGAUGAG X CGAA ICAGGGCU 2219
471 GCCCUGCC c UGGCUCCU 445 AGGAGCCA CUGAUGAG X CGAA IGCAGGGC 2220
472 CCCUGCCC u GGCUCCUG 446 CAGGAGCC CUGAUGAG X CGAA IGGCAGGG 2221
476 GCCCUGGC u CCUGCUGU 447 ACAGCAGG CUGAUGAG X CGAA ICCAGGGC 2222
478 CCUGGCUC c UGCUGUGG 448 CCACAGCA CUGAUGAG X CGAA IAGCCAGG 2223
479 CUGGCUCC u GCUGUGGA 449 UCCACAGC CUGAUGAG X CGAA IGAGCCAG 2224
482 GCUCCUGC u GUGGAUGG 450 CCAUCCAC CUGAUGAG X CGAA ICAGGAGC 2225
503 GGGAGUGC u GCCUGCCC 451 GGGCAGGC CUGAUGAG X CGAA ICACUCCC 2226
506 AGUGCUGC c UGCCCACG 452 CGUGGGCA CUGAUGAG X CGAA ICAGCACU 2227
507 GUGCUGCC u GCCCACGG 453 CCGUGGGC CUGAUGAG X CGAA IGCAGCAC 2228
510 CUGCCUGC c CACGGCAC 454 GUGCCGUG CUGAUGAG X CGAA ICAGGCAG 2229
511 UGCCUGCC c ACGGCACC 455 GGUGCCGU CUGAUGAG X CGAA IGCAGGCA 2230
512 GCCUGCCC A CGGCACCC 456 GGGUGCCG CUGAUGAG X CGAA IGGCAGGC 2231
517 CCCACGGC A CCCAGCAC 457 GUGCUGGG CUGAUGAG X CGAA ICCGUGGG 2232
519 CACGGCAC C CAGCACGG 458 CCGUGCUG CUGAUGAG X CGAA IUGCCGUG 2233
520 ACGGCACC C AGCACGGC 459 GCCGUGCU CUGAUGAG X CGAA IGUGCCGU 2234
521 CGGCACCC A GCACGGCA 460 UGCCGUGC CUGAUGAG X CGAA IGGUGCCG 2235
524 CACCCAGC A CGGCAUCC 461 GGAUGCCG CUGAUGAG X CGAA ICUGGGUG 2236
529 AGCACGGC A UCCGGCUG 462 CAGCCGGA CUGAUGAG X CGAA ICCGUGCU 2237
532 ACGGCAUC C GGCUGCCC 463 GGGCAGCC CUGAUGAG X CGAA IAUGCCGU 2238
536 CAUCCGGC U GCCCCUGC 464 GCAGGGGC CUGAUGAG X CGAA ICCGGAUG 2239
539 CCGGCUGC C CCUGCGCA 465 UGCGCAGG CUGAUGAG X CGAA ICAGCCGG 2240
540 CGGCUGCC C CUGCGCAG 466 CUGCGCAG CUGAUGAG X CGAA IGCAGCCG 2241
541 GGCUGCCC C UGCGCAGC 467 GCUGCGCA CUGAUGAG X CGAA IGGCAGCC 2242
542 GCUGCCCC U GCGCAGCG 468 CGCUGCGC CUGAUGAG X CGAA IGGGCAGC 2243
547 CCCUGCGC A GCGGCCUG 469 CAGGCCGC CUGAUGAG X CGAA ICGCAGGG 2244
553 GCAGCGGC C UGGGGGGC 470 GCCCCCCA CUGAUGAG X CGAA ICCGCUGC 2245
554 CAGCGGCC u GGGGGGCG 471 CGCCCCCC CUGAUGAG X CGAA IGCCGCUG 2246
564 GGGGGCGC c CCCCUGGG 472 CCCAGGGG CUGAUGAG X CGAA ICGCCCCC 2247
565 GGGGCGCC c CCCUGGGG 473 CCCCAGGG CUGAUGAG X CGAA IGCGCCCC 2248
566 GGGCGCCC c CCUGGGGC 474 GCCCCAGG CUGAUGAG X CGAA IGGCGCCC 2249
567 GGCGCCCC c CUGGGGCU 475 AGCCCCAG CUGAUGAG X CGAA IGGGCGCC 2250
568 GCGCCCCC c UGGGGCUG 476 CAGCCCCA CUGAUGAG X CGAA IGGGGCGC ~ 2251
569 CGCCCCCC u GGGGCUGC 477 GCAGCCCC CUGAUGAG X CGAA IGGGGGCG 2252
575 CCUGGGGC u GCGGCUGC 478 GCAGCCGC CUGAUGAG X CGAA ICCCCAGG 2253
581 GCUGCGGC u GCCCCGGG 479 CCCGGGGC CUGAUGAG X CGAA ICCGCAGC 2254
584 GCGGCUGC c CCGGGAGA 480 UCUCCCGG CUGAUGAG X CGAA ICAGCCGC 2255
585 CGGCUGCC c CGGGAGAC 481 GUCUCCCG CUGAUGAG X CGAA IGCAGCCG 2256
586 GGCUGCCC c GGGAGACC 482 GGUCUCCC CUGAUGAG X CGAA IGGCAGCC 2257
594 CGGGAGAC c GACGAAGA 483 UCUUCGUC CUGAUGAG X CGAA IUCUCCCG 2258
605 CGAAGAGC c CGAGGAGC 484 σcuccucG CUGAUGAG X CGAA ICUCUUCG 2259
Table 19
606 GAAGAGCC C GAGGAGCC 485 GGCUCCUC CUGAUGAG X CGAA IGCUCUUC 2260
614 CGAGGAGC C CGGCCGGA 486 UCCGGCCG CUGAUGAG X CGAA ICUCCUCG 2261
615 GAGGAGCC C GGCCGGAG 487 CUCCGGCC CUGAUGAG X CGAA IGCUCCUC 2262
619 AGCCCGGC C GGAGGGGC 488 GCCCCUCC CUGAUGAG X CGAA ICCGGGCU 2263
628 GGAGGGGC A GCUUUGUG 489 CACAAAGC CUGAUGAG X CGAA ICCCCUCC 2264
631 GGGGCAGC U UUGUGGAG 490 CUCCACAA CUGAUGAG X CGAA ICUGCCCC 2265
649 UGGUGGAC A ACCUGAGG 491 CCUCAGGU CUGAUGAG X CGAA IUCCACCA 2266
652 UGGACAAC C UGAGGGGC 492 GCCCCUCA CUGAUGAG X CGAA IUUGUCCA 2267
653 GGACAACC U GAGGGGCA 493 UGCCCCUC CUGAUGAG X CGAA IGUUGUCC 2268
661 UGAGGGGC A AGUCGGGG 494 CCCCGACU CUGAUGAG X CGAA ICCCCUCA 2269
671 GUCGGGGC A GGGCUACU 495 AGUAGCCC CUGAUGAG X CGAA ICCCCGAC 2270
676 GGCAGGGC U ACUACGUG 496 CACGUAGU CUGAUGAG X CGAA ICCCUGCC 2271
679 AGGGCUAC U ACGUGGAG 497 CUCCACGU CUGAUGAG X CGAA IUAGCCCU 2272
693 GAGAUGAC C GUGGGCAG 498 CUGCCCAC CUGAUGAG X CGAA IUCAUCUC 2273
700 CCGUGGGC A GCCCCCCG 499 CGGGGGGC CUGAUGAG X CGAA ICCCACGG 2274
703 UGGGCAGC C CCCCGCAG 500 CUGCGGGG CUGAUGAG X CGAA ICUGCCCA 2275
704 GGGCAGCC C CCCGCAGA 501 UCUGCGGG CUGAUGAG X CGAA IGCUGCCC 2276
705 GGCAGCCC C CCGCAGAC 502 GUCUGCGG CUGAUGAG X CGAA IGGCUGCC 2277
706 GCAGCCCC C CGCAGACG 503 CGUCUGCG CUGAUGAG X CGAA IGGGCUGC 2278
707 CAGCCCCC C GCAGACGC 504 GCGUCUGC CUGAUGAG X CGAA IGGGGCUG 2279
710 CCCCCCGC A GACGCUCA 505 UGAGCGUC CUGAUGAG X CGAA ICGGGGGG 2280
716 GCAGACGC U CAACAUCC 506 GGAUGUUG CUGAUGAG X CGAA ICGUCUGC 2281
718 AGACGCUC A ACAUCCUG 507 CAGGAUGU CUGAUGAG X CGAA IAGCGUCU 2282
721 CGCUCAAC A UCCUGGUG 508 CACCAGGA CUGAUGAG X CGAA IUUGAGCG 2283
724 UCAACAUC C UGGUGGAU 509 AUCCACCA CUGAUGAG X CGAA IAUGUUGA 2284
725 CAACAUCC U GGUGGAUA 510 UAUCCACC CUGAUGAG X CGAA IGAUGUUG 2285
735 GUGGAUAC A GGCAGCAG 511 CUGCUGCC CUGAUGAG X CGAA IUAUCCAC 2286
739 AUACAGGC A GCAGUAAC 512 GUUACUGC CUGAUGAG X CGAA ICCUGUAU 2287
742 CAGGCAGC A GUAACUUU 513 AAAGUUAC CUGAUGAG X CGAA ICUGCCUG 2288
748 GCAGUAAC U UUGCAGUG 514 CACUGCAA CUGAUGAG X CGAA IUUACUGC 2289
753 AACUUUGC A GUGGGUGC 515 GCACCCAC CUGAUGAG X CGAA ICAAAGUU 2290
762 GUGGGUGC U GCCCCCCA 516 UGGGGGGC CUGAUGAG X CGAA ICACCCAC 2291
765 GGUGCUGC C CCCCACCC 517 GGGUGGGG CUGAUGAG X CGAA ICAGCACC 2292
766 GUGCUGCC C CCCACCCC 518 GGGGUGGG CUGAUGAG X CGAA IGCAGCAC 2293
767 UGCUGCCC C CCACCCCU 519 AGGGGUGG CUGAUGAG X CGAA IGGCAGCA 2294
768 GCUGCCCC C CACCCCUU 520 AAGGGGUG CUGAUGAG X CGAA IGGGCAGC 2295
769 CUGCCCCC C ACCCCUUC 521 GAAGGGGU CUGAUGAG X CGAA IGGGGCAG 2296
770 UGCCCCCC A CCCCUUCC 522 GGAAGGGG CUGAUGAG X CGAA IGGGGGCA 2297
772 CCCCCCAC C CCUUCCUG 523 CAGGAAGG CUGAUGAG X CGAA IUGGGGGG 2298
773 CCCCCACC C CUUCCUGC 524 GCAGGAAG CUGAUGAG X CGAA IGUGGGGG 2299
774 CCCCACCC C UUCCUGCA 525 UGCAGGAA CUGAUGAG X CGAA IGGUGGGG 2300
775 CCCACCCC U UCCUGCAU 526 AUGCAGGA CUGAUGAG X CGAA IGGGUGGG 2301
778 ACCCCUUC C UGCAUCGC 527 GCGAUGCA CUGAUGAG X CGAA IAAGGGGU 2302
779 CCCCUUCC U GCAUCGCU 528 AGCGAUGC CUGAUGAG X CGAA IGAAGGGG 2303
782 CUUCCUGC A UCGCUACU 529 AGUAGCGA CUGAUGAG X CGAA ICAGGAAG 2304
787 UGCAUCGC U ACUACCAG 530 CUGGUAGU CUGAUGAG X CGAA ICGAUGCA 2305
790 AUCGCUAC U ACCAGAGG 531 CCUCUGGU CUGAUGAG X CGAA IUAGCGAU 2306
793 GCUACUAC C AGAGGCAG 532 CUGCCUCU CUGAUGAG X CGAA IUAGUAGC 2307
794 CUACUACC A GAGGCAGC 533 GCUGCCUC CUGAUGAG X CGAA IGUAGUAG 2308
800 CCAGAGGC A GCUGUCCA 534 UGGACAGC CUGAUGAG X CGAA ICCUCUGG 2309
803 GAGGCAGC U GUCCAGCA 535 UGCUGGAC CUGAUGAG X CGAA ICUGCCUC 2310
Table 19
807 CAGCUGUC C AGCACAUA 536 UAUGUGCU CUGAUGAG X CGAA I CAGCUG 2311
808 AGCUGUCC A GCACAUAC 537 GUAUGUGC CUGAUGAG X CGAA IGACAGCU 2312
811 UGUCCAGC A CAUACCGG 538 CCGGUAUG CUGAUGAG X CGAA ICUGGACA 2313
813 UCCAGCAC A UACCGGGA 539 UCCCGGUA CUGAUGAG X CGAA IUGCUGGA 2314
817 GCACAUAC C GGGACCUC 540 GAGGUCCC CUGAUGAG X CGAA IUAUGUGC 2315
823 ACCGGGAC C UCCGGAAG 541 CUUCCGGA CUGAUGAG X CGAA IUCCCGGU 2316
824 CCGGGACC U CCGGAAGG 542 CCUUCCGG CUGAUGAG X CGAA IGUCCCGG 2317
826 GGGACCUC C GGAAGGGU 543 ACCCUUCC CUGAUGAG X CGAA IAGGUCCC 2318
845 GUAUGUGC C CUACACCC 544 GGGUGUAG CUGAUGAG X CGAA ICACAUAC 2319
846 UAUGUGCC C UACACCCA 545 UGGGUGUA CUGAUGAG X CGAA IGCACAUA 2320
847 AUGUGCCC U ACACCCAG 546 CUGGGUGU CUGAUGAG X CGAA IGGCACAU 2321
850 UGCCCUAC A CCCAGGGC 547 GCCCUGGG CUGAUGAG X CGAA IUAGGGCA 2322
852 CCCUACAC C CAGGGCAA 548 UUGCCCUG CUGAUGAG X CGAA IUGUAGGG 2323
853 CCUACACC C AGGGCAAG 549 CUUGCCCU CUGAUGAG X CGAA IGUGUAGG 2324
854 CUACACCC A GGGCAAGU 550 ACUUGCCC CUGAUGAG X CGAA IGGUGUAG 2325
859 CCCAGGGC A AGUGGGAA 551 UUCCCACU CUGAUGAG X CGAA ICCCUGGG 2326
875 AGGGGAGC U GGGCACCG 552 CGGUGCCC CUGAUGAG X CGAA ICUCCCCU 2327
880 AGCUGGGC A CCGACCUG 553 CAGGUCGG CUGAUGAG X CGAA ICCCAGCU 2328
882 CUGGGCAC C GACCUGGU 554 ACCAGGUC CUGAUGAG X CGAA IUGCCCAG 2329
886 GCACCGAC C UGGUAAGC 555 GCUUACCA CUGAUGAG X CGAA IUCGGUGC 2330
887 CACCGACC U GGUAAGCA 556 UGCUUACC CUGAUGAG X CGAA IGUCGGUG 2331
895 UGGUAAGC A UCCCCCAU 557 AUGGGGGA CUGAUGAG X CGAA ICUUACCA 2332
898 UAAGCAUC C CCCAUGGC 558 GCCAUGGG CUGAUGAG X CGAA IAUGCUUA 2333
899 AAGCAUCC C CCAUGGCC 559 GGCCAUGG CUGAUGAG X CGAA IGAUGCUU 2334
900 AGCAUCCC C CAUGGCCC 560 GGGCCAUG CUGAUGAG X CGAA IGGAUGCU 2335
901 GCAUCCCC C AUGGCCCC 561 GGGGCCAU CUGAUGAG X CGAA IGGGAUGC 2336
902 CAUCCCCC A UGGCCCCA 562 UGGGGCCA CUGAUGAG X CGAA IGGGGAUG 2337
907 CCCAUGGC C CCAACGUC 563 GACGUUGG CUGAUGAG X CGAA ICCAUGGG 2338
908 CCAUGGCC C CAACGUCA 564 UGACGUUG CUGAUGAG X CGAA IGCCAUGG 2339
909 CAUGGCCC C AACGUCAC 565 GUGACGUU CUGAUGAG X CGAA IGGCCAUG 2340
910 AUGGCCCC A ACGUCACU 566 AGUGACGU CUGAUGAG X CGAA IGGGCCAU 2341
916 CCAACGUC A CUGUGCGU 567 ACGCACAG CUGAUGAG X CGAA IACGUUGG 2342
918 AACGUCAC U GUGCGUGC 568 GCACGCAC CUGAUGAG X CGAA IUGACGUU 2343
927 GUGCGUGC c AACAUUGC 569 GCAAUGUU CUGAUGAG X CGAA ICACGCAC 2344
928 UGCGUGCC A ACAUUGCU 570 AGCAAUGU CUGAUGAG X CGAA IGCACGCA 2345
931 GUGCCAAC A UUGCUGCC 571 GGCAGCAA CUGAUGAG X CGAA IUUGGCAC 2346
936 AACAUUGC U GCCAUCAC 572 GUGAUGGC CUGAUGAG X CGAA ICAAUGUU 2347
939 AUUGCUGC C AUCACUGA 573 UCAGUGAU CUGAUGAG X CGAA ICAGCAAU 2348
940 UUGCUGCC A UCACUGAA 574 UUCAGUGA CUGAUGAG X CGAA IGCAGCAA 2349
943 CUGCCAUC A CUGAAUCA 575 UGAUUCAG CUGAUGAG X CGAA IAUGGCAG 2350
945 GCCAUCAC U GAAUCAGA 576 UCUGAUUC CUGAUGAG X CGAA IUGAUGGC 2351
951 ACUGAAUC A GACAAGUU 577 AACUUGUC CUGAUGAG X CGAA IAUUCAGU 2352
955 AAUCAGAC A AGUUCUUC 578 GAAGAACU CUGAUGAG X CGAA IUCUGAUU 2353
961 ACAAGUUC U UCAUCAAC 579 GUUGAUGA CUGAUGAG X CGAA IAACUUGU 2354
964 AGUUCUUC A UCAACGGC 580 GCCGUUGA CUGAUGAG X CGAA IAAGAACU 2355
967 UCUUCAUC A ACGGCUCC 581 GGAGCCGU CUGAUGAG X CGAA IAUGAAGA 2356
973 UCAACGGC U CCAACUGG 582 CCAGUUGG CUGAUGAG X CGAA ICCGUUGA 2357
975 AACGGCUC C AACUGGGA 583 UCCCAGUU CUGAUGAG X CGAA lAGCCGUU 2358
976 ACGGCUCC A ACUGGGAA 584 UUCCCAGU CUGAUGAG X CGAA IGAGCCGU 2359
979 GCUCCAAC U GGGAAGGC 585 GCCUUCCC CUGAUGAG X CGAA IUUGGAGC 2360
988 GGGAAGGC A UCCUGGGG 586 CCCCAGGA CUGAUGAG X CGAA ICCUUCCC 2361
Table 19
991 AAGGCAUC C UGGGGCUG 587 CAGCCCCA CUGAUGAG X CGAA IAUGCCUU 2362
992 AGGCAUCC U GGGGCUGG 588 CCAGCCCC CUGAUGAG X CGAA IGAUGCCU 2363
998 CCUGGGGC U GGCCUAUG 589 CAUAGGCC CUGAUGAG X CGAA ICCCCAGG 2364
1002 GGGCUGGC C UAUGCUGA 590 UCAGCAUA CUGAUGAG X CGAA ICCAGCCC 2365
1003 GGCUGGCC U AUGCUGAG 591 CUCAGCAU CUGAUGAG X CGAA IGCCAGCC 2366
1008 GCCUAUGC U GAGAUUGC 592 GCAAUCUC CUGAUGAG X CGAA ICAUAGGC 2367
1017 GAGAUUGC C AGGCCUGA 593 UCAGGCCU CUGAUGAG X CGAA ICAAUCUC 2368
1018 AGAUUGCC A GGCCUGAC 594 GUCAGGCC CUGAUGAG X CGAA IGCAAUCU 2369
1022 UGCCAGGC C UGACGACU 595 AGUCGUCA CUGAUGAG X CGAA ■ICCUGGCA 2370
1023 GCCAGGCC U GACGACUC 596 GAGUCGUC CUGAUGAG X CGAA IGCCUGGC 2371
1030 CUGACGAC U CCCUGGAG 597 CUCCAGGG CUGAUGAG X CGAA IUCGUCAG 2372
1032 GACGACUC c CUGGAGCC 598 GGCUCCAG CUGAUGAG X CGAA IAGUCGUC 2373
1033 ACGACUCC c UGGAGCCU 599 AGGCUCCA CUGAUGAG X CGAA IGAGUCGU 2374
1034 CGACUCCC u GGAGCCUU 600 AAGGCUCC CUGAUGAG X CGAA IGGAGUCG 2375
1040 CCUGGAGC c UUUCUUUG 601 CAAAGAAA CUGAUGAG X CGAA ICUCCAGG 2376
1041 CUGGAGCC u UUCUUUGA 602 UCAAAGAA CUGAUGAG X CGAA IGCUCCAG 2377
1045 AGCCUUUC u UUGACUCU 603 AGAGUCAA CUGAUGAG X CGAA IAAAGGCU 2378
1051 UCUUUGAC u CUCUGGUA 604 UACCAGAG CUGAUGAG X CGAA IUCAAAGA 2379
1053 UUUGACUC u CUGGUAAA 605 UUUACCAG CUGAUGAG X CGAA IAGUCAAA 2380
1055 UGACUCUC u GGUAAAGC 606 GCUUUACC CUGAUGAG X CGAA IAGAGUCA 2381
1064 GGUAAAGC A GACCCACG 607 CGUGGGUC CUGAUGAG X CGAA ICUUUACC 2382
1068 AAGCAGAC C CACGUUCC 608 GGAACGUG CUGAUGAG X CGAA IUCUGCUU 2383
1069 AGCAGACC c ACGUUCCC 609 GGGAACGU CUGAUGAG X CGAA IGUCUGCU 2384
1070 GCAGACCC A CGUUCCCA 610 UGGGAACG CUGAUGAG X CGAA IGGUCUGC 2385
1076 CCACGUUC C CAACCUCU 611 AGAGGUUG CUGAUGAG X CGAA IAACGUGG 2386
1077 CACGUUCC C AACCUCUU 612 AAGAGGUU CUGAUGAG X CGAA IGAACGUG 2387
1078 ACGUUCCC A ACCUCUUC 613 GAAGAGGU CUGAUGAG X CGAA IGGAACGU 2388
1081 UUCCCAAC C UCUUCUCC 614 GGAGAAGA CUGAUGAG X CGAA IUUGGGAA 2389
1082 UCCCAACC U CUUCUCCC 615 GGGAGAAG CUGAUGAG X CGAA IGUUGGGA 2390
1084 CCAACCUC U UCUCCCUG 616 CAGGGAGA CUGAUGAG X CGAA IAGGUUGG 2391
1087 ACCUCUUC U CCCUGCAG 617 CUGCAGGG CUGAUGAG X CGAA IAAGAGGU 2392
1089 CUCUUCUC C CUGCAGCU 618 AGCUGCAG CUGAUGAG X CGAA IAGAAGAG 2393
1090 UCUUCUCC C UGCAGCUU 619 AAGCUGCA CUGAUGAG X CGAA IGAGAAGA 2394
1091 cuucuccc u GCAGCUUU 620 AAAGCUGC CUGAUGAG X CGAA IGGAGAAG 2395
1094 CUCCCUGC A GCUUUGUG 621 CACAAAGC CUGAUGAG X CGAA ICAGGGAG 2396
1097 CCUGCAGC U UUGUGGUG 622 CACCACAA CUGAUGAG X CGAA ICUGCAGG 2397
1107 UGUGGUGC U GGCUUCCC 623 GGGAAGCC CUGAUGAG X CGAA ICACCACA 2398 llll GUGCUGGC U UCCCCCUC 624 GAGGGGGA CUGAUGAG X CGAA ICCAGCAC 2399
1114 CUGGCUUC C CCCUCAAC 625 GUUGAGGG CUGAUGAG X CGAA IAAGCCAG 2400
1115 UGGCUUCC C CCUCAACC 626 GGUUGAGG CUGAUGAG X CGAA IGAAGCCA 2401
1116 GGCUUCCC C CUCAACCA 627 UGGUUGAG CUGAUGAG X CGAA IGGAAGCC 2402
1117 GCUUCCCC C UCAACCAG 628 CUGGUUGA CUGAUGAG X CGAA IGGGAAGC 2403
1118 CUUCCCCC U CAACCAGU 629 ACUGGUUG CUGAUGAG X CGAA IGGGGAAG 2404
1120 UCCCCCUC A ACCAGUCU 630 AGACUGGU CUGAUGAG X CGAA lAGGGGGA 2405
1123 CCCUCAAC C AGUCUGAA 631 UUCAGACU CUGAUGAG X CGAA IUUGAGGG 2406
1124 CCUCAACC A GUCUGAAG 632 CUUCAGAC CUGAUGAG X CGAA IGUUGAGG 2407
1128 AACCAGUC U GAAGUGCU 633 AGCACUUC CUGAUGAG X CGAA lACUGGUU 2408
1136 UGAAGUGC U GGCCUCUG 634 CAGAGGCC CUGAUGAG X CGAA ICACUUCA 2409
1140 GUGCUGGC C UCUGUCGG 635 CCGACAGA CUGAUGAG X CGAA ICCAGCAC 2410
1141 UGCUGGCC U CUGUCGGA 636 UCCGACAG CUGAUGAG X CGAA IGCCAGCA 2411 •
1143 CUGGCCUC U GUCGGAGG 637 CCUCCGAC CUGAUGAG X CGAA IAGGCCAG 2412
Table 19
1156 GAGGGAGC A UGAUCAUU 638 AAUGAUCA CUGAUGAG X CGAA ICUCCCUC 2413
1162 GCAUGAUC A UUGGAGGU 639 ACCUCCAA CUGAUGAG X CGAA IAUCAUGC 2414
1177 GUAUCGAC C ACUCGCUG 640 CAGCGAGU CUGAUGAG X CGAA IUCGAUAC 2415
1178 UAUCGACC A CUCGCUGU 641 ACAGCGAG CUGAUGAG X CGAA IGUCGAUA 2416
1180 UCGACCAC U CGCUGUAC 642 GUACAGCG CUGAUGAG X CGAA IUGGUCGA 2417
1184 CCACUCGC U GUACACAG 643 CUGUGUAC CUGAUGAG X CGAA ICGAGUGG 2418
1189 CGCUGUAC A CAGGCAGU 644 ACUGCCUG CUGAUGAG X CGAA IUACAGCG 2419
1191 CUGUACAC A GGCAGUCU 645 AGACUGCC CUGAUGAG X CGAA IUGUACAG 2420
1195 ACACAGGC A GUCUCUGG 646 CCAGAGAC CUGAUGAG X CGAA ICCUGUGU 2421
1199 AGGCAGUC U CUGGUAUA 647 UAUACCAG CUGAUGAG X CGAA IACUGCCU 2422
1201 GCAGUCUC U GGUAUACA 648 UGUAUACC CUGAUGAG X CGAA IAGACUGC 2423
1209 UGGUAUAC A CCCAUCCG 649 CGGAUGGG CUGAUGAG X CGAA IUAUACCA 2424
1211 GUAUACAC C CAUCCGGC 650 GCCGGAUG CUGAUGAG X CGAA IUGUAUAC 2425
1212 UAUACACC C AUCCGGCG 651 CGCCGGAU CUGAUGAG X CGAA IGUGUAUA 2426
1213 AUACACCC A UCCGGCGG 652 CCGCCGGA CUGAUGAG X CGAA IGGUGUAU 2427
1216 CACCCAUC C GGCGGGAG 653 CUCCCGCC CUGAUGAG X CGAA IAUGGGUG 2428
1243 AGGUGAUC A UUGUGCGG 654 CCGCACAA CUGAUGAG X CGAA IAUCACCU 2429
1261 UGGAGAUC A AUGGACAG 655 CUGUCCAU CUGAUGAG X CGAA IAUCUCCA 2430
1268 CAAUGGAC A GGAUCUGA 656 UCAGAUCC CUGAUGAG X CGAA IUCCAUUG 2431
1274 ACAGGAUC U GAAAAUGG 657 CCAUUUUC CUGAUGAG X CGAA IAUCCUGU 2432
1285 AAAUGGAC U GCAAGGAG 658 CUCCUUGC CUGAUGAG X CGAA IUCCAUUU 2433
1288 UGGACUGC A AGGAGUAC 659 GUACUCCU CUGAUGAG X CGAA ICAGUCCA 2434
1297 AGGAGUAC A ACUAUGAC 660 GUCAUAGU CUGAUGAG X CGAA IUACUCCU 2435
1300 AGUACAAC U AUGACAAG 661 CUUGUCAU CUGAUGAG X CGAA IUUGUACU 2436
1306 ACUAUGAC A AGAGCAUU 662 AAUGCUCU CUGAUGAG X CGAA IUCAUAGU 2437
1312 ACAAGAGC A UUGUGGAC 663 GUCCACAA CUGAUGAG X CGAA ICUCUUGU 2438
1321 UUGUGGAC A GUGGCACC 664 GGUGCCAC CUGAUGAG X CGAA IUCCACAA 2439
1327 ACAGUGGC A CCACCAAC 665 GUUGGUGG CUGAUGAG X CGAA ICCACUGU 2440
1329 AGUGGCAC C ACCAACCU 666 AGGUUGGU CUGAUGAG X CGAA IUGCCACU 2441
1330 GUGGCACC A CCAACCUU 667 AAGGUUGG CUGAUGAG X CGAA IGUGCCAC 2442
1332 GGCACCAC C AACCUUCG 668 CGAAGGUU CUGAUGAG X CGAA IUGGUGCC 2443
1333 GCACCACC A ACCUUCGU 669 ACGAAGGU CUGAUGAG X CGAA IGUGGUGC 2444
1336 CCACCAAC C UUCGUUUG 670 CAAACGAA CUGAUGAG X CGAA IUUGGUGG 2445
1337 CACCAACC U UCGUUUGC 671 GCAAACGA CUGAUGAG X CGAA IGUUGGUG 2446
1346 UCGUUUGC C CAAGAAAG 672 CUUUCUUG CUGAUGAG X CGAA ICAAACGA 2447
1347 CGUUUGCC C AAGAAAGU 673 ACUUUCUU CUGAUGAG X CGAA IGCAAACG 2448
1348 GUUUGCCC A AGAAAGUG 674 CACUUUCU CUGAUGAG X CGAA IGGCAAAC 2449
1365 UUUGAAGC U GCAGUCAA 675 UUGACUGC CUGAUGAG X CGAA ICUUCAAA 2450
1368 GAAGCUGC A GUCAAAUC 676 GAUUUGAC CUGAUGAG X CGAA ICAGCUUC 2451
1372 CUGCAGUC A AAUCCAUC 677 GAUGGAUU CUGAUGAG X CGAA IACUGCAG 2452
1377 GUCAAAUC C AUCAAGGC 678 GCCUUGAU CUGAUGAG X CGAA IAUUUGAC 2453
1378 UCAAAUCC A UCAAGGCA 679 UGCCUUGA CUGAUGAG X CGAA IGAUUUGA 2454
1381 AAUCCAUC A AGGCAGCC 680 GGCUGCCU CUGAUGAG X CGAA IAUGGAUU 2455
1386 AUCAAGGC A GCCUCCUC 681 GAGGAGGC CUGAUGAG X CGAA ICCUUGAU 2456
1389 AAGGCAGC C UCCUCCAC 682 GUGGAGGA CUGAUGAG X CGAA ICUGCCUU 2457
1390 AGGCAGCC U CCUCCACG 683 CGUGGAGG CUGAUGAG X CGAA IGCUGCCU 2458
1392 GCAGCCUC C UCCACGGA 684 UCCGUGGA CUGAUGAG X CGAA IAGGCUGC 2459
1393 CAGCCUCC U CCACGGAG 685 CUCCGUGG CUGAUGAG X CGAA IGAGGCUG 2460
1395 GCCUCCUC C ACGGAGAA 686 UUCUCCGU CUGAUGAG X CGAA IAGGAGGC 2461
1396 CCUCCUCC A CGGAGAAG 687 CUUCUCCG CUGAUGAG X CGAA IGAGGAGG 2462
1408 AGAAGUUC C CUGAUGGU 688 ACCAUCAG CUGAUGAG X CGAA lAACUUCU 2463
Table 19
1409 GAAGUUCC C UGAUGGUU 689 AACCAUCA CUGAUGAG X CGAA IGAACUUC 2464
1410 AAGUUCCC U GAUGGUUU 690 AAACCAUC CUGAUGAG X CGAA IGGAACUU 2465
1420 AUGGUUUC U GGCUAGGA 691 UCCUAGCC CUGAUGAG X CGAA IAAACCAU 2466
1424 UUUCUGGC U AGGAGAGC 692 GCUCUCCU CUGAUGAG X CGAA ICCAGAAA 2467
1433 AGGAGAGC A GCUGGUGU 693 ACACCAGC CUGAUGAG X CGAA ICUCUCCU 2468
1436 AGAGCAGC U GGUGUGCU 694 AGCACACC CUGAUGAG X CGAA ICUGCUCU 2469
1444 UGGUGUGC U GGCAAGCA 695 UGCUUGCC CUGAUGAG X CGAA ICACACCA 2470
1448 GUGCUGGC A AGCAGGCA 696 UGCCUGCU CUGAUGAG X CGAA ICCAGCAC 2471
1452 UGGCAAGC A GGCACCAC 697 GUGGUGCC CUGAUGAG X CGAA ICUUGCCA 2472
1456 AAGCAGGC A CCACCCCU 698 AGGGGUGG CUGAUGAG X CGAA ICCUGCUU 2473
1458 GCAGGCAC C ACCCCUUG 699 CAAGGGGU CUGAUGAG X CGAA IUGCCUGC 2474
1459 CAGGCACC A CCCCUUGG 700 CCAAGGGG CUGAUGAG X CGAA IGUGCCUG 2475
1461 GGCACCAC C CCUUGGAA 701 UUCCAAGG CUGAUGAG X CGAA IUGGUGCC 2476
1462 GCACCACC C CUUGGAAC 702 GUUCCAAG CUGAUGAG X CGAA IGUGGUGC 2477
1463 CACCACCC C UUGGAACA 703 UGUUCCAA CUGAUGAG X CGAA IGGUGGUG 2478
1464 ACCACCCC U UGGAACAU 704 AUGUUCCA CUGAUGAG X CGAA IGGGUGGU 2479
1471 CUUGGAAC A UUUUCCCA 705 UGGGAAAA CUGAUGAG X CGAA IUUCCAAG 2480
1477 ACAUUUUC C CAGUCAUC 706 GAUGACUG CUGAUGAG X CGAA IAAAAUGU 2481
1478 CAUUUUCC C AGUCAUCU 707 AGAUGACU CUGAUGAG X CGAA IGAAAAUG 2482
1479 AUUUUCCC A GUCAUCUC 708 GAGAUGAC CUGAUGAG X CGAA IGGAAAAU 2483
1483 UCCCAGUC A UCUCACUC 709 GAGUGAGA CUGAUGAG X CGAA IACUGGGA 2484
1486 CAGUCAUC U CACUCUAC 710 GUAGAGUG CUGAUGAG X CGAA IAUGACUG 2485
1488 GUCAUCUC A CUCUACCU 711 AGGUAGAG CUGAUGAG X CGAA IAGAUGAC 2486
1490 CAUCUCAC U CUACCUAA 712 UUAGGUAG CUGAUGAG X CGAA IUGAGAUG 2487
1492 UCUCACUC U ACCUAAUG 713 CAUUAGGU CUGAUGAG X CGAA IAGUGAGA 2488
1495 CACUCUAC C UAAUGGGU 714 ACCCAUUA CUGAUGAG X CGAA IUAGAGUG 2489
1496 ACUCUACC U AAUGGGUG 715 CACCCAUU CUGAUGAG X CGAA IGUAGAGU 2490
1512 GAGGUUAC C AACCAGUC 716 GACUGGUU CUGAUGAG X CGAA IUAACCUC 2491
1513 AGGUUACC A ACCAGUCC 717 GGACUGGU CUGAUGAG X CGAA IGUAACCU 2492
1516 UUACCAAC C AGUCCUUC 718 GAAGGACU CUGAUGAG X CGAA IUUGGUAA 2493
1517 UACCAACC A GUCCUUCC 719 GGAAGGAC CUGAUGAG X CGAA IGUUGGUA 2494
1521 AACCAGUC C UUCCGCAU 720 AUGCGGAA CUGAUGAG X CGAA lACUGGUU 2495
1522 ACCAGUCC U UCCGCAUC 721 GAUGCGGA CUGAUGAG X CGAA IGACUGGU 2496
1525 AGUCCUUC C GCAUCACC 722 GGUGAUGC CUGAUGAG X CGAA IAAGGACU 2497
1528 CCUUCCGC A UCACCAUC 723 GAUGGUGA CUGAUGAG X CGAA ICGGAAGG 2498
1531 UCCGCAUC A CCAUCCUU 724 AAGGAUGG CUGAUGAG X CGAA lAUGCGGA 2499
1533 CGCAUCAC C AUCCUUCC 725 GGAAGGAU CUGAUGAG X CGAA IUGAUGCG 2500
1534 GCAUCACC A UCCUUCCG 726 CGGAAGGA CUGAUGAG X CGAA IGUGAUGC 2501
1537 UCACCAUC C UUCCGCAG 727 CUGCGGAA CUGAUGAG X CGAA IAUGGUGA 2502
1538 CACCAUCC U UCCGCAGC 728 GCUGCGGA CUGAUGAG X CGAA IGAUGGUG 2503
1541 CAUCCUUC C GCAGCAAU 729 AUUGCUGC CUGAUGAG X CGAA IAAGGAUG 2504
1544 CCUUCCGC A GCAAUACC 730 GGUAUUGC CUGAUGAG X CGAA ICGGAAGG 2505
1547 UCCGCAGC A AUACCUGC 731 GCAGGUAU CUGAUGAG X CGAA ICUGCGGA 2506
1552 AGCAAUAC C UGCGGCCA 732 UGGCCGCA CUGAUGAG X CGAA IUAUUGCU 2507
1553 GCAAUACC U GCGGCCAG 733 CUGGCCGC CUGAUGAG X CGAA IGUAUUGC 2508
1559 CCUGCGGC C AGUGGAAG 734 CUUCCACU CUGAUGAG X CGAA ICCGCAGG 2509
1560 CUGCGGCC A GUGGAAGA 735 UCUUCCAC CUGAUGAG X CGAA IGCCGCAG 2510
1575 GAUGUGGC C ACGUCCCA 736 UGGGACGU CUGAUGAG X CGAA ICCACAUC 2511
1576 AUGUGGCC A CGUCCCAA 737 UUGGGACG CUGAUGAG X CGAA IGCCACAU 2512
1581 GCCACGUC C CAAGACGA 738 UCGUCUUG CUGAUGAG X CGAA IACGUGGC 2513
1582 CCACGUCC C AAGACGAC 739 GUCGUCUU CUGAUGAG X CGAA IGACGUGG 2514
Table 19
1583 CACGUCCC A AGACGACU 740 AGUCGUCU CUGAUGAG X CGAA IGGACGUG 2515
1591 AAGACGAC U GUUACAAG 741 CUUGUAAC CUGAUGAG X CGAA IUCGUCUU 2516
1597 ACUGUUAC A AGUUUGCC 742 GGCAAACU CUGAUGAG X CGAA IUAACAGU 2517
1605 AAGUUUGC C AUCUCACA 743 UGUGAGAU CUGAUGAG X CGAA ICAAACUU 2518
1606 AGUUUGCC A UCUCACAG 744 CUGUGAGA CUGAUGAG X CGAA IGCAAACU 2519
1609 UUGCCAUC U CACAGUCA 745 UGACUGUG CUGAUGAG X CGAA IAUGGCAA 2520
1611 GCCAUCUC A CAGUCAUC 746 GAUGACUG CUGAUGAG X CGAA IAGAUGGC 2521
1613 CAUCUCAC A GUCAUCCA 747 UGGAUGAC CUGAUGAG X CGAA IUGAGAUG 2522
1617 UCACAGUC A UCCACGGG 748 CCCGUGGA CUGAUGAG X CGAA IACUGUGA 2523
1620 CAGUCAUC C ACGGGCAC 749 GUGCCCGU CUGAUGAG X CGAA IAUGACUG 2524
1621 AGUCAUCC A CGGGCACU 750 AGUGCCCG CUGAUGAG X CGAA IGAUGACU 2525
1627 CCACGGGC A CUGUUAUG 751 CAUAACAG CUGAUGAG X CGAA ICCCGUGG 2526
1629 ACGGGCAC U GUUAUGGG 752 CCCAUAAC CUGAUGAG X CGAA IUGCCCGU 2527
1641 AUGGGAGC U GUUAUCAU 753 AUGAUAAC CUGAUGAG X CGAA ICUCCCAU 2528
1648 CUGUUAUC A UGGAGGGC 754 GCCCUCCA CUGAUGAG X CGAA IAUAACAG 2529
1657 UGGAGGGC U UCUACGUU 755 AACGUAGA CUGAUGAG X CGAA ICCCUCCA 2530
1660 AGGGCUUC U ACGUUGUC 756 GACAACGU CUGAUGAG X CGAA IAAGCCCU 2531
1669 ACGUUGUC U UUGAUCGG 757 CCGAUCAA CUGAUGAG X CGAA IACAACGU 2532
1680 GAUCGGGC C CGAAAACG 758 CGUUUUCG CUGAUGAG X CGAA ICCCGAUC 2533
1681 AUCGGGCC C GAAAACGA 759 UCGUUUUC CUGAUGAG X CGAA IGCCCGAU 2534
1696 GAAUUGGC U UUGCUGUC 760 GACAGCAA CUGAUGAG X CGAA ICCAAUUC 2535
1701 GGCUUUGC U GUCAGCGC 761 GCGCUGAC CUGAUGAG X CGAA ICAAAGCC 2536
1705 UUGCUGUC A GCGCUUGC 762 GCAAGCGC CUGAUGAG X CGAA lACAGCAA 2537
1710 GUCAGCGC U UGCCAUGU 763 ACAUGGCA CUGAUGAG X CGAA ICGCUGAC 2538
1714 GCGCUUGC C AUGUGCAC 764 GUGCACAU CUGAUGAG X CGAA ICAAGCGC 2539
1715 CGCUUGCC A UGUGCACG 765 CGUGCACA CUGAUGAG X CGAA IGCAAGCG 2540
1721 CCAUGUGC A CGAUGAGU 766 ACUCAUCG CUGAUGAG X CGAA ICACAUGG 2541
1732 AUGAGUUC A GGACGGCA 767 UGCCGUCC CUGAUGAG X CGAA IAACUCAU 2542
1740 AGGACGGC A GCGGUGGA 768 UCCACCGC CUGAUGAG X CGAA ICCGUCCU 2543
1753 UGGAAGGC C CUUUUGUC 769 GACAAAAG CUGAUGAG X CGAA ICCUUCCA 2544
1754 GGAAGGCC C UUUUGUCA 770 UGACAAAA CUGAUGAG X CGAA IGCCUUCC 2545
1755 GAAGGCCC U UUUGUCAC 771 GUGACAAA CUGAUGAG X CGAA IGGCCUUC 2546
1762 CUUUUGUC A CCUUGGAC 772 GUCCAAGG CUGAUGAG X CGAA IACAAAAG 2547
1764 UUUGUCAC C UUGGACAU 773 AUGUCCAA CUGAUGAG X CGAA lUGACAAA 2548
1765 UUGUCACC U UGGACAUG 774 CAUGUCCA CUGAUGAG X CGAA IGUGACAA 2549
1771 CCUUGGAC A UGGAAGAC 775 GUCUUCCA CUGAUGAG X CGAA IUCCAAGG 2550
1780 UGGAAGAC U GUGGCUAC 776 GUAGCCAC CUGAUGAG X CGAA IUCUUCCA 2551
1786 ACUGUGGC U ACAACAUU 777 AAUGUUGU CUGAUGAG X CGAA ICCACAGU 2552
1789 GUGGCUAC A ACAUUCCA 778 UGGAAUGU CUGAUGAG X CGAA IUAGCCAC 2553
1792 GCUACAAC A UUCCACAG 779 CUGUGGAA CUGAUGAG X CGAA IUUGUAGC 2554
1796 CAACAUUC C ACAGACAG 780 CUGUCUGU CUGAUGAG X CGAA IAAUGUUG 2555
1797 AACAUUCC A CAGACAGA 781 UCUGUCUG CUGAUGAG X CGAA IGAAUGUU 2556
1799 CAUUCCAC A GACAGAUG 782 CAUCUGUC CUGAUGAG X CGAA IUGGAAUG 2557
1803 CCACAGAC A GAUGAGUC 783 GACUCAUC CUGAUGAG X CGAA IUCUGUGG 2558
1812 GAUGAGUC A ACCCUCAU 784 AUGAGGGU CUGAUGAG X CGAA IACUCAUC 2559
1815 GAGUCAAC C CUCAUGAC 785 GUCAUGAG CUGAUGAG X CGAA IUUGACUC 2560
1816 AGUCAACC C UCAUGACC 786 GGUCAUGA CUGAUGAG X CGAA IGUUGACU 2561
1817 GUCAACCC U CAUGACCA 787 UGGUCAUG CUGAUGAG X CGAA IGGUUGAC 2562
1819 CAACCCUC A UGACCAUA 788 ' UAUGGUCA CUGAUGAG X CGAA IAGGGUUG 2563
1824 CUCAUGAC C AUAGCCUA 789 UAGGCUAU CUGAUGAG X CGAA IUCAUGAG 2564
1825 UCAUGACC -A UAGCCUAU 790 AUAGGCUA CUGAUGAG X CGAA IGUCAUGA 2565
Table 19
1830 ACCAUAGC C UAUGUCAU 791 AUGACAUA CUGAUGAG X CGAA ICUAUGGU 2566
1831 CCAUAGCC U AUGUCAUG 792 CAUGACAU CUGAUGAG X CGAA IGCUAUGG 2567
1837 CCUAUGUC A UGGCUGCC 793 GGCAGCCA CUGAUGAG X CGAA IACAUAGG 2568
1842 GUCAUGGC U GCCAUCUG 794 CAGAUGGC CUGAUGAG X CGAA ICCAUGAC 2569
1845 AUGGCUGC C AUCUGCGC 795 GCGCAGAU CUGAUGAG X CGAA ICAGCCAU 2570
1846 UGGCUGCC A UCUGCGCC 796 GGCGCAGA CUGAUGAG X CGAA IGCAGCCA 2571
1849 CUGCCAUC U GCGCCCUC 797 GAGGGCGC CUGAUGAG X CGAA IAUGGCAG 2572
1854 AUCUGCGC C CUCUUCAU 798 AUGAAGAG CUGAUGAG X CGAA ICGCAGAU 2573
1855 UCUGCGCC C UCUUCAUG 799 CAUGAAGA CUGAUGAG X CGAA IGCGCAGA 2574
1856 CUGCGCCC U CUUCAUGC 800 GCAUGAAG CUGAUGAG X CGAA IGGCGCAG 2575
1858 GCGCCCUC U UCAUGCUG 801 CAGCAUGA CUGAUGAG X CGAA IAGGGCGC 2576
1861 CCCUCUUC A UGCUGCCA 802 UGGCAGCA CUGAUGAG X CGAA IAAGAGGG 2577
1865 CUUCAUGC U GCCACUCU 803 AGAGUGGC CUGAUGAG X CGAA ICAUGAAG 2578
1868 CAUGCUGC C ACUCUGCC 804 GGCAGAGU CUGAUGAG X CGAA ICAGCAUG 2579
1869 AUGCUGCC A CUCUGCCU 805 AGGCAGAG CUGAUGAG X CGAA IGCAGCAU 2580
1871 GCUGCCAC U CUGCCUCA 806 UGAGGCAG CUGAUGAG X CGAA IUGGCAGC 2581
1873 UGCCACUC U GCCUCAUG 807 CAUGAGGC CUGAUGAG X CGAA lAGUGGCA 2582
1876 CACUCUGC C UCAUGGUG 808 CACCAUGA CUGAUGAG X CGAA ICAGAGUG 2583
1877 ACUCUGCC U CAUGGUGU 809 ACACCAUG CUGAUGAG X CGAA IGCAGAGU 2584
1879 UCUGCCUC A UGGUGUGU 810 ACACACCA CUGAUGAG X CGAA IAGGCAGA 2585
1889 GGUGUGUC A GUGGQGCU 811 AGCGCCAC CUGAUGAG X CGAA IACACACC 2586
1897 AGUGGCGC U GCCUCCGC 812 GCGGAGGC CUGAUGAG X CGAA ICGCCACU 2587
1900 GGCGCUGC C UCCGCUGC 813 GCAGCGGA CUGAUGAG X CGAA ICAGCGCC 2588
1901 GCGCUGCC U CCGCUGCC 814 GGCAGCGG CUGAUGAG X CGAA IGCAGCGC 2589
1903 GCUGCCUC C GCUGCCUG 815 CAGGCAGC CUGAUGAG X CGAA IAGGCAGC 2590
1906 GCCUCCGC u GCCUGCGC 816 GCGCAGGC CUGAUGAG X CGAA ICGGAGGC 2591
1909 UCCGCUGC c UGCGCCAG 817 CUGGCGCA CUGAUGAG X CGAA ICAGCGGA 2592
1910 CCGCUGCC u GCGCCAGC 818 GCUGGCGC CUGAUGAG X CGAA IGCAGCGG 2593
1915 GCCUGCGC c AGCAGCAU 819 AUGCUGCU CUGAUGAG X CGAA ICGCAGGC 2594
1916 CCUGCGCC A GCAGCAUG 820 CAUGCUGC CUGAUGAG X CGAA IGCGCAGG 2595
1919 GCGCCAGC A GCAUGAUG 821 CAUCAUGC CUGAUGAG X CGAA ICUGGCGC 2596
1922 CCAGCAGC A UGAUGACU 822 AGUCAUCA CUGAUGAG X CGAA ICUGCUGG 2597
1930 AUGAUGAC u UUGCUGAU 823 AUCAGCAA CUGAUGAG X CGAA lUCAUCAU 2598
1935 GACUUUGC U GAUGACAU 824 AUGUCAUC CUGAUGAG X CGAA ICAAAGUC 2599
1942 CUGAUGAC A UCUCCCUG '825 CAGGGAGA CUGAUGAG X CGAA IUCAUCAG 2600
1945 AUGACAUC U CCCUGCUG 826 CAGCAGGG CUGAUGAG X CGAA IAUGUCAU 2601
1947 GACAUCUC C CUGCUGAA 827 UUCAGCAG CUGAUGAG X CGAA IAGAUGUC 2602
1948 ACAUCUCC C UGCUGAAG 828 CUUCAGCA CUGAUGAG X CGAA IGAGAUGU 2603
1949 CAUCUCCC U GCUGAAGU 829 ACUUCAGC CUGAUGAG X CGAA IGGAGAUG 2604
1952 CUCCCUGC U GAAGUGAG 830 CUCACUUC CUGAUGAG X CGAA ICAGGGAG 2605
1966 GAGGAGGC C CAUGGGCA 831 UGCCCAUG CUGAUGAG X CGAA ICCUCCUC 2606
1967 AGGAGGCC C AUGGGCAG 832 CUGCCCAU CUGAUGAG X CGAA IGCCUCCU 2607
1968 GGAGGCCC A UGGGCAGA 833 UCUGCCCA CUGAUGAG X CGAA IGGCCUCC 2608
1974 CCAUGGGC A GAAGAUAG 834 CUAUCUUC CUGAUGAG X CGAA ICCCAUGG 2609
1989 AGAGAUUC C CCUGGACC 835 GGUCCAGG CUGAUGAG X CGAA IAAUCUCU 2610
1990 GAGAUUCC C CUGGACCA 836 UGGUCCAG CUGAUGAG X CGAA IGAAUCUC 2611
1991 AGAUUCCC C UGGACCAC 837 GUGGUCCA CUGAUGAG X CGAA IGGAAUCU 2612
1992 GAUUCCCC u GGACCACA 838 UGUGGUCC CUGAUGAG X CGAA IGGGAAUC 2613
1997 CCCUGGAC c ACACCUCC 839 GGAGGUGU CUGAUGAG X CGAA IUCCAGGG 2614
1998 CCUGGACC A CACCUCCG 840 CGGAGGUG CUGAUGAG X CGAA IGUCCAGG 2615
2000 UGGACCAC A CCUCCGUG 841 CACGGAGG CUGAUGAG X CGAA IUGGUCCA 2616
Table 19
2002 GACCACAC C UCCGUGGU 842 ACCACGGA CUGAUGAG X CGAA IUGUGGUC 2617
2003 ACCACACC U CCGUGGUU 843 AACCACGG CUGAUGAG X CGAA IGUGUGGU 2618
2005 CACACCUC C GUGGUUCA 844 UGAACCAC CUGAUGAG X CGAA IAGGUGUG 2619
2013 CGUGGUUC A CUUUGGUC 845 GACCAAAG CUGAUGAG X CGAA IAACCACG 2620
2015 UGGUUCAC U UUGGUCAC 846 GUGACCAA CUGAUGAG X CGAA IUGAACCA 2621
2022 CUUUGGUC A CAAGUAGG 847 CCUACUUG CUGAUGAG X CGAA IACCAAAG 2622
2024 UUGGUCAC A AGUAGGAG 848 CUCCUACU CUGAUGAG X CGAA lUGACCAA 2623
2035 UAGGAGAC A CAGAUGGC 849 GCCAUCUG CUGAUGAG X CGAA IUCUCCUA 2624
2037 GGAGACAC A GAUGGCAC 850 GUGCCAUC CUGAUGAG X CGAA IUGUCUCC 2625
2044 CAGAUGGC A CCUGUGGC 851 GCCACAGG CUGAUGAG X CGAA ICCAUCUG 2626
2046 GAUGGCAC C UGUGGCCA 852 UGGCCACA CUGAUGAG X CGAA IUGCCAUC 2627
2047 AUGGCACC U GUGGCCAG 853 CUGGCCAC CUGAUGAG X CGAA IGUGCCAU 2628
2053 CCUGUGGC C AGAGCACC 854 GGUGCUCU CUGAUGAG X CGAA ICCACAGG 2629
2054 CUGUGGCC A GAGCACCU 855 AGGUGCUC CUGAUGAG X CGAA IGCCACAG 2630
2059 GCCAGAGC A CCUCAGGA 856 UCCUGAGG CUGAUGAG X CGAA ICUCUGGC 2631
2061 CAGAGCAC C UCAGGACC 857 GGUCCUGA CUGAUGAG X CGAA lUGCUCUG 2632
2062 AGAGCACC U CAGGACCC 858 GGGUCCUG CUGAUGAG X CGAA IGUGCUCU 2633
2064 AGCACCUC A GGACCCUC 859 GAGGGUCC CUGAUGAG X CGAA IAGGUGCU 2634
2069 CUCAGGAC C CUCCCCAC 860 GUGGGGAG CUGAUGAG X CGAA IUCCUGAG 2635
2070 UCAGGACC C UCCCCACC 861 GGUGGGGA CUGAUGAG X CGAA IGUCCUGA 2636
2071 CAGGACCC U CCCCACCC 862 GGGUGGGG CUGAUGAG X CGAA IGGUCCUG 2637
2073 GGACCCUC c CCACCCAC 863 GUGGGUGG CUGAUGAG X CGAA IAGGGUCC 2638
2074 GACCCUCC c CACCCACC 864 GGUGGGUG CUGAUGAG X CGAA IGAGGGUC 2639
2075 ACCCUCCC c ACCCACCA 865 UGGUGGGU CUGAUGAG X CGAA IGGAGGGU 2640
2076 CCCUCCCC A CCCACCAA 866 UUGGUGGG CUGAUGAG X CGAA IGGGAGGG 2641
2078 CUCCCCAC C CACCAAAU 867 AUUUGGUG CUGAUGAG X CGAA IUGGGGAG 2642
2079 UCCCCACC C ACCAAAUG 868 CAUUUGGU CUGAUGAG X CGAA IGUGGGGA 2643
2080 CCCCACCC A CCAAAUGC 869 GCAUUUGG CUGAUGAG X CGAA IGGUGGGG 2644
2082 CCACCCAC C AAAUGCCU 870 AGGCAUUU CUGAUGAG X CGAA IUGGGUGG 2645
2083 CACCCACC A AAUGCCUC 871 GAGGCAUU CUGAUGAG X CGAA IGUGGGUG 2646
2089 CCAAAUGC C UCUGCCUU 872 AAGGCAGA CUGAUGAG X CGAA ICAUUUGG 2647
2090 CAAAUGCC U CUGCCUUG 873 CAAGGCAG CUGAUGAG X CGAA IGCAUUUG 2648
2092 AAUGCCUC U GCCUUGAU 874 AUCAAGGC CUGAUGAG X CGAA IAGGCAUU 2649
2095 GCCUCUGC C UUGAUGGA 875 UCCAUCAA CUGAUGAG X CGAA ICAGAGGC 2650
2096 CCUCUGCC U UGAUGGAG 876 CUCCAUCA CUGAUGAG X CGAA IGCAGAGG 2651
2116 GAAAAGGC U GGCAAGGU 877 ACCUUGCC CUGAUGAG X CGAA ICCUUUUC 2652
2120 AGGCUGGC A AGGUGGGU 878 ACCCACCU CUGAUGAG X CGAA ICCAGCCU 2653
2131 GUGGGUUC C AGGGACUG 879 CAGUCCCU CUGAUGAG X CGAA IAACCCAC 2654
2132 UGGGUUCC A GGGACUGU 880 ACAGUCCC CUGAUGAG X CGAA IGAACCCA 2655
2138 CCAGGGAC U GUACCUGU 881 ACAGGUAC CUGAUGAG X CGAA lUCCCUGG 2656
2143 GACUGUAC C UGUAGGAA 882 UUCCUACA CUGAUGAG X CGAA IUACAGUC 2657
2144 ACUGUACC U GUAGGAAA 883 UUUCCUAC CUGAUGAG X CGAA IGUACAGU 2658
2154 UAGGAAAC A GAAAAGAG 884 CUCUUUUC CUGAUGAG X CGAA IUUUCCUA 2659
2174 AAAGAAGC A CUCUGCUG 885 CAGCAGAG CUGAUGAG X CGAA ICUUCUUU 2660
2176 AGAAGCAC U CUGCUGGC 886 GCCAGCAG CUGAUGAG X CGAA IUGCUUCU 2661
2178 AAGCACUC u GCUGGCGG 887 CCGCCAGC CUGAUGAG X CGAA IAGUGCUU 2662
2181 CACUCUGC u GGCGGGAA 888 UUCCCGCC CUGAUGAG X CGAA ICAGAGUG 2663
2193 GGGAAUAC u CUUGGUCA 889 UGACCAAG CUGAUGAG X CGAA IUAUUCCC 2664
2195 GAAUACUC u UGGUCACC 890 GGUGACCA CUGAUGAG X CGAA IAGUAUUC 2665
2201 UCUUGGUC A CCUCAAAU 891 AUUUGAGG CUGAUGAG X CGAA lACCAAGA 2666
2203 UUGGUCAC C UCAAAUUU 892 AAAUUUGA CUGAUGAG X CGAA lUGACCAA 2667
Table 19
2204 UGGUCACC U CAAAUUUA 893 UAAAUUUG CUGAUGAG X CGAA IGUGACCA 2668
2206 GUCACCUC A AAUUUAAG 894 CUUAAAUU CUGAUGAG X CGAA IAGGUGAC 2669
2226 GGAAAUUC U GCUGCUUG 895 CAAGCAGC CUGAUGAG X CGAA IAAUUUCC 2670
2229 AAUUCUGC U GCUUGAAA 896 UUUCAAGC CUGAUGAG X CGAA ICAGAAUU 2671
2232 UCUGCUGC u UGAAACUU 897 AAGUUUCA CUGAUGAG X CGAA ICAGCAGA 2672
2239 CUUGAAAC u UCAGCCCU 898 AGGGCUGA CUGAUGAG X CGAA IUUUCAAG 2673
2242 GAAACUUC A GCCCUGAA 899 UUCAGGGC CUGAUGAG X CGAA IAAGUUUC 2674
2245 ACUUCAGC C CUGAACCU 900 AGGUUCAG CUGAUGAG X CGAA ICUGAAGU 2675
2246 CUUCAGCC C UGAACCUU 901 AAGGUUCA CUGAUGAG X CGAA IGCUGAAG 2676
2247 UUCAGCCC u GAACCUUU 902 AAAGGUUC CUGAUGAG X CGAA IGGCUGAA 2677
2252 CCCUGAAC C UUUGUCCA 903 UGGACAAA CUGAUGAG X CGAA IUUCAGGG 2678
2253 CCUGAACC u UUGUCCAC 904 GUGGACAA CUGAUGAG X CGAA IGUUCAGG 2679
2259 CCUUUGUC c ACCAUUCC 905 GGAAUGGU CUGAUGAG X CGAA IACAAAGG 2680
2260 CUUUGUCC A CCAUUCCU 906 AGGAAUGG CUGAUGAG X CGAA IGACAAAG 2681
2262 UUGUCCAC C AUUCCUUU 907 AAAGGAAU CUGAUGAG X CGAA IUGGACAA 2682
2263 UGUCCACC A UUCCUUUA 908 UAAAGGAA CUGAUGAG X CGAA IGUGGACA 2683
2267 CACCAUUC C UUUAAAUU 909 AAUUUAAA CUGAUGAG X CGAA IAAUGGUG 2684
2268 ACCAUUCC u UUAAAUUC 910 GAAUUUAA CUGAUGAG X CGAA IGAAUGGU 2685
2277 UUAAAUUC u CCAACCCA 911 UGGGUUGG CUGAUGAG X CGAA IAAUUUAA 2686
2279 AAAUUCUC c AACCCAAA 912 UUUGGGUU CUGAUGAG X CGAA IAGAAUUU 2687
2280 AAUUCUCC A ACCCAAAG 913 CUUUGGGU CUGAUGAG X CGAA IGAGAAUU 2688
2283 UCUCCAAC C CAAAGUAU 914 AUACUUUG CUGAUGAG X CGAA IUUGGAGA 2689
2284 CUCCAACC C AAAGUAUU 915 AAUACUUU CUGAUGAG X CGAA IGUUGGAG 2690
2285 UCCAACCC A AAGUAUUC 916 GAAUACUU CUGAUGAG X CGAA IGGUUGGA 2691
2294 AAGUAUUC U UCUUUUCU 917 AGAAAAGA CUGAUGAG X CGAA IAAUACUU 2692
2297 UAUUCUUC U UUUCUUAG 918 CUAAGAAA CUGAUGAG X CGAA IAAGAAUA 2693
2302 UUCUUUUC U UAGUUUCA 919 UGAAACUA CUGAUGAG X CGAA IAAAAGAA 2694
2310 UUAGUUUC A GAAGUACU 920 AGUACUUC CUGAUGAG X CGAA IAAACUAA 2695
2318 AGAAGUAC U GGCAUCAC 921 GUGAUGCC CUGAUGAG X CGAA IUACUUCU 2696
2322 GUACUGGC A UCACACGC 922 GCGUGUGA CUGAUGAG X CGAA ICCAGUAC 2697
2325 CUGGCAUC A CACGCAGG 923 CCUGCGUG CUGAUGAG X CGAA IAUGCCAG 2698
2327 GGCAUCAC A CGCAGGUU 924 AACCUGCG CUGAUGAG X CGAA IUGAUGCC 2699
2331 UCACACGC A GGUUACCU 925 AGGUAACC CUGAUGAG X CGAA ICGUGUGA 2700
2338 CAGGUUAC C UUGGCGUG 926 CACGCCAA CUGAUGAG X CGAA IUAACCUG 2701
2339 AGGUUACC U UGGCGUGU 927 ACACGCCA CUGAUGAG X CGAA IGUAACCU 2702
2351 CGUGUGUC c CUGUGGUA 928 UACCACAG CUGAUGAG X CGAA IACACACG 2703
2352 GUGUGUCC c UGUGGUAC 929 GUACCACA CUGAUGAG X CGAA IGACACAC 2704
2353 UGUGUCCC u GUGGUACC 930 GGUACCAC CUGAUGAG X CGAA IGGACACA 2705
2361 UGUGGUAC c CUGGCAGA 931 UCUGCCAG CUGAUGAG X CGAA IUACCACA 2706
2362 GUGGUACC c UGGCAGAG 932 CUCUGCCA CUGAUGAG X CGAA IGUACCAC 2707
2363 UGGUACCC u GGCAGAGA 933 UCUCUGCC CUGAUGAG X CGAA IGGUACCA 2708
2367 ACCCUGGC A GAGAAGAG 934 CUCUUCUC CUGAUGAG X CGAA ICCAGGGU 2709
2378 GAAGAGAC C AAGCUUGU 935 ACAAGCUU CUGAUGAG X CGAA IUCUCUUC 2710
2379 AAGAGACC A AGCUUGUU 936 AACAAGCU CUGAUGAG X CGAA IGUCUCUU 2711
2383 GACCAAGC U UGUUUCCC 937 GGGAAACA CUGAUGAG X CGAA ICUUGGUC 2712
2390 CUUGUUUC C CUGCUGGC 938 GCCAGCAG CUGAUGAG X CGAA IAAACAAG 2713
2391 UUGUUUCC C UGCUGGCC 939 GGCCAGCA CUGAUGAG X CGAA IGAAACAA 2714
2392 UGUUUCCC u GCUGGCCA 940 UGGCCAGC CUGAUGAG X CGAA IGGAAACA 2715
2395 UUCCCUGC u GGCCAAAG 941 CUUUGGCC CUGAUGAG X CGAA ICAGGGAA 2716
2399 CUGCUGGC c AAAGUCAG 942 CUGACUUU CUGAUGAG X CGAA ICCAGCAG 2717
2400 UGCUGGCC A AAGUCAGU 943 ACUGACUU CUGAUGAG X CGAA IGCCAGCA 2718
Table 19
2406 CCAAAGUC A GUAGGAGA 944 UCUCCUAC CUGAUGAG X CGAA IACUUUGG 2719
2421 GAGGAUGC A CAGUUUGC 945 GCAAACUG CUGAUGAG X CGAA ICAUCCUC 2720
2423 GGAUGCAC A GUUUGCUA 946 UAGCAAAC CUGAUGAG X CGAA IUGCAUCC 2721
2430 CAGUUUGC U AUUUGCUU 947 AAGCAAAU CUGAUGAG X CGAA ICAAACUG 2722
2437 CUAUUUGC U UUAGAGAC 948 GUCUCUAA CUGAUGAG X CGAA ICAAAUAG 2723
2446 UUAGAGAC A GGGACUGU 949 ACAGUCCC CUGAUGAG X CGAA IUCUCUAA 2724
2452 ACAGGGAC U GUAUAAAC 950 GUUUAUAC CUGAUGAG X CGAA IUCCCUGU 2725
2461 GUAUAAAC A AGCCUAAC 951 GUUAGGCU CUGAUGAG X CGAA IUUUAUAC 2726
2465 AAACAAGC C UAACAUUG 952 CAAUGUUA CUGAUGAG X CGAA ICUUGUUU 2727
2466 AACAAGCC U AACAUUGG 953 CCAAUGUU CUGAUGAG X CGAA IGCUUGUU 2728
2470 AGCCUAAC A UUGGUGCA 954 UGCACCAA CUGAUGAG X CGAA lUUAGGCU 2729
2478 AUUGGUGC A AAGAUUGC 955 GCAAUCUU CUGAUGAG X CGAA ICACCAAU 2730
2487 AAGAUUGC C UCUUGAAU 956 AUUCAAGA CUGAUGAG X CGAA ICAAUCUU 2731
2488 AGAUUGCC U CUUGAAUU 957 AAUUCAAG CUGAUGAG X CGAA IGCAAUCU 2732
2490 AUUGCCUC u UGAAUUAA 958 UUAAUUCA CUGAUGAG X CGAA IAGGCAAU 2733
2509 AAAAAAAC u AGAAAAAA 959 UUUUUUCU CUGAUGAG X CGAA IUUUUUUU 2734
Input Sequence = AF190725. Cut Site = G/ .
Stem Length = 8 . Core Sequence = CUGAUGAG X CGAA (X = GCCGUUAGGC or other stem II)
AF190725 (Homo sapiens beta-site APP cleaving enzyme (BACE) mRNA; 2526 bp)
Table 20
Table 20: Human BACE G-cleaver Ribozyme and Target Sequence
Pos Substrate Seq ID Ribozyme Rz Seq
ID
11 ACGCGUCC G CAGCCCGC 960 GCGGGCUG UGAUG GCAUGCACUAUGC GCG GGACGCGU 2735
18 CGCAGCCC G CCCGGGAG 961 CUCCCGGG UGAUG GCAUGCACUAUGC GCG GGGCUGCG 2736
29 CGGGAGCU G CGAGCCGC 962 GCGGCUCG UGAUG GCAUGCACUAUGC GCG AGCUCCCG 2737
31 GGAGCUGC G AGCCGCGA 963 UCGCGGCU UGAUG GCAUGCACUAUGC GCG GCAGCUCC 2738
36 UGCGAGCC G CGAGCUGG 964 CCAGCUCG UGAUG GCAUGCACUAUGC GCG GGCUCGCA 2739
38 CGAGCCGC G AGCUGGAU 965 AUCCAGCU UGAUG GCAUGCACUAUGC GCG GCGGCUCG 2740
58 GGUGGCCU G AGCAGCCA 966 UGGCUGCU UGAUG GCAUGCACUAUGC GCG AGGCCACC 2741
69 CAGCCAAC G CAGCCGCA 967 UGCGGCUG UGAUG GCAUGCACUAUGC GCG GUUGGCUG 2742
75 ACGCAGCC G CAGGAGCC 968 GGCUCCUG UGAUG GCAUGCACUAUGC GCG GGCUGCGU 2743
94 GAGCCCUU G CCCCUGCC 969 GGCAGGGG UGAUG GCAUGCACUAUGC GCG AAGGGCUC 2744
100 UUGCCCCU G CCCGCGCC 970 GGCGCGGG UGAUG GCAUGCACUAUGC GCG AGGGGCAA 2745
104 CCCUGCCC G CGCCGCCG 971 CGGCGGCG UGAUG GCAUGCACUAUGC GCG GGGCAGGG 2746
106 CUGCCCGC G CCGCCGCC 972 GGCGGCGG UGAUG GCAUGCACUAUGC GCG GCGGGCAG 2747
109 CCCGCGCC G CCGCCCGC 973 GCGGGCGG UGAUG GCAUGCACUAUGC GCG GGCGCGGG 2748
112 GCGCCGCC G CCCGCCGG 974 CCGGCGGG UGAUG GCAUGCACUAUGC GCG GGCGGCGC 2749
116 CGCCGCCC G CCGGGGGG 975 CCCCCCGG UGAUG GCAUGCACUAUGC GCG GGGCGGCG 2750
137 GGGAAGCC G CCACCGGC 976 GCCGGUGG UGAUG GCAUGCACUAUGC GCG GGCUUCCC 2751
148 ACCGGCCC G CCAUGCCC 977 GGGCAUGG UGAUG GCAUGCACUAUGC GCG GGGCCGGU 2752
153 CCCGCCAU G CCCGCCCC 978 GGGGCGGG UGAUG GCAUGCACUAUGC GCG AUGGCGGG 2753
157 CCAUGCCC G CCCCUCCC 979 GGGAGGGG UGAUG GCAUGCACUAUGC GCG GGGCAUGG 2754
172 CCAGCCCC G CCGGGAGC 980 GCUCCCGG UGAUG GCAUGCACUAUGC GCG GGGGCUGG 2755
183 GGGAGCCC G CGCCCGCU 981 AGCGGGCG UGAUG GCAUGCACUAUGC GCG GGGCUCCC 2756
185 GAGCCCGC G CCCGCUGC 982 GCAGCGGG UGAUG GCAUGCACUAUGC GCG GCGGGCUC 2757
189 CCGCGCCC G CUGCCCAG 983 CUGGGCAG UGAUG GCAUGCACUAUGC GCG GGGCGCGG 2758
192 CGCCCGCU G CCCAGGCU 984 AGCCUGGG UGAUG GCAUGCACUAUGC GCG AGCGGGCG 2759
205 GGCUGGCC G CCGCCGUG 985 CACGGCGG UGAUG GCAUGCACUAUGC GCG GGCCAGCC 2760
208 UGGCCGCC G CCGUGCCG 986 CGGCACGG UGAUG GCAUGCACUAUGC GCG GGCGGCCA 2761
213 GCCGCCGU G CCGAUGUA 987 UACAUCGG UGAUG GCAUGCACUAUGC GCG ACGGCGGC 2762
216 GCCGUGCC G AUGUAGCG 988 CGCUACAU UGAUG GCAUGCACUAUGC GCG GGCACGGC 2763
250 UCUCCCCU G CUCCCGUG 989 CACGGGAG UGAUG GCAUGCACUAUGC GCG AGGGGAGA 2764
Table 20
258 GCUCCCGU G CUCUGCGG 990 CCGCAGAG UGAUG GCAUGCACUAUGC GCG ACGGGAGC 2765
263 CGUGCUCU G CGGAUCUC 991 GAGAUCCG UGAUG GCAUGCACUAUGC GCG AGAGCACG 2766
276 UCUCCCCU G ACCGCUCU 992 AGAGCGGU UGAUG GCAUGCACUAUGC GςG AGGGGAGA 2767
280 CCCUGACC G CUCUCCAC 993 GUGGAGAG UGAUG GCAUGCACUAUGC GCG GGUCAGGG 2768
320 AGGGCCCU G CAGGCCCU 994 AGGGCCUG UGAUG GCAUGCACUAUGC GCG AGGGCCCU 2769
337 GGCGUCCU G AUGCCCCC 995 GGGGGCAU UGAUG GCAUGCACUAUGC GCG AGGACGCC 2770
340 GUCCUGAU G CCCCCAAG 996 CUUGGGGG UGAUG GCAUGCACUAUGC GCG AUCAGGAC 2771
360 CCUCUCCU G AGAAGCCA 997 UGGCUUCU UGAUG GCAUGCACUAUGC GCG AGGAGAGG 2772
397 GGGCAGGC G CCAGGGAC 998 GUCCCUGG UGAUG GCAUGCACUAUGC GCG GCCUGCCC 2773
420 GGGCCAGU G CGAGCCCA 999 UGGGCUCG UGAUG GCAUGCACUAUGC GCG ACUGGCCC 2774
422 GCCAGUGC G AGCCCAGA 1000 UCUGGGCU UGAUG GCAUGCACUAUGC GCG GCACUGGC 2775
437 GAGGGCCC G AAGGCCGG 1001 CCGGCCUU UGAUG GCAUGCACUAUGC GCG GGGCCCUC 2776
468 CAAGCCCU G CCCUGGCU 1002 AGCCAGGG UGAUG GCAUGCACUAUGC GCG AGGGCUUG 2777
480 UGGCUCCU G CUGUGGAU 1003 AUCCACAG UGAUG GCAUGCACUAUGC GCG AGGAGCCA 2778
493 GGAUGGGC G CGGGAGUG 1004 CACUCCCG UGAUG GCAUGCACUAUGC GCG GCCCAUCC 2779
501 GCGGGAGU G CUGCCUGC 1005 GCAGGCAG UGAUG GCAUGCACUAUGC GCG ACUCCCGC 2780
504 GGAGUGCU G CCUGCCCA 1006 UGGGCAGG UGAUG GCAUGCACUAUGC GCG AGCACUCC 2781
508 UGCUGCCU G CCCACGGC 1007 GCCGUGGG UGAUG GCAUGCACUAUGC GCG AGGCAGCA 2782
537 AUCCGGCU G CCCCUGCG 1008 CGCAGGGG UGAUG GCAUGCACUAUGC GCG AGCCGGAU 2783
543 CUGCCCCU G CGCAGCGG 1009 CCGCUGCG UGAUG GCAUGCACUAUGC GCG AGGGGCAG 2784
545 GCCCCUGC G CAGCGGCC 1010 GGCCGCUG UGAUG GCAUGCACUAUGC GCG GCAGGGGC 2785
562 UGGGGGGC G CCCCCCUG 1011 CAGGGGGG UGAUG GCAUGCACUAUGC GCG GCCCCCCA 2786
576 CUGGGGCU G CGGCUGCC 1012 GGCAGCCG UGAUG GCAUGCACUAUGC GCG AGCCCCAG 2787
582 CUGCGGCU G CCCCGGGA 1013 UCCCGGGG UGAUG GCAUGCACUAUGC GCG AGCCGCAG 2788
595 GGGAGACC G ACGAAGAG 1014 CUCUUCGU UGAUG GCAUGCACUAUGC GCG GGUCUCCC 2789
598 AGACCGAC G AAGAGCCC 1015 GGGCUCUU UGAUG GCAUGCACUAUGC GCG GUCGGUCU 2790
607 AAGAGCCC G AGGAGCCC 1016 GGGCUCCU UGAUG GCAUGCACUAUGC GCG GGGCUCUU 2791
654 GACAACCU G AGGGGCAA 1017 UUGCCCCU UGAUG GCAUGCACUAUGC GCG AGGUUGUC 2792
690 GUGGAGAU G ACCGUGGG 1018 CCCACGGU UGAUG GCAUGCACUAUGC GCG AUCUCCAC 2793
708 AGCCCCCC G CAGACGCU 1019 AGCGUCUG UGAUG GCAUGCACUAUGC GCG GGGGGGCU 2794
714 CCGCAGAC G CUCAACAU 1020 AUGUUGAG UGAUG GCAUGCACUAUGC GCG GUCUGCGG 2795
751 GUAACUUU G CAGUGGGU 1021 ACCCACUG UGAUG GCAUGCACUAUGC GCG AAAGUUAC 2796
760 CAGUGGGU G CUGCCCCC 1022 GGGGGCAG UGAUG GCAUGCACUAUGC GCG ACCCACUG 2797
763 UGGGUGCU G CCCCCCAC 1023 GUGGGGGG UGAUG GCAUGCACUAUGC GCG AGCACCCA 2798
Table 20
780 CCCUUCCU G CAUCGCUA 1024 UAGCGAUG UGAUG GCAUGCACUAUGC GCG AGGAAGGG 2799
785 CCUGCAUC G CUACUACC 1025 GGUAGUAG UGAUG GCAUGCACUAUGC GCG GAUGCAGG 2800
843 GUGUAUGU G CCCUACAC 1026 GUGUAGGG UGAUG GCAUGCACUAUGC GCG ACAUACAC 2801
883 UGGGCACC G ACCUGGUA 1027 UACCAGGU UGAUG GCAUGCACUAUGC GCG GGUGCCCA 2802
921 GUCACUGU G CGUGCCAA 1028 UUGGCACG UGAUG GCAUGCACUAUGC GCG ACAGUGAC 2803
925 CUGUGCGU G CCAACAUU 1029 AAUGUUGG UGAUG GCAUGCACUAUGC GCG ACGCACAG 2804
934 CCAACAUU G CUGCCAUC 1030 GAUGGCAG UGAUG GCAUGCACUAUGC GCG AAUGUUGG 2805
937 ACAUUGCU G CCAUCACU 1031 AGUGAUGG UGAUG GCAUGCACUAUGC GCG AGCAAUGU 2806
946 CCAUCACU G AAUCAGAC 1032 GUCUGAUU UGAUG GCAUGCACUAUGC GCG AGUGAUGG 2807
1006 UGGCCUAU G CUGAGAUU 1033 AAUCUCAG UGAUG GCAUGCACUAUGC GCG AUAGGCCA 2808
1009 CCUAUGCU G AGAUUGCC 1034 GGCAAUCU UGAUG GCAUGCACUAUGC GCG AGCAUAGG 2809
1015 CUGAGAUU G CCAGGCCU 1035 AGGCCUGG UGAUG GCAUGCACUAUGC GCG AAUCUCAG 2810
1024 CCAGGCCU G ACGACUCC 1036 GGAGUCGU UGAUG GCAUGCACUAUGC GCG AGGCCUGG 2811
1027 GGCCUGAC G ACUCCCUG 1037 CAGGGAGU UGAUG GCAUGCACUAUGC GCG GUCAGGCC 2812
1048 CUUUCUUU G ACUCUCUG 1038 CAGAGAGU UGAUG GCAUGCACUAUGC GCG AAAGAAAG 2813
1092 UUCUCCCU G CAGCUUUG 1039 CAAAGCUG UGAUG GCAUGCACUAUGC GCG AGGGAGAA 2814
1105 UUUGUGGU G CUGGCUUC 1040 GAAGCCAG UGAUG GCAUGCACUAUGC GCG ACCACAAA 2815
1129 ACCAGUCU G AAGUGCUG 1041 CAGCACUU UGAUG GCAUGCACUAUGC GCG AGACUGGU 2816
1134 UCUGAAGU G CUGGCCUC 1042 GAGGCCAG UGAUG GCAUGCACUAUGC GCG ACUUCAGA 2817
1158 GGGAGCAU G AUCAUUGG 1043 CCAAUGAU UGAUG GCAUGCACUAUGC GCG AUGCUCCC 2818
1174 GAGGUAUC G ACCACUCG 1044 CGAGUGGU UGAUG GCAUGCACUAUGC GCG GAUACCUC 2819
1182 GACCACUC G CUGUACAC 1045 GUGUACAG UGAUG GCAUGCACUAUGC GCG GAGUGGUC 2820
1234 GGUAUUAU G AGGUGAUC 1046 GAUCACCU UGAUG GCAUGCACUAUGC GCG AUAAUACC 2821
1239 UAUGAGGU G AUCAUUGU 1047 ACAAUGAU UGAUG GCAUGCACUAUGC GCG ACCUCAUA 2822
1248 AUCAUUGU G CGGGUGGA 1048 UCCACCCG UGAUG GCAUGCACUAUGC GCG ACAAUGAU 2823
1275 CAGGAUCU G AAAAUGGA 1049 UCCAUUUU UGAUG GCAUGCACUAUGC GCG AGAUCCUG 2824
1286 AAUGGACU G CAAGGAGU 1050 ACUCCUUG UGAUG GCAUGCACUAUGC GCG AGUCCAUU 2825
1303 ACAACUAU G ACAAGAGC 1051 GCUCUUGU UGAUG GCAUGCACUAUGC GCG AUAGUUGU 2826
1344 CUUCGUUU G CCCAAGAA 1052 UUCUUGGG UGAUG GCAUGCACUAUGC GCG AAACGAAG 2827
1360 AAGUGUUU G AAGCUGCA 1053 UGCAGCUU UGAUG GCAUGCACUAUGC GCG AAACACUU 2828
1366 UUGAAGCU G CAGUCAAA 1054 UUUGACUG UGAUG GCAUGCACUAUGC GCG AGCUUCAA 2829
1411 AGUUCCCU G AUGGUUUC 1055 GAAACCAU UGAUG GCAUGCACUAUGC GCG AGGGAACU 2830
1442 GCUGGUGU G CUGGCAAG 1056 CUUGCCAG UGAUG GCAUGCACUAUGC GCG ACACCAGC 2831
1504 UAAUGGGU G AGGUUACC 1057 GGUAACCU UGAUG GCAUGCACUAUGC GCG ACCCAUUA 2832
Table 20
1526 GUCCUUCC G CAUCACCA 1058 UGGUGAUG UGAUG GCAUGCACUAUGC GCG GGAAGGAC 2833
1542 AUCCUUCC G CAGCAAUA 1059 UAUUGCUG UGAUG GCAUGCACUAUGC GCG GGAAGGAU 2834
1554 CAAUACCU G CGGCCAGU 1060 ACUGGCCG UGAUG GCAUGCACUAUGC GCG AGGUAUUG 2835
1588 CCCAAGAC G ACUGUUAC 1061 GUAACAGU UGAUG GCAUGCACUAUGC GCG GUCUUGGG 2836
1603 ACAAGUUU G CCAUCUCA 1062 UGAGAUGG UGAUG GCAUGCACUAUGC GCG AAACUUGU 2837
1672 UUGUCUUU G AUCGGGCC 1063 GGCCCGAU UGAUG GCAUGCACUAUGC GCG AAAGACAA 2838
1682 UCGGGCCC G AAAACGAA 1064 UUCGUUUU UGAUG GCAUGCACUAUGC GCG GGGCCCGA 2839
1688 CCGAAAAC G AAUUGGCU 1065 AGCCAAUU UGAUG GCAUGCACUAUGC GCG GUUUUCGG 2840
1699 UUGGCUUU G CUGUCAGC 1066 GCUGACAG UGAUG GCAUGCACUAUGC GCG AAAGCCAA 2841
1708 CUGUCAGC G CUUGCCAU 1067 AUGGCAAG UGAUG GCAUGCACUAUGC GCG GCUGACAG 2842
1712 CAGCGCUU G CCAUGUGC 1068 GCACAUGG UGAUG GCAUGCACUAUGC GCG AAGCGCUG 2843
1719 UGCCAUGU G CACGAUGA 1069 UCAUCGUG UGAUG GCAUGCACUAUGC GCG ACAUGGCA 2844
1723 AUGUGCAC G AUGAGUUC 1070 GAACUCAU UGAUG GCAUGCACUAUGC GCG GUGCACAU 2845
1726 UGCACGAU G AGUUCAGG 1071 CCUGAACU UGAUG GCAUGCACUAUGC GCG AUCGUGCA 2846
1807 AGACAGAU G AGUCAACC 1072 GGUUGACU UGAUG GCAUGCACUAUGC GCG AUCUGUCU 2847
1821 ACCCUCAU G ACCAUAGC 1073 GCUAUGGU UGAUG GCAUGCACUAUGC GCG AUGAGGGU 2848
1843 UCAUGGCU G CCAUCUGC 1074 GCAGAUGG UGAUG GCAUGCACUAUGC GCG AGCCAUGA 2849
1850 UGCCAUCU G CGCCCUCU 1075 AGAGGGCG UGAUG GCAUGCACUAUGC GCG AGAUGGCA 2850
1852 CCAUCUGC G CCCUCUUC 1076 GAAGAGGG UGAUG GCAUGCACUAUGC GCG GCAGAUGG 2851
1863 CUCUUCAU G CUGCCACU 1077 AGUGGCAG UGAUG GCAUGCACUAUGC GCG AUGAAGAG 2852
1866 UUCAUGCU G CCACUCUG 1078 CAGAGUGG UGAUG GCAUGCACUAUGC GCG AGCAUGAA 2853
1874 GCCACUCU G CCUCAUGG 1079 CCAUGAGG UGAUG GCAUGCACUAUGC GCG AGAGUGGC 2854
1895 UCAGUGGC G CUGCCUCC 1080 GGAGGCAG UGAUG GCAUGCACUAUGC GCG GCCACUGA 2855
1898 GUGGCGCU G CCUCCGCU 1081 AGCGGAGG UGAUG GCAUGCACUAUGC GCG AGCGCCAC 2856
1904 CUGCCUCC G CUGCCUGC 1082 GCAGGCAG UGAUG GCAUGCACUAUGC GCG GGAGGCAG 2857
1907 CCUCCGCU G CCUGCGCC 1083 GGCGCAGG UGAUG GCAUGCACUAUGC GCG AGCGGAGG 2858
1911 CGCUGCCU G CGCCAGCA 1084 UGCUGGCG UGAUG GCAUGCACUAUGC GCG AGGCAGCG 2859
1913 CUGCCUGC G CCAGCAGC 1085 GCUGCUGG UGAUG GCAUGCACUAUGC GCG GCAGGCAG 2860
1924 AGCAGCAU G AUGACUUU 1086 AAAGUCAU UGAUG GCAUGCACUAUGC GCG AUGCUGCU 2861
1927 AGCAUGAU G ACUUUGCU 1087 AGCAAAGU UGAUG GCAUGCACUAUGC GCG AUCAUGCU 2862
1933 AUGACUUU G CUGAUGAC 1088 GUCAUCAG UGAUG GCAUGCACUAUGC GCG AAAGUCAU 2863
1936 ACUUUGCU G AUGACAUC 1089 GAUGUCAU UGAUG GCAUGCACUAUGC GCG AGCAAAGU 2864
1939 UUGCUGAU G ACAUCUCC 1090 GGAGAUGU UGAUG GCAUGCACUAUGC GCG AUCAGCAA 2865
1950 AUCUCCCU G CUGAAGUG 1091 CACUUCAG UGAUG GCAUGCACUAUGC GCG AGGGAGAU 2866
Table 20
1953 UCCCUGCU G AAGUGAGG 1092 CCUCACUU UGAUG GCAUGCACUAUGC GCG AGCAGGGA 2867
1958 GCUGAAGU G AGGAGGCC 1093 GGCCUCCU UGAUG GCAUGCACUAUGC GCG ACUUCAGC 2868
2087 CACCAAAU G CCUCUGCC 1094 GGCAGAGG UGAUG GCAUGCACUAUGC GCG AUUUGGUG 2869
2093 AUGCCUCU G CCUUGAUG 1095 CAUCAAGG UGAUG GCAUGCACUAUGC GCG AGAGGCAU 2870
2098 UCUGCCUU G AUGGAGAA 1096 UUCUCCAU UGAUG GCAUGCACUAUGC GCG AAGGCAGA 2871
2179 AGCACUCU G CUGGCGGG 1097 CCCGCCAG UGAUG GCAUGCACUAUGC GCG AGAGUGCU 2872
2227 GAAAUUCU G CUGCUUGA 1098 UCAAGCAG UGAUG GCAUGCACUAUGC GCG AGAAUUUC 2873
2230 AUUCUGCU G CUUGAAAC 1099 GUUUCAAG UGAUG GCAUGCACUAUGC GCG AGCAGAAU 2874
2234 UGCUGCUU G AAACUUCA 1100 UGAAGUUU UGAUG GCAUGCACUAUGC GCG AAGCAGCA 2875
2248 UCAGCCCU G AACCUUUG 1101 CAAAGGUU UGAUG GCAUGCACUAUGC GCG AGGGCUGA 2876
2329 CAUCACAC G CAGGUUAC 1102 GUAACCUG UGAUG GCAUGCACUAUGC GCG GUGUGAUG 2877
2393 GUUUCCCU G CUGGCCAA 1103 UUGGCCAG UGAUG GCAUGCACUAUGC GCG AGGGAAAC 2878
2419 GAGAGGAU G CACAGUUU 1104 AAACUGUG UGAUG GCAUGCACUAUGC GCG AUCCUCUC 2879
2428 CACAGUUU G CUAUUUGC 1105 GCAAAUAG UGAUG GCAUGCACUAUGC GCG AAACUGUG 2880
2435 UGCUAUUU G CUUUAGAG 1106 CUCUAAAG UGAUG GCAUGCACUAUGC GCG AAAUAGCA 2881
2476 ACAUUGGU G CAAAGAUU 1107 AAUCUUUG UGAUG GCAUGCACUAUGC GCG ACCAAUGU 2882
2485 CAAAGAUU G CCUCUUGA 1108 UCAAGAGG UGAUG GCAUGCACUAUGC GCG AAUCUUUG 2883
2492 UGCCUCUU G AAUUAAAA 1109 UUUUAAUU UGAUG GCAUGCACUAUGC GCG AAGAGGCA 2884
219 GUGCCGAU G UAGCGGGC 1110 GCCCGCUA UGAUG GCAUGCACUAUGC GCG AUCGGCAC 2885
483 CUCCUGCU G UGGAUGGG llll CCCAUCCA UGAUG GCAUGCACUAUGC GCG AGCAGGAG 2886
634 GCAGCUUU G UGGAGAUG 1112 CAUCUCCA UGAUG GCAUGCACUAUGC GCG AAAGCUGC 2887
804 AGGCAGCU G UCCAGCAC 1113 GUGCUGGA UGAUG GCAUGCACUAUGC GCG AGCUGCCU 2888
835 GGAAGGGU G UGUAUGUG 1114 CACAUACA UGAUG GCAUGCACUAUGC GCG ACCCUUCC 2889
837 AAGGGUGU G UAUGUGCC 1115 GGCACAUA UGAUG GCAUGCACUAUGC GCG ACACCCUU 2890
841 GUGUGUAU G UGCCCUAC 1116 GUAGGGCA UGAUG GCAUGCACUAUGC GCG AUACACAC 2891
919 ACGUCACU G UGCGUGCC 1117 GGCACGCA UGAUG GCAUGCACUAUGC GCG AGUGACGU 2892
1100 GCAGCUUU G UGGUGCUG 1118 CAGCACCA UGAUG GCAUGCACUAUGC GCG AAAGCUGC 2893
1144 UGGCCUCU G UCGGAGGG 1119 CCCUCCGA UGAUG GCAUGCACUAUGC GCG AGAGGCCA 2894
1185 CACUCGCU G UACACAGG 1120 CCUGUGUA UGAUG GCAUGCACUAUGC GCG AGCGAGUG 2895
1246 UGAUCAUU G UGCGGGUG 1121 CACCCGCA UGAUG GCAUGCACUAUGC GCG AAUGAUCA 2896
1315 AGAGCAUU G UGGACAGU 1122 ACUGUCCA UGAUG GCAUGCACUAUGC GCG AAUGCUCU 2897
1356 AAGAAAGU G UUUGAAGC 1123 GCUUCAAA UGAUG GCAUGCACUAUGC GCG ACUUUCUU 2898
1440 CAGCUGGU G UGCUGGCA 1124 UGCCAGCA UGAUG GCAUGCACUAUGC GCG ACCAGCUG 2899
1570 UGGAAGAU G UGGCCACG 1125 CGUGGCCA UGAUG GCAUGCACUAUGC GCG AUCUUCCA 2900
Table 20
1592 AGACGACU G UUACAAGU 1126 ACUUGUAA UGAUG GCAUGCACUAUGC GCG AGUCGUCU 2901
1630 CGGGCACU G UUAUGGGA 1127 UCCCAUAA UGAUG GCAUGCACUAUGC GCG AGUGCCCG 2902
1642 UGGGAGCU G UUAUCAUG 1128 CAUGAUAA UGAUG GCAUGCACUAUGC GCG AGCUCCCA 2903
1666 UCUACGUU G UCUUUGAU 1129 AUCAAAGA UGAUG GCAUGCACUAUGC GCG AACGUAGA 2904
1702 GCUUUGCU G UCAGCGCU 1130 AGCGCUGA UGAUG GCAUGCACUAUGC GCG AGCAAAGC 2905
1717 CUUGCCAU G UGCACGAU 1131 AUCGUGCA UGAUG GCAUGCACUAUGC GCG AUGGCAAG 2906
1759 GCCCUUUU G UCACCUUG 1132 CAAGGUGA UGAUG GCAUGCACUAUGC GCG AAAAGGGC 2907
1781 GGAAGACU G UGGCUACA 1133 UGUAGCCA UGAUG GCAUGCACUAUGC GCG AGUCUUCC 2908
1834 UAGCCUAU G UCAUGGCU 1134 AGCCAUGA UGAUG GCAUGCACUAUGC GCG AUAGGCUA 2909
1884 CUCAUGGU G UGUCAGUG 1135 CACUGACA UGAUG GCAUGCACUAUGC GCG ACCAUGAG 2910
1886 CAUGGUGU G UCAGUGGC 1136 GCCACUGA UGAUG GCAUGCACUAUGC GCG ACACCAUG 2911
2048 UGGCACCU G UGGCCAGA 1137 UCUGGCCA UGAUG GCAUGCACUAUGC GCG AGGUGCCA 2912
2139 CAGGGACU G UACCUGUA 1138 UACAGGUA UGAUG GCAUGCACUAUGC GCG AGUCCCUG 2913
2145 CUGUACCU G UAGGAAAC 1139 GUUUCCUA UGAUG GCAUGCACUAUGC GCG AGGUACAG 2914
2256 GAACCUUU G UCCACCAU 1140 AUGGUGGA UGAUG GCAUGCACUAUGC GCG AAAGGUUC 2915
2346 CUUGGCGU G UGUCCCUG 1141 CAGGGACA UGAUG GCAUGCACUAUGC GCG ACGCCAAG 2916
2348 UGGCGUGU G UCCCUGUG 1142 CACAGGGA UGAUG GCAUGCACUAUGC GCG ACACGCCA 2917
2354 GUGUCCCU G UGGUACCC 1143 GGGUACCA UGAUG GCAUGCACUAUGC GCG AGGGACAC 2918
2385 CCAAGCUU G UUUCCCUG 1144 CAGGGAAA UGAUG GCAUGCACUAUGC GCG AAGCUUGG 2919
2453 CAGGGACU G UAUAAACA 1145 UGUUUAUA UGAUG GCAUGCACUAUGC GCG AGUCCCUG 2920
Input Sequence = AF190725. Cut Site = G/ .
Stem Length = 8. Core Sequence = UGAUG GCAUGCACUAUGC GCG
AF190725 (Homo sapiens beta-site APP cleaving enzyme (BACE) mRNA; 2526 bp)
Table 21 Table 21: Human BACE Zinzyme Ribozyme and Target Sequence
Pos Substrate Seq ID Ribozyme Rz Seq ID
11 ACGCGUCC G CAGCCCGC 960 GCGGGCUG GCCGAAAGGCGAGUCAAGGUCU GGACGCGU 2921
18 CGCAGCCC G CCCGGGAG 961 , CUCCCGGG GCCGAAAGGCGAGUCAAGGUCU GGGCUGCG 2922
29 CGGGAGCU G CGAGCCGC 962 GCGGCUCG GCCGAAAGGCGAGUCAAGGUCU AGCUCCCG 2923
36 UGCGAGCC G CGAGCUGG 964 CCAGCUCG GCCGAAAGGCGAGUCAAGGUCU GGCUCGCA 2924
69 CAGCCAAC G CAGCCGCA 967 UGCGGCUG GCCGAAAGGCGAGUCAAGGUCU GUUGGCUG 2925
75 ACGCAGCC G CAGGAGCC 968 GGCUCCUG GCCGAAAGGCGAGUCAAGGUCU GGCUGCGU 2926
94 GAGCCCUU G CCCCUGCC 969 ■ GGCAGGGG GCCGAAAGGCGAGUCAAGGUCU AAGGGCUC 2927
100 UUGCCCCU G CCCGCGCC 970 GGCGCGGG GCCGAAAGGCGAGUCAAGGUCU AGGGGCAA 2928
104 CCCUGCCC G CGCCGCCG 971 CGGCGGCG GCCGAAAGGCGAGUCAAGGUCU GGGCAGGG 2929
106 CUGCCCGC G CCGCCGCC 972 GGCGGCGG GCCGAAAGGCGAGUCAAGGUCU GCGGGCAG 2930
109 CCCGCGCC G CCGCCCGC 973 GCGGGCGG GCCGAAAGGCGAGUCAAGGUCU GGCGCGGG 2931
112 GCGCCGCC G CCCGCCGG 974 CCGGCGGG GCCGAAAGGCGAGUCAAGGUCU GGCGGCGC 2932
116 CGCCGCCC G CCGGGGGG 975 CCCCCCGG GCCGAAAGGCGAGUCAAGGUCU GGGCGGCG 2933
137 GGGAAGCC G CCACCGGC 976 GCCGGUGG GCCGAAAGGCGAGUCAAGGUCU GGCUUCCC 2934
148 ACCGGCCC G CCAUGCCC 977 GGGCAUGG GCCGAAAGGCGAGUCAAGGUCU GGGCCGGU 2935
153 CCCGCCAU G CCCGCCCC 978 GGGGCGGG GCCGAAAGGCGAGUCAAGGUCU AUGGCGGG 2936
157 CCAUGCCC G CCCCUCCC 979 GGGAGGGG GCCGAAAGGCGAGUCAAGGUCU GGGCAUGG 2937
172 CCAGCCCC G CCGGGAGC 980 GCUCCCGG GCCGAAAGGCGAGUCAAGGUCU GGGGCUGG 2938
183 GGGAGCCC G CGCCCGCU 981 AGCGGGCG GCCGAAAGGCGAGUCAAGGUCU GGGCUCCC 2939
185 GAGCCCGC G CCCGCUGC 982 GCAGCGGG GCCGAAAGGCGAGUCAAGGUCU GCGGGCUC 2940
189 CCGCGCCC G CUGCCCAG 983 CUGGGCAG GCCGAAAGGCGAGUCAAGGUCU GGGCGCGG 2941
192 CGCCCGCU G CCCAGGCU 984 AGCCUGGG GCCGAAAGGCGAGUCAAGGUCU AGCGGGCG 2942
205 GGCUGGCC G CCGCCGUG 985 CACGGCGG GCCGAAAGGCGAGUCAAGGUCU GGCCAGCC 2943
208 UGGCCGCC G CCGUGCCG 986 CGGCACGG GCCGAAAGGCGAGUCAAGGUCU GGCGGCCA 2944
213 GCCGCCGU G CCGAUGUA 987 UACAUCGG GCCGAAAGGCGAGUCAAGGUCU ACGGCGGC 2945
250 UCUCCCCU G CUCCCGUG 989 CACGGGAG GCCGAAAGGCGAGUCAAGGUCU AGGGGAGA 2946
258 GCUCCCGU G CUCUGCGG 990 CCGCAGAG GCCGAAAGGCGAGUCAAGGUCU ACGGGAGC 2947
263 CGUGCUCU G CGGAUCUC 991 GAGAUCCG GCCGAAAGGCGAGUCAAGGUCU AGAGCACG 2948
280 CCCUGACC G CUCUCCAC 993 GUGGAGAG GCCGAAAGGCGAGUCAAGGUCU GGUCAGGG 2949
320 AGGGCCCU G CAGGCCCU 994 AGGGCCUG GCCGAAAGGCGAGUCAAGGUCU AGGGCCCU 2950
340 GUCCUGAU G CCCCCAAG 996 CUUGGGGG GCCGAAAGGCGAGUCAAGGUCU AUCAGGAC 2951
397 GGGCAGGC G CCAGGGAC 998 GUCCCUGG GCCGAAAGGCGAGUCAAGGUCU GCCUGCCC 2952
420 GGGCCAGU G CGAGCCCA 999 UGGGCUCG GCCGAAAGGCGAGUCAAGGUCU ACUGGCCC 2953
468 CAAGCCCU G CCCUGGCU 1002 AGCCAGGG GCCGAAAGGCGAGUCAAGGUCU AGGGCUUG 2954
480 UGGCUCCU G CUGUGGAU 1003 AUCCACAG GCCGAAAGGCGAGUCAAGGUCU AGGAGCCA 2955
493 GGAUGGGC G CGGGAGUG 1004 CACUCCCG GCCGAAAGGCGAGUCAAGGUCU GCCCAUCC * 2956
501 GCGGGAGU G CUGCCUGC 1005 GCAGGCAG GCCGAAAGGCGAGUCAAGGUCU ACUCCCGC 2957
504 GGAGUGCU G CCUGCCCA 1006 UGGGCAGG GCCGAAAGGCGAGUCAAGGUCU AGCACUCC 2958
508 UGCUGCCU G CCCACGGC 1007 GCCGUGGG GCCGAAAGGCGAGUCAAGGUCU AGGCAGCA 2959
537 AUCCGGCU G CCCCUGCG 1008 CGCAGGGG GCCGAAAGGCGAGUCAAGGUCU AGCCGGAU 2960
543 CUGCCCCU G CGCAGCGG 1009 CCGCUGCG GCCGAAAGGCGAGUCAAGGUCU AGGGGCAG 2961
545 GCCCCUGC G CAGCGGCC 1010 GGCCGCUG GCCGAAAGGCGAGUCAAGGUCU GCAGGGGC 2962
562 UGGGGGGC G CCCCCCUG 1011 CAGGGGGG GCCGAAAGGCGAGUCAAGGUCU GCCCCCCA 2963
576 CUGGGGCU G CGGCUGCC 1012 GGCAGCCG GCCGAAAGGCGAGUCAAGGUCU AGCCCCAG 2964
582 CUGCGGCU G CCCCGGGA 1013 UCCCGGGG GCCGAAAGGCGAGUCAAGGUCU AGCCGCAG 2965
708 AGCCCCCC G CAGACGCU 1019 AGCGUCUG GCCGAAAGGCGAGUCAAGGUCU GGGGGGCU 2966
714 CCGCAGAC G CUCAACAU 1020 AUGUUGAG GCCGAAAGGCGAGUCAAGGUCU GUCUGCGG 2967
Table 21
751 GUAACUUU G CAGUGGGU 1021 ACCCACUG GCCGAAAGGCGAGUCAAGGUCU AAAGUUAC 2968
760 CAGUGGGU G CUGCCCCC 1022 GGGGGCAG GCCGAAAGGCGAGUCAAGGUCU ACCCACUG 2969
763 UGGGUGCU G CCCCCCAC 1023 GUGGGGGG GCCGAAAGGCGAGUCAAGGUCU AGCACCCA 2970
780 CCCUUCCU G CAUCGCUA 1024 UAGCGAUG GCCGAAAGGCGAGUCAAGGUCU AGGAAGGG 2971
785 CCUGCAUC G CUACUACC 1025 GGUAGUAG GCCGAAAGGCGAGUCAAGGUCU GAUGCAGG 2972
843 GUGUAUGU G CCCUACAC 1026 GUGUAGGG GCCGAAAGGCGAGUCAAGGUCU ACAUACAC 2973
921 GUCACUGU G CGUGCCAA 1028 UUGGCACG GCCGAAAGGCGAGUCAAGGUCU ACAGUGAC 2974
925 CUGUGCGU G CCAACAUU 1029 AAUGUUGG GCCGAAAGGCGAGUCAAGGUCU ACGCACAG 2975
934 CCAACAUU G CUGCCAUC 1030 GAUGGCAG GCCGAAAGGCGAGUCAAGGUCU AAUGUUGG 2976
937 ACAUUGCU G CCAUCACU 1031 AGUGAUGG GCCGAAAGGCGAGUCAAGGUCU AGCAAUGU 2977
1006 UGGCCUAU G CUGAGAUU 1033 AAUCUCAG GCCGAAAGGCGAGUCAAGGUCU AUAGGCCA 2978
1015 CUGAGAUU G CCAGGCCU 1035 AGGCCUGG GCCGAAAGGCGAGUCAAGGUCU AAUCUCAG 2979
1092 UUCUCCCU G CAGCUUUG 1039 CAAAGCUG GCCGAAAGGCGAGUCAAGGUCU AGGGAGAA 2980
1105 UUUGUGGU G CUGGCUUC 1040 GAAGCCAG GCCGAAAGGCGAGUCAAGGUCU ACCACAAA 2981
1134 UCUGAAGU G CUGGCCUC 1042 GAGGCCAG GCCGAAAGGCGAGUCAAGGUCU ACUUCAGA 2982
1182 GACCACUC G CUGUACAC 1045 GUGUACAG GCCGAAAGGCGAGUCAAGGUCU GAGUGGUC 2983
1248 AUCAUUGU G CGGGUGGA 1048 UCCACCCG GCCGAAAGGCGAGUCAAGGUCU ACAAUGAU 2984
1286 AAUGGACU G CAAGGAGU 1050 ACUCCUUG GCCGAAAGGCGAGUCAAGGUCU AGUCCAUU 2985
1344 CUUCGUUU G CCCAAGAA 1052 UUCUUGGG GCCGAAAGGCGAGUCAAGGUCU AAACGAAG 2986
1366 UUGAAGCU G CAGUCAAA 1054 UUUGACUG GCCGAAAGGCGAGUCAAGGUCU AGCUUCAA 2987
1442 GCUGGUGU G CUGGCAAG 1056 " CUUGCCAG GCCGAAAGGCGAGUCAAGGUCU ACACCAGC 2988
1526 GUCCUUCC G CAUCACCA 1058 UGGUGAUG GCCGAAAGGCGAGUCAAGGUCU GGAAGGAC 2989
1542 AUCCUUCC G CAGCAAUA 1059 UAUUGCUG GCCGAAAGGCGAGUCAAGGUCU GGAAGGAU 2990
1554 CAAUACCU G CGGCCAGU 1060 ACUGGCCG GCCGAAAGGCGAGUCAAGGUCU AGGUAUUG 2991
1603 ACAAGUUU G CCAUCUCA 1062 UGAGAUGG GCCGAAAGGCGAGUCAAGGUCU AAACUUGU 2992
1699 UUGGCUUU G CUGUCAGC 1066 GCUGACAG GCCGAAAGGCGAGUCAAGGUCU AAAGCCAA 2993
1708 CUGUCAGC G CUUGCCAU 1067 AUGGCAAG GCCGAAAGGCGAGUCAAGGUCU GCUGACAG 2994
1712 CAGCGCUU G CCAUGUGC 1068 GCACAUGG GCCGAAAGGCGAGUCAAGGUCU AAGCGCUG 2995
1719 UGCCAUGU G CACGAUGA 1069 UCAUCGUG GCCGAAAGGCGAGUCAAGGUCU ACAUGGCA 2996
1843 UCAUGGCU G CCAUCUGC 1074 GCAGAUGG GCCGAAAGGCGAGUCAAGGUCU AGCCAUGA 2997
1850 UGCCAUCU G CGCCCUCU 1075 AGAGGGCG GCCGAAAGGCGAGUCAAGGUCU AGAUGGCA 2998
1852 CCAUCUGC G CCCUCUUC 1076 GAAGAGGG GCCGAAAGGCGAGUCAAGGUCU GCAGAUGG 2999
1863 CUCUUCAU G CUGCCACU 1077 AGUGGCAG GCCGAAAGGCGAGUCAAGGUCU AUGAAGAG 3000
1866 UUCAUGCU G CCACUCUG 1078 CAGAGUGG GCCGAAAGGCGAGUCAAGGUCU AGCAUGAA 3001
1874 GCCACUCU G CCUCAUGG 1079 CCAUGAGG GCCGAAAGGCGAGUCAAGGUCU AGAGUGGC 3002
1895 UCAGUGGC G CUGCCUCC 1080 GGAGGCAG GCCGAAAGGCGAGUCAAGGUCU GCCACUGA 3003
1898 GUGGCGCU G CCUCCGCU 1081 AGCGGAGG GCCGAAAGGCGAGUCAAGGUCU AGCGCCAC 3004
1904 CUGCCUCC G CUGCCUGC 1082 GCAGGCAG GCCGAAAGGCGAGUCAAGGUCU GGAGGCAG 3005
1907 CCUCCGCU G CCUGCGCC 1083 GGCGCAGG GCCGAAAGGCGAGUCAAGGUCU AGCGGAGG 3006
1911 CGCUGCCU G CGCCAGCA 1084 UGCUGGCG GCCGAAAGGCGAGUCAAGGUCU AGGCAGCG 3007
1913 CUGCCUGC G CCAGCAGC 1085 GCUGCUGG GCCGAAAGGCGAGUCAAGGUCU GCAGGCAG 3008
1933 AUGACUUU G CUGAUGAC 1088 GUCAUCAG GCCGAAAGGCGAGUCAAGGUCU AAAGUCAU 3009
1950 AUCUCCCU G CUGAAGUG 1091 CACUUCAG GCCGAAAGGCGAGUCAAGGUCU AGGGAGAU 3010
2087 CACCAAAU G CCUCUGCC 1094 GGCAGAGG GCCGAAAGGCGAGUCAAGGUCU AUUUGGUG 3011
2093 AUGCCUCU G CCUUGAUG 1095 CAUCAAGG GCCGAAAGGCGAGUCAAGGUCU AGAGGCAU 3012
2179 AGCACUCU G CUGGCGGG 1097 CCCGCCAG GCCGAAAGGCGAGUCAAGGUCU AGAGUGCU 3013
2227 GAAAUUCU G CUGCUUGA 1098 UCAAGCAG GCCGAAAGGCGAGUCAAGGUCU AGAAUUUC 3014
2230 AUUCUGCU G CUUGAAAC 1099 GUUUCAAG GCCGAAAGGCGAGUCAAGGUCU AGCAGAAU 3015
2329 CAUCACAC G CAGGUUAC 1102 GUAACCUG GCCGAAAGGCGAGUCAAGGUCU GUGUGAUG 3016
2393 GUUUCCCU G CUGGCCAA 1103 UUGGCCAG GCCGAAAGGCGAGUCAAGGUCU AGGGAAAC 3017
2419 GAGAGGAU G CACAGUUU 1104 AAACUGUG GCCGAAAGGCGAGUCAAGGUCU AUCCUCUC 3018
Table 21
2428 CACAGUUU G CUAUUUGC 1105 GCAAAUAG GCCGAAAGGCGAGUCAAGGUCU AAACUGUG 3019
2435 UGCUAUUU G CUUUAGAG 1106 CUCUAAAG GCCGAAAGGCGAGUCAAGGUCU AAAUAGCA 3020
2476 ACAUUGGU G CAAAGAUU 1107 AAUCUUUG GCCGAAAGGCGAGUCAAGGUCU ACCAAUGU 3021
2485 CAAAGAUU G CCUCUUGA 1108 UCAAGAGG GCCGAAAGGCGAGUCAAGGUCU AAUCUUUG 3022
219 GUGCCGAU G UAGCGGGC 1110 GCCCGCUA GCCGAAAGGCGAGUCAAGGUCU AUCGGCAC 3023
483 CUCCUGCU G UGGAUGGG llll CCCAUCCA GCCGAAAGGCGAGUCAAGGUCU AGCAGGAG 3024
634 GCAGCUUU G UGGAGAUG 1112 CAUCUCCA GCCGAAAGGCGAGUCAAGGUCU AAAGCUGC 3025
804 AGGCAGCU G UCCAGCAC 1113 GUGCUGGA GCCGAAAGGCGAGUCAAGGUCU AGCUGCCU 3026
835 GGAAGGGU G UGUAUGUG 1114 CACAUACA GCCGAAAGGCGAGUCAAGGUCU ACCCUUCC 3027
837 AAGGGUGU G UAUGUGCC 1115 GGCACAUA GCCGAAAGGCGAGUCAAGGUCU ACACCCUU 3028
841 GUGUGUAU G UGCCCUAC 1116 GUAGGGCA GCCGAAAGGCGAGUCAAGGUCU AUACACAC 3029
919 ACGUCACU G UGCGUGCC 1117 GGCACGCA GCCGAAAGGCGAGUCAAGGUCU AGUGACGU 3030
1100 GCAGCUUU G UGGUGCUG 1118 CAGCACCA GCCGAAAGGCGAGUCAAGGUCU AAAGCUGC 3031
1144 UGGCCUCU G UCGGAGGG 1119 CCCUCCGA GCCGAAAGGCGAGUCAAGGUCU AGAGGCCA 3032
1185 CACUCGCU G UACACAGG 1120 CCUGUGUA GCCGAAAGGCGAGUCAAGGUCU AGCGAGUG 3033
1246 UGAUCAUU G UGCGGGUG 1121 CACCCGCA GCCGAAAGGCGAGUCAAGGUCU AAUGAUCA 3034
1315 AGAGCAUU G UGGACAGU 1122 ACUGUCCA GCCGAAAGGCGAGUCAAGGUCU AAUGCUCU 3035
1356 AAGAAAGU G UUUGAAGC 1123 GCUUCAAA GCCGAAAGGCGAGUCAAGGUCU ACUUUCUU 3036
1440 CAGCUGGU G UGCUGGCA 1124 UGCCAGCA GCCGAAAGGCGAGUCAAGGUCU ACCAGCUG 3037
1570 UGGAAGAU G UGGCCACG 1125 CGUGGCCA GCCGAAAGGCGAGUCAAGGUCU AUCUUCCA 3038
1592 AGACGACU G UUACAAGU 1126 ACUUGUAA GCCGAAAGGCGAGUCAAGGUCU AGUCGUCU 3039
1630 CGGGCACU G UUAUGGGA 1127 UCCCAUAA GCCGAAAGGCGAGUCAAGGUCU AGUGCCCG 3040
1642 UGGGAGCU G UUAUCAUG 1128 CAUGAUAA GCCGAAAGGCGAGUCAAGGUCU AGCUCCCA 3041
1666 UCUACGUU G UCUUUGAU 1129 AUCAAAGA GCCGAAAGGCGAGUCAAGGUCU AACGUAGA 3042
1702 GCUUUGCU G UCAGCGCU 1130 AGCGCUGA GCCGAAAGGCGAGUCAAGGUCU AGCAAAGC 3043
1717 CUUGCCAU G UGCACGAU 1131 AUCGUGCA GCCGAAAGGCGAGUCAAGGUCU AUGGCAAG 3044
1759 GCCCUUUU G UCACCUUG 1132 CAAGGUGA GCCGAAAGGCGAGUCAAGGUCU AAAAGGGC 3045
1781 GGAAGACU G UGGCUACA 1133 UGUAGCCA GCCGAAAGGCGAGUCAAGGUCU AGUCUUCC 3046
1834 UAGCCUAU G UCAUGGCU 1134 AGCCAUGA GCCGAAAGGCGAGUCAAGGUCU AUAGGCUA 3047
1884 CUCAUGGU G UGUCAGUG 1135 CACUGACA GCCGAAAGGCGAGUCAAGGUCU ACCAUGAG 3048
1886 CAUGGUGU G UCAGUGGC 1136 GCCACUGA GCCGAAAGGCGAGUCAAGGUCU ACACCAUG 3049
2048 UGGCACCU G UGGCCAGA 1137 UCUGGCCA GCCGAAAGGCGAGUCAAGGUCU AGGUGCCA 3050
2139 CAGGGACU G UACCUGUA 1138 UACAGGUA GCCGAAAGGCGAGUCAAGGUCU AGUCCCUG 3051
2145 CUGUACCU G UAGGAAAC 1139 GUUUCCUA GCCGAAAGGCGAGUCAAGGUCU AGGUACAG 3052
2256 GAACCUUU G UCCACCAU 1140 AUGGUGGA GCCGAAAGGCGAGUCAAGGUCU AAAGGUUC 3053
2346 CUUGGCGU G UGUCCCUG 1141 CAGGGACA GCCGAAAGGCGAGUCAAGGUCU ACGCCAAG 3054
2348 UGGCGUGU G UCCCUGUG 1142 CACAGGGA GCCGAAAGGCGAGUCAAGGUCU ACACGCCA 3055
2354 GUGUCCCU G UGGUACCC 1143 GGGUACCA GCCGAAAGGCGAGUCAAGGUCU AGGGACAC 3056
2385 CCAAGCUU G UUUCCCUG 1144 CAGGGAAA GCCGAAAGGCGAGUCAAGGUCU AAGCUUGG 3057
2453 CAGGGACU G UAUAAACA 1145 UGUUUAUA GCCGAAAGGCGAGUCAAGGUCU AGUCCCUG 3058
14 CGUCCGCA G CCCGCCCG 1146 CGGGCGGG GCCGAAAGGCGAGUCAAGGUCU UGCGGACG 3059
26 GCCCGGGA G CUGCGAGC 1147 GCUCGCAG GCCGAAAGGCGAGUCAAGGUCU UCCCGGGC 3060
33 AGCUGCGA G CCGCGAGC 1148 GCUCGCGG GCCGAAAGGCGAGUCAAGGUCU UCGCAGCU 3061
40 AGCCGCGA G CUGGAUUA 1149 UAAUCCAG GCCGAAAGGCGAGUCAAGGUCU UCGCGGCU 3062
51 GGAUUAUG G UGGCCUGA 1150 UCAGGCCA GCCGAAAGGCGAGUCAAGGUCU CAUAAUCC 3063
54 UUAUGGUG G CCUGAGCA 1151 UGCUCAGG GCCGAAAGGCGAGUCAAGGUCU CACCAUAA 3064
60 UGGCCUGA G CAGCCAAC 1152 GUUGGCUG GCCGAAAGGCGAGUCAAGGUCU UCAGGCCA 3065
63 CCUGAGCA G CCAACGCA 1153 UGCGUUGG GCCGAAAGGCGAGUCAAGGUCU UGCUCAGG 3066
72 CCAACGCA G CCGCAGGA 1154 UCCUGCGG GCCGAAAGGCGAGUCAAGGUCU UGCGUUGG 3067
81 CCGCAGGA G CCCGGAGC 1155 GCUCCGGG GCCGAAAGGCGAGUCAAGGUCU UCCUGCGG 3068
88 AGCCCGGA G CCCUUGCC 1156 GGCAAGGG GCCGAAAGGCGAGUCAAGGUCU UCCGGGCU 3069
Table 21
134 CCAGGGAA G CCGCCACC 1157 GGUGGCGG GCCGAAAGGCGAGUCAAGGUCU UUCCCUGG 3070
144 CGCCACCG G CCCGCCAU 1158 AUGGCGGG GCCGAAAGGCGAGUCAAGGUCU CGGUGGCG 3071
167 CCCUCCCA G CCCCGCCG 1159 CGGCGGGG GCCGAAAGGCGAGUCAAGGUCU UGGGAGGG 3072
179 CGCCGGGA G CCCGCGCC 1160 GGCGCGGG GCCGAAAGGCGAGUCAAGGUCU UCCCGGCG 3073
198 CUGCCCAG G CUGGCCGC 1161 GCGGCCAG GCCGAAAGGCGAGUCAAGGUCU CUGGGCAG 3074
202 CCAGGCUG G CCGCCGCC 1162 GGCGGCGG GCCGAAAGGCGAGUCAAGGUCU CAGCCUGG 3075
211 CCGCCGCC G UGCCGAUG 1163 CAUCGGCA GCCGAAAGGCGAGUCAAGGUCU GGCGGCGG 3076
222 CCGAUGUA G CGGGCUCC 1164 GGAGCCCG GCCGAAAGGCGAGUCAAGGUCU UACAUCGG 3077
226 UGUAGCGG G CUCCGGAU 1165 AUCCGGAG GCCGAAAGGCGAGUCAAGGUCU CCGCUACA 3078
239 GGAUCCCA G CCUCUCCC 1166 GGGAGAGG GCCGAAAGGCGAGUCAAGGUCU UGGGAUCC 3079
256 CUGCUCCC G UGCUCUGC 1167 GCAGAGCA GCCGAAAGGCGAGUCAAGGUCU GGGAGCAG 3080
290 UCUCCACA G CCCGGACC 1168 GGUCCGGG GCCGAAAGGCGAGUCAAGGUCU UGUGGAGA 3081
304 ACCCGGGG G CUGGCCCA 1169 UGGGCCAG GCCGAAAGGCGAGUCAAGGUCU CCCCGGGU 3082
308 GGGGGCUG G CCCAGGGC 1170 GCCCUGGG GCCGAAAGGCGAGUCAAGGUCU CAGCCCCC 3083
315 GGCCCAGG G CCCUGCAG 1171 CUGCAGGG GCCGAAAGGCGAGUCAAGGUCU CCUGGGCC 3084
324 CCCUGCAG G CCCUGGCG 1172 CGCCAGGG GCCGAAAGGCGAGUCAAGGUCU CUGCAGGG 3085
330 AGGCCCUG G CGUCCUGA 1173 UCAGGACG GCCGAAAGGCGAGUCAAGGUCU CAGGGCCU 3086
332 GCCCUGGC G UCCUGAUG 1174 CAUCAGGA GCCGAAAGGCGAGUCAAGGUCU GCCAGGGC 3087
348 GCCCCCAA G CUCCCUCU 1175 AGAGGGAG GCCGAAAGGCGAGUCAAGGUCU UUGGGGGC 3088
365 CCUGAGAA G CCACCAGC 1176 GCUGGUGG GCCGAAAGGCGAGUCAAGGUCU UUCUCAGG 3089
372 AGCCACCA G CACCACCC 1177 GGGUGGUG GCCGAAAGGCGAGUCAAGGUCU UGGUGGCU 3090
391 ACUUGGGG G CAGGCGCC 1178 GGCGCCUG GCCGAAAGGCGAGUCAAGGUCU CCCCAAGU 3091
395 GGGGGCAG G CGCCAGGG 1179 CCCUGGCG GCCGAAAGGCGAGUCAAGGUCU CUGCCCCC 3092
410 GGACGGAC G UGGGCCAG 1180 CUGGCCCA GCCGAAAGGCGAGUCAAGGUCU GUCCGUCC 3093
414 GGACGUGG G CCAGUGCG 1181 CGCACUGG GCCGAAAGGCGAGUCAAGGUCU CCACGUCC 3094
418 GUGGGCCA G UGCGAGCC 1182 GGCUCGCA GCCGAAAGGCGAGUCAAGGUCU UGGCCCAC 3095
424 CAGUGCGA G CCCAGAGG 1183 CCUCUGGG GCCGAAAGGCGAGUCAAGGUCU UCGCACUG 3096
433 CCCAGAGG G CCCGAAGG 1184 CCUUCGGG GCCGAAAGGCGAGUCAAGGUCU CCUCUGGG 3097
441 GCCCGAAG G CCGGGGCC 1185 GGCCCCGG GCCGAAAGGCGAGUCAAGGUCU CUUCGGGC 3098
447 AGGCCGGG G CCCACCAU 1186 AUGGUGGG GCCGAAAGGCGAGUCAAGGUCU CCCGGCCU 3099
457 CCACCAUG G CCCAAGCC 1187 GGCUUGGG GCCGAAAGGCGAGUCAAGGUCU CAUGGUGG 3100
463 UGGCCCAA G CCCUGCCC 1188 GGGCAGGG GCCGAAAGGCGAGUCAAGGUCU UUGGGCCA 3101
474 CUGCCCUG G CUCCUGCU 1189 AGCAGGAG GCCGAAAGGCGAGUCAAGGUCU CAGGGCAG 3102
491 GUGGAUGG G CGCGGGAG 1190 CUCCCGCG GCCGAAAGGCGAGUCAAGGUCU CCAUCCAC 3103
499 GCGCGGGA G UGCUGCCU 1191 AGGCAGCA GCCGAAAGGCGAGUCAAGGUCU UCCCGCGC 3104
515 UGCCCACG G CACCCAGC 1192 GCUGGGUG GCCGAAAGGCGAGUCAAGGUCU CGUGGGCA 3105
522 GGCACCCA G CACGGCAU 1193 AUGCCGUG GCCGAAAGGCGAGUCAAGGUCU UGGGUGCC 3106
527 CCAGCACG G CAUCCGGC 1194 GCCGGAUG GCCGAAAGGCGAGUCAAGGUCU CGUGCUGG 3107
534 GGCAUCCG G CUGCCCCU 1195 AGGGGCAG GCCGAAAGGCGAGUCAAGGUCU CGGAUGCC 3108
548 CCUGCGCA G CGGCCUGG 1196 CCAGGCCG GCCGAAAGGCGAGUCAAGGUCU UGCGCAGG 3109
551 GCGCAGCG G CCUGGGGG 1197 CCCCCAGG GCCGAAAGGCGAGUCAAGGUCU CGCUGCGC 3110
560 CCUGGGGG G CGCCCCCC 1198 GGGGGGCG GCCGAAAGGCGAGUCAAGGUCU CCCCCAGG 3111
573 CCCCUGGG G CUGCGGCU 1199 AGCCGCAG GCCGAAAGGCGAGUCAAGGUCU CCCAGGGG 3112
579 GGGCUGCG G CUGCCCCG 1200 CGGGGCAG GCCGAAAGGCGAGUCAAGGUCU CGCAGCCC 3113
603 GACGAAGA G CCCGAGGA 1201 UCCUCGGG GCCGAAAGGCGAGUCAAGGUCU UCUUCGUC 3114
612 CCCGAGGA G CCCGGCCG 1202 CGGCCGGG GCCGAAAGGCGAGUCAAGGUCU UCCUCGGG 3115
617 GGAGCCCG G CCGGAGGG 1203 CCCUCCGG GCCGAAAGGCGAGUCAAGGUCU CGGGCUCC 3116
626 CCGGAGGG G CAGCUUUG 1204 CAAAGCUG GCCGAAAGGCGAGUCAAGGUCU CCCUCCGG 3117
629 GAGGGGCA G CUUUGUGG 1205 CCACAAAG GCCGAAAGGCGAGUCAAGGUCU UGCCCCUC 3118
643 UGGAGAUG G UGGACAAC 1206 GUUGUCCA GCCGAAAGGCGAGUCAAGGUCU CAUCUCCA 3119
659 CCUGAGGG G CAAGUCGG 1207 CCGACUUG GCCGAAAGGCGAGUCAAGGUCU CCCUCAGG 3120
Table 21
663 AGGGGCAA G UCGGGGCA 1208- UGCCCCGA GCCGAAAGGCGAGUCAAGGUCU UUGCCCCU 3121
669 AAGUCGGG G CAGGGCUA 1209 UAGCCCUG GCCGAAAGGCGAGUCAAGGUCU CCCGACUU 3122
674 GGGGCAGG G CUACUACG 1210 CGUAGUAG GCCGAAAGGCGAGUCAAGGUCU CCUGCCCC 3123
682 GCUACUAC G UGGAGAUG 1211 CAUCUCCA GCCGAAAGGCGAGUCAAGGUCU GUAGUAGC 3124
694 AGAUGACC G UGGGCAGC 1212 GCUGCCCA GCCGAAAGGCGAGUCAAGGUCU GGUCAUCU 3125
698 GACCGUGG G CAGCCCCC 1213 GGGGGCUG GCCGAAAGGCGAGUCAAGGUCU CCACGGUC 3126
701 CGUGGGCA G CCCCCCGC 1214 GCGGGGGG GCCGAAAGGCGAGUCAAGGUCU UGCCCACG 3127
727 ACAUCCUG G UGGAUACA 1215 UGUAUCCA GCCGAAAGGCGAGUCAAGGUCU CAGGAUGU 3128
737 GGAUACAG G CAGCAGUA 1216 UACUGCUG GCCGAAAGGCGAGUCAAGGUCU CUGUAUCC 3129
740 UACAGGCA G CAGUAACU 1217 AGUUACUG GCCGAAAGGCGAGUCAAGGUCU UGCCUGUA 3130
743 AGGCAGCA G UAACUUUG 1218 CAAAGUUA GCCGAAAGGCGAGUCAAGGUCU UGCUGCCU 3131
754 ACUUUGCA G UGGGUGCU 1219 AGCACCCA GCCGAAAGGCGAGUCAAGGUCU UGCAAAGU 3132
758 UGCAGUGG G UGCUGCCC 1220 GGGCAGCA GCCGAAAGGCGAGUCAAGGUCU CCACUGCA 3133
798 UACCAGAG G CAGCUGUC 1221 GACAGCUG GCCGAAAGGCGAGUCAAGGUCU CUCUGGUA 3134
801 CAGAGGCA G CUGUCCAG 1222 CUGGACAG GCCGAAAGGCGAGUCAAGGUCU UGCCUCUG 3135
809 GCUGUCCA G CACAUACC 1223 GGUAUGUG GCCGAAAGGCGAGUCAAGGUCU UGGACAGC 3136
833 CCGGAAGG G UGUGUAUG 1224 CAUACACA GCCGAAAGGCGAGUCAAGGUCU CCUUCCGG 3137
857 CACCCAGG G CAAGUGGG 1225 CCCACUUG GCCGAAAGGCGAGUCAAGGUCU CCUGGGUG 3138
861 CAGGGCAA G UGGGAAGG 1226 CCUUCCCA GCCGAAAGGCGAGUCAAGGUCU UUGCCCUG 3139
873 GAAGGGGA G CUGGGCAC 1227 GUGCCCAG GCCGAAAGGCGAGUCAAGGUCU UCCCCUUC 3140
878 GGAGCUGG G CACCGACC 1228 GGUCGGUG GCCGAAAGGCGAGUCAAGGUCU CCAGCUCC 3141
889 CCGACCUG G UAAGCAUC 1229 GAUGCUUA GCCGAAAGGCGAGUCAAGGUCU CAGGUCGG 3142
893 CCUGGUAA G CAUCCCCC 1230 GGGGGAUG GCCGAAAGGCGAGUCAAGGUCU UUACCAGG 3143
905 CCCCCAUG G CCCCAACG 1231 CGUUGGGG GCCGAAAGGCGAGUCAAGGUCU CAUGGGGG 3144
913 GCCCCAAC G UCACUGUG 1232 CACAGUGA GCCGAAAGGCGAGUCAAGGUCU GUUGGGGC 3145
923 CACUGUGC G UGCCAACA 1233 UGUUGGCA GCCGAAAGGCGAGUCAAGGUCU GCACAGUG 3146
957 UCAGACAA G UUCUUCAU 1234 AUGAAGAA GCCGAAAGGCGAGUCAAGGUCU UUGUCUGA 3147
971 CAUCAACG G CUCCAACU 1235 AGUUGGAG GCCGAAAGGCGAGUCAAGGUCU CGUUGAUG 3148
986 CUGGGAAG G CAUCCUGG 1236 CCAGGAUG GCCGAAAGGCGAGUCAAGGUCU CUUCCCAG 3149
996 AUCCUGGG G CUGGCCUA 1237 UAGGCCAG GCCGAAAGGCGAGUCAAGGUCU CCCAGGAU 3150
1000 UGGGGCUG G CCUAUGCU 1238 AGCAUAGG GCCGAAAGGCGAGUCAAGGUCU CAGCCCCA 3151
1020 AUUGCCAG G CCUGACGA 1239 UCGUCAGG GCCGAAAGGCGAGUCAAGGUCU CUGGCAAU 3152
1038 UCCCUGGA G CCUUUCUU 1240 AAGAAAGG GCCGAAAGGCGAGUCAAGGUCU UCCAGGGA 3153
1057 ACUCUCUG G UAAAGCAG 1241 CUGCUUUA GCCGAAAGGCGAGUCAAGGUCU CAGAGAGU 3154
1062 CUGGUAAA G CAGACCCA 1242 UGGGUCUG GCCGAAAGGCGAGUCAAGGUCU UUUACCAG 3155
1072 AGACCCAC G UUCCCAAC 1243 GUUGGGAA GCCGAAAGGCGAGUCAAGGUCU GUGGGUCU 3156
1095 UCCCUGCA G CUUUGUGG 1244 CCACAAAG GCCGAAAGGCGAGUCAAGGUCU UGCAGGGA 3157
1103 GCUUUGUG G UGCUGGCU 1245 AGCCAGCA GCCGAAAGGCGAGUCAAGGUCU CACAAAGC 3158
1109 UGGUGCUG G CUUCCCCC 1246 GGGGGAAG GCCGAAAGGCGAGUCAAGGUCU CAGCACCA 3159
1125 CUCAACCA G UCUGAAGU 1247 ACUUCAGA GCCGAAAGGCGAGUCAAGGUCU UGGUUGAG 3160
1132 AGUCUGAA G UGCUGGCC 1248 GGCCAGCA GCCGAAAGGCGAGUCAAGGUCU UUCAGACU 3161
1138 AAGUGCUG G CCUCUGUC 1249 GACAGAGG GCCGAAAGGCGAGUCAAGGUCU CAGCACUU 3162
1154 CGGAGGGA G CAUGAUCA 1250 UGAUCAUG GCCGAAAGGCGAGUCAAGGUCU UCCCUCCG 3163
1169 CAUUGGAG G UAUCGACC 1251 GGUCGAUA GCCGAAAGGCGAGUCAAGGUCU CUCCAAUG 3164
1193 GUACACAG G CAGUCUCU 1252 AGAGACUG GCCGAAAGGCGAGUCAAGGUCU CUGUGUAC 3165
1196 CACAGGCA G UCUCUGGU 1253 ACCAGAGA GCCGAAAGGCGAGUCAAGGUCU UGCCUGUG 3166
1203 AGUCUCUG G UAUACACC 1254 GGUGUAUA GCCGAAAGGCGAGUCAAGGUCU CAGAGACU 3167
1218 CCCAUCCG G CGGGAGUG 1255 CACUCCCG GCCGAAAGGCGAGUCAAGGUCU CGGAUGGG 3168
1224 CGGCGGGA G UGGUAUUA 1256 UAAUACCA GCCGAAAGGCGAGUCAAGGUCU UCCCGCCG 3169
1227 CGGGAGUG G UAUUAUGA 1257 UCAUAAUA GCCGAAAGGCGAGUCAAGGUCU CACUCCCG 3170
1237 AUUAUGAG G UGAUCAUU 1258 AAUGAUCA GCCGAAAGGCGAGUCAAGGUCU CUCAUAAU 3171
Table 21
1252 UUGUGCGG G UGGAGAUC 1259 GAUCUCCA GCCGAAAGGCGAGUCAAGGUCU CCGCACAA 3172
1293 UGCAAGGA G UACAACUA 1260 UAGUUGUA GCCGAAAGGCGAGUCAAGGUCU UCCUUGCA 3173
1310 UGACAAGA G CAUUGUGG 1261 CCACAAUG GCCGAAAGGCGAGUCAAGGUCU UCUUGUCA 3174
1322 UGUGGACA G UGGCACCA 1262 UGGUGCCA GCCGAAAGGCGAGUCAAGGUCU UGUCCACA 3175
1325 GGACAGUG G CACCACCA 1263 UGGUGGUG GCCGAAAGGCGAGUCAAGGUCU CACUGUCC 3176
1340 CAACCUUC G UUUGCCCA 1264 UGGGCAAA GCCGAAAGGCGAGUCAAGGUCU GAAGGUUG 3177
1354 CCAAGAAA G UGUUUGAA 1265 UUCAAACA GCCGAAAGGCGAGUCAAGGUCU UUUCUUGG 3178
1363 UGUUUGAA G CUGCAGUC 1266 GACUGCAG GCCGAAAGGCGAGUCAAGGUCU UUCAAACA 3179
1369 AAGCUGCA G UCAAAUCC 1267 GGAUUUGA GCCGAAAGGCGAGUCAAGGUCU UGCAGCUU 3180
1384 CCAUCAAG G CAGCCUCC 1268 GGAGGCUG GCCGAAAGGCGAGUCAAGGUCU CUUGAUGG 3181
1387 UCAAGGCA G CCUCCUCC 1269 GGAGGAGG GCCGAAAGGCGAGUCAAGGUCU UGCCUUGA 3182
1404 ACGGAGAA G UUCCCUGA 1270 UCAGGGAA GCCGAAAGGCGAGUCAAGGUCU UUCUCCGU 3183
1415 CCCUGAUG G UUUCUGGC 1271 GCCAGAAA GCCGAAAGGCGAGUCAAGGUCU CAUCAGGG 3184
1422 GGUUUCUG G CUAGGAGA 1272 UCUCCUAG GCCGAAAGGCGAGUCAAGGUCU CAGAAACC 3185
1431 CUAGGAGA G CAGCUGGU 1273 ACCAGCUG GCCGAAAGGCGAGUCAAGGUCU UCUCCUAG 3186
1434 GGAGAGCA G CUGGUGUG 1274 CACACCAG GCCGAAAGGCGAGUCAAGGUCU UGCUCUCC 3187
1438 AGCAGCUG G UGUGCUGG 1275 CCAGCACA GCCGAAAGGCGAGUCAAGGUCU CAGCUGCU 3188
1446 GUGUGCUG G CAAGCAGG 1276 CCUGCUUG GCCGAAAGGCGAGUCAAGGUCU CAGCACAC 3189
1450 GCUGGCAA G CAGGCACC 1277 GGUGCCUG GCCGAAAGGCGAGUCAAGGUCU UUGCCAGC 3190
1454 GCAAGCAG G CACCACCC 1278 GGGUGGUG GCCGAAAGGCGAGUCAAGGUCU CUGCUUGC 3191
1480 UUUUCCCA G UCAUCUCA 1279 UGAGAUGA GCCGAAAGGCGAGUCAAGGUCU UGGGAAAA 3192
1502 CCUAAUGG G UGAGGUUA 1280 UAACCUCA GCCGAAAGGCGAGUCAAGGUCU CCAUUAGG 3193
1507 UGGGUGAG G UUACCAAC 1281 GUUGGUAA GCCGAAAGGCGAGUCAAGGUCU CUCACCCA 3194
1518 ACCAACCA G UCCUUCCG 1282 CGGAAGGA GCCGAAAGGCGAGUCAAGGUCU UGGUUGGU 3195
1545 CUUCCGCA G CAAUACCU 1283 AGGUAUUG GCCGAAAGGCGAGUCAAGGUCU UGCGGAAG 3196
1557 UACCUGCG G CCAGUGGA 1284 UCCACUGG GCCGAAAGGCGAGUCAAGGUCU CGCAGGUA 3197
1561 UGCGGCCA G UGGAAGAU 1285 AUCUUCCA GCCGAAAGGCGAGUCAAGGUCU UGGCCGCA 3198
1573 AAGAUGUG G CCACGUCC 1286 GGACGUGG GCCGAAAGGCGAGUCAAGGUCU CACAUCUU 3199
1578 GUGGCCAC G UCCCAAGA 1287 UCUUGGGA GCCGAAAGGCGAGUCAAGGUCU GUGGCCAC 3200
1599 UGUUACAA G UUUGCCAU 1288 AUGGCAAA GCCGAAAGGCGAGUCAAGGUCU UUGUAACA 3201
1614 AUCUCACA G UCAUCCAC 1289 GUGGAUGA GCCGAAAGGCGAGUCAAGGUCU UGUGAGAU 3202
1625 AUCCACGG G CACUGUUA 1290 UAACAGUG GCCGAAAGGCGAGUCAAGGUCU CCGUGGAU 3203
1639 UUAUGGGA G CUGUUAUC 1291 GAUAACAG GCCGAAAGGCGAGUCAAGGUCU UCCCAUAA 3204
1655 CAUGGAGG G CUUCUACG 1292 CGUAGAAG GCCGAAAGGCGAGUCAAGGUCU CCUCCAUG 3205
1663 GCUUCUAC G uuσucuuu 1293 AAAGACAA GCCGAAAGGCGAGUCAAGGUCU GUAGAAGC 3206
1678 UUGAUCGG G CCCGAAAA 1294 UUUUCGGG GCCGAAAGGCGAGUCAAGGUCU CCGAUCAA 3207
1694 ACGAAUUG G CUUUGCUG 1295 CAGCAAAG GCCGAAAGGCGAGUCAAGGUCU CAAUUCGU 3208
1706 UGCUGUCA G CGCUUGCC 1296 GGCAAGCG GCCGAAAGGCGAGUCAAGGUCU UGACAGCA 3209
1728 CACGAUGA G UUCAGGAC 1297 GUCCUGAA GCCGAAAGGCGAGUCAAGGUCU UCAUCGUG 3210
1738 UCAGGACG G CAGCGGUG 1298 CACCGCUG GCCGAAAGGCGAGUCAAGGUCU CGUCCUGA 3211
1741 GGACGGCA G CGGUGGAA 1299 UUCCACCG GCCGAAAGGCGAGUCAAGGUCU UGCCGUCC 3212
1744 CGGCAGCG G UGGAAGGC 1300 GCCUUCCA GCCGAAAGGCGAGUCAAGGUCU CGCUGCCG 3213
1751 GGUGGAAG G CCCUUUUG 1301 CAAAAGGG GCCGAAAGGCGAGUCAAGGUCU CUUCCACC 3214
1784 AGACUGUG G CUACAACA 1302 UGUUGUAG GCCGAAAGGCGAGUCAAGGUCU CACAGUCU 3215
1809 ACAGAUGA G UCAACCCU 1303 AGGGUUGA GCCGAAAGGCGAGUCAAGGUCU UCAUCUGU 3216
1828 UGACCAUA G CCUAUGUC 1304 GACAUAGG GCCGAAAGGCGAGUCAAGGUCU UAUGGUCA 3217
1840 AUGUCAUG G CUGCCAUC 1305 GAUGGCAG GCCGAAAGGCGAGUCAAGGUCU CAUGACAU 3218
1882 GCCUCAUG G UGUGUCAG 1306 CUGACACA GCCGAAAGGCGAGUCAAGGUCU CAUGAGGC 3219
1890 GUGUGUCA G UGGCGCUG 1307 CAGCGCCA GCCGAAAGGCGAGUCAAGGUCU UGACACAC 3220
1893 UGUCAGUG G CGCUGCCU 1308 AGGCAGCG GCCGAAAGGCGAGUCAAGGUCU CACUGACA 3221
1917 CUGCGCCA G CAGCAUGA 1309 UCAUGCUG GCCGAAAGGCGAGUCAAGGUCU UGGCGCAG 3222
Table 21
1920 CGCCAGCA G CAUGAUGA 1310 UCAUCAUG GCCGAAAGGCGAGUCAAGGUCU UGCUGGCG 3223
1956 CUGCUGAA G UGAGGAGG 1311 CCUCCUCA GCCGAAAGGCGAGUCAAGGUCU UUCAGCAG 3224
1964 GUGAGGAG G CCCAUGGG 1312 CCCAUGGG GCCGAAAGGCGAGUCAAGGUCU CUCCUCAC 3225
1972 GCCCAUGG G CAGAAGAU 1313 AUCUUCUG GCCGAAAGGCGAGUCAAGGUCU CCAUGGGC 3226
2006 ACACCUCC G UGGUUCAC 1314 GUGAACCA GCCGAAAGGCGAGUCAAGGUCU GGAGGUGU 3227
2009 CCUCCGUG G UUCACUUU 1315 AAAGUGAA GCCGAAAGGCGAGUCAAGGUCU CACGGAGG 3228
2019 UCACUUUG G UCACAAGU 1316 ACUUGUGA GCCGAAAGGCGAGUCAAGGUCU CAAAGUGA 3229
2026 GGUCACAA G UAGGAGAC 1317 GUCUCCUA GCCGAAAGGCGAGUCAAGGUCU UUGUGACC 3230
2042 CACAGAUG G CACCUGUG 1318 CACAGGUG GCCGAAAGGCGAGUCAAGGUCU CAUCUGUG 3231
2051 CACCUGUG G CCAGAGCA 1319 UGCUCUGG GCCGAAAGGCGAGUCAAGGUCU CACAGGUG 3232
2057 UGGCCAGA G CACCUCAG 1320 CUGAGGUG GCCGAAAGGCGAGUCAAGGUCU UCUGGCCA 3233
2114 AGGAAAAG G CUGGCAAG 1321 CUUGCCAG GCCGAAAGGCGAGUCAAGGUCU CUUUUCCU 3234
2118 AAAGGCUG G CAAGGUGG 1322 CCACCUUG GCCGAAAGGCGAGUCAAGGUCU CAGCCUUU 3235
2123 CUGGCAAG G UGGGUUCC 1323 GGAACCCA GCCGAAAGGCGAGUCAAGGUCU CUUGCCAG 3236
2127 CAAGGUGG G UUCCAGGG 1324 CCCUGGAA GCCGAAAGGCGAGUCAAGGUCU CCACCUUG 3237
2172 AGAAAGAA G CACUCUGC 1325 GCAGAGUG GCCGAAAGGCGAGUCAAGGUCU UUCUUUCU 3238
2183 CUCUGCUG G CGGGAAUA 1326 UAUUCCCG GCCGAAAGGCGAGUCAAGGUCU CAGCAGAG 3239
2198 UACUCUUG G UCACCUCA 1327 UGAGGUGA GCCGAAAGGCGAGUCAAGGUCU CAAGAGUA 3240
2214 AAAUUUAA G UCGGGAAA 1328 UUUCCCGA GCCGAAAGGCGAGUCAAGGUCU UUAAAUUU 3241
2243 AAACUUCA G CCCUGAAC 1329 GUUCAGGG GCCGAAAGGCGAGUCAAGGUCU UGAAGUUU 3242
2288 AACCCAAA G UAUUCUUC 1330 GAAGAAUA GCCGAAAGGCGAGUCAAGGUCU UUUGGGUU 3243
2305 UUUUCUUA G UUUCAGAA 1331 UUCUGAAA GCCGAAAGGCGAGUCAAGGUCU UAAGAAAA 3244
2314 UUUCAGAA G UACUGGCA 1332 UGCCAGUA GCCGAAAGGCGAGUCAAGGUCU UUCUGAAA 3245
2320 AAGUACUG G CAUCACAC 1333 GUGUGAUG GCCGAAAGGCGAGUCAAGGUCU CAGUACUU 3246
2333 ACACGCAG G UUACCUUG 1334 CAAGGUAA GCCGAAAGGCGAGUCAAGGUCU CUGCGUGU 3247
2342 UUACCUUG G CGUGUGUC 1335 GACACACG GCCGAAAGGCGAGUCAAGGUCU CAAGGUAA 3248
2344 ACCUUGGC G UGUGUCCC 1336 GGGACACA GCCGAAAGGCGAGUCAAGGUCU GCCAAGGU 3249
2357 UCCCUGUG G UACCCUGG 1337 CCAGGGUA GCCGAAAGGCGAGUCAAGGUCU CACAGGGA 3250
2365 GUACCCUG G CAGAGAAG 1338 CUUCUCUG GCCGAAAGGCGAGUCAAGGUCU CAGGGUAC 3251
2381 GAGACCAA G CUUGUUUC 1339 GAAACAAG GCCGAAAGGCGAGUCAAGGUCU UUGGUCUC 3252
2397 CCCUGCUG G CCAAAGUC 1340 GACUUUGG GCCGAAAGGCGAGUCAAGGUCU CAGCAGGG 3253
2403 UGGCCAAA G UCAGUAGG 1341 CCUACUGA GCCGAAAGGCGAGUCAAGGUCU UUUGGCCA 3254
2407 CAAAGUCA G UAGGAGAG 1342 CUCUCCUA GCCGAAAGGCGAGUCAAGGUCU UGACUUUG 3255
2424 GAUGCACA G UUUGCUAU 1343 AUAGCAAA GCCGAAAGGCGAGUCAAGGUCU UGUGCAUC 3256
2463 AUAAACAA G CCUAACAU 1344 AUGUUAGG GCCGAAAGGCGAGUCAAGGUCU UUGUUUAU 3257
2474 UAACAUUG G UGCAAAGA 1345 UCUUUGCA GCCGAAAGGCGAGUCAAGGUCU CAAUGUUA 3258
Input Sequence = AF190725. Cut Site = G/ .
Stem Length = 8 . Core Sequence = GCcgaaagGCGaGuCaaGGuCu
AF190725 (Homo sapiens beta-site APP cleaving enzyme (BACE) mRNA; 2526 bp)
Table 22 Table 22: Human BACE DNAzyme and Target Sequence
Pos Substrate Seq ID Ribozyme Rz Seq ID
48 GCUGGAUU A UGGUGGCC 3 GGCCACCA GGCTAGCTACAACGA AATCCAGC 3259
677 GCAGGGCU A CUACGUGG 27 CCACGTAG GGCTAGCTACAACGA AGCCCTGC 3260
680 GGGCUACU A CGUGGAGA 28 TCTCCACG GGCTAGCTACAACGA AGTAGCCC 3261
733 UGGUGGAU A CAGGCAGC 31 GCTGCCTG GGCTAGCTACAACGA ATCCACCA 3262
788 GCAUCGCU A CUACCAGA 38 TCTGGTAG GGCTAGCTACAACGA AGCGATGC 3263
791 UCGCUACU A CCAGAGGC 39 GCCTCTGG GGCTAGCTACAACGA AGTAGCGA 3264
815 CAGCACAU A CCGGGACC 41 GGTCCCGG GGCTAGCTACAACGA ATGTGCTG 3265
839 GGGUGUGU A UGUGCCCU 43 AGGGCACA GGCTAGCTACAACGA ACACACCC 3266
848 UGUGCCCU A CACCCAGG 44 CCTGGGTG GGCTAGCTACAACGA AGGGCACA 3267
1004 GCUGGCCU A UGCUGAGA 58 TCTCAGCA GGCTAGCTACAACGA AGGCCAGC 3268
1171 UUGGAGGU A UCGACCAC 85 GTGGTCGA GGCTAGCTACAACGA ACCTCCAA 3269
1187 CUCGCUGU A CACAGGCA 88 TGCCTGTG GGCTAGCTACAACGA ACAGCGAG 3270
1205 UCUCUGGU A UACACCCA 91 TGGGTGTA GGCTAGCTACAACGA ACCAGAGA 3271
1207 UCUGGUAU A CACCCAUC 92 GATGGGTG GGCTAGCTACAACGA ATACCAGA 3272
1229 GGAGUGGU A UUAUGAGG 94 CCTCATAA GGCTAGCTACAACGA ACCACTCC 3273
1232 GUGGUAUU A UGAGGUGA 96 TCACCTCA GGCTAGCTACAACGA AATACCAC 3274
1295 CAAGGAGU A CAACUAUG 101 CATAGTTG GGCTAGCTACAACGA ACTCCTTG 3275
1301 GUACAACU A UGACAAGA 102 TCTTGTCA GGCTAGCTACAACGA AGTTGTAC 3276
1493 CUCACUCU A CCUAAUGG 130 CCATTAGG GGCTAGCTACAACGA AGAGTGAG 3277
1510 GUGAGGUU A CCAACCAG 133 CTGGTTGG GGCTAGCTACAACGA AACCTCAC 3278
1550 GCAGCAAU A CCUGCGGC 141 GCCGCAGG GGCTAGCTACAACGA ATTGCTGC 3279
1595 CGACUGUU A CAAGUUUG 144 CAAACTTG GGCTAGCTACAACGA AACAGTCG 3280
1633 GCACUGUU A UGGGAGCU 152 AGCTCCCA GGCTAGCTACAACGA AACAGTGC 3281
1645 GAGCUGUU A UCAUGGAG 154 CTCCATGA GGCTAGCTACAACGA AACAGCTC 3282
1661 GGGCUUCU A CGUUGUCU 158 AGACAACG GGCTAGCTACAACGA AGAAGCCC 3283
1787 CUGUGGCU A CAACAUUC 176 GAATGTTG GGCTAGCTACAACGA AGCCACAG 3284
1832 CAUAGCCU A UGUCAUGG 182 CCATGACA GGCTAGCTACAACGA AGGCTATG 3285
2141 GGGACUGU A CCUGUAGG 212 CCTACAGG GGCTAGCTACAACGA ACAGTCCC 3286
2191 GCGGGAAU A CUCUUGGU 215 ACCAAGAG GGCTAGCTACAACGA ATTCCCGC 3287
2290 CCCAAAGU A UUCUUCUU 240 AAGAAGAA GGCTAGCTACAACGA ACTTTGGG 3288
2316 UCAGAAGU A CUGGCAUC 254 GATGCCAG GGCTAGCTACAACGA ACTTCTGA 3289
2336 CGCAGGUU A CCUUGGCG 257 CGCCAAGG GGCTAGCTACAACGA AACCTGCG 3290
2359 CCUGUGGU A CCCUGGCA 260 TGCCAGGG GGCTAGCTACAACGA ACCACAGG 3291
2431 AGUUUGCU A UUUGCUUU 269 AAAGCAAA GGCTAGCTACAACGA AGCAAACT 3292
2455 GGGACUGU A UAAACAAG 275 CTTGTTTA GGCTAGCTACAACGA ACAGTCCC 3293
140 AAGCCGCC A CCGGCCCG 322 CGGGCCGG GGCTAGCTACAACGA GGCGGCTT 3294
151 GGCCCGCC A UGCCCGCC 327 GGCGGGCA GGCTAGCTACAACGA GGCGGGCC 3295
287 CGCUCUCC A CAGCCCGG 380 CCGGGCTG GGCTAGCTACAACGA GGAGAGCG 3296
368 GAGAAGCC A CCAGCACC 412 GGTGCTGG GGCTAGCTACAACGA GGCTTCTC 3297
374 CCACCAGC A CCACCCAG 415 CTGGGTGG GGCTAGCTACAACGA GCTGGTGG 3298
377 CCAGCACC A CCCAGACU 417 AGTCTGGG GGCTAGCTACAACGA GGTGCTGG 3299
451 CGGGGCCC A CCAUGGCC 435 GGCCATGG GGCTAGCTACAACGA GGGCCCCG 3300
454 GGCCCACC A UGGCCCAA 437 TTGGGCCA GGCTAGCTACAACGA GGTGGGCC 3301
512 GCCUGCCC A CGGCACCC 456 GGGTGCCG GGCTAGCTACAACGA GGGCAGGC 3302
517 CCCACGGC A CCCAGCAC 457 GTGCTGGG GGCTAGCTACAACGA GCCGTGGG 3303
524 CACCCAGC A CGGCAUCC 461 GGATGCCG GGCTAGCTACAACGA GCTGGGTG 3304
529 AGCACGGC A UCCGGCUG 462 CAGCCGGA GGCTAGCTACAACGA GCCGTGCT 3305
Table 22
721 CGCUCAAC A UCCUGGUG 508 CACCAGGA GGCTAGCTACAACGA GTTGAGCG 3306
770 UGCCCCCC A CCCCUUCC 522 GGAAGGGG GGCTAGCTACAACGA GGGGGGCA , 3307
782 CUUCCUGC A UCGCUACU 529 AGTAGCGA GGCTAGCTACAACGA GCAGGAAG 3308
811 UGUCCAGC A CAUACCGG 538 CCGGTATG GGCTAGCTACAACGA GCTGGACA 3309
813 UCCAGCAC A UACCGGGA 539 TCCCGGTA GGCTAGCTACAACGA GTGCTGGA 3310
850 UGCCCUAC A CCCAGGGC 547 GCCCTGGG GGCTAGCTACAACGA GTAGGGCA 3311
880 AGCUGGGC A CCGACCUG 553 CAGGTCGG GGCTAGCTACAACGA GCCCAGCT 3312
895 UGGUAAGC A UCCCCCAU 557 ATGGGGGA GGCTAGCTACAACGA GCTTACCA 3313
902 CAUCCCCC A UGGCCCCA 562 TGGGGCCA GGCTAGCTACAACGA GGGGGATG 3314
916 CCAACGUC A CUGUGCGU 567 ACGCACAG GGCTAGCTACAACGA GACGTTGG 3315
931 GUGCCAAC A UUGCUGCC 571 GGCAGCAA GGCTAGCTACAACGA GTTGGCAC 3316
940 UUGCUGCC A UCACUGAA 574 TTCAGTGA GGCTAGCTACAACGA GGCAGCAA 3317
943 CUGCCAUC A CUGAAUCA 575 TGATTCAG GGCTAGCTACAACGA GATGGCAG 3318
964 AGUUCUUC A UCAACGGC 580 GCCGTTGA GGCTAGCTACAACGA GAAGAACT 3319
988 GGGAAGGC A UCCUGGGG 586 CCCCAGGA GGCTAGCTACAACGA GCCTTCCC 3320
1070 GCAGACCC A CGUUCCCA 610 TGGGAACG GGCTAGCTACAACGA GGGTCTGC 3321
1156 GAGGGAGC A UGAUCAUU 638 AATGATCA GGCTAGCTACAACGA GCTCCCTC 3322
1162 GCAUGAUC A UUGGAGGU 639 ACCTCCAA GGCTAGCTACAACGA GATCATGC 3323
1178 UAUCGACC A CUCGCUGU 641 ACAGCGAG GGCTAGCTACAACGA GGTCGATA 3324
1189 CGCUGUAC A CAGGCAGU 644 ACTGCCTG GGCTAGCTACAACGA GTACAGCG 3325
1209 UGGUAUAC A CCCAUCCG 649 CGGATGGG GGCTAGCTACAACGA GTATACCA 3326
1213 AUACACCC A UCCGGCGG 652 CCGCCGGA GGCTAGCTACAACGA GGGTGTAT 3327
1243 AGGUGAUC A UUGUGCGG 654 CCGCACAA GGCTAGCTACAACGA GATCACCT 3328
1312 ACAAGAGC A UUGUGGAC 663 GTCCACAA GGCTAGCTACAACGA GCTCTTGT 3329
1327 ACAGUGGC A CCACCAAC 665 GTTGGTGG GGCTAGCTACAACGA GCCACTGT 3330
1330 GUGGCACC A CCAACCUU 667 AAGGTTGG GGCTAGCTACAACGA GGTGCCAC 3331
1378 UCAAAUCC A UCAAGGCA 679 TGCCTTGA GGCTAGCTACAACGA GGATTTGA 3332
1396 CCUCCUCC A CGGAGAAG 687 CTTCTCCG GGCTAGCTACAACGA GGAGGAGG 3333
1456 AAGCAGGC A CCACCCCU 698 AGGGGTGG GGCTAGCTACAACGA GCCTGCTT 3334
1459 CAGGCACC A CCCCUUGG 700 CCAAGGGG GGCTAGCTACAACGA GGTGCCTG 3335
1471 CUUGGAAC A UUUUCCCA 705 TGGGAAAA GGCTAGCTACAACGA GTTCCAAG 3336
1483 UCCCAGUC A UCUCACUC 709 GAGTGAGA GGCTAGCTACAACGA GACTGGGA 3337
1488 GUCAUCUC A CUCUACCU 711 AGGTAGAG GGCTAGCTACAACGA GAGATGAC 3338
1528 CCUUCCGC A UCACCAUC 723 GATGGTGA GGCTAGCTACAACGA GCGGAAGG 3339
1531 UCCGCAUC A CCAUCCUU 724 AAGGATGG GGCTAGCTACAACGA GATGCGGA 3340
1534 GCAUCACC A UCCUUCCG 726 CGGAAGGA GGCTAGCTACAACGA GGTGATGC 3341
1576 AUGUGGCC A CGUCCCAA 737 TTGGGACG GGCTAGCTACAACGA GGCCACAT 3342
1606 AGUUUGCC A UCUCACAG 744 CTGTGAGA GGCTAGCTACAACGA GGCAAACT 3343
1611 GCCAUCUC A CAGUCAUC 746 GATGACTG GGCTAGCTACAACGA GAGATGGC 3344
1617 UCACAGUC A UCCACGGG 748 CCCGTGGA GGCTAGCTACAACGA GACTGTGA 3345
1621 AGUCAUCC A CGGGCACU 750 AGTGCCCG GGCTAGCTACAACGA GGATGACT 3346
1627 CCACGGGC A CUGUUAUG 751 CATAACAG GGCTAGCTACAACGA GCCCGTGG 3347
1648 CUGUUAUC A UGGAGGGC 754 GCCCTCCA GGCTAGCTACAACGA GATAACAG 3348
1715 CGCUUGCC A UGUGCACG 765 CGTGCACA GGCTAGCTACAACGA GGCAAGCG 3349
1721 CCAUGUGC A CGAUGAGU 766 ACTCATCG GGCTAGCTACAACGA GCACATGG 3350
1762 CUUUUGUC A CCUUGGAC 772 GTCCAAGG GGCTAGCTACAACGA GACAAAAG 3351
1771 CCUUGGAC A UGGAAGAC 775 GTCTTCCA GGCTAGCTACAACGA GTCCAAGG 3352
1792 GCUACAAC A UUCCACAG 779 CTGTGGAA GGCTAGCTACAACGA GTTGTAGC 3353
1797 AACAUUCC A CAGACAGA 781 TCTGTCTG GGCTAGCTACAACGA GGAATGTT 3354
1819 CAACCCUC A UGACCAUA 788 TATGGTCA GGCTAGCTACAACGA GAGGGTTG 3355
1825 UCAUGACC A UAGCCUAU 790 ATAGGCTA GGCTAGCTACAACGA GGTCATGA 3356
Table 22
1837 CCUAUGUC A UGGCUGCC 793 GGCAGCCA GGCTAGCTACAACGA GACATAGG 3357
1846 UGGCUGCC A UCUGCGCC 796 GGCGCAGA GGCTAGCTACAACGA GGCAGCCA 3358
1861 CCCUCUUC A UGCUGCCA 802 TGGCAGCA GGCTAGCTACAACGA GAAGAGGG 3359
1869 AUGCUGCC A CUCUGCCU 805 AGGCAGAG GGCTAGCTACAACGA GGCAGCAT 3360
1879 UCUGCCUC A UGGUGUGU 810 ACACACCA GGCTAGCTACAACGA GAGGCAGA 3361
1922 CCAGCAGC A UGAUGACU 822 AGTCATCA GGCTAGCTACAACGA GCTGCTGG 3362
1942 CUGAUGAC A UCUCCCUG 825 CAGGGAGA GGCTAGCTACAACGA GTCATCAG 3363
1968 GGAGGCCC A UGGGCAGA 833 TCTGCCCA GGCTAGCTACAACGA GGGCCTCC 3364
1998 CCUGGACC A CACCUCCG 840 CGGAGGTG GGCTAGCTACAACGA GGTCCAGG 3365
2000 UGGACCAC A CCUCCGUG 841 CACGGAGG GGCTAGCTACAACGA GTGGTCCA 3366
2013 CGUGGUUC A CUUUGGUC 845 GACCAAAG GGCTAGCTACAACGA GAACCACG 3367
2022 CUUUGGUC A CAAGUAGG 847 CCTACTTG GGCTAGCTACAACGA GACCAAAG 3368
2035 UAGGAGAC A CAGAUGGC 849 GCCATCTG GGCTAGCTACAACGA GTCTCCTA 3369
2044 CAGAUGGC A CCUGUGGC 851 GCCACAGG GGCTAGCTACAACGA GCCATCTG 3370
2059 GCCAGAGC A CCUCAGGA 856 TCCTGAGG GGCTAGCTACAACGA GCTCTGGC 3371
2076 CCCUCCCC A CCCACCAA 866 TTGGTGGG GGCTAGCTACAACGA GGGGAGGG 3372
2080 CCCCACCC A CCAAAUGC 869 GCATTTGG GGCTAGCTACAACGA GGGTGGGG 3373
2174 AAAGAAGC A CUCUGCUG 885 CAGCAGAG GGCTAGCTACAACGA GCTTCTTT 3374
2201 UCUUGGUC A CCUCAAAU 891 ATTTGAGG GGCTAGCTACAACGA GACCAAGA 3375
2260 CUUUGUCC A CCAUUCCU 906 AGGAATGG GGCTAGCTACAACGA GGACAAAG 3376
2263 UGUCCACC A UUCCUUUA 908 TAAAGGAA GGCTAGCTACAACGA GGTGGACA 3377
2322 GUACUGGC A UCACACGC 922 GCGTGTGA GGCTAGCTACAACGA GCCAGTAC 3378
2325 CUGGCAUC A CACGCAGG 923 CCTGCGTG GGCTAGCTACAACGA GATGCCAG 3379
2327 GGCAUCAC A CGCAGGUU 924 AACCTGCG GGCTAGCTACAACGA GTGATGCC 3380
2421 GAGGAUGC A CAGUUUGC 945 GCAAACTG GGCTAGCTACAACGA GCATCCTC 3381
2470 AGCCUAAC A UUGGUGCA 954 TGCACCAA GGCTAGCTACAACGA GTTAGGCT 3382
11 ACGCGUCC G CAGCCCGC 960 GCGGGCTG GGCTAGCTACAACGA GGACGCGT 3383
18 CGCAGCCC G CCCGGGAG 961 CTCCCGGG GGCTAGCTACAACGA GGGCTGCG 3384
29 CGGGAGCU G CGAGCCGC 962 GCGGCTCG GGCTAGCTACAACGA AGCTCCCG 3385
36 UGCGAGCC G CGAGCUGG 964 CCAGCTCG GGCTAGCTACAACGA GGCTCGCA 3386
69 CAGCCAAC G CAGCCGCA 967 TGCGGCTG GGCTAGCTACAACGA GTTGGCTG 3387
75 ACGCAGCC G CAGGAGCC 968 GGCTCCTG GGCTAGCTACAACGA GGCTGCGT 3388
94 GAGCCCUU G CCCCUGCC 969 GGCAGGGG GGCTAGCTACAACGA AAGGGCTC 3389
100 UUGCCCCU G CCCGCGCC 970 GGCGCGGG GGCTAGCTACAACGA AGGGGCAA 3390
104 CCCUGCCC G CGCCGCCG 971 CGGCGGCG GGCTAGCTACAACGA GGGCAGGG 3391
106 CUGCCCGC G CCGCCGCC 972 GGCGGCGG GGCTAGCTACAACGA GCGGGCAG 3392
109 CCCGCGCC G CCGCCCGC 973 GCGGGCGG GGCTAGCTACAACGA GGCGCGGG 3393
112 GCGCCGCC G CCCGCCGG 974 CCGGCGGG GGCTAGCTACAACGA GGCGGCGC 3394
116 CGCCGCCC G CCGGGGGG 975 CCCCCCGG GGCTAGCTACAACGA GGGCGGCG 3395
137 GGGAAGCC G CCACCGGC 976 GCCGGTGG GGCTAGCTACAACGA GGCTTCCC 3396
148 ACCGGCCC G CCAUGCCC 977 GGGCATGG GGCTAGCTACAACGA GGGCCGGT 3397
153 CCCGCCAU G CCCGCCCC 978 GGGGCGGG GGCTAGCTACAACGA ATGGCGGG 3398
157 CCAUGCCC G CCCCUCCC 979 GGGAGGGG GGCTAGCTACAACGA GGGCATGG 3399
172 CCAGCCCC G CCGGGAGC 980 GCTCCCGG GGCTAGCTACAACGA GGGGCTGG 3400
183 GGGAGCCC G CGCCCGCU 981 AGCGGGCG GGCTAGCTACAACGA GGGCTCCC 3401
185 GAGCCCGC G CCCGCUGC 982 GCAGCGGG GGCTAGCTACAACGA GCGGGCTC 3402
189 CCGCGCCC G CUGCCCAG 983 CTGGGCAG GGCTAGCTACAACGA GGGCGCGG 3403
192 CGCCCGCU G CCCAGGCU 984 AGCCTGGG GGCTAGCTACAACGA AGCGGGCG 3404
205 GGCUGGCC G CCGCCGUG 985 CACGGCGG GGCTAGCTACAACGA GGCCAGCC 3405
208 UGGCCGCC G CCGUGCCG 986 CGGCACGG GGCTAGCTACAACGA GGCGGCCA 3406
213 GCCGCCGU G CCGAUGUA 987 TACATCGG GGCTAGCTACAACGA ACGGCGGC 3407
Table 22
250 UCUCCCCU G CUCCCGUG 989 CACGGGAG GGCTAGCTACAACGA AGGGGAGA 3408
258 GCUCCCGU G CUCUGCGG 990 CCGCAGAG GGCTAGCTACAACGA ACGGGAGC 3409
263 CGUGCUCU G CGGAUCUC 991 GAGATCCG GGCTAGCTACAACGA AGAGCACG 3410
280 CCCUGACC G CUCUCCAC 993 GTGGAGAG GGCTAGCTACAACGA GGTCAGGG 3411
320 AGGGCCCU G CAGGCCCU 994 AGGGCCTG GGCTAGCTACAACGA AGGGCCCT 3412
340 GUCCUGAU G CCCCCAAG 996 CTTGGGGG GGCTAGCTACAACGA ATCAGGAC 3413
397 GGGCAGGC G CCAGGGAC 998 GTCCCTGG GGCTAGCTACAACGA GCCTGCCC 3414
420 GGGCCAGU G CGAGCCCA 999 TGGGCTCG GGCTAGCTACAACGA ACTGGCCC 3415
468 CAAGCCCU G CCCUGGCU 1002 AGCCAGGG GGCTAGCTACAACGA AGGGCTTG 3416
480 UGGCUCCU G CUGUGGAU 1003 ATCCACAG GGCTAGCTACAACGA AGGAGCCA 3417
493 GGAUGGGC G CGGGAGUG 1004 CACTCCCG GGCTAGCTACAACGA GCCCATCC 3418
501 GCGGGAGU G CUGCCUGC 1005 GCAGGCAG GGCTAGCTACAACGA ACTCCCGC 3419
504 GGAGUGCU G CCUGCCCA 1006 TGGGCAGG GGCTAGCTACAACGA AGCACTCC 3420
508 UGCUGCCU G CCCACGGC 1007 GCCGTGGG GGCTAGCTACAACGA AGGCAGCA 3421
537 AUCCGGCU G CCCCUGCG 1008 CGCAGGGG GGCTAGCTACAACGA AGCCGGAT 3422
543 CUGCCCCU G CGCAGCGG 1009 CCGCTGCG GGCTAGCTACAACGA AGGGGCAG 3423
545 GCCCCUGC G CAGCGGCC 1010 GGCCGCTG GGCTAGCTACAACGA GCAGGGGC 3424
562 UGGGGGGC G CCCCCCUG 1011 CAGGGGGG GGCTAGCTACAACGA GCCCCCCA 3425
576 CUGGGGCU G CGGCUGCC 1012 GGCAGCCG GGCTAGCTACAACGA AGCCCCAG 3426
582 CUGCGGCU G CCCCGGGA 1013 TCCCGGGG GGCTAGCTACAACGA AGCCGCAG 3427
708 AGCCCCCC G CAGACGCU 1019 AGCGTCTG GGCTAGCTACAACGA GGGGGGCT 3428
714 CCGCAGAC G CUCAACAU 1020 ATGTTGAG GGCTAGCTACAACGA GTCTGCGG 3429
751 GUAACUUU G CAGUGGGU 1021 ACCCACTG GGCTAGCTACAACGA AAAGTTAC 3430
760 CAGUGGGU G CUGCCCCC 1022 GGGGGCAG GGCTAGCTACAACGA ACCCACTG 3431
763 UGGGUGCU G CCCCCCAC 1023 GTGGGGGG GGCTAGCTACAACGA AGCACCCA 3432
780 CCCUUCCU G CAUCGCUA 1024 TAGCGATG GGCTAGCTACAACGA AGGAAGGG 3433
785 CCUGCAUC G CUACUACC 1025 GGTAGTAG GGCTAGCTACAACGA GATGCAGG 3434
843 GUGUAUGU G CCCUACAC 1026 GTGTAGGG GGCTAGCTACAACGA ACATACAC 3435
921 GUCACUGU G CGUGCCAA 1028 TTGGCACG GGCTAGCTACAACGA ACAGTGAC 3436
925 CUGUGCGU G CCAACAUU 1029 AATGTTGG GGCTAGCTACAACGA ACGCACAG 3437
934 CCAACAUU G CUGCCAUC 1030 GATGGCAG GGCTAGCTACAACGA AATGTTGG 3438
937 ACAUUGCU G CCAUCACU 1031 AGTGATGG GGCTAGCTACAACGA AGCAATGT 3439
1006 UGGCCUAU G CUGAGAUU 1033 AATCTCAG GGCTAGCTACAACGA ATAGGCCA 3440
1015 CUGAGAUU G CCAGGCCU 1035 AGGCCTGG GGCTAGCTACAACGA AATCTCAG 3441
1092 UUCUCCCU G CAGCUUUG 1039 CAAAGCTG GGCTAGCTACAACGA AGGGAGAA 3442
1105 UUUGUGGU G CUGGCUUC 1040 GAAGCCAG GGCTAGCTACAACGA ACCACAAA 3443
1134 UCUGAAGU G CUGGCCUC 1042 GAGGCCAG GGCTAGCTACAACGA ACTTCAGA 3444
1182 GACCACUC G CUGUACAC 1045 GTGTACAG GGCTAGCTACAACGA GAGTGGTC 3445
1248 AUCAUUGU G CGGGUGGA 1048 TCCACCCG GGCTAGCTACAACGA ACAATGAT 3446
1286 AAUGGACU G CAAGGAGU 1050 ACTCCTTG GGCTAGCTACAACGA AGTCCATT 3447
1344 CUUCGUUU G CCCAAGAA 1052 TTCTTGGG GGCTAGCTACAACGA AAACGAAG 3448
1366 UUGAAGCU G CAGUCAAA 1054 TTTGACTG GGCTAGCTACAACGA AGCTTCAA 3449
1442 GCUGGUGU G CUGGCAAG 1056 CTTGCCAG GGCTAGCTACAACGA ACACCAGC 3450
1526 GUCCUUCC G CAUCACCA 1058 TGGTGATG GGCTAGCTACAACGA GGAAGGAC 3451
1542 AUCCUUCC G CAGCAAUA 1059 TATTGCTG GGCTAGCTACAACGA GGAAGGAT 3452
1554 CAAUACCU G CGGCCAGU 1060 ACTGGCCG GGCTAGCTACAACGA AGGTATTG 3453
1603 ACAAGUUU G CCAUCUCA 1062 TGAGATGG GGCTAGCTACAACGA AAACTTGT 3454
1699 UUGGCUUU G CUGUCAGC 1066 GCTGACAG GGCTAGCTACAACGA AAAGCCAA 3455
1708 CUGUCAGC G CUUGCCAU 1067 ATGGCAAG GGCTAGCTACAACGA GCTGACAG 3456
1712 CAGCGCUU G CCAUGUGC 1068 GCACATGG GGCTAGCTACAACGA AAGCGCTG 3457
1719 UGCCAUGU G CACGAUGA 1069 TCATCGTG GGCTAGCTACAACGA ACATGGCA 3458
Table 22
1843 UCAUGGCU G CCAUCUGC 1074 GCAGATGG GGCTAGCTACAACGA AGCCATGA 3459
1850 UGCCAUCU G CGCCCUCU 1075 AGAGGGCG GGCTAGCTACAACGA AGATGGCA 3460
1852 CCAUCUGC G CCCUCUUC 1076 GAAGAGGG GGCTAGCTACAACGA GCAGATGG 3461
1863 CUCUUCAU G CUGCCACU 1077 AGTGGCAG GGCTAGCTACAACGA ATGAAGAG 3462
1866 UUCAUGCU G CCACUCUG 1078 CAGAGTGG GGCTAGCTACAACGA AGCATGAA 3463
1874 GCCACUCU G CCUCAUGG 1079 CCATGAGG GGCTAGCTACAACGA AGAGTGGC 3464
1895 UCAGUGGC G CUGCCUCC 1080 GGAGGCAG GGCTAGCTACAACGA GCCACTGA 3465
1898 GUGGCGCU G CCUCCGCU 1081 AGCGGAGG GGCTAGCTACAACGA AGCGCCAC 3466
1904 CUGCCUCC G CUGCCUGC 1082 GCAGGCAG GGCTAGCTACAACGA GGAGGCAG 3467
1907 CCUCCGCU G CCUGCGCC 1083 GGCGCAGG GGCTAGCTACAACGA AGCGGAGG 3468
1911 CGCUGCCU G CGCCAGCA 1084 TGCTGGCG GGCTAGCTACAACGA AGGCAGCG 3469
1913 CUGCCUGC G CCAGCAGC 1085 GCTGCTGG GGCTAGCTACAACGA GCAGGCAG 3470
1933 AUGACUUU G CUGAUGAC 1088 GTCATCAG GGCTAGCTACAACGA AAAGTCAT 3471
1950 AUCUCCCU G CUGAAGUG 1091 CACTTCAG GGCTAGCTACAACGA AGGGAGAT 3472
2087 CACCAAAU G CCUCUGCC 1094 GGCAGAGG GGCTAGCTACAACGA ATTTGGTG 3473
2093 AUGCCUCU G CCUUGAUG 1095 CATCAAGG GGCTAGCTACAACGA AGAGGCAT 3474
2179 AGCACUCU G CUGGCGGG 1097 CCCGCCAG GGCTAGCTACAACGA AGAGTGCT 3475
2227 GAAAUUCU G CUGCUUGA 1098 TCAAGCAG GGCTAGCTACAACGA AGAATTTC 3476
2230 AUUCUGCU σ CUUGAAAC 1099 GTTTCAAG GGCTAGCTACAACGA AGCAGAAT 3477
2329 CAUCACAC G CAGGUUAC 1102 GTAACCTG GGCTAGCTACAACGA GTGTGATG 3478
2393 GUUUCCCU G CUGGCCAA 1103 TTGGCCAG GGCTAGCTACAACGA AGGGAAAC 3479
2419 GAGAGGAU G CACAGUUU 1104 AAACTGTG GGCTAGCTACAACGA ATCCTCTC 3480
2428 CACAGUUU G CUAUUUGC 1105 GCAAATAG GGCTAGCTACAACGA AAACTGTG 3481
2435 UGCUAUUU G CUUUAGAG 1106 CTCTAAAG GGCTAGCTACAACGA AAATAGCA 3482
2476 ACAUUGGU G CAAAGAUU 1107 AATCTTTG GGCTAGCTACAACGA ACCAATGT 3483
2485 CAAAGAUU G CCUCUUGA 1108 TCAAGAGG GGCTAGCTACAACGA AATCTTTG 3484
219 GUGCCGAU G UAGCGGGC 1110 GCCCGCTA GGCTAGCTACAACGA ATCGGCAC 3485
483 CUCCUGCU G UGGAUGGG llll CCCATCCA GGCTAGCTACAACGA AGCAGGAG 3486
634 GCAGCUUU G UGGAGAUG 1112 CATCTCCA GGCTAGCTACAACGA AAAGCTGC 3487
804 AGGCAGCU G UCCAGCAC 1113 GTGCTGGA GGCTAGCTACAACGA AGCTGCCT 3488
835 GGAAGGGU G UGUAUGUG 1114 CACATACA GGCTAGCTACAACGA ACCCTTCC 3489
837 AAGGGUGU G UAUGUGCC 1115 GGCACATA GGCTAGCTACAACGA ACACCCTT 3490
841 GUGUGUAU G UGCCCUAC 1116 GTAGGGCA GGCTAGCTACAACGA ATACACAC 3491
919 ACGUCACU G UGCGUGCC 1117 GGCACGCA GGCTAGCTACAACGA AGTGACGT 3492
1100 GCAGCUUU G UGGUGCUG 1118 CAGCACCA GGCTAGCTACAACGA AAAGCTGC 3493
1144 UGGCCUCU G UCGGAGGG 1119 CCCTCCGA GGCTAGCTACAACGA AGAGGCCA 3494
1185 CACUCGCU G UACACAGG 1120 CCTGTGTA GGCTAGCTACAACGA AGCGAGTG 3495
1246 UGAUCAUU G UGCGGGUG 1121 CACCCGCA GGCTAGCTACAACGA AATGATCA 3496
1315 AGAGCAUU G UGGACAGU 1122 ACTGTCCA GGCTAGCTACAACGA AATGCTCT 3497
1356 AAGAAAGU G UUUGAAGC 1123 GCTTCAAA GGCTAGCTACAACGA ACTTTCTT 3498
1440 CAGCUGGU G UGCUGGCA 1124 TGCCAGCA GGCTAGCTACAACGA ACCAGCTG 3499
1570 UGGAAGAU G UGGCCACG 1125 CGTGGCCA GGCTAGCTACAACGA ATCTTCCA 3500
1592 AGACGACU G UUACAAGU 1126 ACTTGTAA GGCTAGCTACAACGA AGTCGTCT 3501
1630 CGGGCACU G UUAUGGGA 1127 TCCCATAA GGCTAGCTACAACGA AGTGCCCG 3502
1642 UGGGAGCU G UUAUCAUG 1128 CATGATAA GGCTAGCTACAACGA AGCTCCCA 3503
1666 UCUACGUU G UCUUUGAU 1129 ATCAAAGA GGCTAGCTACAACGA AACGTAGA 3504
1702 GCUUUGCU G UCAGCGCU 1130 AGCGCTGA GGCTAGCTACAACGA AGCAAAGC 3505
1717 CUUGCCAU G UGCACGAU 1131 ATCGTGCA GGCTAGCTACAACGA ATGGCAAG 3506
1759 GCCCUUUU G UCACCUUG 1132 CAAGGTGA GGCTAGCTACAACGA AAAAGGGC 3507
1781 GGAAGACU G UGGCUACA 1133 TGTAGCCA GGCTAGCTACAACGA AGTCTTCC 3508
1834 UAGCCUAU G UCAUGGCU 1134 AGCCATGA GGCTAGCTACAACGA ATAGGCTA 3509
Table 22
1884 CUCAUGGU G UGUCAGUG 1135 CACTGACA GGCTAGCTACAACGA ACCATGAG 3510
1886 CAUGGUGU G UCAGUGGC 1136 GCCACTGA GGCTAGCTACAACGA ACACCATG 3511
2048 UGGCACCU G UGGCCAGA 1137 TCTGGCCA GGCTAGCTACAACGA AGGTGCCA 3512
2139 CAGGGACU G UACCUGUA 1138 TACAGGTA GGCTAGCTACAACGA AGTCCCTG 3513
2145 CUGUACCU G UAGGAAAC 1139 GTTTCCTA GGCTAGCTACAACGA AGGTACAG 3514
2256 GAACCUUU G UCCACCAU 1140 ATGGTGGA GGCTAGCTACAACGA AAAGGTTC 3515
2346 CUUGGCGU G UGUCCCUG 1141 CAGGGACA GGCTAGCTACAACGA ACGCCAAG 3516
2348 UGGCGUGU G UCCCUGUG 1142 CACAGGGA GGCTAGCTACAACGA ACACGCCA 3517
2354 GUGUCCCU G UGGUACCC 1143 GGGTACCA GGCTAGCTACAACGA AGGGACAC 3518
2385 CCAAGCUU G UUUCCCUG 1144 CAGGGAAA GGCTAGCTACAACGA AAGCTTGG 3519
2453 CAGGGACU G UAUAAACA 1145 TGTTTATA GGCTAGCTACAACGA AGTCCCTG 3520
14 CGUCCGCA G CCCGCCCG 1146 CGGGCGGG GGCTAGCTACAACGA TGCGGACG 3521
26 GCCCGGGA G CUGCGAGC 1147 GCTCGCAG GGCTAGCTACAACGA TCCCGGGC 3522
33 AGCUGCGA G CCGCGAGC 1148 GCTCGCGG GGCTAGCTACAACGA TCGCAGCT 3523
40 AGCCGCGA G CUGGAUUA 1149 TAATCCAG GGCTAGCTACAACGA TCGCGGCT 3524
51 GGAUUAUG G UGGCCUGA 1150 TCAGGCCA GGCTAGCTACAACGA CATAATCC 3525
54 UUAUGGUG G CCUGAGCA 1151 TGCTCAGG GGCTAGCTACAACGA CACCATAA 3526
60 UGGCCUGA G CAGCCAAC 1152 GTTGGCTG GGCTAGCTACAACGA TCAGGCCA 3527
63 CCUGAGCA G CCAACGCA 1153 TGCGTTGG GGCTAGCTACAACGA TGCTCAGG 3528
72 CCAACGCA G CCGCAGGA 1154 TCCTGCGG GGCTAGCTACAACGA TGCGTTGG 3529
81 CCGCAGGA G CCCGGAGC 1155 GCTCCGGG GGCTAGCTACAACGA TCCTGCGG 3530
88 AGCCCGGA G CCCUUGCC 1156 GGCAAGGG GGCTAGCTACAACGA TCCGGGCT 3531
134 CCAGGGAA G CCGCCACC 1157 GGTGGCGG GGCTAGCTACAACGA TTCCCTGG 3532
144 CGCCACCG G CCCGCCAU 1158 ATGGCGGG GGCTAGCTACAACGA CGGTGGCG 3533
167 CCCUCCCA G CCCCGCCG 1159 CGGCGGGG GGCTAGCTACAACGA TGGGAGGG 3534
179 CGCCGGGA G CCCGCGCC 1160 GGCGCGGG GGCTAGCTACAACGA TCCCGGCG 3535
198 CUGCCCAG G CUGGCCGC 1161 GCGGCCAG GGCTAGCTACAACGA CTGGGCAG 3536
202 CCAGGCUG G CCGCCGCC 1162 GGCGGCGG GGCTAGCTACAACGA CAGCCTGG 3537
211 CCGCCGCC G UGCCGAUG 1163 CATCGGCA GGCTAGCTACAACGA GGCGGCGG 3538
222 CCGAUGUA G CGGGCUCC 1164 GGAGCCCG GGCTAGCTACAACGA TACATCGG 3539
226 UGUAGCGG G CUCCGGAU 1165 ATCCGGAG GGCTAGCTACAACGA CCGCTACA 3540
239 GGAUCCCA G CCUCUCCC 1166 GGGAGAGG GGCTAGCTACAACGA TGGGATCC 3541
256 CUGCUCCC G UGCUCUGC 1167 GCAGAGCA GGCTAGCTACAACGA GGGAGCAG 3542
290 UCUCCACA G CCCGGACC 1168 GGTCCGGG GGCTAGCTACAACGA TGTGGAGA 3543
304 ACCCGGGG G CUGGCCCA 1169 TGGGCCAG GGCTAGCTACAACGA CCCCGGGT 3544
308 GGGGGCUG G CCCAGGGC 1170 GCCCTGGG GGCTAGCTACAACGA CAGCCCCC 3545
315 GGCCCAGG G CCCUGCAG 1171 CTGCAGGG GGCTAGCTACAACGA CCTGGGCC 3546
324 CCCUGCAG G CCCUGGCG 1172 CGCCAGGG GGCTAGCTACAACGA CTGCAGGG 3547
330 AGGCCCUG G CGUCCUGA 1173 TCAGGACG GGCTAGCTACAACGA CAGGGCCT 3548
332 GCCCUGGC G UCCUGAUG 1174 CATCAGGA GGCTAGCTACAACGA GCCAGGGC 3549
348 GCCCCCAA G CUCCCUCU 1175 AGAGGGAG GGCTAGCTACAACGA TTGGGGGC 3550
365 CCUGAGAA G CCACCAGC 1176 GCTGGTGG GGCTAGCTACAACGA TTCTCAGG 3551
372 AGCCACCA G CACCACCC 1177 GGGTGGTG GGCTAGCTACAACGA TGGTGGCT 3552
391 ACUUGGGG G CAGGCGCC 1178 GGCGCCTG GGCTAGCTACAACGA CCCCAAGT 3553
395 GGGGGCAG G CGCCAGGG 1179 CCCTGGCG GGCTAGCTACAACGA CTGCCCCC 3554
410 GGACGGAC G UGGGCCAG 1180 CTGGCCCA GGCTAGCTACAACGA GTCCGTCC 3555
414 GGACGUGG G CCAGUGCG 1181 CGCACTGG GGCTAGCTACAACGA CCACGTCC 3556
418 GUGGGCCA G UGCGAGCC 1182 GGCTCGCA GGCTAGCTACAACGA TGGCCCAC 3557
424 CAGUGCGA G CCCAGAGG 1183 CCTCTGGG GGCTAGCTACAACGA TCGCACTG 3558
433 CCCAGAGG G CCCGAAGG 1184 CCTTCGGG GGCTAGCTACAACGA CCTCTGGG 3559
441 GCCCGAAG G CCGGGGCC 1185 GGCCCCGG GGCTAGCTACAACGA CTTCGGGC 3560
Table 22
447 AGGCCGGG G CCCACCAU 1186 ATGGTGGG GGCTAGCTACAACGA CCCGGCCT 3561
457 CCACCAUG G CCCAAGCC 1187 GGCTTGGG GGCTAGCTACAACGA CATGGTGG 3562
463 UGGCCCAA G CCCUGCCC 1188 GGGCAGGG GGCTAGCTACAACGA TTGGGCCA 3563
474 CUGCCCUG G CUCCUGCU 1189 AGCAGGAG GGCTAGCTACAACGA CAGGGCAG 3564
491 GUGGAUGG G CGCGGGAG 1190 CTCCCGCG GGCTAGCTACAACGA CCATCCAC 3565
499 GCGCGGGA G UGCUGCCU 1191 AGGCAGCA GGCTAGCTACAACGA TCCCGCGC 3566
515 UGCCCACG G CACCCAGC 1192 GCTGGGTG GGCTAGCTACAACGA CGTGGGCA 3567
522 GGCACCCA G CACGGCAU 1193 ATGCCGTG GGCTAGCTACAACGA TGGGTGCC 3568
527 CCAGCACG G CAUCCGGC 1194 GCCGGATG GGCTAGCTACAACGA CGTGCTGG 3569
534 GGCAUCCG G CUGCCCCU 1195 AGGGGCAG GGCTAGCTACAACGA CGGATGCC 3570
548 CCUGCGCA G CGGCCUGG 1196 CCAGGCCG GGCTAGCTACAACGA TGCGCAGG 3571
551 GCGCAGCG G CCUGGGGG 1197 CCCCCAGG GGCTAGCTACAACGA CGCTGCGC 3572
560 CCUGGGGG G CGCCCCCC 1198 GGGGGGCG GGCTAGCTACAACGA CCCCCAGG 3573
573 CCCCUGGG G CUGCGGCU 1199 AGCCGCAG GGCTAGCTACAACGA CCCAGGGG 3574
579 GGGCUGCG G CUGCCCCG 1200 CGGGGCAG GGCTAGCTACAACGA CGCAGCCC 3575
603 GACGAAGA G CCCGAGGA 1201 TCCTCGGG GGCTAGCTACAACGA TCTTCGTC 3576
612 CCCGAGGA G CCCGGCCG 1202 CGGCCGGG GGCTAGCTACAACGA TCCTCGGG 3577
617 GGAGCCCG G CCGGAGGG 1203 CCCTCCGG GGCTAGCTACAACGA CGGGCTCC 3578
626 CCGGAGGG G CAGCUUUG 1204 CAAAGCTG GGCTAGCTACAACGA CCCTCCGG 3579
629 GAGGGGCA G CUUUGUGG 1205 CCACAAAG GGCTAGCTACAACGA TGCCCCTC 3580
643 UGGAGAUG G UGGACAAC 1206 GTTGTCCA GGCTAGCTACAACGA CATCTCCA 3581
659 CCUGAGGG G CAAGUCGG 1207 CCGACTTG GGCTAGCTACAACGA CCCTCAGG 3582
663 AGGGGCAA G UCGGGGCA 1208 TGCCCCGA GGCTAGCTACAACGA TTGCCCCT 3583
669 AAGUCGGG G CAGGGCUA 1209 TAGCCCTG GGCTAGCTACAACGA CCCGACTT 3584
674 GGGGCAGG G CUACUACG 1210 CGTAGTAG GGCTAGCTACAACGA CCTGCCCC 3585
682 GCUACUAC G UGGAGAUG 1211 CATCTCCA GGCTAGCTACAACGA GTAGTAGC 3586
694 AGAUGACC G UGGGCAGC 1212 GCTGCCCA GGCTAGCTACAACGA GGTCATCT 3587
698 GACCGUGG G CAGCCCCC 1213 GGGGGCTG GGCTAGCTACAACGA CCACGGTC 3588
701 CGUGGGCA G CCCCCCGC 1214 GCGGGGGG GGCTAGCTACAACGA TGCCCACG 3589
727 ACAUCCUG G UGGAUACA 1215 TGTATCCA GGCTAGCTACAACGA CAGGATGT 3590
737 GGAUACAG G CAGCAGUA 1216 TACTGCTG GGCTAGCTACAACGA CTGTATCC 3591
740 UACAGGCA G CAGUAACU 1217 AGTTACTG GGCTAGCTACAACGA TGCCTGTA 3592
743 AGGCAGCA G UAACUUUG 1218 CAAAGTTA GGCTAGCTACAACGA TGCTGCCT 3593
754 ACUUUGCA G UGGGUGCU 1219 AGCACCCA GGCTAGCTACAACGA TGCAAAGT 3594
758 UGCAGUGG G UGCUGCCC 1220 GGGCAGCA GGCTAGCTACAACGA CCACTGCA 3595
798 UACCAGAG G CAGCUGUC 1221 GACAGCTG GGCTAGCTACAACGA CTCTGGTA ' 3596
801 CAGAGGCA G CUGUCCAG 1222 CTGGACAG GGCTAGCTACAACGA TGCCTCTG 3597
809 GCUGUCCA G CACAUACC 1223 GGTATGTG GGCTAGCTACAACGA TGGACAGC 3598
833 CCGGAAGG G UGUGUAUG 1224 CATACACA GGCTAGCTACAACGA CCTTCCGG 3599
857 CACCCAGG G CAAGUGGG 1225 CCCACTTG GGCTAGCTACAACGA CCTGGGTG 3600
861 CAGGGCAA G UGGGAAGG 1226 CCTTCCCA GGCTAGCTACAACGA TTGCCCTG 3601
873 GAAGGGGA G CUGGGCAC 1227 GTGCCCAG GGCTAGCTACAACGA TCCCCTTC 3602
878 GGAGCUGG G CACCGACC 1228 GGTCGGTG GGCTAGCTACAACGA CCAGCTCC 3603
889 CCGACCUG G UAAGCAUC 1229 GATGCTTA GGCTAGCTACAACGA CAGGTCGG 3604
893 CCUGGUAA G CAUCCCCC 1230 GGGGGATG GGCTAGCTACAACGA TTACCAGG 3605
905 CCCCCAUG G CCCCAACG 1231 CGTTGGGG GGCTAGCTACAACGA CATGGGGG 3606
913 GCCCCAAC G UCACUGUG 1232 CACAGTGA GGCTAGCTACAACGA GTTGGGGC 3607
923 CACUGUGC G UGCCAACA 1233 TGTTGGCA GGCTAGCTACAACGA GCACAGTG 3608
957 UCAGACAA G UUCUUCAU 1234 ATGAAGAA GGCTAGCTACAACGA TTGTCTGA 3609
971 CAUCAACG G CUCCAACU 1235 AGTTGGAG GGCTAGCTACAACGA CGTTGATG 3610
986 CUGGGAAG G CAUCCUGG 1236 CCAGGATG GGCTAGCTACAACGA CTTCCCAG 3611
Table 22
996 AUCCUGGG G CUGGCCUA 1237 TAGGCCAG GGCTAGCTACAACGA CCCAGGAT 3612
1000 UGGGGCUG G CCUAUGCU 1238 AGCATAGG GGCTAGCTACAACGA CAGCCCCA 3613
1020 AUUGCCAG G CCUGACGA 1239 TCGTCAGG GGCTAGCTACAACGA CTGGCAAT 3614
1038 UCCCUGGA G CCUUUCUU 1240 AAGAAAGG GGCTAGCTACAACGA TCCAGGGA 3615
1057 ACUCUCUG G UAAAGCAG 1241 CTGCTTTA GGCTAGCTACAACGA CAGAGAGT 3616
1062 CUGGUAAA G CAGACCCA 1242 TGGGTCTG GGCTAGCTACAACGA TTTACCAG 3617
1072 AGACCCAC G UUCCCAAC 1243 GTTGGGAA GGCTAGCTACAACGA GTGGGTCT 3618
1095 UCCCUGCA G CUUUGUGG 1244 CCACAAAG GGCTAGCTACAACGA TGCAGGGA 3619
1103 GCUUUGUG G UGCUGGCU 1245 AGCCAGCA GGCTAGCTACAACGA CACAAAGC 3620
1109 UGGUGCUG G CUUCCCCC 1246 GGGGGAAG GGCTAGCTACAACGA CAGCACCA 3621
1125 CUCAACCA G UCUGAAGU 1247 ACTTCAGA GGCTAGCTACAACGA TGGTTGAG 3622
1132 AGUCUGAA G UGCUGGCC 1248 GGCCAGCA GGCTAGCTACAACGA TTCAGACT 3623
1138 AAGUGCUG G CCUCUGUC 1249 GACAGAGG GGCTAGCTACAACGA CAGCACTT 3624
1154 CGGAGGGA G CAUGAUCA 1250 TGATCATG GGCTAGCTACAACGA TCCCTCCG 3625
1169 CAUUGGAG G UAUCGACC 1251 GGTCGATA GGCTAGCTACAACGA CTCCAATG .3626
1193 GUACACAG G CAGUCUCU 1252 AGAGACTG GGCTAGCTACAACGA CTGTGTAC 3627
1196 CACAGGCA G UCUCUGGU 1253 ACCAGAGA GGCTAGCTACAACGA TGCCTGTG 3628
1203 AGUCUCUG G UAUACACC 1254 GGTGTATA GGCTAGCTACAACGA CAGAGACT 3629
1218 CCCAUCCG G CGGGAGUG 1255 CACTCCCG GGCTAGCTACAACGA CGGATGGG 3630
1224 CGGCGGGA G UGGUAUUA 1256 TAATACCA GGCTAGCTACAACGA TCCCGCCG 3631
1227 CGGGAGUG G UAUUAUGA 1257 TCATAATA GGCTAGCTACAACGA CACTCCCG 3632
1237 AUUAUGAG G UGAUCAUU 1258 AATGATCA GGCTAGCTACAACGA CTCATAAT 3633
1252 UUGUGCGG G UGGAGAUC 1259 GATCTCCA GGCTAGCTACAACGA CCGCACAA 3634
1293 UGCAAGGA G UACAACUA 1260 TAGTTGTA GGCTAGCTACAACGA TCCTTGCA 3635
1310 UGACAAGA G CAUUGUGG 1261 CCACAATG GGCTAGCTACAACGA TCTTGTCA 3636
1322 UGUGGACA G UGGCACCA 1262 TGGTGCCA GGCTAGCTACAACGA TGTCCACA 3637
1325 GGACAGUG G CACCACCA 1263 TGGTGGTG GGCTAGCTACAACGA CACTGTCC 3638
1340 CAACCUUC G UUUGCCCA 1264 TGGGCAAA GGCTAGCTACAACGA GAAGGTTG 3639
1354 CCAAGAAA G UGUUUGAA 1265 TTCAAACA GGCTAGCTACAACGA TTTCTTGG 3640
1363 UGUUUGAA G CUGCAGUC 1266 GACTGCAG GGCTAGCTACAACGA TTCAAACA 3641
1369 AAGCUGCA G UCAAAUCC 1267 GGATTTGA GGCTAGCTACAACGA TGCAGCTT 3642
1384 CCAUCAAG G CAGCCUCC 1268 GGAGGCTG GGCTAGCTACAACGA CTTGATGG 3643
1387 UCAAGGCA G CCUCCUCC 1269 GGAGGAGG GGCTAGCTACAACGA TGCCTTGA 3644
1404 ACGGAGAA G UUCCCUGA 1270 TCAGGGAA GGCTAGCTACAACGA TTCTCCGT 3645
1415 CCCUGAUG G UUUCUGGC 1271 GCCAGAAA GGCTAGCTACAACGA CATCAGGG 3646
1422 GGUUUCUG G CUAGGAGA 1272 TCTCCTAG GGCTAGCTACAACGA CAGAAACC 3647
1431 CUAGGAGA G CAGCUGGU 1273 ACCAGCTG GGCTAGCTACAACGA TCTCCTAG 3648
1434 GGAGAGCA G CUGGUGUG 1274 CACACCAG GGCTAGCTACAACGA TGCTCTCC 3649
1438 AGCAGCUG G UGUGCUGG 1275 CCAGCACA GGCTAGCTACAACGA CAGCTGCT 3650
1446 GUGUGCUG G CAAGCAGG 1276 CCTGCTTG GGCTAGCTACAACGA CAGCACAC 3651
1450 GCUGGCAA G CAGGCACC 1277 GGTGCCTG GGCTAGCTACAACGA TTGCCAGC 3652
1454 GCAAGCAG G CACCACCC 1278 GGGTGGTG GGCTAGCTACAACGA CTGCTTGC 3653
1480 UUUUCCCA G UCAUCUCA 1279 TGAGATGA GGCTAGCTACAACGA TGGGAAAA 3654
1502 CCUAAUGG G UGAGGUUA 1280 TAACCTCA GGCTAGCTACAACGA CCATTAGG 3655
1507 UGGGUGAG G UUACCAAC 1281 GTTGGTAA GGCTAGCTACAACGA CTCACCCA 3656
1518 ACCAACCA G UCCUUCCG 1282 CGGAAGGA GGCTAGCTACAACGA TGGTTGGT 3657
1545 CUUCCGCA G CAAUACCU 1283 AGGTATTG GGCTAGCTACAACGA TGCGGAAG 3658
1557 UACCUGCG G CCAGUGGA 1284 TCCACTGG GGCTAGCTACAACGA CGCAGGTA 3659
1561 UGCGGCCA G UGGAAGAU 1285 ATCTTCCA GGCTAGCTACAACGA TGGCCGCA 3660
1573 AAGAUGUG G CCACGUCC 1286 GGACGTGG GGCTAGCTACAACGA CACATCTT 3661
1578 GUGGCCAC G UCCCAAGA 1287 TCTTGGGA GGCTAGCTACAACGA GTGGCCAC 3662
Table 22
1599 UGUUACAA G UUUGCCAU 1288 ATGGCAAA GGCTAGCTACAACGA TTGTAACA 3663
1614 AUCUCACA G UCAUCCAC 1289 GTGGATGA GGCTAGCTACAACGA TGTGAGAT 3664
1625 AUCCACGG G CACUGUUA 1290 TAACAGTG GGCTAGCTACAACGA CCGTGGAT 3665
1639 UUAUGGGA G CUGUUAUC 1291 GATAACAG GGCTAGCTACAACGA TCCCATAA 3666
1655 CAUGGAGG G CUUCUACG 1292 CGTAGAAG GGCTAGCTACAACGA CCTCCATG 3667
1663 GCUUCUAC G UUGUCUUU 1293 AAAGACAA GGCTAGCTACAACGA GTAGAAGC 3668
1678 UUGAUCGG G CCCGAAAA 1294 TTTTCGGG GGCTAGCTACAACGA CCGATCAA 3669
1694 ACGAAUUG G CUUUGCUG 1295 CAGCAAAG GGCTAGCTACAACGA CAATTCGT 3670
1706 UGCUGUCA G CGCUUGCC 1296 GGCAAGCG GGCTAGCTACAACGA TGACAGCA 3671
1728 CACGAUGA G UUCAGGAC ' 1297 GTCCTGAA GGCTAGCTACAACGA TCATCGTG 3672
1738 UCAGGACG G CAGCGGUG 1298 CACCGCTG GGCTAGCTACAACGA CGTCCTGA 3673
1741 GGACGGCA G CGGUGGAA 1299 TTCCACCG GGCTAGCTACAACGA TGCCGTCC 3674
1744 CGGCAGCG G UGGAAGGC 1300 GCCTTCCA GGCTAGCTACAACGA CGCTGCCG. 3675
1751 GGUGGAAG G CCCUUUUG 1301 CAAAAGGG GGCTAGCTACAACGA CTTCCACC 3676
1784 AGACUGUG G CUACAACA 1302 TGTTGTAG GGCTAGCTACAACGA CACAGTCT 3677
1809 ACAGAUGA G UCAACCCU 1303 AGGGTTGA GGCTAGCTACAACGA TCATCTGT 3678
1828 UGACCAUA G CCUAUGUC 1304 GACATAGG GGCTAGCTACAACGA TATGGTCA 3679
1840 AUGUCAUG G CUGCCAUC 1305 GATGGCAG GGCTAGCTACAACGA CATGACAT 3680
1882 GCCUCAUG G UGUGUCAG 1306 CTGACACA GGCTAGCTACAACGA CATGAGGC 3681
1890 GUGUGUCA G UGGCGCUG 1307 CAGCGCCA GGCTAGCTACAACGA TGACACAC 3682
1893 UGUCAGUG G CGCUGCCU 1308 AGGCAGCG GGCTAGCTACAACGA CACTGACA 3683
1917 CUGCGCCA G CAGCAUGA 1309 TCATGCTG GGCTAGCTACAACGA TGGCGCAG 3684
1920 CGCCAGCA G CAUGAUGA 1310 TCATCATG GGCTAGCTACAACGA TGCTGGCG 3685
1956 CUGCUGAA G UGAGGAGG 1311 CCTCCTCA GGCTAGCTACAACGA TTCAGCAG 3686
1964 GUGAGGAG G CCCAUGGG 1312 CCCATGGG GGCTAGCTACAACGA CTCCTCAC 3687
1972 GCCCAUGG G CAGAAGAU 1313 ATCTTCTG GGCTAGCTACAACGA CCATGGGC 3688
2006 ACACCUCC G UGGUUCAC 1314 GTGAACCA GGCTAGCTACAACGA GGAGGTGT 3689
2009 CCUCCGUG G UUCACUUU 1315 AAAGTGAA GGCTAGCTACAACGA CACGGAGG 3690
2019 UCACUUUG G UCACAAGU 1316 ACTTGTGA GGCTAGCTACAACGA CAAAGTGA 3691
2026 GGUCACAA G UAGGAGAC 1317 GTCTCCTA GGCTAGCTACAACGA TTGTGACC 3692
2042 CACAGAUG G CACCUGUG 1318 CACAGGTG GGCTAGCTACAACGA CATCTGTG 3693
2051 CACCUGUG G CCAGAGCA 1319 TGCTCTGG GGCTAGCTACAACGA CACAGGTG 3694
2057 UGGCCAGA G CACCUCAG 1320 CTGAGGTG GGCTAGCTACAACGA TCTGGCCA 3695
2114 AGGAAAAG G CUGGCAAG 1321 CTTGCCAG GGCTAGCTACAACGA CTTTTCCT 3696
2118 AAAGGCUG G CAAGGUGG 1322 CCACCTTG GGCTAGCTACAACGA CAGCCTTT 3697
2123 CUGGCAAG G UGGGUUCC 1323 GGAACCCA GGCTAGCTACAACGA CTTGCCAG 3698
2127 CAAGGUGG G UUCCAGGG 1324 ■ CCCTGGAA GGCTAGCTACAACGA CCACCTTG 3699
2172 AGAAAGAA G CACUCUGC 1325 GCAGAGTG GGCTAGCTACAACGA TTCTTTCT 3700
2183 CUCUGCUG G CGGGAAUA 1326 TATTCCCG GGCTAGCTACAACGA CAGCAGAG 3701
2198 UACUCUUG G UCACCUCA 1327 TGAGGTGA GGCTAGCTACAACGA CAAGAGTA 3702
2214 AAAUUUAA G UCGGGAAA 1328 TTTCCCGA GGCTAGCTACAACGA TTAAATTT 3703
2243 AAACUUCA G CCCUGAAC 1329 GTTCAGGG GGCTAGCTACAACGA TGAAGTTT 3704
2288 AACCCAAA G UAUUCUUC 1330 GAAGAATA GGCTAGCTACAACGA TTTGGGTT 3705
2305 UUUUCUUA G UUUCAGAA 1331 TTCTGAAA GGCTAGCTACAACGA TAAGAAAA 3706
2314 UUUCAGAA G UACUGGCA 1332 TGCCAGTA GGCTAGCTACAACGA TTCTGAAA 3707
2320 AAGUACUG G CAUCACAC 1333 GTGTGATG GGCTAGCTACAACGA CAGTACTT 3708
2333 ACACGCAG G UUACCUUG 1334 CAAGGTAA GGCTAGCTACAACGA CTGCGTGT 3709
2342 UUACCUUG G CGUGUGUC 1335 GACACACG GGCTAGCTACAACGA CAAGGTAA 3710
2344 ACCUUGGC G UGUGUCCC 1336 GGGACACA GGCTAGCTACAACGA GCCAAGGT 3711
2357 UCCCUGUG G UACCCUGG 1337 CCAGGGTA GGCTAGCTACAACGA CACAGGGA 3712
2365 GUACCCUG G CAGAGAAG 1338 CTTCTCTG GGCTAGCTACAACGA CAGGGTAC 3713
Table 22
2381 GAGACCAA G CUUGUUUC 1339 GAAACAAG GGCTAGCTACAACGA TTGGTCTC 3714
2397 CCCUGCUG G CCAAAGUC 1340 GACTTTGG GGCTAGCTACAACGA CAGCAGGG 3715
2403 UGGCCAAA G UCAGUAGG 1341 CCTACTGA GGCTAGCTACAACGA TTTGGCCA 3716
2407 CAAAGUCA G UAGGAGAG 1342 CTCTCCTA GGCTAGCTACAACGA TGACTTTG 3717
2424 GAUGCACA G UUUGCUAU 1343 ATAGCAAA GGCTAGCTACAACGA TGTGCATC 3718
2463 AUAAACAA G CCUAACAU 1344 ATGTTAGG GGCTAGCTACAACGA TTGTTTAT 3719
2474 UAACAUUG G UGCAAAGA 1345 TCTTTGCA GGCTAGCTACAACGA CAATGTTA 3720
45 CGAGCUGG A UUAUGGUG 1346 CACCATAA GGCTAGCTACAACGA CCAGCTCG 3721
67 AGCAGCCA A CGCAGCCG 1347 CGGCTGCG GGCTAGCTACAACGA TGGCTGCT 3722
125 CCGGGGGG A CCAGGGAA 1348 TTCCCTGG GGCTAGCTACAACGA CCCCCCGG 3723
217 CCGUGCCG A UGUAGCGG 1349 CCGCTACA GGCTAGCTACAACGA CGGCACGG 3724
233 GGCUCCGG A UCCCAGCC 1350 GGCTGGGA GGCTAGCTACAACGA CCGGAGCC 3725
267 CUCUGCGG A UCUCCCCU 1351 AGGGGAGA GGCTAGCTACAACGA CCGCAGAG 3726
277 CUCCCCUG A CCGCUCUC 1352 GAGAGCGG GGCTAGCTACAACGA CAGGGGAG 3727
296 CAGCCCGG A CCCGGGGG 1353 CCCCCGGG GGCTAGCTACAACGA CCGGGCTG 3728
338 GCGUCCUG A UGCCCCCA 1354 TGGGGGCA GGCTAGCTACAACGA CAGGACGC 3729
383 CCACCCAG A CUUGGGGG 1355 CCCCCAAG GGCTAGCTACAACGA CTGGGTGG 3730
404 CGCCAGGG A CGGACGUG 1356 CACGTCCG GGCTAGCTACAACGA CCCTGGCG 3731
408 AGGGACGG A CGUGGGCC 1357 GGCCCACG GGCTAGCTACAACGA CCGTCCCT 3732
487 UGCUGUGG A UGGGCGCG 1358 CGCGCCCA GGCTAGCTACAACGA CCACAGCA 3733
592 CCCGGGAG A CCGACGAA 1359 TTCGTCGG GGCTAGCTACAACGA CTCCCGGG 3734
596 GGAGACCG A CGAAGAGC 1360 GCTCTTCG GGCTAGCTACAACGA CGGTCTCC 3735
640 UUGUGGAG A UGGUGGAC 1361 GTCCACCA GGCTAGCTACAACGA CTCCACAA 3736
647 GAUGGUGG A CAACCUGA 1362 TCAGGTTG GGCTAGCTACAACGA CCACCATC 3737
650 GGUGGACA A CCUGAGGG 1363 CCCTCAGG GGCTAGCTACAACGA TGTCCACC 3738
688 ACGUGGAG A UGACCGUG 1364 CACGGTCA GGCTAGCTACAACGA CTCCACGT 3739
691 UGGAGAUG A CCGUGGGC 1365 GCCCACGG GGCTAGCTACAACGA CATCTCCA 3740
712 CCCCGCAG A CGCUCAAC 1366 GTTGAGCG GGCTAGCTACAACGA CTGCGGGG 3741
719 GACGCUCA A CAUCCUGG 1367 CCAGGATG GGCTAGCTACAACGA TGAGCGTC 3742
731 CCUGGUGG A UACAGGCA 1368 TGCCTGTA GGCTAGCTACAACGA CCACCAGG 3743
746 CAGCAGUA A CUUUGCAG 1369 CTGCAAAG GGCTAGCTACAACGA TACTGCTG 3744
821 AUACCGGG A CCUCCGGA 1370 TCCGGAGG GGCTAGCTACAACGA CCCGGTAT 3745
884 GGGCACCG A CCUGGUAA 1371 TTACCAGG GGCTAGCTACAACGA CGGTGCCC 3746
911 UGGCCCCA A CGUCACUG 1372 CAGTGACG GGCTAGCTACAACGA TGGGGCCA 3747
929 GCGUGCCA A CAUUGCUG 1373 CAGCAATG GGCTAGCTACAACGA TGGCACGC 3748
948 AUCACUGA A UCAGACAA 1374 TTGTCTGA GGCTAGCTACAACGA TCAGTGAT 3749
953 UGAAUCAG A CAAGUUCU 1375 AGAACTTG GGCTAGCTACAACGA CTGATTCA 3750
968 CUUCAUCA A CGGCUCCA 1376 TGGAGCCG GGCTAGCTACAACGA TGATGAAG 3751
977 CGGCUCCA A CUGGGAAG 1377 CTTCCCAG GGCTAGCTACAACGA TGGAGCCG 3752
1012 AUGCUGAG A UUGCCAGG 1378 CCTGGCAA GGCTAGCTACAACGA CTCAGCAT 3753
1025 CAGGCCUG A CGACUCCC 1379 GGGAGTCG GGCTAGCTACAACGA CAGGCCTG 3754
1028 GCCUGACG A CUCCCUGG 1380 CCAGGGAG GGCTAGCTACAACGA CGTCAGGC 3755
1049 UUUCUUUG A CUCUCUGG 1381 CCAGAGAG GGCTAGCTACAACGA CAAAGAAA 3756
1066 UAAAGCAG A CCCACGUU 1382 AACGTGGG GGCTAGCTACAACGA CTGCTTTA 3757
1079 CGUUCCCA A CCUCUUCU 1383 AGAAGAGG GGCTAGCTACAACGA TGGGAACG 3758
1121 CCCCCUCA A CCAGUCUG 1384 CAGACTGG GGCTAGCTACAACGA TGAGGGGG 3759
1159 GGAGCAUG A UCAUUGGA 1385 TCCAATGA GGCTAGCTACAACGA CATGCTCC 3760
1175 AGGUAUCG A CCACUCGC 1386 GCGAGTGG GGCTAGCTACAACGA CGATACCT 3761
1240 AUGAGGUG A UCAUUGUG 1387 CACAATGA GGCTAGCTACAACGA CACCTCAT 3762
1258 GGGUGGAG A UCAAUGGA 1388 TCCATTGA GGCTAGCTACAACGA CTCCACCC 3763
1262 GGAGAUCA A UGGACAGG 1389 CCTGTCCA GGCTAGCTACAACGA TGATCTCC 3764
Table 22
1266 AUCAAUGG A CAGGAUCU 1390 AGATCCTG GGCTAGCTACAACGA CCATTGAT 3765
1271 UGGACAGG A UCUGAAAA 1391 TTTTCAGA GGCTAGCTACAACGA CCTGTCCA 3766
1279 AUCUGAAA A UGGACUGC 1392 GCAGTCCA GGCTAGCTACAACGA TTTCAGAT 3767
1283 GAAAAUGG A CUGCAAGG 1393 CCTTGCAG GGCTAGCTACAACGA CCATTTTC 3768
1298 GGAGUACA A CUAUGACA 1394 TGTCATAG GGCTAGCTACAACGA TGTACTCC 3769
1304 CAACUAUG A CAAGAGCA 1395 TGCTCTTG GGCTAGCTACAACGA CATAGTTG 3770
1319 CAUUGUGG A CAGUGGCA 1396 TGCCACTG GGCTAGCTACAACGA CCACAATG 3771
1334 CACCACCA A CCUUCGUU 1397 AACGAAGG GGCTAGCTACAACGA TGGTGGTG 3772
1374 GCAGUCAA A UCCAUCAA 1398 TTGATGGA GGCTAGCTACAACGA TTGACTGC 3773
1412 GUUCCCUG A UGGUUUCU 1399 AGAAACCA GGCTAGCTACAACGA CAGGGAAC 3774
1469 CCCUUGGA A CAUUUUCC 1400 GGAAAATG GGCTAGCTACAACGA TCCAAGGG 3775
1498 UCUACCUA A UGGGUGAG 1401 CTCACCCA GGCTAGCTACAACGA TAGGTAGA 3776
1514 GGUUACCA A CCAGUCCU 1402 AGGACTGG GGCTAGCTACAACGA TGGTAACC 3777
1548 CCGCAGCA A UACCUGCG 1403 CGCAGGTA GGCTAGCTACAACGA TGCTGCGG 3778
1568 AGUGGAAG A UGUGGCCA 1404 TGGCCACA GGCTAGCTACAACGA CTTCCACT 3779
1586 GUCCCAAG A CGACUGUU 1405 AACAGTCG GGCTAGCTACAACGA CTTGGGAC 3780
1589 CCAAGACG A CUGUUACA 1406 TGTAACAG GGCTAGCTACAACGA CGTCTTGG 3781
1673 UGUCUUUG A UCGGGCCC 1407 GGGCCCGA GGCTAGCTACAACGA CAAAGACA 3782
1686 GCCCGAAA A CGAAUUGG 1408 CCAATTCG GGCTAGCTACAACGA TTTCGGGC 3783
1690 GAAAACGA A UUGGCUUU 1409 AAAGCCAA GGCTAGCTACAACGA TCGTTTTC 3784
1724 UGUGCACG A UGAGUUCA 1410 TGAACTCA GGCTAGCTACAACGA CGTGCACA 3785
1735 AGUUCAGG A CGGCAGCG 1411 CGCTGCCG GGCTAGCTACAACGA CCTGAACT 3786
1769 CACCUUGG A CAUGGAAG 1412 CTTCCATG GGCTAGCTACAACGA CCAAGGTG 3787
1778 CAUGGAAG A CUGUGGCU 1413 AGCCACAG GGCTAGCTACAACGA CTTCCATG 3788
1790 UGGCUACA A CAUUCCAC 1414 GTGGAATG GGCTAGCTACAACGA TGTAGCCA 3789
1801 UUCCACAG A CAGAUGAG 1415 CTCATCTG GGCTAGCTACAACGA CTGTGGAA 3790
1805 ACAGACAG A UGAGUCAA 1416 TTGACTCA GGCTAGCTACAACGA CTGTCTGT 3791
1813 AUGAGUCA A CCCUCAUG 1417 CATGAGGG GGCTAGCTACAACGA TGACTCAT 3792
1822 CCCUCAUG A CCAUAGCC 1418 GGCTATGG GGCTAGCTACAACGA CATGAGGG 3793
1925 GCAGCAUG A UGACUUUG 1419 CAAAGTCA GGCTAGCTACAACGA CATGCTGC 3794
1928 GCAUGAUG A CUUUGCUG 1420 CAGCAAAG GGCTAGCTACAACGA CATCATGC 3795
1937 CUUUGCUG A UGACAUCU 1421 AGATGTCA GGCTAGCTACAACGA CAGCAAAG 3796
1940 UGCUGAUG A CAUCUCCC 1422 GGGAGATG GGCTAGCTACAACGA CATCAGCA 3797
1979 GGCAGAAG A UAGAGAUU 1423 AATCTCTA GGCTAGCTACAACGA CTTCTGCC 3798
1985 AGAUAGAG A UUCCCCUG 1424 CAGGGGAA GGCTAGCTACAACGA CTCTATCT 3799
1995 UCCCCUGG A CCACACCU 1425 AGGTGTGG GGCTAGCTACAACGA CCAGGGGA 3800
2033 AGUAGGAG A CACAGAUG 1426 CATCTGTG GGCTAGCTACAACGA CTCCTACT 3801
2039 AGACACAG A UGGCACCU 1427 AGGTGCCA GGCTAGCTACAACGA CTGTGTCT 3802
2067 ACCUCAGG A CCCUCCCC 1428 GGGGAGGG GGCTAGCTACAACGA CCTGAGGT 3803
2085 CCCACCAA A UGCCUCUG 1429 CAGAGGCA GGCTAGCTACAACGA TTGGTGGG 3804
2099 CUGCCUUG A UGGAGAAG 1430 CTTCTCCA GGCTAGCTACAACGA CAAGGCAG 3805
2136 UUCCAGGG A CUGUACCU 1431 AGGTACAG GGCTAGCTACAACGA CCCTGGAA 3806
2152 UGUAGGAA A CAGAAAAG 1432 CTTTTCTG GGCTAGCTACAACGA TTCCTACA 3807
2189 UGGCGGGA A UACUCUUG 1433 CAAGAGTA GGCTAGCTACAACGA TCCCGCCA 3808
2208 CACCUCAA A UUUAAGUC 1434 GACTTAAA GGCTAGCTACAACGA TTGAGGTG 3809
2222 GUCGGGAA A UUCUGCUG 1435 CAGCAGAA GGCTAGCTACAACGA TTCCCGAC 3810
2237 UGCUUGAA A CUUCAGCC " 1436 GGCTGAAG GGCTAGCTACAACGA TTCAAGCA 3811
2250 AGCCCUGA A CCUUUGUC 1437 GACAAAGG GGCTAGCTACAACGA TCAGGGCT 3812
2273 UCCUUUAA A UUCUCCAA 1438 TTGGAGAA GGCTAGCTACAACGA TTAAAGGA 3813
2281 AUUCUCCA A CCCAAAGU 1439 ACTTTGGG GGCTAGCTACAACGA TGGAGAAT 3814
2376 GAGAAGAG A CCAAGCUU 1440 AAGCTTGG GGCTAGCTACAACGA CTCTTCTC 3815
Table 22
2417 AGGAGAGG A UGCACAGU 1441 ACTGTGCA GGCTAGCTACAACGA CCTCTCCT 3816
2444 CUUUAGAG A CAGGGACU 1442 AGTCCCTG GGCTAGCTACAACGA CTCTAAAG 3817
2450 AGACAGGG A CUGUAUAA 1443 TTATACAG GGCTAGCTACAACGA CCCTGTCT 3818
2459 CUGUAUAA A CAAGCCUA 1444 TAGGCTTG GGCTAGCTACAACGA TTATACAG 3819
2468 CAAGCCUA A CAUUGGUG 1445 CACCAATG GGCTAGCTACAACGA TAGGCTTG 3820
2482 GUGCAAAG A UUGCCUCU 1446 AGAGGCAA GGCTAGCTACAACGA CTTTGCAC 3821
2494 CCUCUUGA A UUAAAAAA 1447 TTTTTTAA GGCTAGCTACAACGA TCAAGAGG 3822
2507 AAAAAAAA A CUAGAAAA 1448 TTTTCTAG GGCTAGCTACAACGA TTTTTTTT 3823
Input Sequence = AF190725. Cut Site = G/ .
Stem Length = 8 . Core Sequence = GGCTAGCTACAACGA
AF190725 (Homo sapiens beta-site APP cleaving enzyme (BACE) mRNA; 2526 bp)
Table 23
Table 23: Human BACE Amberzyme Ribozyme and Target Sequence
Pos Substrate Seq ID Ribozyme Rz Seq ID
11 ACGCGUCC G CAGCCCGC 960 GCGGGCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGACGCGU 3260
18 CGCAGCCC G CCCGGGAG 961 CUCCCGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGGCUGCG 3261
29 CGGGAGCU G CGAGCCGC 962 GCGGCUCG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCUCCCG 3262
31 GGAGCUGC G AGCCGCGA 963 UCGCGGCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCAGCUCC 3263
36 UGCGAGCC G CGAGCUGG 964 CCAGCUCG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGCUCGCA 3264
38 CGAGCCGC G AGCUGGAU 965 AUCCAGCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCGGCUCG 3265
58 GGUGGCCU G AGCAGCCA 966 UGGCUGCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGCCACC 3266
69 CAGCCAAC G CAGCCGCA 967 UGCGGCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUUGGCUG 3267
75 ACGCAGCC G CAGGAGCC 968 GGCUCCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGCUGCGU 3268
94 GAGCCCUU G CCCCUGCC 969 GGCAGGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAGGGCUC 3269
100 UUGCCCCU G CCCGCGCC 970 GGCGCGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGGGCAA 3270
104 CCCUGCCC G CGCCGCCG 971 CGGCGGCG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGGCAGGG 3271
106 CUGCCCGC G CCGCCGCC 972 GGCGGCGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCGGGCAG 3272
109 CCCGCGCC G CCGCCCGC 973 GCGGGCGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGCGCGGG 3273
112 GCGCCGCC G CCCGCCGG 974 CCGGCGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGCGGCGC 3274
116 CGCCGCCC G CCGGGGGG 975 CCCCCCGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGCGGCG 3275
137 GGGAAGCC G CCACCGGC 976 GCCGGUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGCUUCCC 3276
148 ACCGGCCC G CCAUGCCC 977 GGGCAUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGCCGGU 3277
153 CCCGCCAU G CCCGCCCC 978 GGGGCGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGGCGGG 3278
157 CCAUGCCC G CCCCUCCC 979 GGGAGGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGCAUGG 3279
172 CCAGCCCC G CCGGGAGC 980 GCUCCCGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGGCUGG 3280
183 GGGAGCCC G CGCCCGCU 981 AGCGGGCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGCUCCC 3281
185 GAGCCCGC G CCCGCUGC 982 GCAGCGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCGGGCUC 3282
189 CCGCGCCC G CUGCCCAG 983 CUGGGCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGCGCGG 3283
192 CGCCCGCU G CCCAGGCU 984 AGCCUGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCGGGCG 3284
205 GGCUGGCC G CCGCCGUG 985 CACGGCGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGCCAGCC 3285
208 UGGCCGCC G CCGUGCCG 986 CGGCACGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGCGGCCA 3286
213 GCCGCCGU G CCGAUGUA 987 UACAUCGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACGGCGGC 3287
216 GCCGUGCC G AUGUAGCG 988 CGCUACAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGCACGGC 3288
250 UCUCCCCU G CUCCCGUG 989 CACGGGAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGGGAGA 3289
Table 23
258 GCUCCCGU G CUCUGCGG 990 CCGCAGAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACGGGAGC 3290
263 CGUGCUCU G CGGAUCUC 991 GAGAUCCG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGAGCACG 3291
276 UCUCCCCU G ACCGCUCU 992 AGAGCGGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGGGAGA 3292
280 CCCUGACC G CUCUCCAC 993 GUGGAGAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGUCAGGG 3293
320 AGGGCCCU G CAGGCCCU 994 AGGGCCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGGCCCU 3294
337 GGCGUCCU G AUGCCCCC 995 GGGGGCAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGACGCC 3295
340 GUCCUGAU G CCCCCAAG 996 CUUGGGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUCAGGAC 3296
360 CCUCUCCU G AGAAGCCA 997 UGGCUUCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGAGAGG 3297
397 GGGCAGGC G CCAGGGAC 998 GUCCCUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCCUGCCC 3298
420 GGGCCAGU G CGAGCCCA 999 UGGGCUCG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACUGGCCC 3299
422 GCCAGUGC G AGCCCAGA 1000 UCUGGGCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCACUGGC 3300
437 GAGGGCCC G AAGGCCGG 1001 CCGGCCUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGCCCUC 3301
468 CAAGCCCU G CCCUGGCU 1002 AGCCAGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGGCUUG 3302
480 UGGCUCCU G CUGUGGAU 1003 AUCCACAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGAGCCA 3303
493 GGAUGGGC G CGGGAGUG 1004 CACUCCCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCCCAUCC 3304
501 GCGGGAGU G CUGCCUGC 1005 GCAGGCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACUCCCGC 3305
504 GGAGUGCU G CCUGCCCA 1006 UGGGCAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCACUCC 3306
508 UGCUGCCU G CCCACGGC 1007 GCCGUGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGCAGCA 3307
537 AUCCGGCU G CCCCUGCG 1008 CGCAGGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCCGGAU 3308
543 CUGCCCCU G CGCAGCGG 1009 CCGCUGCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGGGCAG 3309
545 GCCCCUGC G CAGCGGCC 1010 GGCCGCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCAGGGGC 3310
562 UGGGGGGC G CCCCCCUG 1011 CAGGGGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCCCCCCA 3311
576 CUGGGGCU G CGGCUGCC 1012 GGCAGCCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCCCCAG 3312
582 CUGCGGCU G CCCCGGGA 1013 UCCCGGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCCGCAG 3313
595 GGGAGACC G ACGAAGAG 1014 CUCUUCGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGUCUCCC 3314
598 AGACCGAC G AAGAGCCC 1015 GGGCUCUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUCGGUCU 3315
607 AAGAGCCC G AGGAGCCC 1016 GGGCUCCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGCUCUU 3316
654 GACAACCU G AGGGGCAA 1017 UUGCCCCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGUUGUC 3317
690 GUGGAGAU G ACCGUGGG 1018 CCCACGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCUCCAC 3318
708 AGCCCCCC G CAGACGCU 1019 AGCGUCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGGGGCU 3319
714 CCGCAGAC G CUCAACAU 1020 AUGUUGAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUCUGCGG 3320
751 GUAACUUU G CAGUGGGU 1021 ACCCACUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAGUUAC 3321
760 CAGUGGGU G CUGCCCCC 1022 GGGGGCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCCACUG 3322
763 UGGGUGCU G CCCCCCAC 1023 GUGGGGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCACCCA 3323
Table 23
780 CCCUUCCU G CAUCGCUA 1024 UAGCGAUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGAAGGG 3324
785 CCUGCAUC G CUACUACC 1025 GGUAGUAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAUGCAGG 3325
843 GUGUAUGU G CCCUACAC 1026 GUGUAGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACAUACAC 3326
883 UGGGCACC G ACCUGGUA 1027 UACCAGGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGUGCCCA 3327
921 GUCACUGU G CGUGCCAA 1028 UUGGCACG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACAGUGAC 3328
925 CUGUGCGU G CCAACAUU 1029 AAUGUUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACGCACAG 3329
934 CCAACAUU G CUGCCAUC 1030 GAUGGCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAUGUUGG 3330
937 ACAUUGCU G CCAUCACU 1031 AGUGAUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCAAUGU 3331
946 CCAUCACU G AAUCAGAC 1032 GUCUGAUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGUGAUGG 3332
1006 UGGCCUAU G CUGAGAUU 1033 AAUCUCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUAGGCCA 3333
1009 CCUAUGCU G AGAUUGCC 1034 GGCAAUCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCAUAGG 3334
1015 CUGAGAUU G CCAGGCCU 1035 AGGCCUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAUCUCAG 3335
1024 CCAGGCCU G ACGACUCC 1036 GGAGUCGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGCCUGG 3336
1027 GGCCUGAC G ACUCCCUG 1037 CAGGGAGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUCAGGCC 3337
1048 CUUUCUUU G ACUCUCUG 1038 CAGAGAGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAGAAAG 3338
1092 UUCUCCCU G CAGCUUUG 1039 CAAAGCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGGAGAA 3339
1105 UUUGUGGU G CUGGCUUC 1040 GAAGCCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACCACAAA 3340
1129 ACCAGUCU G AAGUGCUG 1041 CAGCACUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGACUGGU 3341
1134 UCUGAAGU G CUGGCCUC 1042 GAGGCCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACUUCAGA 3342
1158 GGGAGCAU G AUCAUUGG 1043 CCAAUGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGCUCCC 3343
1174 GAGGUAUC G ACCACUCG 1044 CGAGUGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GAUACCUC 3344
1182 GACCACUC G CUGUACAC 1045 GUGUACAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GAGUGGUC 3345
1234 GGUAUUAU G AGGUGAUC 1046 GAUCACCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUAAUACC 3346
1239 UAUGAGGU G AUCAUUGU 1047 ACAAUGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCUCAUA 3347
1248 AUCAUUGU G CGGGUGGA 1048 UCCACCCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAAUGAU 3348
1275 CAGGAUCU G AAAAUGGA 1049 UCCAUUUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAUCCUG 3349
1286 AAUGGACU G CAAGGAGU 1050 ACUCCUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUCCAUU 3350
1303 ACAACUAU G ACAAGAGC 1051 GCUCUUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUAGUUGU 3351
1344 CUUCGUUU G CCCAAGAA 1052 UUCUUGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAACGAAG 3352
1360 AAGUGUUU G AAGCUGCA 1053 UGCAGCUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAACACUU 3353
1366 UUGAAGCU G CAGUCAAA 1054 UUUGACUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCUUCAA 3354
1411 AGUUCCCU G AUGGUUUC 1055 GAAACCAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGGAACU 3355
1442 GCUGGUGU G CUGGCAAG 1056 CUUGCCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACACCAGC 3356
1504 UAAUGGGU G AGGUUACC 1057 GGUAACCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCCAUUA 3357
Table 23
1526 GUCCUUCC G CAUCACCA 1058 UGGUGAUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGAAGGAC 3358
1542 AUCCUUCC G CAGCAAUA 1059 UAUUGCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGAAGGAU 3359
1554 CAAUACCU G CGGCCAGU 1060 ACUGGCCG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGUAUUG 3360
1588 CCCAAGAC G ACUGUUAC 1061 GUAACAGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUCUUGGG 3361
1603 ACAAGUUU G CCAUCUCA 1062 UGAGAUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAACUUGU 3362
1672 UUGUCUUU G AUCGGGCC 1063 GGCCCGAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAGACAA 3363
1682 UCGGGCCC G AAAACGAA 1064 UUCGUUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGGCCCGA 3364
1688 CCGAAAAC G AAUUGGCU 1065 AGCCAAUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUUUUCGG 3365
1699 UUGGCUUU G CUGUCAGC 1066 GCUGACAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAGCCAA 3366
1708 CUGUCAGC G CUUGCCAU 1067 AUGGCAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCUGACAG 3367
1712 CAGCGCUU G CCAUGUGC 1068 GCACAUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAGCGCUG 3368
1719 UGCCAUGU G CACGAUGA 1069 UCAUCGUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACAUGGCA 3369
1723 AUGUGCAC G AUGAGUUC 1070 GAACUCAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUGCACAU 3370
1726 UGCACGAU G AGUUCAGG 1071 CCUGAACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCGUGCA 3371
1807 AGACAGAU G AGUCAACC 1072 GGUUGACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCUGUCU 3372
1821 ACCCUCAU G ACCAUAGC 1073 GCUAUGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGAGGGU 3373
1843 UCAUGGCU G CCAUCUGC 1074 GCAGAUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCCAUGA 3374
1850 UGCCAUCU G CGCCCUCU 1075 AGAGGGCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAUGGCA 3375
1852 CCAUCUGC G CCCUCUUC 1076 GAAGAGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCAGAUGG 3376
1863 CUCUUCAU G CUGCCACU 1077 AGUGGCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGAAGAG 3377
1866 UUCAUGCU G CCACUCUG 1078 CAGAGUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCAUGAA 3378
1874 GCCACUCU G CCUCAUGG 1079 CCAUGAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAGUGGC 3379
1895 UCAGUGGC G CUGCCUCC 1080 GGAGGCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCCACUGA 3380
1898 GUGGCGCU G CCUCCGCU 1081 AGCGGAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCGCCAC 3381
1904 CUGCCUCC G CUGCCUGC 1082 GCAGGCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGAGGCAG 3382
1907 CCUCCGCU G CCUGCGCC 1083 GGCGCAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCGGAGG 3383
1911 CGCUGCCU G CGCCAGCA 1084 UGCUGGCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGCAGCG 3384
1913 CUGCCUGC G CCAGCAGC 1085 GCUGCUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCAGGCAG 3385
1924 AGCAGCAU G AUGACUUU 1086 AAAGUCAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGCUGCU 3386
1927 AGCAUGAU G ACUUUGCU 1087 AGCAAAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCAUGCU 3387
1933 AUGACUUU G CUGAUGAC 1088 GUCAUCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAGUCAU 3388
1936 ACUUUGCU G AUGACAUC 1089 GAUGUCAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCAAAGU 3389
1939 UUGCUGAU G ACAUCUCC 1090 GGAGAUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCAGCAA 3390
1950 AUCUCCCU G CUGAAGUG 1091 CACUUCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGGAGAU 3391
Table 23
1953 UCCCUGCU G AAGUGAGG 1092 CCUCACUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCAGGGA 3392
1958 GCUGAAGU G AGGAGGCC 1093 GGCCUCCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACUUCAGC 3393
2087 CACCAAAU G CCUCUGCC 1094 GGCAGAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUUUGGUG 3394
2093 AUGCCUCU G CCUUGAUG 1095 CAUCAAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG 'AGAGGCAU 3395
2098 UCUGCCUU G AUGGAGAA 1096 UUCUCCAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAGGCAGA 3396
2179 AGCACUCU G CUGGCGGG 1097 CCCGCCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGAGUGCU 3397
2227 GAAAUUCU G CUGCUUGA 1098 UCAAGCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGAAUUUC 3398
2230 AUUCUGCU G CUUGAAAC 1099 GUUUCAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCAGAAU 3399
2234 UGCUGCUU G AAACUUCA 1100 UGAAGUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAGCAGCA 3400
2248 UCAGCCCU G AACCUUUG 1101 CAAAGGUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGGCUGA 3401
2329 CAUCACAC G CAGGUUAC 1102 GUAACCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUGUGAUG 3402
2393 GUUUCCCU G CUGGCCAA 1103 UUGGCCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGGAAAC 3403
2419 GAGAGGAU G CACAGUUU 1104 AAACUGUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUCCUCUC 3404
2428 CACAGUUU G CUAUUUGC 1105 GCAAAUAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAACUGUG 3405
2435 UGCUAUUU G CUUUAGAG 1106 CUCUAAAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAUAGCA 3406
2476 ACAUUGGU G CAAAGAUU 1107 AAUCUUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCAAUGU 3407
2485 CAAAGAUU G CCUCUUGA 1108 UCAAGAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUCUUUG 3408
2492 UGCCUCUU G AAUUAAAA 1109 UUUUAAUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGAGGCA 3409
219 GUGCCGAU G UAGCGGGC 1110 GCCCGCUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCGGCAC 3410
483 CUCCUGCU G UGGAUGGG llll CCCAUCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCAGGAG 3411
634 GCAGCUUU G UGGAGAUG 1112 CAUCUCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAGCUGC 3412
804 AGGCAGCU G UCCAGCAC 1113 GUGCUGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCUGCCU 3413
835 GGAAGGGU G UGUAUGUG 1114 CACAUACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCCUUCC 3414
837 AAGGGUGU G UAUGUGCC 1115 GGCACAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACACCCUU 3415
841 GUGUGUAU G UGCCCUAC 1116 GUAGGGCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUACACAC 3416
919 ACGUCACU G UGCGUGCC 1117 GGCACGCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUGACGU 3417
1100 GCAGCUUU G UGGUGCUG 1118 CAGCACCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAGCUGC 3418
1144 UGGCCUCU G UCGGAGGG 1119 CCCUCCGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAGGCCA 3419
1185 CACUCGCU G UACACAGG 1120 CCUGUGUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCGAGUG 3420
1246 UGAUCAUU G UGCGGGUG 1121 CACCCGCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUGAUCA 3421
1315 AGAGCAUU G UGGACAGU 1122 ACUGUCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUGCUCU 3422
1356 AAGAAAGU G UUUGAAGC 1123 GCUUCAAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACUUUCUU 3423
1440 CAGCUGGU G UGCUGGCA 1124 UGCCAGCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCAGCUG 3424
1570 UGGAAGAU G UGGCCACG 1125 CGUGGCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCUUCCA 3425
Table 23
1592 AGACGACU G UUACAAGU 1126 ACUUGUAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGUCGUCU 3426
1630 CGGGCACU G UUAUGGGA 1127 UCCCAUAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGUGCCCG 3427
1642 UGGGAGCU G UUAUCAUG 1128 CAUGAUAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCUCCCA 3428
1666 UCUACGUU G UCUUUGAU 1129 AUCAAAGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AACGUAGA 3429
1702 GCUUUGCU G UCAGCGCU 1130 AGCGCUGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCAAAGC 3430
1717 CUUGCCAU G UGCACGAU 1131 AUCGUGCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUGGCAAG 3431
1759 GCCCUUUU G UCACCUUG 1132 CAAGGUGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAAGGGC 3432
1781 GGAAGACU G UGGCUACA 1133 UGUAGCCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGUCUUCC 3433
1834 UAGCCUAU G UCAUGGCU 1134 AGCCAUGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUAGGCUA 3434
1884 CUCAUGGU G UGUCAGUG 1135 CACUGACA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACCAUGAG 3435
1886 CAUGGUGU G UCAGUGGC 1136 GCCACUGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACACCAUG 3436
2048 UGGCACCU G UGGCCAGA 1137 UCUGGCCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGUGCCA 3437
2139 CAGGGACU G UACCUGUA 1138 UACAGGUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGUCCCUG 3438
2145 CUGUACCU G UAGGAAAC 1139 GUUUCCUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGUACAG 3439
2256 GAACCUUU G UCCACCAU 1140 AUGGUGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAGGUUC 3440
2346 CUUGGCGU G UGUCCCUG 1141 CAGGGACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACGCCAAG 3441
2348 UGGCGUGU G UCCCUGUG 1142 CACAGGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACACGCCA 3442
2354 GUGUCCCU G UGGUACCC 1143 GGGUACCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGGACAC 3443
2385 CCAAGCUU G UUUCCCUG 1144 CAGGGAAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGCUUGG 3444
2453 CAGGGACU G UAUAAACA 1145 UGUUUAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUCCCUG 3445
14 CGUCCGCA G CCCGCCCG 1146 CGGGCGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCGGACG 3446
26 GCCCGGGA G CUGCGAGC 1147 GCUCGCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCCGGGC 3447
33 AGCUGCGA G CCGCGAGC 1148 GCUCGCGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCGCAGCU 3448
40 AGCCGCGA G CUGGAUUA 1149 UAAUCCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCGCGGCU 3449
51 GGAUUAUG G UGGCCUGA 1150 UCAGGCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUAAUCC 3450
54 UUAUGGUG G CCUGAGCA 1151 UGCUCAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACCAUAA 3451
60 UGGCCUGA G CAGCCAAC 1152 GUUGGCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCAGGCCA 3452
63 CCUGAGCA G CCAACGCA 1153 UGCGUUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCUCAGG 3453
72 CCAACGCA G CCGCAGGA 1154 UCCUGCGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCGUUGG 3454
81 CCGCAGGA G CCCGGAGC 1155 GCUCCGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCUGCGG 3455
88 AGCCCGGA G CCCUUGCC 1156 GGCAAGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCGGGCU 3456
134 CCAGGGAA G CCGCCACC 1157 GGUGGCGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCCCUGG 3457
144 CGCCACCG G CCCGCCAU 1158 AUGGCGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGGUGGCG 3458
167 CCCUCCCA G CCCCGCCG 1159 CGGCGGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGGAGGG 3459
Table 23
179 CGCCGGGA G CCCGCGCC 1160 GGCGCGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCCCGGCG 3460
198 CUGCCCAG G CUGGCCGC 1161 GCGGCCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUGGGCAG 3461
202 CCAGGCUG G CCGCCGCC 1162 GGCGGCGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGCCUGG 3462
211 CCGCCGCC G UGCCGAUG 1163 CAUCGGCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGCGGCGG 3463
222 CCGAUGUA G CGGGCUCC 1164 GGAGCCCG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UACAUCGG 3464
226 UGUAGCGG G CUCCGGAU 1165 AUCCGGAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCGCUACA 3465
239 GGAUCCCA G CCUCUCCC 1166 GGGAGAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGGGAUCC 3466
256 CUGCUCCC G UGCUCUGC 1167 GCAGAGCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGGAGCAG 3467
290 UCUCCACA G CCCGGACC 1168 GGUCCGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGUGGAGA 3468
304 ACCCGGGG G CUGGCCCA 1169 UGGGCCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCCGGGU 3469
308 GGGGGCUG G CCCAGGGC 1170 GCCCUGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGCCCCC 3470
315 GGCCCAGG G CCCUGCAG 1171 CUGCAGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCUGGGCC 3471
324 CCCUGCAG G CCCUGGCG 1172 CGCCAGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGCAGGG 3472
330 AGGCCCUG G CGUCCUGA 1173 UCAGGACG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGGGCCU 3473
332 GCCCUGGC G UCCUGAUG 1174 CAUCAGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCCAGGGC 3474
348 GCCCCCAA G CUCCCUCU 1175 AGAGGGAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGGGGGC 3475
365 CCUGAGAA G CCACCAGC 1176 GCUGGUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCUCAGG 3476
372 AGCCACCA G CACCACCC 1177 GGGUGGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGUGGCU 3477
391 ACUUGGGG G CAGGCGCC 1178 GGCGCCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCCAAGU 3478
395 GGGGGCAG G CGCCAGGG 1179 CCCUGGCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGCCCCC 3479
410 GGACGGAC G UGGGCCAG 1180 CUGGCCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUCCGUCC 3480
414 GGACGUGG G CCAGUGCG 1181 CGCACUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCACGUCC 3481
418 GUGGGCCA G UGCGAGCC 1182 GGCUCGCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGCCCAC 3482
424 CAGUGCGA G CCCAGAGG 1183 CCUCUGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCGCACUG 3483
433 CCCAGAGG G CCCGAAGG 1184 CCUUCGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCUCUGGG 3484
441 GCCCGAAG G CCGGGGCC 1185 GGCCCCGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUCGGGC 3485
447 AGGCCGGG G CCCACCAU 1186 AUGGUGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCGGCCU 3486
457 CCACCAUG G CCCAAGCC 1187 GGCUUGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUGGUGG 3487
463 UGGCCCAA G CCCUGCCC 1188 GGGCAGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGGGCCA 3488
474 CUGCCCUG G CUCCUGCU 1189 AGCAGGAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGGGCAG 3489
491 GUGGAUGG G CGCGGGAG 1190 CUCCCGCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAUCCAC 3490
499 GCGCGGGA G UGCUGCCU 1191 AGGCAGCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCCGCGC 3491
515 UGCCCACG G CACCCAGC 1192 GCUGGGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGUGGGCA 3492
522 GGCACCCA G CACGGCAU 1193 AUGCCGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGGUGCC 3493
Table 23
527 CCAGCACG G CAUCCGGC 1194 GCCGGAUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGUGCUGG 3494
534 GGCAUCCG G CUGCCCCU 1195 AGGGGCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGGAUGCC 3495
548 CCUGCGCA G CGGCCUGG 1196 CCAGGCCG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCGCAGG 3496
551 GCGCAGCG G CCUGGGGG 1197 CCCCCAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGCUGCGC 3497
560 CCUGGGGG G CGCCCCCC 1198 GGGGGGCG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCCCAGG 3498
573 CCCCUGGG G CUGCGGCU 1199 AGCCGCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCAGGGG 3499
579 GGGCUGCG G CUGCCCCG 1200 CGGGGCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGCAGCCC 3500
603 GACGAAGA G CCCGAGGA 1201 UCCUCGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCUUCGUC 3501
612 CCCGAGGA G CCCGGCCG 1202 CGGCCGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCCUCGGG 3502
617 GGAGCCCG G CCGGAGGG 1203 CCCUCCGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGGGCUCC 3503
626 CCGGAGGG G CAGCUUUG 1204 CAAAGCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCUCCGG 3504
629 GAGGGGCA G CUUUGUGG 1205 CCACAAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCCCCUC 3505
643 UGGAGAUG G UGGACAAC 1206 GUUGUCCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUCUCCA 3506
659 CCUGAGGG G CAAGUCGG 1207 CCGACUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCUCAGG 3507
663 AGGGGCAA G UCGGGGCA 1208 UGCCCCGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGCCCCU 3508
669 AAGUCGGG G CAGGGCUA 1209 UAGCCCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCGACUU 3509
674 GGGGCAGG G CUACUACG 1210 CGUAGUAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCUGCCCC 3510
682 GCUACUAC G UGGAGAUG 1211 CAUCUCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUAGUAGC 3511
694 AGAUGACC G UGGGCAGC 1212 GCUGCCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGUCAUCU 3512
698 GACCGUGG G CAGCCCCC 1213 GGGGGCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCACGGUC 3513
701 CGUGGGCA G CCCCCCGC 1214 GCGGGGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCCCACG 3514
727 ACAUCCUG G UGGAUACA 1215 UGUAUCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGGAUGU 3515
737 GGAUACAG G CAGCAGUA 1216 UACUGCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGUAUCC 3516
740 UACAGGCA G CAGUAACU 1217 AGUUACUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCCUGUA 3517
743 AGGCAGCA G UAACUUUG 1218 CAAAGUUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCUGCCU 3518
754 ACUUUGCA G UGGGUGCU 1219 AGCACCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCAAAGU 3519
758 UGCAGUGG G UGCUGCCC 1220 GGGCAGCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCACUGCA 3520
798 UACCAGAG G CAGCUGUC 1221 GACAGCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCUGGUA 3521
801 CAGAGGCA G CUGUCCAG 1222 CUGGACAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCCUCUG 3522
809 GCUGUCCA G CACAUACC 1223 GGUAUGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGACAGC 3523
833 CCGGAAGG G UGUGUAUG 1224 CAUACACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCUUCCGG 3524
857 CACCCAGG G CAAGUGGG 1225 CCCACUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCUGGGUG 3525
861 CAGGGCAA G UGGGAAGG 1226 CCUUCCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGCCCUG 3526
873 GAAGGGGA G CUGGGCAC 1227 GUGCCCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCCCUUC 3527
Table 23
878 GGAGCUGG G CACCGACC 1228 GGUCGGUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCAGCUCC 3528
889 CCGACCUG G UAAGCAUC 1229 GAUGCUUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGGUCGG 3529
893 CCUGGUAA G CAUCCCCC 1230 GGGGGAUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUACCAGG 3530
905 CCCCCAUG G CCCCAACG 1231 CGUUGGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUGGGGG 3531
913 GCCCCAAC G UCACUGUG 1232 CACAGUGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUUGGGGC 3532
923 CACUGUGC G UGCCAACA 1233 UGUUGGCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCACAGUG 3533
957 UCAGACAA G UUCUUCAU 1234 AUGAAGAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUGUCUGA 3534
971 CAUCAACG G CUCCAACU 1235 AGUUGGAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGUUGAUG 3535
986 CUGGGAAG G CAUCCUGG 1236 CCAGGAUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUUCCCAG 3536
996 AUCCUGGG G CUGGCCUA 1237 UAGGCCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCAGGAU 3537
1000 UGGGGCUG G CCUAUGCU 1238 AGCAUAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGCCCCA 3538
1020 AUUGCCAG G CCUGACGA 1239 UCGUCAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUGGCAAU 3539
1038 UCCCUGGA G CCUUUCUU 1240 AAGAAAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCCAGGGA 3540
1057 ACUCUCUG G UAAAGCAG 1241 CUGCUUUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGAGAGU 3541
1062 CUGGUAAA G CAGACCCA 1242 UGGGUCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUACCAG 3542
1072 AGACCCAC G UUCCCAAC 1243 GUUGGGAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUGGGUCU 3543
1095 UCCCUGCA G CUUUGUGG 1244 CCACAAAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCAGGGA 3544
1103 GCUUUGUG G UGCUGGCU 1245 AGCCAGCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACAAAGC 3545
1109 UGGUGCUG G CUUCCCCC 1246 GGGGGAAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGCACCA 3546
1125 CUCAACCA G UCUGAAGU 1247 ACUUCAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGUUGAG 3547
1132 AGUCUGAA G UGCUGGCC 1248 GGCCAGCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCAGACU 3548
1138 AAGUGCUG G CCUCUGUC 1249 GACAGAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGCACUU 3549
1154 CGGAGGGA G CAUGAUCA 1250 UGAUCAUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCCUCCG 3550
1169 CAUUGGAG G UAUCGACC 1251 GGUCGAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCCAAUG 3551
1193 GUACACAG G CAGUCUCU 1252 AGAGACUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGUGUAC 3552
1196 CACAGGCA G UCUCUGGU 1253 ACCAGAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCCUGUG 3553
1203 AGUCUCUG G UAUACACC 1254 GGUGUAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGAGACU 3554
1218 CCCAUCCG G CGGGAGUG 1255 CACUCCCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGGAUGGG 3555
1224 CGGCGGGA G UGGUAUUA 1256 UAAUACCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCCGCCG 3556
1227 CGGGAGUG G UAUUAUGA 1257 UCAUAAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACUCCCG 3557
1237 AUUAUGAG G UGAUCAUU 1258 AAUGAUCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCAUAAU 3558
1252 UUGUGCGG G UGGAGAUC 1259 GAUCUCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGCACAA 3559
1293 UGCAAGGA G UACAACUA 1260 UAGUUGUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCUUGCA 3560
1310 UGACAAGA G CAUUGUGG 1261 CCACAAUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCUUGUCA 3561
Table 23
1322 UGUGGACA G UGGCACCA 1262 UGGUGCCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGUCCACA 3562
1325 GGACAGUG G CACCACCA 1263 UGGUGGUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACUGUCC 3563
1340 CAACCUUC G UUUGCCCA 1264 UGGGCAAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAAGGUUG 3564
1354 CCAAGAAA G UGUUUGAA 1265 UUCAAACA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUUCUUGG 3565
1363 UGUUUGAA G CUGCAGUC 1266 GACUGCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUCAAACA 3566
1369 AAGCUGCA G UCAAAUCC 1267 GGAUUUGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCAGCUU 3567
1384 CCAUCAAG G CAGCCUCC 1268 GGAGGCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUUGAUGG 3568
1387 UCAAGGCA G CCUCCUCC 1269 GGAGGAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCCUUGA 3569
1404 ACGGAGAA G UUCCCUGA 1270 UCAGGGAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUCUCCGU 3570
1415 CCCUGAUG G UUUCUGGC 1271 GCCAGAAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUCAGGG 3571
1422 GGUUUCUG G CUAGGAGA 1272 UCUCCUAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGAAACC 3572
1431 CUAGGAGA G CAGCUGGU 1273 ACCAGCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCUCCUAG 3573
1434 GGAGAGCA G CUGGUGUG 1274 CACACCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCUCUCC 3574
1438 AGCAGCUG G UGUGCUGG 1275 CCAGCACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGCUGCU 3575
1446 GUGUGCUG G CAAGCAGG 1276 CCUGCUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGCACAC 3576
1450 GCUGGCAA G CAGGCACC 1277 GGUGCCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGCCAGC 3577
1454 GCAAGCAG G CACCACCC 1278 GGGUGGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGCUUGC 3578
1480 UUUUCCCA G UCAUCUCA 1279 UGAGAUGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGGAAAA 3579
1502 CCUAAUGG G UGAGGUUA 1280 UAACCUCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAUUAGG 3580
1507 UGGGUGAG G UUACCAAC 1281 GUUGGUAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCACCCA 3581
1518 ACCAACCA G UCCUUCCG 1282 CGGAAGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGUUGGU 3582
1545 CUUCCGCA G CAAUACCU 1283 AGGUAUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCGGAAG 3583
1557 UACCUGCG G CCAGUGGA 1284 UCCACUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGCAGGUA 3584
1561 UGCGGCCA G UGGAAGAU 1285 AUCUUCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGCCGCA 3585
1573 AAGAUGUG G CCACGUCC 1286 GGACGUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACAUCUU 3586
1578 GUGGCCAC G UCCCAAGA 1287 UCUUGGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUGGCCAC 3587
1599 UGUUACAA G UUUGCCAU 1288 AUGGCAAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGUAACA 3588
1614 AUCUCACA G UCAUCCAC 1289 GUGGAUGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUGAGAU 3589
1625 AUCCACGG G CACUGUUA 1290 UAACAGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGUGGAU 3590
1639 UUAUGGGA G CUGUUAUC 1291 GAUAACAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCCAUAA 3591
1655 CAUGGAGG G CUUCUACG 1292 CGUAGAAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCUCCAUG 3592
1663 GCUUCUAC G UUGUCUUU 1293 AAAGACAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUAGAAGC 3593
1678 UUGAUCGG G CCCGAAAA 1294 UUUUCGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGAUCAA 3594
1694j ACGAAUUG G CUUUGCUG 1295 CAGCAAAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAUUCGU 3595
Table 23
1706 UGCUGUCA G CGCUUGCC 1296 GGCAAGCG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGACAGCA 3596
1728 CACGAUGA G UUCAGGAC 1297 GUCCUGAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCAUCGUG 3597
1738 UCAGGACG G CAGCGGUG 1298 CACCGCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGUCCUGA 3598
1741 GGACGGCA G CGGUGGAA 1299 UUCCACCG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCCGUCC 3599
1744 CGGCAGCG G UGGAAGGC 1300 GCCUUCCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGCUGCCG 3600
1751 GGUGGAAG G CCCUUUUG 1301 CAAAAGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUUCCACC 3601
1784 AGACUGUG G CUACAACA 1302 UGUUGUAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACAGUCU 3602
1809 ACAGAUGA G UCAACCCU 1303 AGGGUUGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCAUCUGU 3603
1828 UGACCAUA G CCUAUGUC 1304 GACAUAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAUGGUCA 3604
1840 AUGUCAUG G CUGCCAUC 1305 GAUGGCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUGACAU 3605
1882 GCCUCAUG G UGUGUCAG 1306 CUGACACA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUGAGGC 3606
1890 GUGUGUCA G UGGCGCUG 1307 CAGCGCCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGACACAC 3607
1893 UGUCAGUG G CGCUGCCU 1308 AGGCAGCG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACUGACA 3608
1917 CUGCGCCA G CAGCAUGA 1309 UCAUGCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGCGCAG 3609
1920 CGCCAGCA G CAUGAUGA 1310 UCAUCAUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCUGGCG 3610
1956 CUGCUGAA G UGAGGAGG 1311 CCUCCUCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCAGCAG 3611
1964 GUGAGGAG G CCCAUGGG 1312 CCCAUGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCCUCAC 3612
1972 GCCCAUGG G CAGAAGAU 1313 AUCUUCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAUGGGC 3613
2006 ACACCUCC G UGGUUCAC 1314 GUGAACCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGAGGUGU 3614
2009 CCUCCGUG G UUCACUUU 1315 AAAGUGAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACGGAGG 3615
2019 UCACUUUG G UCACAAGU 1316 ACUUGUGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAAGUGA 3616
2026 GGUCACAA G UAGGAGAC 1317 GUCUCCUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGUGACC 3617
2042 CACAGAUG G CACCUGUG 1318 CACAGGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUCUGUG 3618
2051 CACCUGUG G CCAGAGCA 1319 UGCUCUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACAGGUG 3619
2057 UGGCCAGA G CACCUCAG 1320 CUGAGGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCUGGCCA 3620
2114 AGGAAAAG G CUGGCAAG 1321 CUUGCCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUUUCCU 3621
2118 AAAGGCUG G CAAGGUGG 1322 CCACCUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGCCUUU 3622
2123 CUGGCAAG G UGGGUUCC 1323 GGAACCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUGCCAG 3623
2127 CAAGGUGG G UUCCAGGG 1324 CCCUGGAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCACCUUG 3624
2172 AGAAAGAA G CACUCUGC 1325 GCAGAGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCUUUCU 3625
2183 CUCUGCUG G CGGGAAUA 1326 UAUUCCCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGCAGAG 3626
2198 UACUCUUG G UCACCUCA 1327 UGAGGUGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAGAGUA 3627
2214 AAAUUUAA G UCGGGAAA 1328 UUUCCCGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUAAAUUU 3628
2243 AAACUUCA G CCCUGAAC 1329 GUUCAGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGAAGUUU 3629
Table 23
2288 AACCCAAA G UAUUCUUC 1330 GAAGAAUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUUGGGUU 3630
2305 UUUUCUUA G UUUCAGAA 1331 UUCUGAAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAAGAAAA 3631
2314 UUUCAGAA G UACUGGCA 1332 UGCCAGUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUCUGAAA 3632
2320 AAGUACUG G CAUCACAC 1333 GUGUGAUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGUACUU 3633
2333 ACACGCAG G UUACCUUG 1334 CAAGGUAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUGCGUGU 3634
2342 UUACCUUG G CGUGUGUC 1335 GACACACG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAAGGUAA 3635
2344 ACCUUGGC G UGUGUCCC 1336 GGGACACA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCCAAGGU 3636
2357 UCCCUGUG G UACCCUGG 1337 CCAGGGUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACAGGGA 3637
2365 GUACCCUG G CAGAGAAG 1338 CUUCUCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGGGUAC 3638
2381 GAGACCAA G CUUGUUUC 1339 GAAACAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUGGUCUC 3639
2397 CCCUGCUG G CCAAAGUC 1340 GACUUUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGCAGGG 3640
2403 UGGCCAAA G UCAGUAGG 1341 CCUACUGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUUGGCCA 3641
2407 CAAAGUCA G UAGGAGAG 1342 CUCUCCUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGACUUUG 3642
2424 GAUGCACA G UUUGCUAU 1343 AUAGCAAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGUGCAUC 3643
2463 AUAAACAA G CCUAACAU 1344 AUGUUAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGUUUAU 3644
2474 UAACAUUG G UGCAAAGA 1345 UCUUUGCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAUGUUA 3645
22 GCCCGCCC G GGAGCUGC 1449 GCAGCUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGCGGGC 3646
23 CCCGCCCG G GAGCUGCG 1450 CGCAGCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGGGCGGG 3647
24 CCGCCCGG G AGCUGCGA 1451 UCGCAGCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGGGCGG 3648
43 CGCGAGCU G GAUUAUGG 1452 CCAUAAUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCUCGCG 3649
44 GCGAGCUG G AUUAUGGU 1453 ACCAUAAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGCUCGC 3650
50 UGGAUUAU G GUGGCCUG 1454 CAGGCCAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUAAUCCA 3651
53 AUUAUGGU G GCCUGAGC 1455 GCUCAGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCAUAAU 3652
78 CAGCCGCA G GAGCCCGG 1456 CCGGGCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCGGCUG 3653
79 AGCCGCAG G AGCCCGGA 1457 UCCGGGCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGCGGCU 3654
85 AGGAGCCC G GAGCCCUU 1458 AAGGGCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGCUCCU 3655
86 GGAGCCCG G AGCCCUUG 1459 CAAGGGCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGGGCUCC 3656
119 CGCCCGCC G GGGGGACC 1460 GGUCCCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGCGGGCG 3657
120 GCCCGCCG G GGGGACCA 1461 UGGUCCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGGCGGGC 3658
121 CCCGCCGG G GGGACCAG 1462 CUGGUCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGGCGGG 3659
122 CCGCCGGG G GGACCAGG 1463 CCUGGUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCGGCGG 3660
123 CGCCGGGG G GACCAGGG 1464 CCCUGGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCCGGCG 3661
124 GCCGGGGG G ACCAGGGA 1465 UCCCUGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCCCGGC 3662
129 GGGGACCA G GGAAGCCG 1466 CGGCUUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGUCCCC 3663
Table 23
130 GGGACCAG G GAAGCCGC 1467 GCGGCUUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUGGUCCC 3664
131 GGACCAGG G AAGCCGCC 1468 GGCGGCUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCUGGUCC 3665
143 CCGCCACC G GCCCGCCA 1469 UGGCGGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGUGGCGG 3666
175 GCCCCGCC G GGAGCCCG 1470 CGGGCUCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGCGGGGC 3667
176 CCCCGCCG G GAGCCCGC 1471 GCGGGCUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGGCGGGG 3668
177 CCCGCCGG G AGCCCGCG 1472 CGCGGGCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCGGCGGG 3669
197 GCUGCCCA G GCUGGCCG 1473 CGGCCAGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGGGCAGC 3670
201 CCCAGGCU G GCCGCCGC 1474 GCGGCGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCCUGGG 3671
224 GAUGUAGC G GGCUCCGG 1475 CCGGAGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCUACAUC 3672
225 AUGUAGCG G GCUCCGGA 1476 UCCGGAGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGCUACAU 3673
231 CGGGCUCC G GAUCCCAG 1477 CUGGGAUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGAGCCCG 3674
232 GGGCUCCG G AUCCCAGC 1478 GCUGGGAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGGAGCCC 3675
265 UGCUCUGC G GAUCUCCC 1479 GGGAGAUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCAGAGCA 3676
266 GCUCUGCG G AUCUCCCC 1480 GGGGAGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGCAGAGC 3677
294 CACAGCCC G GACCCGGG 1481 CCCGGGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGCUGUG 3678
295 ACAGCCCG G ACCCGGGG 1482 CCCCGGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGGGCUGU 3679
300 CCGGACCC G GGGGCUGG 1483 CCAGCCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGUCCGG 3680
301 CGGACCCG G GGGCUGGC 1484 GCCAGCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGGGUCCG 3681
302 GGACCCGG G GGCUGGCC 1485 GGCCAGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGGGUCC 3682
303 GACCCGGG G GCUGGCCC 1486 GGGCCAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCGGGUC 3683
307 CGGGGGCU G GCCCAGGG 1487 CCCUGGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCCCCCG 3684
313 CUGGCCCA G GGCCCUGC 1488 GCAGGGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGGCCAG 3685
314 UGGCCCAG G GCCCUGCA 1489 UGCAGGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGGGCCA 3686
323 GCCCUGCA G GCCCUGGC 1490 GCCAGGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCAGGGC 3687
329 CAGGCCCU G GCGUCCUG 1491 CAGGACGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGGCCUG 3688
362 UCUCCUGA G AAGCCACC 1492 GGUGGCUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCAGGAGA 3689
382 ACCACCCA G ACUUGGGG 1493 CCCCAAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGGUGGU 3690
387 CCAGACUU G GGGGCAGG 1494 CCUGCCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGUCUGG 3691
388 CAGACUUG G GGGCAGGC 1495 GCCUGCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAGUCUG 3692
389 AGACUUGG G GGCAGGCG 1496 CGCCUGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAAGUCU 3693
390 GACUUGGG G GCAGGCGC 1497 GCGCCUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCAAGUC 3694
394 UGGGGGCA G GCGCCAGG 1498 CCUGGCGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCCCCCA 3695
401 AGGCGCCA G GGACGGAC 1499 GUCCGUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGCGCCU 3696
402 GGCGCCAG G GACGGACG 1500 CGUCCGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGGCGCC 3697
Table 23
403 GCGCCAGG G ACGGACGU 1501 ACGUCCGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCUGGCGC 3698
406 CCAGGGAC G GACGUGGG 1502 CCCACGUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUCCCUGG 3699
407 CAGGGACG G ACGUGGGC 1503 GCCCACGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGUCCCUG 3700
412 ACGGACGU G GGCCAGUG 1504 CACUGGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACGUCCGU 3701
413 CGGACGUG G GCCAGUGC 1505 GCACUGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACGUCCG 3702
429 CGAGCCCA G AGGGCCCG 1506 CGGGCCCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGGGCUCG 3703
431 AGCCCAGA G GGCCCGAA 1507 UUCGGGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCUGGGCU 3704
432 GCCCAGAG G GCCCGAAG 1508 CUUCGGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUCUGGGC 3705
440 GGCCCGAA G GCCGGGGC 1509 GCCCCGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUCGGGCC 3706
444 CGAAGGCC G GGGCCCAC 1510 GUGGGCCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGCCUUCG 3707
445 GAAGGCCG G GGCCCACC 1511 GGUGGGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGGCCUUC 3708
446 AAGGCCGG G GCCCACCA 1512 UGGUGGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCGGCCUU 3709
456 CCCACCAU G GCCCAAGC 1513 GCUUGGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUGGUGGG 3710
473 CCUGCCCU G GCUCCUGC 1514 GCAGGAGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGGCAGG 3711
485 CCUGCUGU G GAUGGGCG 1515 CGCCCAUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAGCAGG 3712
486 CUGCUGUG G AUGGGCGC 1516 GCGCCCAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACAGCAG 3713
489 CUGUGGAU G GGCGCGGG 1517 CCCGCGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCCACAG 3714
490 UGUGGAUG G GCGCGGGA 1518 UCCCGCGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUCCACA 3715
495 AUGGGCGC G GGAGUGCU 1519 AGCACUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCGCCCAU 3716
496 UGGGCGCG G GAGUGCUG 1520 CAGCACUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGCGCCCA 3717
497 GGGCGCGG G AGUGCUGC 1521 GCAGCACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGCGCCC 3718
514 CUGCCCAC G GCACCCAG 1522 CUGGGUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUGGGCAG 3719
526 CCCAGCAC G GCAUCCGG 1523 CCGGAUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUGCUGGG 3720
533 CGGCAUCC G GCUGCCCC 1524 GGGGCAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGAUGCCG 3721
550 UGCGCAGC G GCCUGGGG 1525 CCCCAGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCUGCGCA 3722
555 AGCGGCCU G GGGGGCGC 1526 GCGCCCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGCCGCU 3723
556 GCGGCCUG G GGGGCGCC 1527 GGCGCCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGGCCGC 3724
557 CGGCCUGG G GGGCGCCC 1528 GGGCGCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAGGCCG 3725
558 GGCCUGGG G GGCGCCCC 1529 GGGGCGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCAGGCC 3726
559 GCCUGGGG G GCGCCCCC 1530 GGGGGCGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCCAGGC 3727
570 GCCCCCCU G GGGCUGCG 1531 CGCAGCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGGGGGC 3728
571 CCCCCCUG G GGCUGCGG 1532 CCGCAGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGGGGGG 3729
572 CCCCCUGG G GCUGCGGC 1533 GCCGCAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAGGGGG 3730
578 GGGGCUGC G GCUGCCCC 1534 GGGGCAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCAGCCCC 3731
Table 23
587 GCUGCCCC G GGAGACCG 1535 CGGUCUCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGGGCAGC 3732
588 CUGCCCCG G GAGACCGA 1536 UCGGUCUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGGGGCAG 3733
589 UGCCCCGG G AGACCGAC 1537 GUCGGUCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCGGGGCA 3734
591 CCCCGGGA G ACCGACGA 1538 UCGUCGGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCCCGGGG 3735
601 CCGACGAA G AGCCCGAG 1539 CUCGGGCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUCGUCGG 3736
609 GAGCCCGA G GAGCCCGG 1540 CCGGGCUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCGGGCUC 3737
610 AGCCCGAG G AGCCCGGC 1541 GCCGGGCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUCGGGCU 3738
616 AGGAGCCC G GCCGGAGG 1542 CCUCCGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGGCUCCU 3739
620 GCCCGGCC G GAGGGGCA 1543 UGCCCCUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGCCGGGC 3740
621 CCCGGCCG G AGGGGCAG 1544 CUGCCCCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGGCCGGG 3741
623 CGGCCGGA G GGGCAGCU 1545 AGCUGCCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCCGGCCG 3742
624 GGCCGGAG G GGCAGCUU 1546 AAGCUGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUCCGGCC 3743
625 GCCGGAGG G GCAGCUUU 1547 AAAGCUGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCUCCGGC 3744
636 AGCUUUGU G GAGAUGGU 1548 ACCAUCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAAAGCU 3745
637 GCUUUGUG G AGAUGGUG 1549 CACCAUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACAAAGC 3746
639 UUUGUGGA G AUGGUGGA 1550 UCCACCAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCACAAA 3747
642 GUGGAGAU G GUGGACAA 1551 UUGUCCAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCUCCAC 3748
645 GAGAUGGU G GACAACCU 1552 AGGUUGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCAUCUC 3749
646 AGAUGGUG G ACAACCUG 1553 CAGGUUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACCAUCU 3750
656 CAACCUGA G GGGCAAGU 1554 ACUUGCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCAGGUUG 3751
657 AACCUGAG G GGCAAGUC 1555 GACUUGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCAGGUU 3752
658 ACCUGAGG G GCAAGUCG 1556 CGACUUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCUCAGGU 3753
666 GGCAAGUC G GGGCAGGG 1557 CCCUGCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GACUUGCC 3754
667 GCAAGUCG G GGCAGGGC 1558 GCCCUGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGACUUGC 3755
668 CAAGUCGG G GCAGGGCU 1559 AGCCCUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGACUUG 3756
672 UCGGGGCA G GGCUACUA 1560 UAGUAGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCCCCGA 3757
673 CGGGGCAG G GCUACUAC 1561 GUAGUAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGCCCCG 3758
684 UACUACGU G GAGAUGAC 1562 GUCAUCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACGUAGUA 3759
685 ACUACGUG G AGAUGACC 1563 GGUCAUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACGUAGU 3760
687 UACGUGGA G AUGACCGU 1564 ACGGUCAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCACGUA 3761
696 AUGACCGU G GGCAGCCC 1565 GGGCUGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACGGUCAU 3762
697 UGACCGUG G GCAGCCCC 1566 GGGGCUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACGGUCA 3763
711 CCCCCGCA G ACGCUCAA 1567 UUGAGCGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCGGGGG 3764
726 AACAUCCU G GUGGAUAC 1568 GUAUCCAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGAUGUU 3765
Table 23
729 AUCCUGGU G GAUACAGG 1569 CCUGUAUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACCAGGAU 3766
730 UCCUGGUG G AUACAGGC 1570. GCCUGUAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACCAGGA 3767
736 UGGAUACA G GCAGCAGU 1571 ACUGCUGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGUAUCCA 3768
756 UUUGCAGU G GGUGCUGC 1572 GCAGCACC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACUGCAAA 3769
757 UUGCAGUG G GUGCUGCC 1573 GGCAGCAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACUGCAA 3770
795 UACUACCA G AGGCAGCU 1574 AGCUGCCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGGUAGUA 3771
797 CUACCAGA G GCAGCUGU 1575 ACAGCUGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCUGGUAG 3772
818 CACAUACC G GGACCUCC 1576 GGAGGUCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGUAUGUG 3773
819 ' ACAUACCG G GACCUCCG 1577 CGGAGGUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGGUAUGU 3774
820 CAUACCGG G ACCUCCGG 1578 CCGGAGGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCGGUAUG 3775
827 GGACCUCC G GAAGGGUG 1579 CACCCUUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGAGGUCC 3776
828 GACCUCCG G AAGGGUGU 1580 ACACCCUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGGAGGUC 3777
831 CUCCGGAA G GGUGUGUA 1581 UACACACC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUCCGGAG 3778
832 UCCGGAAG G GUGUGUAU 1582 AUACACAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUCCGGA 3779
855 UACACCCA G GGCAAGUG 1583 CACUUGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGGUGUA 3780
856 ACACCCAG G GCAAGUGG 1584 CCACUUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGGGUGU 3781
863 GGGCAAGU G GGAAGGGG 1585 CCCCUUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACUUGCCC 3782
864 GGCAAGUG G GAAGGGGA 1586 UCCCCUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACUUGCC 3783
865 GCAAGUGG G AAGGGGAG 1587 cuccccuu GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCACUUGC 3784
868 AGUGGGAA G GGGAGCUG 1588 CAGCUCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCCCACU 3785
869 GUGGGAAG G GGAGCUGG 1589 CCAGCUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUCCCAC 3786
870 UGGGAAGG G GAGCUGGG 1590 CCCAGCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCUUCCCA 3787
871 GGGAAGGG G AGCUGGGC 1591 GCCCAGCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCUUCCC 3788
876 GGGGAGCU G GGCACCGA 1592 UCGGUGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCUCCCC 3789
877 GGGAGCUG G GCACCGAC 1593 GUCGGUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGCUCCC 3790
888 ACCGACCU G GUAAGCAU 1594 AUGCUUAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGUCGGU 3791
904 UCCCCCAU G GCCCCAAC 1595 GUUGGGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGGGGGA 3792
952 CUGAAUCA G ACAAGUUC 1596 GAACUUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGAUUCAG 3793
970 UCAUCAAC G GCUCCAAC 1597 GUUGGAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUUGAUGA 3794
980 CUCCAACU G GGAAGGCA 1598 UGCCUUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUUGGAG 3795
981 UCCAACUG G GAAGGCAU 1599 AUGCCUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGUUGGA 3796
982 CCAACUGG G AAGGCAUC 1600 GAUGCCUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAGUUGG 3797
985 ACUGGGAA G GCAUCCUG 1601 CAGGAUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCCCAGU 3798
993 GGCAUCCU G GGGCUGGC 1602 GCCAGCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGAUGCC 3799
Table 23
994 GCAUCCUG G GGCUGGCC 1603 GGCCAGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGGAUGC 3800
995 CAUCCUGG G GCUGGCCU 1604 AGGCCAGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCAGGAUG 3801
999 CUGGGGCU G GCCUAUGC 1605 GCAUAGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCCCCAG 3802
1011 UAUGCUGA G AUUGCCAG 1606 CUGGCAAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCAGCAUA 3803
1019 GAUUGCCA G GCCUGACG 1607 CGUCAGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGGCAAUC 3804
1035 GACUCCCU G GAGCCUUU 1608 AAAGGCUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGGAGUC 3805
1036 ACUCCCUG G AGCCUUUC 1609 GAAAGGCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGGGAGU 3806
1056 GACUCUCU G GUAAAGCA 1610 UGCUUUAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGAGAGUC 3807
1065 GUAAAGCA G ACCCACGU 1611 ACGUGGGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCUUUAC 3808
1102 AGCUUUGU G GUGCUGGC 1612 GCCAGCAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACAAAGCU 3809
1108 GUGGUGCU G GCUUCCCC 1613 GGGGAAGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCACCAC 3810
1137 GAAGUGCU G GCCUCUGU 1614 ACAGAGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCACUUC 3811
1147 CCUCUGUC G GAGGGAGC 1615 GCUCCCUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GACAGAGG 3812
1148 CUCUGUCG G AGGGAGCA 1616 UGCUCCCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGACAGAG 3813
1150 CUGUCGGA G GGAGCAUG 1617 CAUGCUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCGACAG 3814
1151 UGUCGGAG G GAGCAUGA 1618 UCAUGCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCCGACA 3815
1152 GUCGGAGG G AGCAUGAU 1619 AUCAUGCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCUCCGAC 3816
1165 UGAUCAUU G GAGGUAUC 1620 GAUACCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUGAUCA 3817
1166 GAUCAUUG G AGGUAUCG 1621 CGAUACCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAUGAUC 3818
1168 UCAUUGGA G GUAUCGAC 1622 GUCGAUAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCAAUGA 3819
1192 UGUACACA G GCAGUCUC 1623 GAGACUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUGUACA 3820
1202 CAGUCUCU G GUAUACAC 1624 GUGUAUAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAGACUG 3821
1217 ACCCAUCC G GCGGGAGU 1625 ACUCCCGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGAUGGGU 3822
1220 CAUCCGGC G GGAGUGGU 1626 ACCACUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCCGGAUG 3823
1221 AUCCGGCG G GAGUGGUA 1627 UACCACUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGCCGGAU 3824
1222 UCCGGCGG G AGUGGUAU 1628 AUACCACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGCCGGA 3825
1226 GCGGGAGU G GUAUUAUG 1629 CAUAAUAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACUCCCGC 3826
1236 UAUUAUGA G GUGAUCAU 1630 AUGAUCAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCAUAAUA 3827
1250 CAUUGUGC G GGUGGAGA 1631 UCUCCACC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCACAAUG 3828
1251 AUUGUGCG G GUGGAGAU 1632 AUCUCCAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGCACAAU 3829
1254 GUGCGGGU G GAGAUCAA 1633 UUGAUCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCCGCAC 3830
1255 UGCGGGUG G AGAUCAAU 1634 AUUGAUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACCCGCA 3831
1257 CGGGUGGA G AUCAAUGG 1635 CCAUUGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCACCCG 3832
1264 AGAUCAAU G GACAGGAU 1636 AUCCUGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUUGAUCU 3833
Table 23
1265 GAUCAAUG G ACAGGAUC 1637 GAUCCUGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUUGAUC 3834
1269 AAUGGACA G GAUCUGAA 1638 UUCAGAUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGUCCAUU 3835
1270 AUGGACAG G AUCUGAAA 1639 UUUCAGAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUGUCCAU 3836
1281 CUGAAAAU G GACUGCAA 1640 UUGCAGUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUUUUCAG 3837
1282 UGAAAAUG G ACUGCAAG 1641 CUUGCAGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUUUUCA 3838
1290 GACUGCAA G GAGUACAA 1642 UUGUACUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUGCAGUC 3839
1291 ACUGCAAG G AGUACAAC 1643 GUUGUACU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUUGCAGU 3840
1308 UAUGACAA G AGCAUUGU 1644 ACAAUGCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUGUCAUA 3841
1317 AGCAUUGU G GACAGUGG 1645 CCACUGUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACAAUGCU 3842
1318 GCAUUGUG G ACAGUGGC 1646 GCCACUGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACAAUGC 3843
1324 UGGACAGU G GCACCACC 1647 GGUGGUGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACUGUCCA 3844
1350 UUGCCCAA G AAAGUGUU 1648 AACACUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUGGGCAA 3845
1383 UCCAUCAA G GCAGCCUC 1649 GAGGCUGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUGAUGGA 3846
1398 UCCUCCAC G GAGAAGUU 1650 AACUUCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUGGAGGA 3847
1399 CCUCCACG G AGAAGUUC 1651 GAACUUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGUGGAGG 3848
1401 UCCACGGA G AAGUUCCC 1652 GGGAACUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCGUGGA 3849
1414 UCCCUGAU G GUUUCUGG 1653 CCAGAAAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCAGGGA 3850
1421 UGGUUUCU G GCUAGGAG 1654 CUCCUAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAAACCA 3851
1426 UCUGGCUA G GAGAGCAG 1655 CUGCUCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAGCCAGA 3852
1427 CUGGCUAG G AGAGCAGC 1656 GCUGCUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUAGCCAG 3853
1429 GGCUAGGA G AGCAGCUG 1657 CAGCUGCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCUAGCC 3854
1437 GAGCAGCU G GUGUGCUG 1658 CAGCACAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCUGCUC 3855
1445 GGUGUGCU G GCAAGCAG 1659 CUGCUUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCACACC 3856
1453 GGCAAGCA G GCACCACC 1660 GGUGGUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCUUGCC 3857
1466 CACCCCUU G GAACAUUU 1661 AAAUGUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGGGGUG 3858
1467 ACCCCUUG G AACAUUUU 1662 AAAAUGUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAGGGGU 3859
1500 UACCUAAU G GGUGAGGU 1663 ACCUCACC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUUAGGUA 3860
1501 ACCUAAUG G GUGAGGUU 1664 AACCUCAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUUAGGU 3861
1506 AUGGGUGA G GUUACCAA 1665 UUGGUAAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCACCCAU 3862
1556 AUACCUGC G GCCAGUGG 1666 CCACUGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCAGGUAU 3863
1563 CGGCCAGU G GAAGAUGU 1667 ACAUCUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACUGGCCG 3864
1564 GGCCAGUG G AAGAUGUG 1668 CACAUCUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACUGGCC 3865
1567 CAGUGGAA G AUGUGGCC 1669 GGCCACAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCCACUG 3866
1572 GAAGAUGU G GCCACGUC 1670 GACGUGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAUCUUC 3867
Table 23
1585 CGUCCCAA G ACGACUGU 1671 ACAGUCGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUGGGACG 3868
1623 UCAUCCAC G GGCACUGU 1672 ACAGUGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUGGAUGA 3869
1624 CAUCCACG G GCACUGUU 1673 AACAGUGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGUGGAUG 3870
1635 ACUGUUAU G GGAGCUGU 1674 ACAGCUCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUAACAGU 3871
1636 CUGUUAUG G GAGCUGUU 1675 AACAGCUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUAACAG 3872
1637 UGUUAUGG G AGCUGUUA 1676 UAACAGCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCAUAACA 3873
1650 GUUAUCAU G GAGGGCUU 1677 AAGCCCUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUGAUAAC 3874
1651 UUAUCAUG G AGGGCUUC 1678 GAAGCCCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUGAUAA 3875
1653 AUCAUGGA G GGCUUCUA 1679 UAGAAGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCCAUGAU 3876
1654 UCAUGGAG G GCUUCUAC 1680 GUAGAAGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUCCAUGA 3877
1676 CUUUGAUC G GGCCCGAA 1681 UUCGGGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAUCAAAG 3878
1677 UUUGAUCG G GCCCGAAA 1682 UUUCGGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGAUCAAA 3879
1693 AACGAAUU G GCUUUGCU 1683 AGCAAAGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAUUCGUU 3880
1733 UGAGUUCA G GACGGCAG 1684 CUGCCGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGAACUCA 3881
1734 GAGUUCAG G ACGGCAGC 1685 GCUGCCGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGAACUC 3882
1737 UUCAGGAC G GCAGCGGU 1686 ACCGCUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUCCUGAA 3883
1743 ACGGCAGC G GUGGAAGG 1687 CCUUCCAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCUGCCGU 3884
1746 GCAGCGGU G GAAGGCCC 1688 GGGCCUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCGCUGC 3885
1747 CAGCGGUG G AAGGCCCU 1689 AGGGCCUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACCGCUG 3886
1750 CGGUGGAA G GCCCUUUU 1690 AAAAGGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCCACCG 3887
1767 GUCACCUU G GACAUGGA 1691 UCCAUGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGGUGAC 3888
1768 UCACCUUG G ACAUGGAA 1692 UUCCAUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAGGUGA 3889
1773 UUGGACAU G GAAGACUG 1693 CAGUCUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGUCCAA 3890
1774 UGGACAUG G AAGACUGU 1694 ACAGUCUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUGUCCA 3891
1777 ACAUGGAA G ACUGUGGC 1695 GCCACAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCCAUGU 3892
1783 AAGACUGU G GCUACAAC 1696 GUUGUAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAGUCUU 3893
1800 AUUCCACA G ACAGAUGA 1697 UCAUCUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUGGAAU 3894
1804 CACAGACA G AUGAGUCA 1698 UGACUCAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUCUGUG 3895
1839 UAUGUCAU G GCUGCCAU 1699 AUGGCAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGACAUA 3896 '
1881 UGCCUCAU G GUGUGUCA 1700 UGACACAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGAGGCA 3897
1892 GUGUCAGU G GCGCUGCC 1701 GGCAGCGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACUGACAC 3898
1960 UGAAGUGA G GAGGCCCA 1702 UGGGCCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCACUUCA 3899
1961 GAAGUGAG G AGGCCCAU 1703 AUGGGCCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCACUUC 3900
1963 AGUGAGGA G GCCCAUGG 1704 CCAUGGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCUCACU 3901
Table 23
1970 AGGCCCAU G GGCAGAAG 1705 CUUCUGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUGGGCCU 3902
1971 GGCCCAUG G GCAGAAGA 1706 UCUUCUGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUGGGCC 3903
1975 CAUGGGCA G AAGAUAGA 1707 UCUAUCUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCCCAUG 3904
1978 GGGCAGAA G AUAGAGAU 1708 AUCUCUAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUCUGCCC 3905
1982 AGAAGAUA G AGAUUCCC 1709 GGGAAUCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAUCUUCU 3906
1984 AAGAUAGA G AUUCCCCU 1710 AGGGGAAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCUAUCUU 3907
1993 AUUCCCCU G GACCACAC 1711 GUGUGGUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGGGAAU 3908
1994 UUCCCCUG G ACCACACC 1712 GGUGUGGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGGGGAA 3909
2008 ACCUCCGU G GUUCACUU 1713 AAGUGAAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACGGAGGU 3910
2018 UUCACUUU G GUCACAAG 1714 CUUGUGAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAGUGAA 3911
2029 CACAAGUA G GAGACACA 1715 UGUGUCUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UACUUGUG 3912
2030 ACAAGUAG G AGACACAG 1716 CUGUGUCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUACUUGU 3913
2032 AAGUAGGA G ACACAGAU 1717 AUCUGUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCUACUU 3914
2038 GAGACACA G AUGGCACC 1718 GGUGCCAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUGUCUC 3915
2041 ACACAGAU G GCACCUGU 1719 ACAGGUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCUGUGU 3916
2050 GCACCUGU G GCCAGAGC 1720 GCUCUGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAGGUGC 3917
2055 UGUGGCCA G AGCACCUC 1721 GAGGUGCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGCCACA 3918
2065 GCACCUCA G GACCCUCC 1722 GGAGGGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGAGGUGC 3919
2066 CACCUCAG G ACCCUCCC 1723 GGGAGGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGAGGUG 3920
2101 GCCUUGAU G GAGAAGGA 1724 UCCUUCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCAAGGC 3921
2102 CCUUGAUG G AGAAGGAA 1725 UUCCUUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUCAAGG 3922
2104 UUGAUGGA G AAGGAAAA 1726 UUUUCCUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCAUCAA 3923
2107 AUGGAGAA G GAAAAGGC 1727 . GCCUUUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCUCCAU 3924
2108 UGGAGAAG G AAAAGGCU 1728 AGCCUUUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUCUCCA 3925
2113 AAGGAAAA G GCUGGCAA 1729 UUGCCAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUUCCUU 3926
2117 AAAAGGCU G GCAAGGUG 1730 CACCUUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCCUUUU 3927
2122 GCUGGCAA G GUGGGUUC 1731 GAACCCAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGCCAGC 3928
2125 GGCAAGGU G GGUUCCAG 1732 CUGGAACC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCUUGCC 3929
2126 GCAAGGUG G GUUCCAGG 1733 CCUGGAAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACCUUGC 3930
2133 GGGUUCCA G GGACUGUA 1734 UACAGUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGAACCC 3931
2134 GGUUCCAG G GACUGUAC 1735 GUACAGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGGAACC 3932
2135 GUUCCAGG G ACUGUACC 1736 GGUACAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCUGGAAC 3933
2148 UACCUGUA G GAAACAGA 1737 UCUGUUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UACAGGUA 3934
2149 ACCUGUAG G AAACAGAA 1738 UUCUGUUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUACAGGU 3935
Laϋi zό
2155 AGGAAACA G AAAAGAGA 1739 UCUCUUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGUUUCCU 3936
2160 ACAGAAAA G AGAAGAAA 1740 UUUCUUCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUUUCUGU 3937
2162 AGAAAAGA G AAGAAAGA 1741 UCUUUCUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCUUUUCU 3938
2165 AAAGAGAA G AAAGAAGC 1742 GCUUCUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUCUCUUU 3939
2169 AGAAGAAA G AAGCACUC 1743 GAGUGCUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUUCUUCU 3940
2182 ACUCUGCU G GCGGGAAU 1744 AUUCCCGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCAGAGU 3941
2185 CUGCUGGC G GGAAUACU 1745 AGUAUUCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCCAGCAG 3942
2186 UGCUGGCG G GAAUACUC 1746 GAGUAUUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGCCAGCA 3943
2187 GCUGGCGG G AAUACUCU 1747 AGAGUAUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCGCCAGC 3944
2197 AUACUCUU G GUCACCUC 1748 GAGGUGAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAGAGUAU 3945
2217 UUUAAGUC G GGAAAUUC 1749 GAAUUUCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GACUUAAA 3946
2218 UUAAGUCG G GAAAUUCU 1750 AGAAUUUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGACUUAA 3947
2219 UAAGUCGG G AAAUUCUG 1751 CAGAAUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCGACUUA 3948
2311 UAGUUUCA G AAGUACUG 1752 CAGUACUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGAAACUA 3949
2319 GAAGUACU G GCAUCACA 1753 UGUGAUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUACUUC 3950
2332 CACACGCA G GUUACCUU 1754 AAGGUAAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCGUGUG 3951
2341 GUUACCUU G GCGUGUGU 1755 ACACACGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGGUAAC 3952
2356 GUCCCUGU G GUACCCUG 1756 CAGGGUAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAGGGAC 3953
2364 GGUACCCU G GCAGAGAA 1757 UUCUCUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGGUACC 3954
2368 CCCUGGCA G AGAAGAGA 1758 UCUCUUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCCAGGG 3955
2370 CUGGCAGA G AAGAGACC 1759 GGUCUCUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCUGCCAG 3956
2373 GCAGAGAA G AGACCAAG 1760 CUUGGUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCUCUGC 3957
2375 AGAGAAGA G ACCAAGCU 1761 AGCUUGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCUUCUCU 3958
2396 UCCCUGCU G GCCAAAGU 1762 ACUUUGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCAGGGA 3959
2410 AGUCAGUA G GAGAGGAU 1763 AUCCUCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UACUGACU 3960
2411 GUCAGUAG G AGAGGAUG 1764 CAUCCUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUACUGAC 3961
2413 CAGUAGGA G AGGAUGCA 1765 UGCAUCCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCUACUG 3962
2415 GUAGGAGA G GAUGCACA 1766 UGUGCAUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCUCCUAC 3963
2416 UAGGAGAG G AUGCACAG 1767 CUGUGCAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCUCCUA 3964
2441 UUGCUUUA G AGACAGGG 1768 CCCUGUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAAAGCAA 3965
2443 GCUUUAGA G ACAGGGAC 1769 GUCCCUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCUAAAGC 3966
2447 UAGAGACA G GGACUGUA 1770 UACAGUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUCUCUA 3967
2448 AGAGACAG G GACUGUAU 1771 AUACAGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGUCUCU 3968
2449 GAGACAGG G ACUGUAUA 1772 UAUACAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCUGUCUC 3969
Table 23
Input Sequence = AF190725. Cut Site = G/ .
Stem Length = 8. Core Sequence = GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG
AF190725 (Homo sapiens beta-site APP cleaving enzyme (BACE) mRNA; 2526 bp)
Table 24
Table 24: Human Phospholamban (PLN) Hammerhead Ribozyme and Target Sequence
Pos Substrate Seq ID Ribozyme Rz Seq ID
16 AGAAAACU C CCCAGCUA 1 UAGCUGGG CUGAUGAG X CGAA AGUUUUCU 1137
24 CCCCAGCU A AACACCCG 2 CGGGUGUU CUGAUGAG X CGAA AGCUGGGG 1138
34 ACACCCGU A AGACUUCA 3 UGAAGUCU CUGAUGAG X CGAA ACGGGUGU 1139
40 GUAAGACU U CAUACAAC 4 GUUGUAUG CUGAUGAG X CGAA AGUCUUAC 1140
41 UAAGACUU c AUACAACA 5 UGUUGUAU CUGAUGAG X CGAA AAGUCUUA 1141
44 GACUUCAU A CAACACAA 6 UUGUGUUG CUGAUGAG X CGAA AUGAAGUC 1142
54 AACACAAU A CUCUAUAC 7 GUAUAGAG CUGAUGAG X CGAA AUUGUGUU 1143
57 ACAAUACU C UAUACUGU 8 ACAGUAUA CUGAUGAG X CGAA AGUAUUGU 1144
59 AAUACUCU A UACUGUGA 9 UCACAGUA CUGAUGAG X CGAA AGAGUAUU 1145
61 UACUCUAU A CUGUGAUG 10 CAUCACAG CUGAUGAG X CGAA AUAGAGUA 1146
72 GUGAUGAU C ACAGCUGC 11 GCAGCUGU CUGAUGAG X CGAA AUCAUCAC 1147
88 CCAAGGCU A CCUAAAAG 12 CUUUUAGG CUGAUGAG X CGAA AGCCUUGG 1148
92 GGCUACCU A AAAGAAGA 13 UCUUCUUU CUGAUGAG X CGAA AGGUAGCC 1149
105 AAGACAGU U AUCUCAUA 14 UAUGAGAU CUGAUGAG X CGAA ACUGUCUU 1150
106 AGACAGUU A UCUCAUAU 15 AUAUGAGA CUGAUGAG X CGAA AACUGUCU 1151
108 ACAGUUAU C UCAUAUUU 16 AAAUAUGA CUGAUGAG X CGAA AUAACUGU 1152
110 AGUUAUCU C AUAUUUGG 17 CCAAAUAU CUGAUGAG X CGAA AGAUAACU 1153
113 UAUCUCAU A UUUGGCUG 18 CAGCCAAA CUGAUGAG X CGAA AUGAGAUA 1154
115 UCUCAUAU U UGGCUGCC 19 GGCAGCCA CUGAUGAG X CGAA AUAUGAGA 1155
116 CUCAUAUU U GGCUGCCA 20 UGGCAGCC CUGAUGAG X CGAA AAUAUGAG 1156
128 UGCCAGCU U UUUAUCUU 21 AAGAUAAA CUGAUGAG X CGAA AGCUGGCA 1157
129 GCCAGCUU U UUAUCUUU 22 AAAGAUAA CUGAUGAG X CGAA AAGCUGGC 1158
130 CCAGCUUU U UAUCUUUC 23 GAAAGAUA CUGAUGAG X CGAA AAAGCUGG 1159
131 CAGCUUUU U AUCUUUCU 24 AGAAAGAU CUGAUGAG X CGAA AAAAGCUG 1160
132 AGCUUUUU A UCUUUCUC 25 GAGAAAGA CUGAUGAG X CGAA AAAAAGCU 1161
134 CUUUUUAU C UUUCUCUC 26 GAGAGAAA CUGAUGAG X CGAA AUAAAAAG 1162
136 UUUUAUCU U UCUCUCGA 27 UCGAGAGA CUGAUGAG X CGAA AGAUAAAA 1163
137 UUUAUCUU U CUCUCGAC 28 GUCGAGAG CUGAUGAG X CGAA AAGAUAAA 1164
138 UUAUCUUU C UCUCGACC 29 GGUCGAGA CUGAUGAG X CGAA AAAGAUAA 1165
140 AUCUUUCU C UCGACCAC 30 GUGGUCGA CUGAUGAG X CGAA AGAAAGAU 1166
142 cuuucucu C GACCACUU 31 AAGUGGUC CUGAUGAG X CGAA AGAGAAAG 1167
150 CGACCACU U AAAACUUC 32 GAAGUUUU CUGAUGAG X CGAA AGUGGUCG 1168
151 GACCACUU A AAACUUCA 33 UGAAGUUU CUGAUGAG X CGAA AAGUGGUC 1169
157 UUAAAACU U CAGACUUC 34 GAAGUCUG CUGAUGAG X CGAA AGUUUUAA 1170
158 UAAAACUU C AGACUUCC 35 GGAAGUCU CUGAUGAG X CGAA AAGUUUUA 1171
164 UUCAGACU U CCUGUCCU 36 AGGACAGG CUGAUGAG X CGAA AGUCUGAA 1172
165 UCAGACUU C CUGUCCUG 37 CAGGACAG CUGAUGAG X CGAA AAGUCUGA 1173
170 CUUCCUGU C CUGCUGGU 38 ACCAGCAG CUGAUGAG X CGAA ACAGGAAG 1174
179 CUGCUGGU A UCAUGGAG 39 CUCCAUGA CUGAUGAG X CGAA ACCAGCAG 1175
181 GCUGGUAU C AUGGAGAA 40 UUCUCCAU CUGAUGAG X CGAA AUACCAGC 1176
193 GAGAAAGU C CAAUACCU 41 AGGUAUUG CUGAUGAG X CGAA ACUUUCUC 1177
198 AGUCCAAU A CCUCACUC 42 GAGUGAGG CUGAUGAG X CGAA AUUGGACU 1178
Table 24
202 CAAUACCU C ACUCGCUC 43 GAGCGAGU CUGAUGAG X CGAA AGGUAUUG 1179
206 ACCUCACU C GCUCAGCU 44 AGCUGAGC CUGAUGAG X CGAA AGUGAGGU 1180
210 CACUCGCU C AGCUAUAA 45 UUAUAGCU CUGAUGAG X CGAA AGCGAGUG 1181
215 GCUCAGCU A UAAGAAGA 46 UCUUCUUA CUGAUGAG X CGAA AGCUGAGC 1182
217 UCAGCUAU A AGAAGAGC 47 GCUCUUCU CUGAUGAG X CGAA AUAGCUGA 1183
228 AAGAGCCU C AACCAUUG 48 CAAUGGUU CUGAUGAG X CGAA AGGCUCUU 1184
235 UCAACCAU U GAAAUGCC 49 GGCAUUUC CUGAUGAG X CGAA AUGGUUGA 1185
245 AAAUGCCU C AACAAGCA 50 UGCUUGUU CUGAUGAG X CGAA AGGCAUUU 1186
257 AAGCACGU C AAAAGCUA 51 UAGCUUUU CUGAUGAG X CGAA ACGUGCUU 1187
265 CAAAAGCU A CAGAAUCU 52 AGAUUCUG CUGAUGAG X CGAA AGCUUUUG 1188
272 UACAGAAU C UAUUUAUC 53 GAUAAAUA CUGAUGAG X CGAA AUUCUGUA 1189
274 CAGAAUCU A UUUAUCAA 54 UUGAUAAA CUGAUGAG X CGAA AGAUUCUG 1190
276 GAAUCUAU U UAUCAAUU 55 AAUUGAUA CUGAUGAG X CGAA AUAGAUUC 1191
277 AAUCUAUU U AUCAAUUU 56 AAAUUGAU CUGAUGAG X CGAA AAUAGAUU 1192
278 AUCUAUUU A UCAAUUUC 57 GAAAUUGA CUGAUGAG X CGAA AAAUAGAU 1193
280 CUAUUUAU C AAUUUCUG 58 CAGAAAUU CUGAUGAG X CGAA AUAAAUAG 1194
284 UUAUCAAU U UCUGUCUC 59 GAGACAGA CUGAUGAG X CGAA AUUGAUAA 1195
285 UAUCAAUU U CUGUCUCA 60 UGAGACAG CUGAUGAG X CGAA AAUUGAUA 1196
286 AUCAAUUU C UGUCUCAU 61 AUGAGACA CUGAUGAG X CGAA AAAUUGAU 1197
290 AUUUCUGU C UCAUCUUA 62 UAAGAUGA CUGAUGAG X CGAA ACAGAAAU 1198
292 UUCUGUCU C AUCUUAAU 63 AUUAAGAU CUGAUGAG X CGAA AGACAGAA 1199
295 UGUCUCAU C UUAAUAUG 64 CAUAUUAA CUGAUGAG X CGAA AUGAGACA 1200
297 UCUCAUCU U AAUAUGUC 65 GACAUAUU CUGAUGAG X CGAA AGAUGAGA 1201
298 CUCAUCUU A AUAUGUCU 66 AGACAUAU CUGAUGAG X CGAA AAGAUGAG 1202
301 AUCUUAAU A UGUCUCUU 67 AAGAGACA CUGAUGAG X CGAA AUUAAGAU 1203
305 UAAUAUGU C UCUUGCUG 68 CAGCAAGA CUGAUGAG X CGAA ACAUAUUA 1204
307 AUAUGUCU C UUGCUGAU 69 AUCAGCAA CUGAUGAG X CGAA AGACAUAU 1205
309 AUGUCUCU U GCUGAUCU 70 AGAUCAGC CUGAUGAG X CGAA AGAGACAU 1206
316 UUGCUGAU C UGUAUCAU 71 AUGAUACA CUGAUGAG X CGAA AUCAGCAA 1207
320 UGAUCUGU A UCAUCGUG 72 CACGAUGA CUGAUGAG X CGAA ACAGAUCA 1208
322 AUCUGUAU C AUCGUGAU 73 AUCACGAU CUGAUGAG X CGAA AUACAGAU 1209
325 UGUAUCAU C GUGAUGCU 74 AGCAUCAC CUGAUGAG X CGAA AUGAUACA 1210
334 GUGAUGCU U CUCUGAAG 75 CUUCAGAG CUGAUGAG X CGAA AGCAUCAC 1211
335 UGAUGCUU C UCUGAAGU 76 ACUUCAGA CUGAUGAG X CGAA AAGCAUCA 1212
337 AUGCUUCU C UGAAGUUC 77 GAACUUCA CUGAUGAG X CGAA AGAAGCAU 1213
344 UCUGAAGU U CUGCUACA 78 UGUAGCAG CUGAUGAG X CGAA ACUUCAGA 1214
345 CUGAAGUU C UGCUACAA 79 UUGUAGCA CUGAUGAG X CGAA AACUUCAG 1215
350 GUUCUGCU A CAACCUCU 80 AGAGGUUG CUGAUGAG X CGAA AGCAGAAC 1216
357 UACAACCU C UAGAUCUG 81 CAGAUCUA CUGAUGAG X CGAA AGGUUGUA 1217
359 CAACCUCU A GAUCUGCA 82 UGCAGAUC CUGAUGAG X CGAA AGAGGUUG 1218
363 CUCUAGAU C UGCAGCUU 83 AAGCUGCA CUGAUGAG X CGAA AUCUAGAG 1219
371 CUGCAGCU U GCCACAUC 84 GAUGUGGC CUGAUGAG X CGAA AGCUGCAG 1220
379 UGCCACAU C AGCUUAAA 85 UUUAAGCU CUGAUGAG X CGAA AUGUGGCA 1221
384 CAUCAGCU U AAAAUCUG 86 CAGAUUUU CUGAUGAG X CGAA AGCUGAUG 1222
385 AUCAGCUU A AAAUCUGU 87 ACAGAUUU CUGAUGAG X CGAA AAGCUGAU 1223
390 CUUAAAAU C UGUCAUCC 88 GGAUGACA CUGAUGAG X CGAA AUUUUAAG 1224
394 AAAUCUGU C AUCCCAUG 89 CAUGGGAU CUGAUGAG X CGAA ACAGAUUU 1225
Table 24
397 UCUGUCAU C CCAUGCAG 90 CUGCAUGG CUGAUGAG X CGAA AUGACAGA 1226
419 AAAACAAU A UUGUAUAA 91 UUAUACAA CUGAUGAG X CGAA AUUGUUUU 1227
421 AACAAUAU U GUAUAACA 92 UGUUAUAC CUGAUGAG X CGAA AUAUUGUU 1228
424 AAUAUUGU A UAACAGAC 93 GUCUGUUA CUGAUGAG X CGAA ACAAUAUU 1229
426 UAUUGUAU A ACAGACCA 94 UGGUCUGU CUGAUGAG X CGAA AUACAAUA 1230
437 AGACCACU U CCUGAGUA 95 UACUCAGG CUGAUGAG X CGAA AGUGGUCU 1231
438 GACCACUU C CUGAGUAG 96 CUACUCAG CUGAUGAG X CGAA AAGUGGUC 1232
445 UCCUGAGU A GAAGAGUU 97 AACUCUUC CUGAUGAG X CGAA ACUCAGGA 1233
453 AGAAGAGU U UCUUUGUG 98 CACAAAGA CUGAUGAG X CGAA ACUCUUCU 1234
454 GAAGAGUU U CUUUGUGA 99 UCACAAAG CUGAUGAG X CGAA AACUCUUC 1235
455 AAGAGUUU C UUUGUGAA 100 UUCACAAA CUGAUGAG X CGAA AAACUCUU 1236
457 GAGUUUCU U UGUGAAAA 101 UUUUCACA CUGAUGAG X CGAA AGAAACUC 1237
458 AGUUUCUU U GUGAAAAG 102 CUUUUCAC CUGAUGAG X CGAA AAGAAACU 1238
469 GAAAAGGU C AAGAUUAA 103 UUAAUCUU CUGAUGAG X CGAA ACCUUUUC 1239
475 GUCAAGAU U AAGACUAA 104 UUAGUCUU CUGAUGAG X CGAA AUCUUGAC 1240
476 UCAAGAUU A AGACUAAA 105 UUUAGUCU CUGAUGAG X CGAA AAUCUUGA 1241
482 UUAAGACU A AAACUUAU 106 AUAAGUUU CUGAUGAG X CGAA AGUCUUAA 1242
488 CUAAAACU U AUUGUUAC 107 GUAACAAU CUGAUGAG X CGAA AGUUUUAG 1243
489 UAAAACUU A UUGUUACC 108 GGUAACAA CUGAUGAG X CGAA AAGUUUUA 1244
491 AAACUUAU U GUUACCAU 109 AUGGUAAC CUGAUGAG X CGAA AUAAGUUU 1245
494 CUUAUUGU U ACCAUAUG 110 CAUAUGGU CUGAUGAG X CGAA ACAAUAAG 1246
495 UUAUUGUU A CCAUAUGU 111 ACAUAUGG CUGAUGAG X CGAA AACAAUAA 1247
500 GUUACCAU A UGUAUUCA 112 UGAAUACA CUGAUGAG X CGAA AUGGUAAC 1248
504 CCAUAUGU A UUCAUCUG 113 CAGAUGAA CUGAUGAG X CGAA ACAUAUGG 1249
506 AUAUGUAU U CAUCUGUU 114 AACAGAUG CUGAUGAG X CGAA AUACAUAU 1250
507 UAUGUAUU C AUCUGUUG 115 CAACAGAU CUGAUGAG X CGAA AAUACAUA 1251
510 GUAUUCAU C UGUUGGAU 116 AUCCAACA CUGAUGAG X CGAA AUGAAUAC 1252
514 UCAUCUGU U GGAUCUUG 117 CAAGAUCC CUGAUGAG X CGAA ACAGAUGA 1253
519 UGUUGGAU C UUGUAAAC 118 GUUUACAA CUGAUGAG X CGAA AUCCAACA 1254
521 UUGGAUCU U GUAAACAU 119 AUGUUUAC CUGAUGAG X CGAA AGAUCCAA 1255
524 GAUCUUGU A AACAUGAA 120 UUCAUGUU CUGAUGAG X CGAA ACAAGAUC 1256
540 AAAGGGCU U UAUUUUCA 121 UGAAAAUA CUGAUGAG X CGAA AGCCCUUU 1257
541 AAGGGCUU U AUUUUCAA 122 UUGAAAAU CUGAUGAG X CGAA AAGCCCUU 1258
542 AGGGCUUU A UUUUCAAA 123 UUUGAAAA CUGAUGAG X CGAA AAAGCCCU 1259
544 GGCUUUAU U UUCAAAAA 124 UUUUUGAA CUGAUGAG X CGAA AUAAAGCC 1260
545 GCUUUAUU U UCAAAAAU 125 AUUUUUGA CUGAUGAG X CGAA AAUAAAGC 1261
546 CUUUAUUU U CAAAAAUU 126 AAUUUUUG CUGAUGAG X CGAA AAAUAAAG 1262
547 UUUAUUUU C AAAAAUUA 127 UAAUUUUU CUGAUGAG X CGAA AAAAUAAA 1263
554 UCAAAAAU U AACUUCAA 128 UUGAAGUU CUGAUGAG X CGAA AUUUUUGA 1264
555 CAAAAAUU A ACUUCAAA 129 UUUGAAGU CUGAUGAG X CGAA AAUUUUUG 1265
559 AAUUAACU U CAAAAUAA 130 UUAUUUUG CUGAUGAG X CGAA AGUUAAUU 1266
560 AUUAACUU C AAAAUAAG 131 CUUAUUUU CUGAUGAG X CGAA AAGUUAAU 1267
566 UUCAAAAU A AGUGUAUA 132 UAUACACU CUGAUGAG X CGAA AUUUUGAA 1268
572 AUAAGUGU A UAAAAUGC 133 GCAUUUUA CUGAUGAG X CGAA ACACUUAU 1269
574 AAGUGUAU A AAAUGCAA 134 UUGCAUUU CUGAUGAG X CGAA AUACACUU 1270
587 GCAACUGU U GAUUUCCU 135 AGGAAAUC CUGAUGAG X CGAA ACAGUUGC 1271
591 CUGUUGAU U UCCUCAAC 136 GUUGAGGA CUGAUGAG X CGAA AUCAACAG 1272
Table 24
592 UGUUGAUU U CCUCAACA 137 UGUUGAGG CUGAUGAG X CGAA AAUCAACA 1273
593 GUUGAUUU C CUCAACAU 138 AUGUUGAG CUGAUGAG X CGAA AAAUCAAC 1274
596 GAUUUCCU C AACAUGGC 139 GCCAUGUU CUGAUGAG X CGAA AGGAAAUC 1275
606 ACAUGGCU C ACAAAUUU 140 AAAUUUGU CUGAUGAG X CGAA AGCCAUGU 1276
613 UCACAAAU U UCUAUCCC 141 GGGAUAGA CUGAUGAG X CGAA AUUUGUGA 1277
614 CACAAAUU U CUAUCCCA 142 UGGGAUAG CUGAUGAG X CGAA AAUUUGUG 1278
615 ACAAAUUU C UAUCCCAA 143 UUGGGAUA CUGAUGAG X CGAA AAAUUUGU 1279
617 AAAUUUCU A UCCCAAAU 144 AUUUGGGA CUGAUGAG X CGAA AGAAAUUU 1280
619 AUUUCUAU C CCAAAUCU 145 AGAUUUGG CUGAUGAG X CGAA AUAGAAAU 1281
626 UCCCAAAU C UUUUCUGA 146 UCAGAAAA CUGAUGAG X CGAA AUUUGGGA 1282
628 CCAAAUCU U UUCUGAAG 147 CUUCAGAA CUGAUGAG X CGAA AGAUUUGG 1283
629 CAAAUCUU u UCUGAAGA 148 UCUUCAGA CUGAUGAG X CGAA AAGAUUUG 1284
630 AAAUCUUU u CUGAAGAU 149 AUCUUCAG CUGAUGAG X CGAA AAAGAUUU 1285
631 AAUCUUUU c UGAAGAUG 150 CAUCUUCA CUGAUGAG X CGAA AAAAGAUU 1286
646 UGAAGAGU u UAGUUUUA 151 UAAAACUA CUGAUGAG X CGAA ACUCUUCA 1287
647 GAAGAGUU u AGUUUUAA 152 UUAAAACU CUGAUGAG X CGAA AACUCUUC 1288
648 AAGAGUUU A GUUUUAAA 153 UUUAAAAC CUGAUGAG X CGAA AAACUCUU 1289
651 AGUUUAGU U UUAAAACU 154 AGUUUUAA CUGAUGAG X CGAA ACUAAACU 1290
652 GUUUAGUU U UAAAACUG 155 CAGUUUUA CUGAUGAG X CGAA AACUAAAC 1291
653 UUUAGUUU U AAAACUGC 156 GCAGUUUU CUGAUGAG X CGAA AAACUAAA 1292
654 UUAGUUUU A AAACUGCA 157 UGCAGUUU CUGAUGAG X CGAA AAAACUAA 1293
675 CAACAAGU U CACUUCAU 158 AUGAAGUG CUGAUGAG X CGAA ACUUGUUG 1294
676 AACAAGUU C ACUUCAUA 159 UAUGAAGU CUGAUGAG X CGAA AACUUGUU 1295
680 AGUUCACU U CAUAUAUA 160 UAUAUAUG CUGAUGAG X CGAA AGUGAACU 1296
681 GUUCACUU C AUAUAUAA 161 UUAUAUAU CUGAUGAG X CGAA AAGUGAAC 1297
684 CACUUCAU A UAUAAAGC 162 GCUUUAUA CUGAUGAG X CGAA AUGAAGUG 1298
686 CUUCAUAU A UAAAGCAU 163 AUGCUUUA CUGAUGAG X CGAA AUAUGAAG 1299
688 UCAUAUAU A AAGCAUUA 164 UAAUGCUU CUGAUGAG X CGAA AUAUAUGA 1300
695 UAAAGCAU U AUUUUUAC 165 GUAAAAAU CUGAUGAG X CGAA AUGCUUUA 1301
696 AAAGCAUU A UUUUUACU 166 AGUAAAAA CUGAUGAG X CGAA AAUGCUUU 1302
698 AGCAUUAU U UUUACUCU 167 AGAGUAAA CUGAUGAG X CGAA AUAAUGCU 1303
699 GCAUUAUU U UUACUCUU 168 AAGAGUAA CUGAUGAG X CGAA AAUAAUGC 1304
700 CAUUAUUU U UACUCUUU 169 AAAGAGUA CUGAUGAG X CGAA AAAUAAUG 1305
701 AUUAUUUU U ACUCUUUU 170 AAAAGAGU CUGAUGAG X CGAA AAAAUAAU 1306
702 UUAUUUUU A CUCUUUUG 171 CAAAAGAG CUGAUGAG X CGAA AAAAAUAA 1307
705 UUUUUACU C UUUUGAGG 172 CCUCAAAA CUGAUGAG X CGAA AGUAAAAA 1308
707 UUUACUCU U UUGAGGUG 173 CACCUCAA CUGAUGAG X CGAA AGAGUAAA 1309
708 UUACUCUU U UGAGGUGA 174 UCACCUCA CUGAUGAG X CGAA AAGAGUAA 1310
709 UACUCUUU U GAGGUGAA 175 UUCACCUC CUGAUGAG X CGAA AAAGAGUA 1311
719 AGGUGAAU A UAAUUUAU 176 AUAAAUUA CUGAUGAG X CGAA AUUCACCU 1312
721 GUGAAUAU A AUUUAUAU 177 AUAUAAAU CUGAUGAG X CGAA AUAUUCAC 1313
724 AAUAUAAU U UAUAUUAC 178 GUAAUAUA CUGAUGAG X CGAA AUUAUAUU 1314
725 AUAUAAUU U AUAUUACA 179 UGUAAUAU CUGAUGAG X CGAA AAUUAUAU 1315
726 UAUAAUUU A UAUUACAA 180 UUGUAAUA CUGAUGAG X CGAA AAAUUAUA 1316
728 UAAUUUAU A UUACAAUG 181 CAUUGUAA CUGAUGAG X CGAA AUAAAUUA 1317
730 AUUUAUAU U ACAAUGUA 182 UACAUUGU CUGAUGAG X CGAA AUAUAAAU 1318
731 UUUAUAUU A CAAUGUAA 183 UUACAUUG CUGAUGAG X CGAA AAUAUAAA 1319
Table 24
738 UACAAUGU A AAAGCUUC 184 GAAGCUUU CUGAUGAG X CGAA ACAUUGUA 1320
745 UAAAAGCU U CUUUAAUA 185 UAUUAAAG CUGAUGAG X CGAA AGCUUUUA 1321
746 AAAAGCUU C UUUAAUAC 186 GUAUUAAA CUGAUGAG X CGAA AAGCUUUU 1322
748 AAGCUUCU U UAAUACUA 187 UAGUAUUA CUGAUGAG X CGAA AGAAGCUU 1323
749 AGCUUCUU U AAUACUAA 188 UUAGUAUU CUGAUGAG X CGAA AAGAAGCU 1324
750 GCUUCUUU A AUACUAAG 189 CUUAGUAU CUGAUGAG X CGAA AAAGAAGC 1325
753 UCUUUAAU A CUAAGUAU 190 AUACUUAG CUGAUGAG X CGAA AUUAAAGA 1326
756 UUAAUACU A AGUAUUUU 191 AAAAUACU CUGAUGAG X CGAA AGUAUUAA 1327
760 UACUAAGU A UUUUUCAG 192 CUGAAAAA CUGAUGAG X CGAA ACUUAGUA 1328
762 CUAAGUAU U UUUCAGGU 193 ACCUGAAA CUGAUGAG X CGAA AUACUUAG 1329
763 UAAGUAUU U UUCAGGUC 194 GACCUGAA CUGAUGAG X CGAA AAUACUUA 1330
764 AAGUAUUU U UCAGGUCU 195 AGACCUGA CUGAUGAG X CGAA AAAUACUU 1331
765 AGUAUUUU U CAGGUCUU 196 AAGACCUG CUGAUGAG X CGAA AAAAUACU 1332
766 GUAUUUUU C AGGUCUUC 197 GAAGACCU CUGAUGAG X CGAA AAAAAUAC 1333
771 UUUCAGGU C UUCACCAA 198 UUGGUGAA CUGAUGAG X CGAA ACCUGAAA 1334
773 UCAGGUCU U CACCAAGU 199 ACUUGGUG CUGAUGAG X CGAA AGACCUGA 1335
774 CAGGUCUU C ACCAAGUA 200 UACUUGGU CUGAUGAG X CGAA AAGACCUG 1336
782 CACCAAGU A UCAAAGUA 201 UACUUUGA CUGAUGAG X CGAA ACUUGGUG 1337
784 CCAAGUAU C AAAGUAAU 202 AUUACUUU CUGAUGAG X CGAA AUACUUGG 1338
790 AUCAAAGU A AUAACACA 203 UGUGUUAU CUGAUGAG X CGAA ACUUUGAU 1339
793 AAAGUAAU A ACACAAAU 204 AUUUGUGU CUGAUGAG X CGAA AUUACUUU 1340
809 UGAAGUGU C AUUAUUGA 205 UGAAUAAU CUGAUGAG X CGAA ACACUUCA 1341
812 AGUGUCAU U AUUCAAAA 206 UUUUGAAU CUGAUGAG X CGAA AUGACACU 1342
813 GUGUCAUU A UUCAAAAU 207 AUUUUGAA CUGAUGAG X CGAA AAUGACAC 1343
815 GUCAUUAU U CAAAAUAG 208 CUAUUUUG CUGAUGAG X CGAA AUAAUGAC 1344
816 UCAUUAUU C AAAAUAGU 209 ACUAUUUU CUGAUGAG X CGAA AAUAAUGA 1345
822 UUCAAAAU A GUCCACUG 210 CAGUGGAC CUGAUGAG X CGAA AUUUUGAA 1346
825 AAAAUAGU C CACUGACU 211 AGUCAGUG CUGAUGAG X CGAA ACUAUUUU 1347
834 CACUGACU C CUCACAUC 212 GAUGUGAG CUGAUGAG X CGAA AGUCAGUG 1348
837 UGACUCCU C ACAUCUGU 213 ACAGAUGU CUGAUGAG X CGAA AGGAGUCA 1349
842 CCUCACAU C UGUUAUCU 214 AGAUAACA CUGAUGAG X CGAA AUGUGAGG 1350
846 ACAUCUGU U AUCUUAUU 215 AAUAAGAU CUGAUGAG X CGAA ACAGAUGU 1351
847 CAUCUGUU A UCUUAUUA 216 UAAUAAGA CUGAUGAG X CGAA AACAGAUG 1352
849 UCUGUUAU C UUAUUAUA 217 UAUAAUAA CUGAUGAG X CGAA AUAACAGA 1353
851 UGUUAUCU U AUUAUAAA 218 UUUAUAAU CUGAUGAG X CGAA AGAUAACA 1354
852 GUUAUCUU A UUAUAAAG 219 CUUUAUAA CUGAUGAG X CGAA AAGAUAAC 1355
854 UAUCUUAU U AUAAAGAA 220 UUCUUUAU CUGAUGAG X CGAA AUAAGAUA 1356
855 AUCUUAUU A UAAAGAAC 221 GUUCUUUA CUGAUGAG X CGAA AAUAAGAU 1357
857 CUUAUUAU A AAGAACUA 222 UAGUUCUU CUGAUGAG X CGAA AUAAUAAG 1358
865 AAAGAACU A UUUGUAGU 223 ACUACAAA CUGAUGAG X CGAA AGUUCUUU 1359
867 AGAACUAU U UGUAGUAA 224 UUACUACA CUGAUGAG X CGAA AUAGUUCU 1360
868 GAACUAUU U GUAGUAAC 225 GUUACUAC CUGAUGAG X CGAA AAUAGUUC 1361
871 CUAUUUGU A GUAACUAU 226 AUAGUUAC CUGAUGAG X CGAA ACAAAUAG 1362
874 UUUGUAGU A ACUAUCAG 227 CUGAUAGU CUGAUGAG X CGAA ACUACAAA 1363
878 UAGUAACU A UCAGAAUC 228 GAUUCUGA CUGAUGAG X CGAA AGUUACUA 1364
880 GUAACUAU C AGAAUCUA 229 UAGAUUCU CUGAUGAG X CGAA AUAGUUAC 1365
886 AUCAGAAU C UACAUUCU 230 AGAAUGUA CUGAUGAG X CGAA AUUCUGAU 1366
Table 24
888 CAGAAUCU A CAUUCUAA 231 UUAGAAUG CUGAUGAG X CGAA AGAUUCUG 1367
892 AUCUACAU U CUAAAACA 232 UGUUUUAG CUGAUGAG X CGAA AUGUAGAU 1368
893 UCUACAUU C UAAAACAG 233 CUGUUUUA CUGAUGAG X CGAA AAUGUAGA 1369
895 UACAUUCU A AAACAGAA 234 UUCUGUUU CUGAUGAG X CGAA AGAAUGUA 1370
906 ACAGAAAU U GUAUUUUU 235 AAAAAUAC CUGAUGAG X CGAA AUUUCUGU 1371
909 GAAAUUGU A UUUUUUCU 236 AGAAAAAA CUGAUGAG X CGAA ACAAUUUC 1372
911 AAUUGUAU U UUUUCUAU 237 AUAGAAAA CUGAUGAG X CGAA AUACAAUU 1373
912 AUUGUAUU U UUUCUAUG 238 CAUAGAAA CUGAUGAG X CGAA AAUACAAU 1374
913 UUGUAUUU U UUCUAUGC 239 GCAUAGAA CUGAUGAG X CGAA AAAUACAA 1375
914 UGUAUUUU U UCUAUGCC 240 GGCAUAGA CUGAUGAG X CGAA AAAAUACA 1376
915 GUAUUUUU U CUAUGCCA 241 UGGCAUAG CUGAUGAG X CGAA AAAAAUAC 1377
916 UAUUUUUU C UAUGCCAC 242 GUGGCAUA CUGAUGAG X CGAA AAAAAAUA 1378
918 UUUUUUCU A UGCCACAU 243 AUGUGGCA CUGAUGAG X CGAA AGAAAAAA 1379
927 UGCCACAU U AACAUCUU 244 AAGAUGUU CUGAUGAG X CGAA AUGUGGCA 1380
928 GCCACAUU A ACAUCUUU 245 AAAGAUGU CUGAUGAG X CGAA AAUGUGGC 1381
933 AUUAACAU C UUUUAAAG 246 CUUUAAAA CUGAUGAG X CGAA AUGUUAAU 1382
935 UAACAUCU U UUAAAGUU 247 AACUUUAA CUGAUGAG X CGAA AGAUGUUA 1383
936 AACAUCUU U UAAAGUUG 248 CAACUUUA CUGAUGAG X CGAA AAGAUGUU 1384
937 ACAUCUUU U AAAGUUGA 249 UCAACUUU CUGAUGAG X CGAA AAAGAUGU 1385
938 CAUCUUUU A AAGUUGAU 250 AUCAACUU CUGAUGAG X CGAA AAAAGAUG 1386
943 UUUAAAGU U GAUGAGAA 251 UUCUCAUC CUGAUGAG X CGAA ACUUUAAA 1387
953 AUGAGAAU C AAGUAUGG 252 CCAUACUU CUGAUGAG X CGAA AUUCUCAU 1388
958 AAUCAAGU A UGGAAAAG 253 CUUUUCCA CUGAUGAG X CGAA ACUUGAUU 1389
968 GGAAAAGU A AGGCCAUA 254 UAUGGCCU CUGAUGAG X CGAA ACUUUUCC 1390
976 AAGGCCAU A CUCUUACA 255 UGUAAGAG CUGAUGAG X CGAA AUGGCCUU 1391
979 GCCAUACU C UUACAUAA 256 UUAUGUAA CUGAUGAG X CGAA AGUAUGGC 1392
981 CAUACUCU U ACAUAAUA 257 UAUUAUGU CUGAUGAG X CGAA AGAGUAUG 1393
982 AUACUCUU A CAUAAUAA 258 UUAUUAUG CUGAUGAG X CGAA AAGAGUAU 1394
986 UCUUACAU A AUAAAAUU 259 AAUUUUAU CUGAUGAG X CGAA AUGUAAGA 1395
989 UACAUAAU A AAAUUCCU 260 AGGAAUUU CUGAUGAG X CGAA AUUAUGUA 1396
994 AAUAAAAU U CCUUUUAA 261 UUAAAAGG CUGAUGAG X CGAA AUUUUAUU 1397
995 AUAAAAUU C CUUUUAAG 262 CUUAAAAG CUGAUGAG X CGAA AAUUUUAU 1398
998 AAAUUCCU U UUAAGUAA 263 UUACUUAA CUGAUGAG X CGAA AGGAAUUU 1399
999 AAUUCCUU U UAAGUAAU 264 AUUACUUA CUGAUGAG X CGAA AAGGAAUU 1400
1000 AUUCCUUU U AAGUAAUU 265 AAUUACUU CUGAUGAG X CGAA AAAGGAAU 1401
1001 UUCCUUUU A AGUAAUUU 266 AAAUUACU CUGAUGAG X CGAA AAAAGGAA 1402
1005 UUUUAAGU A AUUUUUUC 267 GAAAAAAU CUGAUGAG X CGAA ACUUAAAA 1403
1008 UAAGUAAU U UUUUCAAA 268 UUUGAAAA CUGAUGAG X CGAA AUUACUUA 1404
1009 AAGUAAUU U UUUCAAAG 269 CUUUGAAA CUGAUGAG X CGAA AAUUACUU 1405
1010 AGUAAUUU U UUCAAAGA 270 UCUUUGAA CUGAUGAG X CGAA AAAUUACU 1406
1011 GUAAUUUU U UCAAAGAA 271 UUCUUUGA CUGAUGAG X CGAA AAAAUUAC 1407
1012 UAAUUUUU U CAAAGAAU 272 AUUCUUUG CUGAUGAG X CGAA AAAAAUUA 1408
1013 AAUUUUUU C AAAGAAUC 273 GAUUCUUU CUGAUGAG X CGAA AAAAAAUU 1409
1021 CAAAGAAU C ACAGAAUU 274 AAUUCUGU CUGAUGAG X CGAA AUUCUUUG 1410
1029 CACAGAAU U CUAGUACA 275 UGUACUAG CUGAUGAG X CGAA AUUCUGUG 1411
1030 ACAGAAUU C UAGUACAU 276 AUGUACUA CUGAUGAG X CGAA AAUUCUGU 1412
1032 AGAAUUCU A GUACAUGU 277 ACAUGUAC CUGAUGAG X CGAA AGAAUUCU 1413
Table 24
1035 AUUCUAGU A CAUGUAGG 278 CCUACAUG CUGAUGAG X CGAA ACUAGAAU 1414
1041 GUACAUGU A GGUAAAUC 279 GAUUUACC CUGAUGAG X CGAA ACAUGUAC 1415
1045 AUGUAGGU A AAUCAUAA 280 UUAUGAUU CUGAUGAG X CGAA ACCUACAU 1416
1049 AGGUAAAU C AUAAAUCU 281 AGAUUUAU CUGAUGAG X CGAA AUUUACCU 1417
1052 UAAAUCAU A AAUCUGUU 282 AACAGAUU CUGAUGAG X CGAA AUGAUUUA 1418
1056 UCAUAAAU C UGUUCUAA 283 UUAGAACA CUGAUGAG X CGAA AUUUAUGA 1419
1060 AAAUCUGU U CUAAGACA 284 UGUCUUAG CUGAUGAG X CGAA ACAGAUUU 1420
1061 AAUCUGUU C UAAGACAU 285 AUGUCUUA CUGAUGAG X CGAA AACAGAUU 1421
1063 UCUGUUCU A AGACAUAU 286 AUAUGUCU CUGAUGAG X CGAA AGAACAGA 1422
1070 UAAGACAU A UGAUCAAC 287 GUUGAUCA CUGAUGAG X CGAA AUGUCUUA 1423
1075 CAUAUGAU C AACAGAUG 288 CAUCUGUU CUGAUGAG X CGAA AUCAUAUG 1424
1096 CUGGUGGU U AAUAUGUG 289 CACAUAUU CUGAUGAG X CGAA ACCACCAG 1425
1097 UGGUGGUU A AUAUGUGA 290 UCACAUAU CUGAUGAG X CGAA AACCACCA 1426
1100 UGGUUAAU A UGUGACAG 291 CUGUCACA CUGAUGAG X CGAA AUUAACCA 1427
1115 AGUGAGAU U AGUCAUAU 292 AUAUGACU CUGAUGAG X CGAA AUCUCACU 1428
1116 GUGAGAUU A GUCAUAUC 293 GAUAUGAC CUGAUGAG X CGAA AAUCUCAC 1429
1119 AGAUUAGU C AUAUCACU 294 AGUGAUAU CUGAUGAG X CGAA ACUAAUCU 1430
1122 UUAGUCAU A UCACUAAU 295 AUUAGUGA CUGAUGAG X CGAA AUGACUAA 1431
1124 AGUCAUAU C ACUAAUAU 296 AUAUUAGU CUGAUGAG X CGAA AUAUGACU 1432
1128 AUAUCACU A AUAUACUA 297 UAGUAUAU CUGAUGAG X CGAA AGUGAUAU 1433
1131 UCACUAAU A UACUAACA 298 UGUUAGUA CUGAUGAG X CGAA AUUAGUGA 1434
1133 ACUAAUAU A CUAACAAC 299 GUUGUUAG CUGAUGAG X CGAA AUAUUAGU 1435
1136 AAUAUACU A ACAACAGA 300 UCUGUUGU CUGAUGAG X CGAA AGUAUAUU 1436
1147 AACAGAAU C UAAUCUUC 301 GAAGAUUA CUGAUGAG X CGAA AUUCUGUU 1437
1149 CAGAAUCU A AUCUUCAU 302 AUGAAGAU CUGAUGAG X CGAA AGAUUCUG 1438
1152 AAUCUAAU C UUCAUUUA 303 UAAAUGAA CUGAUGAG X CGAA AUUAGAUU 1439
1154 UCUAAUCU U CAUUUAAG 304 CUUAAAUG CUGAUGAG X CGAA AGAUUAGA 1440
1155 CUAAUCUU C AUUUAAGG 305 CCUUAAAU CUGAUGAG X CGAA AAGAUUAG 1441
1158 AUCUUCAU U UAAGGCAC 306 GUGCCUUA CUGAUGAG X CGAA AUGAAGAU 1442
1159 UCUUCAUU U AAGGCACU 307 AGUGCCUU CUGAUGAG X CGAA AAUGAAGA 1443
1160 CUUCAUUU A AGGCACUG 308 CAGUGCCU CUGAUGAG X CGAA AAAUGAAG 1444
1170 GGCACUGU A GUGAAUUA 309 UAAUUCAC CUGAUGAG X CGAA ACAGUGCC 1445
1177 UAGUGAAU U AUCUGAGC 310 GCUCAGAU CUGAUGAG X CGAA AUUCACUA 1446
1178 AGUGAAUU A UCUGAGCU 311 AGCUCAGA CUGAUGAG X CGAA AAUUCACU 1447
1180 UGAAUUAU C UGAGCUAG 312 CUAGCUCA CUGAUGAG X CGAA AUAAUUCA 1448
1187 UCUGAGCU A GAGUUACC 313 GGUAACUC CUGAUGAG X CGAA AGCUCAGA 1449
1192 GCUAGAGU U ACCUAGCU 314 AGCUAGGU CUGAUGAG X CGAA ACUCUAGC 1450
1193 CUAGAGUU A CCUAGCUU 315 AAGCUAGG CUGAUGAG X CGAA AACUCUAG 1451
1197 AGUUACCU A GCUUACCA 316 UGGUAAGC CUGAUGAG X CGAA AGGUAACU 1452
1201 ACCUAGCU U ACCAUACU 317 AGUAUGGU CUGAUGAG X CGAA AGCUAGGU 1453
1202 CCUAGCUU A CCAUACUA 318 UAGUAUGG CUGAUGAG X CGAA AAGCUAGG 1454
1207 CUUACCAU A CUAUAUCU 319 AGAUAUAG CUGAUGAG X CGAA AUGGUAAG 1455
1210 ACCAUACU A UAUCUUUG 320 CAAAGAUA CUGAUGAG X CGAA AGUAUGGU 1456
1212 CAUACUAU A UCUUUGGA 321 UCCAAAGA CUGAUGAG X CGAA AUAGUAUG 1457
1214 UACUAUAU C UUUGGAAU 322 AUUCCAAA CUGAUGAG X CGAA AUAUAGUA 1458
1216 CUAUAUCU U UGGAAUCA 323 UGAUUCCA CUGAUGAG X CGAA AGAUAUAG 1459
1217 UAUAUCUU U GGAAUCAU 324 AUGAUUCC CUGAUGAG X CGAA AAGAUAUA 1460
Table 24
1223 UUUGGAAU C AUGAAACC 325 GGUUUCAU CUGAUGAG X CGAA AUUCCAAA 1461
1233 UGAAACCU U AAGACUUC 326 GAAGUCUU CUGAUGAG X CGAA AGGUUUCA 1462
1234 GAAACCUU A AGACUUCA 327 UGAAGUCU CUGAUGAG X CGAA AAGGUUUC 1463
1240 UUAAGACU U CAGAAUGA 328 UCAUUCUG CUGAUGAG X CGAA AGUCUUAA 1464
1241 UAAGACUU C AGAAUGAU 329 AUCAUUCU CUGAUGAG X CGAA AAGUCUUA 1465
1250 AGAAUGAU U UUGCAGGU 330 ACCUGCAA CUGAUGAG X CGAA AUCAUUCU 1466
1251 GAAUGAUU U UGCAGGUU 331 AACCUGCA CUGAUGAG X CGAA AAUCAUUC 1467
1252 AAUGAUUU U GCAGGUUG 332 CAACCUGC CUGAUGAG X CGAA AAAUCAUU 1468
1259 UUGGAGGU U GUCUUCCA 333 UGGAAGAC CUGAUGAG X CGAA ACCUGCAA 1469
1262 CAGGUUGU C UUCCAUUC 334 GAAUGGAA CUGAUGAG X CGAA ACAACCUG 1470
1264 GGUUGUCU U CCAUUCCA 335 UGGAAUGG CUGAUGAG X CGAA AGACAACC 1471
1265 GUUGUCUU C CAUUCCAG 336 CUGGAAUG CUGAUGAG X CGAA AAGACAAC 1472
1269 UCUUCCAU U CCAGCCUA 337 UAGGCUGG CUGAUGAG X CGAA AUGGAAGA 1473
1270 CUUCCAUU C CAGCCUAA 338 UUAGGCUG CUGAUGAG X CGAA AAUGGAAG 1474
1277 UCCAGCCU A ACAUCCAA 339 UUGGAUGU CUGAUGAG X CGAA AGGCUGGA 1475
1282 CCUAACAU C CAAUGCAG 340 CUGCAUUG CUGAUGAG X CGAA AUGUUAGG 1476
1302 AGGAAAAU A AAAGAUUU 341 AAAUCUUU CUGAUGAG X CGAA AUUUUCCU 1477
1309 UAAAAGAU U UCCAGUGA 342 UCACUGGA CUGAUGAG X CGAA AUCUUUUA 1478
1310 AAAAGAUU U CCAGUGAC 343 GUCACUGG CUGAUGAG X CGAA AAUCUUUU 1479
1311 AAAGAUUU C CAGUGACA 344 UGUCACUG CUGAUGAG X CGAA AAAUCUUU 1480
1327 AGAAAAAU A UAUUAUCU 345 AGAUAAUA CUGAUGAG X CGAA AUUUUUCU 1481
1329 AAAAAUAU A UUAUCUCA 346 UGAGAUAA CUGAUGAG X CGAA AUAUUUUU 1482
1331 AAAUAUAU U AUCUCAAG 347 CUUGAGAU CUGAUGAG X CGAA AUAUAUUU 1483
1332 AAUAUAUU A UCUCAAGU 348 ACUUGAGA CUGAUGAG X CGAA AAUAUAUU 1484
1334 UAUAUUAU C UCAAGUAU 349 AUACUUGA CUGAUGAG X CGAA AUAAUAUA 1485
1336 UAUUAUCU C AAGUAUUU 350 AAAUACUU CUGAUGAG X CGAA AGAUAAUA 1486
1341 UCUCAAGU A UUUUUUAA 351 UUAAAAAA CUGAUGAG X CGAA ACUUGAGA 1487
1343 UCAAGUAU U UUUUAAAA 352 UUUUAAAA CUGAUGAG X CGAA AUACUUGA 1488
1344 CAAGUAUU U UUUAAAAA 353 UUUUUAAA CUGAUGAG X CGAA AAUACUUG 1489
1345 AAGUAUUU U UUAAAAAU 354 AUUUUUAA CUGAUGAG X CGAA AAAUACUU 1490
1346 AGUAUUUU U UAAAAAUA 355 UAUUUUUA CUGAUGAG X CGAA AAAAUACU 1491
1347 GUAUUUUU U AAAAAUAU 356 AUAUUUUU CUGAUGAG X CGAA AAAAAUAC 1492
1348 UAUUUUUU A AAAAUAUA 357 UAUAUUUU CUGAUGAG X CGAA AAAAAAUA 1493
1354 UUAAAAAU A UAUGAAUU 358 AAUUCAUA CUGAUGAG X CGAA AUUUUUAA 1494
1356 AAAAAUAU A UGAAUUCU 359 AGAAUUCA CUGAUGAG X CGAA AUAUUUUU 1495
1362 AUAUGAAU U CUCUCUCC 360 GGAGAGAG CUGAUGAG X CGAA AUUCAUAU 1496
1363 UAUGAAUU C UCUCUCCA 361 UGGAGAGA CUGAUGAG X CGAA AAUUCAUA 1497
1365 UGAAUUCU C UCUCCAAA 362 UUUGGAGA CUGAUGAG X CGAA AGAAUUCA 1498
1367 AAUUCUCU C UCCAAAUA 363 UAUUUGGA CUGAUGAG X CGAA AGAGAAUU 1499
1369 UUCUCUCU C CAAAUAUU 364 AAUAUUUG CUGAUGAG X CGAA AGAGAGAA 1500
1375 CUCCAAAU A UUAACUAA 365 UUAGUUAA CUGAUGAG X CGAA AUUUGGAG 1501
1377 CCAAAUAU U AACUAAUU 366 AAUUAGUU CUGAUGAG X CGAA AUAUUUGG 1502
1378 CAAAUAUU A ACUAAUUA 367 UAAUUAGU CUGAUGAG X CGAA AAUAUUUG 1503
1382 UAUUAACU A AUUAUUAG 368 CUAAUAAU CUGAUGAG X CGAA AGUUAAUA 1504
1385 UAACUAAU U AUUAGAUU 369 AAUCUAAU CUGAUGAG X CGAA AUUAGUUA 1505
1386 AACUAAUU A UUAGAUUA 370 UAAUCUAA CUGAUGAG X CGAA AAUUAGUU 1506
1388 CUAAUUAU U AGAUUAUA 371 UAUAAUCU CUGAUGAG X CGAA AUAAUUAG 1507
Table 24
1389 UAAUUAUU A GAUUAUAU 372 AUAUAAUC CUGAUGAG X CGAA AAUAAUUA 1508
1393 UAUUAGAU U AUAUUUUG 373 CAAAAUAU CUGAUGAG X CGAA AUCUAAUA 1509
1394 AUUAGAUU A UAUUUUGA 374 UCAAAAUA CUGAUGAG X CGAA AAUCUAAU 1510
1396 UAGAUUAU A UUUUGAAA 375 UUUCAAAA CUGAUGAG X CGAA AUAAUCUA 1511
1398 GAUUAUAU U UUGAAAUG 376 CAUUUCAA CUGAUGAG X CGAA AUAUAAUC 1512
1399 AUUAUAUU U UGAAAUGA 377 UCAUUUCA CUGAUGAG X CGAA AAUAUAAU 1513
1400 UUAUAUUU U GAAAUGAA 378 UUCAUUUC CUGAUGAG X CGAA AAAUAUAA 1514
1411 AAUGAACU U GUUGGCCC 379 GGGCCAAC CUGAUGAG X CGAA AGUUCAUU 1515
1414 GAACUUGU U GGCCCAUC 380 GAUGGGCC CUGAUGAG X CGAA ACAAGUUC 1516
1422 UGGCCCAU C UAUUACAU 381 AUGUAAUA CUGAUGAG X CGAA AUGGGCCA 1517
1424 GCCCAUCU A UUACAUCU 382 AGAUGUAA CUGAUGAG X CGAA AGAUGGGC 1518
1426 CCAUCUAU U ACAUCUAC 383 GUAGAUGU CUGAUGAG X CGAA AUAGAUGG 1519
1427 CAUCUAUU A CAUCUACA 384 UGUAGAUG CUGAUGAG X CGAA AAUAGAUG 1520
1431 UAUUACAU C UACAGCUG 385 CAGCUGUA CUGAUGAG X CGAA AUGUAAUA 1521
1433 UUACAUCU A CAGCUGAC 386 GUCAGCUG CUGAUGAG X CGAA AGAUGUAA 1522
1445 CUGACCCU U GAACAUGG 387 CCAUGUUC CUGAUGAG X CGAA AGGGUCAG 1523
1458 AUGGGGGU U AGGGGAGC 388 GCUCCCCU CUGAUGAG X CGAA ACCCCCAU 1524
1459 UGGGGGUU A GGGGAGCU 389 AGCUCCCC CUGAUGAG X CGAA AACCCCCA 1525
1474 CUGACAAU U CGUGGGUC 390 GACCCACG CUGAUGAG X CGAA AUUGUCAG 1526
1475 UGACAAUU C GUGGGUCC 391 GGACCCAC CUGAUGAG X CGAA AAUUGUCA 1527
1482 UCGUGGGU C CGCAAAAU 392 AUUUUGCG CUGAUGAG X CGAA ACCCACGA 1528
1491 CGCAAAAU C UUAACUAC 393 GUAGUUAA CUGAUGAG X CGAA AUUUUGCG 1529
1493 CAAAAUCU U AACUACCU 394 AGGUAGUU CUGAUGAG X CGAA AGAUUUUG 1530
1494 AAAAUCUU A ACUACCUA 395 UAGGUAGU CUGAUGAG X CGAA AAGAUUUU 1531
1498 UCUUAACU A CCUAAUAG 396 CUAUUAGG CUGAUGAG X CGAA AGUUAAGA 1532
1502 AACUACCU A AUAGCCUA 397 UAGGCUAU CUGAUGAG X CGAA AGGUAGUU 1533
1505 UACCUAAU A GCCUACUA 398 UAGUAGGC CUGAUGAG X CGAA AUUAGGUA 1534
1510 AAUAGCCU A CUAUUGAC 399 GUCAAUAG CUGAUGAG X CGAA AGGCUAUU 1535
1513 AGCCUACU A UUGACCAU 400 AUGGUCAA CUGAUGAG X CGAA AGUAGGCU 1536
1515 CCUACUAU U GACCAUAA 401 UUAUGGUC CUGAUGAG X CGAA AUAGUAGG 1537
1522 UUGACCAU A AACCUUAC 402 GUAAGGUU CUGAUGAG X CGAA AUGGUCAA 1538
1528 AUAAACCU U ACUGAUAA 403 UUAUCAGU CUGAUGAG X CGAA AGGUUUAU 1539
1529 UAAACCUU A CUGAUAAC 404 GUUAUCAG CUGAUGAG X CGAA AAGGUUUA 1540
1535 UUACUGAU A ACAUAAAC 405 GUUUAUGU CUGAUGAG X CGAA AUCAGUAA 1541
1540 GAUAACAU A AACAGUAA 406 UUACUGUU CUGAUGAG X CGAA AUGUUAUC 1542
1547 UAAACAGU A AAUUAACA 407 UGUUAAUU CUGAUGAG X CGAA ACUGUUUA 1543
1551 CAGUAAAU U AACACAUA 408 UAUGUGUU CUGAUGAG X CGAA AUUUACUG 1544
1552 AGUAAAUU A ACACAUAU 409 AUAUGUGU CUGAUGAG X CGAA AAUUUACU 1545
1559 UAACACAU A UUUUGCGU 410 ACGCAAAA CUGAUGAG X CGAA AUGUGUUA 1546
1561 ACACAUAU U UUGCGUGU 411 ACACGCAA CUGAUGAG X CGAA AUAUGUGU 1547
1562 CACAUAUU U UGCGUGUU 412 AACACGCA CUGAUGAG X CGAA AAUAUGUG 1548
1563 ACAUAUUU U GCGUGUUA 413 UAACACGC CUGAUGAG X CGAA AAAUAUGU 1549
1570 UUGCGUGU U AUAUGUAU 414 AUACAUAU CUGAUGAG X CGAA ACACGCAA 1550
1571 UGCGUGUU A UAUGUAUU 415 AAUACAUA CUGAUGAG X CGAA AACACGCA 1551
1573 CGUGUUAU A UGUAUUAU 416 AUAAUACA CUGAUGAG X CGAA AUAACACG 1552
1577 UUAUAUGU A UUAUACAC 417 GUGUAUAA CUGAUGAG X CGAA ACAUAUAA 1553
1579 AUAUGUAU U AUACACUA 418 UAGUGUAU CUGAUGAG X CGAA AUACAUAU 1554
Table 24
1580 UAUGUAUU A UACACUAU 419 AUAGUGUA CUGAUGAG X CGAA AAUACAUA 1555
1582 UGUAUUAU A CACUAUAU 420 AUAUAGUG CUGAUGAG X CGAA AUAAUACA 1556
1587 UAUACACU A UAUUCCUA 421 UAGGAAUA CUGAUGAG X CGAA AGUGUAUA 1557
1589 UACACUAU A UUCCUACA 422 UGUAGGAA CUGAUGAG X CGAA AUAGUGUA 1558
1591 CACUAUAU U CCUACAAU 423 AUUGUAGG CUGAUGAG X CGAA AUAUAGUG 1559
1592 ACUAUAUU C CUACAAUA 424 UAUUGUAG CUGAUGAG X CGAA AAUAUAGU 1560
1595 AUAUUCCU A CAAUAAAG 425 CUUUAUUG CUGAUGAG X CGAA AGGAAUAU 1561
1600 CCUACAAU A AAGUAAGC 426 GCUUACUU CUGAUGAG X CGAA AUUGUAGG 1562
1605 AAUAAAGU A AGCUAGAG 427 CUCUAGCU CUGAUGAG X CGAA ACUUUAUU 1563
1610 AGUAAGCU A GAGAAAAU 428 AUUUUCUC CUGAUGAG X CGAA AGCUUACU 1564
1621 GAAAAUGU U AUUUAGAA 429 UUCUAAAU CUGAUGAG X CGAA ACAUUUUC 1565
1622 AAAAUGUU A UUUAGAAA 430 UUUCUAAA CUGAUGAG X CGAA AACAUUUU 1566
1624 AAUGUUAU U UAGAAAAU 431 AUUUUCUA CUGAUGAG X CGAA AUAACAUU 1567
1625 AUGUUAUU U AGAAAAUC 432 GAUUUUCU CUGAUGAG X CGAA AAUAACAU 1568
1626 UGUUAUUU A GAAAAUCA 433 UGAUUUUC CUGAUGAG X CGAA AAAUAACA 1569
Input Sequence = PLN. Cut Site = UH/.
Stem Length = 8 . Core Sequence = CUGAUGAG X CGAA (X = GCCGUUAGGC or other stem II)
PLN (Homo sapiens phospholamban (PLN) mRNA.; 1635 bp)
Table 25 Table 25: Human Phospholamban (PLN) NCH Ribozyme and Target Sequence
Pos Substrate Seq ID Ribozyme Rz Seq ID
15 CAGAAAAC U AGCUAAAC 434 GUUUAGCU CUGAUGAG X CGAA IUUUUCUG 1570
17 GAAAACUC C CUAAACAC 435 GUGUUUAG CUGAUGAG X CGAA IAGUUUUC 1571
18 AAAACUCC C UAAACACC 436 GGUGUUUA CUGAUGAG X CGAA IGAGUUUU 1572
19 AAACUCCC C AAACACCC 437 GGGUGUUU CUGAUGAG X CGAA IGGAGUUU 1573
20 AACUCCCC A AACACCCG 438 CGGGUGUU CUGAUGAG X CGAA IGGGAGUU 1574
23 UCCCCAGC U ACCCGUAA 439 UUACGGGU CUGAUGAG X CGAA ICUGGGGA 1575
28 AGCUAAAC A UAAGACUU 440 AAGUCUUA CUGAUGAG X CGAA IUUUAGCU 1576
' 30 CUAAACAC C AGACUUCA 441 UGAAGUCU CUGAUGAG X CGAA IUGUUUAG 1577
31 UAAACACC C GACUUCAU 442 AUGAAGUC CUGAUGAG X CGAA IGUGUUUA 1578
39 CGUAAGAC U ACAACACA 443 UGUGUUGU CUGAUGAG X CGAA IUCUUACG 1579
42 AAGACUUC A ACACAAUA 444 UAUUGUGU CUGAUGAG X CGAA IAAGUCUU 1580
46 CUUCAUAC A AAUACUCU 445 AGAGUAUU CUGAUGAG X CGAA IUAUGAAG 1581
49 CAUACAAC A ACUCUAUA 446 UAUAGAGU CUGAUGAG X CGAA IUUGUAUG 1582
51 UACAACAC A UCUAUACU 447 AGUAUAGA CUGAUGAG X CGAA IUGUUGUA 1583
56 CACAAUAC U ACUGUGAU 448 AUCACAGU CUGAUGAG X CGAA IUAUUGUG 1584
58 CAAUACUC U UGUGAUGA 449 UCAUCACA CUGAUGAG X CGAA IAGUAUUG 1585
63 CUCUAUAC U UGAUCACA 450 UGUGAUCA CUGAUGAG X CGAA IUAUAGAG 1586
73 UGAUGAUC A UGCCAAGG 451 CCUUGGCA CUGAUGAG X CGAA IAUCAUCA 1587
75 AUGAUCAC A CCAAGGCU 452 AGCCUUGG CUGAUGAG X CGAA IUGAUCAU 1588
78 AUCACAGC U AGGCUACC 453 GGUAGCCU CUGAUGAG X CGAA ICUGUGAU 1589
81 ACAGCUGC C CUACCUAA 454 UUAGGUAG CUGAUGAG X CGAA ICAGCUGU 1590
82 CAGCUGCC A UACCUAAA 455 UUUAGGUA CUGAUGAG X CGAA IGCAGCUG 1591
87 GCCAAGGC U AAAAGAAG 456 CUUCUUUU CUGAUGAG X CGAA ICCUUGGC 1592
90 AAGGCUAC C AGAAGACA 457 UGUCUUCU CUGAUGAG X CGAA IUAGCCUU 1593
91 AGGCUACC U GAAGACAG 458 CUGUCUUC CUGAUGAG X CGAA IGUAGCCU 1594
102 AAGAAGAC A UCUCAUAU 459 AUAUGAGA CUGAUGAG X CGAA IUCUUCUU 1595
109 CAGUUAUC U UUUGGCUG 460 CAGCCAAA CUGAUGAG X CGAA IAUAACUG 1596
111 GUUAUCUC A UGGCUGCC 461 GGCAGCCA CUGAUGAG X CGAA IAGAUAAC 1597
120 UAUUUGGC U GCUUUUUA 462 UAAAAAGC CUGAUGAG X CGAA ICCAAAUA 1598
123 UUGGCUGC C UUUUAUCU 463 AGAUAAAA CUGAUGAG X CGAA ICAGCCAA 1599
124 UGGCUGCC A UUUAUCUU 464 AAGAUAAA CUGAUGAG X CGAA IGCAGCCA 1600
127 CUGCCAGC U AUCUUUCU 465 AGAAAGAU CUGAUGAG X CGAA ICUGGCAG 1601
135 UUUUUAUC U CUCGACCA 466 UGGUCGAG CUGAUGAG X CGAA IAUAAAAA 1602
139 UAUCUUUC U ACCACUUA 467 UAAGUGGU CUGAUGAG X CGAA IAAAGAUA 1603
141 UCUUUCUC U CACUUAAA 468 UUUAAGUG CUGAUGAG X CGAA IAGAAAGA 1604
146 CUCUCGAC C AAAACUUC 469 GAAGUUUU CUGAUGAG X CGAA IUCGAGAG 1605
147 UCUCGACC A AAACUUCA 470 UGAAGUUU CUGAUGAG X CGAA IGUCGAGA 1606
149 UCGACCAC U ACUUCAGA 471 UCUGAAGU CUGAUGAG X CGAA IUGGUCGA 1607
156 CUUAAAAC U ACUUCCUG 472 CAGGAAGU CUGAUGAG X CGAA IUUUUAAG 1608
159 AAAACUUC A UCCUGUCC 473 GGACAGGA CUGAUGAG X CGAA IAAGUUUU 1609
163 CUUCAGAC U GUCCUGCU 474 AGCAGGAC CUGAUGAG X CGAA IUCUGAAG 1610
166 CAGACUUC C CUGCUGGU 475 ACCAGCAG CUGAUGAG X CGAA IAAGUCUG 1611
167 AGACUUCC U UGCUGGUA 476 UACCAGCA CUGAUGAG X CGAA IGAAGUCU 1612
171 UUCCUGUC C GGUAUCAU 477 AUGAUACC CUGAUGAG X CGAA lACAGGAA 1613
Table 25
172 UCCUGUCC U GUAUCAUG 478 CAUGAUAC CUGAUGAG X CGAA IGACAGGA 1614
175 UGUCCUGC U UCAUGGAG 479 CUCCAUGA CUGAUGAG X CGAA ICAGGACA 1615
182 CUGGUAUC A GAAAGUCC 480 GGACUUUC CUGAUGAG X CGAA IAUACCAG 1616
194 AGAAAGUC C CCUCACUC 481 GAGUGAGG CUGAUGAG X CGAA IACUUUCU 1617
195 GAAAGUCC A CUCACUCG 482 CGAGUGAG CUGAUGAG X CGAA IGACUUUC 1618
200 UCCAAUAC C UCGCUCAG 483 CUGAGCGA CUGAUGAG X CGAA IUAUUGGA 1619
201 CCAAUACC U CGCUCAGC 484 GCUGAGCG CUGAUGAG X CGAA IGUAUUGG 1620
203 AAUACCUC A CUCAGCUA 485 UAGCUGAG CUGAUGAG X CGAA IAGGUAUU 1621
205 UACCUCAC U CAGCUAUA 486 UAUAGCUG CUGAUGAG X CGAA IUGAGGUA 1622
209 UCACUCGC U UAUAAGAA 487 UUCUUAUA CUGAUGAG X CGAA ICGAGUGA 1623
211 ACUCGCUC A UAAGAAGA 488 UCUUCUUA CUGAUGAG X CGAA IAGCGAGU 1624
214 CGCUCAGC U GAAGAGCC 489 GGCUCUUC CUGAUGAG X CGAA ICUGAGCG 1625
226 AGAAGAGC C CCAUUGAA 490 UUCAAUGG CUGAUGAG X CGAA ICUCUUCU 1626
227 GAAGAGCC U CAUUGAAA 491 UUUCAAUG CUGAUGAG X CGAA IGCUCUUC 1627
229 AGAGCCUC A UUGAAAUG 492 CAUUUCAA CUGAUGAG X CGAA IAGGCUCU 1628
232 GCCUCAAC C AAAUGCCU 493 AGGCAUUU CUGAUGAG X CGAA IUUGAGGC 1629
233 CCUCAACC A AAUGCCUC 494 GAGGCAUU CUGAUGAG X CGAA IGUUGAGG 1630
243 UGAAAUGC C CAAGCACG 495 CGUGCUUG CUGAUGAG X CGAA ICAUUUCA 1631
244 GAAAUGCC U AAGCACGU 496 ACGUGCUU CUGAUGAG X CGAA IGCAUUUC 1632
246 AAUGCCUC A GCACGUCA 497 UGACGUGC CUGAUGAG X CGAA IAGGCAUU 1633
249 GCCUCAAC A CGUCAAAA 498 UUUUGACG CUGAUGAG X CGAA IUUGAGGC 1634
253 CAACAAGC A AAAAGCUA 499 UAGCUUUU CUGAUGAG X CGAA ICUUGUUG 1635
258 AGCACGUC A CUACAGAA 500 UUCUGUAG CUGAUGAG X CGAA IACGUGCU 1636
264 UCAAAAGC U AAUCUAUU 501 AAUAGAUU CUGAUGAG X CGAA ICUUUUGA 1637
267 AAAGCUAC A CUAUUUAU 502 AUAAAUAG CUGAUGAG X CGAA lUAGCUUU 1638
273 ACAGAAUC U AUCAAUUU 503 AAAUUGAU CUGAUGAG X CGAA IAUUCUGU 1639
281 UAUUUAUC A CUGUCUCA 504 UGAGACAG CUGAUGAG X CGAA IAUAAAUA 1640
287 UCAAUUUC U CAUCUUAA 505 UUAAGAUG CUGAUGAG X CGAA IAAAUUGA 1641
291 uuucuσuc U UUAAUAUG 506 CAUAUUAA CUGAUGAG X CGAA IACAGAAA 1642
293 UCUGUCUC A AAUAUGUC 507 GACAUAUU CUGAUGAG X CGAA IAGACAGA 1643
296 GUCUCAUC U AUGUCUCU 508 AGAGACAU CUGAUGAG X CGAA IAUGAGAC 1644
306 AAUAUGUC U CUGAUCUG 509 CAGAUCAG CUGAUGAG X CGAA IACAUAUU 1645
308 UAUGUCUC U GAUCUGUA 510 UACAGAUC CUGAUGAG X CGAA IAGACAUA 1646
312 UCUCUUGC U UGUAUCAU 511 AUGAUACA CUGAUGAG X CGAA ICAAGAGA 1647
317 UGCUGAUC U CAUCGUGA 512 - UCACGAUG CUGAUGAG X CGAA IAUCAGCA 1648
323 UCUGUAUC A GAUGCUUC 513 GAAGCAUC CUGAUGAG X CGAA IAUACAGA 1649
333 CGUGAUGC U UGAAGUUC 514 GAACUUCA CUGAUGAG X CGAA ICAUCACG 1650
336 GAUGCUUC U AGUUCUGC 515 GCAGAACU CUGAUGAG X CGAA IAAGCAUC 1651
338 UGCUUCUC U UUCUGCUA 516 UAGCAGAA CUGAUGAG X CGAA lAGAAGCA 1652
346 UGAAGUUC U CAACCUCU 517 AGAGGUUG CUGAUGAG X CGAA lAACUUCA 1653
349 AGUUCUGC U CCUCUAGA 518 UCUAGAGG CUGAUGAG X CGAA ICAGAACU 1654
352 UCUGCUAC A CUAGAUCU 519 AGAUCUAG CUGAUGAG X CGAA lUAGCAGA 1655
355 GCUACAAC C GAUCUGCA 520 UGCAGAUC CUGAUGAG X CGAA IUUGUAGC 1656
356 CUACAACC U AUCUGCAG 521 CUGCAGAU CUGAUGAG X CGAA IGUUGUAG 1657
358 ACAACCUC U CUGCAGCU 522 AGCUGCAG CUGAUGAG X CGAA IAGGUUGU 1658
364 UCUAGAUC U CUUGCCAC 523 GUGGCAAG CUGAUGAG X CGAA IAUCUAGA 1659
367 AGAUCUGC A GCCACAUC 524 GAUGUGGC CUGAUGAG X CGAA ICAGAUCU 1660
Table 25
370 UCUGCAGC U ACAUCAGC 525 GCUGAUGU CUGAUGAG X CGAA ICUGCAGA 1661
374 CAGCUUGC C CAGCUUAA 526 UUAAGCUG CUGAUGAG X CGAA ICAAGCUG 1662
375 AGCUUGCC A AGCUUAAA 527 UUUAAGCU CUGAUGAG X CGAA IGCAAGCU 1663
377 CUUGCCAC A CUUAAAAU 528 AUUUUAAG CUGAUGAG X CGAA IUGGCAAG 1664
380 GCCACAUC A AAAAUCUG 529 CAGAUUUU CUGAUGAG X CGAA IAUGUGGC 1665
383 ACAUCAGC U AUCUGUCA 530 UGACAGAU CUGAUGAG X CGAA ICUGAUGU 1666
391 UUAAAAUC U UCCCAUGC 531 GCAUGGGA CUGAUGAG X CGAA IAUUUUAA 1667
395 AAUCUGUC A AUGCAGAC 532 GUCUGCAU CUGAUGAG X CGAA IACAGAUU 1668
398 CUGUCAUC C CAGACAGG 533 CCUGUCUG CUGAUGAG X CGAA IAUGACAG 1669
399 UGUCAUCC C AGACAGGA 534 UCCUGUCU CUGAUGAG X CGAA IGAUGACA 1670
400 GUCAUCCC A GACAGGAA 535 UUCCUGUC CUGAUGAG X CGAA IGGAUGAC 1671
404 UCCCAUGC A GGAAAACA 536 UGUUUUCC CUGAUGAG X CGAA ICAUGGGA 1672
408 AUGCAGAC A AACAAUAU 537 AUAUUGUU CUGAUGAG X CGAA IUCUGCAU 1673
416 AGGAAAAC A UGUAUAAC 538 GUUAUACA CUGAUGAG X CGAA IUUUUCCU 1674
429 UGUAUAAC A ACUUCCUG 539 CAGGAAGU CUGAUGAG X CGAA IUUAUACA 1675
433 UAACAGAC C CCUGAGUA 540 UACUCAGG CUGAUGAG X CGAA IUCUGUUA 1676
434 AACAGACC A CUGAGUAG 541 CUACUCAG CUGAUGAG X CGAA IGUCUGUU 1677
436 CAGACCAC U GAGUAGAA 542 UUCUACUC CUGAUGAG X CGAA IUGGUCUG 1678
439 ACCACUUC C UAGAAGAG 543 CUCUUCUA CUGAUGAG X CGAA IAAGUGGU 1679
440 CCACUUCC U AGAAGAGU 544 ACUCUUCU CUGAUGAG X CGAA IGAAGUGG 1680
456 AGAGUUUC U GAAAAGGU 545 ACCUUUUC CUGAUGAG X CGAA IAAACUCU 1681
470 AAAAGGUC A UAAGACUA 546 UAGUCUUA CUGAUGAG X CGAA IACCUUUU 1682
481 AUUAAGAC U CUUAUUGU 547 ACAAUAAG CUGAUGAG X CGAA IUCUUAAU 1683
487 ACUAAAAC U GUUACCAU 548 AUGGUAAC CUGAUGAG X CGAA IUUUUAGU 1684
497 AUUGUUAC C GUAUUCAU 549 AUGAAUAC CUGAUGAG X CGAA IUAACAAU 1685
498 UUGUUACC A UAUUCAUC 550 GAUGAAUA CUGAUGAG X CGAA IGUAACAA 1686
508 AUGUAUUC A UUGGAUCU 551 AGAUCCAA CUGAUGAG X CGAA IAAUACAU 1687
511 UAUUCAUC U GAUCUUGU 552 ACAAGAUC CUGAUGAG X CGAA lAUGAAUA 1688
520 GUUGGAUC U AACAUGAA 553 UUCAUGUU CUGAUGAG X CGAA IAUCCAAC 1689
528 UUGUAAAC A AAGGGCUU 554 AAGCCCUU CUGAUGAG X CGAA IUUUACAA 1690
539 AAAAGGGC U UUUCAAAA 555 UUUUGAAA CUGAUGAG X CGAA ICCCUUUU 1691
548 UUAUUUUC A UUAACUUC 556 GAAGUUAA CUGAUGAG X CGAA IAAAAUAA 1692
558 AAAUUAAC U AAUAAGUG 557 CACUUAUU CUGAUGAG X CGAA IUUAAUUU 1693
561 UUAACUUC A AAGUGUAU 558 AUACACUU CUGAUGAG X CGAA IAAGUUAA 1694
581 UAAAAUGC A UUGAUUUC 559 GAAAUCAA CUGAUGAG X CGAA ICAUUUUA 1695
584 AAUGCAAC U AUUUCCUC 560 GAGGAAAU CUGAUGAG X CGAA IUUGCAUU 1696
594 UUGAUUUC C CAUGGCUC 561 GAGCCAUG CUGAUGAG X CGAA IAAAUCAA 1697
595 UGAUUUCC U AUGGCUCA 562 UGAGCCAU CUGAUGAG X CGAA IGAAAUCA 1698
597 AUUUCCUC A GGCUCACA 563 UGUGAGCC CUGAUGAG X CGAA IAGGAAAU 1699
600 UCCUCAAC A UCACAAAU 564 AUUUGUGA CUGAUGAG X CGAA IUUGAGGA 1700
605 AACAUGGC U AAUUUCUA 565 UAGAAAUU CUGAUGAG X CGAA ICCAUGUU 1701
607 CAUGGCUC A UUUCUAUC 566 GAUAGAAA CUGAUGAG X CGAA IAGCCAUG 1702
609 UGGCUCAC A UCUAUCCC 567 GGGAUAGA CUGAUGAG X CGAA IUGAGCCA 1703
616 CAAAUUUC U CAAAUCUU 568 AAGAUUUG CUGAUGAG X CGAA IAAAUUUG 1704
620 UUUCUAUC C UCUUUUCU 569 AGAAAAGA CUGAUGAG X CGAA lAUAGAAA 1705
621 UUCUAUCC C CUUUUCUG 570 CAGAAAAG CUGAUGAG X CGAA IGAUAGAA 1706
622 UCUAUCCC A UUUUCUGA 571 UCAGAAAA CUGAUGAG X CGAA IGGAUAGA 1707
Table 25
627 CCCAAAUC U UGAAGAUG 572 CAUCUUCA CUGAUGAG X CGAA IAUUUGGG 1708
632 AUCUUUUC U AUGAAGAG 573 CUCUUCAU CUGAUGAG X CGAA IAAAAGAU 1709
659 UUUAAAAC U UGCCAACA 574 UGUUGGCA CUGAUGAG X CGAA IUUUUAAA 1710
662 AAAACUGC A CAACAAGU 575 ACUUGUUG CUGAUGAG X CGAA ICAGUUUU 1711
664 AACUGCAC U ACAAGUUC 576 GAACUUGU CUGAUGAG X CGAA IUGCAGUU 1712
667 UGCACUGC C AGUUCACU 577 AGUGAACU CUGAUGAG X CGAA ICAGUGCA 1713
668 GCACUGCC A GUUCACUU 578 AAGUGAAC CUGAUGAG X CGAA IGCAGUGC 1714
671 CUGCCAAC A CACUUCAU 579 AUGAAGUG CUGAUGAG X CGAA IUUGGCAG 1715
677 ACAAGUUC A AUAUAUAA 580 UUAUAUAU CUGAUGAG X CGAA IAACUUGU 1716
679 AAGUUCAC U AUAUAAAG 581 CUUUAUAU CUGAUGAG X CGAA IUGAACUU 1717
682 UUCACUUC A UAAAGCAU 582 AUGCUUUA CUGAUGAG X CGAA IAAGUGAA 1718
693 UAUAAAGC A UUUUACUC 583 GAGUAAAA CUGAUGAG X CGAA ICUUUAUA 1719
704 AUUUUUAC U UGAGGUGA 584 UCACCUCA CUGAUGAG X CGAA IUAAAAAU 1720
706 UUUUACUC U AGGUGAAU 585 AUUCACCU CUGAUGAG X CGAA IAGUAAAA 1721
733 UAUAUUAC A AAAAGCUU 586 AAGCUUUU CUGAUGAG X CGAA IUAAUAUA 1722
744 GUAAAAGC U UAAUACUA 587 UAGUAUUA CUGAUGAG X CGAA ICUUUUAC 1723
747 AAAGCUUC U UACUAAGU 588 ACUUAGUA CUGAUGAG X CGAA IAAGCUUU 1724
755 UUUAAUAC U AUUUUUCA 589 UGAAAAAU CUGAUGAG X CGAA IUAUUAAA 1725
767 UAUUUUUC A UUCACCAA 590 UUGGUGAA CUGAUGAG X CGAA IAAAAAUA 1726
772 UUCAGGUC U CAAGUAUC 591 GAUACUUG CUGAUGAG X CGAA IACCUGAA 1727
775 AGGUCUUC A GUAUCAAA 592 UUUGAUAC CUGAUGAG X CGAA IAAGACCU 1728
777 GUCUUCAC C AUCAAAGU 593 ACUUUGAU CUGAUGAG X CGAA IUGAAGAC 1729
778 UCUUCACC A UCAAAGUA 594 UACUUUGA CUGAUGAG X CGAA IGUGAAGA 1730
785 CAAGUAUC A AAUAACAC 595 GUGUUAUU CUGAUGAG X CGAA IAUACUUG 1731
796 GUAAUAAC A UGAAGUGU 596 ACACUUCA CUGAUGAG X CGAA IUUAUUAC 1732
798 AAUAACAC A AAGUGUCA 597 UGACACUU CUGAUGAG X CGAA IUGUUAUU 1733
810 GAAGUGUC A UCAAAAUA 598 UAUUUUGA CUGAUGAG X CGAA IACACUUC 1734
817 CAUUAUUC A AGUCCACU 599 AGUGGACU CUGAUGAG X CGAA IAAUAAUG 1735
826 AAAUAGUC C ACUCCUCA 600 UGAGGAGU CUGAUGAG X CGAA IACUAUUU 1736
827 AAUAGUCC A CUCCUCAC 601 GUGAGGAG CUGAUGAG X CGAA IGACUAUU 1737
829 UAGUCCAC U CCUCACAU 602 AUGUGAGG CUGAUGAG X CGAA lUGGACUA 1738
833 CCACUGAC U ACAUCUGU 603 ACAGAUGU CUGAUGAG X CGAA IUCAGUGG 1739
835 ACUGACUC C AUCUGUUA 604 UAACAGAU CUGAUGAG X CGAA IAGUCAGU 1740
836 CUGACUCC U UCUGUUAU 605 AUAACAGA CUGAUGAG X CGAA IGAGUCAG 1741
838 GACUCCUC A UGUUAUCU 606 AGAUAACA CUGAUGAG X CGAA IAGGAGUC 1742
840 CUCCUCAC A UUAUCUUA 607 UAAGAUAA CUGAUGAG X CGAA IUGAGGAG 1743
843 CUCACAUC U UCUUAUUA 608 UAAUAAGA CUGAUGAG X CGAA IAUGUGAG 1744
850 CUGUUAUC U AUAAAGAA 609 UUCUUUAU CUGAUGAG X CGAA IAUAACAG 1745
864 UAAAGAAC U GUAGUAAC 610 GUUACUAC CUGAUGAG X CGAA IUUCUUUA 1746
877 GUAGUAAC U GAAUCUAC 611 GUAGAUUC CUGAUGAG X CGAA IUUACUAC 1747
881 UAACUAUC A CUACAUUC 612 GAAUGUAG CUGAUGAG X CGAA IAUAGUUA 1748
887 UCAGAAUC U UCUAAAAC 613 GUUUUAGA CUGAUGAG X CGAA lAUUCUGA 1749
890 GAAUCUAC A AAAACAGA 614 UCUGUUUU CUGAUGAG X CGAA IUAGAUUC 1750
894 CUACAUUC U CAGAAAUU 615 AAUUUCUG CUGAUGAG X CGAA IAAUGUAG 1751
900 UCUAAAAC A UUGUAUUU 616 AAAUACAA CUGAUGAG X CGAA IUUUUAGA 1752
917 AUUUUUUC U CACAUUAA 617 UUAAUGUG CUGAUGAG X CGAA IAAAAAAU 1753
922 UUCUAUGC C UAACAUCU 618 AGAUGUUA CUGAUGAG X CGAA ICAUAGAA 1754
Table 25
923 UCUAUGCC A AACAUCUU 619 AAGAUGUU CUGAUGAG X CGAA IGCAUAGA 1755
925 UAUGCCAC A CAUCUUUU 620 AAAAGAUG CUGAUGAG X CGAA IUGGCAUA 1756
931 ACAUUAAC A UUAAAGUU 621 AACUUUAA CUGAUGAG X CGAA IUUAAUGU 1757
934 UUAACAUC U AAGUUGAU 622 AUCAACUU CUGAUGAG X CGAA IAUGUUAA 1758
954 UGAGAAUC A UGGAAAAG 623 CUUUUCCA CUGAUGAG X CGAA IAUUCUCA 1759
973 AGUAAGGC C UCUUACAU 624 AUGUAAGA CUGAUGAG X CGAA ICCUUACU 1760
974 GUAAGGCC A CUUACAUA 625 UAUGUAAG CUGAUGAG X CGAA IGCCUUAC 1761
978 GGCCAUAC U CAUAAUAA 626 UUAUUAUG CUGAUGAG X CGAA IUAUGGCC 1762
980 CCAUACUC U UAAUAAAA 627 UUUUAUUA CUGAUGAG X CGAA IAGUAUGG 1763
984 ACUCUUAC A AAAAUUCC 628 GGAAUUUU CUGAUGAG X CGAA IUAAGAGU 1764
996 UAAAAUUC C AAGUAAUU 629 AAUUACUU CUGAUGAG X CGAA IAAUUUUA 1765
997 AAAAUUCC U AGUAAUUU 630 AAAUUACU CUGAUGAG X CGAA IGAAUUUU 1766
1014 AUUUUUUC A AUCACAGA 631 UCUGUGAU CUGAUGAG X CGAA IAAAAAAU 1767
1022 AAAGAAUC A AUUCUAGU 632 ACUAGAAU CUGAUGAG X CGAA IAUUCUUU 1768
1024 AGAAUCAC A UCUAGUAC 633 GUACUAGA CUGAUGAG X CGAA IUGAUUCU 1769
1031 CAGAAUUC U CAUGUAGG 634 CCUACAUG CUGAUGAG X CGAA IAAUUCUG 1770
1037 UCUAGUAC A GGUAAAUC 635 GAUUUACC CUGAUGAG X CGAA IUACUAGA 1771
1050 GGUAAAUC A UCUGUUCU 636 AGAACAGA CUGAUGAG X CGAA IAUUUACC 1772
1057 CAUAAAUC U UAAGACAU 637 AUGUCUUA CUGAUGAG X CGAA IAUUUAUG 1773
1062 AUCUGUUC U CAUAUGAU 638 AUCAUAUG CUGAUGAG X CGAA IAACAGAU 1774
1068 UCUAAGAC A AUCAACAG 639 CUGUUGAU CUGAUGAG X CGAA IUCUUAGA 1775
1076 AUAUGAUC A AUGAGAAC 640 GUUCUCAU CUGAUGAG X CGAA IAUCAUAU 1776
1079 UGAUCAAC A AGAACUGG 641 CCAGUUCU CUGAUGAG X CGAA IUUGAUCA 1777
1089 AUGAGAAC U GUUAAUAU 642 AUAUUAAC CUGAUGAG X CGAA IUUCUCAU 1778
1107 UAUGUGAC A GAUUAGUC 643 GACUAAUC CUGAUGAG X CGAA lUCACAUA 1779
1120 GAUUAGUC A ACUAAUAU 644 AUAUUAGU CUGAUGAG X CGAA IACUAAUC 1780
1125 GUCAUAUC A UAUACUAA 645 UUAGUAUA CUGAUGAG X CGAA IAUAUGAC 1781
1127 CAUAUCAC U UACUAACA 646 UGUUAGUA CUGAUGAG X CGAA IUGAUAUG 1782
1135 UAAUAUAC U ACAGAAUC 647 GAUUCUGU CUGAUGAG X CGAA IUAUAUUA 1783
1139 AUACUAAC A AAUCUAAU 648 AUUAGAUU CUGAUGAG X CGAA IUUAGUAU 1784
1142 CUAACAAC A CUAAUCUU 649 AAGAUUAG CUGAUGAG X CGAA IUUGUUAG 1785
1148 ACAGAAUC U UUCAUUUA 650 UAAAUGAA CUGAUGAG X CGAA IAUUCUGU 1786
1153 AUCUAAUC U UUAAGGCA 651 UGCCUUAA CUGAUGAG X CGAA IAUUAGAU 1787
1156 UAAUCUUC A AGGCACUG 652 CAGUGCCU CUGAUGAG X CGAA IAAGAUUA 1788
1165 UUUAAGGC A AGUGAAUU 653 AAUUCACU CUGAUGAG X CGAA ICCUUAAA 1789
1167 UAAGGCAC U UGAAUUAU 654 AUAAUUCA CUGAUGAG X CGAA IUGCCUUA 1790
1181 GAAUUAUC U UAGAGUUA 655 UAACUCUA CUGAUGAG X CGAA IAUAAUUC 1791
1186 AUCUGAGC U UUACCUAG 656 CUAGGUAA CUGAUGAG X CGAA ICUCAGAU 1792
1195 AGAGUUAC C UUACCAUA 657 UAUGGUAA CUGAUGAG X CGAA IUAACUCU 1793
1196 GAGUUACC U UACCAUAC 658 GUAUGGUA CUGAUGAG X CGAA IGUAACUC 1794
1200 UACCUAGC U AUACUAUA 659 UAUAGUAU CUGAUGAG X CGAA ICUAGGUA 1795
1204 UAGCUUAC C UAUAUCUU 660 AAGAUAUA CUGAUGAG X CGAA lUAAGCUA 1796
1205 AGCUUACC A AUAUCUUU 661 AAAGAUAU CUGAUGAG X CGAA IGUAAGCU 1797
1209 UACCAUAC U CUUUGGAA 662 UUCCAAAG CUGAUGAG X CGAA IUAUGGUA 1798
1215 ACUAUAUC U AAUCAUGA 663 UCAUGAUU CUGAUGAG X CGAA IAUAUAGU 1799
1224 UUGGAAUC A ACCUUAAG 664 CUUAAGGU CUGAUGAG X CGAA IAUUCCAA 1800
1231 CAUGAAAC C GACUUCAG 665 CUGAAGUC CUGAUGAG X CGAA IUUUCAUG 1801
Table 25
1232 AUGAAACC U ACUUCAGA 666 UCUGAAGU CUGAUGAG X CGAA IGUUUCAU 1802
1239 CUUAAGAC U AAUGAUUU 667 AAAUCAUU CUGAUGAG X CGAA IUCUUAAG 1803
1242 AAGACUUC A GAUUUUGC 668 GCAAAAUC CUGAUGAG X CGAA IAAGUCUU 1804
1255 GAUUUUGC A GUCUUCCA 669 UGGAAGAC CUGAUGAG X CGAA ICAAAAUC 1805 ■
1263 AGGUUGUC U UUCCAGCC 670 GGCUGGAA CUGAUGAG X CGAA IACAACCU 1806
1266 UUGUCUUC C CAGCCUAA 671 UUAGGCUG CUGAUGAG X CGAA IAAGACAA 1807
1267 UGUCUUCC A AGCCUAAC 672 GUUAGGCU CUGAUGAG X CGAA IGAAGACA 1808
1271 UUCCAUUC C UAACAUCC 673 GGAUGUUA CUGAUGAG X CGAA IAAUGGAA 1809
1272 UCCAUUCC A AACAUCCA 674 UGGAUGUU CUGAUGAG X CGAA IGAAUGGA 1810
1275 AUUCCAGC C AUCCAAUG 675 CAUUGGAU CUGAUGAG X CGAA ICUGGAAU 1811
1276 UUCCAGCC U UCCAAUGC 676 GCAUUGGA CUGAUGAG X CGAA IGCUGGAA 1812
1280 AGCCUAAC A AUGCAGGC 677 GCCUGCAU CUGAUGAG X CGAA lUUAGGCU 1813
1283 CUAACAUC C CAGGCAAG 678 CUUGCCUG CUGAUGAG X CGAA IAUGUUAG 1814
1284 UAACAUCC A AGGCAAGG 679 CCUUGCCU CUGAUGAG X CGAA IGAUGUUA 1815
1289 UCCAAUGC A AGGAAAAU 680 AUUUUCCU CUGAUGAG X CGAA ICAUUGGA 1816
1293 AUGCAGGC A AAAUAAAA 681 UUUUAUUU CUGAUGAG X CGAA ICCUGCAU 1817
1312 AAGAUUUC C ACAGAAAA 682 uuuucuσu CUGAUGAG X CGAA IAAAUCUU 1818
1313 AGAUUUCC A CAGAAAAA 683 UUUUUCUG CUGAUGAG X CGAA IGAAAUCU 1819
1319 CCAGUGAC A AAUAUAUU 684 AAUAUAUU CUGAUGAG X CGAA IUCACUGG 1820
1335 AUAUUAUC U UAUUUUUU 685 AAAAAAUA CUGAUGAG X CGAA IAUAAUAU 1821
1337 AUUAUCUC A UUUUUUAA 686 UUAAAAAA CUGAUGAG X CGAA IAGAUAAU 1822
1364 AUGAAUUC U CCAAAUAU 687 AUAUUUGG CUGAUGAG X CGAA IAAUUCAU 1823
1366 GAAUUCUC U AAAUAUUA 688 UAAUAUUU CUGAUGAG X CGAA IAGAAUUC 1824
1368 AUUCUCUC U AUAUUAAC 689 GUUAAUAU CUGAUGAG X CGAA IAGAGAAU 1825
1370 UCUCUCUC C AUUAACUA 690 UAGUUAAU CUGAUGAG X CGAA IAGAGAGA 1826
1371 CUCUCUCC A UUAACUAA 691 UUAGUUAA CUGAUGAG X CGAA IGAGAGAG 1827
1381 AUAUUAAC U AUUAGAUU 692 AAUCUAAU CUGAUGAG X CGAA IUUAAUAU 1828
1410 AAAUGAAC U GGCCCAUC 693 GAUGGGCC CUGAUGAG X CGAA IUUCAUUU 1829
1418 UUGUUGGC C UAUUACAU 694 AUGUAAUA CUGAUGAG X CGAA ICCAACAA 1830
1419 UGUUGGCC C AUUACAUC 695 GAUGUAAU CUGAUGAG X CGAA IGCCAACA 1831
1420 GUUGGCCC A UUACAUCU 696 AGAUGUAA CUGAUGAG X CGAA IGGCCAAC 1832
1423 GGCCCAUC U CAUCUACA 697 UGUAGAUG CUGAUGAG X CGAA IAUGGGCC 1833
1429 UCUAUUAC A CAGCUGAC 698 GUCAGCUG CUGAUGAG X CGAA IUAAUAGA 1834
1432 AUUACAUC U CUGACCCU 699 AGGGUCAG CUGAUGAG X CGAA IAUGUAAU 1835
1435 ACAUCUAC A ACCCUUGA 700 UCAAGGGU CUGAUGAG X CGAA IUAGAUGU 1836
1438 UCUACAGC U CUUGAACA 701 UGUUCAAG CUGAUGAG X CGAA ICUGUAGA 1837
1442 CAGCUGAC C AACAUGGG 702 CCCAUGUU CUGAUGAG X CGAA IUCAGCUG 1838
1443 AGCUGACC C ACAUGGGG 703 CCCCAUGU CUGAUGAG X CGAA IGUCAGCU 1839
1444 GCUGACCC u CAUGGGGG 704 CCCCCAUG CUGAUGAG X CGAA IGGUCAGC 1840
1450 CCUUGAAC A GGUUAGGG 705 CCCUAACC CUGAUGAG X CGAA IUUCAAGG 1841
1467 AGGGGAGC U AUUCGUGG 706 CCACGAAU CUGAUGAG X CGAA ICUCCCCU 1842
1471 GAGCUGAC A GUGGGUCC 707 GGACCCAC CUGAUGAG X CGAA IUCAGCUC 1843
1483 CGUGGGUC C AAUCUUAA 708 UUAAGAUU CUGAUGAG X CGAA IACCCACG 1844
1486 GGGUCCGC A CUUAACUA 709 UAGUUAAG CUGAUGAG X CGAA ICGGACCC 1845
1492 GCAAAAUC U UACCUAAU 710 AUUAGGUA CUGAUGAG X CGAA IAUUUUGC 1846
1497 AUCUUAAC U AAUAGCCU 711 AGGCUAUU CUGAUGAG X CGAA lUUAAGAU 1847
1500 UUAACUAC C AGCCUACU 712 AGUAGGCU CUGAUGAG X CGAA lUAGUUAA 1848
Table 25
1501 UAACUACC U GCCUACUA 713 UAGUAGGC CUGAUGAG X CGAA IGUAGUUA 1849
1508 CUAAUAGC C AUUGACCA 714 UGGUCAAU CUGAUGAG X CGAA ICUAUUAG 1850
1509 UAAUAGCC U UUGACCAU 715 AUGGUCAA CUGAUGAG X CGAA IGCUAUUA 1851
1512 UAGCCUAC U ACCAUAAA 716 UUUAUGGU CUGAUGAG X CGAA IUAGGCUA 1852
1519 CUAUUGAC C ACCUUACU 717 AGUAAGGU CUGAUGAG X CGAA IUCAAUAG 1853
1520 UAUUGACC A CCUUACUG 718 CAGUAAGG CUGAUGAG X CGAA IGUCAAUA 1854
1526 CCAUAAAC C UGAUAACA 719 UGUUAUCA CUGAUGAG X CGAA IUUUAUGG 1855
1527 CAUAAACC U GAUAACAU 720 AUGUUAUC CUGAUGAG X CGAA IGUUUAUG 1856
1531 AACCUUAC U ACAUAAAC 721 GUUUAUGU CUGAUGAG X CGAA IUAAGGUU 1857
1538 CUGAUAAC A CAGUAAAU 722 AUUUACUG CUGAUGAG X CGAA IUUAUCAG 1858
1544 ACAUAAAC A AUUAACAC 723 GUGUUAAU CUGAUGAG X CGAA IUUUAUGU 1859
1555 AAAUUAAC A UUUUGCGU 724 ACGCAAAA CUGAUGAG X CGAA IUUAAUUU 1860
1557 AUUAACAC A UUGCGUGU 725 ACACGCAA CUGAUGAG X CGAA IUGUUAAU 1861
1584 UAUUAUAC A AUUCCUAC 726 GUAGGAAU CUGAUGAG X CGAA IUAUAAUA 1862
1586 UUAUACAC U UCCUACAA 727 UUGUAGGA CUGAUGAG X CGAA IUGUAUAA 1863
1593 CUAUAUUC C AUAAAGUA 728 UACUUUAU CUGAUGAG X CGAA IAAUAUAG 1864
1594 UAUAUUCC U UAAAGUAA 729 UUACUUUA CUGAUGAG X CGAA IGAAUAUA 1865
1597 AUUCCUAC A AGUAAGCU 730 AGCUUACU CUGAUGAG X CGAA IUAGGAAU 1866
1609 AAGUAAGC U AAAAUGUU 731 AACAUUUU CUGAUGAG X CGAA ICUUACUU 1867
Input Sequence = PLN. Cut Site = CH/.
Stem Length = 8 . Core Sequence = CUGAUGAG X CGAA (X = GCCGUUAGGC or other stem II)
PLN (Homo sapiens phospholamban (PLN) mRNA.; 1635 bp)
Table 26
Table 26: Human Phospholamban (PLN) G-cleaver Ribozyme and Target Sequence
Pos Substrate Seq ID Ribozyme Rz Seq ID
64 UCUAUACU G UGAUGAUC 732 GAUCAUCA UGAUG GCAUGCACUAUGC GCG AGUAUAGA 1868
66 UAUACUGU G AUGAUCAC 733 GUGAUCAU UGAUG GCAUGCACUAUGC GCG ACAGUAUA 1869
69 ACUGUGAU G AUCACAGC 734 GCUGUGAU UGAUG GCAUGCACUAUGC GCG AUCACAGU 1870
79 UCACAGCU G CCAAGGCU 735 AGCCUUGG UGAUG GCAUGCACUAUGC GCG AGCUGUGA 1871
121 AUUUGGCU G CCAGCUUU 736 AAAGCUGG UGAUG GCAUGCACUAUGC GCG AGCCAAAU 1872
143 UUUCUCUC G ACCACUUA 737 UAAGUGGU UGAUG GCAUGCACUAUGC GCG GAGAGAAA 1873
168 GACUUCCU G UCCUGCUG 738 CAGCAGGA UGAUG GCAUGCACUAUGC GCG AGGAAGUC 1874
173 CCUGUCCU G CUGGUAUC 739 GAUACCAG UGAUG GCAUGCACUAUGC GCG AGGACAGG 1875
207 CCUCACUC G CUCAGCUA 740 UAGCUGAG UGAUG GCAUGCACUAUGC GCG GAGUGAGG 1876
236 CAACCAUU G AAAUGCCU 741 AGGCAUUU UGAUG GCAUGCACUAUGC GCG AAUGGUUG 1877
241 AUUGAAAU G CCUCAACA 742 UGUUGAGG UGAUG GCAUGCACUAUGC GCG AUUUCAAU 1878
288 CAAUUUCU G UCUCAUCU 743 AGAUGAGA UGAUG GCAUGCACUAUGC GCG AGAAAUUG 1879
303 CUUAAUAU G UCUCUUGC 744 GCAAGAGA UGAUG GCAUGCACUAUGC GCG AUAUUAAG 1880
310 UGUCUCUU G CUGAUCUG 745 CAGAUCAG UGAUG GCAUGCACUAUGC GCG AAGAGACA 1881
313 CUCUUGCU G AUCUGUAU 746 AUACAGAU UGAUG GCAUGCACUAUGC GCG AGCAAGAG 1882
318 GCUGAUCU G UAUCAUCG 747 CGAUGAUA UGAUG GCAUGCACUAUGC GCG AGAUCAGC 1883
328 AUCAUCGU G AUGCUUCU 748 AGAAGCAU UGAUG GCAUGCACUAUGC GCG ACGAUGAU 1884
331 AUCGUGAU G CUUCUCUG 749 CAGAGAAG UGAUG GCAUGCACUAUGC GCG AUCACGAU 1885
339 GCUUCUCU G AAGUUCUG 750 CAGAACUU UGAUG GCAUGCACUAUGC GCG AGAGAAGC 1886
347 GAAGUUCU G CUACAACC 751 GGUUGUAG UGAUG GCAUGCACUAUGC GCG AGAACUUC 1887
365 CUAGAUCU G CAGCUUGC 752 GCAAGCUG UGAUG GCAUGCACUAUGC GCG AGAUCUAG 1888
372 UGCAGCUU G CCACAUCA 753 UGAUGUGG UGAUG GCAUGCACUAUGC GCG AAGCUGCA 1889
392 UAAAAUCU G UCAUCCCA 754 UGGGAUGA UGAUG GCAUGCACUAUGC GCG AGAUUUUA 1890
402 CAUCCCAU G CAGACAGG 755 CCUGUCUG UGAUG GCAUGCACUAUGC GCG AUGGGAUG 1891
422 ACAAUAUU G UAUAACAG 756 CUGUUAUA UGAUG GCAUGCACUAUGC GCG AAUAUUGU 1892
441 CACUUCCU G AGUAGAAG 757 CUUCUACU UGAUG GCAUGCACUAUGC GCG AGGAAGUG 1893
459 GUUUCUUU G UGAAAAGG 758 CCUUUUCA UGAUG GCAUGCACUAUGC GCG AAAGAAAC 1894
461 UUCUUUGU G AAAAGGUC 759 GACCUUUU UGAUG GCAUGCACUAUGC GCG ACAAAGAA 1895
Table 26
492 AACUUAUU G UUACCAUA 760 UAUGGUAA UGAUG GCAUGCACUAUGC GCG AAUAAGUU 1896
502 UACCAUAU G UAUUCAUC 761 GAUGAAUA UGAUG GCAUGCACUAUGC GCG AUAUGGUA 1897
512 AUUCAUCU G UUGGAUCU 762 AGAUCCAA UGAUG GCAUGCACUAUGC GCG AGAUGAAU 1898
522 UGGAUCUU G UAAACAUG 763 CAUGUUUA UGAUG GCAUGCACUAUGC GCG AAGAUCCA 1899
530 GUAAACAU G AAAAGGGC 764 GCCCUUUU UGAUG GCAUGCACUAUGC GCG AUGUUUAC 1900
570 AAAUAAGU G UAUAAAAU 765 AUUUUAUA UGAUG GCAUGCACUAUGC GCG ACUUAUUU 1901
579 UAUAAAAU G CAACUGUU 766 AACAGUUG UGAUG GCAUGCACUAUGC GCG AUUUUAUA 1902
585 AUGCAACU G UUGAUUUC 767 GAAAUCAA UGAUG GCAUGCACUAUGC GCG AGUUGCAU 1903
588 CAACUGUU G AUUUCCUC 768 GAGGAAAU UGAUG GCAUGCACUAUGC GCG AACAGUUG 1904
633 UCUUUUCU G AAGAUGAA 769 UUCAUCUU UGAUG GCAUGCACUAUGC GCG AGAAAAGA 1905
639 CUGAAGAU G AAGAGUUU 770 AAACUCUU UGAUG GCAUGCACUAUGC GCG AUCUUCAG 1906
660 UUAAAACU G CACUGCCA 771 UGGCAGUG UGAUG GCAUGCACUAUGC GCG AGUUUUAA 1907
665 ACUGCACU G CCAACAAG 772 CUUGUUGG UGAUG GCAUGCACUAUGC GCG AGUGCAGU 1908
710 ACUCUUUU G AGGUGAAU 773 AUUCACCU UGAUG GCAUGCACUAUGC GCG AAAAGAGU 1909
715 UUUGAGGU G AAUAUAAU 774 AUUAUAUU UGAUG GCAUGCACUAUGC GCG ACCUCAAA 1910
736 AUUACAAU G UAAAAGCU 775 AGCUUUUA UGAUG GCAUGCACUAUGC GCG AUUGUAAU 1911
802 ACACAAAU G AAGUGUCA 776 UGACACUU UGAUG GCAUGCACUAUGC GCG AUUUGUGU 1912
807 AAUGAAGU G UCAUUAUU 777 AAUAAUGA UGAUG GCAUGCACUAUGC GCG ACUUCAUU 1913
830 AGUCCACU G ACUCCUCA 778 UGAGGAGU UGAUG GCAUGCACUAUGC GCG AGUGGACU 1914
844 UCACAUCU G UUAUCUUA 779 UAAGAUAA UGAUG GCAUGCACUAUGC GCG AGAUGUGA 1915
869 AACUAUUU G UAGUAACU 780 AGUUACUA UGAUG GCAUGCACUAUGC GCG AAAUAGUU 1916
907 CAGAAAUU G UAUUUUUU 781 AAAAAAUA UGAUG GCAUGCACUAUGC GCG AAUUUCUG 1917
920 UUUUCUAU G CCACAUUA 782 UAAUGUGG UGAUG GCAUGCACUAUGC GCG AUAGAAAA 1918
944 UUAAAGUU G AUGAGAAU 783 AUUCUCAU UGAUG GCAUGCACUAUGC GCG AACUUUAA 1919
947 AAGUUGAU G AGAAUCAA 784 UUGAUUCU UGAUG GCAUGCACUAUGC GCG AUCAACUU 1920
1039 UAGUACAU G UAGGUAAA 785 UUUACCUA UGAUG GCAUGCACUAUGC GCG AUGUACUA 1921
1058 AUAAAUCU G UUCUAAGA 786 UCUUAGAA UGAUG GCAUGCACUAUGC GCG AGAUUUAU 1922
1072 AGACAUAU G AUCAACAG 787 CUGUUGAU UGAUG GCAUGCACUAUGC GCG AUAUGUCU 1923
1083 CAACAGAU G AGAACUGG 788 CCAGUUCU UGAUG GCAUGCACUAUGC GCG AUCUGUUG 1924
1102 GUUAAUAU G UGACAGUG 789 CACUGUCA UGAUG GCAUGCACUAUGC GCG AUAUUAAC 1925
1104 UAAUAUGU G ACAGUGAG 790 CUCACUGU UGAUG GCAUGCACUAUGC GCG ACAUAUUA 1926
Table 26
1110 GUGACAGU G AGAUUAGU 791 ACUAAUCU UGAUG GCAUGCACUAUGC GCG ACUGUCAC 1927
1168 AAGGCACU G UAGUGAAU 792 AUUCACUA UGAUG GCAUGCACUAUGC GCG AGUGCCUU 1928
1173 ACUGUAGU G AAUUAUCU 793 AGAUAAUU UGAUG GCAUGCACUAUGC GCG ACUACAGU 1929
1182 AAUUAUCU G AGCUAGAG 794 CUCUAGCU UGAUG GCAUGCACUAUGC GCG AGAUAAUU 1930
1226 GGAAUCAU G AAACCUUA 795 UAAGGUUU UGAUG GCAUGCACUAUGC GCG AUGAUUCC 1931
1247 UUCAGAAU G AUUUUGCA 796 UGCAAAAU UGAUG GCAUGCACUAUGC GCG AUUCUGAA 1932
1253 AUGAUUUU G CAGGUUGU 797 ACAACCUG UGAUG GCAUGCACUAUGC GCG AAAAUCAU 1933
1260 UGCAGGUU G UCUUCCAU 798 AUGGAAGA UGAUG GCAUGCACUAUGC GCG AACCUGCA 1934
1287 CAUCCAAU G CAGGCAAG 799 CUUGCCUG UGAUG GCAUGCACUAUGC GCG AUUGGAUG 1935
1316 UUUCCAGU G ACAGAAAA 800 UUUUCUGU UGAUG GCAUGCACUAUGC GCG ACUGGAAA 1936
1358 AAAUAUAU G AAUUCUCU 801 AGAGAAUU UGAUG GCAUGCACUAUGC GCG AUAUAUUU 1937
1401 UAUAUUUU G AAAUGAAC 802 GUUCAUUU UGAUG GCAUGCACUAUGC GCG AAAAUAUA 1938
1406 UUUGAAAU G AACUUGUU 803 AACAAGUU UGAUG GCAUGCACUAUGC GCG AUUUCAAA 1939
1412 AUGAACUU G UUGGCCCA 804 UGGGCCAA UGAUG GCAUGCACUAUGC GCG AAGUUCAU 1940
1439 CUACAGCU G ACCCUUGA 805 UCAAGGGU UGAUG GCAUGCACUAUGC GCG AGCUGUAG 1941
1446 UGACCCUU G AACAUGGG 806 CCCAUGUU UGAUG GCAUGCACUAUGC GCG AAGGGUCA 1942
1468 GGGGAGCU G ACAAUUCG 807 CGAAUUGU UGAUG GCAUGCACUAUGC GCG AGCUCCCC 1943
1484 GUGGGUCC G CAAAAUCU 808 AGAUUUUG UGAUG GCAUGCACUAUGC GCG GGACCCAC 1944
1516 CUACUAUU G ACCAUAAA 809 UUUAUGGU UGAUG GCAUGCACUAUGC GCG AAUAGUAG 1945
1532 ■ ACCUUACU G AUAACAUA 810 UAUGUUAU UGAUG GCAUGCACUAUGC GCG AGUAAGGU 1946
1564 CAUAUUUU G CGUGUUAU 811 AUAACACG UGAUG GCAUGCACUAUGC GCG AAAAUAUG 1947
1568 UUUUGCGU G UUAUAUGU 812 ACAUAUAA UGAUG GCAUGCACUAUGC GCG ACGCAAAA 1948
1575 UGUUAUAU G UAUUAUAC 813 GUAUAAUA UGAUG GCAUGCACUAUGC GCG AUAUAACA 1949
1619 GAGAAAAU G UUAUUUAG 814 CUAAAUAA UGAUG GCAUGCACUAUGC GCG AUUUUCUC 1950
Input Sequence = PLN. Cut Site = YG/M or UG/U.
Stem Length = 8. Core Sequence = UGAUG GCAUGCACUAUGC GCG
PLN (Homo sapiens phospholamban (PLN) mRNA.; 1635 bp)
Table 27
Table 27: Human Phospholamban (PLN) zinzyme Ribozyme and Target Sequence
Pos Substrate Seq ID Ribozyme Rz Seq ID
64 UCUAUACU G UGAUGAUC 732 GAUCAUCA GCCGAAAGGCGAGUCAAGGUCU AGUAUAGA 1951
79 UCACAGCU G CCAAGGCU 735 AGCCUUGG GCCGAAAGGCGAGUCAAGGUCU AGCUGUGA 1952
121 AUUUGGCU G CCAGCUUU 736 AAAGCUGG GCCGAAAGGCGAGUCAAGGUCU AGCCAAAU 1953
168 GACUUCCU G UCCUGCUG 738 CAGCAGGA GCCGAAAGGCGAGUCAAGGUCU AGGAAGUC 1954
173 CCUGUCCU G CUGGUAUC 739 GAUACCAG GCCGAAAGGCGAGUCAAGGUCU AGGACAGG 1955
207 CCUCACUC G CUCAGCUA 740 UAGCUGAG GCCGAAAGGCGAGUCAAGGUCU GAGUGAGG 1956
241 AUUGAAAU G CCUCAACA 742 UGUUGAGG GCCGAAAGGCGAGUCAAGGUCU AUUUCAAU 1957
288 CAAUUUCU G UCUCAUCU 743 AGAUGAGA GCCGAAAGGCGAGUCAAGGUCU AGAAAUUG 1958
303 CUUAAUAU G UCUCUUGC 744 GCAAGAGA GCCGAAAGGCGAGUCAAGGUCU AUAUUAAG 1959
310 UGUCUCUU G CUGAUCUG 745 CAGAUCAG GCCGAAAGGCGAGUCAAGGUCU AAGAGACA 1960
318 GCUGAUCU G UAUCAUCG 747 CGAUGAUA GCCGAAAGGCGAGUCAAGGUCU AGAUCAGC 1961
331 AUCGUGAU G CUUCUCUG 749 CAGAGAAG GCCGAAAGGCGAGUCAAGGUCU AUCACGAU 1962
347 GAAGUUCU G CUACAACC 751 GGUUGUAG GCCGAAAGGCGAGUCAAGGUCU AGAACUUC 1963
365 CUAGAUCU G CAGCUUGC 752 GCAAGCUG GCCGAAAGGCGAGUCAAGGUCU AGAUCUAG 1964
372 UGCAGCUU G CCACAUCA 753 UGAUGUGG GCCGAAAGGCGAGUCAAGGUCU AAGCUGCA 1965
392 UAAAAUCU G UCAUCCCA 754 UGGGAUGA GCCGAAAGGCGAGUCAAGGUCU AGAUUUUA 1966
402 CAUCCCAU G CAGACAGG 755 CCUGUCUG GCCGAAAGGCGAGUCAAGGUCU AUGGGAUG 1967
422 ACAAUAUU G UAUAACAG 756 CUGUUAUA GCCGAAAGGCGAGUCAAGGUCU AAUAUUGU 1968
459 GUUUCUUU G UGAAAAGG 758 CCUUUUCA GCCGAAAGGCGAGUCAAGGUCU AAAGAAAC 1969
492 AACUUAUU G UUACCAUA 760 UAUGGUAA GCCGAAAGGCGAGUCAAGGUCU AAUAAGUU 1970
502 UACCAUAU G UAUUCAUC 761 GAUGAAUA GCCGAAAGGCGAGUCAAGGUCU AUAUGGUA 1971
512 AUUCAUCU G UUGGAUCU 762 AGAUCCAA GCCGAAAGGCGAGUCAAGGUCU AGAUGAAU 1972
522 UGGAUCUU G UAAACAUG 763 CAUGUUUA GCCGAAAGGCGAGUCAAGGUCU AAGAUCCA 1973
570 AAAUAAGU G UAUAAAAU 765 AUUUUAUA GCCGAAAGGCGAGUCAAGGUCU ACUUAUUU 1974
579 UAUAAAAU G CAACUGUU 766 AACAGUUG GCCGAAAGGCGAGUCAAGGUCU AUUUUAUA 1975
585 AUGCAACU G UUGAUUUC 767 GAAAUCAA GCCGAAAGGCGAGUCAAGGUCU AGUUGCAU 1976
660 UUAAAACU G CACUGCCA 771 UGGCAGUG GCCGAAAGGCGAGUCAAGGUCU AGUUUUAA 1977
665 ACUGCACU G CCAACAAG 772 CUUGUUGG GCCGAAAGGCGAGUCAAGGUCU AGUGCAGU 1978
Table 27
736 AUUACAAU G UAAAAGCU 775 AGCUUUUA GCCGAAAGGCGAGUCAAGGUCU AUUGUAAU 1979
807 AAUGAAGU G UCAUUAUU 777 AAUAAUGA GCCGAAAGGCGAGUCAAGGUCU ACUUCAUU 1980
844 UCACAUCU G UUAUCUUA 779 UAAGAUAA GCCGAAAGGCGAGUCAAGGUCU AGAUGUGA 1981
869 AACUAUUU G UAGUAACU 780 AGUUACUA GCCGAAAGGCGAGUCAAGGUCU AAAUAGUU 1982
907 CAGAAAUU G UAUUUUUU 781 AAAAAAUA GCCGAAAGGCGAGUCAAGGUCU AAUUUCUG 1983
920 UUUUCUAU G CCACAUUA 782 UAAUGUGG GCCGAAAGGCGAGUCAAGGUCU AUAGAAAA 1984
1039 UAGUACAU G UAGGUAAA 785 UUUACCUA GCCGAAAGGCGAGUCAAGGUCU AUGUACUA 1985
1058 AUAAAUCU G UUCUAAGA 786 UCUUAGAA GCCGAAAGGCGAGUCAAGGUCU AGAUUUAU 1986
1102 GUUAAUAU G UGACAGUG 789 CACUGUCA GCCGAAAGGCGAGUCAAGGUCU AUAUUAAC 1987
1168 AAGGCACU G UAGUGAAU 792 AUUCACUA GCCGAAAGGCGAGUCAAGGUCU AGUGCCUU 1988
1253 AUGAUUUU G CAGGUUGU 797 ACAACCUG GCCGAAAGGCGAGUCAAGGUCU AAAAUCAU 1989
1260 UGCAGGUU G UCUUCCAU 798 AUGGAAGA GCCGAAAGGCGAGUCAAGGUCU AACCUGCA 1990
1287 CAUCCAAU G CAGGCAAG 799 CUUGCCUG GCCGAAAGGCGAGUCAAGGUCU AUUGGAUG 1991
1412 AUGAACUU G UUGGCCCA 804 UGGGCCAA GCCGAAAGGCGAGUCAAGGUCU AAGUUCAU 1992
1484 GUGGGUCC G CAAAAUCU 808 AGAUUUUG GCCGAAAGGCGAGUCAAGGUCU GGACCCAC 1993
1564 CAUAUUUU G CGUGUUAU 811 AUAACACG GCCGAAAGGCGAGUCAAGGUCU AAAAUAUG 1994
1568 UUUUGCGU G UUAUAUGU 812 ACAUAUAA GCCGAAAGGCGAGUCAAGGUCU ACGCAAAA 1995
1575 UGUUAUAU G UAUUAUAC 813 GUAUAAUA GCCGAAAGGCGAGUCAAGGUCU AUAUAACA 1996
1619 GAGAAAAU G UUAUUUAG 814 CUAAAUAA GCCGAAAGGCGAGUCAAGGUCU AUUUUCUC 1997
21 ACUCCCCA G CUAAACAC 815 GUGUUUAG GCCGAAAGGCGAGUCAAGGUCU UGGGGAGU 1998
32 AAACACCC G UAAGACUU 816 AAGUCUUA GCCGAAAGGCGAGUCAAGGUCU GGGUGUUU 1999
76 UGAUCACA G CUGCCAAG 817 CUUGGCAG GCCGAAAGGCGAGUCAAGGUCU UGUGAUCA 2000
85 CUGCCAAG G CUACCUAA 818 UUAGGUAG GCCGAAAGGCGAGUCAAGGUCU CUUGGCAG 2001
103 AGAAGACA G UUAUCUCA 819 UGAGAUAA GCCGAAAGGCGAGUCAAGGUCU UGUCUUCU 2002
118 CAUAUUUG G CUGCCAGC 820 GCUGGCAG GCCGAAAGGCGAGUCAAGGUCU CAAAUAUG 2003
125 GGCUGCCA G CUUUUUAU 821 AUAAAAAG GCCGAAAGGCGAGUCAAGGUCU UGGCAGCC 2004
177 UCCUGCUG G UAUCAUGG 822 CCAUGAUA GCCGAAAGGCGAGUCAAGGUCU CAGCAGGA 2005
191 UGGAGAAA G UCCAAUAC 823 GUAUUGGA GCCGAAAGGCGAGUCAAGGUCU UUUCUCCA 2006
212 CUCGCUCA G CUAUAAGA 824 UCUUAUAG GCCGAAAGGCGAGUCAAGGUCU UGAGCGAG 2007
224 UAAGAAGA G CCUCAACC 825 GGUUGAGG GCCGAAAGGCGAGUCAAGGUCU UCUUCUUA 2008
251 CUCAACAA G CACGUCAA 826 UUGACGUG GCCGAAAGGCGAGUCAAGGUCU UUGUUGAG 2009
Table 27
255 ACAAGCAC G UCAAAAGC 827 GCUUUUGA GCCGAAAGGCGAGUCAAGGUCU GUGCUUGU 2010
262 CGUCAAAA G CUACAGAA 828 UUCUGUAG GCCGAAAGGCGAGUCAAGGUCU UUUUGACG 2011
326 GUAUCAUC G UGAUGCUU 829 AAGCAUCA GCCGAAAGGCGAGUCAAGGUCU GAUGAUAC 2012
342 UCUCUGAA G UUCUGCUA 830 UAGCAGAA GCCGAAAGGCGAGUCAAGGUCU UUCAGAGA 2013
368 GAUCUGCA G CUUGCCAC 831 GUGGCAAG GCCGAAAGGCGAGUCAAGGUCU UGCAGAUC 2014
381 CCACAUCA G CUUAAAAU 832 AUUUUAAG GCCGAAAGGCGAGUCAAGGUCU UGAUGUGG 2015
443 CUUCCUGA G UAGAAGAG 833 CUCUUCUA GCCGAAAGGCGAGUCAAGGUCU UCAGGAAG 2016
451 GUAGAAGA G UUUCUUUG 834 CAAAGAAA GCCGAAAGGCGAGUCAAGGUCU UCUUCUAC 2017
467 GUGAAAAG G UCAAGAUU 835 AAUCUUGA GCCGAAAGGCGAGUCAAGGUCU CUUUUCAC 2018
537 UGAAAAGG G CUUUAUUU 836 AAAUAAAG GCCGAAAGGCGAGUCAAGGUCU CCUUUUCA 2019
568 CAAAAUAA G UGUAUAAA 837 UUUAUACA GCCGAAAGGCGAGUCAAGGUCU UUAUUUUG 2020
603 UCAACAUG G CUCACAAA 838 UUUGUGAG GCCGAAAGGCGAGUCAAGGUCU CAUGUUGA 2021
644 GAUGAAGA G UUUAGUUU 839 AAACUAAA GCCGAAAGGCGAGUCAAGGUCU UCUUCAUC 2022
649 AGAGUUUA G UUUUAAAA 840 UUUUAAAA GCCGAAAGGCGAGUCAAGGUCU UAAACUCU 2023
673 GCCAACAA G UUCACUUC 841 GAAGUGAA GCCGAAAGGCGAGUCAAGGUCU UUGUUGGC 2024
691 UAUAUAAA G CAUUAUUU 842 AAAUAAUG GCCGAAAGGCGAGUCAAGGUCU UUUAUAUA 2025
713 CUUUUGAG G UGAAUAUA 843 UAUAUUCA GCCGAAAGGCGAGUCAAGGUCU CUCAAAAG 2026
742 AUGUAAAA G CUUCUUUA 844 UAAAGAAG GCCGAAAGGCGAGUCAAGGUCU UUUUACAU 2027
758 AAUACUAA G UAUUUUUC 845 GAAAAAUA GCCGAAAGGCGAGUCAAGGUCU UUAGUAUU 2028
769 UUUUUCAG G UCUUCACC 846 GGUGAAGA GCCGAAAGGCGAGUCAAGGUCU CUGAAAAA 2029
780 UUCACCAA G UAUCAAAG 847 CUUUGAUA GCCGAAAGGCGAGUCAAGGUCU UUGGUGAA 2030
788 GUAUCAAA G UAAUAACA 848 UGUUAUUA GCCGAAAGGCGAGUCAAGGUCU UUUGAUAC 2031
805 CAAAUGAA G UGUCAUUA 849 UAAUGACA GCCGAAAGGCGAGUCAAGGUCU UUCAUUUG 2032
823 UCAAAAUA G UCCACUGA 850 UCAGUGGA GCCGAAAGGCGAGUCAAGGUCU UAUUUUGA 2033
872 UAUUUGUA G UAACUAUC 851 GAUAGUUA GCCGAAAGGCGAGUCAAGGUCU UACAAAUA 2034
941 CUUUUAAA G UUGAUGAG 852 CUCAUCAA GCCGAAAGGCGAGUCAAGGUCU UUUAAAAG 2035
956 AGAAUCAA G UAUGGAAA 853 UUUCCAUA GCCGAAAGGCGAGUCAAGGUCU UUGAUUCU 2036
966 AUGGAAAA G UAAGGCCA 854 UGGCCUUA GCCGAAAGGCGAGUCAAGGUCU UUUUCCAU 2037
971 AAAGUAAG G CCAUACUC 855 GAGUAUGG GCCGAAAGGCGAGUCAAGGUCU CUUACUUU 2038
1003 CCUUUUAA G UAAUUUUU 856 AAAAAUUA GCCGAAAGGCGAGUCAAGGUCU UUAAAAGG 2039
1033 GAAUUCUA G UACAUGUA 857 UACAUGUA GCCGAAAGGCGAGUCAAGGUCU UAGAAUUC 2040
Table 27
1043 ACAUGUAG G UAAAUCAU 858 AUGAUUUA GCCGAAAGGCGAGUCAAGGUCU CUACAUGU 2041
1091 GAGAACUG G UGGUUAAU 859 AUUAACCA GCCGAAAGGCGAGUCAAGGUCU CAGUUCUC 2042
1094 AACUGGUG G UUAAUAUG 860 CAUAUUAA GCCGAAAGGCGAGUCAAGGUCU CACCAGUU 2043
1108 AUGUGACA G UGAGAUUA 861 UAAUCUCA GCCGAAAGGCGAGUCAAGGUCU UGUCACAU 2044
1117 UGAGAUUA G UCAUAUCA 862 UGAUAUGA GCCGAAAGGCGAGUCAAGGUCU UAAUCUCA 2045
1163 CAUUUAAG G CACUGUAG 863 CUACAGUG GCCGAAAGGCGAGUCAAGGUCU CUUAAAUG 2046
1171 GCACUGUA G UGAAUUAU 864 AUAAUUCA GCCGAAAGGCGAGUCAAGGUCU UACAGUGC 2047
1184 UUAUCUGA G CUAGAGUU 865 AACUCUAG GCCGAAAGGCGAGUCAAGGUCU UCAGAUAA 2048
1190 GAGCUAGA G UUACCUAG 866 CUAGGUAA GCCGAAAGGCGAGUCAAGGUCU UCUAGCUC 2049
1198 GUUACCUA G CUUACCAU 867 AUGGUAAG GCCGAAAGGCGAGUCAAGGUCU UAGGUAAC 2050
1257 UUUUGCAG G UUGUCUUC 868 GAAGACAA GCCGAAAGGCGAGUCAAGGUCU CUGCAAAA 2051
1273 CCAUUCCA G CCUAACAU 869 AUGUUAGG GCCGAAAGGCGAGUCAAGGUCU UGGAAUGG 2052
1291 CAAUGCAG G CAAGGAAA 870 UUUCCUUG GCCGAAAGGCGAGUCAAGGUCU CUGCAUUG 2053
1314 GAUUUCCA G UGACAGAA 871 UUCUGUCA GCCGAAAGGCGAGUCAAGGUCU UGGAAAUC 2054
1339 UAUCUCAA G UAUUUUUU 872 AAAAAAUA GCCGAAAGGCGAGUCAAGGUCU UUGAGAUA 2055
1416 ACUUGUUG G CCCAUCUA 873 UAGAUGGG GCCGAAAGGCGAGUCAAGGUCU CAACAAGU 2056
1436 CAUCUACA G CUGACCCU 874 AGGGUCAG GCCGAAAGGCGAGUCAAGGUCU UGUAGAUG 2057
1456 ACAUGGGG G UUAGGGGA 875 UCCCCUAA GCCGAAAGGCGAGUCAAGGUCU CCCCAUGU 2058
1465 UUAGGGGA G CUGACAAU 876 AUUGUCAG GCCGAAAGGCGAGUCAAGGUCU UCCCCUAA 2059
1476 GACAAUUC G UGGGUCCG 877 CGGACCCA GCCGAAAGGCGAGUCAAGGUCU GAAUUGUC 2060
1480 AUUCGUGG G UCCGCAAA 878 UUUGCGGA GCCGAAAGGCGAGUCAAGGUCU CCACGAAU 2061
1506 ACCUAAUA G CCUACUAU 879 AUAGUAGG GCCGAAAGGCGAGUCAAGGUCU UAUUAGGU 2062
1545 CAUAAACA G UAAAUUAA 880 UUAAUUUA GCCGAAAGGCGAGUCAAGGUCU UGUUUAUG 2063
1566 UAUUUUGC G UGUUAUAU 881 AUAUAACA GCCGAAAGGCGAGUCAAGGUCU GCAAAAUA 2064
1603 ACAAUAAA G UAAGCUAG 882 CUAGCUUA GCCGAAAGGCGAGUCAAGGUCU UUUAUUGU 2065
1607 UAAAGUAA G CUAGAGAA 883 UUCUCUAG GCCGAAAGGCGAGUCAAGGUCU UUACUUUA 2066
Input Sequence = PLN. Cut Site = G Y
Stem Length = 8 . Core Sequence = GCcgaaagGCGaGuCaaGGuCu
PLN (Homo sapiens phospholamban (PLN) mRNA.; 1635 bp)
Table 28
Table 28: Human Phospholamban (PLN) DNAzyme and Target Sequence
Pos Substrate Seq ID Ribozyme Rz Seq ID
44 GACUUCAU A CAACACAA 6 TTGTGTTG GGCTAGCTACAACGA ATGAAGTC 2067
54 AACACAAU A CUCUAUAC 7 GTATAGAG GGCTAGCTACAACGA ATTGTGTT 2068
59 AAUACUCU A UACUGUGA 9 TCACAGTA GGCTAGCTACAACGA AGAGTATT 2069
61 UACUCUAU A CUGUGAUG 10 CATCACAG GGCTAGCTACAACGA ATAGAGTA 2070
88 CCAAGGCU A CCUAAAAG 12 CTTTTAGG GGCTAGCTACAACGA AGCCTTGG 2071
106 AGACAGUU A UCUCAUAU 15 ATATGAGA GGCTAGCTACAACGA AACTGTCT 2072
113 UAUCUCAU A UUUGGCUG 18 CAGCCAAA GGCTAGCTACAACGA ATGAGATA 2073
132 AGCUUUUU A UCUUUCUC 25 GAGAAAGA GGCTAGCTACAACGA AAAAAGCT 2074
179 CUGCUGGU A UCAUGGAG 39 CTCCATGA GGCTAGCTACAACGA ACCAGCAG 2075
198 AGUCCAAU A CCUCACUC 42 GAGTGAGG GGCTAGCTACAACGA ATTGGACT 2076
215 GCUCAGCU A UAAGAAGA 46 TCTTCTTA GGCTAGCTACAACGA AGCTGAGC 2077
265 CAAAAGCU A CAGAAUCU 52 AGATTCTG GGCTAGCTACAACGA AGCTTTTG 2078
274 CAGAAUCU A UUUAUCAA 54 TTGATAAA GGCTAGCTACAACGA AGATTCTG 2079
278 AUCUAUUU A UCAAUUUC 57 GAAATTGA GGCTAGCTACAACGA AAATAGAT 2080
301 AUCUUAAU A UGUCUCUU 67 AAGAGACA GGCTAGCTACAACGA ATTAAGAT 2081
320 UGAUCUGU A UCAUCGUG 72 CACGATGA GGCTAGCTACAACGA ACAGATCA 2082
350 GUUCUGCU A CAACCUCU 80 AGAGGTTG GGCTAGCTACAACGA AGCAGAAC 2083
419 AAAACAAU A UUGUAUAA 91 TTATACAA GGCTAGCTACAACGA ATTGTTTT 2084
424 AAUAUUGU A UAACAGAC 93 GTCTGTTA GGCTAGCTACAACGA ACAATATT 2085
489 UAAAACUU A UUGUUACC 108 GGTAACAA GGCTAGCTACAACGA AAGTTTTA 2086
495 UUAUUGUU A CCAUAUGU 111 ACATATGG GGCTAGCTACAACGA AACAATAA 2087
500 GUUACCAU A UGUAUUCA 112 TGAATACA GGCTAGCTACAACGA ATGGTAAC 2088
504 CCAUAUGU A UUCAUCUG 113 CAGATGAA GGCTAGCTACAACGA ACATATGG 2089
542 AGGGCUUU A UUUUCAAA 123 TTTGAAAA GGCTAGCTACAACGA AAAGCCCT 2090
572 AUAAGUGU A UAAAAUGC 133 GCATTTTA GGCTAGCTACAACGA ACACTTAT 2091
617 AAAUUUCU A UCCCAAAU 144 ATTTGGGA GGCTAGCTACAACGA AGAAATTT 2092
684 CACUUCAU A UAUAAAGC 162 GCTTTATA GGCTAGCTACAACGA ATGAAGTG 2093
686 CUUCAUAU A UAAAGCAU 163 ATGCTTTA GGCTAGCTACAACGA ATATGAAG 2094
696 AAAGCAUU A UUUUUACU 166 AGTAAAAA GGCTAGCTACAACGA AATGCTTT 2095
702 UUAUUUUU A CUCUUUUG 171 CAAAAGAG GGCTAGCTACAACGA AAAAATAA 2096
719 AGGUGAAU A UAAUUUAU 176 ATAAATTA GGCTAGCTACAACGA ATTCACCT 2097
726 UAUAAUUU A UAUUACAA 180 TTGTAATA GGCTAGCTACAACGA AAATTATA 2098
728 UAAUUUAU A UUACAAUG 181 CATTGTAA GGCTAGCTACAACGA ATAAATTA 2099
731 UUUAUAUU A CAAUGUAA 183 TTACATTG GGCTAGCTACAACGA AATATAAA 2100
753 UCUUUAAU A CUAAGUAU 190 ATACTTAG GGCTAGCTACAACGA ATTAAAGA 2101
760 UACUAAGU A UUUUUCAG 192 CTGAAAAA GGCTAGCTACAACGA ACTTAGTA 2102
782 CACCAAGU A UCAAAGUA 201 TACTTTGA GGCTAGCTACAACGA ACTTGGTG 2103
813 GUGUCAUU A UUCAAAAU 207 ATTTTGAA GGCTAGCTACAACGA AATGACAC 2104
847 CAUCUGUU A UCUUAUUA 216 TAATAAGA GGCTAGCTACAACGA AACAGATG 2105
852 GUUAUCUU A UUAUAAAG 219 CTTTATAA GGCTAGCTACAACGA AAGATAAC 2106
855 AUCUUAUU A UAAAGAAC 221 GTTCTTTA GGCTAGCTACAACGA AATAAGAT 2107
865 AAAGAACU A UUUGUAGU 223 ACTACAAA GGCTAGCTACAACGA AGTTCTTT 2108
878 UAGUAACU A UCAGAAUC 228 GATTCTGA GGCTAGCTACAACGA AGTTACTA 2109
888 CAGAAUCU A CAUUCUAA 231 TTAGAATG GGCTAGCTACAACGA AGATTCTG 2110
Table 28
909 GAAAUUGU A UUUUUUCU 236 AGAAAAAA GGCTAGCTACAACGA ACAATTTC 2111
918 UUUUUUCU A UGCCACAU 243 ATGTGGCA GGCTAGCTACAACGA AGAAAAAA 2112
958 AAUCAAGU A UGGAAAAG 253 CTTTTCCA GGCTAGCTACAACGA ACTTGATT 2113
976 AAGGCCAU A CUCUUACA 255 TGTAAGAG GGCTAGCTACAACGA ATGGCCTT 2114
982 AUACUCUU A CAUAAUAA 258 TTATTATG GGCTAGCTACAACGA AAGAGTAT 2115
1035 AUUCUAGU A CAUGUAGG 278 CCTACATG GGCTAGCTACAACGA ACTAGAAT 2116
1070 UAAGACAU A UGAUCAAC 287 GTTGATCA GGCTAGCTACAACGA ATGTCTTA 2117
1100 UGGUUAAU A UGUGACAG 291 CTGTCACA GGCTAGCTACAACGA ATTAACCA 2118
1122 UUAGUCAU A UCACUAAU 295 ATTAGTGA GGCTAGCTACAACGA ATGACTAA 2119
1131 UCACUAAU A UACUAACA 298 TGTTAGTA GGCTAGCTACAACGA ATTAGTGA 2120
1133 ACUAAUAU A CUAACAAC 299 GTTGTTAG GGCTAGCTACAACGA ATATTAGT 2121
1178 AGUGAAUU A UCUGAGCU 311 AGCTCAGA GGCTAGCTACAACGA AATTCACT 2122
1193 CUAGAGUU A CCUAGCUU 315 AAGCTAGG GGCTAGCTACAACGA AACTCTAG 2123
1202 CCUAGCUU A CCAUACUA 318 TAGTATGG GGCTAGCTACAACGA AAGCTAGG 2124
1207 CUUACCAU A CUAUAUCU 319 AGATATAG GGCTAGCTACAACGA ATGGTAAG 2125
1210 ACCAUACU A UAUCUUUG 320 CAAAGATA GGCTAGCTACAACGA AGTATGGT 2126
1212 CAUACUAU A UCUUUGGA 321 TCCAAAGA GGCTAGCTACAACGA ATAGTATG 2127
1327 AGAAAAAU A UAUUAUCU 345 AGATAATA GGCTAGCTACAACGA ATTTTTCT 2128
1329 AAAAAUAU A UUAUCUCA 346 TGAGATAA GGCTAGCTACAACGA ATATTTTT 2129
1332 AAUAUAUU A UCUCAAGU 348 ACTTGAGA GGCTAGCTACAACGA AATATATT 2130
1341 UCUCAAGU A UUUUUUAA 351 TTAAAAAA GGCTAGCTACAACGA ACTTGAGA 2131
1354 UUAAAAAU A UAUGAAUU 358 AATTCATA GGCTAGCTACAACGA ATTTTTAA 2132
1356 AAAAAUAU A UGAAUUCU 359 AGAATTCA GGCTAGCTACAACGA ATATTTTT 2133
1375 CUCCAAAU A UUAACUAA 365 TTAGTTAA GGCTAGCTACAACGA ATTTGGAG 2134
1386- AACUAAUU A UUAGAUUA 370 TAATCTAA GGCTAGCTACAACGA AATTAGTT 2135
1394 AUUAGAUU A UAUUUUGA 374 TCAAAATA GGCTAGCTACAACGA AATCTAAT 2136
1396 UAGAUUAU A UUUUGAAA 375 TTTCAAAA GGCTAGCTACAACGA ATAATCTA 2137
1424 GCCCAUCU A UUACAUCU 382 AGATGTAA GGCTAGCTACAACGA AGATGGGC 2138
1427 CAUCUAUU A CAUCUACA 384 TGTAGATG GGCTAGCTACAACGA AATAGATG 2139
1433 UUACAUCU A CAGCUGAC 386 GTCAGCTG GGCTAGCTACAACGA AGATGTAA 2140
1498 UCUUAACU A CCUAAUAG 396 CTATTAGG GGCTAGCTACAACGA AGTTAAGA 2141
1510 AAUAGCCU A CUAUUGAC 399 GTCAATAG GGCTAGCTACAACGA AGGCTATT 2142
1513 AGCCUACU A UUGACCAU 400 ATGGTCAA GGCTAGCTACAACGA AGTAGGCT 2143
1529 UAAACCUU A CUGAUAAC 404 GTTATCAG GGCTAGCTACAACGA AAGGTTTA 2144
1559 UAACACAU A UUUUGCGU 410 ACGCAAAA GGCTAGCTACAACGA ATGTGTTA 2145
1571 UGCGUGUU A UAUGUAUU 415 AATACATA GGCTAGCTACAACGA AACACGCA 2146
1573 CGUGUUAU A UGUAUUAU 416 ATAATACA GGCTAGCTACAACGA ATAACACG 2147
1577 UUAUAUGU A UUAUACAC 417 GTGTATAA GGCTAGCTACAACGA ACATATAA 2148
1580 UAUGUAUU A UACACUAU 419 ATAGTGTA GGCTAGCTACAACGA AATACATA 2149
1582 UGUAUUAU A CACUAUAU 420 ATATAGTG GGCTAGCTACAACGA ATAATACA 2150
1587 UAUACACU A UAUUCCUA 421 TAGGAATA GGCTAGCTACAACGA AGTGTATA 2151
1589 UACACUAU A UUCCUACA 422 TGTAGGAA GGCTAGCTACAACGA ATAGTGTA 2152
1595 AUAUUCCU A CAAUAAAG 425 CTTTATTG GGCTAGCTACAACGA AGGAATAT 2153
1622 AAAAUGUU A UUUAGAAA 430 TTTCTAAA GGCTAGCTACAACGA AACATTTT 2154
64 UCUAUACU G UGAUGAUC 732 GATCATCA GGCTAGCTACAACGA AGTATAGA 2155
79 UCACAGCU G CCAAGGCU 735 AGCCTTGG GGCTAGCTACAACGA AGCTGTGA 2156
121 AUUUGGCU G CCAGCUUU 736 AAAGCTGG GGCTAGCTACAACGA AGCCAAAT 2157
Table 28
168 GACUUCCU G UCCUGCUG 738 CAGCAGGA GGCTAGCTACAACGA AGGAAGTC 2158
173 CCUGUCCU G CUGGUAUC 739 GATACCAG GGCTAGCTACAACGA AGGACAGG 2159
207 CCUCACUC G CUCAGCUA 740 TAGCTGAG GGCTAGCTACAACGA GAGTGAGG 2160
241 AUUGAAAU G CCUCAACA 742 TGTTGAGG GGCTAGCTACAACGA ATTTCAAT 2161
288 CAAUUUCU G UCUCAUCU 743 AGATGAGA GGCTAGCTACAACGA AGAAATTG 2162
303 CUUAAUAU G UCUCUUGC 744 GCAAGAGA GGCTAGCTACAACGA ATATTAAG 2163
310 UGUCUCUU G CUGAUCUG 745 CAGATCAG GGCTAGCTACAACGA AAGAGACA 2164
318 GCUGAUCU G UAUCAUCG 747 CGATGATA GGCTAGCTACAACGA AGATCAGC 2165
331 AUCGUGAU G CUUCUCUG 749 CAGAGAAG GGCTAGCTACAACGA ATCACGAT 2166
347 GAAGUUCU G CUACAACC 751 GGTTGTAG GGCTAGCTACAACGA AGAACTTC 2167
365 CUAGAUCU G CAGCUUGC 752 GCAAGCTG GGCTAGCTACAACGA AGATCTAG 2168
372 UGCAGCUU G CCACAUCA 753 TGATGTGG GGCTAGCTACAACGA AAGCTGCA 2169
392 UAAAAUCU G UCAUCCCA 754 TGGGATGA GGCTAGCTACAACGA AGATTTTA 2170
402 CAUCCCAU G CAGACAGG 755 CCTGTCTG GGCTAGCTACAACGA ATGGGATG 2171
422 ACAAUAUU G UAUAACAG 756 CTGTTATA GGCTAGCTACAACGA AATATTGT 2172
459 GUUUCUUU G UGAAAAGG 758 CCTTTTCA GGCTAGCTACAACGA AAAGAAAC 2173
492 AACUUAUU G UUACCAUA 760 TATGGTAA GGCTAGCTACAACGA AATAAGTT 2174
502 UACCAUAU G UAUUCAUC 761 GATGAATA GGCTAGCTACAACGA ATATGGTA 2175
512 AUUCAUCU G UUGGAUCU 762 AGATCCAA GGCTAGCTACAACGA AGATGAAT 2176
522 UGGAUCUU G UAAACAUG 763 CATGTTTA GGCTAGCTACAACGA AAGATCCA 2177
570 AAAUAAGU G UAUAAAAU 765 ATTTTATA GGCTAGCTACAACGA ACTTATTT 2178
579 UAUAAAAU G CAACUGUU 766 AACAGTTG GGCTAGCTACAACGA ATTTTATA 2179
585 AUGCAACU G UUGAUUUC 767 GAAATCAA GGCTAGCTACAACGA AGTTGCAT 2180
660 UUAAAACU G CACUGCCA 771 TGGCAGTG GGCTAGCTACAACGA AGTTTTAA 2181
665 ACUGCACU G CCAACAAG 772 CTTGTTGG GGCTAGCTACAACGA AGTGCAGT 2182
736 AUUACAAU G UAAAAGCU 775 AGCTTTTA GGCTAGCTACAACGA ATTGTAAT 2183
807 AAUGAAGU G UCAUUAUU 777 AATAATGA GGCTAGCTACAACGA ACTTCATT 2184
844 UCACAUCU G UUAUCUUA 779' TAAGATAA GGCTAGCTACAACGA AGATGTGA 2185
869 AACUAUUU G UAGUAACU 780 AGTTACTA GGCTAGCTACAACGA AAATAGTT 2186
907 CAGAAAUU G UAUUUUUU 781 AAAAAATA GGCTAGCTACAACGA AATTTCTG 2187
920 UUUUCUAU G CCACAUUA 782 TAATGTGG GGCTAGCTACAACGA ATAGAAAA 2188
1039 UAGUACAU G UAGGUAAA 785 TTTACCTA GGCTAGCTACAACGA ATGTACTA 2189
1058 AUAAAUCU G UUCUAAGA 786 TCTTAGAA GGCTAGCTACAACGA AGATTTAT 2190
1102 GUUAAUAU G UGACAGUG 789 CACTGTCA GGCTAGCTACAACGA ATATTAAC 2191
1168 AAGGCACU G UAGUGAAU 792 ATTCACTA GGCTAGCTACAACGA AGTGCCTT 2192
1253 AUGAUUUU G CAGGUUGU 797 ACAACCTG GGCTAGCTACAACGA AAAATCAT 2193
1260 UGCAGGUU G UCUUCCAU 798 ATGGAAGA GGCTAGCTACAACGA AACCTGCA 2194
1287 CAUCCAAU G CAGGCAAG 799 CTTGCCTG GGCTAGCTACAACGA ATTGGATG 2195
1412 AUGAACUU G UUGGCCCA 804 TGGGCCAA GGCTAGCTACAACGA AAGTTCAT 2196
1484 GUGGGUCC G CAAAAUCU 808 AGATTTTG GGCTAGCTACAACGA GGACCCAC 2197
1564 CAUAUUUU G CGUGUUAU 811 ATAACACG GGCTAGCTACAACGA AAAATATG 2198
1568 UUUUGCGU G UUAUAUGU 812 ACATATAA GGCTAGCTACAACGA ACGCAAAA 2199
1575 UGUUAUAU G UAUUAUAC 813 GTATAATA GGCTAGCTACAACGA ATATAACA 2200
1619 GAGAAAAU G UUAUUUAG 814 CTAAATAA GGCTAGCTACAACGA ATTTTCTC 2201
21 ACUCCCCA G CUAAACAC 815 GTGTTTAG GGCTAGCTACAACGA TGGGGAGT 2202
32 AAACACCC G UAAGACUU 816 AAGTCTTA GGCTAGCTACAACGA GGGTGTTT 2203
76 UGAUCACA G CUGCCAAG 817 CTTGGCAG GGCTAGCTACAACGA TGTGATCA 2204
Table 28
85 CUGCCAAG G CUACCUAA 818 TTAGGTAG GGCTAGCTACAACGA CTTGGCAG 2205
103 AGAAGACA G UUAUCUCA 819 TGAGATAA GGCTAGCTACAACGA TGTCTTCT 2206
118 CAUAUUUG G CUGCCAGC 820 GCTGGCAG GGCTAGCTACAACGA CAAATATG 2207
125 GGCUGCCA G CUUUUUAU 821 ATAAAAAG GGCTAGCTACAACGA TGGCAGCC 2208
177 UCCUGCUG G UAUCAUGG 822 CCATGATA GGCTAGCTACAACGA CAGCAGGA 2209
191 UGGAGAAA G UCCAAUAC 823 GTATTGGA GGCTAGCTACAACGA TTTCTCCA 2210
212 CUCGCUCA G CUAUAAGA 824 TCTTATAG GGCTAGCTACAACGA TGAGCGAG 2211
224 UAAGAAGA G CCUCAACC 825 GGTTGAGG GGCTAGCTACAACGA TCTTCTTA 2212
251 CUCAACAA G CACGUCAA 826 TTGACGTG GGCTAGCTACAACGA TTGTTGAG 2213
255 ACAAGCAC G UCAAAAGC 827 GCTTTTGA GGCTAGCTACAACGA GTGCTTGT 2214
262 CGUCAAAA G CUACAGAA 828 TTCTGTAG GGCTAGCTACAACGA TTTTGACG 2215
326 GUAUCAUC G UGAUGCUU 829 AAGCATCA GGCTAGCTACAACGA GATGATAC 2216
342 UCUCUGAA G UUCUGCUA 830 TAGCAGAA GGCTAGCTACAACGA TTCAGAGA 2217
368 GAUCUGCA G CUUGCCAC 831 GTGGCAAG GGCTAGCTACAACGA TGCAGATC 2218
381 CCACAUCA G CUUAAAAU 832 ATTTTAAG GGCTAGCTACAACGA TGATGTGG 2219
443 CUUCCUGA G UAGAAGAG 833 CTCTTCTA GGCTAGCTACAACGA TCAGGAAG 2220
451 GUAGAAGA G UUUCUUUG 834 CAAAGAAA GGCTAGCTACAACGA TCTTCTAC 2221
467 GUGAAAAG G UCAAGAUU 835 AATCTTGA GGCTAGCTACAACGA CTTTTCAC 2222
537 UGAAAAGG G CUUUAUUU 836 AAATAAAG GGCTAGCTACAACGA CCTTTTCA 2223
568 CAAAAUAA G UGUAUAAA 837 TTTATACA GGCTAGCTACAACGA TTATTTTG 2224
603 UCAACAUG G CUCACAAA 838 TTTGTGAG GGCTAGCTACAACGA CATGTTGA 2225
644 GAUGAAGA G UUUAGUUU 839 AAACTAAA GGCTAGCTACAACGA TCTTCATC 2226
649 AGAGUUUA G UUUUAAAA 840 TTTTAAAA GGCTAGCTACAACGA TAAACTCT 2227
673 GCCAACAA G UUCACUUC 841 GAAGTGAA GGCTAGCTACAACGA TTGTTGGC 2228
691 UAUAUAAA G CAUUAUUU 842 AAATAATG GGCTAGCTACAACGA TTTATATA 2229
713 CUUUUGAG G UGAAUAUA 843 TATATTCA GGCTAGCTACAACGA CTCAAAAG 2230
742 AUGUAAAA G CUUCUUUA 844 TAAAGAAG GGCTAGCTACAACGA TTTTACAT 2231
758 AAUACUAA G UAUUUUUC 845 GAAAAATA GGCTAGCTACAACGA TTAGTATT 2232
769 UUUUUCAG G UCUUCACC 846 GGTGAAGA GGCTAGCTACAACGA CTGAAAAA 2233
780 UUCACCAA G UAUCAAAG 847 CTTTGATA GGCTAGCTACAACGA TTGGTGAA 2234
788 GUAUCAAA G UAAUAACA 848 TGTTATTA GGCTAGCTACAACGA TTTGATAC 2235
805 CAAAUGAA G UGUCAUUA 849 TAATGACA GGCTAGCTACAACGA TTCATTTG 2236
823 UCAAAAUA G UCCACUGA 850 TCAGTGGA GGCTAGCTACAACGA TATTTTGA 2237
872 UAUUUGUA G UAACUAUC 851 GATAGTTA GGCTAGCTACAACGA TACAAATA 2238
941 CUUUUAAA G UUGAUGAG 852 CTCATCAA GGCTAGCTACAACGA TTTAAAAG 2239
956 AGAAUCAA G UAUGGAAA 853 TTTCCATA GGCTAGCTACAACGA TTGATTCT 2240
966 AUGGAAAA G UAAGGCCA 854 TGGCCTTA GGCTAGCTACAACGA TTTTCCAT 2241
971 AAAGUAAG G CCAUACUC 855 GAGTATGG GGCTAGCTACAACGA CTTACTTT 2242
1003 CCUUUUAA G UAAUUUUU 856 AAAAATTA GGCTAGCTACAACGA TTAAAAGG 2243
1033 GAAUUCUA G UACAUGUA 857 TACATGTA GGCTAGCTACAACGA TAGAATTC 2244
1043 ACAUGUAG G UAAAUCAU 858 ATGATTTA GGCTAGCTACAACGA CTACATGT 2245
1091 GAGAACUG G UGGUUAAU 859 ATTAACCA GGCTAGCTACAACGA CAGTTCTC 2246
1094 AACUGGUG G UUAAUAUG 860 CATATTAA GGCTAGCTACAACGA CACCAGTT 2247
1108 AUGUGACA G UGAGAUUA 861 TAATCTCA GGCTAGCTACAACGA TGTCACAT 2248
1117 UGAGAUUA G UCAUAUCA 862 TGATATGA GGCTAGCTACAACGA TAATCTCA 2249
1163 CAUUUAAG G CACUGUAG 863 CTACAGTG GGCTAGCTACAACGA CTTAAATG 2250
1171 GCACUGUA G UGAAUUAU 864 ATAATTCA GGCTAGCTACAACGA TACAGTGC 2251
Table 28
1184 UUAUCUGA G CUAGAGUU 865 AACTCTAG GGCTAGCTACAACGA TCAGATAA 2252
1190 GAGCUAGA G UUACCUAG 866 CTAGGTAA GGCTAGCTACAACGA TCTAGCTC 2253
1198 GUUACCUA G CUUACCAU 867 ATGGTAAG GGCTAGCTACAACGA TAGGTAAC 2254
1257 UUUUGCAG G UUGUCUUC 868 GAAGACAA GGCTAGCTACAACGA CTGCAAAA 2255
1273 CCAUUCCA G CCUAACAU 869 ATGTTAGG GGCTAGCTACAACGA TGGAATGG 2256
1291 CAAUGCAG G CAAGGAAA 870 TTTCCTTG GGCTAGCTACAACGA CTGCATTG 2257
1314 GAUUUCCA G UGACAGAA 871 TTCTGTCA GGCTAGCTACAACGA TGGAAATC 2258
1339 UAUCUCAA G UAUUUUUU 872 AAAAAATA GGCTAGCTACAACGA TTGAGATA 2259
1416 ACUUGUUG G CCCAUCUA 873 TAGATGGG GGCTAGCTACAACGA CAACAAGT 2260
1436 CAUCUACA G CUGACCCU 874 AGGGTCAG GGCTAGCTACAACGA TGTAGATG 2261
1456 ACAUGGGG G UUAGGGGA 875 TCCCCTAA GGCTAGCTACAACGA CCCCATGT 2262
1465 UUAGGGGA G CUGACAAU 876 ATTGTCAG GGCTAGCTACAACGA TCCCCTAA 2263
1476 GACAAUUC G UGGGUCCG 877 CGGACCCA GGCTAGCTACAACGA GAATTGTC 2264
1480 AUUCGUGG G UCCGCAAA 878 TTTGCGGA GGCTAGCTACAACGA CCACGAAT 2265
1506 ACCUAAUA G CCUACUAU 879 ATAGTAGG GGCTAGCTACAACGA TATTAGGT 2266
1545 CAUAAACA G UAAAUUAA 880 TTAATTTA GGCTAGCTACAACGA TGTTTATG 2267
1566 UAUUUUGC G UGUUAUAU 881 ATATAACA GGCTAGCTACAACGA GCAAAATA 2268
1603 ACAAUAAA G UAAGCUAG 882 CTAGCTTA GGCTAGCTACAACGA TTTATTGT 2269
1607 UAAAGUAA G CUAGAGAA 883 TTCTCTAG GGCTAGCTACAACGA TTACTTTA 2270
13 GUCAGAAA A CUCCCCAG 884 CTGGGGAG GGCTAGCTACAACGA TTTCTGAC 2271
26 CCAGCUAA A CACCCGUA 885 TACGGGTG GGCTAGCTACAACGA TTAGCTGG 2272
28 AGCUAAAC A CCCGUAAG 886 CTTACGGG GGCTAGCTACAACGA GTTTAGCT 2273
37 CCCGUAAG A CUUCAUAC 887 GTATGAAG GGCTAGCTACAACGA CTTACGGG 2274
42 AAGACUUC A UACAACAC 888 GTGTTGTA GGCTAGCTACAACGA GAAGTCTT 2275
47 UUCAUACA A CACAAUAC 889 GTATTGTG GGCTAGCTACAACGA TGTATGAA 2276
49 CAUACAAC A CAAUACUC 890 GAGTATTG GGCTAGCTACAACGA GTTGTATG 2277
52 ACAACACA A UACUCUAU 891 ATAGAGTA GGCTAGCTACAACGA TGTGTTGT 2278
67 AUACUGUG A UGAUCACA 892 TGTGATCA GGCTAGCTACAACGA CACAGTAT 2279
70 CUGUGAUG A UCACAGCU 893 AGCTGTGA GGCTAGCTACAACGA CATCACAG 2280
73 UGAUGAUC A CAGCUGCC 894 GGCAGCTG GGCTAGCTACAACGA GATCATCA 2281
100 AAAAGAAG A CAGUUAUC 895 GATAACTG GGCTAGCTACAACGA CTTCTTTT 2282
111 GUUAUCUC A UAUUUGGC 896 GCCAAATA GGCTAGCTACAACGA GAGATAAC 2283
144 UUCUCUCG A CCACUUAA 897 TTAAGTGG GGCTAGCTACAACGA CGAGAGAA 2284
147 UCUCGACC A CUUAAAAC 898 GTTTTAAG GGCTAGCTACAACGA GGTCGAGA 2285
154 CACUUAAA A CUUCAGAC 899 GTCTGAAG GGCTAGCTACAACGA TTTAAGTG 2286
161 AACUUCAG A CUUCCUGU 900 ACAGGAAG GGCTAGCTACAACGA CTGAAGTT 2287
182 CUGGUAUC A UGGAGAAA 901 TTTCTCCA GGCTAGCTACAACGA GATACCAG 2288
196 AAAGUCCA A UACCUCAC 902 GTGAGGTA GGCTAGCTACAACGA TGGACTTT 2289
203 AAUACCUC A CUCGCUCA 903 TGAGCGAG GGCTAGCTACAACGA GAGGTATT 2290
230 GAGCCUCA A CCAUUGAA 904 TTCAATGG GGCTAGCTACAACGA TGAGGCTC 2291
233 CCUCAACC A UUGAAAUG 905 CATTTCAA GGCTAGCTACAACGA GGTTGAGG 2292
239 CCAUUGAA A UGCCUCAA 906 TTGAGGCA GGCTAGCTACAACGA TTCAATGG 2293
247 AUGCCUCA A CAAGCACG 907 CGTGCTTG GGCTAGCTACAACGA TGAGGCAT 2294
253 CAACAAGC A CGUCAAAA 908 TTTTGACG GGCTAGCTACAACGA GCTTGTTG 2295
270 GCUACAGA A UCUAUUUA 909 TAAATAGA GGCTAGCTACAACGA TCTGTAGC 2296
282 AUUUAUCA A UUUCUGUC 910 GACAGAAA GGCTAGCTACAACGA TGATAAAT 2297
293 UCUGUCUC A UCUUAAUA 911 TATTAAGA GGCTAGCTACAACGA GAGACAGA 2298
Table 28
299 UCAUCUUA A UAUGUCUC 912 GAGACATA GGCTAGCTACAACGA TAAGATGA 2299
314 UCUUGCUG A UCUGUAUC 913 GATACAGA GGCTAGCTACAACGA CAGCAAGA 2300
323 UCUGUAUC A UCGUGAUG 914 CATCACGA GGCTAGCTACAACGA GATACAGA 2301
329 UCAUCGUG A UGCUUCUC 915 GAGAAGCA GGCTAGCTACAACGA CACGATGA 2302
353 CUGCUACA A CCUCUAGA 916 TCTAGAGG GGCTAGCTACAACGA TGTAGCAG 2303
361 ACCUCUAG A UCUGCAGC 917 GCTGCAGA GGCTAGCTACAACGA CTAGAGGT 2304
375 AGCUUGCC A CAUCAGCU 918 AGCTGATG GGCTAGCTACAACGA GGCAAGCT 2305
377 CUUGCCAC A UCAGCUUA 919 TAAGCTGA GGCTAGCTACAACGA GTGGCAAG 2306
388 AGCUUAAA A UCUGUCAU 920 ATGACAGA GGCTAGCTACAACGA TTTAAGCT 2307
395 AAUCUGUC A UCCCAUGC 921 GCATGGGA GGCTAGCTACAACGA GACAGATT 2308
400 GUCAUCCC A UGCAGACA 922 TGTCTGCA GGCTAGCTACAACGA GGGATGAC 2309
406 CCAUGCAG A CAGGAAAA 923 TTTTCCTG GGCTAGCTACAACGA CTGCATGG 2310
414 ACAGGAAA A CAAUAUUG 924 CAATATTG GGCTAGCTACAACGA TTTCCTGT 2311
417 GGAAAACA A UAUUGUAU 925 ATACAATA GGCTAGCTACAACGA TGTTTTCC 2312
427 AUUGUAUA A CAGACCAC 926 GTGGTCTG GGCTAGCTACAACGA TATACAAT 2313
431 UAUAACAG A CCACUUCC 927 GGAAGTGG GGCTAGCTACAACGA CTGTTATA 2314
434 AACAGACC A CUUCCUGA 928 TCAGGAAG GGCTAGCTACAACGA GGTCTGTT 2315
473 AGGUCAAG A UUAAGACU 929 AGTCTTAA GGCTAGCTACAACGA CTTGACCT 2316
479 AGAUUAAG A CUAAAACU 930 AGTTTTAG GGCTAGCTACAACGA CTTAATCT 2317
485 AGACUAAA A CUUAUUGU 931 ACAATAAG GGCTAGCTACAACGA TTTAGTCT 2318
498 UUGUUACC A UAUGUAUU 932 AATACATA GGCTAGCTACAACGA GGTAACAA 2319
508 AUGUAUUC A UCUGUUGG 933 CCAACAGA GGCTAGCTACAACGA GAATACAT 2320
517 UCUGUUGG A UCUUGUAA 934 TTACAAGA GGCTAGCTACAACGA CCAACAGA 2321
526 UCUUGUAA A CAUGAAAA 935 TTTTCATG GGCTAGCTACAACGA TTACAAGA 2322
528 UUGUAAAC A UGAAAAGG 936 CCTTTTCA GGCTAGCTACAACGA GTTTACAA 2323
552 UUUCAAAA A UUAACUUC 937 GAAGTTAA GGCTAGCTACAACGA TTTTGAAA 2324
556 AAAAAUUA A CUUCAAAA 938 TTTTGAAG GGCTAGCTACAACGA TAATTTTT 2325
564 ACUUCAAA A UAAGUGUA 939 TACACTTA GGCTAGCTACAACGA TTTGAAGT 2326
577 UGUAUAAA A UGCAACUG 940 CAGTTGCA GGCTAGCTACAACGA TTTATACA 2327
582 AAAAUGCA A CUGUUGAU 941 ATCAACAG GGCTAGCTACAACGA TGCATTTT 2328
589 AACUGUUG A UUUCCUCA 942 TGAGGAAA GGCTAGCTACAACGA CAACAGTT 2329
598 UUUCCUCA A CAUGGCUC 943 GAGCCATG GGCTAGCTACAACGA TGAGGAAA 2330
600 UCCUCAAC A UGGCUCAC 944 GTGAGCCA GGCTAGCTACAACGA GTTGAGGA 2331
607 CAUGGCUC A CAAAUUUC 945 GAAATTTG GGCTAGCTACAACGA GAGCCATG 2332
611 GCUCACAA A UUUCUAUC 946 GATAGAAA GGCTAGCTACAACGA TTGTGAGC 2333
624 UAUCCCAA A UCUUUUCU 947 AGAAAAGA GGCTAGCTACAACGA TTGGGATA 2334
637 UUCUGAAG A UGAAGAGU 948 ACTCTTCA GGCTAGCTACAACGA CTTCAGAA 2335
657 GUUUUAAA A CUGCACUG 949 CAGTGCAG GGCTAGCTACAACGA TTTAAAAC 2336
662 AAAACUGC A CUGCCAAC 950 GTTGGCAG GGCTAGCTACAACGA GCAGTTTT 2337
669 CACUGCCA A CAAGUUCA 951 TGAACTTG GGCTAGCTACAACGA TGGCAGTG 2338
677 ACAAGUUC A CUUCAUAU 952 ATATGAAG GGCTAGCTACAACGA GAACTTGT 2339
682 UUCACUUC A UAUAUAAA 953 TTTATATA GGCTAGCTACAACGA GAAGTGAA 2340
693 UAUAAAGC A UUAUUUUU 954 AAAAATAA GGCTAGCTACAACGA GCTTTATA 2341
717 UGAGGUGA A UAUAAUUU 955 AAATTATA GGCTAGCTACAACGA TCACCTCA 2342
722 UGAAUAUA A UUUAUAUU 956 AATATAAA GGCTAGCTACAACGA TATATTCA 2343
734 AUAUUACA A UGUAAAAG 957 CTTTTACA GGCTAGCTACAACGA TGTAATAT 2344
751 CUUCUUUA A UACUAAGU 958 ACTTAGTA GGCTAGCTACAACGA TAAAGAAG 2345
Table 28
775 AGGUCUUC A CCAAGUAU 959 ATACTTGG GGCTAGCTACAACGA GAAGACCT 2346
791 UCAAAGUA A UAACACAA 960 TTGTGTTA GGCTAGCTACAACGA TACTTTGA 2347
794 AAGUAAUA A CACAAAUG 961 CATTTGTG GGCTAGCTACAACGA TATTACTT 2348
796 GUAAUAAC A CAAAUGAA 962 TTCATTTG GGCTAGCTACAACGA GTTATTAC 2349
800 UAACACAA A UGAAGUGU 963 ACACTTCA GGCTAGCTACAACGA TTGTGTTA 2350
810 GAAGUGUC A UUAUUCAA 964 TTGAATAA GGCTAGCTACAACGA GACACTTC 2351
820 UAUUCAAA A UAGUCCAC 965 GTGGACTA GGCTAGCTACAACGA TTTGAATA 2352
827 AAUAGUCC A CUGACUCC 966 GGAGTCAG GGCTAGCTACAACGA GGACTATT 2353
831 GUCCACUG A CUCCUCAC 967 GTGAGGAG GGCTAGCTACAACGA CAGTGGAC 2354
838 GACUCCUC A CAUCUGUU 968 AACAGATG GGCTAGCTACAACGA GAGGAGTC 2355
840 CUCCUCAC A UCUGUUAU 969 ATAACAGA GGCTAGCTACAACGA GTGAGGAG 2356
862 UAUAAAGA A CUAUUUGU 970 ACAAATAG GGCTAGCTACAACGA TCTTTATA 2357
875 UUGUAGUA A CUAUCAGA 971 TCTGATAG GGCTAGCTACAACGA TACTACAA 2358
884 CUAUCAGA A UCUACAUU 972 AATGTAGA GGCTAGCTACAACGA TCTGATAG 2359
890 GAAUCUAC A UUCUAAAA 973 TTTTAGAA GGCTAGCTACAACGA GTAGATTC 2360
898 AUUCUAAA A CAGAAAUU 974 AATTTCTG GGCTAGCTACAACGA TTTAGAAT 2361
904 AAACAGAA A UUGUAUUU 975 AAATACAA GGCTAGCTACAACGA TTCTGTTT 2362
923 UCUAUGCC A CAUUAACA 976 TGTTAATG GGCTAGCTACAACGA GGCATAGA 2363
925 UAUGCCAC A UUAACAUC 977 GATGTTAA GGCTAGCTACAACGA GTGGCATA 2364
929 CCACAUUA A CAUCUUUU 978 AAAAGATG GGCTAGCTACAACGA TAATGTGG 2365
931 ACAUUAAC A UCUUUUAA 979 TTAAAAGA GGCTAGCTACAACGA GTTAATGT 2366
945 UAAAGUUG A UGAGAAUC 980 GATTCTCA GGCTAGCTACAACGA CAACTTTA 2367
951 UGAUGAGA A UCAAGUAU 981 ATACTTGA GGCTAGCTACAACGA TCTCATCA 2368
974 GUAAGGCC A UACUCUUA 982 TAAGAGTA GGCTAGCTACAACGA GGCCTTAC 2369
984 ACUCUUAC A UAAUAAAA 983 TTTTATTA GGCTAGCTACAACGA GTAAGAGT 2370
987 CUUACAUA A UAAAAUUC 984 GAATTTTA GGCTAGCTACAACGA TATGTAAG 2371
992 AUAAUAAA A UUCCUUUU 985 AAAAGGAA GGCTAGCTACAACGA TTTATTAT 2372
1006 UUUAAGUA A UUUUUUCA 986 TGAAAAAA GGCTAGCTACAACGA TACTTAAA 2373
1019 UUCAAAGA A UCACAGAA 987 TTCTGTGA GGCTAGCTACAACGA TCTTTGAA 2374
1022 AAAGAAUC A CAGAAUUC 988 GAATTCTG GGCTAGCTACAACGA GATTCTTT 2375
1027 AUCACAGA A UUCUAGUA 989 TACTAGAA GGCTAGCTACAACGA TCTGTGAT 2376
1037 UCUAGUAC A UGUAGGUA 990 TACCTACA GGCTAGCTACAACGA GTACTAGA 2377
1047 GUAGGUAA A UCAUAAAU 991 ATTTATGA GGCTAGCTACAACGA TTACCTAC 2378
1050 GGUAAAUC A UAAAUCUG 992 CAGATTTA GGCTAGCTACAACGA GATTTACC 2379
1054 AAUCAUAA A UCUGUUCU 993 AGAACAGA GGCTAGCTACAACGA TTATGATT 2380
1066 GUUCUAAG A CAUAUGAU 994 ATCATATG GGCTAGCTACAACGA CTTAGAAC 2381
1068 UCUAAGAC A UAUGAUCA 995 TGATCATA GGCTAGCTACAACGA GTCTTAGA 2382
1073 GACAUAUG A UCAACAGA 996 TCTGTTGA GGCTAGCTACAACGA CATATGTC 2383
1077 UAUGAUCA A CAGAUGAG 997 CTCATCTG GGCTAGCTACAACGA TGATCATA 2384
1081 AUCAACAG A UGAGAACU 998 AGTTCTCA GGCTAGCTACAACGA CTGTTGAT 2385
1087 AGAUGAGA A CUGGUGGU 999 ACCACCAG GGCTAGCTACAACGA TCTCATCT 2386
1098 GGUGGUUA A UAUGUGAC 1000 GTCACATA GGCTAGCTACAACGA TAACCACC 2387
1105 AAUAUGUG A CAGUGAGA 1001 TCTCACTG GGCTAGCTACAACGA CACATATT 2388
1113 ACAGUGAG A UUAGUCAU 1002 ATGACTAA GGCTAGCTACAACGA CTCACTGT 2389
1120 GAUUAGUC A UAUCACUA 1003 TAGTGATA GGCTAGCTACAACGA GACTAATC 2390
1125 GUCAUAUC A CUAAUAUA 1004 TATATTAG GGCTAGCTACAACGA GATATGAC 2391
1129 UAUCACUA A UAUACUAA 1005 TTAGTATA GGCTAGCTACAACGA TAGTGATA 2392
Table 28
1137 AUAUACUA A CAACAGAA 1006 TTCTGTTG GGCTAGCTACAACGA TAGTATAT 2393
1140 UACUAACA A CAGAAUCU 1007 AGATTCTG GGCTAGCTACAACGA TGTTAGTA 2394
1145 ACAACAGA A UCUAAUCU 1008 AGATTAGA GGCTAGCTACAACGA TCTGTTGT 2395
1150 AGAAUCUA A UCUUCAUU 1009 AATGAAGA GGCTAGCTACAACGA TAGATTCT 2396
1156 UAAUCUUC A UUUAAGGC 1010 GCCTTAAA GGCTAGCTACAACGA GAAGATTA 2397
1165 UUUAAGGC A CUGUAGUG 1011 CACTACAG GGCTAGCTACAACGA GCCTTAAA 2398
1175 UGUAGUGA A UUAUCUGA 1012 TCAGATAA GGCTAGCTACAACGA TCACTACA 2399
1205 . AGCUUACC A UACUAUAU 1013 ATATAGTA GGCTAGCTACAACGA GGTAAGCT 2400
1221 UCUUUGGA A UCAUGAAA 1014 TTTCATGA GGCTAGCTACAACGA TCCAAAGA 2401
1224 UUGGAAUC A UGAAACCU 1015 AGGTTTCA GGCTAGCTACAACGA GATTCCAA 2402
1229 AUCAUGAA A CCUUAAGA 1016 TCTTAAGG GGCTAGCTACAACGA TTCATGAT 2403
1237 ACCUUAAG A CUUCAGAA 1017 TTCTGAAG GGCTAGCTACAACGA CTTAAGGT 2404
1245 ACUUCAGA A UGAUUUUG 1018 CAAAATCA GGCTAGCTACAACGA TCTGAAGT 2405
1248 UCAGAAUG A UUUUGCAG 1019 CTGCAAAA GGCTAGCTACAACGA CATTCTGA 2406
1267 UGUCUUCC A UUCCAGCC 1020 GGCTGGAA GGCTAGCTACAACGA GGAAGACA 2407
1278 CCAGCCUA A CAUCCAAU 1021 ATTGGATG GGCTAGCTACAACGA TAGGCTGG 2408
1280 AGCCUAAC A UCCAAUGC 1022 GCATTGGA GGCTAGCTACAACGA GTTAGGCT 2409
1285 AACAUCCA A UGCAGGCA 1023 TGCCTGCA GGCTAGCTACAACGA TGGATGTT 2410
1300 CAAGGAAA A UAAAAGAU 1024 ATCTTTTA GGCTAGCTACAACGA TTTCCTTG 2411
1307 AAUAAAAG A UUUCCAGU 1025 ACTGGAAA GGCTAGCTACAACGA CTTTTATT 2412
1317 UUCCAGUG A CAGAAAAA 1026 TTTTTCTG GGCTAGCTACAACGA CACTGGAA 2413
1325 ACAGAAAA A UAUAUUAU 1027 ATAATATA GGCTAGCTACAACGA TTTTCTGT 2414
1352 UUUUAAAA A UAUAUGAA 1028 TTCATATA GGCTAGCTACAACGA TTTTAAAA 2415
1360 AUAUAUGA A UUCUCUCU 1029 AGAGAGAA GGCTAGCTACAACGA TCATATAT 2416
1373 CUCUCCAA A UAUUAACU 1030 AGTTAATA GGCTAGCTACAACGA TTGGAGAG 2417
1379 AAAUAUUA A CUAAUUAU 1031 ATAATTAG GGCTAGCTACAACGA TAATATTT 2418
1383 AUUAACUA A UUAUUAGA 1032 TCTAATAA GGCTAGCTACAACGA TAGTTAAT 2419
1391 AUUAUUAG A UUAUAUUU 1033 AAATATAA GGCTAGCTACAACGA CTAATAAT 2420
1404 AUUUUGAA A UGAACUUG 1034 CAAGTTCA GGCTAGCTACAACGA TTCAAAAT 2421
1408 UGAAAUGA A CUUGUUGG 1035 CCAACAAG GGCTAGCTACAACGA TCATTTCA 2422
1420 GUUGGCCC A UCUAUUAC 1036 GTAATAGA GGCTAGCTACAACGA GGGCCAAC 2423
1429 UCUAUUAC A UCUACAGC 1037 GCTGTAGA GGCTAGCTACAACGA GTAATAGA 2424
1440 UACAGCUG A CCCUUGAA 1038 TTCAAGGG GGCTAGCTACAACGA CAGCTGTA 2425
1448 ACCCUUGA A CAUGGGGG 1039 CCCCCATG GGCTAGCTACAACGA TCAAGGGT 2426
1450 CCUUGAAC A UGGGGGUU 1040 AACCCCCA GGCTAGCTACAACGA GTTCAAGG 2427
1469 GGGAGCUG A CAAUUCGU 1041 ACGAATTG GGCTAGCTACAACGA CAGCTCCC 2428
1472 AGCUGACA A UUCGUGGG 1042 CCCACGAA GGCTAGCTACAACGA TGTCAGCT 2429
1489 UCCGCAAA A UCUUAACU 1043 AGTTAAGA GGCTAGCTACAACGA TTTGCGGA 2430
1495 AAAUCUUA A CUACCUAA 1044 TTAGGTAG GGCTAGCTACAACGA TAAGATTT 2431
1503 ACUACCUA A UAGCCUAC 1045 GTAGGCTA GGCTAGCTACAACGA TAGGTAGT 2432
1517 UACUAUUG A CCAUAAAC 1046 GTTTATGG GGCTAGCTACAACGA CAATAGTA 2433
1520 UAUUGACC A UAAACCUU 1047 AAGGTTTA GGCTAGCTACAACGA GGTCAATA 2434
1524 GACCAUAA A CCUUACUG 1048 CAGTAAGG GGCTAGCTACAACGA TTATGGTC 2435
1533 CCUUACUG A UAACAUAA 1049 TTATGTTA GGCTAGCTACAACGA CAGTAAGG 2436
1536 UACUGAUA A CAUAAACA 1050 TGTTTATG GGCTAGCTACAACGA TATCAGTA 2437
1538 CUGAUAAC A UAAACAGU 1051 ACTGTTTA GGCTAGCTACAACGA GTTATCAG 2438
1542 UAACAUAA A CAGUAAAU 1052 ATTTACTG GGCTAGCTACAACGA TTATGTTA 2439
Table 28
Input Sequence = PLN. Cut Site = R/Y
Stem Length = 8 . Core Sequence = GGCTAGCTACAACGA
PLN (Homo sapiens phospholamban (PLN) mRNA.; 1635 bp)
Table 29
Table 29: Human Phospholamban (PLN) amberzyme Ribozyme and Target Sequence
Pos Substrate Seq ID Ribozyme Rz Seq ID
64 UCUAUACU G UGAUGAUC 732 GAUCAUCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGUAUAGA 2447
66 UAUACUGU G AUGAUCAC 733 GUGAUCAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACAGUAUA 2448
69 ACUGUGAU G AUCACAGC 734 GCUGUGAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUCACAGU 2449
79 UCACAGCU G CCAAGGCU 735 AGCCUUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCUGUGA 2450
121 AUUUGGCU G CCAGCUUU 736 AAAGCUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCCAAAU 2451
143 UUUCUCUC G ACCACUUA 737 UAAGUGGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAGAGAAA 2452
168 GACUUCCU G UCCUGCUG 738 CAGCAGGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGAAGUC 2453
173 CCUGUCCU G CUGGUAUC 739 GAUACCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGACAGG 2454
207 CCUCACUC G CUCAGCUA 740 UAGCUGAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAGUGAGG 2455
236 CAACCAUU G AAAUGCCU 741 AGGCAUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAUGGUUG 2456
241 AUUGAAAU G CCUCAACA 742 UGUUGAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUUUCAAU 2457
288 CAAUUUCU G UCUCAUCU 743 AGAUGAGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGAAAUUG 2458
303 CUUAAUAU G UCUCUUGC 744 GCAAGAGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUAUUAAG 2459
310 UGUCUCUU G CUGAUCUG 745 CAGAUCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGAGACA 2460
313 CUCUUGCU G AUCUGUAU 746 AUACAGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCAAGAG 2461
318 GCUGAUCU G UAUCAUCG 747 CGAUGAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAUCAGC 2462
328 AUCAUCGU G AUGCUUCU 748 AGAAGCAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACGAUGAU 2463
331 AUCGUGAU G CUUCUCUG 749 CAGAGAAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCACGAU 2464
339 GCUUCUCU G AAGUUCUG 750 CAGAACUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAGAAGC 2465
347 GAAGUUCU G CUACAACC 751 GGUUGUAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAACUUC 2466
365 CUAGAUCU G CAGCUUGC 752 GCAAGCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAUCUAG 2467
372 UGCAGCUU G CCACAUCA 753 UGAUGUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGCUGCA 2468
392 UAAAAUCU G UCAUCCCA 754 UGGGAUGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAUUUUA 2469
402 CAUCCCAU G CAGACAGG 755 CCUGUCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGGGAUG 2470
422 ACAAUAUU G UAUAACAG 756 CUGUUAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUAUUGU 2471
441 CACUUCCU G AGUAGAAG 757 CUUCUACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGAAGUG 2472
459 GUUUCUUU G UGAAAAGG 758 CCUUUUCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAGAAAC 2473
461 UUCUUUGU G AAAAGGUC 759 GACCUUUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAAAGAA 2474
Table 29
492 AACUUAUU G UUACCAUA 760 UAUGGUAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAUAAGUU 2475
502 UACCAUAU G UAUUCAUC 761 GAUGAAUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUAUGGUA 2476
512 AUUCAUCU G UUGGAUCU 762 AGAUCCAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGAUGAAU 2477
522 UGGAUCUU G UAAACAUG 763 CAUGUUUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAGAUCCA 2478
530 GUAAACAU G AAAAGGGC 764 GCCCUUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUGUUUAC 2479
570 AAAUAAGU G UAUAAAAU 765 AUUUUAUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACUUAUUU 2480
579 UAUAAAAU G CAACUGUU 766 AACAGUUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUUUUAUA 2481
585 AUGCAACU G UUGAUUUC 767 GAAAUCAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGUUGCAU 2482
588 CAACUGUU G AUUUCCUC 768 GAGGAAAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AACAGUUG 2483
633 UCUUUUCU G AAGAUGAA 769 UUCAUCUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAAAAGA 2484
639 CUGAAGAU G AAGAGUUU 770 AAACUCUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCUUCAG 2485
660 UUAAAACU G CACUGCCA 771 UGGCAGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUUUUAA 2486
665 ACUGCACU G CCAACAAG 772 CUUGUUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUGCAGU 2487
710 ACUCUUUU G AGGUGAAU 773 AUUCACCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAAGAGU 2488
715 UUUGAGGU G AAUAUAAU 774 AUUAUAUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCUCAAA 2489
736 AUUACAAU G UAAAAGCU 775 AGCUUUUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUUGUAAU 2490
802 ACACAAAU G AAGUGUCA 776 UGACACUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUUUGUGU 2491
807 AAUGAAGU G UCAUUAUU 777 AAUAAUGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACUUCAUU 2492
830 AGUCCACU G ACUCCUCA 778 UGAGGAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUGGACU 2493
844 UCACAUCU G UUAUCUUA 779 UAAGAUAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAUGUGA 2494
869 AACUAUUU G UAGUAACU 780 AGUUACUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAUAGUU 2495
907 CAGAAAUU G UAUUUUUU 781 AAAAAAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUUUCUG 2496
920 UUUUCUAU G CCACAUUA 782 UAAUGUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUAGAAAA 2497
944 UUAAAGUU G AUGAGAAU 783 AUUCUCAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AACUUUAA 2498
947 AAGUUGAU G AGAAUCAA 784 UUGAUUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCAACUU 2499
1039 UAGUACAU G UAGGUAAA 785 UUUACCUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGUACUA 2500
1058 AUAAAUCU G UUCUAAGA 786 UCUUAGAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAUUUAU 2501
1072 AGACAUAU G AUCAACAG 787 CUGUUGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUAUGUCU 2502
1083 CAACAGAU G AGAACUGG 788 CCAGUUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCUGUUG 2503
1102 GUUAAUAU G UGACAGUG 789 CACUGUCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUAUUAAC 2504
1104 UAAUAUGU G ACAGUGAG 790 CUCACUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAUAUUA 2505
Table 29
1110 GUGACAGU G AGAUUAGU 791 ACUAAUCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACUGUCAC 2506
1168 AAGGCACU G UAGUGAAU 792 AUUCACUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGUGCCUU 2507
1173 ACUGUAGU G AAUUAUCU 793 AGAUAAUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACUACAGU 2508
1182 AAUUAUCU G AGCUAGAG 794 CUCUAGCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGAUAAUU 2509
1226 GGAAUCAU G AAACCUUA 795 UAAGGUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUGAUUCC 2510
1247 UUCAGAAU G AUUUUGCA 796 UGCAAAAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUUCUGAA 2511
1253 AUGAUUUU G CAGGUUGU 797 ACAACCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAAUCAU 2512
1260 UGCAGGUU G UCUUCCAU 798 AUGGAAGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AACCUGCA 2513
1287 CAUCCAAU G CAGGCAAG 799 CUUGCCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUUGGAUG 2514
1316 UUUCCAGU G ACAGAAAA 800 UUUUCUGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACUGGAAA 2515
1358 AAAUAUAU G AAUUCUCU 801 AGAGAAUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUAUAUUU 2516
1401 UAUAUUUU G AAAUGAAC 802 GUUCAUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAAUAUA 2517
1406 UUUGAAAU G AACUUGUU 803 AACAAGUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUUUCAAA 2518
1412 AUGAACUU G UUGGCCCA 804 UGGGCCAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGUUCAU 2519
1439 CUACAGCU G ACCCUUGA 805 UCAAGGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCUGUAG 2520
1446 UGACCCUU G AACAUGGG 806 CCCAUGUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGGGUCA 2521
1468 GGGGAGCU G ACAAUUCG 807 CGAAUUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCUCCCC 2522
1484 GUGGGUCC G CAAAAUCU 808 AGAUUUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGACCCAC 2523
1516 CUACUAUU G ACCAUAAA 809 UUUAUGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUAGUAG 2524
1532 ACCUUACU G AUAACAUA 810 UAUGUUAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUAAGGU 2525
1564 CAUAUUUU G CGUGUUAU 811 AUAACACG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAAUAUG 2526
1568 UUUUGCGU G UUAUAUGU 812 ACAUAUAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACGCAAAA 2527
1575 UGUUAUAU G UAUUAUAC 813 GUAUAAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUAUAACA 2528
1619 GAGAAAAU G UUAUUUAG 814 CUAAAUAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUUUUCUC 2529
21 ACUCCCCA G CUAAACAC 815 GUGUUUAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGGGAGU 2530
32 AAACACCC G UAAGACUU 816 AAGUCUUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGUGUUU 2531
76 UGAUCACA G CUGCCAAG 817 CUUGGCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUGAUCA 2532
85 CUGCCAAG G CUACCUAA 818 UUAGGUAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUGGCAG 2533
103 AGAAGACA G UUAUCUCA 819 UGAGAUAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUCUUCU 2534
118 CAUAUUUG G CUGCCAGC 820 GCUGGCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAAUAUG 2535
125 GGCUGCCA G CUUUUUAU 821 AUAAAAAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGCAGCC 2536
Table 29
177 UCCUGCUG G UAUCAUGG 822 CCAUGAUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGCAGGA 2537
191 UGGAGAAA G UCCAAUAC 823 GUAUUGGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUUCUCCA 2538
212 CUCGCUCA G CUAUAAGA 824 UCUUAUAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGAGCGAG 2539
224 UAAGAAGA G CCUCAACC 825 GGUUGAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCUUCUUA 2540
251 CUCAACAA G CACGUCAA 826 UUGACGUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUGUUGAG 2541
255 ACAAGCAC G UCAAAAGC 827 GCUUUUGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUGCUUGU 2542
262 CGUCAAAA G CUACAGAA 828 UUCUGUAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUUUGACG 2543
326 GUAUCAUC G UGAUGCUU 829 AAGCAUCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAUGAUAC 2544
342 UCUCUGAA G UUCUGCUA 830 UAGCAGAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUCAGAGA 2545
368 GAUCUGCA G CUUGCCAC 831 GUGGCAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCAGAUC 2546
381 CCACAUCA G CUUAAAAU 832 AUUUUAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGAUGUGG 2547
443 CUUCCUGA G UAGAAGAG 833 CUCUUCUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCAGGAAG 2548
451 GUAGAAGA G UUUCUUUG 834 CAAAGAAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCUUCUAC 2549
467 GUGAAAAG G UCAAGAUU 835 AAUCUUGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUUUCAC 2550
537 UGAAAAGG G CUUUAUUU 836 AAAUAAAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCUUUUCA 2551
568 CAAAAUAA G UGUAUAAA 837 UUUAUACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUAUUUUG 2552
603 UCAACAUG G CUCACAAA 838 UUUGUGAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUGUUGA 2553
644 GAUGAAGA G UUUAGUUU 839 AAACUAAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCUUCAUC 2554
649 AGAGUUUA G UUUUAAAA 840 UUUUAAAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAAACUCU 2555
673 GCCAACAA G UUCACUUC 841 GAAGUGAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGUUGGC 2556
691 UAUAUAAA G CAUUAUUU 842 AAAUAAUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUAUAUA 2557
713 CUUUUGAG G UGAAUAUA 843 UAUAUUCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCAAAAG 2558
742 AUGUAAAA G CUUCUUUA 844 UAAAGAAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUUACAU 2559
758 AAUACUAA G UAUUUUUC 845 GAAAAAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUAGUAUU 2560
769 UUUUUCAG G UCUUCACC 846 GGUGAAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGAAAAA 2561
780 UUCACCAA G UAUCAAAG 847 CUUUGAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGGUGAA 2562
788 GUAUCAAA G UAAUAACA 848 UGUUAUUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUGAUAC 2563
805 CAAAUGAA G UGUCAUUA 849 UAAUGACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCAUUUG 2564
823 UCAAAAUA G UCCACUGA 850 UCAGUGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAUUUUGA 2565
872 UAUUUGUA G UAACUAUC 851 GAUAGUUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UACAAAUA 2566
941 CUUUUAAA G UUGAUGAG 852 CUCAUCAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUAAAAG 2567
Table 29
956 AGAAUCAA G UAUGGAAA 853 UUUCCAUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUGAUUCU 2568
966 AUGGAAAA G UAAGGCCA 854 UGGCCUUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUUUCCAU 2569
971 AAAGUAAG G CCAUACUC 855 GAGUAUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUUACUUU 2570
1003 CCUUUUAA G UAAUUUUU 856 AAAAAUUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUAAAAGG 2571
1033 GAAUUCUA G UACAUGUA 857 UACAUGUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAGAAUUC 2572
1043 ACAUGUAG G UAAAUCAU 858 AUGAUUUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUACAUGU 2573
1091 GAGAACUG G UGGUUAAU 859 AUUAACCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGUUCUC 2574
1094 AACUGGUG G UUAAUAUG 860 CAUAUUAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACCAGUU 2575
1108 AUGUGACA G UGAGAUUA 861 UAAUCUCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGUCACAU 2576
1117 UGAGAUUA G UCAUAUCA 862 UGAUAUGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAAUCUCA 2577
1163 CAUUUAAG G CACUGUAG 863 CUACAGUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUUAAAUG 2578
1171 GCACUGUA G UGAAUUAU 864 AUAAUUCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UACAGUGC 2579
1184 UUAUCUGA G CUAGAGUU 865 AACUCUAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCAGAUAA 2580
1190 GAGCUAGA G UUACCUAG 866 CUAGGUAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCUAGCUC 2581
1198 GUUACCUA G CUUACCAU 867 AUGGUAAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAGGUAAC 2582
1257 UUUUGCAG G UUGUCUUC 868 GAAGACAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGCAAAA 2583
1273 CCAUUCCA G CCUAACAU 869 AUGUUAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGAAUGG 2584
1291 CAAUGCAG G CAAGGAAA 870 UUUCCUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGCAUUG 2585
1314 GAUUUCCA G UGACAGAA 871 UUCUGUCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGAAAUC 2586
1339 UAUCUCAA G UAUUUUUU 872 AAAAAAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGAGAUA 2587
1416 ACUUGUUG G CCCAUCUA 873 UAGAUGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAACAAGU 2588
1436 CAUCUACA G CUGACCCU 874 AGGGUCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUAGAUG 2589
1456 ACAUGGGG G UUAGGGGA 875 UCCCCUAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCCAUGU 2590
1465 UUAGGGGA G CUGACAAU 876 AUUGUCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCCCUAA 2591
1476 GACAAUUC G UGGGUCCG 877 CGGACCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GAAUUGUC 2592
1480 AUUCGUGG G UCCGCAAA 878 UUUGCGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCACGAAU 2593
1506 ACCUAAUA G CCUACUAU 879 AUAGUAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAUUAGGU 2594
1545 CAUAAACA G UAAAUUAA 880 UUAAUUUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUUUAUG 2595
1566 UAUUUUGC G UGUUAUAU 881 AUAUAACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCAAAAUA 2596
1603 ACAAUAAA G UAAGCUAG 882 CUAGCUUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUAUUGU 2597
1607 UAAAGUAA G CUAGAGAA 883 UUCUCUAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUACUUUA 2598
Table 29
CAGAGUCA G AAAACUCC 1060 GGAGUUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGACUCUG 2599
ACCCGUAA G ACUUCAUA 1061 UAUGAAGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUACGGGU 2600
GCUGCCAA G GCUACCUA 1062 UAGGUAGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUGGCAGC 2601
ACCUAAAA G AAGACAGU 1063 ACUGUCUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUUUAGGU 2602
UAAAAGAA G ACAGUUAU 1064 AUAACUGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUCUUUUA 2603
UCAUAUUU G GCUGCCAG 1065 CUGGCAGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAUAUGA 2604
AAACUUCA σ ACUUCCUG 1066 CAGGAAGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGAAGUUU 2605
GUCCUGCU G GUAUCAUG 1067 CAUGAUAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCAGGAC 2606
GGUAUCAU G GAGAAAGU 1068 ACUUUCUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUGAUACC 2607
GUAUCAUG G AGAAAGUC 1069 GACUUUCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUGAUAC 2608
AUCAUGGA G AAAGUCCA 1070 UGGACUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCCAUGAU 2609
AGCUAUAA G AAGAGCCU 1071 AGGCUCUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUAUAGCU 2610
UAUAAGAA G AGCCUCAA 1072 UUGAGGCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCUUAUA 2611
AAGCUACA G AAUCUAUU 1073 AAUAGAUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUAGCUU 2612
AACCUCUA G AUCUGCAG 1074 CUGCAGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAGAGGUU 2613
CCCAUGCA G ACAGGAAA 1075 UUUCCUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCAUGGG 2614
UGCAGACA G GAAAACAA 1076 UUGUUUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUCUGCA 2615
GCAGACAG G AAAACAAU 1077 AUUGUUUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGUCUGC 2616
GUAUAACA G ACCACUUC 1078 GAAGUGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUUAUAC 2617
CCUGAGUA G AAGAGUUU 1079 AAACUCUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UACUCAGG 2618
GAGUAGAA G AGUUUCUU 1080 AAGAAACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCUACUC 2619
UGUGAAAA G GUCAAGAU 1081 AUCUUGAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUUCACA 2620
AAGGUCAA G AUUAAGAC 1082 GUCUUAAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGACCUU 2621
AAGAUUAA G ACUAAAAC 1083 GUUUUAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUAAUCUU 2622
CAUCUGUU G GAUCUUGU 1084 ACAAGAUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AACAGAUG 2623
AUCUGUUG G AUCUUGUA 1085 UACAAGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAACAGAU 2624
CAUGAAAA G GGCUUUAU 1086 AUAAAGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUUCAUG 2625
AUGAAAAG G GCUUUAUU 1087 AAUAAAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUUUCAU 2626
CUCAACAU G GCUCACAA 1088 UUGUGAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGUUGAG 2627
UUUCUGAA G AUGAAGAG 1089 CUCUUCAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCAGAAA 2628
AAGAUGAA G AGUUUAGU 1090 ACUAAACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCAUCUU 2629
Table 29
712 UCUUUUGA G GUGAAUAU 1091 AUAUUCAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCAAAAGA 2630
768 AUUUUUCA G GUCUUCAC 1092 GUGAAGAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGAAAAAU 2631
860 AUUAUAAA G AACUAUUU 1093 AAAUAGUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUUAUAAU 2632
882 AACUAUCA G AAUCUACA 1094 UGUAGAUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGAUAGUU 2633
901 CUAAAACA G AAAUUGUA 1095 UACAAUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGUUUUAG 2634
949 GUUGAUGA G AAUCAAGU 1096 ACUUGAUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCAUCAAC 2635
960 UCAAGUAU G GAAAAGUA 1097 UACUUUUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUACUUGA 2636
961 CAAGUAUG G AAAAGUAA 1098 UUACUUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUACUUG 2637
970 AAAAGUAA G GCCAUACU 1099 AGUAUGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUACUUUU 2638
1017 UUUUCAAA G AAUCACAG 1100 CUGUGAUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUUGAAAA 2639
1025 GAAUCACA G AAUUCUAG 1101 CUAGAAUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGUGAUUC 2640
1042 UACAUGUA G GUAAAUCA 1102 UGAUUUAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UACAUGUA 2641
1065 UGUUCUAA G ACAUAUGA 1103 UCAUAUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUAGAACA 2642
1080 GAUCAACA G AUGAGAAC 1104 GUUCUCAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUUGAUC 2643
1085 ACAGAUGA G AACUGGUG 1105 CACCAGUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCAUCUGU 2644
1090 UGAGAACU G GUGGUUAA 1106 UUAACCAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUUCUCA 2645
1093 GAACUGGU G GUUAAUAU 1107 AUAUUAAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCAGUUC 2646
1112 GACAGUGA G AUUAGUCA 1108 UGACUAAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCACUGUC 2647
1143 UAACAACA G AAUCUAAU 1109 AUUAGAUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUUGUUA 2648
1162 UCAUUUAA G GCACUGUA 1110 UACAGUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUAAAUGA 2649
1188 CUGAGCUA G AGUUACCU llll AGGUAACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAGCUCAG 2650
1218 AUAUCUUU G GAAUCAUG 1112 CAUGAUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAGAUAU 2651
1219 UAUCUUUG G AAUCAUGA 1113 UCAUGAUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAAGAUA 2652
1236 AACCUUAA G ACUUCAGA 1114 UCUGAAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUAAGGUU 2653
1243 AGACUUCA G AAUGAUUU 1115 AAAUCAUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGAAGUCU 2654
1256 AUUUUGCA G GUUGUCUU 1116 AAGACAAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCAAAAU 2655
1290 CCAAUGCA G GCAAGGAA 1117 UUCCUUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCAUUGG 2656
1295 GCAGGCAA G GAAAAUAA 1118 UUAUUUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGCCUGC 2657
1296 CAGGCAAG G AAAAUAAA 1119 UUUAUUUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUGCCUG 2658
1306 AAAUAAAA G AUUUCCAG 1120 CUGGAAAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUUAUUU 2659
1320 CAGUGACA G AAAAAUAU 1121 AUAUUUUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUCACUG 2660
Table 29
1390 AAUUAUUA G AUUAUAUU 1122 AAUAUAAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAAUAAUU 2661
1415 AACUUGUU G GCCCAUCU 1123 AGAUGGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AACAAGUU 2662
1452 UUGAACAU G GGGGUUAG 1124 CUAACCCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUGUUCAA 2663
1453 UGAACAUG G GGGUUAGG 1125 CCUAACCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUGUUCA 2664
1454 GAACAUGG G GGUUAGGG 1126 CCCUAACC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCAUGUUC 2665
1455 AACAUGGG G GUUAGGGG 1127 CCCCUAAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCAUGUU 2666
1460 GGGGGUUA G GGGAGCUG 1128 CAGCUCCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAACCCCC 2667
1461 GGGGUUAG G GGAGCUGA 1129 UCAGCUCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUAACCCC 2668
1462 GGGUUAGG G GAGCUGAC 1130 GUCAGCUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCUAACCC 2669
1463 GGUUAGGG G AGCUGACA 1131 UGUCAGCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCUAACC 2670
1478 CAAUUCGU G GGUCCGCA 1132 UGCGGACC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACGAAUUG 2671
1479 AAUUCGUG G GUCCGCAA 1133 UUGCGGAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACGAAUU 2672
1611 GUAAGCUA G AGAAAAUG 1134 CAUUUUCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAGCUUAC 2673
1613 AAGCUAGA G AAAAUGUU 1135 AACAUUUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCUAGCUU 2674
1627 GUUAUUUA G AAAAUCAU 1136 AUGAUUUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAAAUAAC 2675
Input Sequence = PLN. Cut Site = Gl.
Stem Length = 8. Core Sequence = GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG
PLN (Homo sapiens phospholamban (PLN) mRNA.; 1635 bp)
Table 30 Table 30: Human Phospholamban (PLN) Antisense and Target Sequence
Input Sequence = PLN
Oligo Length = 25
PLN (Homo sapiens phospholamban (PLN) mRNA.; 1635 bp)
Table 31
Table 31: Anti-Her2 Ribozyme and Substrate Sequence
RPI nt. Seq ID Seq ID NOS Position Nos Nos
NCH Ribozyme Sequence NCH Substrate Sequence
17214 175 asgscsuscca cUGAuGaggccguuaggccGaa Igugcuc B GAGCACC A UGGAGCU
17215 212 gsasgsgsgcg cUGAuGaggccguuaggccGaa Igaggag B CUCCUCC U CGCCCUC
17216 261 uscsasusguc cUGAuGaggccguuaggccGaa Iugccgg B CCGGCAC A GACAUGA
17217 297 cscsasgsgug cUGAuGaggccguuaggccGaa lucucgg B CCGAGAC C CACCUGG
17218 381 ascsasgsgcu cUGAuGaggccguuaggccGaa Icauugg B CCAAUGC C AGCCUGU
17219 619 cscsususuca cUGAuGaggccguuaggccGaa Iaucucu B AGAGAUC U UGAAAGG
17220 665 csgsusgsucc cUGAuGaggccguuaggccGaa Iguagca B UGCUACC A GGACACG
17221 943 ususgsasagu cUGAuGaggccguuaggccGaa Iaggcag B CUGCCUC C ACUUCAA
17222 944 gsususgsaag cUGAuGaggccguuaggccGaa Igaggca B UGCCUCC A CUUCAAC
17223 946 usgsgsusuga cUGAuGaggccguuaggccGaa Iuggagg B CCUCCAC U UCAACCA
17224 993 usgsususgua cUGAuGaggccguuaggccGaa Iugacca B UGGUCAC C UACAACA
17225 997 uscsusgsugu cUGAuGaggccguuaggccGaa Iuaggug B CACCUAC A ACACAGA
17226 1000 gsusgsuscug cUGAuGaggccguuaggccGaa Iuuguag B CUACAAC A CAGACAC
17227 1017 usgsgsgscau cUGAuGaggccguuaggccGaa lacucaa B UUGAGUC C AUGCCCA
17228 1078 asgsgsusagu cUGAuGaggccguuaggccGaa Iuaggga B UCCCUAC A ACUACCU
17229 1201 asgsascscau cUGAuGaggccguuaggccGaa Icacacu B AGUGUGC U AUGGUCU
17230 1285 csuscscscaa cUGAuGaggccguuaggccGaa Iaucuuc B GAAGAUC U UUGGGAG
17231 1395 gsgsusasacc cUGAuGaggccguuaggccGaa Iugaucu B AGAUCAC A GGUUACC
17232 1413 gscscsasugc cUGAuGaggccguuaggccGaa lagaugu B • ACAUCUC A GCAUGGC
17233 1450 asgsgsusucu cUGAuGaggccguuaggccGaa Iaagacg B CGUCUUC C AGAACCU
17234 1624 asasgsasgcu cUGAuGaggccguuaggccGaa Iucccag B CUGGGAC C AGCUCUU
17235 1787 gscsascsucc CUGAuGaggccguuaggccGaa Igccccg B CGGGGCC A GGAGUGC
17236 2001 gscsasusgua cUGAuGaggccguuaggccGaa Iagaggu B ACCUCUC C UACAUGC
17237 2002 gsgscsasugu cUGAuGaggccguuaggccGaa Igagagg B CCUCUCC U ACAUGCC
17238 2055 asgsususgau cUGAuGaggccguuaggccGaa Iggcaag B CUUGCCC C AUCAACU
Table 31
17239 2056 csasgsusuga cUGAuGaggccguuaggccGaa Igggcaa B UUGCCCC A UCAACUG
17240 2068 csasgsgsagu cUGAuGaggccguuaggccGaa Igugcag B CUGCACC C ACUCCUG
17241 2069 ascsasgsgag cUGAuGaggccguuaggccGaa Iggugca B UGCACCC A CUCCUGU
17242 2295 cscsusgsguu cUGAuGaggccguuaggccGaa Igcaucg B CGAUGCC C AACCAGG
17243 2351 asgsasuscca cUGAuGaggccguuaggccGaa Icaccuu B AAGGUGC U UGGAUCU
17244 2373 usgsusasgac cUGAuGaggccguuaggccGaa Iugccaa B UUGGCAC A GUCUACA
17245 2570 csasusasagc cUGAuGaggccguuaggccGaa Iugucac B GUGACAC A GCUUAUG
17246 2665 asuscscsccu cUGAuGaggccguuaggccGaa Igcaauc B GAUUGCC A AGGGGAU
17247 2702 gsuscscscug cUGAuGaggccguuaggccGaa luacgag B CUCGUAC A CAGGGAC
17248 2771 cscsgsasgcc cUGAuGaggccguuaggccGaa Icccgaa B UUCGGGC U GGCUCGG
18261 2780 gsuscscsagc cUGAuGaggccguuaggccGaa Iccgagc B GCUCGGC U GCUGGAC
17249 2783 asasusgsucc cUGAuGaggccguuaggccGaa Icagccg B CGGCUGC U GGACAUU
18265 2783 asusgsuscc cUGAuGaggccguuaggccGaa Icagcc B GGCUGC U GGACAU
18267 2783 asasusgsucc cUGAuGaggcguuagccGaa ' [cagccg B CGGCUGC U GGACAUU
18269 2783 asusgsuscc cUGAuGaggcguuagccGaa : [cagcc B GGCUGC U GGACAU
18271 2783 asasusgsucc cUGAuGaggccguuaggccGaa Icagccg B CGGCUGC U GGACAUU
18273 2783 asusgsuscc cUGAuGaggccguuaggccGaa Icagcc B GGCUGC U GGACAU
18275 2783 asasusgsucc cUGAuGaggcguuagccGaa [cagccg B CGGCUGC U GGACAUU
18277 2783 asusgsuscc cUGAuGaggcguuagccGaa [cagcc B GGCUGC U GGACAU
18262 2788 uscsgsuscaa cUGAuGaggccguuaggccGaa Iuccagc B GCUGGAC A UUGACGA
17250 2799 gsgsusascuc cUGAuGaggccguuaggccGaa lucucgu B ACGAGAC A GAGUACC
18263 2935 usasasgsguu cUGAuGaggccguuaggccGaa Igcccca B UGGGGCC A AACCUUA
17251 2939 asuscsgsuaa cUGAuGaggccguuaggccGaa Iuuuggc B GCCAAAC C UUACGAU
18266 2939 uscsgsusaa cUGAuGaggccguuaggccGaa Iuuugg B CCAAAC C UUACGA
18268 2939 asuscsgsuaa cUGAuGaggcguuagccGaa [uuuggc B GCCAAAC C UUACGAU
18270 2939 uscsgsusaa cUGAuGaggcguuagccGaa [uuugg B CCAAAC C UUACGA
18272 2939 asuscsgsuaa cUGAuGaggccguuaggccGaa Iuuuggc B GCCAAAC C UUACGAU
18274 2939 uscsgsusaa cUGAuGaggccguuaggccGaa Iuuugg B CCAAAC C UUACGA
18276 2939 asuscsgsuaa cUGAuGaggcguuagccGaa [uuuggc B GCCAAAC C UUACGAU
18278 2939 uscsgsusaa cUGAuGaggcguuagccGaa [uuugg B CCAAAC C UUACGA
18264 2940 csasuscsgua cUGAuGaggccguuaggccGaa Iguuugg B CCAAACC U UACGAUG
Table 31
Table 31
17194 2003 gsgsgscsaug cUGAuGaggccguuaggccGaa Aggagag B CUCUCCU A CAUGCCC
17205 2058 usgscsasguu cUGAuGaggccguuaggccGaa Auggggc B GCCCCAU C AACUGCA
17206 2072 csascsascag cUGAuGaggccguuaggccGaa Agugggu B ACCCACU C CUGUGUG
17195 2352 csasgsasucc cUGAuGaggccguuaggccGaa Agcaccu B AGGUGCU U GGAUCUG
17196 2575 usasgsgsgca cUGAuGaggccguuaggccGaa Aagcugu B ACAGCUU A UGCCCUA
17197 2697 usgsusgsuac cUGAuGaggccguuaggccGaa Agccgca B UGCGGCU C GUACACA
18257 2776 asgscsasgcc cUGAuGaggccguuaggccGaa Agccagc B GCUGGCU C GGCUGCU
18258 2790 uscsuscsguc cUGAuGaggccguuaggccGaa Augucca B UGGACAU U GACGAGA
17207 2926 gscscscscaa cUGAuGaggccguuaggccGaa Agucauc B GAUGACU U UUGGGGC
18259 2928 usgsgscsccc cUGAuGaggccguuaggccGaa Aaaguca B UGACUUU U GGGGCCA
18260 2942 cscscsasucg cUGAuGaggccguuaggccGaa Aagguuu B AAACCUU A CGAUGGG
17198 3089 asgsascsacc cUGAuGaggccguuaggccGaa Acucccg B CGGGAGU U GGUGUCU
17208 3155 usgsgsgsccc cUGAuGaggccguuaggccGaa Aguccuc B GAGGACU U GGGCCCA
17209 3499 csgscsusgua cUGAuGaggccguuaggccGaa Agggcug B CAGCCCU C UACAGCG
17210 3501 ascscsgscug cUGAuGaggccguuaggccGaa Agagggc B GCCCUCU A CAGCGGU
17211 3714 csusususgac cUGAuGaggccguuaggccGaa Accccau B AUGGGGU C GUCAAAG
17199 3802 gscsasgsgag cUGAuGaggccguuaggccGaa Agggugg B CCACCCU C CUCCUGC
17200 3825 gsgsususguc cUGAuGaggccguuaggccGaa Aaggcug B CAGCCUU C GACAACC
17201 3838 uscscscsagu cUGAuGaggccguuaggccGaa Auagagg B CCUCUAU U ACUGGGA
Lowercase = 2'-O-methyl Modifications
Uppercase: 1. Uppercase U under Ribozyme Sequence = 2'-C-Allyl U
2. All other Uppercases = Ribonucleotides I = Inosine
B = 3 '-3' Inverted abasic deoxyribose
Table 32
Table 32: RNA Cleavage by NCH-XYLO Ribozyme (Reaction: 50 mM TRIS-CI pH 7.5, 10 mM Mg2+, 37°C, 500 nM FINAL [Rz], Trace
Substrate)
DATA SUMMARY
Table 33
Table 33: Examples of NCH-Xylo Ribozyme and Substrate Sequences
Uppercase = Ribonucleotides
Lowercase = 2'-0-methyl nucleotides
I = Xylo -Inosine
B = 3'-3' inverted abasic
U = 2'-C-allyl-U
Table 34 Table 34: Anti-HER2 NCH Ribozyme and Target Sequences
118 CGCGCCC C GCGCCCU AGGGCGC CUGAUGAG X CGAA IGGCGCG
123 CCCGCGC C CUCCCAG CUGGGAG CUGAUGAG X CGAA ICGCGGG
124 CCGCGCC C UCCCAGC GCUGGGA CUGAUGAG X CGAA IGCGCGG
125 CGCGCCC U CCCAGCC GGCUGGG CUGAUGAG X CGAA IGGCGCG
127 CGCCCUC C CAGCCGG CCGGCUG CUGAUGAG X CGAA IAGGGCG
128 GCCCUCC C AGCCGGG CCCGGCU CUGAUGAG X CGAA IGAGGGC
129 CCCUCCC A GCCGGGU ACCCGGC CUGAUGAG X CGAA IGGAGGG
132 UCCCAGC C GGGUCCA UGGACCC CUGAUGAG X CGAA ICUGGGA
138 CCGGGUC C AGCCGGA UCCGGCU CUGAUGAG X CGAA IACCCGG
139 CGGGUCC A GCCGGAG CUCCGGC CUGAUGAG X CGAA IGACCCG
142 GUCCAGC C GGAGCCA UGGCUCC CUGAUGAG X CGAA ICUGGAC
148 CCGGAGC C AUGGGGC GCCCCAU CUGAUGAG X CGAA ICUCCGG
149 CGGAGCC A UGGGGCC GGCCCCA CUGAUGAG X CGAA IGCUCCG
156 AUGGGGC C GGAGCCG CGGCUCC CUGAUGAG X CGAA ICCCCAU
162 CCGGAGC C GCAGUGA UCACUGC CUGAUGAG X CGAA ICUCCGG
165 GAGCCGC A GUGAGCA UGCUCAC CUGAUGAG X CGAA ICGGCUC
172 AGUGAGC A CCAUGGA UCCAUGG CUGAUGAG X CGAA ICUCACU
174 UGAGCAC C AUGGAGC GCUCCAU CUGAUGAG X CGAA IUGCUCA
175 GAGCACC A UGGAGCU AGCUCCA CUGAUGAG X CGAA IGUGCUC
182 AUGGAGC U GGCGGCC GGCCGCC CUGAUGAG X CGAA ICUCCAU
189 UGGCGGC C UUGUGCC GGCACAA CUGAUGAG X CGAA ICCGCCA
190 GGCGGCC U UGUGCCG CGGCACA CUGAUGAG X CGAA IGCCGCC
196 CUUGUGC C GCUGGGG CCCCAGC CUGAUGAG X CGAA ICACAAG
199 GUGCCGC U GGGGGCU AGCCCCC CUGAUGAG X CGAA ICGGCAC
206 UGGGGGC U CCUCCUC GAGGAGG CUGAUGAG X CGAA ICCCCCA
208 GGGGCUC C UCCUCGC GCGAGGA CUGAUGAG X CGAA IAGCCCC
209 GGGCUCC U CCUCGCC GGCGAGG CUGAUGAG X CGAA IGAGCCC
211 GCUCCUC C UCGCCCU AGGGCGA CUGAUGAG X CGAA IAGGAGC
212 CUCCUCC U CGCCCUC GAGGGCG CUGAUGAG X CGAA IGAGGAG
216 UCCUCGC C CUCUUGC GCAAGAG CUGAUGAG X CGAA ICGAGGA
217 CCUCGCC C UCUUGCC GGCAAGA CUGAUGAG X CGAA IGCGAGG
218 CUCGCCC U CUUGCCC GGGCAAG CUGAUGAG X CGAA IGGCGAG
220 CGCCCUC U UGCCCCC GGGGGCA CUGAUGAG X CGAA IAGGGCG
224 cucuuσc C CCCCGGA UCCGGGG CUGAUGAG X CGAA ICAAGAG
225 UCUUGCC C CCCGGAG CUCCGGG CUGAUGAG X CGAA IGCAAGA
226 CUUGCCC c CCGGAGC GCUCCGG CUGAUGAG X CGAA IGGCAAG
227 UUGCCCC c CGGAGCC GGCUCCG CUGAUGAG X CGAA IGGGCAA
228 UGCCCCC c GGAGCCG CGGCUCC CUGAUGAG X CGAA IGGGGCA
234 CCGGAGC c GCGAGCA UGCUCGC CUGAUGAG X CGAA ICUCCGG
241 CGCGAGC A CCCAAGU ACUUGGG CUGAUGAG X CGAA ICUCGCG
243 CGAGCAC C CAAGUGU ACACUUG CUGAUGAG X CGAA IUGCUCG
244 GAGCACC C AAGUGUG CACACUU CUGAUGAG X CGAA IGUGCUC
245 AGCACCC A AGUGUGC GCACACU CUGAUGAG X CGAA IGGUGCU
253 AGUGUGC A CCGGCAC GUGCCGG CUGAUGAG X CGAA ICACACU
255 UGUGCAC C GGCACAG CUGUGCC CUGAUGAG X CGAA IUGCACA
259 CACCGGC A CAGACAU AUGUCUG CUGAUGAG X CGAA ICCGGUG
261 CCGGCAC A GACAUGA UCAUGUC CUGAUGAG X CGAA IUGCCGG
Table 34
265 CACAGAC A UGAAGCU AGCUUCA CUGAUGAG X CGAA IUCUGUG
272 AUGAAGC U GCGGCUC GAGCCGC CUGAUGAG X CGAA ICUUCAU
278 CUGCGGC u CCCUGCC GGCAGGG CUGAUGAG X CGAA ICCGCAG
280 GCGGCUC c CUGCCAG CUGGCAG CUGAUGAG X CGAA IAGCCGC
281 CGGCUCC c UGCCAGU ACUGGCA CUGAUGAG X CGAA IGAGCCG
282 GGCUCCC u GCCAGUC GACUGGC CUGAUGAG X CGAA IGGAGCC
285 UCCCUGC c AGUCCCG CGGGACU CUGAUGAG X CGAA ICAGGGA
286 CCCUGCC A GUCCCGA UCGGGAC CUGAUGAG X CGAA IGCAGGG
290 GCCAGUC C CGAGACC GGUCUCG CUGAUGAG X CGAA IACUGGC
291 CCAGUCC C GAGACCC GGGUCUC CUGAUGAG X CGAA IGACUGG
297 CCGAGAC C CACCUGG CCAGGUG CUGAUGAG X CGAA IUCUCGG
298 CGAGACC C ACCUGGA UCCAGGU CUGAUGAG X CGAA IGUCUCG
299 GAGACCC A CCUGGAC GUCCAGG CUGAUGAG X CGAA IGGUCUC
301 GACCCAC C UGGACAU AUGUCCA CUGAUGAG X CGAA IUGGGUC
302 ACCCACC U GGACAUG CAUGUCC CUGAUGAG X CGAA IGUGGGU
307 CCUGGAC A UGCUCCG CGGAGCA CUGAUGAG X CGAA IUCCAGG
311 GACAUGC U CCGCCAC GUGGCGG CUGAUGAG X CGAA ICAUGUC
313 CAUGCUC C GCCACCU AGGUGGC CUGAUGAG X CGAA IAGCAUG
316 GCUCCGC C ACCUCUA UAGAGGU CUGAUGAG X CGAA ICGGAGC
317 CUCCGCC A CCUCUAC GUAGAGG CUGAUGAG X CGAA IGCGGAG
319 CCGCCAC C UCUACCA UGGUAGA CUGAUGAG X CGAA IUGGCGG
320 CGCCACC U CUACCAG CUGGUAG CUGAUGAG X CGAA IGUGGCG
322 CCACCUC U ACCAGGG CCCUGGU CUGAUGAG X CGAA IAGGUGG
325 CCUCUAC C AGGGCUG CAGCCCU CUGAUGAG X CGAA IUAGAGG
326 CUCUACC A GGGCUGC GCAGCCC CUGAUGAG X CGAA IGUAGAG
331 CCAGGGC U GCCAGGU ACCUGGC CUGAUGAG X CGAA ICCCUGG
334 GGGCUGC C AGGUGGU ACCACCU CUGAUGAG X CGAA ICAGCCC
335 GGCUGCC A GGUGGUG CACCACC CUGAUGAG X CGAA IGCAGCC
344 GUGGUGC A GGGAAAC GUUUCCC CUGAUGAG X CGAA ICACCAC
352 GGGAAAC C UGGAACU AGUUCCA CUGAUGAG X CGAA IUUUCCC
353 GGAAACC U GGAACUC GAGUUCC CUGAUGAG X CGAA IGUUUCC
359 CUGGAAC U CACCUAC GUAGGUG CUGAUGAG X CGAA IUUCCAG
361 GGAACUC A CCUACCU AGGUAGG CUGAUGAG X CGAA IAGUUCC
363 AACUCAC C UACCUGC GCAGGUA CUGAUGAG X CGAA IUGAGUU
364 ACUCACC U ACCUGCC GGCAGGU CUGAUGAG X CGAA IGUGAGU
367 CACCUAC C UGCCCAC GUGGGCA CUGAUGAG X CGAA IUAGGUG
368 ACCUACC U GCCCACC GGUGGGC CUGAUGAG X CGAA IGUAGGU
371 UACCUGC C CACCAAU AUUGGUG CUGAUGAG X CGAA ICAGGUA
372 ACCUGCC C ACCAAUG CAUUGGU CUGAUGAG X CGAA IGCAGGU
373 CCUGCCC A CCAAUGC GCAUUGG CUGAUGAG X CGAA IGGCAGG
375 UGCCCAC C AAUGCCA UGGCAUU CUGAUGAG X CGAA IUGGGCA
376 GCCCACC A AUGCCAG CUGGCAU CUGAUGAG X CGAA IGUGGGC
381 CCAAUGC C AGCCUGU ACAGGCU CUGAUGAG X CGAA ICAUUGG
382 CAAUGCC A GCCUGUC GACAGGC CUGAUGAG X CGAA IGCAUUG
385 UGCCAGC C UGUCCUU AAGGACA CUGAUGAG X CGAA ICUGGCA
386 GCCAGCC U GUCCUUC GAAGGAC CUGAUGAG X CGAA IGCUGGC
390 GCCUGUC C UUCCUGC GCAGGAA CUGAUGAG X CGAA IACAGGC
Table J
391 CCUGUCC U UCCUGCA UGCAGGA CUGAUGAG X CGAA IGACAGG
394 GUCCUUC C UGCAGGA UCCUGCA CUGAUGAG X CGAA IAAGGAC
395 UCCUUCC U GCAGGAU AUCCUGC CUGAUGAG X CGAA IGAAGGA
398 UUCCUGC A GGAUAUC GAUAUCC CUGAUGAG X CGAA ICAGGAA
406 GGAUAUC C AGGAGGU ACCUCCU CUGAUGAG X CGAA IAUAUCC
407 GAUAUCC A GGAGGUG CACCUCC CUGAUGAG X CGAA IGAUAUC
416 GAGGUGC A GGGCUAC GUAGCCC CUGAUGAG X CGAA ICACCUC
421 GCAGGGC U ACGUGCU AGCACGU CUGAUGAG X CGAA ICCCUGC
428 UACGUGC U CAUCGCU AGCGAUG CUGAUGAG X CGAA ICACGUA
430 CGUGCUC A UCGCUCA UGAGCGA CUGAUGAG X CGAA IAGCACG
435 UCAUCGC U CACAACC GGUUGUG CUGAUGAG X CGAA ICGAUGA
437 AUCGCUC A CAACCAA UUGGUUG CUGAUGAG X CGAA IAGCGAU
439 CGCUCAC A ACCAAGU ACUUGGU CUGAUGAG X CGAA IUGAGCG
442 UCACAAC C AAGUGAG CUCACUU CUGAUGAG X CGAA IUUGUGA
443 CACAACC A AGUGAGG CCUCACU CUGAUGAG X CGAA IGUUGUG
452 GUGAGGC A GGUCCCA UGGGACC CUGAUGAG X CGAA ICCUCAC
457 GCAGGUC C CACUGCA UGCAGUG CUGAUGAG X CGAA IACCUGC
458 CAGGUCC C ACUGCAG CUGCAGU CUGAUGAG X CGAA IGACCUG
459 AGGUCCC A CUGCAGA UCUGCAG CUGAUGAG X CGAA IGGACCU
461 GUCCCAC U GCAGAGG CCUCUGC CUGAUGAG X CGAA IUGGGAC
464 CCACUGC A GAGGCUG CAGCCUC CUGAUGAG X CGAA ICAGUGG
470 CAGAGGC U GCGGAUU AAUCCGC CUGAUGAG X CGAA ICCUCUG
487 GCGAGGC A CCCAGCU AGCUGGG CUGAUGAG X CGAA ICCUCGC
489 GAGGCAC C CAGCUCU AGAGCUG CUGAUGAG X CGAA IUGCCUC
490 AGGCACC C AGCUCUU AAGAGCU CUGAUGAG X CGAA IGUGCCU
491 GGCACCC A GCUCUUU AAAGAGC CUGAUGAG X CGAA IGGUGCC
494 ACCCAGC U CUUUGAG CUCAAAG CUGAUGAG X CGAA ICUGGGU
496 CCAGCUC U UUGAGGA UCCUCAA CUGAUGAG X CGAA IAGCUGG
505 UGAGGAC A ACUAUGC GCAUAGU CUGAUGAG X CGAA IUCCUCA
508 GGACAAC U AUGCCCU AGGGCAU CUGAUGAG X CGAA IUUGUCC
513 ACUAUGC C CUGGCCG CGGCCAG CUGAUGAG X CGAA ICAUAGU
514 CUAUGCC C UGGCCGU ACGGCCA CUGAUGAG X CGAA IGCAUAG
515 UAUGCCC U GGCCGUG CACGGCC CUGAUGAG X CGAA IGGCAUA
519 CCCUGGC C GUGCUAG CUAGCAC CUGAUGAG X CGAA ICCAGGG
524 GCCGUGC U AGACAAU AUUGUCU CUGAUGAG X CGAA ICACGGC
529 GCUAGAC A AUGGAGA UCUCCAU CUGAUGAG X CGAA IUCUAGC
538 UGGAGAC C CGCUGAA UUCAGCG CUGAUGAG X CGAA IUCUCCA
539 GGAGACC C GCUGAAC GUUCAGC CUGAUGAG X CGAA IGUCUCC
542 GACCCGC U GAACAAU AUUGUUC CUGAUGAG X CGAA ICGGGUC
547 GCUGAAC A AUACCAC GUGGUAU CUGAUGAG X CGAA IUUCAGC
552 ACAAUAC C ACCCCUG CAGGGGU CUGAUGAG X CGAA IUAUUGU
553 CAAUACC A CCCCUGU ACAGGGG CUGAUGAG X CGAA IGUAUUG
555 AUACCAC C CCUGUCA UGACAGG CUGAUGAG X CGAA IUGGUAU
556 UACCACC C CUGUCAC GUGACAG CUGAUGAG X CGAA IGUGGUA
557 ACCACCC C UGUCACA UGUGACA CUGAUGAG X CGAA IGGUGGU
558 CCACCCC U GUCACAG CUGUGAC CUGAUGAG X CGAA IGGGUGG
562 CCCUGUC A CAGGGGC GCCCCUG CUGAUGAG X CGAA IACAGGG
Table 34
564 CUGUCAC A GGGGCCU AGGCCCC CUGAUGAG X CGAA IUGACAG
570 CAGGGGC C UCCCCAG CUGGGGA CUGAUGAG X CGAA ICCCCUG
571 AGGGGCC U CCCCAGG CCUGGGG CUGAUGAG X CGAA IGCCCCU
573 GGGCCUC C CCAGGAG CUCCUGG CUGAUGAG X CGAA IAGGCCC
574 GGCCUCC C CAGGAGG CCUCCUG CUGAUGAG X CGAA IGAGGCC
575 GCCUCCC C AGGAGGC GCCUCCU CUGAUGAG X CGAA IGGAGGC
576 CCUCCCC A GGAGGCC GGCCUCC CUGAUGAG X CGAA IGGGAGG
583 AGGAGGC C UGCGGGA UCCCGCA CUGAUGAG X CGAA ICCUCCU
584 GGAGGCC U GCGGGAG CUCCCGC CUGAUGAG X CGAA IGCCUCC
593 CGGGAGC U GCAGCUU AAGCUGC CUGAUGAG X CGAA ICUCCCG
596 GAGCUGC A GCUUCGA UCGAAGC CUGAUGAG X CGAA ICAGCUC
599 CUGCAGC U UCGAAGC GCUUCGA CUGAUGAG X CGAA ICUGCAG
607 UCGAAGC C UCACAGA UCUGUGA CUGAUGAG X CGAA ICUUCGA
608 CGAAGCC U CACAGAG CUCUGUG CUGAUGAG X CGAA IGCUUCG
610 AAGCCUC A CAGAGAU AUCUCUG CUGAUGAG X CGAA IAGGCUU
612 GCCUCAC A GAGAUCU AGAUCUC CUGAUGAG X CGAA IUGAGGC
619 AGAGAUC U UGAAAGG CCUUUCA CUGAUGAG X CGAA IAUCUCU
634 AGGGGUC U UGAUCCA UGGAUCA CUGAUGAG X CGAA IACCCCU
640 CUUGAUC C AGCGGAA UUCCGCU CUGAUGAG X CGAA IAUCAAG
641 UUGAUCC A GCGGAAC GUUCCGC CUGAUGAG X CGAA IGAUCAA
649 GCGGAAC C CCCAGCU AGCUGGG CUGAUGAG X CGAA IUUCCGC
650 CGGAACC C CCAGCUC GAGCUGG CUGAUGAG X CGAA IGUUCCG
651 GGAACCC C CAGCUCU AGAGCUG CUGAUGAG X CGAA IGGUUCC
652 GAACCCC C AGCUCUG CAGAGCU CUGAUGAG X CGAA IGGGUUC
653 AACCCCC A GCUCUGC GCAGAGC CUGAUGAG X CGAA IGGGGUU
656 CCCCAGC U CUGCUAC GUAGCAG CUGAUGAG X CGAA ICUGGGG
658 CCAGCUC U GCUACCA UGGUAGC CUGAUGAG X CGAA IAGCUGG
661 GCUCUGC U ACCAGGA UCCUGGU CUGAUGAG X CGAA ICAGAGC
664 CUGCUAC C AGGACAC GUGUCCU CUGAUGAG X CGAA IUAGCAG
665 UGCUACC A GGACACG CGUGUCC CUGAUGAG X CGAA IGUAGCA
670 CCAGGAC A CGAUUUU AAAAUCG CUGAUGAG X CGAA IUCCUGG
688 GAAGGAC A UCUUCCA UGGAAGA CUGAUGAG X CGAA IUCCUUC
691 GGACAUC U UCCACAA UUGUGGA CUGAUGAG X CGAA IAUGUCC
694 CAUCUUC C ACAAGAA UUCUUGU CUGAUGAG X CGAA IAAGAUG
695 AUCUUCC A CAAGAAC GUUCUUG CUGAUGAG X CGAA IGAAGAU
697 CUUCCAC A AGAACAA UUGUUCU CUGAUGAG X CGAA IUGGAAG
703 CAAGAAC A ACCAGCU AGCUGGU CUGAUGAG X CGAA IUUCUUG
706 GAACAAC C AGCUGGC GCCAGCU CUGAUGAG X CGAA IUUGUUC
707 AACAACC A GCUGGCU AGCCAGC CUGAUGAG X CGAA IGUUGUU
710 AACCAGC U GGCUCUC GAGAGCC CUGAUGAG X CGAA ICUGGUU
714 AGCUGGC U CUCACAC GUGUGAG CUGAUGAG X CGAA ICCAGCU
716 CUGGCUC U CACACUG CAGUGUG CUGAUGAG X CGAA IAGCCAG
718 GGCUCUC A CACUGAU AUCAGUG CUGAUGAG X CGAA IAGAGCC
720 CUCUCAC A CUGAUAG CUAUCAG CUGAUGAG X CGAA IUGAGAG
722 CUCACAC U GAUAGAC GUCUAUC CUGAUGAG X CGAA IUGUGAG
730 GAUAGAC A CCAACCG CGGUUGG CUGAUGAG X CGAA IUCUAUC
732 UAGACAC C AACCGCU AGCGGUU CUGAUGAG X CGAA IUGUCUA
Table 34
733 AGACACC A ACCGCUC GAGCGGU CUGAUGAG X CGAA IGUGUCU
736 CACCAAC C GCUCUCG CGAGAGC CUGAUGAG X CGAA IUUGGUG
739 CAACCGC U CUCGGGC GCCCGAG CUGAUGAG X CGAA ICGGUUG
741 ACCGCUC U CGGGCCU AGGCCCG CUGAUGAG X CGAA IAGCGGU
747 CUCGGGC C UGCCACC GGUGGCA CUGAUGAG X CGAA ICCCGAG
748 UCGGGCC U GCCACCC GGGUGGC CUGAUGAG X CGAA IGCCCGA
751 GGCCUGC C ACCCCUG CAGGGGU CUGAUGAG X CGAA ICAGGCC
752 GCCUGCC A CCCCUGU ACAGGGG CUGAUGAG X CGAA IGCAGGC
754 CUGCCAC C CCUGUUC GAACAGG CUGAUGAG X CGAA IUGGCAG
755 UGCCACC C CUGUUCU AGAACAG CUGAUGAG X CGAA IGUGGCA
756 GCCACCC C UGUUCUC GAGAACA CUGAUGAG X CGAA IGGUGGC
757 CCACCCC U GUUCUCC GGAGAAC CUGAUGAG X CGAA IGGGUGG
762 CCUGUUC U CCGAUGU ACAUCGG CUGAUGAG X CGAA IAACAGG
764 UGUUCUC C GAUGUGU ACACAUC CUGAUGAG X CGAA IAGAACA
778 UAAGGGC U CCCGCUG CAGCGGG CUGAUGAG X CGAA ICCCUUA
780 AGGGCUC C CGCUGCU AGCAGCG CUGAUGAG X CGAA IAGCCCU
781 GGGCUCC C GCUGCUG CAGCAGC CUGAUGAG X CGAA IGAGCCC
784 CUCCCGC U GCUGGGG CCCCAGC CUGAUGAG X CGAA ICGGGAG
787 CCGCUGC U GGGGAGA UCUCCCC CUGAUGAG X CGAA ICAGCGG
801 AGAGUUC U GAGGAUU AAUCCUC CUGAUGAG X CGAA IAACUCU
812 GAUUGUC A GAGCCUG CAGGCUC CUGAUGAG X CGAA IACAAUC
817 UCAGAGC C UGACGCG CGCGUCA CUGAUGAG X CGAA ICUCUGA
818 CAGAGCC U GACGCGC GCGCGUC CUGAUGAG X CGAA IGCUCUG
826 GACGCGC A CUGUCUG CAGACAG CUGAUGAG X CGAA ICGCGUC
828 CGCGCAC U GUCUGUG CACAGAC CUGAUGAG X CGAA IUGCGCG
832 CACUGUC U GUGCCGG CCGGCAC CUGAUGAG X CGAA IACAGUG
837 UCUGUGC C GGUGGCU AGCCACC CUGAUGAG X CGAA ICACAGA
844 CGGUGGC U GUGCCCG CGGGCAC CUGAUGAG X CGAA ICCACCG
849 GCUGUGC C CGCUGCA UGCAGCG CUGAUGAG X CGAA ICACAGC
850 CUGUGCC C GCUGCAA UUGCAGC CUGAUGAG X CGAA IGCACAG
853 UGCCCGC U GCAAGGG CCCUUGC CUGAUGAG X CGAA ICGGGCA
856 CCGCUGC A AGGGGCC GGCCCCU CUGAUGAG X CGAA ICAGCGG
863 AAGGGGC C ACUGCCC GGGCAGU CUGAUGAG X CGAA ICCCCUU
864 AGGGGCC A CUGCCCA UGGGCAG CUGAUGAG X CGAA IGCCCCU
866 GGGCCAC U GCCCACU AGUGGGC CUGAUGAG X CGAA IUGGCCC
869 CCACUGC C CACUGAC _ GUCAGUG CUGAUGAG X CGAA ICAGUGG
870 ' CACUGCC C ACUGACU AGUCAGU CUGAUGAG X CGAA IGCAGUG
871 ACUGCCC A CUGACUG CAGUCAG CUGAUGAG X CGAA IGGCAGU
873 UGCCCAC U GACUGCU AGCAGUC CUGAUGAG X CGAA IUGGGCA
877 CACUGAC U GCUGCCA UGGCAGC CUGAUGAG X CGAA IUCAGUG
880 UGACUGC U GCCAUGA UCAUGGC CUGAUGAG X CGAA ICAGUCA
883 CUGCUGC C AUGAGCA UGCUCAU CUGAUGAG X CGAA ICAGCAG
884 UGCUGCC A UGAGCAG CUGCUCA CUGAUGAG X CGAA IGCAGCA
890 CAUGAGC A GUGUGCU AGCACAC CUGAUGAG X CGAA ICUCAUG
897 AGUGUGC U GCCGGCU AGCCGGC CUGAUGAG X CGAA ICACACU
900 GUGCUGC C GGCUGCA UGCAGCC CUGAUGAG X CGAA ICAGCAC
904 UGCCGGC U GCACGGG CCCGUGC CUGAUGAG X CGAA ICCGGCA
Table 34
907 CGGCUGC A CGGGCCC GGGCCCG CUGAUGAG X CGAA ICAGCCG
913 CACGGGC C CCAAGCA UGCUUGG CUGAUGAG X CGAA ICCCGUG
914 ACGGGCC C CAAGCAC GUGCUUG CUGAUGAG X CGAA IGCCCGU
915 CGGGCCC C AAGCACU AGUGCUU CUGAUGAG X CGAA IGGCCCG
916 GGGCCCC A AGCACUC GAGUGCU CUGAUGAG X CGAA IGGGCCC
920 CCCAAGC A CUCUGAC GUCAGAG CUGAUGAG X CGAA ICUUGGG
922 CAAGCAC U CUGACUG CAGUCAG CUGAUGAG X CGAA IUGCUUG
924 AGCACUC U GACUGCC GGCAGUC CUGAUGAG X CGAA IAGUGCU
928 CUCUGAC U GCCUGGC GCCAGGC CUGAUGAG X CGAA IUCAGAG
931 UGACUGC C UGGCCUG CAGGCCA CUGAUGAG X CGAA ICAGUCA
932 GACUGCC U GGCCUGC GCAGGCC CUGAUGAG X CGAA IGCAGUC
936 GCCUGGC C UGCCUCC GGAGGCA CUGAUGAG X CGAA ICCAGGC
937 CCUGGCC U GCCUCCA UGGAGGC CUGAUGAG X CGAA IGCCAGG
940 GGCCUGC C UCCACUU AAGUGGA CUGAUGAG X CGAA ICAGGCC
941 GCCUGCC u CCACUUC GAAGUGG CUGAUGAG X CGAA IGCAGGC
943 CUGCCUC c ACUUCAA UUGAAGU CUGAUGAG X CGAA IAGGCAG
944 UGCCUCC A CUUCAAC GUUGAAG CUGAUGAG X CGAA IGAGGCA
946 CCUCCAC U UCAACCA UGGUUGA CUGAUGAG X CGAA IUGGAGG
949 CCACUUC A ACCACAG CUGUGGU CUGAUGAG X CGAA IAAGUGG
952 CUUCAAC C ACAGUGG CCACUGU CUGAUGAG X CGAA IUUGAAG
953 UUCAACC A CAGUGGC GCCACUG CUGAUGAG X CGAA IGUUGAA
955 CAACCAC A GUGGCAU AUGCCAC CUGAUGAG X CGAA IUGGUUG
961 CAGUGGC A UCUGUGA UCACAGA CUGAUGAG X CGAA ICCACUG
964 UGGCAUC U GUGAGCU AGCUCAC CUGAUGAG X CGAA IAUGCCA
971 UGUGAGC U GCACUGC GCAGUGC CUGAUGAG X CGAA ICUCACA
974 GAGCUGC A CUGCCCA UGGGCAG CUGAUGAG X CGAA ICAGCUC
976 GCUGCAC U GCCCAGC GCUGGGC CUGAUGAG X CGAA IUGCAGC
979 GCACUGC C CAGCCCU AGGGCUG CUGAUGAG X CGAA ICAGUGC
980 CACUGCC C AGCCCUG CAGGGCU CUGAUGAG X CGAA IGCAGUG
981 ACUGCCC A GCCCUGG CCAGGGC CUGAUGAG X CGAA IGGCAGU
984 GCCCAGC C CUGGUCA UGACCAG CUGAUGAG X CGAA ICUGGGC
985 CCCAGCC C UGGUCAC GUGACCA CUGAUGAG X CGAA IGCUGGG
986 CCAGCCC U GGUCACC GGUGACC CUGAUGAG X CGAA IGGCUGG
991 CCUGGUC A CCUACAA UUGUAGG CUGAUGAG X CGAA IACCAGG
993 UGGUCAC C UACAACA UGUUGUA CUGAUGAG X CGAA IUGACCA
994 GGUCACC U ACAACAC GUGUUGU CUGAUGAG X CGAA IGUGACC
997 CACCUAC A ACACAGA UCUGUGU CUGAUGAG X CGAA IUAGGUG
1000 CUACAAC A CAGACAC GUGUCUG CUGAUGAG X CGAA IUUGUAG
1002 ACAACAC A GACACGU ACGUGUC CUGAUGAG X CGAA IUGUUGU
1006 CACAGAC A CGUUUGA UCAAACG CUGAUGAG X CGAA IUCUGUG
1017 UUGAGUC C AUGCCCA UGGGCAU CUGAUGAG X CGAA IACUCAA
1018 UGAGUCC A UGCCCAA UUGGGCA CUGAUGAG X CGAA IGACUCA
1022 UCCAUGC C CAAUCCC GGGAUUG CUGAUGAG X CGAA ICAUGGA
1023 CCAUGCC C AAUCCCG CGGGAUU CUGAUGAG X CGAA IGCAUGG
1024 CAUGCCC A AUCCCGA UCGGGAU CUGAUGAG X CGAA IGGCAUG
1028 CCCAAUC C CGAGGGC GCCCUCG CUGAUGAG X CGAA IAUUGGG
1029 CCAAUCC C GAGGGCC GGCCCUC CUGAUGAG X CGAA IGAUUGG
Table 34
1036 CGAGGGC C GGUAUAC GUAUACC CUGAUGAG X CGAA ICCCUCG
1044 GGUAUAC A UUCGGCG CGCCGAA CUGAUGAG X CGAA IUAUACC
1053 UCGGCGC C AGCUGUG CACAGCU CUGAUGAG X CGAA ICGCCGA
1054 CGGCGCC A GCUGUGU ACACAGC CUGAUGAG X CGAA IGCGCCG
1057 CGCCAGC U GUGUGAC GUCACAC CUGAUGAG X CGAA ICUGGCG
1065 GUGUGAC U GCCUGUC GACAGGC CUGAUGAG X CGAA IUCACAC
1068 UGACUGC C UGUCCCU AGGGACA CUGAUGAG X CGAA ICAGUCA
1069 GACUGCC U GUCCCUA UAGGGAC CUGAUGAG X CGAA IGCAGUC
1073 GCCUGUC C CUACAAC GUUGUAG CUGAUGAG X CGAA IACAGGC
1074 CCUGUCC C UACAACU AGUUGUA CUGAUGAG X CGAA IGACAGG
1075 CUGUCCC U ACAACUA UAGUUGU CUGAUGAG X CGAA IGGACAG
1078 UCCCUAC A ACUACCU AGGUAGU CUGAUGAG X CGAA IUAGGGA
1081 CUACAAC U ACCUUUC GAAAGGU CUGAUGAG X CGAA IUUGUAG
1084 CAACUAC C UUUCUAC GUAGAAA CUGAUGAG X CGAA IUAGUUG
1085 AACUACC U UUCUACG CGUAGAA CUGAUGAG X CGAA IGUAGUU
1089 ACCUUUC U ACGGACG CGUCCGU CUGAUGAG X CGAA IAAAGGU
1104 UGGGAUC C UGCACCC GGGUGCA CUGAUGAG X CGAA IAUCCCA
1105 GGGAUCC U GCACCCU AGGGUGC CUGAUGAG X CGAA IGAUCCC
1108 AUCCUGC A CCCUCGU ACGAGGG CUGAUGAG X CGAA ICAGGAU
1110 CCUGCAC C CUCGUCU AGACGAG CUGAUGAG X CGAA IUGCAGG llll CUGCACC C UCGUCUG CAGACGA CUGAUGAG X CGAA IGUGCAG
1112 UGCACCC U CGUCUGC GCAGACG CUGAUGAG X CGAA IGGUGCA
1117 CCUCGUC U GCCCCCU AGGGGGC CUGAUGAG X CGAA IACGAGG
1120 CGUCUGC C CCCUGCA UGCAGGG CUGAUGAG X CGAA ICAGACG
1121 GUCUGCC C CCUGCAC GUGCAGG CUGAUGAG X CGAA IGCAGAC
1122 UCUGCCC C CUGCACA UGUGCAG CUGAUGAG X CGAA IGGCAGA
1123 CUGCCCC C UGCACAA UUGUGCA CUGAUGAG X CGAA IGGGCAG
1124 UGCCCCC U GCACAAC GUUGUGC CUGAUGAG X CGAA IGGGGCA
1127 CCCCUGC A CAACCAA UUGGUUG CUGAUGAG X CGAA ICAGGGG
1129 CCUGCAC A ACCAAGA UCUUGGU CUGAUGAG X CGAA IUGCAGG
1132 GCACAAC C AAGAGGU ACCUCUU CUGAUGAG X CGAA IUUGUGC
1133 CACAACC A AGAGGUG CACCUCU CUGAUGAG X CGAA IGUUGUG
1143 AGGUGAC A GCAGAGG CCUCUGC CUGAUGAG X CGAA IUCACCU
1146 UGACAGC A GAGGAUG CAUCCUC CUGAUGAG X CGAA ICUGUCA
1158 AUGGAAC A CAGCGGU ACCGCUG CUGAUGAG X CGAA IUUCCAU
1160 GGAACAC A GCGGUGU ACACCGC CUGAUGAG X CGAA IUGUUCC
1177 GAAGUGC A GCAAGCC GGCUUGC CUGAUGAG X CGAA ICACUUC
1180 GUGCAGC A AGCCCUG CAGGGCU CUGAUGAG X CGAA ICUGCAC
1184 AGCAAGC C CUGUGCC GGCACAG CUGAUGAG X CGAA ICUUGCU
1185 GCAAGCC C UGUGCCC GGGCACA CUGAUGAG X CGAA IGCUUGC
1186 CAAGCCC U GUGCCCG CGGGCAC CUGAUGAG X CGAA IGGCUUG
1191 CCUGUGC c CGAGUGU ACACUCG CUGAUGAG X CGAA ICACAGG
1192 CUGUGCC c GAGUGUG CACACUC CUGAUGAG X CGAA IGCACAG
1201 AGUGUGC u AUGGUCU AGACCAU CUGAUGAG X CGAA ICACACU
1208 UAUGGUC u GGGCAUG CAUGCCC CUGAUGAG X CGAA IACCAUA
1213 UCUGGGC A UGGAGCA UGCUCCA CUGAUGAG X CGAA ICCCAGA
1220 AUGGAGC A CUUGCGA UCGCAAG CUGAUGAG X CGAA ICUCCAU
Table 34
1222 GGAGCAC U UGCGAGA UCUCGCA CUGAUGAG X CGAA IUGCUCC
1239 UGAGGGC A GUUACCA UGGUAAC CUGAUGAG X CGAA ICCCUCA
1245 CAGUUAC C AGUGCCA UGGCACU CUGAUGAG X CGAA IUAACUG
1246 AGUUACC A GUGCCAA UUGGCAC CUGAUGAG X CGAA IGUAACU
1251 CCAGUGC C AAUAUCC GGAUAUU CUGAUGAG X CGAA ICACUGG
1252 CAGUGCC A AUAUCCA UGGAUAU CUGAUGAG X CGAA IGCACUG
1258 CAAUAUC C AGGAGUU AACUCCU CUGAUGAG X CGAA IAUAUUG
1259 AAUAUCC A GGAGUUU AAACUCC CUGAUGAG X CGAA IGAUAUU
1269 AGUUUGC U GGCUGCA UGCAGCC CUGAUGAG X CGAA ICAAACU
1273 UGCUGGC U GCAAGAA UUCUUGC CUGAUGAG X CGAA ICCAGCA
1276 UGGCUGC A AGAAGAU AUCUUCU CUGAUGAG X CGAA ICAGCCA
1285 GAAGAUC U UUGGGAG CUCCCAA CUGAUGAG X CGAA IAUCUUC
1294 UGGGAGC C UGGCAUU AAUGCCA CUGAUGAG X CGAA ICUCCCA
1295 GGGAGCC U GGCAUUU AAAUGCC CUGAUGAG X CGAA IGCUCCC
1299 GCCUGGC A UUUCUGC GCAGAAA CUGAUGAG X CGAA ICCAGGC
1304 GCAUUUC U GCCGGAG CUCCGGC CUGAUGAG X CGAA IAAAUGC
1307 UUUCUGC C GGAGAGC GCUCUCC CUGAUGAG X CGAA ICAGAAA
1315 GGAGAGC U UUGAUGG CCAUCAA CUGAUGAG X CGAA ICUCUCC
1327 UGGGGAC C CAGCCUC GAGGCUG CUGAUGAG X CGAA IUCCCCA
1328 GGGGACC C AGCCUCC •GGAGGCU CUGAUGAG X CGAA IGUCCCC
1329 GGGACCC A GCCUCCA UGGAGGC CUGAUGAG X CGAA IGGUCCC
1332 ACCCAGC C UCCAACA UGUUGGA CUGAUGAG X CGAA ICUGGGU
1333 CCCAGCC U CCAACAC GUGUUGG CUGAUGAG X CGAA IGCUGGG
1335 CAGCCUC C AACACUG CAGUGUU CUGAUGAG X CGAA IAGGCUG
1336 AGCCUCC A ACACUGC GCAGUGU CUGAUGAG X CGAA IGAGGCU
1339 CUCCAAC A CUGCCCC GGGGCAG CUGAUGAG X CGAA IUUGGAG
1341 CCAACAC U GCCCCGC GCGGGGC CUGAUGAG X CGAA IUGUUGG
1344 ACACUGC C CCGCUCC GGAGCGG CUGAUGAG X CGAA ICAGUGU
1345 CACUGCC C CGCUCCA UGGAGCG CUGAUGAG X CGAA IGCAGUG
1346 ACUGCCC C GCUCCAG CUGGAGC CUGAUGAG X CGAA IGGCAGU
1349 GCCCCGC U CCAGCCA UGGCUGG CUGAUGAG X CGAA ICGGGGC
1351 CCCGCUC C AGCCAGA UCUGGCU CUGAUGAG X CGAA IAGCGGG
1352 CCGCUCC A GCCAGAG CUCUGGC CUGAUGAG X CGAA IGAGCGG
1355 CUCCAGC C AGAGCAG CUGCUCU CUGAUGAG X CGAA ICUGGAG
1356 UCCAGCC A GAGCAGC GCUGCUC CUGAUGAG X CGAA IGCUGGA
1361 CCAGAGC A GCUCCAA UUGGAGC CUGAUGAG X CGAA ICUCUGG
1364 GAGCAGC U CCAAGUG CACUUGG CUGAUGAG X CGAA ICUGCUC
1366 GCAGCUC C AAGUGUU AACACUU CUGAUGAG X CGAA IAGCUGC
1367 CAGCUCC A AGUGUUU AAACACU CUGAUGAG X CGAA IGAGCUG
1380 UUGAGAC U CUGGAAG CUUCCAG CUGAUGAG X CGAA IUCUCAA
1382 GAGACUC U GGAAGAG CUCUUCC CUGAUGAG X CGAA IAGUCUC
1393 AGAGAUC A CAGGUUA UAACCUG CUGAUGAG X CGAA IAUCUCU
1395 AGAUCAC A GGUUACC GGUAACC CUGAUGAG X CGAA IUGAUCU
1402 AGGUUAC C UAUACAU AUGUAUA CUGAUGAG X CGAA IUAACCU
1403 GGUUACC U AUACAUC GAUGUAU CUGAUGAG X CGAA IGUAACC
1408 CCUAUAC A UCUCAGC GCUGAGA CUGAUGAG X CGAA IUAUAGG
1411 AUACAUC U CAGCAUG CAUGCUG CUGAUGAG X CGAA IAUGUAU
Table 34
1413 ACAUCUC A GCAUGGC GCCAUGC CUGAUGAG X CGAA IAGAUGU
1416 UCUCAGC A UGGCCGG CCGGCCA CUGAUGAG X CGAA ICUGAGA
1421 GCAUGGC C GGACAGC GCUGUCC CUGAUGAG X CGAA ICCAUGC
1426 GCCGGAC A GCCUGCC GGCAGGC CUGAUGAG X CGAA IUCCGGC
1429 GGACAGC C UGCCUGA UCAGGCA CUGAUGAG X CGAA ICUGUCC
1430 GACAGCC U GCCUGAC GUCAGGC CUGAUGAG X CGAA IGCUGUC
1433 AGCCUGC C UGACCUC GAGGUCA CUGAUGAG X CGAA ICAGGCU
1434 GCCUGCC U GACCUCA UGAGGUC CUGAUGAG X CGAA IGCAGGC
1438 GCCUGAC C UCAGCGU ACGCUGA CUGAUGAG X CGAA IUCAGGC
1439 CCUGACC U CAGCGUC GACGCUG CUGAUGAG X CGAA IGUCAGG
1441 UGACCUC A GCGUCUU AAGACGC CUGAUGAG X CGAA IAGGUCA
1447 CAGCGUC U UCCAGAA UUCUGGA CUGAUGAG X CGAA IACGCUG
1450 CGUCUUC C AGAACCU AGGUUCU CUGAUGAG X CGAA IAAGACG
1451 GUCUUCC A GAACCUG CAGGUUC CUGAUGAG X CGAA IGAAGAC
1456 CCAGAAC C UGCAAGU ACUUGCA CUGAUGAG X CGAA IUUCUGG
1457 CAGAACC U GCAAGUA UACUUGC CUGAUGAG X CGAA IGUUCUG
1460 AACCUGC A AGUAAUC GAUUACU CUGAUGAG X CGAA ICAGGUU
1468 AGUAAUC C GGGGACG CGUCCCC CUGAUGAG X CGAA IAUUACU
1481 CGAAUUC U GCACAAU AUUGUGC CUGAUGAG X CGAA IAAUUCG
1484 AUUCUGC A CAAUGGC GCCAUUG CUGAUGAG X CGAA ICAGAAU
1486 UCUGCAC A AUGGCGC GCGCCAU CUGAUGAG X CGAA IUGCAGA
1494 AUGGCGC C UACUCGC GCGAGUA CUGAUGAG X CGAA ICGCCAU
1495 UGGCGCC U ACUCGCU AGCGAGU CUGAUGAG X CGAA IGCGCCA
1498 CGCCUAC U CGCUGAC GUCAGCG CUGAUGAG X CGAA IUAGGCG
1502 UACUCGC U GACCCUG CAGGGUC CUGAUGAG X CGAA ICGAGUA
1506 CGCUGAC C CUGCAAG CUUGCAG CUGAUGAG X CGAA IUCAGCG
1507 GCUGACC C UGCAAGG CCUUGCA CUGAUGAG X CGAA IGUCAGC
1508 CUGACCC U GCAAGGG CCCUUGC CUGAUGAG X CGAA IGGUCAG
1511 ACCCUGC A AGGGCUG CAGCCCU CUGAUGAG X CGAA ICAGGGU
1517 CAAGGGC U GGGCAUC GAUGCCC CUGAUGAG X CGAA ICCCUUG
1522 GCUGGGC A UCAGCUG CAGCUGA CUGAUGAG X CGAA ICCCAGC
1525 GGGCAUC A GCUGGCU AGCCAGC CUGAUGAG X CGAA IAUGCCC
1528 CAUCAGC U GGCUGGG CCCAGCC CUGAUGAG X CGAA ICUGAUG
1532 AGCUGGC U GGGGCUG CAGCCCC CUGAUGAG X CGAA ICCAGCU
1538 CUGGGGC U GCGCUCA UGAGCGC CUGAUGAG X CGAA ICCCCAG
1543 GCUGCGC U CACUGAG CUCAGUG CUGAUGAG X CGAA ICGCAGC
1545 UGCGCUC A CUGAGGG CCCUCAG CUGAUGAG X CGAA IAGCGCA
1547 CGCUCAC U GAGGGAA UUCCCUC CUGAUGAG X CGAA IUGAGCG
1556 AGGGAAC U GGGCAGU ACUGCCC CUGAUGAG X CGAA IUUCCCU
1561 ACUGGGC A GUGGACU AGUCCAC CUGAUGAG X CGAA ICCCAGU
1568 AGUGGAC U GGCCCUC GAGGGCC CUGAUGAG X CGAA IUCCACU
1572 GACUGGC C CUCAUCC GGAUGAG CUGAUGAG X CGAA ICCAGUC
1573 ACUGGCC C UCAUCCA UGGAUGA CUGAUGAG X CGAA IGCCAGU
1574 CUGGCCC U CAUCCAC GUGGAUG CUGAUGAG X CGAA IGGCCAG
1576 GGCCCUC A UCCACCA UGGUGGA CUGAUGAG X CGAA IAGGGCC
1579 CCUCAUC C ACCAUAA UUAUGGU CUGAUGAG X CGAA IAUGAGG
1580 CUCAUCC A CCAUAAC GUUAUGG CUGAUGAG X CGAA IGAUGAG
Table 34
1582 CAUCCAC C AUAACAC GUGUUAU CUGAUGAG X CGAA IUGGAUG
1583 AUCCACC A UAACACC GGUGUUA CUGAUGAG X CGAA IGUGGAU
1588 CCAUAAC A CCCACCU AGGUGGG CUGAUGAG X CGAA IUUAUGG
1590 AUAACAC C CACCUCU AGAGGUG CUGAUGAG X CGAA IUGUUAU
1591 UAACACC C ACCUCUG CAGAGGU CUGAUGAG X CGAA IGUGUUA
1592 AACACCC A CCUCUGC GCAGAGG CUGAUGAG X CGAA IGGUGUU
1594 CACCCAC C UCUGCUU AAGCAGA CUGAUGAG X CGAA IUGGGUG
1595 ACCCACC U CUGCUUC GAAGCAG CUGAUGAG X CGAA IGUGGGU
1597 CCACCUC u GCUUCGU ACGAAGC CUGAUGAG X CGAA IAGGUGG
1600 CCUCUGC u UCGUGCA UGCACGA CUGAUGAG X CGAA ICAGAGG
1607 UUCGUGC A CACGGUG CACCGUG CUGAUGAG X CGAA ICACGAA
1609 CGUGCAC A CGGUGCC GGCACCG CUGAUGAG X CGAA IUGCACG
1616 ACGGUGC C CUGGGAC GUCCCAG CUGAUGAG X CGAA ICACCGU
1617 CGGUGCC C UGGGACC GGUCCCA CUGAUGAG X CGAA IGCACCG
1618 GGUGCCC U GGGACCA UGGUCCC CUGAUGAG X CGAA IGGCACC
1624 CUGGGAC C AGCUCUU AAGAGCU CUGAUGAG X CGAA IUCCCAG
1625 UGGGACC A GCUCUUU AAAGAGC CUGAUGAG X CGAA IGUCCCA
1628 GACCAGC U CUUUCGG CCGAAAG CUGAUGAG X CGAA ICUGGUC
1630 CCAGCUC U UUCGGAA UUCCGAA CUGAUGAG X CGAA IAGCUGG
1639 UCGGAAC C CGCACCA UGGUGCG CUGAUGAG X CGAA IUUCCGA
1640 CGGAACC C GCACCAA UUGGUGC CUGAUGAG X CGAA IGUUCCG
1643 AACCCGC A CCAAGCU AGCUUGG CUGAUGAG X CGAA ICGGGUU
1645 CCCGCAC C AAGCUCU AGAGCUU CUGAUGAG X CGAA IUGCGGG
1646 CCGCACC A AGCUCUG CAGAGCU CUGAUGAG X CGAA IGUGCGG
1650 ACCAAGC U CUGCUCC GGAGCAG CUGAUGAG X CGAA ICUUGGU
1652 CAAGCUC U GCUCCAC GUGGAGC CUGAUGAG X CGAA IAGCUUG
1655 GCUCUGC U CCACACU AGUGUGG CUGAUGAG X CGAA ICAGAGC
1657 UCUGCUC C ACACUGC GCAGUGU CUGAUGAG X CGAA IAGCAGA
1658 CUGCUCC A CACUGCC GGCAGUG CUGAUGAG X CGAA IGAGCAG
1660 GCUCCAC A CUGCCAA UUGGCAG CUGAUGAG X CGAA IUGGAGC
1662 UCCACAC U GCCAACC GGUUGGC CUGAUGAG X CGAA IUGUGGA
1665 ACACUGC C AACCGGC GCCGGUU CUGAUGAG X CGAA ICAGUGU
1666 CACUGCC A ACCGGCC GGCCGGU CUGAUGAG X CGAA IGCAGUG
1669 UGCCAAC C GGCCAGA UCUGGCC CUGAUGAG X CGAA IUUGGCA
1673 AACCGGC C AGAGGAC GUCCUCU CUGAUGAG X CGAA ICCGGUU
1674 ACCGGCC A GAGGACG CGUCCUC CUGAUGAG X CGAA IGCCGGU
1699 CGAGGGC C UGGCCUG CAGGCCA CUGAUGAG X CGAA ICCCUCG
1700 GAGGGCC U GGCCUGC GCAGGCC CUGAUGAG X CGAA IGCCCUC
1704 GCCUGGC C UGCCACC GGUGGCA CUGAUGAG X CGAA ICCAGGC
1705 CCUGGCC U GCCACCA UGGUGGC CUGAUGAG X CGAA IGCCAGG
1708 GGCCUGC C ACCAGCU AGCUGGU CUGAUGAG X CGAA ICAGGCC
1709 GCCUGCC A CCAGCUG CAGCUGG CUGAUGAG X CGAA IGCAGGC
1711 CUGCCAC C AGCUGUG CACAGCU CUGAUGAG X CGAA IUGGCAG
1712 UGCCACC A GCUGUGC GCACAGC CUGAUGAG X CGAA IGUGGCA
1715 CACCAGC U GUGCGCC GGCGCAC CUGAUGAG X CGAA ICUGGUG
1722 UGUGCGC C CGAGGGC GCCCUCG CUGAUGAG X CGAA ICGCACA
1723 GUGCGCC c GAGGGCA UGCCCUC CUGAUGAG X CGAA IGCGCAC
Table 34
1730 CGAGGGC A CUGCUGG CCAGCAG CUGAUGAG X CGAA ICCCUCG
1732 AGGGCAC U GCUGGGG CCCCAGC CUGAUGAG X CGAA IUGCCCU
1735 GCACUGC U GGGGUCC GGACCCC CUGAUGAG X CGAA ICAGUGC
1742 UGGGGUC C AGGGCCC GGGCCCU CUGAUGAG X CGAA IACCCCA
1743 GGGGUCC A GGGCCCA UGGGCCC CUGAUGAG X CGAA IGACCCC
1748 CCAGGGC C CACCCAG CUGGGUG CUGAUGAG X CGAA ICCCUGG
1749 CAGGGCC C ACCCAGU ACUGGGU CUGAUGAG X CGAA IGCCCUG
1750 AGGGCCC A CCCAGUG CACUGGG CUGAUGAG X CGAA IGGCCCU
1752 GGCCCAC C CAGUGUG CACACUG CUGAUGAG X CGAA IUGGGCC
1753 GCCCACC C AGUGUGU ACACACU CUGAUGAG X CGAA IGUGGGC
1754 CCCACCC A GUGUGUC GACACAC CUGAUGAG X CGAA IGGUGGG
1762 GUGUGUC A ACUGCAG CUGCAGU CUGAUGAG X CGAA IACACAC
1765 UGUCAAC U GCAGCCA UGGCUGC CUGAUGAG X CGAA IUUGACA
1768 CAACUGC A GCCAGUU AACUGGC CUGAUGAG X CGAA ICAGUUG
1771 CUGCAGC C AGUUCCU AGGAACU CUGAUGAG X CGAA ICUGCAG
1772 UGCAGCC A GUUCCUU AAGGAAC CUGAUGAG X CGAA IGCUGCA
1777 CCAGUUC C UUCGGGG CCCCGAA CUGAUGAG X CGAA IAACUGG
1778 CAGUUCC U UCGGGGC GCCCCGA CUGAUGAG X CGAA IGAACUG
1786 UCGGGGC C AGGAGUG CACUCCU CUGAUGAG X CGAA ICCCCGA
1787 CGGGGCC A GGAGUGC GCACUCC CUGAUGAG X CGAA IGCCCCG
1807 GGAAUGC C GAGUACU AGUACUC CUGAUGAG X CGAA ICAUUCC
1814 CGAGUAC U GCAGGGG CCCCUGC CUGAUGAG X CGAA IUACUCG
1817 GUACUGC A GGGGCUC GAGCCCC CUGAUGAG X CGAA ICAGUAC
1823 CAGGGGC U CCCCAGG CCUGGGG CUGAUGAG X CGAA ICCCCUG
1825 GGGGCUC C CCAGGGA UCCCUGG CUGAUGAG X CGAA IAGCCCC
1826 GGGCUCC C CAGGGAG CUCCCUG CUGAUGAG X CGAA IGAGCCC
1827 GGCUCCC C AGGGAGU ACUCCCU CUGAUGAG X CGAA IGGAGCC
1828 GCUCCCC A GGGAGUA UACUCCC CUGAUGAG X CGAA IGGGAGC
1845 UGAAUGC C AGGCACU AGUGCCU CUGAUGAG X CGAA ICAUUCA
1846 GAAUGCC A GGCACUG CAGUGCC CUGAUGAG X CGAA IGCAUUC
1850 GCCAGGC A CUGUUUG CAAACAG CUGAUGAG X CGAA ICCUGGC
1852 CAGGCAC U GUUUGCC GGCAAAC CUGAUGAG X CGAA IUGCCUG
1859 UGUUUGC C GUGCCAC GUGGCAC CUGAUGAG X CGAA ICAAACA
1864 GCCGUGC C ACCCUGA UCAGGGU CUGAUGAG X CGAA ICACGGC
1865 CCGUGCC A CCCUGAG CUCAGGG CUGAUGAG X CGAA IGCACGG
1867 GUGCCAC C CUGAGUG CACUCAG CUGAUGAG X CGAA IUGGCAC
1868 UGCCACC C UGAGUGU ACACUCA CUGAUGAG X CGAA IGUGGCA
1869 GCCACCC U GAGUGUC GACACUC CUGAUGAG X CGAA IGGUGGC
1877 GAGUGUC A GCCCCAG CUGGGGC CUGAUGAG X CGAA IACACUC
1880 UGUCAGC C CCAGAAU AUUCUGG CUGAUGAG X CGAA ICUGACA
1881 GUCAGCC C CAGAAUG CAUUCUG CUGAUGAG X CGAA IGCUGAC
1882 UCAGCCC C AGAAUGG CCAUUCU CUGAUGAG X CGAA IGGCUGA
1883 CAGCCCC A GAAUGGC GCCAUUC CUGAUGAG X CGAA IGGGCUG
1891 GAAUGGC U CAGUGAC GUCACUG CUGAUGAG X CGAA ICCAUUC
1893 AUGGCUC A GUGACCU AGGUCAC CUGAUGAG X CGAA IAGCCAU
1899 CAGUGAC C UGUUUUG CAAAACA CUGAUGAG X CGAA IUCACUG
1900 AGUGACC U GUUUUGG CCAAAAC CUGAUGAG X CGAA IGUCACU
Table 34
1910 UUUGGAC C GGAGGCU AGCCUCC CUGAUGAG X CGAA IUCCAAA
1917 CGGAGGC U GACCAGU ACUGGUC CUGAUGAG X CGAA ICCUCCG
1921 GGCUGAC C AGUGUGU ACACACU CUGAUGAG X CGAA IUCAGCC
1922 GCUGACC A GUGUGUG CACACAC CUGAUGAG X CGAA IGUCAGC
1932 GUGUGGC C UGUGCCC GGGCACA CUGAUGAG X CGAA ICCACAC
1933 UGUGGCC U GUGCCCA UGGGCAC CUGAUGAG X CGAA IGCCACA
1938 CCUGUGC C CACUAUA UAUAGUG CUGAUGAG X CGAA ICACAGG
1939 CUGUGCC C ACUAUAA UUAUAGU CUGAUGAG X CGAA IGCACAG
1940 UGUGCCC A CUAUAAG CUUAUAG CUGAUGAG X CGAA IGGCACA
1942 UGCCCAC U AUAAGGA UCCUUAU CUGAUGAG X CGAA IUGGGCA
1951 UAAGGAC C CUCCCUU AAGGGAG CUGAUGAG X CGAA IUCCUUA
1952 AAGGACC C UCCCUUC GAAGGGA CUGAUGAG X CGAA IGUCCUU
1953 AGGACCC U CCCUUCU AGAAGGG CUGAUGAG X CGAA IGGUCCU
1955 GACCCUC C CUUCUGC GCAGAAG CUGAUGAG X CGAA IAGGGUC
1956 ACCCUCC C UUCUGCG CGCAGAA CUGAUGAG X CGAA IGAGGGU
1957 CCCUCCC U UCUGCGU ACGCAGA CUGAUGAG X CGAA IGGAGGG
1960 UCCCUUC U GCGUGGC GCCACGC CUGAUGAG X CGAA IAAGGGA
1968 GCGUGGC C CGCUGCC GGCAGCG CUGAUGAG X CGAA ICCACGC
1969 CGUGGCC c GCUGCCC GGGCAGC CUGAUGAG X CGAA IGCCACG
1972 GGCCCGC u GCCCCAG CUGGGGC CUGAUGAG X CGAA ICGGGCC
1975 CCGCUGC c CCAGCGG CCGCUGG CUGAUGAG X CGAA ICAGCGG
1976 CGCUGCC c CAGCGGU ACCGCUG CUGAUGAG X CGAA IGCAGCG
1977 GCUGCCC c AGCGGUG CACCGCU CUGAUGAG X CGAA IGGCAGC
1978 CUGCCCC A GCGGUGU ACACCGC CUGAUGAG X CGAA IGGGCAG
1991 GUGAAAC C UGACCUC GAGGUCA CUGAUGAG X CGAA IUUUCAC
1992 UGAAACC U GACCUCU AGAGGUC CUGAUGAG X CGAA IGUUUCA
1996 ACCUGAC C UCUCCUA UAGGAGA CUGAUGAG X CGAA IUCAGGU
1997 CCUGACC u CUCCUAC GUAGGAG CUGAUGAG X CGAA IGUCAGG
1999 UGACCUC u CCUACAU AUGUAGG CUGAUGAG X CGAA IAGGUCA
2001 ACCUCUC c UACAUGC GCAUGUA CUGAUGAG X CGAA IAGAGGU
2002 CCUCUCC u ACAUGCC GGCAUGU CUGAUGAG X CGAA IGAGAGG
2005 CUCCUAC A UGCCCAU AUGGGCA CUGAUGAG X CGAA IUAGGAG
2009 UACAUGC C CAUCUGG CCAGAUG CUGAUGAG X CGAA ICAUGUA
2010 ACAUGCC C AUCUGGA UCCAGAU CUGAUGAG X CGAA IGCAUGU
2011 CAUGCCC A UCUGGAA UUCCAGA CUGAUGAG X CGAA IGGCAUG
2014 GCCCAUC U GGAAGUU AACUUCC CUGAUGAG X CGAA IAUGGGC
2024 AAGUUUC C AGAUGAG CUCAUCU CUGAUGAG X CGAA IAAACUU
2025 AGUUUCC A GAUGAGG CCUCAUC CUGAUGAG X CGAA IGAAACU
2040 AGGGCGC A UGCCAGC GCUGGCA CUGAUGAG X CGAA ICGCCCU
2044 CGCAUGC C AGCCUUG CAAGGCU CUGAUGAG X CGAA ICAUGCG
2045 GCAUGCC A GCCUUGC GCAAGGC CUGAUGAG X CGAA IGCAUGC
2048 UGCCAGC C UUGCCCC GGGGCAA CUGAUGAG X CGAA ICUGGCA
2049 GCCAGCC U UGCCCCA UGGGGCA CUGAUGAG X CGAA IGCUGGC
2053 GCCUUGC C CCAUCAA UUGAUGG CUGAUGAG X CGAA ICAAGGC
2054 CCUUGCC C CAUCAAC GUUGAUG CUGAUGAG X CGAA IGCAAGG
2055 CUUGCCC C AUCAACU AGUUGAU CUGAUGAG X CGAA IGGCAAG
2056 UUGCCCC A UCAACUG CAGUUGA CUGAUGAG X CGAA IGGGCAA
Table 34
2059 CCCCAUC A ACUGCAC GUGCAGU CUGAUGAG X CGAA IAUGGGG
2062 CAUCAAC U GCACCCA UGGGUGC CUGAUGAG X CGAA IUUGAUG
2065 CAACUGC A CCCACUC GAGUGGG CUGAUGAG X CGAA ICAGUUG
2067 ACUGCAC C CACUCCU AGGAGUG CUGAUGAG X CGAA IUGCAGU
2068 CUGCACC C ACUCCUG CAGGAGU CUGAUGAG X CGAA IGUGCAG
2069 UGCACCC A CUCCUGU ACAGGAG CUGAUGAG X CGAA IGGUGCA
2071 CACCCAC U CCUGUGU ACACAGG CUGAUGAG X CGAA IUGGGUG
2073 ' CCCACUC C UGUGUGG CCACACA CUGAUGAG X CGAA IAGUGGG
2074 CCACUCC U GUGUGGA UCCACAC CUGAUGAG X CGAA IGAGUGG
2083 UGUGGAC C UGGAUGA UCAUCCA CUGAUGAG X CGAA IUCCACA
2084 GUGGACC U GGAUGAC GUCAUCC CUGAUGAG X CGAA IGUCCAC
2092 GGAUGAC A AGGGCUG CAGCCCU CUGAUGAG X CGAA IUCAUCC
2098 CAAGGGC U GCCCCGC GCGGGGC CUGAUGAG X CGAA ICCCUUG
2101 GGGCUGC C CCGCCGA UCGGCGG CUGAUGAG X CGAA ICAGCCC
2102 GGCUGCC C CGCCGAG CUCGGCG CUGAUGAG X CGAA IGCAGCC
2103 GCUGCCC C GCCGAGC GCUCGGC CUGAUGAG X CGAA IGGCAGC
2106 GCCCCGC C GAGCAGA UCUGCUC CUGAUGAG X CGAA ICGGGGC
2111 GCCGAGC A GAGAGCC GGCUCUC CUGAUGAG X CGAA ICUCGGC
2118 AGAGAGC C AGCCCUC GAGGGCU CUGAUGAG X CGAA ICUCUCU
2119 GAGAGCC A GCCCUCU AGAGGGC CUGAUGAG X CGAA IGCUCUC
2122 AGCCAGC C CUCUGAC GUCAGAG CUGAUGAG X CGAA ICUGGCU
2123 GCCAGCC C UCUGACG CGUCAGA CUGAUGAG X CGAA IGCUGGC
2124 CCAGCCC u CUGACGU ACGUCAG CUGAUGAG X CGAA IGGCUGG
2126 AGCCCUC u GACGUCC GGACGUC CUGAUGAG X CGAA IAGGGCU
2133 UGACGUC c AUCAUCU AGAUGAU CUGAUGAG X CGAA IACGUCA
2134 GACGUCC A UCAUCUC GAGAUGA CUGAUGAG X CGAA IGACGUC
2137 GUCCAUC A UCUCUGC GCAGAGA CUGAUGAG X CGAA IAUGGAC
2140 CAUCAUC U CUGCGGU ACCGCAG CUGAUGAG X CGAA IAUGAUG
2142 UCAUCUC U GCGGUGG CCACCGC CUGAUGAG X CGAA IAGAUGA
2155 GGUUGGC A UUCUGCU AGCAGAA CUGAUGAG X CGAA ICCAACC
2159 GGCAUUC U GCUGGUC GACCAGC CUGAUGAG X CGAA IAAUGCC
2162 AUUCUGC U GGUCGUG CACGACC CUGAUGAG X CGAA ICAGAAU
2173 CGUGGUC U UGGGGGU ACCCCCA CUGAUGAG X CGAA IACCACG
2185 GGUGGUC U UUGGGAU AUCCCAA CUGAUGAG X CGAA IACCACC
2194 UGGGAUC C UCAUCAA UUGAUGA CUGAUGAG X CGAA IAUCCCA
2195 GGGAUCC U CAUCAAG CUUGAUG CUGAUGAG X CGAA IGAUCCC
2197 GAUCCUC A UCAAGCG CGCUUGA CUGAUGAG X CGAA IAGGAUC
2200 CCUCAUC A AGCGACG CGUCGCU CUGAUGAG X CGAA IAUGAGG
2210 CGACGGC A GCAGAAG CUUCUGC CUGAUGAG X CGAA ICCGUCG
2213 CGGCAGC A GAAGAUC GAUCUUC CUGAUGAG X CGAA ICUGCCG
2221 GAAGAUC C GGAAGUA UACUUCC CUGAUGAG X CGAA IAUCUUC
2230 GAAGUAC A CGAUGCG CGCAUCG CUGAUGAG X CGAA IUACUUC
2243 CGGAGAC U GCUGCAG CUGCAGC CUGAUGAG X CGAA IUCUCCG
2246 AGACUGC U GCAGGAA UUCCUGC CUGAUGAG X CGAA ICAGUCU
2249 CUGCUGC A GGAAACG CGUUUCC CUGAUGAG X CGAA ICAGCAG
2261 ACGGAGC U GGUGGAG CUCCACC CUGAUGAG X CGAA ICUCCGU
2270 GUGGAGC C GCUGACA UGUCAGC CUGAUGAG X CGAA ICUCCAC
Table .34
2273 GAGCCGC U GACACCU AGGUGUC CUGAUGAG X CGAA ICGGCUC
2277 CGCUGAC A CCUAGCG CGCUAGG CUGAUGAG X CGAA IUCAGCG
2279 CUGACAC C UAGCGGA UCCGCUA CUGAUGAG X CGAA IUGUCAG
2280 UGACACC U AGCGGAG CUCCGCU CUGAUGAG X CGAA IGUGUCA
2294 GCGAUGC C CAACCAG CUGGUUG CUGAUGAG X CGAA ICAUCGC
2295 CGAUGCC C AACCAGG CCUGGUU CUGAUGAG X CGAA IGCAUCG
2296 GAUGCCC A ACCAGGC GCCUGGU CUGAUGAG X CGAA IGGCAUC
2299 GCCCAAC C AGGCGCA UGCGCCU CUGAUGAG X CGAA IUUGGGC
2300 CCCAACC A GGCGCAG CUGCGCC CUGAUGAG X CGAA IGUUGGG
2306 CAGGCGC A GAUGCGG CCGCAUC CUGAUGAG X CGAA ICGCCUG
2317 GCGGAUC C UGAAAGA UCUUUCA CUGAUGAG X CGAA IAUCCGC
2318 CGGAUCC U GAAAGAG CUCUUUC CUGAUGAG X CGAA IGAUCCG
2333 ACGGAGC U GAGGAAG CUUCCUC CUGAUGAG X CGAA ICUCCGU
2351 AAGGUGC U UGGAUCU AGAUCCA CUGAUGAG X CGAA ICACCUU
2358 UUGGAUC U GGCGCUU AAGCGCC CUGAUGAG X CGAA IAUCCAA
2364 CUGGCGC U UUUGGCA UGCCAAA CUGAUGAG X CGAA ICGCCAG
2371 UUUUGGC A CAGUCUA UAGACUG CUGAUGAG X CGAA ICCAAAA
2373 UUGGCAC A GUCUACA UGUAGAC CUGAUGAG X CGAA IUGCCAA
2377 CACAGUC U ACAAGGG CCCUUGU CUGAUGAG X CGAA IACUGUG
2380 AGUCUAC A AGGGCAU AUGCCCU CUGAUGAG X CGAA IUAGACU
2386 CAAGGGC A UCUGGAU AUCCAGA CUGAUGAG X CGAA ICCCUUG
2389 GGGCAUC U GGAUCCC GGGAUCC CUGAUGAG X CGAA IAUGCCC
2395 CUGGAUC C CUGAUGG CCAUCAG CUGAUGAG X CGAA IAUCCAG
2396 UGGAUCC C UGAUGGG CCCAUCA CUGAUGAG X CGAA IGAUCCA
2397 GGAUCCC U GAUGGGG CCCCAUC CUGAUGAG X CGAA IGGAUCC
2420 AAAAUUC C AGUGGCC GGCCACU CUGAUGAG X CGAA IAAUUUU
2421 AAAUUCC A GUGGCCA UGGCCAC CUGAUGAG X CGAA IGAAUUU
2427 CAGUGGC C AUCAAAG CUUUGAU CUGAUGAG X CGAA ICCACUG
2428 AGUGGCC A UCAAAGU ACUUUGA CUGAUGAG X CGAA IGCCACU
2431 GGCCAUC A AAGUGUU AACACUU CUGAUGAG X CGAA IAUGGCC
2449 GGAAAAC A CAUCCCC GGGGAUG CUGAUGAG X CGAA IUUUUCC
2451 AAAACAC A UCCCCCA UGGGGGA CUGAUGAG X CGAA IUGUUUU
2454 ACACAUC C CCCAAAG CUUUGGG CUGAUGAG X CGAA IAUGUGU
2455 CACAUCC C CCAAAGC GCUUUGG CUGAUGAG X CGAA IGAUGUG
2456 ACAUCCC C CAAAGCC GGCUUUG CUGAUGAG X CGAA IGGAUGU
2457 CAUCCCC C AAAGCCA UGGCUUU CUGAUGAG X CGAA IGGGAUG
2458 AUCCCCC A AAGCCAA UUGGCUU CUGAUGAG X CGAA IGGGGAU
2463 CCAAAGC C AACAAAG CUUUGUU CUGAUGAG X CGAA ICUUUGG
2464 CAAAGCC A ACAAAGA UCUUUGU CUGAUGAG X CGAA IGCUUUG
2467 AGCCAAC A AAGAAAU AUUUCUU CUGAUGAG X CGAA IUUGGCU
2476 AGAAAUC U UAGACGA UCGUCUA CUGAUGAG X CGAA IAUUUCU
2487 ACGAAGC A UACGUGA UCACGUA CUGAUGAG X CGAA ICUUCGU
2499 UGAUGGC U GGUGUGG CCACACC CUGAUGAG X CGAA ICCAUCA
2509 UGUGGGC U CCCCAUA UAUGGGG CUGAUGAG X CGAA ICCCACA
2511 UGGGCUC C CCAUAUG CAUAUGG CUGAUGAG X CGAA IAGCCCA
2512 GGGCUCC C CAUAUGU ACAUAUG CUGAUGAG X CGAA IGAGCCC
2513 GGCUCCC C AUAUGUC GACAUAU CUGAUGAG X CGAA IGGAGCC
Table 34
2514 GCUCCCC A UAUGUCU AGACAUA CUGAUGAG X CGAA IGGGAGC
2521 AUAUGUC U CCCGCCU AGGCGGG CUGAUGAG X CGAA IACAUAU
2523 AUGUCUC C CGCCUUC GAAGGCG CUGAUGAG X CGAA IAGACAU
2524 UGUCUCC C GCCUUCU AGAAGGC CUGAUGAG X CGAA IGAGACA
2527 CUCCCGC C UUCUGGG CCCAGAA CUGAUGAG X CGAA ICGGGAG
2528 UCCCGCC U UCUGGGC GCCCAGA CUGAUGAG X CGAA IGCGGGA
2531 CGCCUUC U GGGCAUC GAUGCCC CUGAUGAG X CGAA IAAGGCG
2536 UCUGGGC A UCUGCCU AGGCAGA CUGAUGAG X CGAA ICCCAGA
2539 GGGCAUC U GCCUGAC GUCAGGC CUGAUGAG X CGAA IAUGCCC
2542 CAUCUGC C UGACAUC GAUGUCA CUGAUGAG X CGAA ICAGAUG
2543 AUCUGCC U GACAUCC GGAUGUC CUGAUGAG X CGAA IGCAGAU
2547 GCCUGAC A UCCACGG CCGUGGA CUGAUGAG X CGAA IUCAGGC
2550 UGACAUC C ACGGUGC GCACCGU CUGAUGAG X CGAA IAUGUCA
2551 GACAUCC A CGGUGCA UGCACCG CUGAUGAG X CGAA IGAUGUC
2558 ACGGUGC A GCUGGUG CACCAGC CUGAUGAG X CGAA ICACCGU
2561 GUGCAGC U GGUGACA UGUCACC CUGAUGAG X CGAA ICUGCAC
2568 UGGUGAC A CAGCUUA UAAGCUG CUGAUGAG X CGAA IUCACCA
2570 GUGACAC A GCUUAUG CAUAAGC CUGAUGAG X CGAA IUGUCAC
2573 ACACAGC U UAUGCCC GGGCAUA CUGAUGAG X CGAA ICUGUGU
2579 CUUAUGC C CUAUGGC GCCAUAG CUGAUGAG X CGAA ICAUAAG
2580 UUAUGCC C UAUGGCU AGCCAUA CUGAUGAG X CGAA IGCAUAA
2581 UAUGCCC U AUGGCUG CAGCCAU CUGAUGAG X CGAA IGGCAUA
2587 CUAUGGC U GCCUCUU AAGAGGC CUGAUGAG X CGAA ICCAUAG
2590 UGGCUGC C UCUUAGA UCUAAGA CUGAUGAG X CGAA ICAGCCA
2591 GGCUGCC U CUUAGAC GUCUAAG CUGAUGAG X CGAA IGCAGCC
2593 CUGCCUC u UAGACCA UGGUCUA CUGAUGAG X CGAA IAGGCAG
2599 CUUAGAC c AUGUCCG CGGACAU CUGAUGAG X CGAA IUCUAAG
2600 UUAGACC A UGUCCGG CCGGACA CUGAUGAG X CGAA IGUCUAA
2605 CCAUGUC C GGGAAAA UUUUCCC CUGAUGAG X CGAA IACAUGG
2614 GGAAAAC C GCGGACG CGUCCGC CUGAUGAG X CGAA IUUUUCC
2623 CGGACGC C UGGGCUC GAGCCCA CUGAUGAG X CGAA ICGUCCG
2624 GGACGCC U GGGCUCC GGAGCCC CUGAUGAG X CGAA IGCGUCC
2629 CCUGGGC U CCCAGGA UCCUGGG CUGAUGAG X CGAA ICCCAGG
2631 UGGGCUC C CAGGACC GGUCCUG CUGAUGAG X CGAA IAGCCCA
2632 GGGCUCC C AGGACCU AGGUCCU CUGAUGAG X CGAA IGAGCCC
2633 GGCUCCC A GGACCUG CAGGUCC CUGAUGAG X CGAA IGGAGCC
2638 CCAGGAC C UGCUGAA UUCAGCA CUGAUGAG X CGAA IUCCUGG
2639 CAGGACC U GCUGAAC GUUCAGC CUGAUGAG X CGAA IGUCCUG
2642 GACCUGC U GAACUGG CCAGUUC CUGAUGAG X CGAA ICAGGUC
2647 GCUGAAC U GGUGUAU AUACACC CUGAUGAG X CGAA IUUCAGC
2657 UGUAUGC A GAUUGCC GGCAAUC CUGAUGAG X CGAA ICAUACA
2664 AGAUUGC C AAGGGGA UCCCCUU CUGAUGAG X CGAA ICAAUCU
2665 GAUUGCC A AGGGGAU AUCCCCU CUGAUGAG X CGAA IGCAAUC
2677 GAUGAGC U ACCUGGA UCCAGGU CUGAUGAG X CGAA ICUCAUC
2680 GAGCUAC C UGGAGGA UCCUCCA CUGAUGAG X CGAA IUAGCUC
2681 AGCUACC U GGAGGAU AUCCUCC CUGAUGAG X CGAA IGUAGCU
2696 GUGCGGC U CGUACAC GUGUACG CUGAUGAG X CGAA ICCGCAC
Table 34
2702 CUCGUAC A CAGGGAC GUCCCUG CUGAUGAG X CGAA IUACGAG
2704 CGUACAC A GGGACUU AAGUCCC CUGAUGAG X CGAA IUGUACG
2710 CAGGGAC U UGGCCGC GCGGCCA CUGAUGAG X CGAA IUCCCUG
2715 ACUUGGC C GCUCGGA UCCGAGC CUGAUGAG X CGAA ICCAAGU
2718 UGGCCGC U CGGAACG CGUUCCG CUGAUGAG X CGAA ICGGCCA
2729 AACGUGC U GGUCAAG CUUGACC CUGAUGAG X CGAA ICACGUU
2734 GCUGGUC A AGAGUCC GGACUCU CUGAUGAG X CGAA IACCAGC
2741 AAGAGUC C CAACCAU AUGGUUG CUGAUGAG X CGAA IACUCUU
2742 AGAGUCC C AACCAUG CAUGGUU CUGAUGAG X CGAA IGACUCU
2743 GAGUCCC A ACCAUGU ACAUGGU CUGAUGAG X CGAA IGGACUC
2746 UCCCAAC C AUGUCAA UUGACAU CUGAUGAG X CGAA IUUGGGA
2747 CCCAACC A UGUCAAA UUUGACA CUGAUGAG X CGAA IGUUGGG
2752 CCAUGUC A AAAUUAC GUAAUUU CUGAUGAG X CGAA IACAUGG
2760 AAAUUAC A GACUUCG CGAAGUC CUGAUGAG X CGAA IUAAUUU
2764 UACAGAC U UCGGGCU AGCCCGA CUGAUGAG X CGAA IUCUGUA
2771 UUCGGGC U GGCUCGG CCGAGCC CUGAUGAG X CGAA ICCCGAA
2775 GGCUGGC U CGGCUGC GCAGCCG CUGAUGAG X CGAA ICCAGCC
2780 GCUCGGC U GCUGGAC GUCCAGC CUGAUGAG X CGAA ICCGAGC
2783 CGGCUGC U GGACAUU AAUGUCC CUGAUGAG X CGAA ICAGCCG
2788 GCUGGAC A UUGACGA UCGUCAA CUGAUGAG X CGAA IUCCAGC
2799 ACGAGAC A GAGUACC GGUACUC CUGAUGAG X CGAA IUCUCGU
2806 AGAGUAC C AUGCAGA UCUGCAU CUGAUGAG X CGAA IUACUCU
2807 GAGUACC A UGCAGAU AUCUGCA CUGAUGAG X CGAA IGUACUC
2811 ACCAUGC A GAUGGGG CCCCAUC CUGAUGAG X CGAA ICAUGGU
2821 UGGGGGC A AGGUGCC GGCACCU CUGAUGAG X CGAA ICCCCCA
2828 AAGGUGC C CAUCAAG CUUGAUG CUGAUGAG X CGAA ICACCUU
2829 AGGUGCC C AUCAAGU ACUUGAU CUGAUGAG X CGAA IGCACCU
2830 GGUGCCC A UCAAGUG CACUUGA CUGAUGAG X CGAA IGGCACC
2833 GCCCAUC A AGUGGAU AUCCACU CUGAUGAG X CGAA IAUGGGC
2846 AUGGCGC U GGAGUCC GGACUCC CUGAUGAG X CGAA ICGCCAU
2853 UGGAGUC C AUUCUCC GGAGAAU CUGAUGAG X CGAA IACUCCA
2854 GGAGUCC A UUCUCCG CGGAGAA CUGAUGAG X CGAA IGACUCC
2858 UCCAUUC U CCGCCGG CCGGCGG CUGAUGAG X CGAA IAAUGGA
2860 CAUUCUC C GCCGGCG CGCCGGC CUGAUGAG X CGAA IAGAAUG
2863 UCUCCGC C GGCGGUU AACCGCC CUGAUGAG X CGAA ICGGAGA
2872 GCGGUUC A CCCACCA UGGUGGG CUGAUGAG X CGAA IAACCGC
2874 GGUUCAC C CACCAGA UCUGGUG CUGAUGAG X CGAA IUGAACC
2875 GUUCACC C ACCAGAG CUCUGGU CUGAUGAG X CGAA IGUGAAC
2876 UUCACCC A CCAGAGU ACUCUGG CUGAUGAG X CGAA IGGUGAA
2878 CACCCAC C AGAGUGA UCACUCU CUGAUGAG X CGAA IUGGGUG
2879 ACCCACC A GAGUGAU AUCACUC CUGAUGAG X CGAA IGUGGGU
2907 GUGUGAC U GUGUGGG CCCACAC CUGAUGAG X CGAA IUCACAC
2918 UGGGAGC U GAUGACU AGUCAUC CUGAUGAG X CGAA ICUCCCA
2925 UGAUGAC U UUUGGGG CCCCAAA CUGAUGAG X CGAA IUCAUCA
2934 UUGGGGC C AAACCUU AAGGUUU CUGAUGAG X CGAA ICCCCAA
2935 UGGGGCC A AACCUUA UAAGGUU CUGAUGAG X CGAA IGCCCCA
2939 GCCAAAC C UUACGAU AUCGUAA CUGAUGAG X CGAA IUUUGGC
Table 34
2940 CCAAACC U UACGAUG CAUCGUA CUGAUGAG X CGAA IGUUUGG
2953 UGGGAUC C CAGCCCG CGGGCUG CUGAUGAG X CGAA IAUCCCA
2954 GGGAUCC C AGCCCGG CCGGGCU CUGAUGAG X CGAA IGAUCCC
2955 GGAUCCC A GCCCGGG CCCGGGC CUGAUGAG X CGAA IGGAUCC
2958 UCCCAGC C CGGGAGA UCUCCCG CUGAUGAG X CGAA ICUGGGA
2959 CCCAGCC C GGGAGAU AUCUCCC CUGAUGAG X CGAA IGCUGGG
2968 GGAGAUC C CUGACCU AGGUCAG CUGAUGAG X CGAA IAUCUCC
2969 GAGAUCC C UGACCUG CAGGUCA CUGAUGAG X CGAA IGAUCUC
2970 AGAUCCC U GACCUGC GCAGGUC CUGAUGAG X CGAA IGGAUCU
2974 CCCUGAC C UGCUGGA UCCAGCA CUGAUGAG X CGAA IUCAGGG
2975 CCUGACC U GCUGGAA UUCCAGC CUGAUGAG X CGAA IGUCAGG
2978 GACCUGC U GGAAAAG CUUUUCC CUGAUGAG X CGAA ICAGGUC
2996 GAGCGGC U GCCCCAG CUGGGGC CUGAUGAG X CGAA ICCGCUC
2999 CGGCUGC C CCAGCCC GGGCUGG CUGAUGAG X CGAA ICAGCCG
3000 GGCUGCC C CAGCCCC GGGGCUG CUGAUGAG X CGAA IGCAGCC
3001 GCUGCCC C AGCCCCC GGGGGCU CUGAUGAG X CGAA IGGCAGC
3002 CUGCCCC A GCCCCCC GGGGGGC CUGAUGAG X CGAA IGGGCAG
3005 CCCCAGC C CCCCAUC GAUGGGG CUGAUGAG X CGAA ICUGGGG
3006 CCCAGCC C CCCAUCU AGAUGGG CUGAUGAG X CGAA IGCUGGG
3007 CCAGCCC C CCAUCUG CAGAUGG CUGAUGAG X CGAA IGGCUGG
3008 CAGCCCC C CAUCUGC GCAGAUG CUGAUGAG X CGAA IGGGCUG
3009 AGCCCCC C AUCUGCA UGCAGAU CUGAUGAG X CGAA IGGGGCU
3010 GCCCCCC A UCUGCAC GUGCAGA CUGAUGAG X CGAA IGGGGGC
3013 CCCCAUC U GCACCAU AUGGUGC CUGAUGAG X CGAA IAUGGGG
3016 CAUCUGC A CCAUUGA UCAAUGG CUGAUGAG X CGAA ICAGAUG
3018 UCUGCAC C AUUGAUG CAUCAAU CUGAUGAG X CGAA IUGCAGA
3019 CUGCACC A UUGAUGU ACAUCAA CUGAUGAG X CGAA IGUGCAG
3028 UGAUGUC U ACAUGAU AUCAUGU CUGAUGAG X CGAA IACAUCA
3031 UGUCUAC A UGAUCAU AUGAUCA CUGAUGAG X CGAA IUAGACA
3037 CAUGAUC A UGGUCAA UUGACCA CUGAUGAG X CGAA IAUCAUG
3043 CAUGGUC A AAUGUUG CAACAUU CUGAUGAG X CGAA IACCAUG
3061 GAUUGAC U CUGAAUG CAUUCAG CUGAUGAG X CGAA IUCAAUC
3063 UUGACUC U GAAUGUC GACAUUC CUGAUGAG X CGAA IAGUCAA
3074 UGUCGGC C AAGAUUC GAAUCUU CUGAUGAG X CGAA ICCGACA
3075 GUCGGCC A AGAUUCC GGAAUCU CUGAUGAG X CGAA IGCCGAC
3082 AAGAUUC C GGGAGUU AACUCCC CUGAUGAG X CGAA IAAUCUU
3096 UGGUGUC U GAAUUCU AGAAUUC CUGAUGAG X CGAA IACACCA
3103 UGAAUUC U CCCGCAU AUGCGGG CUGAUGAG X CGAA IAAUUCA
3105 AAUUCUC C CGCAUGG CCAUGCG CUGAUGAG X CGAA IAGAAUU
3106 AUUCUCC C GCAUGGC GCCAUGC CUGAUGAG X CGAA IGAGAAU
3109 CUCCCGC A UGGCCAG CUGGCCA CUGAUGAG X CGAA ICGGGAG
3114 GCAUGGC C AGGGACC GGUCCCU CUGAUGAG X CGAA ICCAUGC
3115 CAUGGCC A GGGACCC GGGUCCC CUGAUGAG X CGAA IGCCAUG
3121 CAGGGAC C CCCAGCG CGCUGGG CUGAUGAG X CGAA IUCCCUG
3122 AGGGACC C CCAGCGC GCGCUGG CUGAUGAG X CGAA IGUCCCU
3123 GGGACCC C CAGCGCU AGCGCUG CUGAUGAG X CGAA IGGUCCC
3124 GGACCCC C AGCGCUU AAGCGCU CUGAUGAG X CGAA IGGGUCC
Table 34
3125 GACCCCC A GCGCUUU AAAGCGC CUGAUGAG X CGAA IGGGGUC
3130 CCAGCGC U UUGUGGU ACCACAA CUGAUGAG X CGAA ICGCUGG
3139 UGUGGUC A UCCAGAA UUCUGGA CUGAUGAG X CGAA IACCACA
3142 GGUCAUC C AGAAUGA UCAUUCU CUGAUGAG X CGAA IAUGACC
3143 GUCAUCC A GAAUGAG CUCAUUC CUGAUGAG X CGAA IGAUGAC
3154 UGAGGAC U UGGGCCC GGGCCCA CUGAUGAG X CGAA IUCCUCA
3160 CUUGGGC C CAGCCAG CUGGCUG CUGAUGAG X CGAA ICCCAAG
3161 UUGGGCC C AGCCAGU ACUGGCU CUGAUGAG X CGAA IGCCCAA
3162 UGGGCCC A GCCAGUC GACUGGC CUGAUGAG X CGAA IGGCCCA
3165 GCCCAGC C AGUCCCU AGGGACU CUGAUGAG X CGAA ICUGGGC
3166 CCCAGCC A GUCCCUU AAGGGAC CUGAUGAG X CGAA IGCUGGG
3170 GCCAGUC C CUUGGAC GUCCAAG CUGAUGAG X CGAA IACUGGC
3171 CCAGUCC C UUGGACA UGUCCAA CUGAUGAG X CGAA IGACUGG
3172 CAGUCCC U UGGACAG CUGUCCA CUGAUGAG X CGAA IGGACUG
3178 CUUGGAC A GCACCUU AAGGUGC CUGAUGAG X CGAA IUCCAAG
3181 GGACAGC A CCUUCUA UAGAAGG CUGAUGAG X CGAA ICUGUCC
3183 ACAGCAC C UUCUACC GGUAGAA CUGAUGAG X CGAA IUGCUGU
3184 CAGCACC U UCUACCG CGGUAGA CUGAUGAG X CGAA IGUGCUG
3187 CACCUUC U ACCGCUC GAGCGGU CUGAUGAG X CGAA IAAGGUG
3190 CUUCUAC C GCUCACU AGUGAGC CUGAUGAG X CGAA IUAGAAG
3193 CUACCGC U CACUGCU AGCAGUG CUGAUGAG X CGAA ICGGUAG
3195 ACCGCUC A CUGCUGG CCAGCAG CUGAUGAG X CGAA IAGCGGU
3197 CGCUCAC U GCUGGAG CUCCAGC CUGAUGAG X CGAA IUGAGCG
3200 UCACUGC U GGAGGAC GUCCUCC CUGAUGAG X CGAA ICAGUGA
3214 CGAUGAC A UGGGGGA UCCCCCA CUGAUGAG X CGAA IUCAUCG
3223 GGGGGAC C UGGUGGA UCCACCA CUGAUGAG X CGAA IUCCCCC
3224 GGGGACC U GGUGGAU AUCCACC CUGAUGAG X CGAA IGUCCCC
3234 UGGAUGC U GAGGAGU ACUCCUC CUGAUGAG X CGAA ICAUCCA
3245 GAGUAUC U GGUACCC GGGUACC CUGAUGAG X CGAA IAUACUC
3251 CUGGUAC C CCAGCAG CUGCUGG CUGAUGAG X CGAA IUACCAG
3252 UGGUACC C CAGCAGG CCUGCUG CUGAUGAG X CGAA IGUACCA
3253 GGUACCC G AGCAGGG CCCUGCU CUGAUGAG X CGAA IGGUACC
3254 GUACCCC A GCAGGGC GCCCUGC CUGAUGAG X CGAA IGGGUAC
3257 CCCCAGC A GGGCUUC GAAGCCC CUGAUGAG X CGAA ICUGGGG
3262 GCAGGGC U UCUUCUG CAGAAGA CUGAUGAG X CGAA ICCCUGC
3265 GGGCUUC U UCUGUCC GGACAGA CUGAUGAG X CGAA IAAGCCC
3268 CUUCUUC U GUCCAGA UCUGGAC CUGAUGAG X CGAA IAAGAAG
3272 UUCUGUC C AGACCCU AGGGUCU CUGAUGAG X CGAA IACAGAA
3273 UCUGUCC A GACCCUG CAGGGUC CUGAUGAG X CGAA IGACAGA
3277 UCCAGAC C CUGCCCC GGGGCAG CUGAUGAG X CGAA IUCUGGA
3278 CCAGACC C UGCCCCG CGGGGCA CUGAUGAG X CGAA IGUCUGG
3279 CAGACCC U GCCCCGG CCGGGGC CUGAUGAG X CGAA IGGUCUG
3282 ACCCUGC C CCGGGCG CGCCCGG CUGAUGAG X CGAA ICAGGGU
3283 CCCUGCC C CGGGCGC GCGCCCG CUGAUGAG X CGAA IGCAGGG
3284 CCUGCCC C GGGCGCU AGCGCCC CUGAUGAG X CGAA IGGCAGG
3291 CGGGCGC U GGGGGCA UGCCCCC CUGAUGAG X CGAA ICGCCCG
3298 UGGGGGC A UGGUCCA UGGACCA CUGAUGAG X CGAA ICCCCCA
Table j4
3304 CAUGGUC C ACCACAG CUGUGGU CUGAUGAG X CGAA IACCAUG
3305 AUGGUCC A CCACAGG CCUGUGG CUGAUGAG X CGAA IGACCAU
3307 GGUCCAC C ACAGGCA UGCCUGU CUGAUGAG X CGAA IUGGACC
3308 GUCCACC A CAGGCAC GUGCCUG CUGAUGAG X CGAA IGUGGAC
3310 CCACCAC A GGCACCG CGGUGCC CUGAUGAG X CGAA IUGGUGG
3314 CACAGGC A CCGCAGC GCUGCGG CUGAUGAG X CGAA ICCUGUG
3316 CAGGCAC C GCAGCUC GAGCUGC CUGAUGAG X CGAA IUGCCUG
3319 GCACCGC A GCUCAUC GAUGAGC CUGAUGAG X CGAA ICGGUGC
3322 CCGCAGC U CAUCUAC GUAGAUG CUGAUGAG •X CGAA ICUGCGG
3324 GCAGCUC A UCUACCA UGGUAGA CUGAUGAG X CGAA IAGCUGC
3327 GCUCAUC U ACCAGGA UCCUGGU CUGAUGAG X CGAA IAUGAGC
3330 CAUCUAC C AGGAGUG CACUCCU CUGAUGAG X CGAA IUAGAUG
3331 AUCUACC A GGAGUGG CCACUCC CUGAUGAG X CGAA IGUAGAU
3349 UGGGGAC C UGACACU AGUGUCA CUGAUGAG X CGAA IUCCCCA
3350 GGGGACC U GACACUA UAGUGUC CUGAUGAG X CGAA IGUCCCC
3354 ACCUGAC A CUAGGGC GCCCUAG CUGAUGAG X CGAA IUCAGGU
3356 CUGACAC U AGGGCUG CAGCCCU CUGAUGAG X CGAA IUGUCAG
3362 CUAGGGC U GGAGCCC GGGCUCC CUGAUGAG X CGAA ICCCUAG
3368 CUGGAGC C CUCUGAA UUCAGAG CUGAUGAG X CGAA ICUCCAG
3369 UGGAGCC C UCUGAAG CUUCAGA CUGAUGAG X CGAA IGCUCCA
3370 GGAGCCC U CUGAAGA UCUUCAG CUGAUGAG X CGAA IGGCUCC
3372 AGCCCUC U GAAGAGG CCUCUUC CUGAUGAG X CGAA IAGGGCU
3384 AGGAGGC C CCCAGGU ACCUGGG CUGAUGAG X CGAA ICCUCCU
3385 GGAGGCC C CCAGGUC GACCUGG CUGAUGAG X CGAA IGCCUCC
3386 GAGGCCC C CAGGUCU AGACCUG CUGAUGAG X CGAA IGGCCUC
3387 AGGCCCC C AGGUCUC GAGACCU CUGAUGAG X CGAA IGGGCCU
3388 GGCCCCC A GGUCUCC GGAGACC CUGAUGAG X CGAA IGGGGCC
3393 CCAGGUC U CCACUGG CCAGUGG CUGAUGAG X CGAA IACCUGG
3395 AGGUCUC C ACUGGCA UGCCAGU CUGAUGAG X CGAA IAGACCU
3396 GGUCUCC A CUGGCAC GUGCCAG CUGAUGAG X CGAA IGAGACC
3398 UCUCCAC U GGCACCC GGGUGCC CUGAUGAG X CGAA IUGGAGA
3402 CACUGGC A CCCUCCG CGGAGGG CUGAUGAG X CGAA ICCAGUG
3404 CUGGCAC C CUCCGAA UUCGGAG CUGAUGAG X CGAA IUGCCAG
3405 UGGCACC C UCCGAAG CUUCGGA CUGAUGAG X CGAA IGUGCCA
3406 GGCACCC U CCGAAGG CCUUCGG CUGAUGAG X CGAA IGGUGCC
3408 CACCCUC C GAAGGGG CCCCUUC CUGAUGAG X CGAA IAGGGUG
3417 AAGGGGC U GGCUCCG CGGAGCC CUGAUGAG X CGAA ICCCCUU
3421 GGCUGGC U CCGAUGU ACAUCGG CUGAUGAG X CGAA ICCAGCC
3423 CUGGCUC C GAUGUAU AUACAUC CUGAUGAG X CGAA IAGCCAG
3442 UGGUGAC C UGGGAAU AUUCCCA CUGAUGAG X CGAA IUCACCA
3443 GGUGACC U GGGAAUG CAUUCCC CUGAUGAG X CGAA IGUCACC
3456 UGGGGGC A GCCAAGG CCUUGGC CUGAUGAG X CGAA ICCCCCA
3459 GGGCAGC C AAGGGGC GCCCCUU CUGAUGAG X CGAA ICUGCCC
3460 GGCAGCC A AGGGGCU AGCCCCU CUGAUGAG X CGAA IGCUGCC
3467 AAGGGGC U GCAAAGC GCUUUGC CUGAUGAG X CGAA ICCCCUU
3470 GGGCUGC A AAGCCUC GAGGCUU CUGAUGAG X CGAA ICAGCCC
3475 GCAAAGC C UCCCCAC GUGGGGA CUGAUGAG X CGAA ICUUUGC
Table 34
3476 CAAAGCC U CCCCACA UGUGGGG CUGAUGAG X CGAA IGCUUUG
3478 AAGCCUC C CCACACA UGUGUGG CUGAUGAG X CGAA IAGGCUU
3479 AGCCUCC C CACACAU AUGUGUG CUGAUGAG X CGAA IGAGGCU
3480 GCCUCCC C ACACAUG CAUGUGU CUGAUGAG X CGAA IGGAGGC
3481 CCUCCCC A CACAUGA UCAUGUG CUGAUGAG X CGAA IGGOAGG
3483 UCCCCAC A CAUGACC GGUCAUG CUGAUGAG X CGAA IUGGGGA
3485 CCCACAC A UGACCCC GGGGUCA CUGAUGAG X CGAA IUGUGGG
3490 ACAUGAC C CCAGCCC GGGCUGG CUGAUGAG X CGAA IUCAUGU
3491 CAUGACC C CAGCCCU AGGGCUG CUGAUGAG X CGAA IGUCAUG
3492 AUGACCC C AGCCCUC GAGGGCU CUGAUGAG X CGAA IGGUCAU
3493 UGACCCC A GCCCUCU AGAGGGC CUGAUGAG X CGAA IGGGUCA
3496 CCCCAGC C CUCUACA UGUAGAG CUGAUGAG X CGAA ICUGGGG
3497 CCCAGCC C UCUACAG CUGUAGA CUGAUGAG X CGAA IGCUGGG
3498 CCAGCCC U CUACAGC GCUGUAG CUGAUGAG X CGAA IGGCUGG
3500 AGCCCUC U ACAGCGG CCGCUGU CUGAUGAG X CGAA IAGGGCU
3503 CCUCUAC A GCGGUAC GUACCGC CUGAUGAG X CGAA IUAGAGG
3511 GCGGUAC A GUGAGGA UCCUCAC CUGAUGAG X CGAA IUACCGC
3520 UGAGGAC C CCACAGU ACUGUGG CUGAUGAG X CGAA IUCCUCA
3521 GAGGACC C CACAGUA UACUGUG CUGAUGAG X CGAA IGUCCUC
3522 AGGACCC C ACAGUAC GUACUGU CUGAUGAG X CGAA IGGUCCU
3523 GGACCCC A CAGUACC GGUACUG CUGAUGAG X CGAA IGGGUCC
3525 ACCCCAC A GUACCCC GGGGUAC CUGAUGAG X CGAA IUGGGGU
3530 ACAGUAC C CCUGCCC GGGCAGG CUGAUGAG X CGAA IUACUGU
3531 CAGUACC C CUGCCCU AGGGCAG CUGAUGAG X CGAA IGUACUG
3532 AGUACCC C UGCCCUC GAGGGCA CUGAUGAG X CGAA IGGUACU
3533 GUACCCC U GCCCUCU AGAGGGC CUGAUGAG X CGAA IGGGUAC
3536 CCCCUGC C CUCUGAG CUCAGAG CUGAUGAG X CGAA ICAGGGG
3537 CCCUGCC C UCUGAGA UCUCAGA CUGAUGAG X CGAA IGCAGGG
3538 CCUGCCC U CUGAGAC GUCUCAG CUGAUGAG X CGAA IGGCAGG
3540 UGCCCUC U GAGACUG CAGUCUC CUGAUGAG X CGAA IAGGGCA
3546 CUGAGAC U GAUGGCU AGCCAUC CUGAUGAG X CGAA IUCUCAG
3553 UGAUGGC U ACGUUGC GCAACGU CUGAUGAG X CGAA ICCAUCA
3561 ACGUUGC C CCCCUGA UCAGGGG CUGAUGAG X CGAA ICAACGU
3562 CGUUGCC C CCCUGAC GUCAGGG CUGAUGAG X CGAA IGCAACG
3563 GUUGCCC C CCUGACC GGUCAGG CUGAUGAG X CGAA IGGCAAC
3564 UUGCCCC C CUGACCU AGGUCAG CUGAUGAG X CGAA IGGGCAA
3565 UGCCCCC C UGACCUG CAGGUCA CUGAUGAG X CGAA IGGGGCA
3566 GCCCCCC U GACCUGC GCAGGUC CUGAUGAG X CGAA IGGGGGC
3570 CCCUGAC C UGCAGCC GGCUGCA CUGAUGAG X CGAA IUCAGGG
3571 CCUGACC U GCAGCCC GGGCUGC CUGAUGAG X CGAA IGUCAGG
3574 GACCUGC A GCCCCCA UGGGGGC CUGAUGAG X CGAA ICAGGUC
3577 CUGCAGC C CCCAGCC GGCUGGG CUGAUGAG X CGAA ICUGCAG
3578 UGCAGCC C CCAGCCU AGGCUGG CUGAUGAG X CGAA IGCUGCA
3579 GCAGCCC C CAGCCUG CAGGCUG CUGAUGAG X CGAA IGGCUGC
3580 CAGCCCC C AGCCUGA UCAGGCU CUGAUGAG X CGAA IGGGCUG
3581 AGCCCCC A GCCUGAA UUCAGGC CUGAUGAG X CGAA IGGGGCU
3584 CCCCAGC C UGAAUAU AUAUUCA CUGAUGAG X CGAA ICUGGGG
Table 34
3585 CCCAGCC U GAAUAUG CAUAUUC CUGAUGAG X CGAA IGCUGGG
3598 UGUGAAC C AGCCAGA UCUGGCU CUGAUGAG X CGAA IUUCACA
3599 GUGAACC A GCCAGAU AUCUGGC CUGAUGAG X CGAA IGUUCAC
3602 AACCAGC C AGAUGUU AACAUCU CUGAUGAG X CGAA ICUGGUU
3603 ACCAGCC A GAUGUUC GAACAUC CUGAUGAG X CGAA IGCUGGU
3614 GUUCGGC C CCAGCCC GGGCUGG CUGAUGAG X CGAA ICCGAAC
3615 UUCGGCC C CAGCCCC GGGGCUG CUGAUGAG X CGAA IGCCGAA
3616 UCGGCCC C AGCCCCC GGGGGCU CUGAUGAG X CGAA IGGCCGA
3617 CGGCCCC A GCCCCCU AGGGGGC CUGAUGAG X CGAA IGGGCCG
3620 CCCCAGC C CCCUUCG CGAAGGG CUGAUGAG X CGAA ICUGGGG
3621 CCCAGCC C CCUUCGC GCGAAGG CUGAUGAG X CGAA IGCUGGG
3622 CCAGCCC C CUUCGCC GGCGAAG CUGAUGAG X CGAA IGGCUGG
3623 CAGCCCC C UUCGCCC GGGCGAA CUGAUGAG X CGAA IGGGCUG
3624 AGCCCCC U UCGCCCC GGGGCGA CUGAUGAG X CGAA IGGGGCU
3629 CCUUCGC C CCGAGAG CUCUCGG CUGAUGAG X CGAA ICGAAGG
3630 CUUCGCC C CGAGAGG CCUCUCG CUGAUGAG X CGAA IGCGAAG
3631 UUCGCCC C GAGAGGG CCCUCUC CUGAUGAG X CGAA IGGCGAA
3640 AGAGGGC C CUCUGCC GGCAGAG CUGAUGAG X CGAA ICCCUCU
3641 GAGGGCC C UCUGCCU AGGCAGA CUGAUGAG X CGAA IGCCCUC
3642 AGGGCCC U CUGCCUG CAGGCAG CUGAUGAG X CGAA IGGCCCU
3644 GGCCCUC U GCCUGCU AGCAGGC CUGAUGAG X CGAA IAGGGCC
3647 CCUCUGC C UGCUGCC GGCAGCA CUGAUGAG X CGAA ICAGAGG
3648 CUCUGCC U GCUGCCC GGGCAGC CUGAUGAG X CGAA IGCAGAG
3651 UGCCUGC U GCCCGAC GUCGGGC CUGAUGAG X CGAA ICAGGCA
3654 CUGCUGC c CGACCUG CAGGUCG CUGAUGAG X CGAA ICAGCAG
3655 UGCUGCC c GACCUGC GCAGGUC CUGAUGAG X CGAA IGCAGCA
3659 GCCCGAC c UGCUGGU ACCAGCA CUGAUGAG X CGAA IUCGGGC
3660 CCCGACC u GCUGGUG CACCAGC CUGAUGAG X CGAA IGUCGGG
3663 GACCUGC u GGUGCCA UGGCACC CUGAUGAG X CGAA ICAGGUC
3669 CUGGUGC c ACUCUGG CCAGAGU CUGAUGAG X CGAA ICACCAG
3670 UGGUGCC A CUCUGGA UCCAGAG CUGAUGAG X CGAA IGCACCA
3672 GUGCCAC U CUGGAAA UUUCCAG CUGAUGAG X CGAA IUGGCAC
3674 GCCACUC U GGAAAGG CCUUUCC CUGAUGAG X CGAA IAGUGGC
3683 GAAAGGC C CAAGACU AGUCUUG CUGAUGAG X CGAA ICCUUUC
3684 AAAGGCC c AAGACUC GAGUCUU CUGAUGAG X CGAA IGCCUUU
3685 AAGGCCC A AGACUCU AGAGUCU CUGAUGAG X CGAA IGGCCUU
3690 CCAAGAC U CUCUCCC GGGAGAG CUGAUGAG X CGAA IUCUUGG
3692 AAGACUC U CUCCCCA UGGGGAG CUGAUGAG X CGAA IAGUCUU
3694 GACUCUC U CCCCAGG CCUGGGG CUGAUGAG X CGAA IAGAGUC
3696 CUCUCUC C CCAGGGA UCCCUGG CUGAUGAG X CGAA IAGAGAG
3697 UCUCUCC C CAGGGAA UUCCCUG CUGAUGAG X CGAA IGAGAGA
3698 CUCUCCC C AGGGAAG CUUCCCU CUGAUGAG X CGAA IGGAGAG
3699 ucucccc A GGGAAGA UCUUCCC CUGAUGAG X CGAA IGGGAGA
3718 GGUCGUC A AAGACGU ACGUCUU CUGAUGAG X CGAA IACGACC
3732 UUUUUGC C UUUGGGG CCCCAAA CUGAUGAG X CGAA ICAAAAA
3733 UUUUGCC U UUGGGGG CCCCCAA CUGAUGAG X CGAA IGCAAAA
3744 GGGGUGC C GUGGAGA UCUCCAC CUGAUGAG X CGAA ICACCCC
Table 34
3754 GGAGAAC C CCGAGUA UACUCGG CUGAUGAG X CGAA IUUCUCC
3755 GAGAACC C CGAGUAC GUACUCG CUGAUGAG X CGAA IGUUCUC
3756 AGAACCC C GAGUACU AGUACUC CUGAUGAG X CGAA IGGUUCU
3763 CGAGUAC U UGACACC GGUGUCA CUGAUGAG X CGAA IUACUCG
3768 ACUUGAC A CCCCAGG CCUGGGG CUGAUGAG X CGAA IUCAAGU
3770 UUGACAC C CCAGGGA UCCCUGG CUGAUGAG X CGAA IUGUCAA
3771 UGACACC C CAGGGAG CUCCCUG CUGAUGAG X CGAA IGUGUCA
3772 GACACCC C AGGGAGG CCUCCCU CUGAUGAG X CGAA IGGUGUC
3773 ACACCCC A GGGAGGA UCCUCCC CUGAUGAG X CGAA IGGGUGU
3783 GAGGAGC U GCCCCUC GAGGGGC CUGAUGAG X CGAA ICUCCUC
3786 GAGCUGC C CCUCAGC GCUGAGG CUGAUGAG X CGAA ICAGCUC
3787 AGCUGCC C CUCAGCC GGCUGAG CUGAUGAG X CGAA IGCAGCU
3788 GCUGCCC C UCAGCCC GGGCUGA CUGAUGAG X CGAA IGGCAGC
3789 CUGCCCC U CAGCCCC GGGGCUG CUGAUGAG X CGAA IGGGCAG
3791 GCCCCUC A GCCCCAC GUGGGGC CUGAUGAG X CGAA IAGGGGC
3794 CCUCAGC C CCACCCU AGGGUGG CUGAUGAG X CGAA ICUGAGG
3795 CUCAGCC C CACCCUC GAGGGUG CUGAUGAG X CGAA IGCUGAG
3796 UCAGCCC C ACCCUCC GGAGGGU CUGAUGAG X CGAA IGGCUGA
3797 CAGCCCC A CCCUCCU AGGAGGG CUGAUGAG X CGAA IGGGCUG
3799 GCCCCAC C CUCCUCC GGAGGAG CUGAUGAG X CGAA IUGGGGC
3800 CCCCACC C uccuccu AGGAGGA CUGAUGAG X CGAA IGUGGGG
3801 CCCACCC U CCUCCUG CAGGAGG CUGAUGAG X CGAA IGGUGGG
3803 CACCCUC C UCCUGCC GGCAGGA CUGAUGAG X CGAA IAGGGUG
3804 ACCCUCC U CCUGCCU AGGCAGG CUGAUGAG X CGAA IGAGGGU
3806 CCUCCUC C UGCCUUC GAAGGCA CUGAUGAG X CGAA IAGGAGG
3807 cuccucc U GCCUUCA UGAAGGC CUGAUGAG X CGAA IGAGGAG
3810 CUCCUGC C UUCAGCC GGCUGAA CUGAUGAG X CGAA ICAGGAG
3811 UCCUGCC U UCAGCCC GGGCUGA CUGAUGAG X CGAA IGCAGGA
3814 UGCCUUC A GCCCAGC GCUGGGC CUGAUGAG X CGAA IAAGGCA
3817 CUUCAGC C CAGCCUU AAGGCUG CUGAUGAG X CGAA ICUGAAG
3818 UUCAGCC C AGCCUUC GAAGGCU CUGAUGAG X CGAA IGCUGAA
3819 UCAGCCC A GCCUUCG CGAAGGC CUGAUGAG X CGAA IGGCUGA
3822 GCCCAGC C UUCGACA UGUCGAA CUGAUGAG X CGAA ICUGGGC
3823 CCCAGCC U UCGACAA UUGUCGA CUGAUGAG X CGAA IGCUGGG
3829 CUUCGAC A ACCUCUA UAGAGGU CUGAUGAG X CGAA IUCGAAG
3832 CGACAAC C UCUAUUA UAAUAGA CUGAUGAG X CGAA IUUGUCG
3833 GACAACC U CUAUUAC GUAAUAG CUGAUGAG X CGAA IGUUGUC
3835 CAACCUC U AUUACUG CAGUAAU CUGAUGAG X CGAA IAGGUUG
3841 CUAUUAC U GGGACCA UGGUCCC CUGAUGAG X CGAA IUAAUAG
3847 CUGGGAC C AGGACCC GGGUCCU CUGAUGAG X CGAA IUCCCAG
3848 UGGGACC A GGACCCA UGGGUCC CUGAUGAG X CGAA IGUCCCA
3853 CCAGGAC C CACCAGA UCUGGUG CUGAUGAG X CGAA IUCCUGG
3854 CAGGACC C ACCAGAG CUCUGGU CUGAUGAG X CGAA IGUCCUG
3855 AGGACCC A CCAGAGC GCUCUGG CUGAUGAG X CGAA IGGUCCU
3857 GACCCAC C AGAGCGG CCGCUCU CUGAUGAG X CGAA IUGGGUC
3858 ACCCACC A GAGCGGG CCCGCUC CUGAUGAG X CGAA IGUGGGU
3870 GGGGGGC U CCACCCA UGGGUGG CUGAUGAG X CGAA ICCCCCC
Table 34
3872 GGGGCUC C ACCCAGC GCUGGGU CUGAUGAG X CGAA IAGCCCC
3873 GGGCUCC A CCCAGCA UGCUGGG CUGAUGAG X CGAA IGAGCCC
3875 GCUCCAC C CAGCACC GGUGCUG CUGAUGAG X CGAA IUGGAGC
3876 CUCCACC C AGCACCU AGGUGCU CUGAUGAG X CGAA IGUGGAG
3877 UCCACCC A GCACCUU AAGGUGC CUGAUGAG X CGAA IGGUGGA
3880 ACCCAGC A CCUUCAA UUGAAGG CUGAUGAG X CGAA ICUGGGU
3882 CCAGCAC C UUCAAAG CUUUGAA CUGAUGAG X CGAA IUGCUGG
3883 CAGCACC U UCAAAGG CCUUUGA CUGAUGAG X CGAA IGUGCUG
3886 CACCUUC A AAGGGAC GUCCCUU CUGAUGAG X CGAA IAAGGUG
3894 AAGGGAC A CCUACGG CCGUAGG CUGAUGAG X CGAA IUCCCUU
3896 GGGACAC C UACGGCA UGCCGUA CUGAUGAG X CGAA IUGUCCC
3897 GGACACC U ACGGCAG CUGCCGU CUGAUGAG X CGAA IGUGUCC
3903 CUACGGC A GAGAACC GGUUCUC CUGAUGAG X CGAA ICCGUAG
3910 AGAGAAC C CAGAGUA UACUCUG CUGAUGAG X CGAA IUUCUCU
3911 GAGAACC C AGAGUAC GUACUCU CUGAUGAG X CGAA IGUUCUC
3912 AGAACCC A GAGUACC GGUACUC CUGAUGAG X CGAA IGGUUCU
3919 AGAGUAC C UGGGUCU AGACCCA CUGAUGAG X CGAA IUACUCU
3920 GAGUACC U GGGUCUG CAGACCC CUGAUGAG X CGAA IGUACUC
3926 CUGGGUC U GGACGUG CACGUCC CUGAUGAG X CGAA IACCCAG
3935 GACGUGC C AGUGUGA UCACACU CUGAUGAG X CGAA ICACGUC
3936 ACGUGCC A GUGUGAA UUCACAC CUGAUGAG X CGAA IGCACGU
3945 UGUGAAC C AGAAGGC GCCUUCU CUGAUGAG X CGAA IUUCACA
3946 GUGAACC A GAAGGCC GGCCUUC CUGAUGAG X CGAA IGUUCAC
3953 AGAAGGC C AAGUCCG CGGACUU CUGAUGAG X CGAA ICCUUCU
3954 GAAGGCC A AGUCCGC GCGGACU CUGAUGAG X CGAA IGCCUUC
3959 CCAAGUC C GCAGAAG CUUCUGC CUGAUGAG X CGAA IACUUGG
3962 AGUCCGC A GAAGCCC GGGCUUC CUGAUGAG X CGAA ICGGACU
3968 CAGAAGC C CUGAUGU ACAUCAG CUGAUGAG X CGAA ICUUCUG
3969 AGAAGCC C UGAUGUG CACAUCA CUGAUGAG X CGAA IGCUUCU
3970 GAAGCCC U GAUGUGU ACACAUC CUGAUGAG X CGAA IGGCUUC
3979 AUGUGUC C UCAGGGA UCCCUGA CUGAUGAG X CGAA IACACAU
3980 UGUGUCC U CAGGGAG CUCCCUG CUGAUGAG X CGAA IGACACA
3982 UGUCCUC A GGGAGCA UGCUCCC CUGAUGAG X CGAA IAGGACA
3989 AGGGAGC A GGGAAGG CCUUCCC CUGAUGAG X CGAA ICUCCCU
3998 GGAAGGC C UGACUUC GAAGUCA CUGAUGAG X CGAA ICCUUCC
3999 GAAGGCC U GACUUCU AGAAGUC CUGAUGAG X CGAA IGCCUUC
4003 GCCUGAC U UCUGCUG CAGCAGA CUGAUGAG X CGAA IUCAGGC
4006 UGACUUC U GCUGGCA UGCCAGC CUGAUGAG X CGAA IAAGUCA
4009 CUUCUGC U GGCAUCA UGAUGCC CUGAUGAG X CGAA ICAGAAG
4013 UGCUGGC A UCAAGAG CUCUUGA CUGAUGAG X CGAA ICCAGCA
4016 UGGCAUC A AGAGGUG CACCUCU CUGAUGAG X CGAA IAUGCCA
4031 GGAGGGC C CUCCGAC GUCGGAG CUGAUGAG X CGAA ICCCUCC
4032 GAGGGCC C UCCGACC GGUCGGA CUGAUGAG X CGAA IGCCCUC
4033 AGGGCCC U CCGACCA UGGUCGG CUGAUGAG X CGAA IGGCCCU
4035 GGCCCUC C GACCACU AGUGGUC CUGAUGAG X CGAA IAGGGCC
4039 CUCCGAC C ACUUCCA UGGAAGU CUGAUGAG X CGAA IUCGGAG
4040 UCCGACC A CUUCCAG CUGGAAG CUGAUGAG X CGAA IGUCGGA
Table 34
4042 CGACCAC U UCCAGGG CCCUGGA CUGAUGAG X CGAA IUGGUCG
4045 CCACUUC C AGGGGAA UUCCCCU CUGAUGAG X CGAA IAAGUGG
4046 CACUUCC A GGGGAAC GUUCCCC CUGAUGAG X CGAA IGAAGUG
4054 GGGGAAC C UGCCAUG CAUGGCA CUGAUGAG X CGAA IUUCCCC
4055 GGGAACC U GCCAUGC GCAUGGC CUGAUGAG X CGAA IGUUCCC
4058 AACCUGC C AUGCCAG CUGGCAU CUGAUGAG X CGAA ICAGGUU
4059 ACCUGCC A UGCCAGG CCUGGCA CUGAUGAG X CGAA IGCAGGU
4063 GCCAUGC C AGGAACC GGUUCCU CUGAUGAG X CGAA ICAUGGC
4064 CCAUGCC A GGAACCU AGGUUCC CUGAUGAG X CGAA IGCAUGG
4070 CAGGAAC C UGUCCUA UAGGACA CUGAUGAG X CGAA IUUCCUG
4071 AGGAACC U GUCCUAA UUAGGAC CUGAUGAG X CGAA IGUUCCU
4075 ACCUGUC C UAAGGAA UUCCUUA CUGAUGAG X CGAA IACAGGU
4076 CCUGUCC U AAGGAAC GUUCCUU CUGAUGAG X CGAA IGACAGG
4084 AAGGAAC C UUCCUUC GAAGGAA CUGAUGAG X CGAA IUUCCUU
4085 AGGAACC U UCCUUCC GGAAGGA CUGAUGAG X CGAA IGUUCCU
4088 AACCUUC C UUCCUGC GCAGGAA CUGAUGAG X CGAA IAAGGUU
4089 ACCUUCC U UCCUGCU AGCAGGA CUGAUGAG X CGAA IGAAGGU
4092 UUCCUUC C UGCUUGA UCAAGCA CUGAUGAG X CGAA IAAGGAA
4093 UCCUUCC U GCUUGAG CUCAAGC CUGAUGAG X CGAA IGAAGGA
4096 UUCCUGC U UGAGUUC GAACUCA CUGAUGAG X CGAA ICAGGAA
4104 UGAGUUC C CAGAUGG CCAUCUG CUGAUGAG X CGAA IAACUCA
4105 GAGUUCC C AGAUGGC GCCAUCU CUGAUGAG X CGAA IGAACUC
4106 AGUUCCC A GAUGGCU AGCCAUC CUGAUGAG X CGAA IGGAACU
4113 AGAUGGC U GGAAGGG CCCUUCC CUGAUGAG X CGAA ICCAUCU
4124 AGGGGUC C AGCCUCG CGAGGCU CUGAUGAG X CGAA IACCCCU
4125 GGGGUCC A GCCUCGU ACGAGGC CUGAUGAG X CGAA IGACCCC
4128 GUCCAGC C UCGUUGG CCAACGA CUGAUGAG X CGAA ICUGGAC
4129 UCCAGCC U CGUUGGA UCCAACG CUGAUGAG X CGAA IGCUGGA
4145 GAGGAAC A GCACUGG CCAGUGC CUGAUGAG X CGAA IUUCCUC
4148 GAACAGC A CUGGGGA UCCCCAG CUGAUGAG X CGAA ICUGUUC
4150 ACAGCAC U GGGGAGU ACUCCCC CUGAUGAG X CGAA IUGCUGU
4159 GGGAGUC U UUGUGGA UCCACAA CUGAUGAG X CGAA IACUCCC
4170 UGGAUUC U GAGGCCC GGGCCUC CUGAUGAG X CGAA IAAUCCA
4176 CUGAGGC C CUGCCCA UGGGCAG CUGAUGAG X CGAA ICCUCAG
4177 UGAGGCC C UGCCCAA UUGGGCA CUGAUGAG X CGAA IGCCUCA
4178 GAGGCCC U GCCCAAU AUUGGGC CUGAUGAG X CGAA IGGCCUC
4181 GCCCUGC C CAAUGAG CUCAUUG CUGAUGAG X CGAA ICAGGGC
4182 CCCUGCC C AAUGAGA UCUCAUU CUGAUGAG X CGAA IGCAGGG
4183 CCUGCCC A AUGAGAC GUCUCAU CUGAUGAG X CGAA IGGCAGG
4191 AUGAGAC U CUAGGGU ACCCUAG CUGAUGAG X CGAA IUCUCAU
4193 GAGACUC U AGGGUCC GGACCCU CUGAUGAG X CGAA IAGUCUC
4200 UAGGGUC C AGUGGAU AUCCACU CUGAUGAG X CGAA IACCCUA
4201 AGGGUCC A GUGGAUG CAUCCAC CUGAUGAG X CGAA IGACCCU
4210 UGGAUGC C ACAGCCC GGGCUGU CUGAUGAG X CGAA ICAUCCA
4211 GGAUGCC A CAGCCCA UGGGCUG CUGAUGAG X CGAA IGCAUCC
4213 AUGCCAC A GCCCAGC GCUGGGC CUGAUGAG X CGAA IUGGCAU
4216 CCACAGC C CAGCUUG CAAGCUG CUGAUGAG X CGAA ICUGUGG
Table 34
4217 CACAGCC C AGCUUGG CCAAGCU CUGAUGAG X CGAA IGCUGUG
4218 ACAGCCC A GCUUGGC GCCAAGC CUGAUGAG X CGAA IGGCUGU
4221 GCCCAGC U UGGCCCU AGGGCCA CUGAUGAG X CGAA ICUGGGC
4226 GCUUGGC C CUUUCCU AGGAAAG CUGAUGAG X CGAA ICCAAGC
4227 CUUGGCC C UUUCCUU AAGGAAA CUGAUGAG X CGAA IGCCAAG
4228 UUGGCCC U UUCCUUC GAAGGAA CUGAUGAG X CGAA IGGCCAA
4232 CCCUUUC C UUCCAGA UCUGGAA CUGAUGAG X CGAA IAAAGGG
4233 CCUUUCC U UCCAGAU AUCUGGA CUGAUGAG X CGAA IGAAAGG
4236 UUCCUUC C AGAUCCU AGGAUCU CUGAUGAG X CGAA IAAGGAA
4237 UCCUUCC A GAUCCUG CAGGAUC CUGAUGAG X CGAA IGAAGGA
4242 CCAGAUC C UGGGUAC GUACCCA CUGAUGAG X CGAA IAUCUGG
4243 CAGAUCC U GGGUACU AGUACCC CUGAUGAG X CGAA IGAUCUG
4250 UGGGUAC U GAAAGCC GGCUUUC CUGAUGAG X CGAA IUACCCA
4257 UGAAAGC C UUAGGGA UCCCUAA CUGAUGAG X CGAA ICUUUCA
4258 GAAAGCC u UAGGGAA UUCCCUA CUGAUGAG X CGAA IGCUUUC
4268 GGGAAGC u GGCCUGA UCAGGCC CUGAUGAG X CGAA ICUUCCC
4272 AGCUGGC c UGAGAGG CCUCUCA CUGAUGAG X CGAA ICCAGCU
4273 GCUGGCC u GAGAGGG CCCUCUC CUGAUGAG X CGAA IGCCAGC
4289 AAGCGGC c CUAAGGG CCCUUAG CUGAUGAG X CGAA ICCGCUU
4290 AGCGGCC c UAAGGGA UCCCUUA CUGAUGAG X CGAA IGCCGCU
4291 GCGGCCC u AAGGGAG CUCCCUU CUGAUGAG X CGAA IGGCCGC
4303 GAGUGUC u AAGAACA UGUUCUU CUGAUGAG X CGAA IACACUC
4310 UAAGAAC A AAAGCGA UCGCUUU CUGAUGAG X CGAA IUUCUUA
4319 AAGCGAC C CAUUCAG CUGAAUG CUGAUGAG X CGAA IUCGCUU
4320 AGCGACC C AUUCAGA UCUGAAU CUGAUGAG X CGAA IGUCGCU
4321 GCGACCC A UUCAGAG CUCUGAA CUGAUGAG X CGAA IGGUCGC
4325 CCCAUUC A GAGACUG CAGUCUC CUGAUGAG X CGAA IAAUGGG
4331 CAGAGAC U GUCCCUG CAGGGAC CUGAUGAG X CGAA IUCUCUG
4335 GACUGUC C CUGAAAC GUUUCAG CUGAUGAG X CGAA IACAGUC
4336 ACUGUCC C UGAAACC GGUUUCA CUGAUGAG X CGAA IGACAGU
4337 CUGUCCC U GAAACCU AGGUUUC CUGAUGAG X CGAA IGGACAG
4343 CUGAAAC G UAGUACU AGUACUA CUGAUGAG X CGAA IUUUCAG
4344 UGAAACC U AGUACUG CAGUACU CUGAUGAG X CGAA IGUUUCA
4350 CUAGUAC U GCCCCCC GGGGGGC CUGAUGAG X CGAA IUACUAG
4353 GUACUGC C CCCCAUG CAUGGGG CUGAUGAG X CGAA ICAGUAC
4354 UACUGCC C CCCAUGA UCAUGGG CUGAUGAG X CGAA IGCAGUA
4355 ACUGCCC C CCAUGAG CUCAUGG CUGAUGAG X CGAA IGGCAGU
4356 CUGCCCC C CAUGAGG CCUCAUG CUGAUGAG X CGAA IGGGCAG
4357 UGCCCCC C AUGAGGA UCCUCAU CUGAUGAG X CGAA IGGGGCA
4358 GCCCCCC A UGAGGAA UUCCUCA CUGAUGAG X CGAA IGGGGGC
4371 AAGGAAC A GCAAUGG CCAUUGC CUGAUGAG X CGAA IUUCCUU
4374 GAACAGC A AUGGUGU ACACCAU CUGAUGAG X CGAA ICUGUUC
4383 UGGUGUC A GUAUCCA UGGAUAC CUGAUGAG X CGAA IACACCA
4389 CAGUAUC C AGGCUUU AAAGCCU CUGAUGAG X CGAA IAUACUG
4390 AGUAUCC A GGCUUUG CAAAGCC CUGAUGAG X CGAA IGAUACU
4394 UCCAGGC U UUGUACA UGUACAA CUGAUGAG X CGAA ICCUGGA
4401 UUUGUAC A GAGUGCU AGCACUC CUGAUGAG X CGAA IUACAAA
Table 34
Seq Accession No. = HSERB2R (Human c-erb-B-2 mRNA; 4473 bp) Core Sequence = CUGAUGAG X CGAA (X = GCCGAAAGGC or other stem II)
Table 35 Table 35: HBV Strains and Accession numbers
Table 36: HBV Substrate Sequence
LOCUS HPBADR1CG 3221 bp DNA circular VRL
06-MAR-1995
DEFINITION Hepatitis B virus , complete genome.
ACCESSION M38454
*The nucleotide number referred to in that table is the position of the 5' end of the oligo in this sequence.
Table 37 Table 37: Human HBV Hammerhead Ribozyme and Target Sequence
Pos Substrate Seq ID Ribozyme Rz Seq ID
13 CCACCACT T TCCACCAA 34 UUGGUGGA CUGAUGAG X CGAA AGUGGUGG 2543
14 CACCACTT T CCACCAAA 35 UUUGGUGG CUGAUGAG X CGAA AAGUGGUG 2544
15 ACCACTTT C CACCAAAC 36 GUUUGGUG CUGAUGAG X CGAA AAAGUGGU 2545
25 ACCAAACT C TTCAAGAT 37 AUCUUGAA CUGAUGAG X CGAA AGUUUGGU 2546
27 CAAACTCT T CAAGATCC 38 GGAUCUUG CUGAUGAG X CGAA AGAGUUUG 2547
28 AAACTCTT C AAGATCCC 39 GGGAUCUU CUGAUGAG X CGAA AAGAGUUU 2548
34 TTCAAGAT C CCAGAGTC 40 GACUCUGG CUGAUGAG X CGAA AUCUUGAA 2549
42 CCCAGAGT C AGGGCCCT 41 AGGGCCCU CUGAUGAG X CGAA ACUCUGGG 2550
53 GGCCCTGT A CTTTCCTG 42 CAGGAAAG CUGAUGAG X CGAA ACAGGGCC 2551
56 CCTGTACT T TCCTGCTG 43 CAGCAGGA CUGAUGAG X CGAA AGUACAGG 2552
57 CTGTACTT T CCTGCTGG 44 CCAGCAGG CUGAUGAG X CGAA AAGUACAG 2553
58 TGTACTTT C CTGCTGGT 45 ACCAGCAG CUGAUGAG X CGAA AAAGUACA 2554
71 TGGTGGCT C CAGTTCAG 46 CUGAACUG CUGAUGAG X CGAA AGCCACCA 2555
76 GCTCCAGT T CAGGAACA 47 UGUUCCUG CUGAUGAG X CGAA ACUGGAGC 2556
77 CTCCAGTT C AGGAACAG 48 CUGUUCCU CUGAUGAG X CGAA AACUGGAG 2557
97 GCCCTGCT C AGAATACT 49 AGUAUUCU CUGAUGAG X CGAA AGCAGGGC 2558
103 CTCAGAAT A CTGTCTCT 50 AGAGACAG CUGAUGAG X CGAA AUUCUGAG 2559
108 AATACTGT C TCTGCCAT 51 AUGGCAGA CUGAUGAG X CGAA ACAGUAUU 2560
110 TACTGTCT C TGCCATAT 52 AUAUGGCA CUGAUGAG X CGAA AGACAGUA 2561
117 TCTGCCAT A TCGTCAAT 53 AUUGACGA CUGAUGAG X CGAA AUGGCAGA 2562
119 TGCCATAT C GTCAATCT 54 AGAUUGAC CUGAUGAG X CGAA AUAUGGCA 2563
122 CATATCGT C AATCTTAT 55 AUAAGAUU CUGAUGAG X CGAA ACGAUAUG 2564
126 TCGTCAAT C TTATCGAA 56 UUCGAUAA CUGAUGAG X CGAA AUUGACGA 2565
128 GTCAATCT T ATCGAAGA 57 UCUUCGAU CUGAUGAG X CGAA AGAUUGAC 2566
129 TCAATCTT A TCGAAGAC 58 GUCUUCGA CUGAUGAG X CGAA AAGAUUGA 2567
131 AATCTTAT C GAAGACTG 59 CAGUCUUC CUGAUGAG X CGAA AUAAGAUU 2568
150 GACCCTGT A CCGAACAT 60 AUGUUCGG CUGAUGAG X CGAA ACAGGGUC 2569
168 GAGAACAT C GCATCAGG 61 CCUGAUGC CUGAUGAG X CGAA AUGUUCUC 2570
173 CATCGCAT C AGGACTCC 62 GGAGUCCU CUGAUGAG X CGAA AUGCGAUG 2571
180 TCAGGACT C CTAGGACC 63 GGUCCUAG CUGAUGAG X CGAA AGUCCUGA 2572
183 GGACTCCT A GGACCCCT 64 AGGGGUCC CUGAUGAG X CGAA AGGAGUCC 2573
195 CCCCTGCT C GTGTTACA 65 UGUAACAC CUGAUGAG X CGAA AGCAGGGG 2574
200 GCTCGTGT T ACAGGCGG 66 CCGCCUGU CUGAUGAG X CGAA ACACGAGC 2575
201 CTCGTGTT A CAGGCGGG 67 CCCGCCUG CUGAUGAG X CGAA AACACGAG 2576
212 GGCGGGGT T TTTCTTGT 68 ACAAGAAA CUGAUGAG X CGAA ACCCCGCC 2577
213 GCGGGGTT T TTCTTGTT 69 AACAAGAA CUGAUGAG X CGAA AACCCCGC 2578
214 CGGGGTTT T TCTTGTTG 70 CAACAAGA CUGAUGAG X CGAA AAACCCCG 2579
215 GGGGTTTT T CTTGTTGA 71 UCAACAAG CUGAUGAG X CGAA AAAACCCC 2580
216 GGGTTTTT c TTGTTGAC 72 GUCAACAA CUGAUGAG X CGAA AAAAACCC 2581
218 GTTTTTCT T GTTGACAA 73 UUGUCAAC CUGAUGAG X CGAA AGAAAAAC 2582
221 TTTCTTGT T GACAAAAA 74 UUUUUGUC CUGAUGAG X CGAA ACAAGAAA 2583
231 ACAAAAAT c CTCACAAT 75 AUUGUGAG CUGAUGAG X CGAA AUUUUUGU 2584
234 AAAATCCT c ACAATACC 76 GGUAUUGU CUGAUGAG X CGAA AGGAUUUU 2585
240 CTCACAAT A CCACAGAG 77 CUCUGUGG CUGAUGAG X CGAA AUUGUGAG 2586
250 CACAGAGT C TAGACTCG 78 CGAGUCUA CUGAUGAG X CGAA ACUCUGUG 2587
252 CAGAGTCT A GACTCGTG 79 CACGAGUC CUGAUGAG X CGAA AGACUCUG 2588
257 TCTAGACT C GTGGTGGA 80 UCCACCAC CUGAUGAG X CGAA AGUCUAGA 2589
268 GGTGGACT T CTCTCAAT 81 AUUGAGAG CUGAUGAG X CGAA AGUCCACC 2590
Table 37
269 GTGGACTT C TCTCAATT 82 AAUUGAGA CUGAUGAG X CGAA AAGUCCAC 2591
271 GGACTTCT C TCAATTTT 83 AAAAUUGA CUGAUGAG X CGAA AGAAGUCC 2592
273 ACTTCTCT C AATTTTCT 84 AGAAAAUU CUGAUGAG X CGAA AGAGAAGU 2593
277 CTCTCAAT T TTCTAGGG 85 CCCUAGAA CUGAUGAG X CGAA AUUGAGAG 2594
278 TCTCAATT T TCTAGGGG 86 CCCCUAGA CUGAUGAG X CGAA AAUUGAGA 2595
279 CTCAATTT T CTAGGGGG 87 CCCCCUAG CUGAUGAG X CGAA AAAUUGAG 2596
280 TCAATTTT C TAGGGGGA 88 UCCCCCUA CUGAUGAG X CGAA AAAAUUGA 2597
282 AATTTTCT A GGGGGAAC 89 GUUCCCCC CUGAUGAG X CGAA AGAAAAUU 2598
301 CCGTGTGT C TTGGCCAA 90 UUGGCCAA CUGAUGAG X CGAA ACACACGG 2599
303 GTGTGTCT T GGCCAAAA 91 UUUUGGCC CUGAUGAG X CGAA AGACACAC 2600
313 GCCAAAAT T CGCAGTCC 92 GGACUGCG CUGAUGAG X CGAA AUUUUGGC 2601
314 CCAAAATT c GCAGTCCC 93 GGGACUGC CUGAUGAG X CGAA AAUUUUGG 2602
320 TTCGCAGT c CCAAATCT 94 AGAUUUGG CUGAUGAG X CGAA ACUGCGAA 2603
327 TCCCAAAT c TCCAGTCA 95 UGACUGGA CUGAUGAG X CGAA AUUUGGGA 2604
329 CCAAATCT c CAGTCACT 96 AGUGACUG CUGAUGAG X CGAA AGAUUUGG 2605
334 TCTCCAGT c ACTCACCA 97 UGGUGAGU CUGAUGAG X CGAA ACUGGAGA 2606
338 CAGTCACT c ACCAACCT 98 AGGUUGGU CUGAUGAG X CGAA AGUGACUG 2607
349 CAACCTGT T GTCCTCCA 99 UGGAGGAC CUGAUGAG X CGAA ACAGGUUG 2608
352 CCTGTTGT c CTCCAATT 100 AAUUGGAG CUGAUGAG X CGAA ACAACAGG 2609
355 GTTGTCCT c CAATTTGT 101 ACAAAUUG CUGAUGAG X CGAA AGGACAAC 2610
360 CCTCCAAT T TGTCCTGG 102 CCAGGACA CUGAUGAG X CGAA AUUGGAGG 2611
361 CTCCAATT T GTCCTGGT 103 ACCAGGAC CUGAUGAG X CGAA AAUUGGAG 2612
364 CAATTTGT c CTGGTTAT 104 AUAACCAG CUGAUGAG X CGAA ACAAAUUG 2613
370 GTCCTGGT T ATCGCTGG 105 CCAGCGAU CUGAUGAG X CGAA ACCAGGAC 2614
371 TCCTGGTT A TCGCTGGA 106 UCCAGCGA CUGAUGAG X CGAA AACCAGGA 2615
373 CTGGTTAT c GCTGGATG 107 CAUCCAGC CUGAUGAG X CGAA AUAACCAG 2616
385 GGATGTGT c TGCGGCGT 108 ACGCCGCA CUGAUGAG X CGAA ACACAUCC 2617
394 TGCGGCGT T TTATCATC 109 GAUGAUAA CUGAUGAG X CGAA ACGCCGCA 2618
395 GCGGCGTT T TATCATCT 110 AGAUGAUA CUGAUGAG X CGAA AACGCCGC 2619
396 CGGCGTTT T ATCATCTT 111 AAGAUGAU CUGAUGAG X CGAA AAACGCCG 2620
397 GGCGTTTT A TCATCTTC 112 GAAGAUGA CUGAUGAG X CGAA AAAACGCC 2621
399 CGTTTTAT C ATCTTCCT 113 AGGAAGAU CUGAUGAG X CGAA AUAAAACG 2622
402 TTTATCAT c TTCCTCTG 114 CAGAGGAA CUGAUGAG X CGAA AUGAUAAA 2623
404 TATCATCT T CCTCTGCA 115 UGCAGAGG CUGAUGAG X CGAA AGAUGAUA 2624
405 ATCATCTT c CTCTGCAT 116 AUGCAGAG CUGAUGAG X CGAA AAGAUGAU 2625
408 ATCTTCCT c TGCATCCT 117 AGGAUGCA CUGAUGAG X CGAA AGGAAGAU 2626
414 CTCTGCAT c CTGCTGCT 118 AGCAGCAG CUGAUGAG X CGAA AUGCAGAG 2627
423 CTGCTGCT A TGCCTCAT 119 AUGAGGCA CUGAUGAG X CGAA AGCAGCAG 2628
429 CTATGCCT C ATCTTCTT 120 AAGAAGAU CUGAUGAG X CGAA AGGCAUAG 2629
432 TGCCTCAT C TTCTTGTT 121 AACAAGAA CUGAUGAG X CGAA AUGAGGCA 2630
434 CCTCATCT T CTTGTTGG 122 CCAACAAG CUGAUGAG X CGAA AGAUGAGG 2631
435 CTCATCTT c TTGTTGGT 123 ACCAACAA CUGAUGAG X CGAA AAGAUGAG 2632
437 CATCTTCT T GTTGGTTC 124 GAACCAAC CUGAUGAG X CGAA AGAAGAUG 2633
440 CTTCTTGT T GGTTCTTC 125 GAAGAACC CUGAUGAG X CGAA ACAAGAAG 2634
444 TTGTTGGT T CTTCTGGA 126 UCCAGAAG CUGAUGAG X CGAA ACCAACAA 2635
445 TGTTGGTT c TTCTGGAC 127 GUCCAGAA CUGAUGAG X CGAA AACCAACA 2636
447 TTGGTTCT T CTGGACTA 128 UAGUCCAG CUGAUGAG X CGAA AGAACCAA 2637
448 TGGTTCTT c TGGACTAT 129 AUAGUCCA CUGAUGAG X CGAA AAGAACCA 2638
455 TCTGGACT A TCAAGGTA 130 UACCUUGA CUGAUGAG X CGAA AGUCCAGA 2639
457 TGGACTAT C AAGGTATG 131 CAUACCUU CUGAUGAG X CGAA AUAGUCCA 2640
463 ATCAAGGT A TGTTGCCC 132 GGGCAACA CUGAUGAG X CGAA ACCUUGAU 2641
Table 3/
467 AGGTATGT T GCCCGTTT 133 AAACGGGC CUGAUGAG X CGAA ACAUACCU 2642
474 TTGCCCGT T TGTCCTCT 134 AGAGGACA CUGAUGAG X CGAA ACGGGCAA 2643
475 TGCCCGTT T GTCCTCTA 135 UAGAGGAC CUGAUGAG X CGAA AACGGGCA 2644
478 CCGTTTGT c CTCTAATT 136 AAUUAGAG CUGAUGAG X CGAA ACAAACGG 2645
481 TTTGTCCT c TAATTCCA 137 UGGAAUUA CUGAUGAG X CGAA AGGACAAA 2646
483 TGTCCTCT A ATTCCAGG 138 CCUGGAAU CUGAUGAG X CGAA AGAGGACA 2647
486 CCTCTAAT T CCAGGATC 139 GAUCCUGG CUGAUGAG X CGAA AUUAGAGG 2648
487 CTCTAATT C CAGGATCA 140 UGAUCCUG CUGAUGAG X CGAA AAUUAGAG 2649
494 TCCAGGAT C ATCAACAA 141 UUGUUGAU CUGAUGAG X CGAA AUCCUGGA 2650
497 AGGATCAT C AACAACCA 142 UGGUUGUU CUGAUGAG X CGAA AUGAUCCU 2651
535 GCACAACT C CTGCTCAA 143 UUGAGCAG CUGAUGAG X CGAA AGUUGUGC 2652
541 CTCCTGCT C AAGGAACC 144 GGUUCCUU CUGAUGAG X CGAA AGCAGGAG 2653
551 AGGAACCT C TATGTTTC 145 GAAACAUA CUGAUGAG X CGAA AGGUUCCU 2654
553 GAACCTCT A TGTTTCCC 146 GGGAAACA CUGAUGAG X CGAA AGAGGUUC 2655
557 CTCTATGT T TCCCTCAT 147 AUGAGGGA CUGAUGAG X CGAA ACAUAGAG 2656
558 TCTATGTT T CCCTCATG 148 CAUGAGGG CUGAUGAG X CGAA AACAUAGA 2657
559 CTATGTTT C CCTCATGT 149 ACAUGAGG CUGAUGAG X CGAA AAACAUAG 2658
563 GTTTCCCT C ATGTTGCT 150 AGCAACAU CUGAUGAG X CGAA AGGGAAAC 2659
568 CCTCATGT T GCTGTACA 151 UGUACAGC CUGAUGAG X CGAA ACAUGAGG 2660
574 GTTGCTGT A CAAAACCT 152 AGGUUUUG CUGAUGAG X CGAA ACAGCAAC 2661
583 CAAAACCT A CGGACGGA 153 UCCGUCCG CUGAUGAG X CGAA AGGUUUUG 2662
604 GCACCTGT A TTCCCATC 154 GAUGGGAA CUGAUGAG X CGAA ACAGGUGC 2663
606 ACCTGTAT T CCCATCCC 155 GGGAUGGG CUGAUGAG X CGAA AUACAGGU 2664
607 CCTGTATT C CCATCCCA 156 UGGGAUGG CUGAUGAG X CGAA AAUACAGG 2665
612 ATTCCCAT C CCATCATC 157 GAUGAUGG CUGAUGAG X CGAA AUGGGAAU 2666
617 CATCCCAT C ATCTTGGG 158 CCCAAGAU CUGAUGAG X CGAA AUGGGAUG 2667
620 CCCATCAT C TTGGGCTT 159 AAGCCCAA CUGAUGAG X CGAA AUGAUGGG 2668
622 CATCATCT T GGGCTTTC 160 GAAAGCCC CUGAUGAG X CGAA AGAUGAUG 2669
628 CTTGGGCT T TCGCAAAA 161 UUUUGCGA CUGAUGAG X CGAA AGCCCAAG 2670
629 TTGGGCTT T CGCAAAAT 162 AUUUUGCG CUGAUGAG X CGAA AAGCCCAA 2671
630 TGGGCTTT C GCAAAATA 163 UAUUUUGC CUGAUGAG X CGAA AAAGCCCA 2672
638 CGCAAAAT A CCTATGGG 164 CCCAUAGG CUGAUGAG X CGAA AUUUUGCG 2673
642 AAATACCT A TGGGAGTG 165 CACUCCCA CUGAUGAG X CGAA AGGUAUUU 2674
656 GTGGGCCT C AGTCCGTT 166 AACGGACU CUGAUGAG X CGAA AGGCCCAC 2675
660 GCCTCAGT C CGTTTCTC 167 GAGAAACG CUGAUGAG X CGAA ACUGAGGC 2676
664 CAGTCCGT T TCTCTTGG 168 CCAAGAGA CUGAUGAG X CGAA ACGGACUG 2677
665 AGTCCGTT T CTCTTGGC 169 GCCAAGAG CUGAUGAG X CGAA AACGGACU 2678
666 GTCCGTTT c TCTTGGCT 170 AGCCAAGA CUGAUGAG X CGAA AAACGGAC 2679
668 CCGTTTCT c TTGGCTCA 171 UGAGCCAA CUGAUGAG X CGAA AGAAACGG 2680
670 GTTTCTCT T GGCTCAGT 172 ACUGAGCC CUGAUGAG X CGAA AGAGAAAC 2681
675 TCTTGGCT c AGTTTACT 173 AGUAAACU CUGAUGAG X CGAA AGCCAAGA 2682
679 GGCTCAGT T TACTAGTG 174 CACUAGUA CUGAUGAG X CGAA ACUGAGCC 2683
680 GCTCAGTT T ACTAGTGC 175 GCACUAGU CUGAUGAG X CGAA AACUGAGC 2684
681 CTCAGTTT A CTAGTGCC 176 GGCACUAG CUGAUGAG X CGAA AAACUGAG 2685
684 AGTTTACT A GTGCCATT 177 AAUGGCAC CUGAUGAG X CGAA AGUAAACU 2686
692 AGTGCCAT T TGTTCAGT 178 ACUGAACA CUGAUGAG X CGAA AUGGCACU 2687
693 GTGCCATT T GTTCAGTG 179 CACUGAAC CUGAUGAG X CGAA AAUGGCAC 2688
696 CCATTTGT T CAGTGGTT 180 AACCACUG CUGAUGAG X CGAA ACAAAUGG 2689
697 CATTTGTT C AGTGGTTC 181 GAACCACU CUGAUGAG X CGAA AACAAAUG 2690
704 TCAGTGGT T CGTAGGGC 182 GCCCUACG CUGAUGAG X CGAA ACCACUGA 2691
705 CAGTGGTT C GTAGGGCT 183 AGCCCUAC CUGAUGAG X CGAA AACCACUG 2692
Table 37
708 TGGTTCGT A GGGCTTTC 184 GAAAGCCC CUGAUGAG X CGAA ACGAACCA 2693
714 GTAGGGCT T TCCCCCAC 185 GUGGGGGA CUGAUGAG X CGAA AGCCCUAC 2694
715 TAGGGCTT T CCCCCACT 186 AGUGGGGG CUGAUGAG X CGAA AAGCCCUA 2695
716 AGGGCTTT C CCCCACTG 187 CAGUGGGG CUGAUGAG X CGAA AAAGCCCU 2696
726 CCCACTGT C TGGCTTTC 188 GAAAGCCA CUGAUGAG X CGAA ACAGUGGG 2697
732 GTCTGGCT T TCAGTTAT 189 AUAACUGA CUGAUGAG X CGAA AGCCAGAC 2698
733 TCTGGCTT T CAGTTATA 190 UAUAACUG CUGAUGAG X CGAA AAGCCAGA 2699
734 CTGGCTTT C AGTTATAT 191 AUAUAACU CUGAUGAG X CGAA AAAGCCAG 2700
738 CTTTCAGT T ATATGGAT 192 AUCCAUAU CUGAUGAG X CGAA ACUGAAAG 2701
739 TTTCAGTT A TATGGATG 193 CAUCCAUA CUGAUGAG X CGAA AACUGAAA 2702
741 TCAGTTAT A TGGATGAT 194 AUCAUCCA CUGAUGAG X CGAA AUAACUGA 2703
755 GATGTGGT T TTGGGGGC 195 GCCCCCAA CUGAUGAG X CGAA ACCACAUC 2704
756 ATGTGGTT T TGGGGGCC 196 GGCCCCCA CUGAUGAG X CGAA AACCACAU 2705
757 TGTGGTTT T GGGGGCCA 197 UGGCCCCC CUGAUGAG X CGAA AAACCACA 2706
769 GGCCAAGT C TGTACAAC 198 GUUGUACA CUGAUGAG X CGAA ACUUGGCC 2707
773 AAGTCTGT A CAACATCT 199 AGAUGUUG CUGAUGAG X CGAA ACAGACUU 2708
780 TACAACAT C TTGAGTCC 200 GGACUCAA CUGAUGAG X CGAA AUGUUGUA 2709
782 CAACATCT T GAGTCCCT 201 AGGGACUC CUGAUGAG X CGAA AGAUGUUG 2710
787 TCTTGAGT C CCTTTATG 202 CAUAAAGG CUGAUGAG X CGAA ACUCAAGA 2711
791 GAGTCCCT T TATGCCGC 203 GCGGCAUA CUGAUGAG X CGAA AGGGACUC 2712
792 AGTCCCTT T ATGCCGCT 204 AGCGGCAU CUGAUGAG X CGAA AAGGGACU 2713
793 GTCCCTTT A TGCCGCTG 205 CAGCGGCA CUGAUGAG X CGAA AAAGGGAC 2714
803 GCCGCTGT T ACCAATTT 206 AAAUUGGU CUGAUGAG X CGAA ACAGCGGC 2715
804 CCGCTGTT A CCAATTTT 207 AAAAUUGG CUGAUGAG X CGAA AACAGCGG 2716
810 TTACCAAT T TTCTTTTG 208 CAAAAGAA CUGAUGAG X CGAA AUUGGUAA 2717
811 TACCAATT T TCTTTTGT 209 ACAAAAGA CUGAUGAG X CGAA AAUUGGUA 2718
812 ACCAATTT T CTTTTGTC 210 GACAAAAG CUGAUGAG X CGAA AAAUUGGU 2719
813 CCAATTTT C TTTTGTCT 211 AGACAAAA CUGAUGAG X CGAA AAAAUUGG 2720
815 AATTTTCT T TTGTCTTT 212 AAAGACAA CUGAUGAG X CGAA AGAAAAUU 2721
816 ATTTTCTT T TGTCTTTG 213 CAAAGACA CUGAUGAG X CGAA AAGAAAAU 2722
817 TTTTCTTT T GTCTTTGG 214 CCAAAGAC CUGAUGAG X CGAA AAAGAAAA 2723
820 TCTTTTGT c TTTGGGTA 215 UACCCAAA CUGAUGAG X CGAA ACAAAAGA 2724
822 TTTTGTCT T TGGGTATA 216 UAUACCCA CUGAUGAG X CGAA AGACAAAA 2725
823 TTTGTCTT T GGGTATAC 217 GUAUACCC CUGAUGAG X CGAA AAGACAAA 2726
828 CTTTGGGT A TACATTTA 218 UAAAUGUA CUGAUGAG X CGAA ACCCAAAG 2727
830 TTGGGTAT A CATTTAAA 219 UUUAAAUG CUGAUGAG X CGAA AUACCCAA 2728
834 GTATACAT T TAAACCCT 220 AGGGUUUA CUGAUGAG X CGAA AUGUAUAC 2729
835 TATACATT T AAACCCTC 221 GAGGGUUU CUGAUGAG X CGAA AAUGUAUA 2730
836 ATACATTT A AACCCTCA 222 UGAGGGUU CUGAUGAG X CGAA AAAUGUAU 2731
843 TAAACCCT C ACAAAACA 223 UGUUUUGU CUGAUGAG X CGAA AGGGUUUA 2732
865 ATGGGGAT A TTCCCTTA 224 UAAGGGAA CUGAUGAG X CGAA AUCCCCAU 2733
867 GGGGATAT T CCCTTAAC 225 GUUAAGGG CUGAUGAG X CGAA AUAUCCCC 2734
868 GGGATATT C CCTTAACT 226 AGUUAAGG CUGAUGAG X CGAA AAUAUCCC 2735
872 TATTCCCT T AACTTCAT 227 AUGAAGUU CUGAUGAG X CGAA AGGGAAUA 2736
873 ATTCCCTT A ACTTCATG 228 CAUGAAGU CUGAUGAG X CGAA AAGGGAAU 2737
877 CCTTAACT T CATGGGAT 229 AUCCCAUG CUGAUGAG X CGAA AGUUAAGG 2738
878 CTTAACTT C ATGGGATA 230 UAUCCCAU CUGAUGAG X CGAA AAGUUAAG 2739
886 CATGGGAT A TGTAATTG 231 CAAUUACA CUGAUGAG X CGAA AUCCCAUG 2740
890 GGATATGT A ATTGGGAG 232 CUCCCAAU CUGAUGAG X CGAA ACAUAUCC 2741
893 TATGTAAT T GGGAGTTG 233 CAACUCCC CUGAUGAG X CGAA AUUACAUA 2742
900 TTGGGAGT T GGGGCACA 234 UGUGCCCC CUGAUGAG X CGAA ACUCCCAA 2743
Table 5/
910 GGGCACAT T GCCACAGG 235 CCUGUGGC CUGAUGAG X CGAA AUGUGCCC 2744
924 AGGAACAT A TTGTACAA 236 UUGUACAA CUGAUGAG X CGAA AUGUUCCU 2745
926 GAACATAT T GTACAAAA 237 UUUUGUAC CUGAUGAG X CGAA AUAUGUUC 2746
929 CATATTGT A CAAAAAAT 238 AUUUUUUG CUGAUGAG X CGAA ACAAUAUG 2747
938 CAAAAAAT C AAAATGTG 239 CACAUUUU CUGAUGAG X CGAA AUUUUUUG 2748
948 AAATGTGT T TTAGGAAA 240 UUUCCUAA CUGAUGAG X CGAA ACACAUUU 2749
949 AATGTGTT T TAGGAAAC 241 GUUUCCUA CUGAUGAG X CGAA AACACAUU 2750
950 ATGTGTTT T AGGAAACT 242 AGUUUCCU CUGAUGAG X CGAA AAACACAU 2751
951 TGTGTTTT A GGAAACTT 243 AAGUUUCC CUGAUGAG X CGAA AAAACACA 2752
959 AGGAAACT T CCTGTAAA 244 UUUACAGG CUGAUGAG X CGAA AGUUUCCU 2753
960 GGAAACTT C CTGTAAAC 245 GUUUACAG CUGAUGAG X CGAA AAGUUUCC 2754
965 CTTCCTGT A AACAGGCC 246 GGCCUGUU CUGAUGAG X CGAA ACAGGAAG 2755
975 ACAGGCCT A TTGATTGG 247 CCAAUCAA CUGAUGAG X CGAA AGGCCUGU 2756
977 AGGCCTAT T GATTGGAA 248 UUCCAAUC CUGAUGAG X CGAA AUAGGCCU 2757
981 CTATTGAT T GGAAAGTA 249 UACUUUCC CUGAUGAG X CGAA AUCAAUAG 2758
989 TGGAAAGT A TGTCAACG 250 CGUUGACA CUGAUGAG X CGAA ACUUUCCA 2759
993 AAGTATGT C AACGAATT 251 AAUUCGUU CUGAUGAG X CGAA ACAUACUU 2760
1001 CAACGAAT T GTGGGTCT 252 AGACCCAC CUGAUGAG X CGAA AUUCGUUG 2761
1008 TTGTGGGT C TTTTGGGG 253 CCCCAAAA CUGAUGAG X CGAA ACCCACAA 2762
1010 GTGGGTCT T TTGGGGTT 254 AACCCCAA CUGAUGAG X CGAA AGACCCAC 2763
1011 TGGGTCTT T TGGGGTTT 255 AAACCCCA CUGAUGAG X CGAA AAGACCCA 2764
1012 GGGTCTTT T GGGGTTTG 256 CAAACCCC CUGAUGAG X CGAA AAAGACCC 2765
1018 TTTGGGGT T TGCCGCCC 257 GGGCGGCA CUGAUGAG X CGAA ACCCCAAA 2766
1019 TTGGGGTT T GCCGCCCC 258 GGGGCGGC CUGAUGAG X CGAA AACCCCAA 2767
1029 CCGCCCCT T TCACGCAA 259 UUGCGUGA CUGAUGAG X CGAA AGGGGCGG 2768
1030 CGCCCCTT T CACGCAAT 260 AUUGCGUG CUGAUGAG X CGAA AAGGGGCG 2769
1031 GCCCCTTT c ACGCAATG 261' CAUUGCGU CUGAUGAG X CGAA AAAGGGGC 2770
1045 ATGTGGAT A TTCTGCTT 262 AAGCAGAA CUGAUGAG X CGAA AUCCACAU 2771
1047 GTGGATAT T CTGCTTTA 263 UAAAGCAG CUGAUGAG X CGAA AUAUCCAC 2772
1048 TGGATATT C TGCTTTAA 264 UUAAAGCA CUGAUGAG X CGAA AAUAUCCA 2773
1053 ATTCTGCT T TAATGCCT 265 AGGCAUUA CUGAUGAG X CGAA AGCAGAAU 2774
1054 TTCTGCTT T AATGCCTT 266 AAGGCAUU CUGAUGAG X CGAA AAGCAGAA 2775
1055 TCTGCTTT A ATGCCTTT 267 AAAGGCAU CUGAUGAG X CGAA AAAGCAGA 2776
1062 TAATGCCT T TATATGCA 268 UGCAUAUA CUGAUGAG X CGAA AGGCAUUA 2777
1063 AATGCCTT T ATATGCAT 269 AUGCAUAU CUGAUGAG X CGAA AAGGCAUU 2778
1064 ATGCCTTT A TATGCATG 270 CAUGCAUA CUGAUGAG X CGAA AAAGGCAU 2779
1066 GCCTTTAT A TGCATGCA 271 UGCAUGCA CUGAUGAG X CGAA AUAAAGGC 2780
1076 GCATGCAT A CAAGCAAA 272 UUUGCUUG CUGAUGAG X CGAA AUGCAUGC 2781
1092 AACAGGCT T TTACTTTC 273 GAAAGUAA CUGAUGAG X CGAA AGCCUGUU 2782
1093 ACAGGCTT T TACTTTCT 274 AGAAAGUA CUGAUGAG X CGAA AAGCCUGU 2783
1094 CAGGCTTT T ACTTTCTC 275 GAGAAAGU CUGAUGAG X CGAA AAAGCCUG 2784
1095 AGGCTTTT A CTTTCTCG 276 CGAGAAAG CUGAUGAG X CGAA AAAAGCCU 2785
1098 CTTTTACT T TCTCGCCA 277 UGGCGAGA CUGAUGAG X CGAA AGUAAAAG 2786
1099 TTTTACTT T CTCGCCAA 278 UUGGCGAG CUGAUGAG X CGAA AAGUAAAA 2787
1100 TTTACTTT C TCGCCAAC 279 GUUGGCGA CUGAUGAG X CGAA AAAGUAAA 2788
1102 TACTTTCT C GCCAACTT 280 AAGUUGGC CUGAUGAG X CGAA AGAAAGUA 2789
1110 CGCCAACT T ACAAGGCC 281 GGCCUUGU CUGAUGAG X CGAA AGUUGGCG 2790 llll GCCAACTT A CAAGGCCT 282 AGGCCUUG CUGAUGAG X CGAA AAGUUGGC 2791
1120 CAAGGCCT T TCTAAGTA 283 UACUUAGA CUGAUGAG X CGAA AGGCCUUG 2792
1121 AAGGCCTT T CTAAGTAA 284 UUACUUAG CUGAUGAG X CGAA AAGGCCUU 2793
1122 AGGCCTTT C TAAGTAAA 285 UUUACUUA CUGAUGAG X CGAA AAAGGCCU 2794
Table 37
1124 GCCTTTCT A AGTAAACA 286 UGUUUACU CUGAUGAG X CGAA AGAAAGGC 2795
1128 TTCTAAGT A AACAGTAT 287 AUACUGUU CUGAUGAG X CGAA ACUUAGAA 2796
1135 TAAACAGT A TGTGAACC 288 GGUUCACA CUGAUGAG X CGAA ACUGUUUA 2797
1145 GTGAACCT T TACCCCGT 289 ACGGGGUA CUGAUGAG X CGAA AGGUUCAC 2798
1146 TGAACCTT T ACCCCGTT 290 AACGGGGU CUGAUGAG X CGAA AAGGUUCA 2799
1147 GAACCTTT A CCCCGTTG 291 CAACGGGG CUGAUGAG X CGAA AAAGGUUC 2800
1154 TACCCCGT T GCTCGGCA 292 UGCCGAGC CUGAUGAG X CGAA ACGGGGUA 2801
1158 CCGTTGCT C GGCAACGG 293 CCGUUGCC CUGAUGAG X CGAA AGCAACGG 2802
1173 GGCCTGGT C TATGCCAA 294 UUGGCAUA CUGAUGAG X CGAA ACCAGGCC 2803
1175 CCTGGTCT A TGCCAAGT 295 ACUUGGCA CUGAUGAG X CGAA AGACCAGG 2804
1186 CCAAGTGT T TGCTGACG 296 CGUCAGCA CUGAUGAG X CGAA ACACUUGG 2805
1187 CAAGTGTT T GCTGACGC 297 GCGUCAGC CUGAUGAG X CGAA AACACUUG 2806
1209 CCACTGGT T GGGGCTTG 298 CAAGCCCC CUGAUGAG X CGAA ACCAGUGG 2807
1216 TTGGGGCT T GGCCATAG 299 CUAUGGCC CUGAUGAG X CGAA AGCCCCAA 2808
1223 TTGGCCAT A GGCCATCA 300 UGAUGGCC CUGAUGAG X CGAA AUGGCCAA 2809
1230 TAGGCCAT C AGCGCATG 301 CAUGCGCU CUGAUGAG X CGAA AUGGCCUA 2810
1249 TGGAACCT T TGTGTCTC 302 GAGACACA CUGAUGAG X CGAA AGGUUCCA 2811
1250 GGAACCTT T GTGTCTCC 303 GGAGACAC CUGAUGAG X CGAA AAGGUUCC 2812
1255 CTTTGTGT C TCCTCTGC 304 GCAGAGGA CUGAUGAG X CGAA ACACAAAG 2813
1257 TTGTGTCT C CTCTGCCG 305 CGGCAGAG CUGAUGAG X CGAA AGACACAA 2814
1260 TGTCTCCT C TGCCGATC 306 GAUCGGCA CUGAUGAG X CGAA AGGAGACA 2815
1268 CTGCCGAT C CATACCGC 307 GCGGUAUG CUGAUGAG X CGAA AUCGGCAG 2816
1272 CGATCCAT A CCGCGGAA 308 UUCCGCGG CUGAUGAG X CGAA AUGGAUCG 2817
1283 GCGGAACT C CTAGCCGC 309 GCGGCUAG CUGAUGAG X CGAA AGUUCCGC 2818
1286 GAACTCCT A GCCGCTTG 310 CAAGCGGC CUGAUGAG X CGAA AGGAGUUC 2819
1293 TAGCCGCT T GTTTTGCT 311 AGCAAAAC CUGAUGAG X CGAA AGCGGCUA 2820
1296 CCGCTTGT T TTGCTCGC 312 GCGAGCAA CUGAUGAG X CGAA ACAAGCGG 2821
1297 CGCTTGTT T TGCTCGCA 313 UGCGAGCA CUGAUGAG X CGAA AACAAGCG 2822
1298 GCTTGTTT T GCTCGCAG 314 CUGCGAGC CUGAUGAG X CGAA AAACAAGC 2823
1302 GTTTTGCT C GCAGCAGG 315 CCUGCUGC CUGAUGAG X CGAA AGCAAAAC 2824
1312 CAGCAGGT C TGGGGCAA 316 UUGCCCCA CUGAUGAG X CGAA ACCUGCUG 2825
1325 GCAAAACT C ATCGGGAC 317 GUCCCGAU CUGAUGAG X CGAA AGUUUUGC 2826
1328 AAACTCAT C GGGACTGA 318 UCAGUCCC CUGAUGAG X CGAA AUGAGUUU 2827
1341 CTGACAAT T CTGTCGTG 319 CACGACAG CUGAUGAG X CGAA AUUGUCAG 2828
1342 TGACAATT c TGTCGTGC 320 GCACGACA CUGAUGAG X CGAA AAUUGUCA 2829
1346 AATTCTGT c GTGCTCTC 321 GAGAGCAC CUGAUGAG X CGAA ACAGAAUU 2830
1352 GTCGTGCT c TCCCGCAA 322 UUGCGGGA CUGAUGAG X CGAA AGCACGAC 2831
1354 CGTGCTCT c CCGCAAAT 323 AUUUGCGG CUGAUGAG X CGAA AGAGCACG 2832
1363 CCGCAAAT A TACATCAT 324 AUGAUGUA CUGAUGAG X CGAA AUUUGCGG 2833
1365 GCAAATAT A CATCATTT 325 AAAUGAUG CUGAUGAG X CGAA AUAUUUGC 2834
1369 ATATACAT C ATTTCCAT 326 AUGGAAAU CUGAUGAG X CGAA AUGUAUAU 2835
1372 TACATCAT T TCCATGGC 327 GCCAUGGA CUGAUGAG X CGAA AUGAUGUA 2836
1373 ACATCATT T CCATGGCT 328 AGCCAUGG CUGAUGAG X CGAA AAUGAUGU 2837
1374 CATCATTT c CATGGCTG 329 CAGCCAUG CUGAUGAG X CGAA AAAUGAUG 2838
1385 TGGCTGCT A GGCTGTGC 330 GCACAGCC CUGAUGAG X CGAA AGCAGCCA 2839
1406 AACTGGAT C CTACGCGG 331 CCGCGUAG CUGAUGAG X CGAA AUCCAGUU 2840
1409 TGGATCCT A CGCGGGAC 332 GUCCCGCG CUGAUGAG X CGAA AGGAUCCA 2841
1420 CGGGACGT C CTTTGTTT 333 AAACAAAG CUGAUGAG X CGAA ACGUCCCG 2842
1423 GACGTCCT T TGTTTACG 334 CGUAAACA CUGAUGAG X CGAA AGGACGUC 2843
1424 ACGTCCTT T GTTTACGT 335 ACGUAAAC CUGAUGAG X CGAA AAGGACGU 2844
1427 TCCTTTGT T TACGTCCC 336 GGGACGUA CUGAUGAG X CGAA ACAAAGGA 2845
Table 37
1428 CCTTTGTT T ACGTCCCG 337 CGGGACGU CUGAUGAG X CGAA AACAAAGG 2846
1429 CTTTGTTT A CGTCCCGT 338 ACGGGACG CUGAUGAG X CGAA AAACAAAG 2847
1433 GTTTACGT C CCGTCGGC 339 GCCGACGG CUGAUGAG X CGAA ACGUAAAC 2848
1438 CGTCCCGT C GGCGCTGA 340 UCAGCGCC CUGAUGAG X CGAA ACGGGACG 2849
1449 CGCTGAAT C CCGCGGAC 341 GUCCGCGG CUGAUGAG X CGAA AUUCAGCG 2850
1465 CGACCCCT C CCGGGGCC 342 GGCCCCGG CUGAUGAG X CGAA AGGGGUCG 2851
1477 GGGCCGCT T GGGGCTCT 343 AGAGCCCC CUGAUGAG X CGAA AGCGGCCC 2852
1484 TTGGGGCT C TACCGCCC 344 GGGCGGUA CUGAUGAG X CGAA AGCCCCAA 2853
1486 GGGGCTCT A CCGCCCGC 345 GCGGGCGG CUGAUGAG X CGAA AGAGCCCC 2854
1496 CGCCCGCT T CTCCGCCT 346 AGGCGGAG CUGAUGAG X CGAA AGCGGGCG 2855
1497 GCCCGCTT c TCCGCCTA 347 UAGGCGGA CUGAUGAG X CGAA AAGCGGGC 2856
1499 CCGCTTCT c CGCCTATT 348 AAUAGGCG CUGAUGAG X CGAA AGAAGCGG 2857
1505 CTCCGCCT A TTGTACCG 349 CGGUACAA CUGAUGAG X CGAA AGGCGGAG 2858
1507 CCGCCTAT T GTACCGAC 350 GUCGGUAC CUGAUGAG X CGAA AUAGGCGG 2859
1510 CCTATTGT A CCGACCGT 351 ACGGUCGG CUGAUGAG X CGAA ACAAUAGG 2860
1519 CCGACCGT C CACGGGGC 352 GCCCCGUG CUGAUGAG X CGAA ACGGUCGG 2861
1534 GCGCACCT C TCTTTACG 353 CGUAAAGA CUGAUGAG X CGAA AGGUGCGC 2862
1536 GCACCTCT C TTTACGCG 354 CGCGUAAA CUGAUGAG X CGAA AGAGGUGC 2863
1538 ACCTCTCT T TACGCGGA 355 UCCGCGUA CUGAUGAG X CGAA AGAGAGGU 2864
1539 CCTCTCTT T ACGCGGAC 356 GUCCGCGU CUGAUGAG X CGAA AAGAGAGG 2865
1540 CTCTCTTT A CGCGGACT 357 AGUCCGCG CUGAUGAG X CGAA AAAGAGAG 2866
1549 CGCGGACT C CCCGTCTG 358 CAGACGGG CUGAUGAG X CGAA AGUCCGCG 2867
1555 CTCCCCGT C TGTGCCTT 359 AAGGCACA CUGAUGAG X CGAA ACGGGGAG 2868
1563 CTGTGCCT T CTCATCTG 360 CAGAUGAG CUGAUGAG X CGAA AGGCACAG 2869
1564 TGTGCCTT c TCATCTGC 361 GCAGAUGA CUGAUGAG X CGAA AAGGCACA 2870
1566 TGCCTTCT c ATCTGCCG 362 CGGCAGAU CUGAUGAG .X CGAA AGAAGGCA 2871
1569 CTTCTCAT c TGCCGGAC 363 GUCCGGCA CUGAUGAG X CGAA AUGAGAAG 2872
1588 TGTGCACT T CGCTTCAC 364 GUGAAGCG CUGAUGAG X CGAA AGUGCACA 2873
1589 GTGCACTT c GCTTCACC 365 GGUGAAGC CUGAUGAG X CGAA AAGUGCAC 2874
1593 ACTTCGCT T CACCTCTG 366 CAGAGGUG CUGAUGAG X CGAA AGCGAAGU 2875
1594 CTTCGCTT c ACCTCTGC 367 GCAGAGGU CUGAUGAG X CGAA AAGCGAAG 2876
1599 CTTCACCT c TGCACGTC 368 GACGUGCA CUGAUGAG X CGAA AGGUGAAG 2877
1607 CTGCACGT c GCATGGAG 369 CUCCAUGC CUGAUGAG X CGAA ACGUGCAG 2878
1651 CCCAAGGT c TTGCATAA 370 UUAUGCAA CUGAUGAG X CGAA ACCUUGGG 2879
1653 CAAGGTCT T GCATAAGA 371 UCUUAUGC CUGAUGAG X CGAA AGACCUUG 2880
1658 TCTTGCAT A AGAGGACT 372 AGUCCUCU CUGAUGAG X CGAA AUGCAAGA 2881
1667 AGAGGACT c TTGGACTT 373 AAGUCCAA CUGAUGAG X CGAA AGUCCUCU 2882
1669 AGGACTCT T GGACTTTC 374 GAAAGUCC CUGAUGAG X CGAA AGAGUCCU 2883
1675 CTTGGACT T TCAGCAAT 375 AUUGCUGA CUGAUGAG X CGAA AGUCCAAG 2884
1676 TTGGACTT T CAGCAATG 376 CAUUGCUG CUGAUGAG X CGAA AAGUCCAA 2885
1677 TGGACTTT c AGCAATGT 377 ACAUUGCU CUGAUGAG X CGAA AAAGUCCA 2886
1686 AGCAATGT c AACGACCG 378 CGGUCGUU CUGAUGAG X CGAA ACAUUGCU 2887
1699 ACCGACCT T GAGGCATA 379 UAUGCCUC CUGAUGAG X CGAA AGGUCGGU 2888
1707 TGAGGCAT A CTTCAAAG 380 CUUUGAAG CUGAUGAG X CGAA AUGCCUCA 2889
1710 GGCATACT T CAAAGACT 381 AGUCUUUG CUGAUGAG X CGAA AGUAUGCC 2890
1711 GCATACTT c AAAGACTG 382 CAGUCUUU CUGAUGAG X CGAA AAGUAUGC 2891
1725 CTGTGTGT T TAATGAGT 383 ACUCAUUA CUGAUGAG X CGAA ACACACAG 2892
1726 TGTGTGTT T AATGAGTG 384 CACUCAUU CUGAUGAG X CGAA AACACACA 2893
1727 GTGTGTTT A ATGAGTGG 385 CCACUCAU CUGAUGAG X CGAA AAACACAC 2894
1743 GGAGGAGT T GGGGGAGG 386 CCUCCCCC CUGAUGAG X CGAA ACUCCUCC 2895
1756 GAGGAGGT T AGGTTAAA 387 UUUAACCU CUGAUGAG X CGAA ACCUCCUC 2896
Table 37
1757 AGGAGGTT A GGTTAAAG 388 CUUUAACC CUGAUGAG X CGAA AACCUCCU 2897
1761 GGTTAGGT T AAAGGTCT 389 AGACCUUU CUGAUGAG X CGAA ACCUAACC 2898
1762 GTTAGGTT A AAGGTCTT 390 AAGACCUU CUGAUGAG X CGAA AACCUAAC 2899
1768 TTAAAGGT C TTTGTACT 391 AGUACAAA CUGAUGAG X CGAA ACCUUUAA 2900
1770 AAAGGTCT T TGTACTAG 392 CUAGUACA CUGAUGAG X CGAA AGACCUUU 2901
1771 AAGGTCTT T GTACTAGG 393 CCUAGUAC CUGAUGAG X CGAA AAGACCUU 2902
1774 GTCTTTGT A CTAGGAGG 394 CCUCCUAG CUGAUGAG X CGAA ACAAAGAC 2903
1777 TTTGTACT A GGAGGCTG 395 CAGCCUCC CUGAUGAG X CGAA AGUACAAA 2904
1787 GAGGCTGT A GGCATAAA 396 UUUAUGCC CUGAUGAG X CGAA ACAGCCUC 2905
1793 GTAGGCAT A AATTGGTG 397 CACCAAUU CUGAUGAG X CGAA AUGCCUAC 2906
1797 GCATAAAT T GGTGTGTT 398 AACACACC CUGAUGAG X CGAA AUUUAUGC 2907
1805 TGGTGTGT T CACCAGCA 399 UGCUGGUG CUGAUGAG X CGAA ACACACCA 2908
1806 GGTGTGTT C ACCAGCAC 400 GUGCUGGU CUGAUGAG X CGAA AACACACC 2909
1824 ATGCAACT T TTTCACCT 401 AGGUGAAA CUGAUGAG X CGAA AGUUGCAU 2910
1825 TGCAACTT T TTCACCTC 402 GAGGUGAA CUGAUGAG X CGAA AAGUUGCA 2911
1826 GCAACTTT T TCACCTCT 403 AGAGGUGA CUGAUGAG X CGAA AAAGUUGC 2912
1827 CAACTTTT T CACCTCTG 404 CAGAGGUG CUGAUGAG X CGAA AAAAGUUG 2913
1828 AACTTTTT C ACCTCTGC 405 GCAGAGGU CUGAUGAG X CGAA AAAAAGUU 2914
1833 TTTCACCT C TGCCTAAT 406 AUUAGGCA CUGAUGAG X CGAA AGGUGAAA 2915
1839 CTCTGCCT A ATCATCTC 407 GAGAUGAU CUGAUGAG X CGAA AGGCAGAG 2916
1842 TGCCTAAT C ATCTCATG 408 CAUGAGAU CUGAUGAG X CGAA AUUAGGCA 2917
1845 CTAATCAT C TCATGTTC 409 GAACAUGA CUGAUGAG X CGAA AUGAUUAG 2918
1847 AATCATCT C ATGTTCAT 410 AUGAACAU CUGAUGAG X CGAA AGAUGAUU 2919
1852 TCTCATGT T CATGTCCT 411 AGGACAUG CUGAUGAG X CGAA ACAUGAGA 2920
1853 CTCATGTT C ATGTCCTA 412 UAGGACAU CUGAUGAG X CGAA AACAUGAG 2921
1858 GTTCATGT C CTACTGTT 413 AACAGUAG CUGAUGAG X CGAA ACAUGAAC 2922
1861 CATGTCCT A CTGTTCAA 414 UUGAACAG CUGAUGAG X CGAA AGGACAUG 2923
1866 CCTACTGT T CAAGCCTC 415 GAGGCUUG CUGAUGAG X CGAA ACAGUAGG 2924
1867 CTACTGTT C AAGCCTCC 416 GGAGGCUU CUGAUGAG X CGAA AACAGUAG 2925
1874 TCAAGCCT C CAAGCTGT 417 ACAGCUUG CUGAUGAG X CGAA AGGCUUGA 2926
1887 CTGTGCCT T GGGTGGCT 418 AGCCACCC CUGAUGAG X CGAA AGGCACAG 2927
1896 GGGTGGCT T TGGGGCAT 419 AUGCCCCA CUGAUGAG X CGAA AGCCACCC 2928
1897 GGTGGCTT T GGGGCATG 420 CAUGCCCC CUGAUGAG X CGAA AAGCCACC 2929
1911 ATGGACAT T GACCCGTA 421 UACGGGUC CUGAUGAG X CGAA AUGUCCAU 2930
1919 TGACCCGT A TAAAGAAT 422 AUUCUUUA CUGAUGAG X CGAA ACGGGUCA 2931
1921 ACCCGTAT A AAGAATTT 423 AAAUUCUU CUGAUGAG X CGAA AUACGGGU 2932
1928 TAAAGAAT T TGGAGCTT 424 AAGCUCCA CUGAUGAG X CGAA AUUCUUUA 2933
1929 AAAGAATT T GGAGCTTC 425 GAAGCUCC CUGAUGAG X CGAA AAUUCUUU 2934
1936 TTGGAGCT T CTGTGGAG 426 CUCCACAG CUGAUGAG X CGAA AGCUCCAA 2935
1937 TGGAGCTT c TGTGGAGT 427 ACUCCACA CUGAUGAG X CGAA AAGCUCCA 2936
1946 TGTGGAGT T ACTCTCTT 428 AAGAGAGU CUGAUGAG X CGAA ACUCCACA 2937
1947 GTGGAGTT A CTCTCTTT 429 AAAGAGAG CUGAUGAG X CGAA AACUCCAC 2938
1950 GAGTTACT C TCTTTTTT 430 AAAAAAGA CUGAUGAG X CGAA AGUAACUC 2939
1952 GTTACTCT C TTTTTTGC 431 GCAAAAAA CUGAUGAG X CGAA AGAGUAAC 2940
1954 TACTCTCT T TTTTGCCT 432 AGGCAAAA CUGAUGAG X CGAA AGAGAGUA 2941
1955 ACTCTCTT T TTTGCCTT 433 AAGGCAAA CUGAUGAG X CGAA AAGAGAGU 2942
1956 CTCTCTTT T TTGCCTTC 434 GAAGGCAA CUGAUGAG X CGAA AAAGAGAG 2943
1957 TCTCTTTT T TGCCTTCT 435 AGAAGGCA CUGAUGAG X CGAA AAAAGAGA 2944
1958 CTCTTTTT T GCCTTCTG 436 CAGAAGGC CUGAUGAG X CGAA AAAAAGAG 2945
1963 TTTTGCCT T CTGACTTC 437 GAAGUCAG CUGAUGAG X CGAA AGGCAAAA 2946
1964 TTTGCCTT c TGACTTCT 438 AGAAGUCA CUGAUGAG X CGAA AAGGCAAA 2947
Table 37
1970 TTCTGACT T CTTTCCTT 439 AAGGAAAG CUGAUGAG X CGAA AGUCAGAA 2948
1971 TCTGACTT C TTTCCTTC 440 GAAGGAAA CUGAUGAG X CGAA AAGUCAGA 2949
1973 TGACTTCT T TCCTTCTA 441 UAGAAGGA CUGAUGAG X CGAA AGAAGUCA 2950
1974 GACTTCTT T CCTTCTAT 442 AUAGAAGG CUGAUGAG-X CGAA AAGAAGUC 2951
1975 ACTTCTTT c CTTCTATT 443 AAUAGAAG CUGAUGAG X CGAA AAAGAAGU 2952
1978 TCTTTCCT T CTATTCGA 444 UCGAAUAG CUGAUGAG X CGAA AGGAAAGA 2953
1979 CTTTCCTT c TATTCGAG 445 CUCGAAUA CUGAUGAG X CGAA AAGGAAAG 2954
1981 TTCCTTCT A TTCGAGAT 446 AUCUCGAA CUGAUGAG X CGAA AGAAGGAA 2955
1983 CCTTCTAT T CGAGATCT 447 AGAUCUCG CUGAUGAG X CGAA AUAGAAGG 2956
1984 CTTCTATT C GAGATCTC 448 GAGAUCUC CUGAUGAG X CGAA AAUAGAAG 2957
1990 TTCGAGAT C TCCTCGAC 449 GUCGAGGA CUGAUGAG X CGAA AUCUCGAA 2958
1992 CGAGATCT C CTCGACAC 450 GUGUCGAG CUGAUGAG X CGAA AGAUCUCG 2959
1995 GATCTCCT c GACACCGC 451 GCGGUGUC CUGAUGAG X CGAA AGGAGAUC 2960
2006 CACCGCCT c TGCTCTGT 452 ACAGAGCA CUGAUGAG X CGAA AGGCGGUG 2961
2011 CCTCTGCT c TGTATCGG 453 CCGAUACA CUGAUGAG X CGAA AGCAGAGG 2962
2015 TGCTCTGT A TCGGGGGG 454 CCCCCCGA CUGAUGAG X CGAA ACAGAGCA 2963
2017 CTCTGTAT C GGGGGGCC 455 GGCCCCCC CUGAUGAG X CGAA AUACAGAG 2964
2027 GGGGGCCT T AGAGTCTC 456 GAGACUCU CUGAUGAG X CGAA AGGCCCCC 2965
2028 GGGGCCTT A GAGTCTCC 457 GGAGACUC CUGAUGAG X CGAA AAGGCCCC 2966
2033 CTTAGAGT C TCCGGAAC 458 GUUCCGGA CUGAUGAG X CGAA ACUCUAAG 2967
2035 TAGAGTCT C CGGAACAT 459 AUGUUCCG CUGAUGAG X CGAA AGACUCUA 2968
2044 CGGAACAT T GTTCACCT 460 AGGUGAAC CUGAUGAG X CGAA AUGUUCCG 2969
2047 AACATTGT T CACCTCAC 461 GUGAGGUG CUGAUGAG X CGAA ACAAUGUU 2970
2048 ACATTGTT C ACCTCACC 462 GGUGAGGU CUGAUGAG X CGAA AACAAUGU 2971
2053 GTTCACCT C ACCATACG 463 CGUAUGGU CUGAUGAG X CGAA AGGUGAAC 2972
2059 CTCACCAT A CGGCACTC 464 GAGUGCCG CUGAUGAG X CGAA AUGGUGAG 2973
2067 ACGGCACT C AGGCAAGC 465 GCUUGCCU CUGAUGAG X CGAA AGUGCCGU 2974
2077 GGCAAGCT A TTCTGTGT 466 ACACAGAA CUGAUGAG X CGAA AGCUUGCC 2975
2079 CAAGCTAT T CTGTGTTG 467 CAACACAG CUGAUGAG X CGAA AUAGCUUG 2976
2080 AAGCTATT C TGTGTTGG 468 CCAACACA CUGAUGAG X CGAA AAUAGCUU 2977
2086 TTCTGTGT T GGGGTGAG 469 j CUCACCCC CUGAUGAG X CGAA ACACAGAA 2978
2096 GGGTGAGT T GATGAATC 470 GAUUCAUC CUGAUGAG X CGAA ACUCACCC 2979
2104 TGATGAAT C TAGCCACC 471 GGUGGCUA CUGAUGAG X CGAA AUUCAUCA 2980
2106 ATGAATCT A GCCACCTG 472 CAGGUGGC CUGAUGAG X CGAA AGAUUCAU 2981
2125 TGGGAAGT A ATTTGGAA 473 UUCCAAAU CUGAUGAG X CGAA ACUUCCCA 2982
2128 GAAGTAAT T TGGAAGAT 474 AUCUUCCA CUGAUGAG X CGAA AUUACUUC 2983
2129 AAGTAATT T GGAAGATC 475 GAUCUUCC CUGAUGAG X CGAA AAUUACUU 2984
2137 TGGAAGAT C CAGCATCC 476 GGAUGCUG CUGAUGAG X CGAA AUCUUCCA 2985
2144 TCCAGCAT C CAGGGAAT 477 AUUCCCUG CUGAUGAG X CGAA AUGCUGGA 2986
2153 CAGGGAAT T AGTAGTCA 478 UGACUACU CUGAUGAG X CGAA AUUCCCUG 2987
2154 AGGGAATT A GTAGTCAG 479 CUGACUAC CUGAUGAG X CGAA AAUUCCCU 2988
2157 GAATTAGT A GTCAGCTA 480 UAGCUGAC CUGAUGAG X CGAA ACUAAUUC 2989
2160 TTAGTAGT C AGCTATGT 481 ACAUAGCU CUGAUGAG X CGAA ACUACUAA 2990
2165 AGTCAGCT A TGTCAACG 482 CGUUGACA CUGAUGAG X CGAA AGCUGACU 2991
2169 AGCTATGT C AACGTTAA 483 UUAACGUU CUGAUGAG X CGAA ACAUAGCU 2992
2175 GTCAACGT T AATATGGG 484 CCCAUAUU CUGAUGAG X CGAA ACGUUGAC 2993
2176 TCAACGTT A ATATGGGC 485 GCCCAUAU CUGAUGAG X CGAA AACGUUGA 2994
2179 ACGTTAAT A TGGGCCTA 486 UAGGCCCA CUGAUGAG X CGAA AUUAACGU 2995
2187 ATGGGCCT A AAAATCAG 487 CUGAUUUU CUGAUGAG X CGAA AGGCCCAU 2996
2193 CTAAAAAT C AGACAACT 488 AGUUGUCU CUGAUGAG X CGAA AUUUUUAG 2997
2202 AGACAACT A TTGTGGTT 489 AACCACAA CUGAUGAG X CGAA AGUUGUCU 2998
Tauie __)/
2204 ACAACTAT T GTGGTTTC 490 GAAACCAC CUGAUGAG X CGAA AUAGUUGU 2999
2210 ATTGTGGT T TCACATTT 491 AAAUGUGA CUGAUGAG X CGAA ACCACAAU 3000
2211 TTGTGGTT T CACATTTC 492 GAAAUGUG CUGAUGAG X CGAA AACCACAA 3001
2212 TGTGGTTT C ACATTTCC 493 GGAAAUGU CUGAUGAG X CGAA AAACCACA 3002
2217 TTTCACAT T TCCTGTCT 494 AGACAGGA CUGAUGAG X CGAA AUGUGAAA 3003
2218 TTCACATT T CCTGTCTT 495 AAGACAGG CUGAUGAG X CGAA AAUGUGAA 3004
2219 TCACATTT C CTGTCTTA 496 UAAGACAG CUGAUGAG X CGAA AAAUGUGA 3005
2224 TTTCCTGT C TTACTTTT 497 AAAAGUAA CUGAUGAG X CGAA ACAGGAAA 3006
2226 TCCTGTCT T ACTTTTGG 498 CCAAAAGU CUGAUGAG X CGAA AGACAGGA 3007
2227 CCTGTCTT A CTTTTGGG 499 CCCAAAAG CUGAUGAG X CGAA AAGACAGG 3008
2230 GTCTTACT T TTGGGCGA 500 UCGCCCAA CUGAUGAG X CGAA AGUAAGAC 3009
2231 TCTTACTT T TGGGCGAG 501 CUCGCCCA CUGAUGAG X CGAA AAGUAAGA 3010
2232 CTTACTTT T GGGCGAGA 502 UCUCGCCC CUGAUGAG X CGAA AAAGUAAG 3011
2247 GAAACTGT T CTTGAATA 503 UAUUCAAG CUGAUGAG X CGAA ACAGUUUC 3012
2248 AAACTGTT C TTGAATAT 504 AUAUUCAA CUGAUGAG X CGAA AACAGUUU 3013
2250 ACTGTTCT T GAATATTT 505 AAAUAUUC CUGAUGAG X CGAA AGAACAGU 3014
2255 TCTTGAAT A TTTGGTGT 506 ACACCAAA CUGAUGAG X CGAA AUUCAAGA 3015
2257 TTGAATAT T TGGTGTCT 507 AGACACCA CUGAUGAG X CGAA AUAUUCAA 3016
2258 TGAATATT T GGTGTCTT 508 AAGACACC CUGAUGAG X CGAA AAUAUUCA 3017
2264 TTTGGTGT C TTTTGGAG 509 CUCCAAAA CUGAUGAG X CGAA ACACCAAA 3018
2266 TGGTGTCT T TTGGAGTG 510 CACUCCAA CUGAUGAG X CGAA AGACACCA 3019
2267 GGTGTCTT T TGGAGTGT 511 ACACUCCA CUGAUGAG X CGAA AAGACACC 3020
2268 GTGTCTTT T GGAGTGTG 512 CACACUCC CUGAUGAG X CGAA AAAGACAC 3021
2280 GTGTGGAT T CGCACTCC 513 GGAGUGCG CUGAUGAG X CGAA AUCCACAC 3022
2281 TGTGGATT C GCACTCCT 514 AGGAGUGC CUGAUGAG X CGAA AAUCCACA 3023
2287 TTCGCACT C CTCCTGCA 515 UGCAGGAG CUGAUGAG X CGAA AGUGCGAA 3024 x2290 GCACTCCT c CTGCATAT 516 AUAUGCAG CUGAUGAG X CGAA AGGAGUGC 3025
2297 TCCTGCAT A TAGACCAC 517 GUGGUCUA CUGAUGAG X CGAA AUGCAGGA 3026
2299 CTGCATAT A GACCACCA 518 UGGUGGUC CUGAUGAG X CGAA AUAUGCAG 3027
2317 ATGCCCCT A TCTTATCA 519 UGAUAAGA CUGAUGAG X CGAA AGGGGCAU 3028
2319 GCCCCTAT C TTATCAAC 520 GUUGAUAA CUGAUGAG X CGAA AUAGGGGC 3029
2321 CCCTATCT T ATCAACAC 521 GUGUUGAU CUGAUGAG X CGAA AGAUAGGG 3030
2322 CCTATCTT A TCAACACT 522 AGUGUUGA CUGAUGAG X CGAA AAGAUAGG 3031
2324 TATCTTAT C AACACTTC 523 GAAGUGUU CUGAUGAG X CGAA AUAAGAUA 3032
2331 TCAACACT T CCGGAAAC 524 GUUUCCGG CUGAUGAG X CGAA AGUGUUGA 3033
2332 CAACACTT C CGGAAACT 525 AGUUUCCG CUGAUGAG X CGAA AAGUGUUG 3034
2341 CGGAAACT A CTGTTGTT 526 AACAACAG CUGAUGAG X CGAA AGUUUCCG 3035
2346 ACTACTGT T GTTAGACG 527 CGUCUAAC CUGAUGAG X CGAA ACAGUAGU 3036
2349 ACTGTTGT T AGACGAAG 528 CUUCGUCU CUGAUGAG X CGAA ACAACAGU 3037
2350 CTGTTGTT A GACGAAGA 529 UCUUCGUC CUGAUGAG X CGAA AACAACAG 3038
2366 AGGCAGGT C CCCTAGAA 530 UUCUAGGG CUGAUGAG X CGAA ACCUGCCU 3039
2371 GGTCCCCT A GAAGAAGA 531 UCUUCUUC CUGAUGAG X CGAA AGGGGACC 3040
2383 GAAGAACT C CCTCGCCT 532 AGGCGAGG CUGAUGAG X CGAA AGUUCUUC 3041
2387 AACTCCCT C GCCTCGCA 533 UGCGAGGC CUGAUGAG X CGAA AGGGAGUU 3042
2392 CCTCGCCT C GCAGACGA 534 UCGUCUGC CUGAUGAG X CGAA AGGCGAGG 3043
2405 ACGAAGGT C TCAATCGC 535 GCGAUUGA CUGAUGAG X CGAA ACCUUCGU 3044
2407 GAAGGTCT C AATCGCCG 536 CGGCGAUU CUGAUGAG X CGAA AGACCUUC 3045
2411 GTCTCAAT C GCCGCGTC 537 GACGCGGC CUGAUGAG X CGAA AUUGAGAC 3046
2419 CGCCGCGT C GCAGAAGA 538 UCUUCUGC CUGAUGAG X CGAA ACGCGGCG 3047
2429 CAGAAGAT C TCAATCTC 539 GAGAUUGA CUGAUGAG X CGAA AUCUUCUG 3048
2431 GAAGATCT C AATCTCGG 540 CCGAGAUU CUGAUGAG X CGAA AGAUCUUC 3049
Table 3/
2435 ATCTCAAT C TCGGGAAT 541 AUUCCCGA CUGAUGAG X CGAA AUUGAGAU 3050
2437 CTCAATCT C GGGAATCT 542 AGAUUCCC CUGAUGAG X CGAA AGAUUGAG 3051
2444 TCGGGAAT C TCAATGTT 543 AACAUUGA CUGAUGAG X CGAA AUUCCCGA 3052
2446 GGGAATCT C AATGTTAG 544 CUAACAUU CUGAUGAG X CGAA AGAUUCCC 3053
2452 CTCAATGT T AGTATTCC 545 GGAAUACU CUGAUGAG X CGAA ACAUUGAG 3054
2453 TCAATGTT A GTATTCCT 546 AGGAAUAC CUGAUGAG X CGAA AACAUUGA 3055
2456 ATGTTAGT A TTCCTTGG 547 CCAAGGAA CUGAUGAG X CGAA ACUAACAU 3056
2458 GTTAGTAT T CCTTGGAC 548 GUCCAAGG CUGAUGAG X CGAA AUACUAAC 3057
2459 TTAGTATT C CTTGGACA 549 UGUCCAAG CUGAUGAG X CGAA AAUACUAA 3058
2462 GTATTCCT T GGACACAT 550 AUGUGUCC CUGAUGAG X CGAA AGGAAUAC 3059
2471 GGACACAT A AGGTGGGA 551 UCCCACCU CUGAUGAG X CGAA AUGUGUCC 3060
2484 GGGAAACT T TACGGGGC 552 GCCCCGUA CUGAUGAG X CGAA AGUUUCCC 3061
2485 GGAAACTT T ACGGGGCT 553 AGCCCCGU CUGAUGAG X CGAA AAGUUUCC 3062
2486 GAAACTTT A CGGGGCTT 554 AAGCCCCG CUGAUGAG X CGAA AAAGUUUC 3063
2494 ACGGGGCT T TATTCTTC 555 GAAGAAUA CUGAUGAG X CGAA AGCCCCGU 3064
2495 CGGGGCTT T ATTCTTCT 556 AGAAGAAU CUGAUGAG X CGAA AAGCCCCG 3065
2496 GGGGCTTT A TTCTTCTA 557 UAGAAGAA CUGAUGAG X CGAA AAAGCCCC 3066
2498 GGCTTTAT T CTTCTACG 558 CGUAGAAG CUGAUGAG X CGAA AUAAAGCC 3067
2499 GCTTTATT C TTCTACGG 559 CCGUAGAA CUGAUGAG X CGAA AAUAAAGC 3068
2501 TTTATTCT T CTACGGTA 560 UACCGUAG CUGAUGAG X CGAA AGAAUAAA 3069
2502 TTATTCTT C TACGGTAC 561 GUACCGUA CUGAUGAG X CGAA AAGAAUAA 3070
2504 ATTCTTCT A CGGTACCT 562 AGGUACCG CUGAUGAG X CGAA AGAAGAAU 3071
2509 TCTACGGT A CCTTGCTT 563 AAGCAAGG CUGAUGAG X CGAA ACCGUAGA 3072
2513 CGGTACCT T GCTTTAAT 564 AUUAAAGC CUGAUGAG X CGAA AGGUACCG 3073
2517 ACCTTGCT T TAATCCTA 565 UAGGAUUA CUGAUGAG X CGAA AGCAAGGU 3074
2518 CCTTGCTT T AATCCTAA 566 UUAGGAUU CUGAUGAG X CGAA AAGCAAGG 3075
2519 CTTGCTTT A ATCCTAAA 567 UUUAGGAU CUGAUGAG X CGAA AAAGCAAG 3076
2522 GCTTTAAT C CTAAATGG 568 CCAUUUAG CUGAUGAG X CGAA AUUAAAGC 3077
2525 TTAATCCT A AATGGCAA 569 UUGCCAUU CUGAUGAG X CGAA AGGAUUAA 3078
2537 GGCAAACT C CTTCTTTT 570 AAAAGAAG CUGAUGAG X CGAA AGUUUGCC 3079
2540 AAACTCCT T CTTTTCCT 571 AGGAAAAG CUGAUGAG X CGAA AGGAGUUU 3080
2541 AACTCCTT C TTTTCCTG 572 CAGGAAAA CUGAUGAG X CGAA AAGGAGUU 3081
2543 CTCCTTCT T TTCCTGAC 573 GUCAGGAA CUGAUGAG X CGAA AGAAGGAG 3082
2544 TCCTTCTT T TCCTGACA 574 UGUCAGGA CUGAUGAG X CGAA AAGAAGGA 3083
2545 CCTTCTTT T CCTGACAT 575 AUGUCAGG CUGAUGAG X CGAA AAAGAAGG 3084
2546 CTTCTTTT C CTGACATT 576 AAUGUCAG CUGAUGAG X CGAA AAAAGAAG 3085
2554 CCTGACAT T CATTTGCA 577 UGCAAAUG CUGAUGAG X CGAA AUGUCAGG 3086
2555 CTGACATT C ATTTGCAG 578 CUGCAAAU CUGAUGAG X CGAA AAUGUCAG 3087
2558 ACATTCAT T TGCAGGAG 579 CUCCUGCA CUGAUGAG X CGAA AUGAAUGU 3088
2559 CATTCATT T GCAGGAGG 580 CCUCCUGC CUGAUGAG X CGAA AAUGAAUG 3089
2572 GAGGACAT T GTTGATAG 581 CUAUCAAC CUGAUGAG X CGAA AUGUCCUC 3090
2575 GACATTGT T GATAGATG 582 CAUCUAUC CUGAUGAG X CGAA ACAAUGUC 3091
2579 TTGTTGAT A GATGTAAG 583 CUUACAUC CUGAUGAG X CGAA AUCAACAA 3092
2585 ATAGATGT A AGCAATTT 584 AAAUUGCU CUGAUGAG X CGAA ACAUCUAU 3093
2592 TAAGCAAT T TGTGGGGC 585 GCCCCACA CUGAUGAG X CGAA AUUGCUUA 3094
2593 AAGCAATT T GTGGGGCC 586 GGCCCCAC CUGAUGAG X CGAA AAUUGCUU 3095
2605 GGGCCCCT T ACAGTAAA 587 UUUACUGU CUGAUGAG X CGAA AGGGGCCC 3096
2606 GGCCCCTT A CAGTAAAT 588 AUUUACUG CUGAUGAG X CGAA AAGGGGCC 3097
2611 CTTACAGT A AATGAAAA 589 UUUUCAUU CUGAUGAG X CGAA ACUGUAAG 3098
2629 AGGAGACT T AAATTAAC 590 GUUAAUUU CUGAUGAG X CGAA AGUCUCCU 3099
2630 GGAGACTT A AATTAACT 591 AGUUAAUU CUGAUGAG X CGAA AAGUCUCC 3100
Table 37
2634 ACTTAAAT T AACTATGC 592 GCAUAGUU CUGAUGAG X CGAA AUUUAAGU 3101
2635 CTTAAATT A ACTATGCC 593 GGCAUAGU CUGAUGAG X CGAA AAUUUAAG 3102
2639 AATTAACT A TGCCTGCT 594 AGCAGGCA CUGAUGAG X CGAA AGUUAAUU 3103
2648 TGCCTGCT A GGTTTTAT 595 AUAAAACC CUGAUGAG X CGAA AGCAGGCA 3104
2652 TGCTAGGT T TTATCCCA 596 UGGGAUAA CUGAUGAG X CGAA ACCUAGCA 3105
2653 GCTAGGTT T TATCCCAA 597 UUGGGAUA CUGAUGAG X CGAA AACCUAGC 3106
2654 CTAGGTTT T ATCCCAAT 598 AUUGGGAU CUGAUGAG X CGAA AAACCUAG 3107
2655 TAGGTTTT A TCCCAATG 599 CAUUGGGA CUGAUGAG X CGAA AAAACCUA 3108
2657 GGTTTTAT C CCAATGTT 600 AACAUUGG CUGAUGAG X CGAA AUAAAACC 3109
2665 CCCAATGT T ACTAAATA 601 UAUUUAGU CUGAUGAG X CGAA ACAUUGGG 3110
2666 CCAATGTT A CTAAATAT 602 AUAUUUAG CUGAUGAG X CGAA AACAUUGG 3111
2669 ATGTTACT A AATATTTG 603 CAAAUAUU CUGAUGAG X CGAA AGUAACAU 3112
2673 TACTAAAT A TTTGCCCT 604 AGGGCAAA CUGAUGAG X CGAA AUUUAGUA 3113
2675 CTAAATAT T TGCCCTTA 605 UAAGGGCA CUGAUGAG X CGAA AUAUUUAG 3114
2676 TAAATATT T GCCCTTAG 606 CUAAGGGC CUGAUGAG X CGAA AAUAUUUA 3115
2682 TTTGCCCT T AGATAAAG 607 CUUUAUCU CUGAUGAG X CGAA AGGGCAAA 3116
2683 TTGCCCTT A GATAAAGG 608 CCUUUAUC CUGAUGAG X CGAA AAGGGCAA 3117
2687 CCTTAGAT A AAGGGATC 609 GAUCCCUU CUGAUGAG X CGAA AUCUAAGG 3118
2695 AAAGGGAT C AAACCGTA 610 UACGGUUU CUGAUGAG X CGAA AUCCCUUU 3119
2703 CAAACCGT A TTATCCAG 611 CUGGAUAA CUGAUGAG X CGAA ACGGUUUG 3120
2705 AACCGTAT T ATCCAGAG 612 CUCUGGAU CUGAUGAG X CGAA AUACGGUU 3121
2706 ACCGTATT A TCCAGAGT 613 ACUCUGGA CUGAUGAG X CGAA AAUACGGU 3122
2708 CGTATTAT C CAGAGTAT 614 AUACUCUG CUGAUGAG X CGAA AUAAUACG 3123
2715 TCCAGAGT A TGTAGTTA 615 UAACUACA CUGAUGAG X CGAA ACUCUGGA 3124
2719 GAGTATGT A GTTAATCA 616 UGAUUAAC CUGAUGAG X CGAA ACAUACUC 3125
2722 TATGTAGT T AATCATTA 617 UAAUGAUU CUGAUGAG X CGAA ACUACAUA 3126
2723 ATGTAGTT A ATCATTAC 618 GUAAUGAU CUGAUGAG X CGAA AACUACAU 3127
2726 TAGTTAAT C ATTACTTC 619 GAAGUAAU CUGAUGAG X CGAA AUUAACUA 3128
2729 TTAATCAT T ACTTCCAG 620 CUGGAAGU CUGAUGAG X CGAA AUGAUUAA 3129
2730 TAATCATT A CTTCCAGA 621 UCUGGAAG CUGAUGAG X CGAA AAUGAUUA 3130
2733 TCATTACT T CCAGACGC 622 GCGUCUGG CUGAUGAG X CGAA AGUAAUGA 3131
2734 CATTACTT c CAGACGCG 623 CGCGUCUG CUGAUGAG X CGAA AAGUAAUG 3132
2747 CGCGACAT T ATTTACAC 624 GUGUAAAU CUGAUGAG X CGAA AUGUCGCG 3133
2748 GCGACATT A TTTACACA 625 UGUGUAAA CUGAUGAG X CGAA AAUGUCGC 3134
2750 GACATTAT T TACACACT 626 AGUGUGUA CUGAUGAG X CGAA AUAAUGUC 3135
2751 ACATTATT T ACACACTC 627 GAGUGUGU CUGAUGAG X CGAA AAUAAUGU 3136
2752 CATTATTT A CACACTCT 628 AGAGUGUG CUGAUGAG X CGAA AAAUAAUG 3137
2759 TACACACT C TTTGGAAG 629 CUUCCAAA CUGAUGAG X CGAA AGUGUGUA 3138
2761 CACACTCT T TGGAAGGC 630 GCCUUCCA CUGAUGAG X CGAA AGAGUGUG 3139
2762 ACACTCTT T GGAAGGCG 631 CGCCUUCC CUGAUGAG X CGAA AAGAGUGU 3140
2776 GCGGGGAT C TTATATAA 632 UUAUAUAA CUGAUGAG X CGAA AUCCCCGC 3141
2778 GGGGATCT T ATATAAAA 633 UUUUAUAU CUGAUGAG X CGAA AGAUCCCC 3142
2779 GGGATCTT A TATAAAAG 634 CUUUUAUA CUGAUGAG X CGAA AAGAUCCC 3143
2781 GATCTTAT A TAAAAGAG 635 CUCUUUUA CUGAUGAG X CGAA AUAAGAUC 3144
2783 TCTTATAT A AAAGAGAG 636 CUCUCUUU CUGAUGAG X CGAA AUAUAAGA 3145
2793 AAGAGAGT C CACACGTA 637 UACGUGUG CUGAUGAG X CGAA ACUCUCUU 3146
2801 CCACACGT A GCGCCTCA 638 UGAGGCGC CUGAUGAG X CGAA ACGUGUGG 3147
2808 TAGCGCCT C ATTTTGCG 639 CGCAAAAU CUGAUGAG X CGAA AGGCGCUA 3148
2811 CGCCTCAT T TTGCGGGT 640 ACCCGCAA CUGAUGAG X CGAA AUGAGGCG 3149
2812 GCCTCATT T TGCGGGTC 641 GACCCGCA CUGAUGAG X CGAA AAUGAGGC 3150
2813 CCTCATTT T GCGGGTCA 642 UGACCCGC CUGAUGAG X CGAA AAAUGAGG 3151
Table 37
2820 TTGCGGGT C ACCATATT 643 AAUAUGGU CUGAUGAG X CGAA ACCCGCAA 3152
2826 GTCACCAT A TTCTTGGG 644 CCCAAGAA CUGAUGAG X CGAA AUGGUGAC 3153
2828 CACCATAT T CTTGGGAA 645 UUCCCAAG CUGAUGAG X CGAA AUAUGGUG 3154
2829 ACCATATT C TTGGGAAC 646 GUUCCCAA CUGAUGAG X CGAA AAUAUGGU 3155
2831 CATATTCT T GGGAACAA 647 UUGUUCCC CUGAUGAG X CGAA AGAAUAUG 3156
2843 AACAAGAT C TACAGCAT 648 AUGCUGUA CUGAUGAG X CGAA AUCUUGUU 3157
2845 CAAGATCT A CAGCATGG 649 CCAUGCUG CUGAUGAG X CGAA AGAUCUUG 3158
2859 TGGGAGGT T GGTCTTCC 650 GGAAGACC CUGAUGAG X CGAA ACCUCCCA 3159
2863 AGGTTGGT c TTCCAAAC 651 GUUUGGAA CUGAUGAG X CGAA ACCAACCU 3160
2865 GTTGGTCT T CCAAACCT 652 AGGUUUGG CUGAUGAG X CGAA AGACCAAC 3161
2866 TTGGTCTT c CAAACCTC 653 GAGGUUUG CUGAUGAG X CGAA AAGACCAA 3162
2874 CCAAACCT c GAAAAGGC 654 GCCUUUUC CUGAUGAG X CGAA AGGUUUGG 3163
2895 GGACAAAT c TTTCTGTC 655 GACAGAAA CUGAUGAG X CGAA AUUUGUCC 3164
2897 ACAAATCT T TCTGTCCC 656 GGGACAGA CUGAUGAG X CGAA AGAUUUGU 3165
2898 CAAATCTT T CTGTCCCC 657 GGGGACAG CUGAUGAG X CGAA AAGAUUUG 3166
2899 AAATCTTT c TGTCCCCA 658 UGGGGACA CUGAUGAG X CGAA AAAGAUUU 3167
2903 CTTTCTGT c CCCAATCC 659 GGAUUGGG CUGAUGAG X CGAA ACAGAAAG 3168
2910 TCCCCAAT c CCCTGGGA 660 UCCCAGGG CUGAUGAG X CGAA AUUGGGGA 3169
2920 CCTGGGAT T CTTCCCCG 661 CGGGGAAG CUGAUGAG X CGAA AUCCCAGG 3170
2921 CTGGGATT c TTCCCCGA 662 UCGGGGAA CUGAUGAG X CGAA AAUCCCAG 3171
2923 GGGATTCT T CCCCGATC 663 GAUCGGGG CUGAUGAG X CGAA AGAAUCCC 3172
2924 GGATTCTT c CCCGATCA 664 UGAUCGGG CUGAUGAG X CGAA AAGAAUCC 3173
2931 TCCCCGAT c ATCAGTTG 665 CAACUGAU CUGAUGAG X CGAA AUCGGGGA 3174
2934 CCGATCAT c AGTTGGAC 666 GUCCAACU CUGAUGAG X CGAA AUGAUCGG 3175
2938 TCATCAGT T GGACCCTG 667 CAGGGUCC CUGAUGAG X CGAA ACUGAUGA 3176
2950 CCCTGCAT T CAAAGCCA 668 UGGCUUUG CUGAUGAG X CGAA AUGCAGGG 3177
2951 CCTGCATT c AAAGCCAA 669 UUGGCUUU CUGAUGAG X CGAA AAUGCAGG 3178
2962 AGCCAACT c AGTAAATC 670 GAUUUACU CUGAUGAG X CGAA AGUUGGCU 3179
2966 AACTCAGT A AATCCAGA 671 UCUGGAUU CUGAUGAG X CGAA ACUGAGUU 3180
2970 CAGTAAAT c CAGATTGG 672 CCAAUCUG CUGAUGAG X CGAA AUUUACUG 3181
2976 ATCCAGAT T GGGACCTC 673 GAGGUCCC CUGAUGAG X CGAA AUCUGGAU 3182
2984 TGGGACCT c AACCCGCA 674 UGCGGGUU CUGAUGAG X CGAA AGGUCCCA 3183
3037 GGGAGCAT T CGGGCCAG 675 CUGGCCCG CUGAUGAG X CGAA AUGCUCCC 3184
3038 GGAGCATT c GGGCCAGG 676 CCUGGCCC CUGAUGAG X CGAA AAUGCUCC 3185
3049 GCCAGGGT T CACCCCTC 677 GAGGGGUG CUGAUGAG X CGAA ACCCUGGC 3186
3050 CCAGGGTT c ACCCCTCC 678 GGAGGGGU CUGAUGAG X CGAA AACCCUGG 3187
3057 TCACCCCT c CCCATGGG 679 CCCAUGGG CUGAUGAG X CGAA AGGGGUGA 3188
3073 GGGACTGT T GGGGTGGA 680 UCCACCCC CUGAUGAG X CGAA ACAGUCCC 3189
3087 GGAGCCCT c ACGCTCAG 681 CUGAGCGU CUGAUGAG X CGAA AGGGCUCC 3190
3093 CTCACGCT c AGGGCCTA 682 UAGGCCCU CUGAUGAG X CGAA AGCGUGAG 3191
3101 CAGGGCCT A CTCACAAC 683 GUUGUGAG CUGAUGAG X CGAA AGGCCCUG 3192
3104 GGCCTACT c ACAACTGT 684 ACAGUUGU CUGAUGAG X CGAA AGUAGGCC 3193
3123 CAGCAGCT c CTCCTCCT 685 AGGAGGAG CUGAUGAG X CGAA AGCUGCUG 3194
3126 CAGCTCCT c CTCCTGCC 686 GGCAGGAG CUGAUGAG X CGAA AGGAGCUG 3195
3129 CTCCTCCT c CTGCCTCC 687 GGAGGCAG CUGAUGAG X CGAA AGGAGGAG 3196
3136 TCCTGCCT c CACCAATC 688 GAUUGGUG CUGAUGAG X CGAA AGGCAGGA 3197
3144 CCACCAAT c GGCAGTCA 689 UGACUGCC CUGAUGAG X CGAA AUUGGUGG 3198
3151 TCGGCAGT c AGGAAGGC 690 GCCUUCCU CUGAUGAG X CGAA ACUGCCGA 3199
3165 GGCAGCCT A CTCCCTTA 691 UAAGGGAG CUGAUGAG X CGAA AGGCUGCC 3200
3168 AGCCTACT c CCTTATCT 692 AGAUAAGG CUGAUGAG X CGAA AGUAGGCU 3201
3172 TACTCCCT T ATCTCCAC 693 GUGGAGAU CUGAUGAG X CGAA AGGGAGUA 3202
Taoie 5/
Input Sequence = AF100308. Cut Site = UH/ .
Stem Length = 8 . Core Sequence = CUGAUGAG X CGAA (X = GCCGUUAGGC or other stem II)
AF100308 (Hepatitis B virus strain 2-18, 3215 bp)
Table 38 Table 38: Human HBV Inozyme and Substrate Sequence
Pos Substrate Seq ID Ribozyme Rz Seq ID
9 AACTCCAC C ACTTTCCA 702 UGGAAAGU CUGAUGAG X CGAA IUGGAGUU 3211
10 ACTCCACC A CTTTCCAC 703 GUGGAAAG CUGAUGAG X CGAA IGUGGAGU 3212
12 TCCACCAC T TTCCACCA 704 UGGUGGAA CUGAUGAG X CGAA IUGGUGGA 3213
16 CCACTTTC c ACCAAACT 705 AGUUUGGU CUGAUGAG X CGAA IAAAGUGG 3214
17 CACTTTCC A CCAAACTC 706 GAGUUUGG CUGAUGAG X CGAA IGAAAGUG 3215
19 CTTTCCAC C AAACTCTT 707 AAGAGUUU CUGAUGAG X CGAA IUGGAAAG 3216
20 TTTCCACC A AACTCTTC 708 GAAGAGUU CUGAUGAG X CGAA IGUGGAAA 3217
24 CACCAAAC T CTTCAAGA 709 UCUUGAAG CUGAUGAG X CGAA IUUUGGUG 3218
26 CCAAACTC T TCAAGATC 710 GAUCUUGA CUGAUGAG X CGAA IAGUUUGG 3219
29 AACTCTTC A AGATCCCA 711 UGGGAUCU CUGAUGAG X CGAA IAAGAGUU 3220
35 TCAAGATC C CAGAGTCA 712 UGACUCUG CUGAUGAG X CGAA IAUCUUGA 3221
36 CAAGATCC C AGAGTCAG 713 CUGACUCU CUGAUGAG X CGAA IGAUCUUG 3222
37 AAGATCCC A GAGTCAGG 714 CCUGACUC CUGAUGAG X CGAA IGGAUCUU 3223
43 CCAGAGTC A GGGCCCTG 715 CAGGGCCC CUGAUGAG X CGAA IACUCUGG 3224
48 GTCAGGGC C CTGTACTT 716 AAGUACAG CUGAUGAG X CGAA ICCCUGAC 3225
49 TCAGGGCC C TGTACTTT 717 AAAGUACA CUGAUGAG X CGAA IGCCCUGA 3226
50 CAGGGCCC T GTACTTTC 718 GAAAGUAC CUGAUGAG X CGAA IGGCCCUG 3227
55 CCCTGTAC T TTCCTGCT 719 AGCAGGAA CUGAUGAG X CGAA IUACAGGG 3228
59 GTACTTTC C TGCTGGTG 720 CACCAGCA CUGAUGAG X CGAA IAAAGUAC 3229
60 TACTTTCC T GCTGGTGG 721 CCACCAGC CUGAUGAG X CGAA IGAAAGUA 3230
63 TTTCCTGC T GGTGGCTC 722 GAGCCACC CUGAUGAG X CGAA ICAGGAAA 3231
70 CTGGTGGC T CCAGTTCA 723 UGAACUGG CUGAUGAG X CGAA ICCACCAG 3232
72 GGTGGCTC C AGTTCAGG 724 CCUGAACU CUGAUGAG X CGAA IAGCCACC 3233
73 GTGGCTCC A GTTCAGGA 725 UCCUGAAC CUGAUGAG X CGAA IGAGCCAC 3234
78 TCCAGTTC A GGAACAGT 726 ACUGUUCC CUGAUGAG X CGAA lAACUGGA 3235
84 TCAGGAAC A GTGAGCCC 727 GGGCUCAC CUGAUGAG X CGAA IUUCCUGA 3236
91 CAGTGAGC C CTGCTCAG 728 CUGAGCAG CUGAUGAG X CGAA ICUCACUG 3237
92 AGTGAGCC C TGCTCAGA 729 UCUGAGCA CUGAUGAG X CGAA IGCUCACU 3238
93 GTGAGCCC T GCTCAGAA 730 UUCUGAGC CUGAUGAG X CGAA IGGCUCAC 3239
96 AGCCCTGC T CAGAATAC 731 GUAUUCUG CUGAUGAG X CGAA ICAGGGCU 3240
98 CCCTGCTC A GAATACTG 732 CAGUAUUC CUGAUGAG X CGAA IAGCAGGG 3241
105 CAGAATAC T GTCTCTGC 733 GCAGAGAC CUGAUGAG X CGAA IUAUUCUG 3242
109 ATACTGTC T CTGCCATA 734 UAUGGCAG CUGAUGAG X CGAA IACAGUAU 3243
111 ACTGTCTC T GCCATATC 735 GAUAUGGC CUGAUGAG X CGAA IAGACAGU 3244
114 GTCTCTGC C ATATCGTC 736 GACGAUAU CUGAUGAG X CGAA ICAGAGAC 3245
115 TCTCTGCC A TATCGTCA 737 UGACGAUA CUGAUGAG X CGAA IGCAGAGA 3246
123 ATATCGTC A ATCTTATC 738 GAUAAGAU CUGAUGAG X CGAA IACGAUAU 3247
127 l CGTCAATC T TATCGAAG 739 CUUCGAUA CUGAUGAG X CGAA IAUUGACG 3248
138 TCGAAGAC T GGGGACCC 740 GGGUCCCC CUGAUGAG X CGAA IUCUUCGA 3249
145 CTGGGGAC C CTGTACCG 741 CGGUACAG CUGAUGAG X CGAA IUCCCCAG 3250
146 TGGGGACC C TGTACCGA 742 UCGGUACA CUGAUGAG X CGAA IGUCCCCA 3251
147 GGGGACCC T GTACCGAA 743 UUCGGUAC CUGAUGAG X CGAA IGGUCCCC 3252
152 CCCTGTAC C GAACATGG 744 CCAUGUUC CUGAUGAG X CGAA IUACAGGG 3253
157 TACCGAAC A TGGAGAAC 745 GUUCUCCA CUGAUGAG X CGAA IUUCGGUA 3254
166 TGGAGAAC A TCGCATCA 746 UGAUGCGA CUGAUGAG X CGAA lUUCUCCA 3255
171 AACATCGC A TCAGGACT 747 AGUCCUGA CUGAUGAG X CGAA ICGAUGUU 3256
174 ATCGCATC A GGACTCCT 748 AGGAGUCC CUGAUGAG X CGAA IAUGCGAU 3257
Table 38
179 ATCAGGAC T CCTAGGAC 749 GUCCUAGG CUGAUGAG X CGAA IUCCUGAU 3258
181 CAGGACTC C TAGGACCC 750 GGGUCCUA CUGAUGAG X CGAA IAGUCCUG 3259
182 AGGACTCC T AGGACCCC 751 GGGGUCCU CUGAUGAG X CGAA IGAGUCCU 3260
188 CCTAGGAC c CCTGCTCG 752 CGAGCAGG CUGAUGAG X CGAA IUCCUAGG 3261
189 CTAGGACC c CTGCTCGT 753 ACGAGCAG CUGAUGAG X CGAA IGUCCUAG 3262
190 TAGGACCC c TGCTCGTG 754 CACGAGCA CUGAUGAG X CGAA IGGUCCUA 3263
191 AGGACCCC T GCTCGTGT 755 ACACGAGC CUGAUGAG X CGAA IGGGUCCU 3264
194 ACCCCTGC T CGTGTTAC 756 GUAACACG CUGAUGAG X CGAA ICAGGGGU 3265
203 CGTGTTAC A GGCGGGGT 757 ACCCCGCC CUGAUGAG X CGAA IUAACACG 3266
217 GGTTTTTC T TGTTGACA 758 UGUCAACA CUGAUGAG X CGAA IAAAAACC 3267
225 TTGTTGAC A AAAATCCT 759 AGGAUUUU CUGAUGAG X CGAA lUCAACAA 3268
232 CAAAAATC C TCACAATA 760 UAUUGUGA CUGAUGAG X CGAA IAUUUUUG 3269
233 AAAAATCC T CACAATAC 761 GUAUUGUG CUGAUGAG X CGAA IGAUUUUU 3270
235 AAATCCTC A CAATACCA 762 UGGUAUUG CUGAUGAG X CGAA IAGGAUUU 3271
237 ATCCTCAC A ATACCACA 763 UGUGGUAU CUGAUGAG X CGAA IUGAGGAU 3272
242 CACAATAC C ACAGAGTC 764 GACUCUGU CUGAUGAG X CGAA IUAUUGUG 3273
243 ACAATACC A CAGAGTCT 765 AGACUCUG CUGAUGAG X CGAA IGUAUUGU 3274
245 AATACCAC A GAGTCTAG 766 CUAGACUC CUGAUGAG X CGAA IUGGUAUU 3275
251 ACAGAGTC T AGACTCGT 767 ACGAGUCU CUGAUGAG X CGAA IACUCUGU 3276
256 GTCTAGAC T CGTGGTGG 768 CCACCACG CUGAUGAG X CGAA IUCUAGAC 3277
267 TGGTGGAC T TCTCTCAA 769 UUGAGAGA CUGAUGAG X CGAA IUCCACCA 3278
270 TGGACTTC T CTCAATTT 770 AAAUUGAG CUGAUGAG X CGAA IAAGUCCA 3279
272 GACTTCTC T CAATTTTC 771 GAAAAUUG CUGAUGAG X CGAA IAGAAGUC 3280
274 CTTCTCTC A ATTTTCTA 772 UAGAAAAU CUGAUGAG X CGAA IAGAGAAG 3281
281 CAATTTTC T AGGGGGAA 773 UUCCCCCU CUGAUGAG X CGAA IAAAAUUG 3282
291 GGGGGAAC A CCCGTGTG 774 CACACGGG CUGAUGAG X CGAA IUUCCCCC 3283
293 GGGAACAC C CGTGTGTC 775 GACACACG CUGAUGAG X CGAA IUGUUCCC 3284
294 GGAACACC C GTGTGTCT 776 AGACACAC CUGAUGAG X CGAA IGUGUUCC 3285
302 CGTGTGTC T TGGCCAAA 777 UUUGGCCA CUGAUGAG X CGAA IACACACG 3286
307 GTCTTGGC C AAAATTCG 778 CGAAUUUU CUGAUGAG X CGAA ICCAAGAC 3287
308 TCTTGGCC A AAATTCGC 779 GCGAAUUU CUGAUGAG X CGAA IGCCAAGA 3288
317 AAATTCGC A GTCCCAAA 780 UUUGGGAC CUGAUGAG X CGAA ICGAAUUU 3289
321 TCGCAGTC C CAAATCTC 781 GAGAUUUG CUGAUGAG X CGAA IACUGCGA 3290
322 CGCAGTCC C AAATCTCC 782 GGAGAUUU CUGAUGAG X CGAA IGACUGCG 3291
323 GCAGTCCC A AATCTCCA 783 UGGAGAUU CUGAUGAG X CGAA IGGACUGC 3292
328 CCCAAATC T CCAGTCAC 784 GUGACUGG CUGAUGAG X CGAA IAUUUGGG 3293
330 CAAATCTC C AGTCACTC 785 GAGUGACU CUGAUGAG X CGAA IAGAUUUG 3294
331 AAATCTCC A GTCACTCA 786 UGAGUGAC CUGAUGAG X CGAA IGAGAUUU 3295
335 CTCCAGTC A CTCACCAA 787 UUGGUGAG CUGAUGAG X CGAA IACUGGAG 3296
337 CCAGTCAC T CACCAACC 788 GGUUGGUG CUGAUGAG X CGAA IUGACUGG 3297
339 AGTCACTC A CCAACCTG 789 CAGGUUGG CUGAUGAG X CGAA IAGUGACU 3298
341 TCACTCAC C AACCTGTT 790 AACAGGUU CUGAUGAG X CGAA lUGAGUGA 3299
342 CACTCACC A ACCTGTTG 791 CAACAGGU CUGAUGAG X CGAA IGUGAGUG 3300
345 TCACCAAC C TGTTGTCC 792 GGACAACA CUGAUGAG X CGAA IUUGGUGA 3301
346 CACCAACC T GTTGTCCT 793 AGGACAAC CUGAUGAG X CGAA IGUUGGUG 3302
353 CTGTTGTC C TCCAATTT 794 AAAUUGGA CUGAUGAG X CGAA lACAACAG 3303
354 TGTTGTCC T CCAATTTG 795 CAAAUUGG CUGAUGAG X CGAA IGACAACA 3304
356 TTGTCCTC C AATTTGTC 796 GACAAAUU CUGAUGAG X CGAA IAGGACAA 3305
357 TGTCCTCC A ATTTGTCC 797 GGACAAAU CUGAUGAG X CGAA IGAGGACA 3306
365 AATTTGTC C TGGTTATC 798 GAUAACCA CUGAUGAG X CGAA IACAAAUU 3307
366 ATTTGTCC T GGTTATCG 799 CGAUAACC CUGAUGAG X CGAA IGACAAAU 3308
Table 38
376 GTTATCGC T GGATGTGT 800 ACACAUCC CUGAUGAG X CGAA ICGAUAAC 3309
386 GATGTGTC T GCGGCGTT 801 AACGCCGC CUGAUGAG X CGAA IACACAUC 3310
400 GTTTTATC A TCTTCCTC 802 GAGGAAGA CUGAUGAG X CGAA IAUAAAAC 3311
403 TTATCATC T TCCTCTGC 803 GCAGAGGA CUGAUGAG X CGAA IAUGAUAA 3312
406 TCATCTTC C TCTGCATC 804 GAUGCAGA CUGAUGAG X CGAA IAAGAUGA 3313
407 CATCTTCC T CTGCATCC 805 GGAUGCAG CUGAUGAG X CGAA IGAAGAUG 3314
409 TCTTCCTC T GCATCCTG 806 CAGGAUGC CUGAUGAG X CGAA IAGGAAGA 3315
412 TCCTCTGC A TCCTGCTG 807 CAGCAGGA CUGAUGAG X CGAA ICAGAGGA 3316
415 TCTGCATC C TGCTGCTA 808 UAGCAGCA CUGAUGAG X CGAA IAUGCAGA 3317
416 CTGCATCC T GCTGCTAT 809 AUAGCAGC CUGAUGAG X CGAA IGAUGCAG 3318
419 CATCCTGC T GCTATGCC 810 GGCAUAGC CUGAUGAG X CGAA ICAGGAUG 3319
422 CCTGCTGC T ATGCCTCA 811 UGAGGCAU CUGAUGAG X CGAA ICAGCAGG 3320
427 TGCTATGC C TCATCTTC 812 GAAGAUGA CUGAUGAG X CGAA ICAUAGCA 3321
428 GCTATGCC T CATCTTCT 813 AGAAGAUG CUGAUGAG X CGAA IGCAUAGC 3322
430 TATGCCTC A TCTTCTTG 814 CAAGAAGA CUGAUGAG X CGAA IAGGCAUA 3323
433 GCCTCATC T TCTTGTTG 815 CAACAAGA CUGAUGAG X CGAA IAUGAGGC 3324
436 TCATCTTC T TGTTGGTT 816 AACCAACA CUGAUGAG X CGAA IAAGAUGA 3325
446 GTTGGTTC T TCTGGACT 817 AGUCCAGA CUGAUGAG X CGAA IAACCAAC 3326
449 GGTTCTTC T GGACTATC 818 GAUAGUCC CUGAUGAG X CGAA IAAGAACC 3327
454 TTCTGGAC T ATCAAGGT 819 ACCUUGAU CUGAUGAG X CGAA IUCCAGAA 3328
458 GGACTATC A AGGTATGT 820 ACAUACCU CUGAUGAG X CGAA IAUAGUCC 3329
470 TATGTTGC C CGTTTGTC 821 GACAAACG CUGAUGAG X CGAA ICAACAUA 3330
471 ATGTTGCC C GTTTGTCC 822 GGACAAAC CUGAUGAG X CGAA IGCAACAU 3331
479 CGTTTGTC C TCTAATTC 823 GAAUUAGA CUGAUGAG X CGAA IACAAACG 3332
480 GTTTGTCC T CTAATTCC 824 GGAAUUAG CUGAUGAG X CGAA IGACAAAC 3333
482 TTGTCCTC T AATTCCAG 825 CUGGAAUU CUGAUGAG X CGAA IAGGACAA 3334
488 TCTAATTC c AGGATCAT 826 AUGAUCCU CUGAUGAG X CGAA lAAUUAGA 3335
489 CTAATTCC A GGATCATC 827 GAUGAUCC CUGAUGAG X CGAA IGAAUUAG 3336
495 CCAGGATC A TCAACAAC j 828 GUUGUUGA CUGAUGAG X CGAA IAUCCUGG 3337
498 GGATCATC A ACAACCAG 829 CUGGUUGU CUGAUGAG X CGAA IAUGAUCC 3338
501 TCATCAAC A ACCAGCAC 830 GUGCUGGU CUGAUGAG X CGAA IUUGAUGA 3339
504 TCAACAAC c AGCACCGG 831 CCGGUGCU CUGAUGAG X CGAA IUUGUUGA 3340
505 CAACAACC A GCACCGGA 832 UCCGGUGC CUGAUGAG X CGAA IGUUGUUG 3341
508 CAACCAGC A CCGGACCA 833 UGGUCCGG CUGAUGAG X CGAA ICUGGUUG 3342
510 ACCAGCAC C GGACCATG 834 CAUGGUCC CUGAUGAG X CGAA IUGCUGGU 3343
515 CACCGGAC C ATGCAAAA 835 UUUUGCAU CUGAUGAG X CGAA IUCCGGUG 3344
516 ACCGGACC A TGCAAAAC 836 GUUUUGCA CUGAUGAG X CGAA IGUCCGGU 3345
520 GACCATGC A AAACCTGC 837 GCAGGUUU CUGAUGAG X CGAA ICAUGGUC 3346
525 TGCAAAAC C TGCACAAC 838 GUUGUGCA CUGAUGAG X CGAA IUUUUGCA 3347
526 GCAAAACC T GCACAACT 839 AGUUGUGC CUGAUGAG X CGAA IGUUUUGC 3348
529 AAACCTGC A CAACTCCT 840 AGGAGUUG CUGAUGAG X CGAA ICAGGUUU 3349
531 ACCTGCAC A ACTCCTGC 841 GCAGGAGU CUGAUGAG X CGAA IUGCAGGU 3350
534 TGCACAAC T CCTGCTCA 842 UGAGCAGG CUGAUGAG X CGAA IUUGUGCA 3351
536 CACAACTC C TGCTCAAG 843 CUUGAGCA CUGAUGAG X CGAA IAGUUGUG 3352
537 ACAACTCC T GCTCAAGG 844 CCUUGAGC CUGAUGAG X CGAA IGAGUUGU 3353
540 ACTCCTGC T CAAGGAAC 845 GUUCCUUG CUGAUGAG X CGAA ICAGGAGU 3354
542 TCCTGCTC A AGGAACCT 846 AGGUUCCU CUGAUGAG X CGAA lAGCAGGA 3355
549 CAAGGAAC C TCTATGTT 847 AACAUAGA CUGAUGAG X CGAA IUUCCUUG 3356
550 AAGGAACC T CTATGTTT 848 AAACAUAG CUGAUGAG X CGAA IGUUCCUU 3357
552 GGAACCTC T ATGTTTCC 849 GGAAACAU CUGAUGAG X CGAA IAGGUUCC 3358
560 TATGTTTC C CTCATGTT 850 AACAUGAG CUGAUGAG X CGAA IAAACAUA 3359
Table 38
561 ATGTTTCC C TCATGTTG 851 CAACAUGA CUGAUGAG X CGAA IGAAACAU 3360
562 TGTTTCCC T CATGTTGC 852 GCAACAUG CUGAUGAG X CGAA IGGAAACA 3361
564 TTTCCCTC A TGTTGCTG 853 CAGCAACA CUGAUGAG X CGAA IAGGGAAA 3362
571 CATGTTGC T GTACAAAA 854 UUUUGUAC CUGAUGAG X CGAA ICAACAUG 3363
576 TGCTGTAC A AAACCTAC 855 GUAGGUUU CUGAUGAG X CGAA IUACAGCA 3364
581 TACAAAAC C TACGGACG 856 CGUCCGUA CUGAUGAG X CGAA IUUUUGUA 3365
582 ACAAAACC T ACGGACGG 857 CCGUCCGU CUGAUGAG X CGAA IGUUUUGU 3366
595 ACGGAAAC T GCACCTGT 858 ACAGGUGC CUGAUGAG X CGAA IUUUCCGU 3367
598 GAAACTGC A CCTGTATT 859 AAUACAGG CUGAUGAG X CGAA ICAGUUUC 3368
600 AACTGCAC C TGTATTCC 860 GGAAUACA CUGAUGAG X CGAA IUGCAGUU 3369
601 ACTGCACC T GTATTCCC 861 GGGAAUAC CUGAUGAG X CGAA IGUGCAGU 3370
608 CTGTATTC C CATCCCAT 862 AUGGGAUG CUGAUGAG X CGAA IAAUACAG 3371
609 TGTATTCC C ATCCCATC 863 GAUGGGAU CUGAUGAG X CGAA IGAAUACA 3372
610 GTATTCCC A TCCCATCA 864 UGAUGGGA CUGAUGAG X CGAA IGGAAUAC 3373
613 TTCCCATC C CATCATCT 865 AGAUGAUG CUGAUGAG X CGAA IAUGGGAA 3374
614 TCCCATCC C ATCATCTT 866 AAGAUGAU CUGAUGAG X CGAA IGAUGGGA 3375
615 CCCATCCC A TCATCTTG 867 CAAGAUGA CUGAUGAG X CGAA IGGAUGGG 3376
618 ATCCCATC A TCTTGGGC 868 GCCCAAGA CUGAUGAG X CGAA IAUGGGAU 3377
621 CCATCATC T TGGGCTTT 869 AAAGCCCA CUGAUGAG X CGAA IAUGAUGG 3378
627 TCTTGGGC T TTCGCAAA 870 UUUGCGAA CUGAUGAG X CGAA ICCCAAGA 3379
633 GCTTTCGC A AAATACCT 871 AGGUAUUU CUGAUGAG X CGAA ICGAAAGC 3380
640 CAAAATAC C TATGGGAG 872 CUCCCAUA CUGAUGAG X CGAA IUAUUUUG 3381
641 AAAATACC T ATGGGAGT 873 ACUCCCAU CUGAUGAG X CGAA IGUAUUUU 3382
654 GAGTGGGC C TCAGTCCG 874 CGGACUGA CUGAUGAG X CGAA ICCCACUC 3383
655 AGTGGGCC T CAGTCCGT 875 ACGGACUG CUGAUGAG X CGAA IGCCCACU 3384
657 TGGGCCTC A GTCCGTTT 876 AAACGGAC CUGAUGAG X CGAA lAGGCCCA 3385
661 CCTCAGTC C GTTTCTCT 877 AGAGAAAC CUGAUGAG X CGAA IACUGAGG 3386
667 TCCGTTTC T CTTGGCTC 878 GAGCCAAG CUGAUGAG X CGAA lAAACGGA 3387
669 CGTTTCTC T TGGCTCAG 879 CUGAGCCA CUGAUGAG X CGAA IAGAAACG 3388
674 CTCTTGGC T CAGTTTAC 880 GUAAACUG CUGAUGAG X CGAA ICCAAGAG 3389
676 CTTGGCTC A GTTTACTA 881 UAGUAAAC CUGAUGAG X CGAA IAGCCAAG 3390
683 CAGTTTAC T AGTGCCAT 882 AUGGCACU CUGAUGAG X CGAA IUAAACUG 3391
689 ACTAGTGC C ATTTGTTC 883 GAACAAAU CUGAUGAG X CGAA ICACUAGU 3392
690 CTAGTGCC A TTTGTTCA 884 UGAACAAA CUGAUGAG X CGAA IGCACUAG 3393
698 ATTTGTTC A GTGGTTCG 885 CGAACCAC CUGAUGAG X CGAA IAACAAAU 3394
713 CGTAGGGC T TTCCCCCA 886 UGGGGGAA CUGAUGAG X CGAA ICCCUACG 3395
717 GGGCTTTC C CCCACTGT 887 ACAGUGGG CUGAUGAG X CGAA IAAAGCCC 3396
718 GGCTTTCC C CCACTGTC 888 GACAGUGG CUGAUGAG X CGAA IGAAAGCC 3397
719 GCTTTCCC C CACTGTCT 889 AGACAGUG CUGAUGAG X CGAA IGGAAAGC 3398
720 CTTTCCCC C ACTGTCTG 890 CAGACAGU CUGAUGAG X CGAA IGGGAAAG 3399
721 TTTCCCCC A CTGTCTGG 891 CCAGACAG CUGAUGAG X CGAA IGGGGAAA 3400
723 TCCCCCAC T GTCTGGCT 892 AGCCAGAC CUGAUGAG X CGAA IUGGGGGA 3401
727 CCACTGTC T GGCTTTCA 893 UGAAAGCC CUGAUGAG X CGAA IACAGUGG 3402
731 TGTCTGGC T TTCAGTTA 894 UAACUGAA CUGAUGAG X CGAA ICCAGACA 3403
735 TGGCTTTC A GTTATATG 895 CAUAUAAC CUGAUGAG X CGAA IAAAGCCA 3404
764 TTGGGGGC C AAGTCTGT 896 ACAGACUU CUGAUGAG X CGAA ICCCCCAA 3405
765 TGGGGGCC A AGTCTGTA 897 UACAGACU CUGAUGAG X CGAA IGCCCCCA 3406
770 GCCAAGTC T GTACAACA 898 UGUUGUAC CUGAUGAG X CGAA IACUUGGC 3407
775 GTCTGTAC A ACATCTTG 899 CAAGAUGU CUGAUGAG X CGAA IUACAGAC 3408
778 TGTACAAC A TCTTGAGT 900 ACUCAAGA CUGAUGAG X CGAA IUUGUACA 3409
781 ACAACATC T TGAGTCCC 901 GGGACUCA CUGAUGAG X CGAA IAUGUUGU 3410
Table 38
788 CTTGAGTC C CTTTATGC 902 GCAUAAAG CUGAUGAG X CGAA IACUCAAG 3411
789 TTGAGTCC C TTTATGCC 903 GGCAUAAA CUGAUGAG X CGAA IGACUCAA 3412
790 TGAGTCCC T TTATGCCG 904 CGGCAUAA CUGAUGAG X CGAA IGGACUCA 3413
797 CTTTATGC C GCTGTTAC 905 GUAACAGC CUGAUGAG X CGAA ICAUAAAG 3414
800 TATGCCGC T GTTACCAA 906 UUGGUAAC CUGAUGAG X CGAA ICGGCAUA 3415
806 GCTGTTAC C AATTTTCT 907 AGAAAAUU CUGAUGAG X CGAA IUAACAGC 3416
807 CTGTTACC A ATTTTCTT 908 AAGAAAAU CUGAUGAG X CGAA IGUAACAG 3417
814 CAATTTTC T TTTGTCTT 909 AAGACAAA CUGAUGAG X CGAA IAAAAUUG 3418
821 CTTTTGTC T TTGGGTAT 910 AUACCCAA CUGAUGAG X CGAA IACAAAAG 3419
832 GGGTATAC A TTTAAACC 911 GGUUUAAA CUGAUGAG X CGAA IUAUACCC 3420
840 ATTTAAAC C CTCACAAA 912 UUUGUGAG CUGAUGAG X CGAA IUUUAAAU 3421
841 TTTAAACC C TCACAAAA 913 UUUUGUGA CUGAUGAG X CGAA IGUUUAAA 3422
842 TTAAACCC T CACAAAAC 914 GUUUUGUG CUGAUGAG X CGAA IGGUUUAA 3423
844 AAACCCTC A CAAAACAA 915 UUGUUUUG CUGAUGAG X CGAA IAGGGUUU 3424
846 ACCCTCAC A AAACAAAA 916 UUUUGUUU CUGAUGAG X CGAA IUGAGGGU 3425
851 CACAAAAC A AAAAGATG 917 CAUCUUUU CUGAUGAG X CGAA IUUUUGUG 3426
869 GGATATTC C CTTAACTT 918 AAGUUAAG CUGAUGAG X CGAA IAAUAUCC 3427
870 GATATTCC C TTAACTTC 919 GAAGUUAA CUGAUGAG X CGAA IGAAUAUC 3428
871 ATATTCCC T TAACTTCA 920 UGAAGUUA CUGAUGAG X CGAA IGGAAUAU 3429
876 CCCTTAAC T TCATGGGA 921 UCCCAUGA CUGAUGAG X CGAA IUUAAGGG 3430
879 TTAACTTC A TGGGATAT 922 AUAUCCCA CUGAUGAG X CGAA IAAGUUAA 3431
906 GTTGGGGC A CATTGCCA 923 UGGCAAUG CUGAUGAG X CGAA ICCCCAAC 3432
908 TGGGGCAC A TTGCCACA 924 UGUGGCAA CUGAUGAG X CGAA IUGCCCCA 3433
913 CACATTGC C ACAGGAAC 925 GUUCCUGU CUGAUGAG X CGAA ICAAUGUG 3434
914 ACATTGCC A CAGGAACA 926 UGUUCCUG CUGAUGAG X CGAA IGCAAUGU 3435
916 ATTGCCAC A GGAACATA 927 UAUGUUCC CUGAUGAG X CGAA lUGGCAAU 3436
922 ACAGGAAC A TATTGTAC 928 GUACAAUA CUGAUGAG X CGAA IUUCCUGU 3437
931 TATTGTAC A AAAAATCA 929 UGAUUUUU CUGAUGAG X CGAA IUACAAUA 3438
939 AAAAAATC A AAATGTGT 930 ACACAUUU CUGAUGAG X CGAA IAUUUUUU 3439
958 TAGGAAAC T TCCTGTAA 931 UUACAGGA CUGAUGAG X CGAA IUUUCCUA 3440
961 GAAACTTC C TGTAAACA 932 UGUUUACA CUGAUGAG X CGAA IAAGUUUC 3441
962 AAACTTCC T GTAAACAG 933 CUGUUUAC CUGAUGAG X CGAA IGAAGUUU 3442
969 CTGTAAAC A GGCCTATT 934 AAUAGGCC CUGAUGAG X CGAA IUUUACAG 3443
973 AAACAGGC C TATTGATT 935 AAUCAAUA CUGAUGAG X CGAA ICCUGUUU 3444
974 AACAGGCC T ATTGATTG 936 CAAUCAAU CUGAUGAG X CGAA IGCCUGUU 3445
994 AGTATGTC A ACGAATTG 937 CAAUUCGU CUGAUGAG X CGAA IACAUACU 3446
1009 TGTGGGTC T TTTGGGGT 938 ACCCCAAA CUGAUGAG X CGAA lACCCACA 3447
1022 GGGTTTGC C GCCCCTTT 939 AAAGGGGC CUGAUGAG X CGAA ICAAACCC 3448
1025 TTTGCCGC C CCTTTCAC 940 GUGAAAGG CUGAUGAG X CGAA ICGGCAAA 3449
1026 TTGCCGCC C CTTTCACG 941 CGUGAAAG CUGAUGAG X CGAA IGCGGCAA 3450
1027 TGCCGCCC C TTTCACGC 942 GCGUGAAA CUGAUGAG X CGAA IGGCGGCA 3451
1028 GCCGCCCC T TTCACGCA 943 UGCGUGAA CUGAUGAG X CGAA IGGGCGGC 3452
1032 CCCCTTTC A CGCAATGT 944 ACAUUGCG CUGAUGAG X CGAA IAAAGGGG 3453
1036 TTTCACGC A ATGTGGAT 945 AUCCACAU CUGAUGAG X CGAA ICGUGAAA 3454
1049 GGATATTC T GCTTTAAT 946 AUUAAAGC CUGAUGAG X CGAA IAAUAUCC 3455
1052 TATTCTGC T TTAATGCC 947 GGCAUUAA CUGAUGAG X CGAA ICAGAAUA 3456
1060 TTTAATGC C TTTATATG 948 CAUAUAAA CUGAUGAG X CGAA ICAUUAAA 3457
1061 TTAATGCC T TTATATGC 949 GCAUAUAA CUGAUGAG X CGAA IGCAUUAA 3458
1070 TTATATGC A TGCATACA 950 UGUAUGCA CUGAUGAG X CGAA ICAUAUAA 3459
1074 ATGCATGC A TACAAGCA 951 UGCUUGUA CUGAUGAG X CGAA ICAUGCAU 3460
1078 ATGCATAC A AGCAAAAC 952 GUUUUGCU CUGAUGAG X CGAA IUAUGCAU 3461
Table 38
1082 ATACAAGC A AAACAGGC 953 GCCUGUUU CUGAUGAG X CGAA ICUUGUAU 3462
1087 AGCAAAAC A GGCTTTTA 954 UAAAAGCC CUGAUGAG X CGAA IUUUUGCU 3463
1091 AAACAGGC T TTTACTTT 955 AAAGUAAA CUGAUGAG X CGAA ICCUGUUU 3464
1097 GCTTTTAC T TTCTCGCC 956 GGCGAGAA CUGAUGAG X CGAA lUAAAAGC 3465
1101 TTACTTTC T CGCCAACT 957 AGUUGGCG CUGAUGAG X CGAA IAAAGUAA 3466
1105 TTTCTCGC C AACTTACA 958 UGUAAGUU CUGAUGAG X CGAA ICGAGAAA 3467
1106 TTCTCGCC A ACTTACAA 959 UUGUAAGU CUGAUGAG X CGAA IGCGAGAA 3468
1109 TCGCCAAC T TACAAGGC 960 GCCUUGUA CUGAUGAG X CGAA IUUGGCGA 3469
1113 CAACTTAC A AGGCCTTT 961 AAAGGCCU CUGAUGAG X CGAA IUAAGUUG 3470
1118 TACAAGGC C TTTCTAAG 962 CUUAGAAA CUGAUGAG X CGAA ICCUUGUA 3471
1119 ACAAGGCC T TTCTAAGT 963 ACUUAGAA CUGAUGAG X CGAA IGCCUUGU 3472
1123 GGCCTTTC T AAGTAAAC 964 GUUUACUU CUGAUGAG X CGAA IAAAGGCC 3473
1132 AAGTAAAC A GTATGTGA 965 UCACAUAC CUGAUGAG X CGAA IUUUACUU 3474
1143 ATGTGAAC C TTTACCCC 966 GGGGUAAA CUGAUGAG X CGAA IUUCACAU 3475
1144 TGTGAACC T TTACCCCG 967 CGGGGUAA CUGAUGAG X CGAA IGUUCACA 3476
1149 ACCTTTAC C CCGTTGCT 968 AGCAACGG CUGAUGAG X CGAA IUAAAGGU 3477
1150 CCTTTACC C CGTTGCTC 969 GAGCAACG CUGAUGAG X CGAA IGUAAAGG 3478
1151 CTTTACCC C GTTGCTCG 970 CGAGCAAC CUGAUGAG X CGAA IGGUAAAG 3479
1157 CCCGTTGC T CGGCAACG 971 CGUUGCCG CUGAUGAG X CGAA ICAACGGG 3480
1162 TGCTCGGC A ACGGCCTG 972 CAGGCCGU CUGAUGAG X CGAA ICCGAGCA 3481
1168 GCAACGGC C TGGTCTAT 973 AUAGACCA CUGAUGAG X CGAA ICCGUUGC 3482
1169 CAACGGCC T GGTCTATG 974 CAUAGACC CUGAUGAG X CGAA IGCCGUUG 3483
1174 GCCTGGTC T ATGCCAAG 975 CUUGGCAU CUGAUGAG X CGAA IACCAGGC 3484
1179 GTCTATGC C AAGTGTTT 976 AAACACUU CUGAUGAG X CGAA ICAUAGAC 3485
1180 TCTATGCC A AGTGTTTG 977 CAAACACU CUGAUGAG X CGAA IGCAUAGA 3486
1190 GTGTTTGC T GACGCAAC 978 GUUGCGUC CUGAUGAG X CGAA ICAAACAC 3487
1196 GCTGACGC A ACCCCCAC 979 GUGGGGGU CUGAUGAG X CGAA ICGUCAGC 3488
1199 GACGCAAC C CCCACTGG 980 CCAGUGGG CUGAUGAG X CGAA IUUGCGUC 3489
1200 ACGCAACC C CCACTGGT 981 ACCAGUGG CUGAUGAG X CGAA IGUUGCGU 3490
1201 CGCAACCC C CACTGGTT 982 AACCAGUG CUGAUGAG X CGAA IGGUUGCG 3491
1202 GCAACCCC C ACTGGTTG 983 CAACCAGU CUGAUGAG X CGAA IGGGUUGC 3492
1203 CAACCCCC A CTGGTTGG 984 CCAACCAG CUGAUGAG X CGAA IGGGGUUG 3493
1205 ACCCCCAC T GGTTGGGG 985 CCCCAACC CUGAUGAG X CGAA IUGGGGGU 3494
1215 GTTGGGGC T TGGCCATA 986 UAUGGCCA CUGAUGAG X CGAA ICCCCAAC 3495
1220 GGCTTGGC C ATAGGCCA 987 UGGCCUAU CUGAUGAG X CGAA ICCAAGCC 3496
1221 GCTTGGCC A TAGGCCAT 988 AUGGCCUA CUGAUGAG X CGAA IGCCAAGC 3497
1227 CCATAGGC C ATCAGCGC 989 GCGCUGAU CUGAUGAG X CGAA ICCUAUGG 3498
1228 CATAGGCC A TCAGCGCA 990 UGCGCUGA CUGAUGAG X CGAA IGCCUAUG 3499
1231 AGGCCATC A GCGCATGC 991 GCAUGCGC CUGAUGAG X CGAA IAUGGCCU 3500
1236 ATCAGCGC A TGCGTGGA 992 UCCACGCA CUGAUGAG X CGAA ICGCUGAU 3501
1247 CGTGGAAC C TTTGTGTC 993 GACACAAA CUGAUGAG X CGAA IUUCCACG 3502
1248 GTGGAACC T TTGTGTCT 994 AGACACAA CUGAUGAG X CGAA IGUUCCAC 3503
1256 TTTGTGTC T CCTCTGCC 995 GGCAGAGG CUGAUGAG X CGAA lACACAAA 3504
1258 TGTGTCTC C TCTGCCGA 996 UCGGCAGA CUGAUGAG X CGAA lAGACACA 3505
1259 GTGTCTCC T CTGCCGAT 997 AUCGGCAG CUGAUGAG X CGAA IGAGACAC 3506
1261 GTCTCCTC T GCCGATCC 998 GGAUCGGC CUGAUGAG X CGAA IAGGAGAC 3507
1264 TCCTCTGC C GATCCATA 999 UAUGGAUC CUGAUGAG X CGAA ICAGAGGA 3508
1269 TGCCGATC C ATACCGCG 1000 CGCGGUAU CUGAUGAG X CGAA IAUCGGCA 3509
1270 GCCGATCC A TACCGCGG 1001 CCGCGGUA CUGAUGAG X CGAA IGAUCGGC 3510
1274 ATCCATAC C GCGGAACT 1002 AGUUCCGC CUGAUGAG X CGAA IUAUGGAU 3511
1282 CGCGGAAC T CCTAGCCG 1003 CGGCUAGG CUGAUGAG X CGAA IUUCCGCG 3512
Table 38
1284 CGGAACTC C TAGCCGCT 1004 AGCGGCUA CUGAUGAG X CGAA IAGUUCCG 3513
1285 GGAACTCC T AGCCGCTT 1005 AAGCGGCU CUGAUGAG X CGAA IGAGUUCC 3514
1289 CTCCTAGC C GCTTGTTT 1006 AAACAAGC CUGAUGAG X CGAA ICUAGGAG 3515
1292 CTAGCCGC T TGTTTTGC 1007 GCAAAACA CUGAUGAG X CGAA ICGGCUAG 3516
1301 TGTTTTGC T CGCAGCAG 1008 CUGCUGCG CUGAUGAG X CGAA ICAAAACA 3517
1305 TTGCTCGC A GCAGGTCT 1009 AGACCUGC CUGAUGAG X CGAA ICGAGCAA 3518
1308 CTCGCAGC A GGTCTGGG 1010 CCCAGACC CUGAUGAG X CGAA ICUGCGAG 3519
1313 AGCAGGTC T GGGGCAAA 1011 UUUGCCCC CUGAUGAG X CGAA IACCUGCU 3520
1319 TCTGGGGC A AAACTCAT 1012 AUGAGUUU CUGAUGAG X CGAA ICCCCAGA 3521
1324 GGCAAAAC T CATCGGGA 1013 UCCCGAUG CUGAUGAG X CGAA IUUUUGCC 3522
1326 CAAAACTC A TCGGGACT 1014 AGUCCCGA CUGAUGAG X CGAA lAGUUUUG 3523
1334 ATCGGGAC T GACAATTC 1015 GAAUUGUC CUGAUGAG X CGAA IUCCCGAU 3524
1338 GGACTGAC A ATTCTGTC 1016 GACAGAAU CUGAUGAG X CGAA IUCAGUCC 3525
1343 GACAATTC T GTCGTGCT 1017 AGCACGAC CUGAUGAG X CGAA IAAUUGUC 3526
1351 TGTCGTGC T CTCCCGCA 1018 UGCGGGAG CUGAUGAG X CGAA ICACGACA 3527
1353 TCGTGCTC T CCCGCAAA 1019 UUUGCGGG CUGAUGAG X CGAA IAGCACGA 3528
1355 GTGCTCTC C CGCAAATA 1020 UAUUUGCG CUGAUGAG X CGAA IAGAGCAC 3529
1356 TGCTCTCC C GCAAATAT 1021 AUAUUUGC CUGAUGAG X CGAA IGAGAGCA 3530
1359 TCTCCCGC A AATATACA 1022 UGUAUAUU CUGAUGAG X CGAA ICGGGAGA 3531
1367 AAATATAC A TCATTTCC 1023 GGAAAUGA CUGAUGAG X CGAA IUAUAUUU 3532
1370 TATACATC A TTTCCATG 1024 CAUGGAAA CUGAUGAG X CGAA IAUGUAUA 3533
1375 ATCATTTC C ATGGCTGC 1025 GCAGCCAU CUGAUGAG X CGAA IAAAUGAU 3534
1376 TCATTTCC A TGGCTGCT 1026 AGCAGCCA CUGAUGAG X CGAA IGAAAUGA 3535
1381 TCCATGGC T GCTAGGCT 1027 AGCCUAGC CUGAUGAG X CGAA ICCAUGGA 3536
1384 ATGGCTGC T AGGCTGTG 1028 CACAGCCU CUGAUGAG X CGAA ICAGCCAU 3537
1389 TGCTAGGC T GTGCTGCC 1029 GGCAGCAC CUGAUGAG X CGAA ICCUAGCA 3538
1394 GGCTGTGC T GCCAACTG 1030 CAGUUGGC CUGAUGAG X CGAA ICACAGCC 3539
1397 TGTGCTGC C AACTGGAT 1031 AUCCAGUU CUGAUGAG X CGAA ICAGCACA 3540
1398 GTGCTGCC A ACTGGATC 1032 GAUCCAGU CUGAUGAG X CGAA IGCAGCAC 3541
1401 CTGCCAAC T GGATCCTA 1033 UAGGAUCC CUGAUGAG X CGAA IUUGGCAG 3542
1407 ACTGGATC C TACGCGGG 1034 CCCGCGUA CUGAUGAG X CGAA IAUCCAGU 3543
1408 CTGGATCC T ACGCGGGA 1035 UCCCGCGU CUGAUGAG X CGAA IGAUCCAG 3544
1421 GGGACGTC C TTTGTTTA 1036 UAAACAAA CUGAUGAG X CGAA IACGUCCC 3545
1422 GGACGTCC T TTGTTTAC 1037 GUAAACAA CUGAUGAG X CGAA IGACGUCC 3546
1434 TTTACGTC c CGTCGGCG 1038 CGCCGACG CUGAUGAG X CGAA IACGUAAA 3547
1435 TTACGTCC c GTCGGCGC 1039 GCGCCGAC CUGAUGAG X CGAA IGACGUAA 3548
1444 GTCGGCGC T GAATCCCG 1040 CGGGAUUC CUGAUGAG X CGAA ICGCCGAC 3549
1450 GCTGAATC c CGCGGACG 1041 CGUCCGCG CUGAUGAG X CGAA IAUUCAGC 3550
1451 CTGAATCC c GCGGACGA 1042 UCGUCCGC CUGAUGAG X CGAA IGAUUCAG 3551
1461 CGGACGAC c CCTCCCGG 1043 CCGGGAGG CUGAUGAG X CGAA IUCGUCCG 3552
1462 GGACGACC c CTCCCGGG 1044 CCCGGGAG CUGAUGAG X CGAA IGUCGUCC 3553
1463 GACGACCC c TCCCGGGG 1045 CCCCGGGA CUGAUGAG X CGAA IGGUCGUC 3554
1464 ACGACCCC T CCCGGGGC 1046 GCCCCGGG CUGAUGAG X CGAA IGGGUCGU 3555
1466 GACCCCTC c CGGGGCCG 1047 CGGCCCCG CUGAUGAG X CGAA IAGGGGUC 3556
1467 ACCCCTCC c GGGGCCGC 1048 GCGGCCCC CUGAUGAG X CGAA IGAGGGGU 3557
1473 CCCGGGGC c GCTTGGGG 1049 CCCCAAGC CUGAUGAG X CGAA ICCCCGGG 3558
1476 GGGGCCGC T TGGGGCTC 1050 GAGCCCCA CUGAUGAG X CGAA ICGGCCCC 3559
1483 CTTGGGGC T CTACCGCC 1051 GGCGGUAG CUGAUGAG X CGAA ICCCCAAG 3560
1485 TGGGGCTC T ACCGCCCG 1052 CGGGCGGU CUGAUGAG X CGAA IAGCCCCA 3561
1488 GGCTCTAC c GCCCGCTT 1053 AAGCGGGC CUGAUGAG X CGAA IUAGAGCC 3562
1491 TCTACCGC c CGCTTCTC 1054 GAGAAGCG CUGAUGAG X CGAA ICGGUAGA 3563
Table 38
1492 CTACCGCC C GCTTCTCC 1055 GGAGAAGC CUGAUGAG X CGAA IGCGGUAG 3564
1495 CCGCCCGC T TCTCCGCC 1056 GGCGGAGA CUGAUGAG X CGAA ICGGGCGG 3565
1498 CCCGCTTC T CCGCCTAT 1057 AUAGGCGG CUGAUGAG X CGAA IAAGCGGG 3566
1500 CGCTTCTC C GCCTATTG 1058 CAAUAGGC CUGAUGAG X CGAA IAGAAGCG 3567
1503 TTCTCCGC C TATTGTAC 1059 GUACAAUA CUGAUGAG X CGAA ICGGAGAA 3568
1504 TCTCCGCC T ATTGTACC 1060 GGUACAAU CUGAUGAG X CGAA IGCGGAGA 3569
1512 TATTGTAC C GACCGTCC 1061 GGACGGUC CUGAUGAG X CGAA IUACAAUA 3570
1516 GTACCGAC C GTCCACGG 1062 CCGUGGAC CUGAUGAG X CGAA IUCGGUAC 3571
1520 CGACCGTC C ACGGGGCG 1063 CGCCCCGU CUGAUGAG X CGAA IACGGUCG 3572
1521 GACCGTCC A CGGGGCGC 1064 GCGCCCCG CUGAUGAG X CGAA IGACGGUC 3573
1530 CGGGGCGC A CCTCTCTT 1065 AAGAGAGG CUGAUGAG X CGAA ICGCCCCG 3574
1532 GGGCGCAC C TCTCTTTA 1066 UAAAGAGA CUGAUGAG X CGAA IUGCGCCC 3575
1533 GGCGCACC T CTCTTTAC 1067 GUAAAGAG' CUGAUGAG X CGAA IGUGCGCC 3576
1535 CGCACCTC T CTTTACGC 1068 GCGUAAAG CUGAUGAG X CGAA IAGGUGCG 3577
1537 CACCTCTC T TTACGCGG 1069 CCGCGUAA CUGAUGAG X CGAA IAGAGGUG 3578
1548 ACGCGGAC T CCCCGTCT 1070 AGACGGGG CUGAUGAG X CGAA IUCCGCGU 3579
1550 GCGGACTC c CCGTCTGT 1071 ACAGACGG CUGAUGAG X CGAA IAGUCCGC 3580
1551 CGGACTCC c CGTCTGTG 1072 CACAGACG CUGAUGAG X CGAA IGAGUCCG 3581
1552 GGACTCCC c GTCTGTGC 1073 GCACAGAC CUGAUGAG X CGAA IGGAGUCC 3582
1556 TCCCCGTC T GTGCCTTC 1074 GAAGGCAC CUGAUGAG X CGAA IACGGGGA 3583
1561 GTCTGTGC c TTCTCATC 1075 GAUGAGAA CUGAUGAG X CGAA ICACAGAC 3584
1562 TCTGTGCC T TCTCATCT 1076 AGAUGAGA CUGAUGAG X CGAA IGCACAGA 3585
1565 GTGCCTTC T CATCTGCC 1077 GGCAGAUG CUGAUGAG X CGAA IAAGGCAC 3586
1567 GCCTTCTC A TCTGCCGG 1078 CCGGCAGA CUGAUGAG X CGAA IAGAAGGC 3587
1570 TTCTCATC T GCCGGACC 1079 GGUCCGGC CUGAUGAG X CGAA IAUGAGAA 3588
1573 TCATCTGC c GGACCGTG 1080 CACGGUCC CUGAUGAG X CGAA ICAGAUGA 3589
1578 TGCCGGAC c GTGTGCAC 1081 GUGCACAC CUGAUGAG X CGAA IUCCGGCA 3590
1585 CCGTGTGC A CTTCGCTT 1082 AAGCGAAG CUGAUGAG X CGAA ICACACGG 3591
1587 GTGTGCAC T TCGCTTCA 1083 UGAAGCGA CUGAUGAG X CGAA IUGCACAC 3592
1592 CACTTCGC T TCACCTCT 1084 AGAGGUGA CUGAUGAG X CGAA ICGAAGUG 3593
1595 TTCGCTTC A CCTCTGCA 1085 UGCAGAGG CUGAUGAG X CGAA IAAGCGAA 3594
1597 CGCTTCAC C TCTGCACG 1086 CGUGCAGA CUGAUGAG X CGAA IUGAAGCG 3595
1598 GCTTCACC T CTGCACGT 1087 ACGUGCAG CUGAUGAG X CGAA IGUGAAGC 3596
1600 TTCACCTC T GCACGTCG 1088 CGACGUGC CUGAUGAG X CGAA IAGGUGAA 3597
1603 ACCTCTGC A CGTCGCAT 1089 AUGCGACG CUGAUGAG X CGAA ICAGAGGU 3598
1610 CACGTCGC A TGGAGACC 1090 GGUCUCCA CUGAUGAG X CGAA ICGACGUG 3599
1618 ATGGAGAC C ACCGTGAA 1091 UUCACGGU CUGAUGAG X CGAA IUCUCCAU 3600
1619 TGGAGACC A CCGTGAAC 1092 GUUCACGG CUGAUGAG X CGAA IGUCUCCA 3601
1621 GAGACCAC C GTGAACGC 1093 GCGUUCAC CUGAUGAG X CGAA IUGGUCUC 3602
1630 GTGAACGC C CACAGGAA 1094 UUCCUGUG CUGAUGAG X CGAA ICGUUCAC 3603
1631 TGAACGCC C ACAGGAAC 1095 GUUCCUGU CUGAUGAG X CGAA IGCGUUCA 3604
1632 GAACGCCC A CAGGAACC 1096 GGUUCCUG CUGAUGAG X CGAA IGGCGUUC 3605
1634 ACGCCCAC A GGAACCTG 1097 CAGGUUCC CUGAUGAG X CGAA IUGGGCGU 3606
1640 ACAGGAAC C TGCCCAAG 1098 CUUGGGCA CUGAUGAG X CGAA IUUCCUGU 3607
1641 CAGGAACC T GCCCAAGG 1099 CCUUGGGC CUGAUGAG X CGAA IGUUCCUG 3608
1644 GAACCTGC C CAAGGTCT 1100 AGACCUUG CUGAUGAG X CGAA ICAGGUUC 3609
1645 AACCTGCC C AAGGTCTT 1101 AAGACCUU CUGAUGAG X CGAA IGCAGGUU 3610
1646 ACCTGCCC A AGGTCTTG 1102 CAAGACCU CUGAUGAG X CGAA IGGCAGGU 3611
1652 CCAAGGTC T TGCATAAG 1103 CUUAUGCA CUGAUGAG X CGAA IACCUUGG 3612
1656 GGTCTTGC A TAAGAGGA 1104 UCCUCUUA CUGAUGAG X CGAA ICAAGACC 3613
1666 AAGAGGAC T CTTGGACT 1105 AGUCCAAG CUGAUGAG X CGAA IUCCUCUU 3614
Table 38
1668 GAGGACTC T TGGACTTT 1106 AAAGUCCA CUGAUGAG X CGAA IAGUCCUC 3615
1674 TCTTGGAC T TTCAGCAA 1107 UUGCUGAA CUGAUGAG X CGAA IUCCAAGA 3616
1678 GGACTTTC A GCAATGTC 1108 GACAUUGC CUGAUGAG X CGAA IAAAGUCC 3617
1681 CTTTCAGC A ATGTCAAC 1109 GUUGACAU CUGAUGAG X CGAA ICUGAAAG 3618
1687 GCAATGTC A ACGACCGA 1110 UCGGUCGU CUGAUGAG X CGAA IACAUUGC 3619
1693 TCAACGAC C GACCTTGA llll UCAAGGUC CUGAUGAG X CGAA IUCGUUGA 3620
1697 CGACCGAC C TTGAGGCA 1112 UGCCUCAA CUGAUGAG X CGAA IUCGGUCG 3621
1698 GACCGACC T TGAGGCAT 1113 AUGCCUCA CUGAUGAG X CGAA IGUCGGUC 3622
1705 CTTGAGGC A TACTTCAA 1114 UUGAAGUA CUGAUGAG X CGAA ICCUCAAG 3623
1709 AGGCATAC T TCAAAGAC 1115 GUCUUUGA CUGAUGAG X CGAA IUAUGCCU 3624
1712 CATACTTC A AAGACTGT 1116 ACAGUCUU CUGAUGAG X CGAA IAAGUAUG 3625
1718 TCAAAGAC T GTGTGTTT 1117 AAACACAC CUGAUGAG X CGAA IUCUUUGA 3626
1769 TAAAGGTC T TTGTACTA 1118 UAGUACAA CUGAUGAG X CGAA IACCUUUA 3627
1776 CTTTGTAC T AGGAGGCT 1119 AGCCUCCU CUGAUGAG X CGAA IUACAAAG 3628
1784 TAGGAGGC T GTAGGCAT 1120 AUGCCUAC CUGAUGAG X CGAA ICCUCCUA 3629
1791 CTGTAGGC A TAAATTGG 1121 CCAAUUUA CUGAUGAG X CGAA ICCUACAG 3630
1807 GTGTGTTC A CCAGCACC 1122 GGUGCUGG CUGAUGAG X CGAA IAACACAC 3631
1809 GTGTTCAC C AGCACCAT 1123 AUGGUGCU CUGAUGAG X CGAA IUGAACAC 3632
1810 TGTTCACC A GCACCATG 1124 CAUGGUGC CUGAUGAG X CGAA IGUGAACA 3633
1813 TCACCAGC A CCATGCAA 1125 UUGCAUGG CUGAUGAG X CGAA ICUGGUGA 3634
1815 ACCAGCAC C ATGCAACT 1126 AGUUGCAU CUGAUGAG X CGAA IUGCUGGU 3635
1816 CCAGCACC A TGCAACTT 1127 AAGUUGCA CUGAUGAG X CGAA IGUGCUGG 3636
1820 CACCATGC A ACTTTTTC 1128 GAAAAAGU CUGAUGAG X CGAA ICAUGGUG 3637
1823 CATGCAAC T TTTTCACC 1129 GGUGAAAA CUGAUGAG X CGAA IUUGCAUG 3638
1829 ACTTTTTC A CCTCTGCC 1130 GGCAGAGG CUGAUGAG X CGAA IAAAAAGU 3639
1831 TTTTTCAC C TCTGCCTA 1131 UAGGCAGA CUGAUGAG X CGAA IUGAAAAA 3640
1832 TTTTCACC T CTGCCTAA 1132 UUAGGCAG CUGAUGAG X CGAA IGUGAAAA 3641
1834 TTCACCTC T GCCTAATC 1133 GAUUAGGC CUGAUGAG X CGAA IAGGUGAA 3642
1837 ACCTCTGC C TAATCATC 1134 GAUGAUUA CUGAUGAG X CGAA ICAGAGGU 3643
1838 CCTCTGCC T AATCATCT 1135 AGAUGAUU CUGAUGAG X CGAA IGCAGAGG 3644
1843 GCCTAATC A TCTCATGT 1136 ACAUGAGA CUGAUGAG X CGAA IAUUAGGC 3645
1846 TAATCATC T CATGTTCA 1137 UGAACAUG CUGAUGAG X CGAA IAUGAUUA 3646
1848 ATCATCTC A TGTTCATG 1138 CAUGAACA CUGAUGAG X CGAA IAGAUGAU 3647
1854 TCATGTTC A TGTCCTAC 1139 GUAGGACA CUGAUGAG X CGAA IAACAUGA 3648
1859 TTCATGTC C TACTGTTC 1140 GAACAGUA CUGAUGAG X CGAA IACAUGAA 3649
1860 TCATGTCC T ACTGTTCA 1141 UGAACAGU CUGAUGAG X CGAA IGACAUGA 3650
1863 TGTCCTAC T GTTCAAGC 1142 GCUUGAAC CUGAUGAG X CGAA IUAGGACA 3651
1868 TACTGTTC A AGCCTCCA 1143 UGGAGGCU CUGAUGAG X CGAA IAACAGUA 3652
1872 GTTCAAGC C TCCAAGCT 1144 AGCUUGGA CUGAUGAG X CGAA ICUUGAAC 3653
1873 TTCAAGCC T CCAAGCTG 1145 CAGCUUGG CUGAUGAG X CGAA IGCUUGAA 3654
1875 CAAGCCTC C AAGCTGTG 1146 CACAGCUU CUGAUGAG X CGAA IAGGCUUG 3655
1876 AAGCCTCC A AGCTGTGC 1147 GCACAGCU CUGAUGAG X CGAA IGAGGCUU 3656
1880 CTCCAAGC T GTGCCTTG 1148 CAAGGCAC CUGAUGAG X CGAA ICUUGGAG 3657
1885 AGCTGTGC C TTGGGTGG 1149 CCACCCAA CUGAUGAG X CGAA ICACAGCU 3658
1886 GCTGTGCC T TGGGTGGC 1150 GCCACCCA CUGAUGAG X CGAA IGCACAGC 3659
1895 TGGGTGGC T TTGGGGCA 1151 UGCCCCAA CUGAUGAG X CGAA ICCACCCA 3660
1903 TTTGGGGC A TGGACATT 1152 AAUGUCCA CUGAUGAG X CGAA ICCCCAAA 3661
1909 GCATGGAC A TTGACCCG 1153 CGGGUCAA CUGAUGAG X CGAA IUCCAUGC 3662
1915 ACATTGAC C CGTATAAA 1154 UUUAUACG CUGAUGAG X CGAA IUCAAUGU 3663
1916 CATTGACC C GTATAAAG 1155 CUUUAUAC CUGAUGAG X CGAA IGUCAAUG 3664
1935 TTTGGAGC T TCTGTGGA 1156 UCCACAGA CUGAUGAG X CGAA ICUCCAAA 3665
Table 38
1938 GGAGCTTC T GTGGAGTT 1157 AACUCCAC CUGAUGAG X CGAA IAAGCUCC 3666
1949 GGAGTTAC T CTCTTTTT 1158 AAAAAGAG CUGAUGAG X CGAA IUAACUCC 3667
1951 AGTTACTC T CTTTTTTG 1159 CAAAAAAG CUGAUGAG X CGAA IAGUAACU 3668
1953 TTACTCTC T TTTTTGCC 1160 GGCAAAAA CUGAUGAG X CGAA IAGAGUAA 3669
1961 TTTTTTGC c TTCTGACT 1161 AGUCAGAA CUGAUGAG X CGAA ICAAAAAA 3670
1962 TTTTTGCC T TCTGACTT 1162 AAGUCAGA CUGAUGAG X CGAA IGCAAAAA 3671
1965 TTGCCTTC T GACTTCTT 1163 AAGAAGUC CUGAUGAG X CGAA IAAGGCAA 3672
1969 CTTCTGAC T TCTTTCCT 1164 AGGAAAGA CUGAUGAG X CGAA IUCAGAAG 3673
1972 CTGACTTC T TTCCTTCT 1165 AGAAGGAA CUGAUGAG X CGAA IAAGUCAG 3674
1976 CTTCTTTC c TTCTATTC 1166 GAAUAGAA CUGAUGAG X CGAA IAAAGAAG 3675
1977 TTCTTTCC T TCTATTCG 1167 CGAAUAGA CUGAUGAG X CGAA IGAAAGAA 3676
1980 TTTCCTTC T ATTCGAGA 1168 UCUCGAAU CUGAUGAG X CGAA IAAGGAAA 3677
1991 TCGAGATC T CCTCGACA 1169 UGUCGAGG CUGAUGAG X CGAA IAUCUCGA 3678
1993 GAGATCTC c TCGACACC 1170 GGUGUCGA CUGAUGAG X CGAA IAGAUCUC 3679
1994 AGATCTCC T CGACACCG 1171 CGGUGUCG CUGAUGAG X CGAA IGAGAUCU 3680
1999 TCCTCGAC A CCGCCTCT 1172 AGAGGCGG CUGAUGAG X CGAA IUCGAGGA 3681
2001 CTCGACAC c GCCTCTGC 1173 GCAGAGGC CUGAUGAG X CGAA IUGUCGAG 3682
2004 GACACCGC c TCTGCTCT 1174 AGAGCAGA CUGAUGAG X CGAA ICGGUGUC 3683
2005 ACACCGCC T CTGCTCTG 1175 CAGAGCAG CUGAUGAG X CGAA IGCGGUGU 3684
2007 ACCGCCTC T GCTCTGTA 1176 UACAGAGC CUGAUGAG X CGAA IAGGCGGU 3685
2010 GCCTCTGC T CTGTATCG 1177 CGAUACAG CUGAUGAG X CGAA ICAGAGGC 3686
2012 CTCTGCTC T GTATCGGG 1178 CCCGAUAC CUGAUGAG X CGAA IAGCAGAG 3687
2025 CGGGGGGC c TTAGAGTC 1179 GACUCUAA CUGAUGAG X CGAA ICCCCCCG 3688
2026 GGGGGGCC T TAGAGTCT 1180 AGACUCUA CUGAUGAG X CGAA IGCCCCCC 3689
2034 TTAGAGTC T CCGGAACA 1181 UGUUCCGG CUGAUGAG X CGAA lACUCUAA 3690
2036 AGAGTCTC c GGAACATT 1182 AAUGUUCC CUGAUGAG X CGAA IAGACUCU 3691
2042 TCCGGAAC A TTGTTCAC ' 1183 GUGAACAA CUGAUGAG X CGAA IUUCCGGA 3692
2049 CATTGTTC A CCTCACCA 1184 UGGUGAGG CUGAUGAG X CGAA IAACAAUG 3693
2051 TTGTTCAC C TCACCATA 1185 UAUGGUGA CUGAUGAG X CGAA IUGAACAA 3694
2052 TGTTCACC T CACCATAC 1186 GUAUGGUG CUGAUGAG X CGAA IGUGAACA 3695
2054 TTCACCTC A CCATACGG 1187 CCGUAUGG CUGAUGAG X CGAA IAGGUGAA 3696
2056 CACCTCAC C ATACGGCA 1188 UGCCGUAU CUGAUGAG X CGAA IUGAGGUG 3697
2057 ACCTCACC A TACGGCAC 1189 GUGCCGUA CUGAUGAG X CGAA IGUGAGGU 3698
2064 CATACGGC A CTCAGGCA 1190 UGCCUGAG CUGAUGAG X CGAA ICCGUAUG 3699
2066 TACGGCAC T CAGGCAAG 1191 CUUGCCUG CUGAUGAG X CGAA IUGCCGUA 3700
2068 CGGCACTC A GGCAAGCT 1192 AGCUUGCC CUGAUGAG X CGAA IAGUGCCG 3701
2072 ACTCAGGC A AGCTATTC 1193 GAAUAGCU CUGAUGAG X CGAA ICCUGAGU 3702
2076 AGGCAAGC T ATTCTGTG 1194 CACAGAAU CUGAUGAG X CGAA ICUUGCCU 3703
2081 AGCTATTC T GTGTTGGG 1195 CCCAACAC CUGAUGAG X CGAA IAAUAGCU 3704
2105 GATGAATC T AGCCACCT 1196 AGGUGGCU CUGAUGAG X CGAA IAUUCAUC 3705
2109 AATCTAGC c ACCTGGGT 1197 ACCCAGGU CUGAUGAG X CGAA ICUAGAUU 3706
2110 ATCTAGCC A CCTGGGTG 1198 CACCCAGG CUGAUGAG X CGAA IGCUAGAU 3707
2112 CTAGCCAC C TGGGTGGG 1199 CCCACCCA CUGAUGAG X CGAA IUGGCUAG 3708
2113 TAGCCACC T GGGTGGGA 1200 UCCCACCC CUGAUGAG X CGAA IGUGGCUA 3709
2138 GGAAGATC C AGCATCCA 1201 UGGAUGCU CUGAUGAG X CGAA IAUCUUCC 3710
2139 GAAGATCC A GCATCCAG 1202 CUGGAUGC CUGAUGAG X CGAA IGAUCUUC 3711
2142 GATCCAGC A TCCAGGGA 1203 UCCCUGGA CUGAUGAG X CGAA ICUGGAUC 3712
2145 CCAGCATC C AGGGAATT 1204 AAUUCCCU CUGAUGAG X CGAA IAUGCUGG 3713
2146 CAGCATCC A GGGAATTA 1205 UAAUUCCC CUGAUGAG X CGAA IGAUGCUG 3714
2161 TAGTAGTC A GCTATGTC 1206 GACAUAGC CUGAUGAG X CGAA IACUACUA 3715
2164 TAGTCAGC T ATGTCAAC 1207 GUUGACAU CUGAUGAG X CGAA ICUGACUA 3716
Table 38
2170 GCTATGTC A ACGTTAAT 1208 AUUAACGU CUGAUGAG X CGAA IACAUAGC 3717
2185 ATATGGGC C TAAAAATC 1209 GAUUUUUA CUGAUGAG X CGAA ICCCAUAU 3718
2186 TATGGGCC T AAAAATCA 1210 UGAUUUUU CUGAUGAG X CGAA IGCCCAUA 3719
2194 TAAAAATC A GACAACTA 1211 UAGUUGUC CUGAUGAG X CGAA IAUUUUUA 3720
2198 AATCAGAC A ACTATTGT 1212 ACAAUAGU CUGAUGAG X CGAA IUCUGAUU 3721
2201 CAGACAAC T ATTGTGGT 1213 ACCACAAU CUGAUGAG X CGAA IUUGUCUG 3722
2213 GTGGTTTC A CATTTCCT 1214 AGGAAAUG CUGAUGAG X CGAA IAAACCAC 3723
2215 GGTTTCAC A TTTCCTGT 1215 ACAGGAAA CUGAUGAG X CGAA IUGAAACC 3724
2220 CACATTTC C TGTCTTAC 1216 GUAAGACA CUGAUGAG X CGAA IAAAUGUG 3725
2221 ACATTTCC T GTCTTACT 1217 AGUAAGAC CUGAUGAG X CGAA IGAAAUGU 3726
2225 TTCCTGTC T TACTTTTG 1218 CAAAAGUA CUGAUGAG X CGAA lACAGGAA 3727
2229 TGTCTTAC T TTTGGGCG 1219 CGCCCAAA CUGAUGAG X CGAA IUAAGACA 3728
2244 CGAGAAAC T GTTCTTGA 1220 UCAAGAAC CUGAUGAG X CGAA IUUUCUCG 3729
2249 AACTGTTC T TGAATATT 1221 AAUAUUCA CUGAUGAG X CGAA IAACAGUU 3730
2265 TTGGTGTC T TTTGGAGT 1222 ACUCCAAA CUGAUGAG X CGAA IACACCAA 3731
2284 GGATTCGC A CTCCTCCT 1223 AGGAGGAG CUGAUGAG X CGAA ICGAAUCC 3732
2286 ATTCGCAC T CCTCCTGC 1224 GCAGGAGG CUGAUGAG X CGAA IUGCGAAU 3733
2288 TCGCACTC C TCCTGCAT 1225 AUGCAGGA CUGAUGAG X CGAA lAGUGCGA 3734
2289 CGCACTCC T CCTGCATA 1226 UAUGCAGG CUGAUGAG X CGAA IGAGUGCG 3735
2291 CACTCCTC c TGCATATA 1227 UAUAUGCA CUGAUGAG X CGAA IAGGAGUG 3736
2292 ACTCCTCC T GCATATAG 1228 CUAUAUGC CUGAUGAG X CGAA IGAGGAGU 3737
2295 CCTCCTGC A TATAGACC 1229 GGUCUAUA CUGAUGAG X CGAA ICAGGAGG 3738
2303 ATATAGAC C ACCAAATG 1230 CAUUUGGU CUGAUGAG X CGAA IUCUAUAU 3739
2304 TATAGACC A CCAAATGC 1231 GCAUUUGG CUGAUGAG X CGAA IGUCUAUA 3740
2306 TAGACCAC C AAATGCCC 1232 GGGCAUUU CUGAUGAG X CGAA IUGGUCUA 3741
2307 AGACCACC A AATGCCCC 1233 GGGGCAUU CUGAUGAG X CGAA IGUGGUCU 3742
2313 CCAAATGC C CCTATCTT 1234 AAGAUAGG CUGAUGAG X CGAA ICAUUUGG 3743
2314 CAAATGCC C CTATCTTA 1235 UAAGAUAG CUGAUGAG X CGAA IGCAUUUG 3744
2315 AAATGCCC C TATCTTAT 1236 AUAAGAUA CUGAUGAG X CGAA IGGCAUUU 3745
2316 AATGCCCC T ATCTTATC 1237 GAUAAGAU CUGAUGAG X CGAA IGGGCAUU 3746
2320 CCCCTATC T TATCAACA 1238 UGUUGAUA CUGAUGAG X CGAA IAUAGGGG 3747
2325 ATCTTATC A ACACTTCC 1239 GGAAGUGU CUGAUGAG X CGAA IAUAAGAU 3748
2328 TTATCAAC A CTTCCGGA 1240 UCCGGAAG CUGAUGAG X CGAA lUUGAUAA 3749
2330 ATCAACAC T TCCGGAAA 1241 UUUCCGGA CUGAUGAG X CGAA IUGUUGAU 3750
2333 AACACTTC C GGAAACTA 1242 UAGUUUCC CUGAUGAG X CGAA IAAGUGUU 3751
2340 CCGGAAAC T ACTGTTGT 1243 ACAACAGU CUGAUGAG X CGAA IUUUCCGG 3752
2343 GAAACTAC T GTTGTTAG 1244 CUAACAAC CUGAUGAG X CGAA IUAGUUUC 3753
2362 GAAGAGGC A GGTCCCCT 1245 AGGGGACC CUGAUGAG X CGAA ICCUCUUC 3754
2367 GGCAGGTC C CCTAGAAG 1246 CUUCUAGG CUGAUGAG X CGAA IACCUGCC 3755
2368 GCAGGTCC c CTAGAAGA 1247 UCUUCUAG CUGAUGAG X CGAA IGACCUGC 3756
2369 CAGGTCCC c TAGAAGAA 1248 UUCUUCUA CUGAUGAG X CGAA IGGACCUG 3757
2370 AGGTCCCC T AGAAGAAG 1249 CUUCUUCU CUGAUGAG X CGAA IGGGACCU 3758
2382 AGAAGAAC T CCCTCGCC 1250 GGCGAGGG CUGAUGAG X CGAA IUUCUUCU 3759
2384 AAGAACTC c CTCGCCTC 1251 GAGGCGAG CUGAUGAG X CGAA IAGUUCUU 3760
2385 AGAACTCC c TCGCCTCG 1252 CGAGGCGA CUGAUGAG X CGAA IGAGUUCU 3761
2386 GAACTCCC T CGCCTCGC 1253 GCGAGGCG CUGAUGAG X CGAA IGGAGUUC 3762
2390 TCCCTCGC c TCGCAGAC 1254 GUCUGCGA CUGAUGAG X CGAA ICGAGGGA 3763
2391 CCCTCGCC T CGCAGACG 1255 CGUCUGCG CUGAUGAG X CGAA IGCGAGGG 3764
2395 CGCCTCGC A GACGAAGG 1256 CCUUCGUC CUGAUGAG X CGAA ICGAGGCG 3765
2406 CGAAGGTC T CAATCGCC 1257 GGCGAUUG CUGAUGAG X CGAA IACCUUCG 3766
2408 AAGGTCTC A ATCGCCGC 1258 GCGGCGAU CUGAUGAG X CGAA IAGACCUU 3767
Table 38
2414 TCAATCGC C GCGTCGCA 1259 UGCGACGC CUGAUGAG X CGAA ICGAUUGA 3768
2422 CGCGTCGC A GAAGATCT 1260 AGAUCUUC CUGAUGAG X CGAA ICGACGCG 3769
2430 AGAAGATC T CAATCTCG 1261 CGAGAUUG CUGAUGAG X CGAA IAUCUUCU 3770
2432 AAGATCTC A ATCTCGGG 1262 CCCGAGAU CUGAUGAG X CGAA IAGAUCUU 3771
2436 TCTCAATC T CGGGAATC 1263 GAUUCCCG CUGAUGAG X CGAA IAUUGAGA 3772
2445 CGGGAATC T CAATGTTA 1264 UAACAUUG CUGAUGAG X CGAA IAUUCCCG 3773
2447 GGAATCTC A ATGTTAGT 1265 ACUAACAU CUGAUGAG X CGAA IAGAUUCC 3774
2460 TAGTATTC C TTGGACAC 1266 GUGUCCAA CUGAUGAG X CGAA IAAUACUA 3775
2461 AGTATTCC T TGGACACA 1267 UGUGUCCA CUGAUGAG X CGAA IGAAUACU 3776
2467 CCTTGGAC A CATAAGGT 1268 ACCUUAUG CUGAUGAG X CGAA IUCCAAGG 3777
2469 TTGGACAC A TAAGGTGG 1269 CCACCUUA CUGAUGAG X CGAA IUGUCCAA 3778
2483 TGGGAAAC T TTACGGGG 1270 CCCCGUAA CUGAUGAG X CGAA IUUUCCCA 3779
2493 TACGGGGC T TTATTCTT 1271 AAGAAUAA CUGAUGAG X CGAA ICCCCGUA 3780
2500 CTTTATTC T TCTACGGT 1272 ACCGUAGA CUGAUGAG X CGAA IAAUAAAG 3781
2503 TATTCTTC T ACGGTACC 1273 GGUACCGU CUGAUGAG X CGAA IAAGAAUA 3782
2511 TACGGTAC C TTGCTTTA 1274 UAAAGCAA CUGAUGAG X CGAA IUACCGUA 3783
2512 ACGGTACC T TGCTTTAA 1275 UUAAAGCA CUGAUGAG X CGAA IGUACCGU 3784
2516 TACCTTGC T TTAATCCT 1276 AGGAUUAA CUGAUGAG X CGAA ICAAGGUA 3785
2523 CTTTAATC c TAAATGGC 1277 GCCAUUUA CUGAUGAG X CGAA IAUUAAAG 3786
2524 TTTAATCC T AAATGGCA 1278 UGCCAUUU CUGAUGAG X CGAA IGAUUAAA 3787
2532 TAAATGGC A AACTCCTT 1279 AAGGAGUU CUGAUGAG X CGAA ICCAUUUA 3788
2536 TGGCAAAC T CCTTCTTT 1280 AAAGAAGG CUGAUGAG X CGAA IUUUGCCA 3789
2538 GCAAACTC C TTCTTTTC 1281 GAAAAGAA CUGAUGAG X CGAA IAGUUUGC 3790
2539 CAAACTCC T TCTTTTCC 1282 GGAAAAGA CUGAUGAG X CGAA IGAGUUUG 3791
2542 ACTCCTTC T TTTCCTGA 1283 UCAGGAAA CUGAUGAG X CGAA IAAGGAGU 3792
2547 TTCTTTTC c TGACATTC 1284 GAAUGUCA CUGAUGAG X CGAA IAAAAGAA 3793
2548 TCTTTTCC T GACATTCA 1285 UGAAUGUC CUGAUGAG X CGAA IGAAAAGA 3794
2552 TTCCTGAC A TTCATTTG 1286 CAAAUGAA CUGAUGAG X CGAA IUCAGGAA 3795
2556 TGACATTC A TTTGCAGG 1287 CCUGCAAA CUGAUGAG X CGAA IAAUGUCA 3796
2562 TCATTTGC A GGAGGACA 1288 UGUCCUCC CUGAUGAG X CGAA ICAAAUGA 3797
2570 AGGAGGAC A TTGTTGAT 1289 AUCAACAA CUGAUGAG X CGAA IUCCUCCU 3798
2589 ATGTAAGC A ATTTGTGG 1290 CCACAAAU CUGAUGAG X CGAA ICUUACAU 3799
2601 TGTGGGGC C CCTTACAG 1291 CUGUAAGG CUGAUGAG X CGAA ICCCCACA 3800
2602 GTGGGGCC C CTTACAGT 1292 ACUGUAAO CUGAUGAG X CGAA IGCCCCAC 3801
2603 TGGGGCCC C TTACAGTA 1293 UACUGUAA CUGAUGAG X CGAA IGGCCCCA 3802
2604 GGGGCCCC T TACAGTAA 1294 UUACUGUA CUGAUGAG X CGAA IGGGCCCC 3803
2608 CCCCTTAC A GTAAATGA 1295 UCAUUUAC CUGAUGAG X CGAA IUAAGGGG 3804
2621 ATGAAAAC A GGAGACTT 1296 AAGUCUCC CUGAUGAG X CGAA lUUUUCAU 3805
2628 CAGGAGAC T TAAATTAA 1297 UUAAUUUA CUGAUGAG X CGAA IUCUCCUG 3806
2638 AAATTAAC T ATGCCTGC 1298 GCAGGCAU CUGAUGAG X CGAA IUUAAUUU 3807
2643 AACTATGC C TGCTAGGT 1299 ACCUAGCA CUGAUGAG X CGAA ICAUAGUU 3808
2644 ACTATGCC T GCTAGGTT 1300 AACCUAGC CUGAUGAG X CGAA IGCAUAGU 3809
2647 ATGCCTGC T AGGTTTTA 1301 UAAAACCU CUGAUGAG X CGAA ICAGGCAU 3810
2658 GTTTTATC C CAATGTTA 1302 UAACAUUG CUGAUGAG X CGAA IAUAAAAC 3811
2659 TTTTATCC C AATGTTAC 1303 GUAACAUU CUGAUGAG X CGAA IGAUAAAA 3812
2660 TTTATCCC A ATGTTACT 1304 AGUAACAU CUGAUGAG X CGAA IGGAUAAA 3813
2668 AATGTTAC T AAATATTT 1305 AAAUAUUU CUGAUGAG X CGAA IUAACAUU 3814
2679 ATATTTGC C CTTAGATA 1306 UAUCUAAG CUGAUGAG X CGAA ICAAAUAU 3815
2680 TATTTGCC C TTAGATAA 1307 UUAUCUAA CUGAUGAG X CGAA IGCAAAUA 3816
2681 ATTTGCCC T TAGATAAA 1308 UUUAUCUA CUGAUGAG X CGAA IGGCAAAU 3817
2696 AAGGGATC A AACCGTAT 1309 AUACGGUU CUGAUGAG X CGAA IAUCCCUU 3818
Table 38
2700 GATCAAAC C GTATTATC 1310 GAUAAUAC CUGAUGAG X CGAA IUUUGAUC 3819
2709 GTATTATC C AGAGTATG 1311 CAUACUCU CUGAUGAG X CGAA IAUAAUAC 3820
2710 TATTATCC A GAGTATGT 1312 ACAUACUC CUGAUGAG X CGAA IGAUAAUA 3821
2727 AGTTAATC A TTACTTCC 1313 GGAAGUAA CUGAUGAG X CGAA IAUUAACU 3822
2732 ATCATTAC T TCCAGACG 1314 CGUCUGGA CUGAUGAG X CGAA IUAAUGAU 3823
2735 ATTACTTC C AGACGCGA 1315 UCGCGUCU CUGAUGAG X CGAA IAAGUAAU 3824
2736 TTACTTCC A GACGCGAC 1316 GUCGCGUC CUGAUGAG X CGAA IGAAGUAA 3825
2745 GACGCGAC A TTATTTAC 1317 GUAAAUAA CUGAUGAG X CGAA IUCGCGUC 3826
2754 TTATTTAC A CACTCTTT 1318 AAAGAGUG CUGAUGAG X CGAA IUAAAUAA 3827
2756 ATTTACAC A CTCTTTGG 1319 CCAAAGAG CUGAUGAG X CGAA IUGUAAAU 3828
2758 TTACACAC T CTTTGGAA 1320 UUCCAAAG CUGAUGAG X CGAA IUGUGUAA 3829
2760 ACACACTC T TTGGAAGG 1321 CCUUCCAA CUGAUGAG X CGAA IAGUGUGU 3830
2777 CGGGGATC T TATATAAA 1322 UUUAUAUA CUGAUGAG X CGAA IAUCCCCG 3831
2794 AGAGAGTC C ACACGTAG 1323 CUACGUGU CUGAUGAG X CGAA IACUCUCU 3832
2795 GAGAGTCC A CACGTAGC 1324 GCUACGUG CUGAUGAG X CGAA IGACUCUC 3833
2797 GAGTCCAC A CGTAGCGC 1325 GCGCUACG CUGAUGAG X CGAA IUGGACUC 3834
2806 CGTAGCGC C TCATTTTG 1326 CAAAAUGA CUGAUGAG X CGAA ICGCUACG 3835
2807 GTAGCGCC T CATTTTGC 1327 GCAAAAUG CUGAUGAG X CGAA IGCGCUAC 3836
2809 AGCGCCTC A TTTTGCGG 1328 CCGCAAAA CUGAUGAG X CGAA IAGGCGCU 3837
2821 TGCGGGTC A CCATATTC 1329 GAAUAUGG CUGAUGAG X CGAA IACCCGCA 3838
2823 CGGGTCAC C ATATTCTT 1330 AAGAAUAU CUGAUGAG X CGAA IUGACCCG 3839
2824 GGGTCACC A TATTCTTG 1331 CAAGAAUA CUGAUGAG X CGAA IGUGACCC 3840
2830 CCATATTC T TGGGAACA 1332 UGUUCCCA CUGAUGAG X CGAA IAAUAUGG 3841
2838 TTGGGAAC A AGATCTAC 1333 GUAGAUCU CUGAUGAG X CGAA IUUCCCAA 3842
2844 ACAAGATC T ACAGCATG 1334 CAUGCUGU CUGAUGAG X CGAA IAUCUUGU 3843
2847 AGATCTAC A GCATGGGA 1335 UCCCAUGC CUGAUGAG X CGAA IUAGAUCU 3844
2850 TCTACAGC. A TGGGAGGT 1336 ACCUCCCA CUGAUGAG X CGAA ICUGUAGA 3845
2864 GGTTGGTC T TCCAAACC 1337 GGUUUGGA CUGAUGAG X CGAA IACCAACC 3846
2867 TGGTCTTC C AAACCTCG 1338 CGAGGUUU CUGAUGAG X CGAA IAAGACCA 3847
2868 GGTCTTCC A AACCTCGA 1339 UCGAGGUU CUGAUGAG X CGAA IGAAGACC 3848
2872 TTCCAAAC C TCGAAAAG 1340 CUUUUCGA CUGAUGAG X CGAA IUUUGGAA 3849
2873 TCCAAACC T CGAAAAGG 1341 CCUUUUCG CUGAUGAG X CGAA IGUUUGGA 3850
2883 GAAAAGGC A TGGGGACA 1342 UGUCCCCA CUGAUGAG X CGAA ICCUUUUC 3851
2891 ATGGGGAC A AATCTTTC 1343 GAAAGAUU CUGAUGAG X CGAA IUCCCCAU 3852
2896 GACAAATC T TTCTGTCC 1344 GGACAGAA CUGAUGAG X CGAA IAUUUGUC 3853
2900 AATCTTTC T GTCCCCAA 1345 UUGGGGAC CUGAUGAG X CGAA IAAAGAUU 3854
2904 TTTCTGTC C CCAATCCC 1346 GGGAUUGG CUGAUGAG X CGAA IACAGAAA 3855
2905 TTCTGTCC C CAATCCCC 1347 GGGGAUUG CUGAUGAG X CGAA IGACAGAA 3856
2906 TCTGTCCC C AATCCCCT 1348 AGGGGAUU CUGAUGAG X CGAA IGGACAGA 3857
2907 CTGTCCCC A ATCCCCTG 1349 CAGGGGAU CUGAUGAG X CGAA IGGGACAG 3858
2911 CCCCAATC C CCTGGGAT 1350 AUCCCAGG CUGAUGAG X CGAA IAUUGGGG 3859
2912 CCCAATCC C CTGGGATT 1351 AAUCCCAG CUGAUGAG X CGAA IGAUUGGG 3860
2913 CCAATCCC C TGGGATTC 1352 GAAUCCCA CUGAUGAG X CGAA IGGAUUGG 3861
2914 CAATCCCC T GGGATTCT 1353 AGAAUCCC CUGAUGAG X CGAA IGGGAUUG 3862
2922 TGGGATTC T TCCCCGAT 1354 AUCGGGGA CUGAUGAG X CGAA IAAUCCCA 3863
2925 GATTCTTC c CCGATCAT 1355 AUGAUCGG CUGAUGAG X CGAA IAAGAAUC 3864
2926 ATTCTTCC c CGATCATC 1356 GAUGAUCG CUGAUGAG X CGAA IGAAGAAU 3865
2927 TTCTTCCC c GATCATCA 1357 UGAUGAUC CUGAUGAG X CGAA IGGAAGAA 3866
2932 CCCCGATC A TCAGTTGG 1358 CCAACUGA CUGAUGAG X CGAA IAUCGGGG 3867
2935 CGATCATC A GTTGGACC 1359 GGUCCAAC CUGAUGAG X CGAA IAUGAUCG 3868
2943 AGTTGGAC C CTGCATTC 1360 GAAUGCAG CUGAUGAG X CGAA IUCCAACU 3869
Table 38
2944 GTTGGACC C TGCATTCA 1361 UGAAUGCA CUGAUGAG X CGAA IGUCCAAC 3870
2945 TTGGACCC T GCATTCAA 1362 UUGAAUGC CUGAUGAG X CGAA IGGUCCAA 3871
2948 GACCCTGC A TTCAAAGC 1363 GCUUUGAA CUGAUGAG X CGAA ICAGGGUC 3872
2952 CTGCATTC A AAGCCAAC 1364 GUUGGCUU CUGAUGAG X CGAA IAAUGCAG 3873
2957 TTCAAAGC C AACTCAGT 1365 ACUGAGUU CUGAUGAG X CGAA ICUUUGAA 3874
2958 TCAAAGCC A ACTCAGTA 1366 UACUGAGU CUGAUGAG X CGAA IGCUUUGA 3875
2961 AAGCCAAC T CAGTAAAT 1367 AUUUACUG CUGAUGAG X CGAA IUUGGCUU 3876
2963 GCCAACTC A GTAAATCC 1368 GGAUUUAC CUGAUGAG X CGAA IAGUUGGC 3877
2971 AGTAAATC C AGATTGGG 1369 CCCAAUCU CUGAUGAG X CGAA IAUUUACU 3878
2972 GTAAATCC A GATTGGGA 1370 UCCCAAUC CUGAUGAG X CGAA IGAUUUAC 3879
2982 ATTGGGAC C TCAACCCG 1371 CGGGUUGA CUGAUGAG X CGAA IUCCCAAU 3880
2983 TTGGGACC T CAACCCGC 1372 GCGGGUUG CUGAUGAG X CGAA IGUCCCAA 3881
2985 GGGACCTC A ACCCGCAC 1373 GUGCGGGU CUGAUGAG X CGAA IAGGUCCC 3882
2988 ACCTCAAC C CGCACAAG 1374 CUUGUGCG CUGAUGAG X CGAA IUUGAGGU 3883
2989 CCTCAACC C GCACAAGG 1375 CCUUGUGC CUGAUGAG X CGAA IGUUGAGG 3884
2992 CAACCCGC A CAAGGACA 1376 UGUCCUUG CUGAUGAG X CGAA ICGGGUUG 3885
2994 ACCCGCAC A AGGACAAC 1377 GUUGUCCU CUGAUGAG X CGAA IUGCGGGU 3886
3000 ACAAGGAC A ACTGGCCG 1378 CGGCCAGU CUGAUGAG X CGAA IUCCUUGU 3887
3003 AGGACAAC T GGCCGGAC 1379 GUCCGGCC CUGAUGAG X CGAA IUUGUCCU 3888
3007 CAACTGGC C GGACGCCA 1380 UGGCGUCC CUGAUGAG X CGAA ICCAGUUG 3889
3014 CCGGACGC C AACAAGGT 1381 ACCUUGUU CUGAUGAG X CGAA ICGUCCGG 3890
3015 CGGACGCC A ACAAGGTG 1382 CACCUUGU CUGAUGAG X CGAA IGCGUCCG 3891
3018 ACGCCAAC A AGGTGGGA 1383 UCCCACCU CUGAUGAG X CGAA IUUGGCGU 3892
3035 GTGGGAGC A TTCGGGCC 1384 GGCCCGAA CUGAUGAG X CGAA ICUCCCAC 3893
3043 ATTCGGGC C AGGGTTCA 1385 UGAACCCU CUGAUGAG X CGAA ICCCGAAU 3894
3044 TTCGGGCC A GGGTTCAC 1386 GUGAACCC CUGAUGAG X CGAA IGCCCGAA 3895
3051 CAGGGTTC A CCCCTCCC 1387 GGGAGGGG CUGAUGAG X CGAA IAACCCUG 3896
3053 GGGTTCAC C CCTCCCCA 1388 UGGGGAGG CUGAUGAG X CGAA IUGAACCC 3897
3054 GGTTCACC C CTCCCCAT 1389 AUGGGGAG CUGAUGAG X CGAA IGUGAACC 3898
3055 GTTCACCC C TCCCCATG 1390 CAUGGGGA CUGAUGAG X CGAA IGGUGAAC 3899
3056 TTCACCCC T CCCCATGG 1391 CCAUGGGG CUGAUGAG X CGAA IGGGUGAA 3900
3058 CACCCCTC C CCATGGGG 1392 CCCCAUGG CUGAUGAG X CGAA IAGGGGUG 3901
3059 ACCCCTCC C CATGGGGG 1393 CCCCCAUG CUGAUGAG X CGAA IGAGGGGU 3902
3060 CCCCTCCC C ATGGGGGA 1394 UCCCCCAU CUGAUGAG X CGAA IGGAGGGG 3903
3061 CCCTCCCC A TGGGGGAC 1395 GUCCCCCA CUGAUGAG X CGAA IGGGAGGG 3904
3070 TGGGGGAC T GTTGGGGT 1396 ACCCCAAC CUGAUGAG X CGAA lUCCCCCA 3905
3084 GGTGGAGC c CTCACGCT 1397 AGCGUGAG CUGAUGAG X CGAA ICUCCACC 3906
3085 GTGGAGCC c TCACGCTC 1398 GAGCGUGA CUGAUGAG X CGAA IGCUCCAC 3907
3086 TGGAGCCC T CACGCTCA 1399 UGAGCGUG CUGAUGAG X CGAA IGGCUCCA 3908
3088 GAGCCCTC A CGCTCAGG 1400 CCUGAGCG CUGAUGAG X CGAA IAGGGCUC 3909
3092 CCTCACGC T CAGGGCCT 1401 AGGCCCUG CUGAUGAG X CGAA ICGUGAGG 3910
3094 TCACGCTC A GGGCCTAC 1402 GUAGGCCC CUGAUGAG X CGAA IAGCGUGA 3911
3099 CTCAGGGC C TACTCACA 1403 UGUGAGUA CUGAUGAG X CGAA ICCCUGAG 3912
3100 TCAGGGCC T ACTCACAA 1404 UUGUGAGU CUGAUGAG X CGAA IGCCCUGA 3913
3103 GGGCCTAC T CACAACTG 1405 CAGUUGUG CUGAUGAG X CGAA IUAGGCCC 3914
3105 GCCTACTC A CAACTGTG 1406 CACAGUUG CUGAUGAG X CGAA IAGUAGGC 3915
3107 CTACTCAC A ACTGTGCC 1407 GGCACAGU CUGAUGAG X CGAA IUGAGUAG 3916
3110 CTCACAAC T GTGCCAGC 1408 GCUGGCAC CUGAUGAG X CGAA IUUGUGAG 3917
3115 AACTGTGC C AGCAGCTC 1409 GAGCUGCU CUGAUGAG X CGAA ICACAGUU 3918
3116 ACTGTGCC A GCAGCTCC 1410 GGAGCUGC CUGAUGAG X CGAA IGCACAGU 3919
3119 GTGCCAGC A GCTCCTCC 1411 GGAGGAGC CUGAUGAG X CGAA ICUGGCAC 3920
Table 38
3122 CCAGCAGC T CCTCCTCC 1412 GGAGGAGG CUGAUGAG X CGAA ICUGCUGG 3921
3124 AGCAGCTC C TCCTCCTG 1413 CAGGAGGA CUGAUGAG X CGAA IAGCUGCU 3922
3125 GCAGCTCC T CCTCCTGC 1414 GCAGGAGG CUGAUGAG X CGAA IGAGCUGC 3923
3127 AGCTCCTC C TCCTGCCT 1415 AGGCAGGA CUGAUGAG X CGAA IAGGAGCU 3924
3128 GCTCCTCC T CCTGCCTC 1416 GAGGCAGG CUGAUGAG X CGAA IGAGGAGC 3925
3130 TCCTCCTC c TGCCTCCA 1417 UGGAGGCA CUGAUGAG X CGAA IAGGAGGA 3926
3131 CCTCCTCC T GCCTCCAC 1418 GUGGAGGC CUGAUGAG X CGAA IGAGGAGG 3927
3134 CCTCCTGC c TCCACCAA 1419 UUGGUGGA CUGAUGAG X CGAA ICAGGAGG 3928
3135 CTCCTGCC T CCACCAAT 1420 AUUGGUGG CUGAUGAG X CGAA IGCAGGAG 3929
3137 CCTGCCTC c ACCAATCG 1421 CGAUUGGU CUGAUGAG X CGAA IAGGCAGG 3930
3138 CTGCCTCC A CCAATCGG 1422 CCGAUUGG CUGAUGAG X CGAA IGAGGCAG 3931
3140 GCCTCCAC C AATCGGCA 1423 UGCCGAUU CUGAUGAG X CGAA IUGGAGGC 3932
3141 CCTCCACC A ATCGGCAG 1424 CUGCCGAU CUGAUGAG X CGAA IGUGGAGG 3933
3148 CAATCGGC A GTCAGGAA 1425 UUCCUGAC CUGAUGAG X CGAA ICCGAUUG 3934
3152 CGGCAGTC A GGAAGGCA 1426 UGCCUUCC CUGAUGAG X CGAA IACUGCCG 3935
3160 AGGAAGGC A GCCTACTC 1427 GAGUAGGC CUGAUGAG X CGAA ICCUUCCU 3936
3163 AAGGCAGC C TACTCCCT 1428 AGGGAGUA CUGAUGAG X CGAA ICUGCCUU 3937
3164 AGGCAGCC T ACTCCCTT 1429 AAGGGAGU CUGAUGAG X CGAA IGCUGCCU 3938
3167 CAGCCTAC T CCCTTATC 1430 GAUAAGGG CUGAUGAG X CGAA IUAGGCUG 3939
3169 GCCTACTC C CTTATCTC 1431 GAGAUAAG CUGAUGAG X CGAA IAGUAGGC 3940
3170 CCTACTCC C TTATCTCC 1432 GGAGAUAA CUGAUGAG X CGAA IGAGUAGG 3941
3171 CTACTCCC T TATCTCCA 1433 UGGAGAUA CUGAUGAG X CGAA IGGAGUAG 3942
3176 CCCTTATC T CCACCTCT 1434 AGAGGUGG CUGAUGAG X CGAA lAUAAGGG 3943
3178 CTTATCTC C ACCTCTAA 1435 UUAGAGGU CUGAUGAG X CGAA IAGAUAAG 3944
3179 TTATCTCC A CCTCTAAG 1436 CUUAGAGG CUGAUGAG X CGAA IGAGAUAA 3945
3181 ATCTCCAC C TCTAAGGG 1437 CCCUUAGA CUGAUGAG X CGAA IUGGAGAU 3946
3182 TCTCCACC T CTAAGGGA 1438 UCCCUUAG CUGAUGAG X CGAA IGUGGAGA 3947
3184 TCCACCTC T AAGGGACA 1439 UGUCCCUU CUGAUGAG X CGAA IAGGUGGA 3948
3192 TAAGGGAC A CTCATCCT 1440 AGGAUGAG CUGAUGAG X CGAA IUCCCUUA 3949
3194 AGGGACAC T CATCCTCA 1441 UGAGGAUG CUGAUGAG X CGAA IUGUCCCU 3950
3196 GGACACTC A TCCTCAGG 1442 CCUGAGGA CUGAUGAG X CGAA IAGUGUCC 3951
3199 CACTCATC C TCAGGCCA 1443 UGGCCUGA CUGAUGAG X CGAA IAUGAGUG 3952
3200 ACTCATCC T CAGGCCAT 1444 AUGGCCUG CUGAUGAG X CGAA IGAUGAGU 3953
3202 TCATCCTC A GGCCATGC 1445 GCAUGGCC CUGAUGAG X CGAA IAGGAUGA 3954
3206 CCTCAGGC C ATGCAGTG 1446 CACUGCAU CUGAUGAG X CGAA ICCUGAGG 3955
3207 CTCAGGCC A TGCAGTGG 1447 CCACUGCA CUGAUGAG X CGAA IGCCUGAG 3956
Input Sequence = AF100308. Cut Site = CH/ .
Stem Length = 8 . Core Sequence = CUGAUGAG X CGAA (X = GCCGUUAGGC or other stem II)
AF100308 (Hepatitis B virus strain 2-18, 3215 bp)
Table 39
Table 39: Human HBV G-cleaver Ribozyme and Substrate Sequence
Pos Substrate Seq ID Ribozyme Rz Seq ID
61 ACUUUCCU G CUGGUGGC 1448 GCCACCAG UGAUG GCAUGCACUAUGC GCG AGGAAAGU 3957
87 GGAACAGU G AGCCCUGC 1449 GCAGGGCU UGAUG GCAUGCACUAUGC GCG ACUGUUCC 3958
94 UGAGCCCU G CUCAGAAU 1450 AUUCUGAG UGAUG GCAUGCACUAUGC GCG AGGGCUCA 3959
112 CUGUCUCU G CCAUAUCG 1451 CGAUAUGG UGAUG GCAUGCACUAUGC GCG AGAGACAG 3960
132 AUCUUAUC G AAGACUGG 1452 CCAGUCUU UGAUG GCAUGCACUAUGC GCG GAUAAGAU 3961
153 CCUGUACC G AACAUGGA 1453 UCCAUGUU UGAUG GCAUGCACUAUGC GCG GGUACAGG 3962
169 AGAACAUC G CAUCAGGA 1454 UCCUGAUG UGAUG GCAUGCACUAUGC GCG GAUGUUCU 3963
192 GGACCCCU G CUCGUGUU 1455 AACACGAG UGAUG GCAUGCACUAUGC GCG AGGGGUCC 3964
222 UUCUUGUU G ACAAAAAU 1456 AUUUUUGU UGAUG GCAUGCACUAUGC GCG AACAAGAA 3965
315 CAAAAUUC G CAGUCCCA 1457 UGGGACUG UGAUG GCAUGCACUAUGC GCG GAAUUUUG 3966
374 UGGUUAUC G CUGGAUGU 1458 ACAUCCAG UGAUG GCAUGCACUAUGC GCG GAUAACCA 3967
387 AUGUGUCU G CGGCGUUU 1459 AAACGCCG UGAUG GCAUGCACUAUGC GCG AGACACAU 3968
410 CUUCCUCU G CAUCCUGC 1460 GCAGGAUG UGAUG GCAUGCACUAUGC GCG AGAGGAAG 3969
417 UGCAUCCU G CUGCUAUG 1461 CAUAGCAG UGAUG GCAUGCACUAUGC GCG AGGAUGCA 3970
420 AUCCUGCU G CUAUGCCU 1462 AGGCAUAG UGAUG GCAUGCACUAUGC GCG AGCAGGAU 3971
425 GCUGCUAU G CCUCAUCU 1463 AGAUGAGG UGAUG GCAUGCACUAUGC GCG AUAGCAGC 3972
468 GGUAUGUU G CCCGUUUG 1464 CAAACGGG UGAUG GCAUGCACUAUGC GCG AACAUACC 3973
518 CGGACCAU G CAAAACCU 1465 AGGUUUUG UGAUG GCAUGCACUAUGC GCG AUGGUCCG 3974
527 CAAAACCU G CACAACUC 1466 GAGUUGUG UGAUG GCAUGCACUAUGC GCG AGGUUUUG 3975
538 CAACUCCU G CUCAAGGA 1467 UCCUUGAG UGAUG GCAUGCACUAUGC GCG AGGAGUUG 3976
569 CUCAUGUU G CUGUACAA 1468 UUGUACAG UGAUG GCAUGCACUAUGC GCG AACAUGAG 3977
596 CGGAAACU G CACCUGUA 1469 UACAGGUG UGAUG GCAUGCACUAUGC GCG AGUUUCCG 3978
631 GGGCUUUC G CAAAAUAC 1470 GUAUUUUG UGAUG GCAUGCACUAUGC GCG GAAAGCCC 3979
687 UUACUAGU G CCAUUUGU 1471 ACAAAUGG UGAUG GCAUGCACUAUGC GCG ACUAGUAA 3980
747 AUAUGGAU G AUGUGGUU 1472 AACCACAU UGAUG GCAUGCACUAUGC GCG AUCCAUAU 3981
783 AACAUCUU G AGUCCCUU 1473 AAGGGACU UGAUG GCAUGCACUAUGC GCG AAGAUGUU 3982
795 CCCUUUAU G CCGCUGUU 1474 AACAGCGG UGAUG GCAUGCACUAUGC GCG AUAAAGGG 3983
798 UUUAUGCC G CUGUUACC 1475 GGUAACAG UGAUG GCAUGCACUAUGC GCG GGCAUAAA 3984
911 GGCACAUU G CCACAGGA 1476 UCCUGUGG UGAUG GCAUGCACUAUGC GCG AAUGUGCC 3985
978 GGCCUAUU G AUUGGAAA 1477 UUUCCAAU UGAUG GCAUGCACUAUGC GCG AAUAGGCC 3986
Table 39
997 AUGUCAAC G AAUUGUGG 1478 CCACAAUU UGAUG GCAUGCACUAUGC GCG GUUGACAU 3987
1020 UGGGGUUU G CCGCCCCU 1479 AGGGGCGG UGAUG GCAUGCACUAUGC GCG AAACCCCA 3988
1023 GGUUUGCC G CCCCUUUC 1480 GAAAGGGG UGAUG GCAUGCACUAUGC GCG GGCAAACC 3989
1034 CCUUUCAC G CAAUGUGG 1481 CCACAUUG UGAUG GCAUGCACUAUGC GCG GUGAAAGG 3990
1050 GAUAUUCU G CUUUAAUG 1482 CAUUAAAG UGAUG GCAUGCACUAUGC GCG AGAAUAUC 3991
1058 GCUUUAAU G CCUUUAUA 1483 UAUAAAGG UGAUG GCAUGCACUAUGC GCG AUUAAAGC 3992
1068 CUUUAUAU G CAUGCAUA 1484 UAUGCAUG UGAUG GCAUGCACUAUGC GCG AUAUAAAG 3993
1072 AUAUGCAU G CAUACAAG 1485 CUUGUAUG UGAUG GCAUGCACUAUGC GCG AUGCAUAU 3994
1103 ACUUUCUC G CCAACUUA 1486 UAAGUUGG UGAUG GCAUGCACUAUGC GCG GAGAAAGU 3995
1139 CAGUAUGU G AACCUUUA 1487 UAAAGGUU UGAUG GCAUGCACUAUGC GCG ACAUACUG 3996
1155 ACCCCGUU G CUCGGCAA 1488 UUGCCGAG UGAUG GCAUGCACUAUGC GCG AACGGGGU 3997
1177 UGGUCUAU G CCAAGUGU 1489 ACACUUGG UGAUG GCAUGCACUAUGC GCG AUAGACCA 3998
1188 AAGUGUUU G CUGACGCA 1490 UGCGUCAG UGAUG GCAUGCACUAUGC GCG AAACACUU 3999
1191 UGUUUGCU G ACGCAACC 1491 GGUUGCGU UGAUG GCAUGCACUAUGC GCG AGCAAACA 4000
1194 UUGCUGAC G CAACCCCC 1492 GGGGGUUG UGAUG GCAUGCACUAUGC GCG GUCAGCAA 4001
1234 CCAUCAGC G CAUGCGUG 1493 CACGCAUG UGAUG GCAUGCACUAUGC GCG GCUGAUGG 4002
1238 CAGCGCAU G CGUGGAAC 1494 GUUCCACG UGAUG GCAUGCACUAUGC GCG AUGCGCUG 4003
1262 UCUCCUCU G CCGAUCCA 1495 UGGAUCGG UGAUG GCAUGCACUAUGC GCG AGAGGAGA 4004
1265 CCUCUGCC G AUCCAUAC 1496 GUAUGGAU UGAUG GCAUGCACUAUGC GCG GGCAGAGG 4005
1275 UCCAUACC G CGGAACUC 1497 GAGUUCCG UGAUG GCAUGCACUAUGC GCG GGUAUGGA 4006
1290 UCCUAGCC G CUUGUUUU 1498 AAAACAAG UGAUG GCAUGCACUAUGC GCG GGCUAGGA 4007
1299 CUUGUUUU G CUCGCAGC 1499 GCUGCGAG UGAUG GCAUGCACUAUGC GCG AAAACAAG 4008
1303 UUUUGCUC G CAGCAGGU 1500 ACCUGCUG UGAUG GCAUGCACUAUGC GCG GAGCAAAA 4009
1335 UCGGGACU G AGAAUUCU 1501 AGAAUUGU UGAUG GCAUGCACUAUGC GCG AGUCCCGA 4010
1349 UCUGUCGU G CUCUCCCG 1502 CGGGAGAG UGAUG GCAUGCACUAUGC GCG ACGACAGA 4011
1357 GCUCUCCC G CAAAUAUA 1503 UAUAUUUG UGAUG GCAUGCACUAUGC GCG GGGAGAGC 4012
1382 CCAUGGCU G CUAGGCUG 1504 CAGCCUAG UGAUG GCAUGCACUAUGC GCG AGCCAUGG 4013
1392 UAGGCUGU G CUGCCAAC 1505 GUUGGCAG UGAUG GCAUGCACUAUGC GCG ACAGCCUA 4014
1395 GCUGUGCU G CCAACUGG 1506 CCAGUUGG UGAUG GCAUGCACUAUGC GCG AGCACAGC 4015
1411 GAUCCUAC G CGGGACGU 1507 ACGUCCCG UGAUG GCAUGCACUAUGC GCG GUAGGAUC 4016
1442 CCGUCGGC G CUGAAUCC 1508 GGAUUCAG UGAUG GCAUGCACUAUGC GCG GCCGACGG 4017
1445 UCGGCGCU G AAUCCCGC 1509 GCGGGAUU UGAUG GCAUGCACUAUGC GCG AGCGCCGA 4018
1452 UGAAUCCC G CGGACGAC 1510 GUCGUCCG UGAUG GCAUGCACUAUGC GCG GGGAUUCA 4019
1458 CCGCGGAC G ACCCCUCC 1511 GGAGGGGU UGAUG GCAUGCACUAUGC GCG GUCCGCGG 4020
Table 39
1474 CCGGGGCC G CUUGGGGC 1512 GCCCCAAG UGAUG GCAUGCACUAUGC GCG GGCCCCGG 4021
1489 GCUCUACC G CCCGCUUC 1513 GAAGCGGG UGAUG GCAUGCACUAUGC GCG GGUAGAGC 4022
1493 UACCGCCC G CUUCUCCG 1514 CGGAGAAG UGAUG GCAUGCACUAUGC GCG GGGCGGUA 4023
1501 GCUUCUCC G CCUAUUGU 1515 ACAAUAGG UGAUG GCAUGCACUAUGC GCG GGAGAAGC 4024
1513 AUUGUACC G ACCGUCCA 1516 UGGACGGU UGAUG GCAUGCACUAUGC GCG GGUACAAU 4025
1528 CACGGGGC G CACCUCUC 1517 GAGAGGUG UGAUG GCAUGCACUAUGC GCG GCCCCGUG 4026
1542 CUCUUUAC G CGGACUCC 1518 GGAGUCCG UGAUG GCAUGCACUAUGC GCG GUAAAGAG 4027
1559 CCGUCUGU G CCUUCUCA 1519 UGAGAAGG UGAUG GCAUGCACUAUGC GCG ACAGACGG 4028
1571 UCUCAUCU G CCGGACCG 1520 CGGUCCGG UGAUG GCAUGCACUAUGC GCG AGAUGAGA 4029
1583 GACCGUGU G CACUUCGC 1521 GCGAAGUG UGAUG GCAUGCACUAUGC GCG ACACGGUC 4030
1590 UGCACUUC G CUUCACCU 1522 AGGUGAAG UGAUG GCAUGCACUAUGC GCG GAAGUGCA 4031
1601 UCACCUCU G CACGUCGC 1523 GCGACGUG UGAUG GCAUGCACUAUGC GCG AGAGGUGA 4032
1608 UGCACGUC G CAUGGAGA 1524 UCUCCAUG UGAUG GCAUGCACUAUGC GCG GACGUGCA 4033
1624 ACCACCGU G AACGCCCA 1525 UGGGCGUU UGAUG GCAUGCACUAUGC GCG ACGGUGGU 4034
1628 CCGUGAAC G CCCACAGG 1526 CCUGUGGG UGAUG GCAUGCACUAUGC GCG GUUCACGG 4035
1642 AGGAACCU G CCCAAGGU 1527 ACCUUGGG UGAUG GCAUGCACUAUGC GCG AGGUUCCU 4036
1654 AAGGUCUU G CAUAAGAG 1528 CUCUUAUG UGAUG GCAUGCACUAUGC GCG AAGACCUU 4037
1690 AUGUCAAC G ACCGACCU 1529 AGGUCGGU UGAUG GCAUGCACUAUGC GCG GUUGACAU 4038
1694 CAACGACC G ACCUUGAG 1530 CUCAAGGU UGAUG GCAUGCACUAUGC GCG GGUCGUUG 4039
1700 CCGACCUU G AGGCAUAC 1531 GUAUGCCU UGAUG GCAUGCACUAUGC GCG AAGGUCGG 4040
1730 UGUUUAAU G AGUGGGAG 1532 CUCCCACU UGAUG GCAUGCACUAUGC GCG AUUAAACA 4041
1818 AGCACCAU G CAACUUUU 1533 AAAAGUUG UGAUG GCAUGCACUAUGC GCG AUGGUGCU 4042
1835 UCACCUCU G CCUAAUCA 1534 UGAUUAGG UGAUG GCAUGCACUAUGC GCG AGAGGUGA 4043
1883 CAAGCUGU G CCUUGGGU 1535 ACCCAAGG UGAUG GCAUGCACUAUGC GCG ACAGCUUG 4044
1912 UGGACAUU G ACCCGUAU 1536 AUACGGGU UGAUG GCAUGCACUAUGC GCG AAUGUCCA 4045
1959 UCUUUUUU G CCUUCUGA 1537 UCAGAAGG UGAUG GCAUGCACUAUGC GCG AAAAAAGA 4046
1966 UGCCUUCU G ACUUCUUU 1538 AAAGAAGU UGAUG GCAUGCACUAUGC GCG AGAAGGCA 4047
1985 UUCUAUUC G AGAUCUCC 1539 GGAGAUCU UGAUG GCAUGCACUAUGC GCG GAAUAGAA 4048
1996 AUCUCCUC G ACACCGCC 1540 GGCGGUGU UGAUG GCAUGCACUAUGC GCG GAGGAGAU 4049
2002 UCGACACC G CCUCUGCU 1541 AGCAGAGG UGAUG GCAUGCACUAUGC GCG GGUGUCGA 4050
2008 CCGCCUCU G CUCUGUAU 1542 AUACAGAG UGAUG GCAUGCACUAUGC GCG AGAGGCGG 4051
2092 GUUGGGGU G AGUUGAUG 1543 CAUCAACU UGAUG GCAUGCACUAUGC GCG ACCCCAAC 4052
2097 GGUGAGUU G AUGAAUCU 1544 AGAUUCAU UGAUG GCAUGCACUAUGC GCG AACUCACC 4053
2100 GAGUUGAU G AAUCUAGC 1545 GCUAGAUU UGAUG GCAUGCACUAUGC GCG AUCAACUC 4054
Table 39
2237 UUUUGGGC G AGAAACUG 1546 CAGUUUCU UGAUG GCAUGCACUAUGC GCG GCCCAAAA 4055
2251 CUGUUCUU G AAUAUUUG 1547 CAAAUAUU UGAUG GCAUGCACUAUGC GCG AAGAACAG 4056
2282 GUGGAUUC G CACUCCUC 1548 GAGGAGUG UGAUG GCAUGCACUAUGC GCG GAAUCCAC 4057
2293 CUCCUCCU G CAUAUAGA 1549 UCUAUAUG UGAUG GCAUGCACUAUGC GCG AGGAGGAG 4058
2311 CACCAAAU G CCCCUAUC 1550 GAUAGGGG UGAUG GCAUGCACUAUGC GCG AUUUGGUG 4059
2354 UGUUAGAC G AAGAGGCA 1551 UGCCUCUU UGAUG GCAUGCACUAUGC GCG GUCUAACA 4060
2388 ACUCCCUC G CCUCGCAG 1552 CUGCGAGG UGAUG GCAUGCACUAUGC GCG GAGGGAGU 4061
2393 CUCGCCUC G CAGACGAA 1553 UUCGUCUG UGAUG GCAUGCACUAUGC GCG GAGGCGAG 4062
2399 UCGCAGAC G AAGGUCUC 1554 GAGACCUU UGAUG GCAUGCACUAUGC GCG GUCUGCGA 4063
2412 UCUCAAUC G CCGCGUCG 1555 CGACGCGG UGAUG GCAUGCACUAUGC GCG GAUUGAGA 4064
2415 CAAUCGCC G CGUCGCAG 1556 CUGCGACG UGAUG GCAUGCACUAUGC GCG GGCGAUUG 4065
2420 GCCGCGUC G CAGAAGAU 1557 AUCUUCUG UGAUG GCAUGCACUAUGC GCG GACGCGGC 4066
2514 GGUACCUU G CUUUAAUC 1558 GAUUAAAG UGAUG GCAUGCACUAUGC GCG AAGGUACC 4067
2549 CUUUUCCU G ACAUUCAU 1559 AUGAAUGU UGAUG GCAUGCACUAUGC GCG AGGAAAAG 4068
2560 AUUCAUUU G CAGGAGGA 1560 UCCUCCUG UGAUG GCAUGCACUAUGC GCG AAAUGAAU 4069
2576 ACAUUGUU G AUAGAUGU 1561 ACAUCUAU UGAUG GCAUGCACUAUGC GCG AACAAUGU 4070
2615 CAGUAAAU G AAAACAGG 1562 CCUGUUUU UGAUG GCAUGCACUAUGC GCG AUUUACUG 4071
2641 UUAACUAU G CCUGCUAG 1563 CUAGCAGG UGAUG GCAUGCACUAUGC GCG AUAGUUAA 4072
2645 CUAUGCCU G CUAGGUUU 1564 AAACCUAG UGAUG GCAUGCACUAUGC GCG AGGCAUAG 4073
2677 AAAUAUUU G CCCUUAGA 1565 UCUAAGGG UGAUG GCAUGCACUAUGC GCG AAAUAUUU 4074
2740 UUCCAGAC G CGACAUUA 1566 UAAUGUCG UGAUG GCAUGCACUAUGC GCG GUCUGGAA 4075
2742 CCAGACGC G ACAUUAUU 1567 AAUAAUGU UGAUG GCAUGCACUAUGC GCG GCGUCUGG 4076
2804 CACGUAGC G CCUCAUUU 1568 AAAUGAGG UGAUG GCAUGCACUAUGC GCG GCUACGUG 4077
2814 CUCAUUUU G CGGGUCAC 1569 GUGACCCG UGAUG GCAUGCACUAUGC GCG AAAAUGAG 4078
2875 CAAACCUC G AAAAGGCA 1570 UGCCUUUU UGAUG GCAUGCACUAUGC GCG GAGGUUUG 4079
2928 UCUUCCCC G AUCAUCAG 1571 CUGAUGAU UGAUG GCAUGCACUAUGC GCG GGGGAAGA 4080
2946 UGGACCCU G CAUUCAAA 1572 UUUGAAUG UGAUG GCAUGCACUAUGC GCG AGGGUCCA 4081
2990 CUCAACCC G CACAAGGA 1573 UCCUUGUG UGAUG GCAUGCACUAUGC GCG GGGUUGAG 4082
3012 GGCCGGAC G CCAACAAG 1574 CUUGUUGG UGAUG GCAUGCACUAUGC GCG GUCCGGCC 4083
3090 GCCCUCAC G CUCAGGGC 1575 GCCCUGAG UGAUG GCAUGCACUAUGC GCG GUGAGGGC 4084
3113 ACAACUGU G CCAGCAGC 1576 GCUGCUGG UGAUG GCAUGCACUAUGC GCG ACAGUUGU 4085
3132 CUCCUCCU G CCUCCACC 1577 GGUGGAGG UGAUG GCAUGCACUAUGC GCG AGGAGGAG 4086
51 AGGGCCCU G UACUUUCC 1578 GGAAAGUA UGAUG GCAUGCACUAUGC GCG AGGGCCCU 4087
106 AGAAUACU G UCUCUGCC 1579 GGCAGAGA UGAUG GCAUGCACUAUGC GCG AGUAUUCU 4088
Table 39
148 GGGACCCU G UACCGAAC 1580 GUUCGGUA UGAUG GCAUGCACUAUGC GCG AGGGUCCC 4089
198 CUGCUCGU G UUACAGGC 1581 GCCUGUAA UGAUG GCAUGCACUAUGC GCG ACGAGCAG 4090
219 UUUUUCUU G UUGACAAA 1582 UUUGUCAA UGAUG GCAUGCACUAUGC GCG AAGAAAAA 4091
297 ACACCCGU G UGUCUUGG 1583 CCAAGACA UGAUG GCAUGCACUAUGC GCG ACGGGUGU 4092
299 ACCCGUGU G UCUUGGCC 1584 GGCCAAGA UGAUG GCAUGCACUAUGC GCG ACACGGGU 4093
347 ACCAACCU G UUGUCCUC 1585 GAGGACAA UGAUG GCAUGCACUAUGC GCG AGGUUGGU 4094
350 AACCUGUU G UCCUCCAA 1586 UUGGAGGA UGAUG GCAUGCACUAUGC GCG AACAGGUU 4095
362 UCCAAUUU G UCCUGGUU 1587 AACCAGGA UGAUG GCAUGCACUAUGC GCG AAAUUGGA 4096
381 CGCUGGAU G UGUCUGCG 1588 CGCAGACA UGAUG GCAUGCACUAUGC GCG AUCCAGCG 4097
383 CUGGAUGU G UCUGCGGC 1589 GCCGCAGA UGAUG GCAUGCACUAUGC GCG ACAUCCAG 4098
438 AUCUUCUU G UUGGUUCU 1590 AGAACCAA UGAUG GCAUGCACUAUGC GCG AAGAAGAU 4099
465 CAAGGUAU G UUGCCCGU 1591 ACGGGCAA UGAUG GCAUGCACUAUGC GCG AUACCUUG 4100
476 GCCCGUUU G UCCUCUAA 1592 UUAGAGGA UGAUG GCAUGCACUAUGC GCG AAACGGGC 4101
555 ACCUCUAU G UUUCCCUC 1593 GAGGGAAA UGAUG GCAUGCACUAUGC GCG AUAGAGGU 4102
566 UCCCUCAU G UUGCUGUA 1594 UACAGCAA UGAUG GCAUGCACUAUGC GCG AUGAGGGA 4103
572 AUGUUGCU G UACAAAAC 1595 GUUUUGUA UGAUG GCAUGCACUAUGC GCG AGCAACAU 4104
602 CUGCACCU G UAUUCCCA 1596 UGGGAAUA UGAUG GCAUGCACUAUGC GCG AGGUGCAG 4105
694 UGCCAUUU G UUCAGUGG 1597 CCACUGAA UGAUG GCAUGCACUAUGC GCG AAAUGGCA 4106
724 CCCCCACU G UCUGGCUU 1598 AAGCCAGA UGAUG GCAUGCACUAUGC GCG AGUGGGGG 4107
750 UGGAUGAU G UGGUUUUG 1599 CAAAACCA UGAUG GCAUGCACUAUGC GCG AUCAUCCA 4108
771 CCAAGUCU G UACAACAU 1600 AUGUUGUA UGAUG GCAUGCACUAUGC GCG AGACUUGG 4109
801 AUGCCGCU G UUACCAAU 1601 AUUGGUAA UGAUG GCAUGCACUAUGC GCG AGCGGCAU 4110
818 UUUCUUUU G UCUUUGGG 1602 CCCAAAGA UGAUG GCAUGCACUAUGC GCG AAAAGAAA 4111
888 UGGGAUAU G UAAUUGGG 1603 CCCAAUUA UGAUG GCAUGCACUAUGC GCG AUAUCCCA 4112
927 AACAUAUU G UACAAAAA 1604 UUUUUGUA UGAUG GCAUGCACUAUGC GCG AAUAUGUU 4113
944 AUCAAAAU G UGUUUUAG 1605 CUAAAACA UGAUG GCAUGCACUAUGC GCG AUUUUGAU 4114
946 CAAAAUGU G UUUUAGGA 1606 UCCUAAAA UGAUG GCAUGCACUAUGC GCG ACAUUUUG 4115
963 AACUUCCU G UAAACAGG 1607 CCUGUUUA UGAUG GCAUGCACUAUGC GCG AGGAAGUU 4116
991 GAAAGUAU G UCAACGAA 1608 UUCGUUGA UGAUG GCAUGCACUAUGC GCG AUACUUUC 4117
1002 AACGAAUU G UGGGUCUU 1609 AAGACCCA UGAUG GCAUGCACUAUGC GCG AAUUCGUU 4118
1039 CACGCAAU G UGGAUAUU 1610 AAUAUCCA UGAUG GCAUGCACUAUGC GCG AUUGCGUG 4119
1137 AACAGUAU G UGAACCUU 1611 AAGGUUCA UGAUG GCAUGCACUAUGC GCG AUACUGUU 4120
1184 UGCCAAGU G UUUGCUGA 1612 UCAGCAAA UGAUG GCAUGCACUAUGC GCG ACUUGGCA 4121
1251 GAACCUUU G UGUCUCCU 1613 AGGAGACA UGAUG GCAUGCACUAUGC GCG AAAGGUUC 4122
Table 39
1253 ACCUUUGU G UCUCCUCU 1614 AGAGGAGA UGAUG GCAUGCACUAUGC GCG ACAAAGGU 4123
1294 AGCCGCUU G UUUUGCUC 1615 GAGCAAAA UGAUG GCAUGCACUAUGC GCG AAGCGGCU 4124
1344 ACAAUUCU G UCGUGCUC 1616 GAGCACGA UGAUG GCAUGCACUAUGC GCG AGAAUUGU 4125
1390 GCUAGGCU G UGCUGCCA 1617 UGGCAGCA UGAUG GCAUGCACUAUGC GCG AGCCUAGC 4126
1425 CGUCCUUU G UUUACGUC 1618 GACGUAAA UGAUG GCAUGCACUAUGC GCG AAAGGACG 4127
1508 CGCCUAUU G UACCGACC 1619 GGUCGGUA UGAUG GCAUGCACUAUGC GCG AAUAGGCG 4128
1557 CCCCGUCU G UGCCUUCU 1620 AGAAGGCA UGAUG GCAUGCACUAUGC GCG AGACGGGG 4129
1581 CGGACCGU G UGCACUUC 1621 GAAGUGCA UGAUG GCAUGCACUAUGC GCG ACGGUCCG 4130
1684 UCAGCAAU G UCAACGAC 1622 GUCGUUGA UGAUG GCAUGCACUAUGC GCG AUUGCUGA 4131
1719 CAAAGACU G UGUGUUUA 1623 UAAACACA UGAUG GCAUGCACUAUGC GCG AGUCUUUG 4132
1721 AAGACUGU G UGUUUAAU 1624 AUUAAACA UGAUG GCAUGCACUAUGC GCG ACAGUCUU 4133
1723 GACUGUGU G UUUAAUGA 1625 UCAUUAAA UGAUG GCAUGCACUAUGC GCG ACACAGUC 4134
1772 AGGUCUUU G UACUAGGA 1626 UCCUAGUA UGAUG GCAUGCACUAUGC GCG AAAGACCU 4135
1785 AGGAGGCU G UAGGCAUA 1627 UAUGCCUA UGAUG GCAUGCACUAUGC GCG AGCCUCCU 4136
1801 AAAUUGGU G UGUUCACC 1628 GGUGAACA UGAUG GCAUGCACUAUGC GCG ACCAAUUU 4137
1803 AUUGGUGU G UUCACCAG 1629 CUGGUGAA UGAUG GCAUGCACUAUGC GCG ACACCAAU 4138
1850 CAUCUCAU G UUCAUGUC 1630 GACAUGAA UGAUG GCAUGCACUAUGC GCG AUGAGAUG 4139
1856 AUGUUCAU G UCCUACUG 1631 CAGUAGGA UGAUG GCAUGCACUAUGC GCG AUGAACAU 4140
1864 GUCCUACU G UUCAAGCC 1632 GGCUUGAA UGAUG GCAUGCACUAUGC GCG AGUAGGAC 4141
1881 UCCAAGCU G UGCCUUGG 1633 CCAAGGCA UGAUG GCAUGCACUAUGC GCG AGCUUGGA 4142
1939 GAGCUUCU G UGGAGUUA 1634 UAACUCCA UGAUG GCAUGCACUAUGC GCG AGAAGCUC 4143
2013 UCUGCUCU G UAUCGGGG 1635 CCCCGAUA UGAUG GCAUGCACUAUGC GCG AGAGCAGA 4144
2045 GGAACAUU G UUCACCUC 1636 GAGGUGAA UGAUG GCAUGCACUAUGC GCG AAUGUUCC 4145
2082 GCUAUUCU G UGUUGGGG 1637 CCCCAACA UGAUG GCAUGCACUAUGC GCG AGAAUAGC 4146
2084 UAUUCUGU G UUGGGGUG 1638 CACCCCAA UGAUG GCAUGCACUAUGC GCG ACAGAAUA 4147
2167 UCAGCUAU G UCAACGUU 1639 AACGUUGA UGAUG GCAUGCACUAUGC GCG AUAGCUGA 4148
2205 CAACUAUU G UGGUUUCA 1640 UGAAACCA UGAUG GCAUGCACUAUGC GCG AAUAGUUG 4149
2222 CAUUUCCU G UCUUACUU 1641 AAGUAAGA UGAUG GCAUGCACUAUGC GCG AGGAAAUG 4150
2245 GAGAAACU G UUCUUGAA 1642 UUCAAGAA UGAUG GCAUGCACUAUGC GCG AGUUUCUC 4151
2262 UAUUUGGU G UCUUUUGG 1643 CCAAAAGA UGAUG GCAUGCACUAUGC GCG ACCAAAUA 4152
2274 UUUGGAGU G UGGAUUCG 1644 CGAAUCCA UGAUG GCAUGCACUAUGC GCG ACUCCAAA 4153
2344 AAACUACU G UUGUUAGA 1645 UCUAACAA UGAUG GCAUGCACUAUGC GCG AGUAGUUU 4154
2347 CUACUGUU G UUAGACGA 1646 UCGUCUAA UGAUG GCAUGCACUAUGC GCG AACAGUAG 4155
2450 AUCUCAAU G UUAGUAUU 1647 AAUACUAA UGAUG GCAUGCACUAUGC GCG AUUGAGAU 4156
Table 39
2573 AGGACAUU G UUGAUAGA 1648 UCUAUCAA UGAUG GCAUGCACUAUGC GCG AAUGUCCU 4157
2583 UGAUAGAU G UAAGCAAU 1649 AUUGCUUA UGAUG GCAUGCACUAUGC GCG AUCUAUCA 4158
2594 AGCAAUUU G UGGGGCCC 1650 GGGCCCCA UGAUG GCAUGCACUAUGC GCG AAAUUGCU 4159
2663 AUCCCAAU G UUACUAAA 1651 UUUAGUAA UGAUG GCAUGCACUAUGC GCG AUUGGGAU 4160
2717 CAGAGUAU G UAGUUAAU 1652 AUUAACUA UGAUG GCAUGCACUAUGC GCG AUACUCUG 4161
2901 AUCUUUCU G UCCCCAAU 1653 AUUGGGGA UGAUG GCAUGCACUAUGC GCG AGAAAGAU 4162
3071 GGGGGACU G UUGGGGUG 1654 CACCCCAA UGAUG GCAUGCACUAUGC GCG AGUCCCCC 4163
3111 UCACAACU G UGCCAGCA 1655 UGCUGGCA UGAUG GCAUGCACUAUGC GCG AGUUGUGA 4164
Input Sequence = AF100308. Cut Site = YG/M or UG/U.
Stem Length = 8. Core Sequence = UGAUG GCAUGCACUAUGC GCG
AF100308 (Hepatitis B virus strain 2-18, 3215 bp)
Table 40
Table 40: Human HBV Zinzyme Ribozyme and Substrate Sequence
Pos Substrate Seq ID Ribozyme Rz Seq
ID
61 ACUUUCCU G CUGGUGGC 1448 GCCACCAG GCcgaaagGCGaGuCaaGGuCu AGGAAAGU 4165
94 UGAGCCCU G CUCAGAAU 1450 AUUCUGAG GCcgaaagGCGaGuCaaGGuCu AGGGCUCA 4166
112 CUGUCUCU G CCAUAUCG 1451 CGAUAUGG GCcgaaagGCGaGuCaaGGuCu AGAGACAG 4167
169 AGAACAUC G CAUCAGGA 1454 UCCUGAUG GCcgaaagGCGaGuCaaGGuCu GAUGUUCU 4168
192 GGACCCCU G CUCGUGUU 1455 AACACGAG GCcgaaagGCGaGuCaaGGuCu AGGGGUCC 4169
315 CAAAAUUC G CAGUCCCA 1457 UGGGACUG GCcgaaagGCGaGuCaaGGuCu GAAUUUUG 4170
374 UGGUUAUC G CUGGAUGU 1458 ACAUCCAG GCcgaaagGCGaGuCaaGGuCu GAUAACCA 4171
387 AUGUGUCU G CGGCGUUU 1459 AAACGCCG GCcgaaagGCGaGuCaaGGuCu AGACACAU 4172
410 CUUCCUCU G CAUCCUGC 1460 GCAGGAUG GCcgaaagGCGaGuCaaGGuCu AGAGGAAG 4173
417 UGCAUCCU G CUGCUAUG 1461 CAUAGCAG GCcgaaagGCGaGuCaaGGuCu AGGAUGCA 4174
420 AUCCUGCU G CUAUGCCU 1462 AGGCAUAG GCcgaaagGCGaGuCaaGGuCu AGCAGGAU 4175
425 GCUGCUAU G CCUCAUCU 1463 AGAUGAGG GCcgaaagGCGaGuCaaGGuCu AUAGCAGC 4176
468 GGUAUGUU G CCCGUUUG 1464 CAAACGGG GCcgaaagGCGaGuCaaGGuCu AACAUACC 4177
518 CGGACCAU G CAAAACCU 1465 AGGUUUUG GCcgaaagGCGaGuCaaGGuCu AUGGUCCG 4178
527 CAAAACCU G CACAACUC 1466 GAGUUGUG GCcgaaagGCGaGuCaaGGuCu AGGUUUUG 4179
538 CAACUCCU G CUCAAGGA 1467 UCCUUGAG GCcgaaagGCGaGuCaaGGuCu AGGAGUUG 4180
569 CUCAUGUU G CUGUACAA 1468 UUGUACAG GCcgaaagGCGaGuCaaGGuCu AACAUGAG 4181
596 CGGAAACU G CACCUGUA 1469 UACAGGUG GCcgaaagGCGaGuCaaGGuCu AGUUUCCG 4182
631 GGGCUUUC G CAAAAUAC 1470 GUAUUUUG GCcgaaagGCGaGuCaaGGuCu GAAAGCCC 4183
687 UUACUAGU G CCAUUUGU 1471 ACAAAUGG GCcgaaagGCGaGuCaaGGuCu ACUAGUAA 4184
795 CCCUUUAU G CCGCUGUU 1474 AACAGCGG GCcgaaagGCGaGuCaaGGuCu AUAAAGGG 4185
798 UUUAUGCC G CUGUUACC 1475 GGUAACAG GCcgaaagGCGaGuCaaGGuCu GGCAUAAA 4186
911 GGCACAUU G CCACAGGA 1476 UCCUGUGG GCcgaaagGCGaGuCaaGGuCu AAUGUGCC 4187
1020 UGGGGUUU G CCGCCCCU 1479 AGGGGCGG GCcgaaagGCGaGuCaaGGuCu AAACCCCA 4188
1023 GGUUUGCC G CCCCUUUC 1480 GAAAGGGG GCcgaaagGCGaGuCaaGGuCu GGCAAACC 4189
1034 CCUUUCAC G CAAUGUGG 1481 CCACAUUG GCcgaaagGCGaGuCaaGGuCu GUGAAAGG 4190
1050 GAUAUUCU G CUUUAAUG 1482 CAUUAAAG GCcgaaagGCGaGuCaaGGuCu AGAAUAUC 4191
1058 GCUUUAAU G CCUUUAUA 1483 UAUAAAGG GCcgaaagGCGaGuCaaGGuCu AUUAAAGC 4192
1068 CUUUAUAU G CAUGCAUA 1484 UAUGCAUG GCcgaaagGCGaGuCaaGGuCu AUAUAAAG 4193
1072 AUAUGCAU G CAUACAAG 1485 CUUGUAUG GCcgaaagGCGaGuCaaGGuCu AUGCAUAU 4194
Table 40
1103 ACUUUCUC G CCAACUUA 1486 UAAGUUGG GCcgaaagGCGaGuCaaGGuCu GAGAAAGU 4195
1155 ACCCCGUU G CUCGGCAA 1488 UUGCCGAG GCcgaaagGCGaGuCaaGGuCu AACGGGGU 4196
1177 UGGUCUAU G CCAAGUGU 1489 ACACUUGG GCcgaaagGCGaGuCaaGGuCu AUAGACCA 4197
1188 AAGUGUUU G CUGACGCA 1490 UGCGUCAG GCcgaaagGCGaGuCaaGGuCu AAACACUU 4198
1194 UUGCUGAC G CAACCCCC 1492 GGGGGUUG GCcgaaagGCGaGuCaaGGuCu GUCAGCAA 4199
1234 CCAUCAGC G CAUGCGUG 1493 CACGCAUG GCcgaaagGCGaGuCaaGGuCu GCUGAUGG 4200
1238 CAGCGCAU G CGUGGAAC 1494 GUUCCACG GCcgaaagGCGaGuCaaGGuCu AUGCGCUG 4201
1262 UCUCCUCU G CCGAUCCA 1495 UGGAUCGG GCcgaaagGCGaGuCaaGGuCu AGAGGAGA 4202
1275 UCCAUACC G CGGAACUC 1497 GAGUUCCG GCcgaaagGCGaGuCaaGGuCu GGUAUGGA 4203
1290 UCCUAGCC G CUUGUUUU 1498 AAAACAAG GCcgaaagGCGaGuCaaGGuCu GGCUAGGA 4204
1299 CUUGUUUU G CUCGCAGC 1499 GCUGCGAG GCcgaaagGCGaGuCaaGGuCu AAAACAAG 4205
1303 UUUUGCUC G CAGCAGGU 1500 ACCUGCUG GCcgaaagGCGaGuCaaGGuCu GAGCAAAA 4206
1349 UCUGUCGU G CUCUCCCG 1502 CGGGAGAG GCcgaaagGCGaGuCaaGGuCu ACGACAGA 4207
1357 GCUCUCCC G CAAAUAUA 1503 UAUAUUUG GCcgaaagGCGaGuCaaGGuCu GGGAGAGC 4208
1382 CCAUGGCU G CUAGGCUG 1504 CAGCCUAG GCcgaaagGCGaGuCaaGGuCu AGCCAUGG 4209
1392 UAGGCUGU G CUGCCAAC 1505 GUUGGCAG GCcgaaagGCGaGuCaaGGuCu ACAGCCUA 4210
1395 GCUGUGCU G CCAACUGG 1506 CCAGUUGG GCcgaaagGCGaGuCaaGGuCu AGCACAGC 4211
1411 GAUCCUAC G CGGGACGU 1507 ACGUCCCG GCcgaaagGCGaGuCaaGGuCu GUAGGAUC 4212
1442 CCGUCGGC G CUGAAUCC 1508 GGAUUCAG GCcgaaagGCGaGuCaaGGuCu GCCGACGG 4213
1452 UGAAUCCC G CGGACGAC 1510 GUCGUCCG GCcgaaagGCGaGuCaaGGuCu GGGAUUCA 4214
1474 CCGGGGCC G CUUGGGGC 1512 GCCCCAAG GCcgaaagGCGaGuCaaGGuCu GGCCCCGG 4215
1489 GCUCUACC G CCCGCUUC 1513 GAAGCGGG GCcgaaagGCGaGuCaaGGuCu GGUAGAGC 4216
1493 UACCGCCC G CUUCUCCG 1514 CGGAGAAG GCcgaaagGCGaGuCaaGGuCu GGGCGGUA 4217
1501 GCUUCUCC G CCUAUUGU 1515 ACAAUAGG GCcgaaagGCGaGuCaaGGuCu GGAGAAGC 4218
1528 CACGGGGC G CACCUCUC 1517 GAGAGGUG GCcgaaagGCGaGuCaaGGuCu GCCCCGUG 4219
1542 CUCUUUAC G CGGACUCC 1518 GGAGUCCG GCcgaaagGCGaGuCaaGGuCu GUAAAGAG 4220
1559 CCGUCUGU G CCUUCUCA 1519 UGAGAAGG GCcgaaagGCGaGuCaaGGuCu ACAGACGG 4221
1571 UCUCAUCU G CCGGACCG 1520 CGGUCCGG GCcgaaagGCGaGuCaaGGuCu AGAUGAGA 4222
1583 GACCGUGU G CACUUCGC 1521 GCGAAGUG GCcgaaagGCGaGuCaaGGuCu ACACGGUC 4223
1590 UGCACUUC G CUUCACCU 1522 AGGUGAAG GCcgaaagGCGaGuCaaGGuCu GAAGUGCA 4224
1601 UCACCUCU G CACGUCGC 1523 GCGACGUG GCcgaaagGCGaGuCaaGGuCu AGAGGUGA 4225
1608 UGCACGUC G CAUGGAGA 1524 UCUCCAUG GCcgaaagGCGaGuCaaGGuCu GACGUGCA 4226
1628 CCGUGAAC G CCCACAGG 1526 CCUGUGGG GCcgaaagGCGaGuCaaGGuCu GUUCACGG 4227
1642 AGGAACCU G CCCAAGGU 1527 ACCUUGGG GCcgaaagGCGaGuCaaGGuCu AGGUUCCU 4228
Table 40
1654 AAGGUCUU G CAUAAGAG 1528 CUCUUAUG GCcgaaagGCGaGuCaaGGuCu AAGACCUU 4229
1818 AGCACCAU G CAACUUUU 1533 AAAAGUUG GCcgaaagGCGaGuCaaGGuCu AUGGUGCU 4230
1835 UCACCUCU G CCUAAUCA 1534 UGAUUAGG GCcgaaagGCGaGuCaaGGuCu AGAGGUGA 4231
1883 CAAGCUGU G CCUUGGGU 1535 ACCCAAGG GCcgaaagGCGaGuCaaGGuCu ACAGCUUG 4232
1959 UCUUUUUU G CCUUCUGA 1537 UCAGAAGG GCcgaaagGCGaGuCaaGGuCu AAAAAAGA 4233
2002 UCGACACC G CCUCUGCU 1541 AGCAGAGG GCcgaaagGCGaGuCaaGGuCu GGUGUCGA 4234
2008 CCGCCUCU G CUCUGUAU 1542 AUACAGAG GCcgaaagGCGaGuCaaGGuCu AGAGGCGG 4235
2282 GUGGAUUC G CACUCCUC 1548 GAGGAGUG GCcgaaagGCGaGuCaaGGuCu GAAUCCAC 4236
2293 CUCCUCCU G CAUAUAGA 1549 UCUAUAUG GCcgaaagGCGaGuCaaGGuCu AGGAGGAG 4237
2311 CACCAAAU G CCCCUAUC 1550 GAUAGGGG GCcgaaagGCGaGuCaaGGuCu AUUUGGUG 4238
2388 ACUCCCUC G CCUCGCAG 1552 CUGCGAGG GCcgaaagGCGaGuCaaGGuCu GAGGGAGU 4239
2393 CUCGCCUC G CAGACGAA 1553 UUCGUCUG GCcgaaagGCGaGuCaaGGuCu GAGGCGAG 4240
2412 UCUCAAUC G CCGCGUCG 1555 CGACGCGG GCcgaaagGCGaGuCaaGGuCu GAUUGAGA 4241
2415 CAAUCGCC G CGUCGCAG 1556 CUGCGACG GCcgaaagGCGaGuCaaGGuCu GGCGAUUG 4242
2420 GCCGCGUC G CAGAAGAU 1557 AUCUUCUG GCcgaaagGCGaGuCaaGGuCu GACGCGGC 4243
2514 GGUACCUU G CUUUAAUC 1558 GAUUAAAG GCcgaaagGCGaGuCaaGGuCu AAGGUACC 4244
2560 AUUCAUUU G CAGGAGGA 1560 UCCUCCUG GCcgaaagGCGaGuCaaGGuCu AAAUGAAU 4245
2641 UUAACUAU G CCUGCUAG 1563 CUAGCAGG GCcgaaagGCGaGuCaaGGuCu AUAGUUAA 4246
2645 CUAUGCCU G CUAGGUUU 1564 AAACCUAG GCcgaaagGCGaGuCaaGGuCu AGGCAUAG 4247
2677 AAAUAUUU G CCCUUAGA 1565 UCUAAGGG GCcgaaagGCGaGuCaaGGuCu AAAUAUUU 4248
2740 UUCCAGAC G CGACAUUA 1566 UAAUGUCG GCcgaaagGCGaGuCaaGGuCu GUCUGGAA 4249
2804 CACGUAGC G CCUCAUUU 1568 AAAUGAGG GCcgaaagGCGaGuCaaGGuCu GCUACGUG 4250
2814 CUCAUUUU G CGGGUCAC 1569 GUGACCCG GCcgaaagGCGaGuCaaGGuCu AAAAUGAG 4251
2946 UGGACCCU G CAUUCAAA 1572 UUUGAAUG GCcgaaagGCGaGuCaaGGuCu AGGGUCCA 4252
2990 CUCAACCC G CACAAGGA 1573 UCCUUGUG GCcgaaagGCGaGuCaaGGuCu GGGUUGAG 4253
3012 GGCCGGAC G CCAACAAG 1574 CUUGUUGG GCcgaaagGCGaGuCaaGGuCu GUCCGGCC 4254
3090 GCCCUCAC G CUCAGGGC 1575 GCCCUGAG GCcgaaagGCGaGuCaaGGuCu GUGAGGGC 4255
3113 ACAACUGU G CCAGCAGC 1576 GCUGCUGG GCcgaaagGCGaGuCaaGGuCu ACAGUUGU 4256
3132 CUCCUCCU G CCUCCACC 1577 GGUGGAGG GCcgaaagGCGaGuCaaGGuCu AGGAGGAG 4257
51 AGGGCCCU G UACUUUCC 1578 GGAAAGUA GCcgaaagGCGaGuCaaGGuCu AGGGCCCU 4258
106 AGAAUACU G UCUCUGCC 1579 GGCAGAGA GCcgaaagGCGaGuCaaGGuCu AGUAUUCU 4259
148 GGGACCCU G UACCGAAC 1580 GUUCGGUA GCcgaaagGCGaGuCaaGGuCu AGGGUCCC 4260
198 CUGCUCGU G UUACAGGC 1581 GCCUGUAA GCcgaaagGCGaGuCaaGGuCu ACGAGCAG 4261
219 UUUUUCUU G UUGACAAA 1582 UUUGUCAA GCcgaaagGCGaGuCaaGGuCu AAGAAAAA 4262
Table 40
297 ACACCCGU G UGUCUUGG 1583 CCAAGACA GCcgaaagGCGaGuCaaGGuCu ACGGGUGU 4263
299 ACCCGUGU G UCUUGGCC 1584 GGCCAAGA GCcgaaagGCGaGuCaaGGuCu ACACGGGU 4264
347 ACCAACCU G UUGUCCUC 1585 GAGGACAA GCcgaaagGCGaGuCaaGGuCu AGGUUGGU 4265
350 AACCUGUU G UCCUCCAA 1586 UUGGAGGA GCcgaaagGCGaGuCaaGGuCu AACAGGUU 4266
362 UCCAAUUU G UCCUGGUU 1587 AACCAGGA GCcgaaagGCGaGuCaaGGuCu AAAUUGGA 4267
381 CGCUGGAU G UGUCUGCG 1588 CGCAGACA GCcgaaagGCGaGuCaaGGuCu AUCCAGCG 4268
383 CUGGAUGU G UCUGCGGC 1589 GCCGCAGA GCcgaaagGCGaGuCaaGGuCu ACAUCCAG 4269
438 AUCUUCUU G UUGGUUCU 1590 AGAACCAA GCcgaaagGCGaGuCaaGGuCu AAGAAGAU 4270
465 CAAGGUAU G UUGCCCGU 1591 ACGGGCAA GCcgaaagGCGaGuCaaGGuCu AUACCUUG 4271
476 GCCCGUUU G UCCUCUAA 1592 UUAGAGGA GCcgaaagGCGaGuCaaGGuCu AAACGGGC 4272
555 ACCUCUAU G UUUCCCUC 1593 GAGGGAAA GCcgaaagGCGaGuCaaGGuCu AUAGAGGU 4273
566 UCCCUCAU G UUGCUGUA 1594 UACAGCAA GCcgaaagGCGaGuCaaGGuCu AUGAGGGA 4274
572 AUGUUGCU G UACAAAAC 1595 GUUUUGUA GCcgaaagGCGaGuCaaGGuCu AGCAACAU 4275
602 CUGCACCU G UAUUCCCA 1596 UGGGAAUA GCcgaaagGCGaGuCaaGGuCu AGGUGCAG 4276
694 UGCCAUUU G UUCAGUGG 1597 CCACUGAA GCcgaaagGCGaGuCaaGGuCu AAAUGGCA 4277
724 CCCCCACU G UCUGGCUU 1598 AAGCCAGA GCcgaaagGCGaGuCaaGGuCu AGUGGGGG 4278
750 UGGAUGAU G UGGUUUUG 1599 CAAAACCA GCcgaaagGCGaGuCaaGGuCu AUCAUCCA 4279
771 CCAAGUCU G UACAACAU 1600 AUGUUGUA GCcgaaagGCGaGuCaaGGuCu AGACUUGG 4280
801 AUGCCGCU G UUACCAAU 1601 AUUGGUAA GCcgaaagGCGaGuCaaGGuCu AGCGGCAU 4281
818 UUUCUUUU G UCUUUGGG 1602 CCCAAAGA ■GCcgaaagGCGaGuCaaGGuCu AAAAGAAA 4282
888 UGGGAUAU G UAAUUGGG 1603 CCCAAUUA GCcgaaagGCGaGuCaaGGuCu AUAUCCCA 4283
927 AACAUAUU G UACAAAAA 1604 UUUUUGUA GCcgaaagGCGaGuCaaGGuCu AAUAUGUU 4284
944 AUCAAAAU G UGUUUUAG 1605 CUAAAACA GCcgaaagGCGaGuCaaGGuCu AUUUUGAU 4285
946 CAAAAUGU G UUUUAGGA 1606 UCCUAAAA GCcgaaagGCGaGuCaaGGuCu ACAUUUUG 4286
963 AACUUCCU G UAAACAGG 1607 CCUGUUUA GCcgaaagGCGaGuCaaGGuCu AGGAAGUU 4287
991 GAAAGUAU G UCAACGAA 1608 UUCGUUGA GCcgaaagGCGaGuCaaGGuCu AUACUUUC 4288
1002 AACGAAUU G UGGGUCUU 1609 AAGACCCA GCcgaaagGCGaGuCaaGGuCu AAUUCGUU 4289
1039 CACGCAAU G UGGAUAUU 1610 AAUAUCCA GCcgaaagGCGaGuCaaGGuCu AUUGCGUG 4290
1137 AACAGUAU G UGAACCUU 1611 AAGGUUCA GCcgaaagGCGaGuCaaGGuCu AUACUGUU 4291
1184 UGCCAAGU G UUUGCUGA 1612 UCAGCAAA GCcgaaagGCGaGuCaaGGuCu ACUUGGCA 4292
1251 GAACCUUU G UGUCUCCU 1613 AGGAGACA GCcgaaagGCGaGuCaaGGuCu AAAGGUUC 4293
1253 ACCUUUGU G UCUCCUCU 1614 AGAGGAGA GCcgaaagGCGaGuCaaGGuCu ACAAAGGU 4294
1294 AGCCGCUU G UUUUGCUC 1615 GAGCAAAA GCcgaaagGCGaGuCaaGGuCu AAGCGGCU 4295
1344 ACAAUUCU G UCGUGCUC 1616 GAGCACGA GCcgaaagGCGaGuCaaGGuCu AGAAUUGU 4296
Table 40
1390 GCUAGGCU G UGCUGCCA 1617 UGGCAGCA GCcgaaagGCGaGuCaaGGuCu AGCCUAGC 4297
1425 CGUCCUUU G UUUACGUC 1618 GACGUAAA GCcgaaagGCGaGuCaaGGuCu AAAGGACG 4298
1508 CGCCUAUU G UACCGACC 1619 GGUCGGUA GCcgaaagGCGaGuCaaGGuCu AAUAGGCG 4299
1557 CCCCGUCU G UGCCUUCU 1620 AGAAGGCA GCcgaaagGCGaGuCaaGGuCu AGACGGGG 4300
1581 CGGACCGU G UGCACUUC 1621 GAAGUGCA GCcgaaagGCGaGuCaaGGuCu ACGGUCCG 4301
1684 UCAGCAAU G UCAACGAC 1622 GUCGUUGA GCcgaaagGCGaGuCaaGGuCu AUUGCUGA 4302
1719 CAAAGACU G UGUGUUUA 1623 UAAACACA GCcgaaagGCGaGuCaaGGuCu AGUCUUUG 4303
1721 AAGACUGU G UGUUUAAU 1624 AUUAAACA GCcgaaagGCGaGuCaaGGuCu ACAGUCUU 4304
1723 GACUGUGU G UUUAAUGA 1625 UCAUUAAA GCcgaaagGCGaGuCaaGGuCu ACACAGUC 4305
1772 AGGUCUUU G UACUAGGA 1626 UCCUAGUA GCcgaaagGCGaGuCaaGGuCu AAAGACCU 4306
1785 AGGAGGCU G UAGGCAUA 1627 UAUGCCUA GCcgaaagGCGaGuCaaGGuCu AGCCUCCU 4307
1801 AAAUUGGU G UGUUCACC 1628 GGUGAACA GCcgaaagGCGaGuCaaGGuCu ACCAAUUU 4308
1803 AUUGGUGU G UUCACCAG 1629 CUGGUGAA GCcgaaagGCGaGuCaaGGuCu ACACCAAU 4309
1850 CAUCUCAU G UUCAUGUC 1630 GACAUGAA GCcgaaagGCGaGuCaaGGuCu AUGAGAUG 4310
1856 AUGUUCAU G UCCUACUG 1631 CAGUAGGA GCcgaaagGCGaGuCaaGGuCu AUGAACAU 4311
1864 GUCCUACU G UUCAAGCC 1632 GGCUUGAA GCcgaaagGCGaGuCaaGGuCu AGUAGGAC 4312
1881 UCCAAGCU G UGCCUUGG 1633 CCAAGGCA GCcgaaagGCGaGuCaaGGuCu AGCUUGGA 4313
1939 GAGCUUCU G UGGAGUUA 1634 UAACUCCA GCcgaaagGCGaGuCaaGGuCu AGAAGCUC 4314
2013 UCUGCUCU G UAUCGGGG 1635 CCCCGAUA GCcgaaagGCGaGuCaaGGuCu AGAGCAGA 4315
2045 GGAACAUU G UUCACCUC 1636 GAGGUGAA GCcgaaagGCGaGuCaaGGuCu AAUGUUCC 4316
2082 GCUAUUCU G UGUUGGGG 1637 CCCCAACA GCcgaaagGCGaGuCaaGGuCu AGAAUAGC 4317
2084 UAUUCUGU G UUGGGGUG 1638 CACCCCAA GCcgaaagGCGaGuCaaGGuCu ACAGAAUA 4318
2167 UCAGCUAU G UCAACGUU 1639 AACGUUGA GCcgaaagGCGaGuCaaGGuCu AUAGCUGA 4319
2205 CAACUAUU G UGGUUUCA 1640 UGAAACCA GCcgaaagGCGaGuCaaGGuCu AAUAGUUG 4320
2222 CAUUUCCU G UCUUACUU 1641 AAGUAAGA GCcgaaagGCGaGuCaaGGuCu AGGAAAUG 4321
2245 GAGAAACU G UUCUUGAA 1642 UUCAAGAA GCcgaaagGCGaGuCaaGGuCu AGUUUCUC 4322
2262 UAUUUGGU G UCUUUUGG 1643 CCAAAAGA GCcgaaagGCGaGuCaaGGuCu ACCAAAUA 4323
2274 UUUGGAGU G UGGAUUCG 1644 CGAAUCCA GCcgaaagGCGaGuCaaGGuCu ACUCCAAA 4324
2344 AAACUACU G UUGUUAGA 1645 UCUAACAA GCcgaaagGCGaGuCaaGGuCu AGUAGUUU 4325
2347 CUACUGUU G UUAGACGA 1646 UCGUCUAA GCcgaaagGCGaGuCaaGGuCu AACAGUAG 4326
2450 AUCUCAAU G UUAGUAUU 1647 AAUACUAA GCcgaaagGCGaGuCaaGGuCu AUUGAGAU 4327
2573 AGGACAUU G UUGAUAGA 1648 UCUAUCAA GCcgaaagGCGaGuCaaGGuCu AAUGUCCU 4328
2583 UGAUAGAU G UAAGCAAU 1649 AUUGCUUA GCcgaaagGCGaGuCaaGGuCu AUCUAUCA 4329
2594 AGCAAUUU G UGGGGCCC 1650 GGGCCCCA GCcgaaagGCGaGuCaaGGuCu AAAUUGCU 4330
Table 40
2663 AUCCCAAU G UUACUAAA 1651 UUUAGUAA GCcgaaagGCGaGuCaaGGuCu AUUGGGAU 4331
2717 CAGAGUAU G UAGUUAAU 1652 AUUAACUA GCcgaaagGCGaGuCaaGGuCu AUACUCUG 4332
2901 AUCUUUCU G UCCCCAAU 1653 AUUGGGGA GCcgaaagGCGaGuCaaGGuCu AGAAAGAU 4333
3071 GGGGGACU G UUGGGGUG 1654 CACCCCAA GCcgaaagGCGaGuCaaGGuCu AGUCCCCC 4334
3111 UCACAACU G UGCCAGCA 1655 UGCUGGCA GCcgaaagGCGaGuCaaGGuCu AGUUGUGA 4335
40 AUCCCAGA G UCAGGGCC 1656 GGCCCUGA GCcgaaagGCGaGuCaaGGuCu UCUGGGAU 4336
46 GAGUCAGG G CCCUGUAC 1657 GUACAGGG GCcgaaagGCGaGuCaaGGuCu CCUGACUC 4337
65 UCCUGCUG G UGGCUCCA 1658 UGGAGCCA GCcgaaagGCGaGuCaaGGuCu CAGCAGGA 4338
68 UGCUGGUG G CUCCAGUU 1659 AACUGGAG GCcgaaagGCGaGuCaaGGuCu CACCAGCA 4339
74 UGGCUCCA G UUCAGGAA 1660 UUCCUGAA GCcgaaagGCGaGuCaaGGuCu UGGAGCCA 4340
85 CAGGAACA G UGAGCCCU 1661 AGGGCUCA GCcgaaagGCGaGuCaaGGuCu UGUUCCUG 4341
89 AACAGUGA G CCCUGCUC 1662 GAGCAGGG GCcgaaagGCGaGuCaaGGuCu UCACUGUU 4342
120 GCCAUAUC G UCAAUCUU 1663 AAGAUUGA GCcgaaagGCGaGuCaaGGuCu GAUAUGGC 4343
196 CCCUGCUC G UGUUACAG 1664 CUGUAACA GCcgaaagGCGaGuCaaGGuCu GAGCAGGG 4344
205 UGUUACAG G CGGGGUUU 1665 AAACCCCG GCcgaaagGCGaGuCaaGGuCu CUGUAACA 4345
210 CAGGCGGG G UUUUUCUU 1666 AAGAAAAA GCcgaaagGCGaGuCaaGGuCu CCCGCCUG 4346
248 ACCACAGA G UCUAGACU 1667 AGUCUAGA GCcgaaagGCGaGuCaaGGuCu UCUGUGGU 4347
258 CUAGACUC G UGGUGGAC 1668 GUCCACCA GCcgaaagGCGaGuCaaGGuCu GAGUCUAG 4348
261 GACUCGUG G UGGACUUC 1669 GAAGUCCA GCcgaaagGCGaGuCaaGGuCu CACGAGUC 4349
295 GAACACCC G UGUGUCUU 1670 AAGACACA GCcgaaagGCGaGuCaaGGuCu GGGUGUUC 4350
305 GUGUCUUG G CCAAAAUU 1671 AAUUUUGG GCcgaaagGCGaGuCaaGGuCu CAAGACAC 4351
318 AAUUCGCA G UCCCAAAU 1672 AUUUGGGA GCcgaaagGCGaGuCaaGGuCu UGCGAAUU 4352
332 AAUCUCCA G UCACUCAC 1673 GUGAGUGA GCcgaaagGCGaGuCaaGGuCu UGGAGAUU 4353
368 UUGUCCUG G UUAUCGCU 1674 AGCGAUAA GCcgaaagGCGaGuCaaGGuCu CAGGACAA 4354
390 UGUCUGCG G CGUUUUAU 1675 AUAAAACG GCcgaaagGCGaGuCaaGGuCu CGCAGACA 4355
392 UCUGCGGC G UUUUAUCA 1676 UGAUAAAA GCcgaaagGCGaGuCaaGGuCu GCCGCAGA 4356
442 UCUUGUUG G UUCUUCUG 1677 CAGAAGAA GCcgaaagGCGaGuCaaGGuCu CAACAAGA 4357
461 CUAUCAAG G UAUGUUGC 1678 GCAACAUA GCcgaaagGCGaGuCaaGGuCu CUUGAUAG 4358
472 UGUUGCCC G UUUGUCCU 1679 AGGACAAA GCcgaaagGCGaGuCaaGGuCu GGGCAACA 4359
506 AACAACCA G CACCGGAC 1680 GUCCGGUG GCcgaaagGCGaGuCaaGGuCu UGGUUGUU 4360
625 CAUCUUGG G CUUUCGCA 1681 UGCGAAAG GCcgaaagGCGaGuCaaGGuCu CCAAGAUG 4361
648 CUAUGGGA G UGGGCCUC 1682 GAGGCCCA GCcgaaagGCGaGuCaaGGuCu UCCCAUAG 4362
652 GGGAGUGG G CCUCAGUC 1683 GACUGAGG GCcgaaagGCGaGuCaaGGuCu CCACUCCC 4363
658 GGGCCUCA G UCCGUUUC 1684 GAAACGGA GCcgaaagGCGaGuCaaGGuCu UGAGGCCC 4364
Table 40
662 CUCAGUCC G UUUCUCUU 1685 AAGAGAAA GCcgaaagGCGaGuCaaGGuCu GGACUGAG 4365
672 UUCUCUUG G CUCAGUUU 1686 AAACUGAG GCcgaaagGCGaGuCaaGGuCu CAAGAGAA 4366
677 UUGGCUCA G UUUACUAG 1687 CUAGUAAA GCcgaaagGCGaGuCaaGGuCu UGAGCCAA 4367
685 GUUUACUA G UGCCAUUU 1688 AAAUGGCA GCcgaaagGCGaGuCaaGGuCu UAGUAAAC 4368
699 UUUGUUCA G UGGUUCGU 1689 ACGAACCA GCcgaaagGCGaGuCaaGGuCu UGAACAAA 4369
702 GUUCAGUG G UUCGUAGG 1690 CCUACGAA GCcgaaagGCGaGuCaaGGuCu CACUGAAC 4370
706 AGUGGUUC G UAGGGCUU 1691 AAGCCCUA GCcgaaagGCGaGuCaaGGuCu GAACCACU 4371
711 UUCGUAGG G CUUUCCCC 1692 GGGGAAAG GCcgaaagGCGaGuCaaGGuCu CCUACGAA 4372
729 ACUGUCUG G CUUUCAGU 1693 ACUGAAAG GCcgaaagGCGaGuCaaGGuCu CAGACAGU 4373
736 GGCUUUCA G UUAUAUGG 1694 CCAUAUAA GCcgaaagGCGaGuCaaGGuCu UGAAAGCC 4374
753 AUGAUGUG G UUUUGGGG 1695 CCCCAAAA GCcgaaagGCGaGuCaaGGuCu CACAUCAU 4375
762 UUUUGGGG G CCAAGUCU 1696 AGACUUGG GCcgaaagGCGaGuCaaGGuCu CCCCAAAA 4376
767 GGGGCCAA G UCUGUACA 1697 UGUACAGA GCcgaaagGCGaGuCaaGGuCu UUGGCCCC 4377
785 CAUCUUGA G UCCCUUUA 1698 UAAAGGGA GCcgaaagGCGaGuCaaGGuCu UCAAGAUG 4378
826 GUCUUUGG G UAUACAUU 1699 AAUGUAUA GCcgaaagGCGaGuCaaGGuCu CCAAAGAC 4379
898 AAUUGGGA G UUGGGGCA 1700 UGCCCCAA GCcgaaagGCGaGuCaaGGuCu UCCCAAUU 4380
904 GAGUUGGG G CACAUUGC 1701 GCAAUGUG GCcgaaagGCGaGuCaaGGuCu CCCAACUC 4381
971 GUAAACAG G CCUAUUGA 1702 UCAAUAGG GCcgaaagGCGaGuCaaGGuCu CUGUUUAC 4382
987 AUUGGAAA G UAUGUCAA 1703 UUGACAUA GCcgaaagGCGaGuCaaGGuCu UUUCCAAU 4383
1006 AAUUGUGG G UCUUUUGG 1704 CCAAAAGA GCcgaaagGCGaGuCaaGGuCu CCACAAUU 4384
1016 CUUUUGGG G UUUGCCGC 1705 GCGGCAAA GCcgaaagGCGaGuCaaGGuCu CCCAAAAG 4385
1080 GCAUACAA G CAAAACAG 1706 CUGUUUUG GCcgaaagGCGaGuCaaGGuCu UUGUAUGC 4386
1089 CAAAACAG G CUUUUACU 1707 AGUAAAAG GCcgaaagGCGaGuCaaGGuCu CUGUUUUG 4387
1116 CUUACAAG G CCUUUCUA 1708 UAGAAAGG GCcgaaagGCGaGuCaaGGuCu CUUGUAAG 4388
1126 CUUUCUAA G UAAACAGU 1709 ACUGUUUA GCcgaaagGCGaGuCaaGGuCu UUAGAAAG 4389
1133 AGUAAACA G UAUGUGAA 1710 UUCACAUA GCcgaaagGCGaGuCaaGGuCu UGUUUACU 4390
1152 UUUACCCC G UUGCUCGG 1711 CCGAGCAA GCcgaaagGCGaGuCaaGGuCu GGGGUAAA 4391
1160 GUUGCUCG G CAACGGCC 1712 GGCCGUUG GCcgaaagGCGaGuCaaGGuCu CGAGCAAC 4392
1166 CGGCAACG G CCUGGUCU 1713 AGACCAGG GCcgaaagGCGaGuCaaGGuCu CGUUGCCG 4393
1171 ACGGCCUG G UCUAUGCC 1714 GGCAUAGA GCcgaaagGCGaGuCaaGGuCu CAGGCCGU 4394
1182 UAUGCCAA G UGUUUGCU 1715 AGCAAACA GCcgaaagGCGaGuCaaGGuCu UUGGCAUA 4395
1207 CCCCACUG G UUGGGGCU 1716 AGCCCCAA GCcgaaagGCGaGuCaaGGuCu CAGUGGGG 4396
1213 UGGUUGGG G CUUGGCCA 1717 UGGCCAAG GCcgaaagGCGaGuCaaGGuCu CCCAACCA 4397
1218 GGGGCUUG G CCAUAGGC 1718 GCCUAUGG GCcgaaagGCGaGuCaaGGuCu CAAGCCCC 4398
Table 40
1225 GGCCAUAG G CCAUCAGC 1719 GCUGAUGG GCcgaaagGCGaGuCaaGGuCu CUAUGGCC 4399
1232 GGCCAUCA G CGCAUGCG 1720 CGCAUGCG GCcgaaagGCGaGuCaaGGuCu UGAUGGCC 4400
1240 GCGCAUGC G UGGAACCU 1721 AGGUUCCA GCcgaaagGCGaGuCaaGGuCu GCAUGCGC 4401
1287 AACUCCUA G CCGCUUGU 1722 ACAAGCGG GCcgaaagGCGaGuCaaGGuCu UAGGAGUU 4402
1306 UGCUCGCA G CAGGUCUG 1723 CAGACCUG GCcgaaagGCGaGuCaaGGuCu UGCGAGCA 4403
1310 CGCAGCAG G UCUGGGGC 1724 GCCCCAGA GCcgaaagGCGaGuCaaGGuCu CUGCUGCG 4404
1317 GGUCUGGG G CAAAACUC 1725 GAGUUUUG GCcgaaagGCGaGuCaaGGuCu CCCAGACC 4405
1347 AUUCUGUC G UGCUCUCC 1726 GGAGAGCA GCcgaaagGCGaGuCaaGGuCu GACAGAAU 4406
1379 UUUCCAUG G CUGCUAGG 1727 CCUAGCAG GCcgaaagGCGaGuCaaGGuCu CAUGGAAA 4407
1387 GCUGCUAG G CUGUGCUG 1728 CAGCACAG GCcgaaagGCGaGuCaaGGuCu CUAGCAGC 4408
1418 CGCGGGAC G UCCUUUGU 1729 ACAAAGGA GCcgaaagGCGaGuCaaGGuCu GUCCCGCG 4409
1431 UUGUUUAC G UCCCGUCG 1730 CGACGGGA GCcgaaagGCGaGuCaaGGuCu GUAAACAA 4410
1436 UACGUCCC G UCGGCGCU 1731 AGCGCCGA GCcgaaagGCGaGuCaaGGuCu GGGACGUA 4411
1440 UCCCGUCG G CGCUGAAU 1732 AUUCAGCG GCcgaaagGCGaGuCaaGGuCu CGACGGGA 4412
1471 CUCCCGGG G CCGCUUGG 1733 CCAAGCGG GCcgaaagGCGaGuCaaGGuCu CCCGGGAG 4413
1481 CGCUUGGG G CUCUACCG 1734 CGGUAGAG GCcgaaagGCGaGuCaaGGuCu CCCAAGCG 4414
1517 UACCGACC G UCCACGGG 1735 CCCGUGGA GCcgaaagGCGaGuCaaGGuCu GGUCGGUA 4415
1526 UCCACGGG G CGCACCUC 1736 GAGGUGCG GCcgaaagGCGaGuCaaGGuCu CCCGUGGA 4416
1553 GACUCCCC G UCUGUGCC 1737 GGCACAGA GCcgaaagGCGaGuCaaGGuCu GGGGAGUC 4417
1579 GCCGGACC G UGUGCACU 1738 AGUGCACA GCcgaaagGCGaGuCaaGGuCu GGUCCGGC 4418
1605 CUCUGCAC G UCGCAUGG 1739 CCAUGCGA GCcgaaagGCGaGuCaaGGuCu GUGCAGAG 4419
1622 AGACCACC G UGAACGCC 1740 GGCGUUCA GCcgaaagGCGaGuCaaGGuCu GGUGGUCU 4420
1649 UGCCCAAG G UCUUGCAU 1741 AUGCAAGA GCcgaaagGCGaGuCaaGGuCu CUUGGGCA 4421
1679 GACUUUCA G CAAUGUCA 1742 UGACAUUG GCcgaaagGCGaGuCaaGGuCu UGAAAGUC 4422
1703 ACCUUGAG G CAUACUUC 1743 GAAGUAUG GCcgaaagGCGaGuCaaGGuCu CUCAAGGU 4423
1732 UUUAAUGA G UGGGAGGA 1744 UCCUCCCA GCcgaaagGCGaGuCaaGGuCu UCAUUAAA 4424
1741 UGGGAGGA G UUGGGGGA 1745 UCCCCCAA GCcgaaagGCGaGuCaaGGuCu UCCUCCCA 4425
1754 GGGAGGAG G UUAGGUUA 1746 UAACCUAA GCcgaaagGCGaGuCaaGGuCu CUCCUCCC 4426
1759 GAGGUUAG G UUAAAGGU 1747 ACCUUUAA GCcgaaagGCGaGuCaaGGuCu CUAACCUC 4427
1766 GGUUAAAG G UCUUUGUA 1748 UACAAAGA GCcgaaagGCGaGuCaaGGuCu CUUUAACC 4428
1782 ACUAGGAG G CUGUAGGC 1749 GCCUACAG GCcgaaagGCGaGuCaaGGuCu CUCCUAGU 4429
1789 GGCUGUAG G CAUAAAUU 1750 AAUUUAUG GCcgaaagGCGaGuCaaGGuCu CUACAGCC 4430
1799 AUAAAUUG G UGUGUUCA 1751 UGAACACA GCcgaaagGCGaGuCaaGGuCu CAAUUUAU 4431
1811 GUUCACCA G CACCAUGC 1752 GCAUGGUG GCcgaaagGCGaGuCaaGGuCu UGGUGAAC 4432
Table 40
1870 CUGUUCAA G CCUCCAAG 1753 CUUGGAGG GCcgaaagGCGaGuCaaGGuCu UUGAACAG 4433
1878 GCCUCCAA G CUGUGCCU 1754 AGGCACAG GCcgaaagGCGaGuCaaGGuCu UUGGAGGC 4434
1890 UGCCUUGG G UGGCUUUG 1755 CAAAGCCA GCcgaaagGCGaGuCaaGGuCu CCAAGGCA 4435
1893 CUUGGGUG G CUUUGGGG 1756 CCCCAAAG GCcgaaagGCGaGuCaaGGuCu CACCCAAG 4436
1901 GCUUUGGG G CAUGGACA 1757 UGUCCAUG GCcgaaagGCGaGuCaaGGuCu CCCAAAGC 4437
1917 AUUGACCC G UAUAAAGA 1758 UCUUUAUA GCcgaaagGCGaGuCaaGGuCu GGGUCAAU 4438
1933 AAUUUGGA G CUUCUGUG 1759 CACAGAAG GCcgaaagGCGaGuCaaGGuCu UCCAAAUU 4439
1944 UCUGUGGA G UUACUCUC 1760 GAGAGUAA GCcgaaagGCGaGuCaaGGuCu UCCACAGA 4440
2023 AUCGGGGG G CCUUAGAG 1761 CUCUAAGG GCcgaaagGCGaGuCaaGGuCu CCCCCGAU 4441
2031 GCCUUAGA G UCUCCGGA 1762 UCCGGAGA GCcgaaagGCGaGuCaaGGuCu UCUAAGGC 4442
2062 ACCAUACG G CACUCAGG 1763 CCUGAGUG GCcgaaagGCGaGuCaaGGuCu CGUAUGGU 4443
2070 GCACUCAG G CAAGCUAU 1764 AUAGCUUG GCcgaaagGCGaGuCaaGGuCu CUGAGUGC 4444
2074 UCAGGCAA G CUAUUCUG 1765 CAGAAUAG GCcgaaagGCGaGuCaaGGuCu UUGCCUGA 4445
2090 GUGUUGGG G UGAGUUGA 1766 UCAACUCA GCcgaaagGCGaGuCaaGGuCu CCCAACAC 4446
2094 UGGGGUGA G UUGAUGAA 1767 UUCAUCAA GCcgaaagGCGaGuCaaGGuCu UCACCCCA 4447
2107 UGAAUCUA G CCACCUGG 1768 CCAGGUGG GCcgaaagGCGaGuCaaGGuCu UAGAUUCA 4448
2116 CCACCUGG G UGGGAAGU 1769 ACUUCCCA GCcgaaagGCGaGuCaaGGuCu CCAGGUGG 4449
2123 GGUGGGAA G UAAUUUGG 1770 CCAAAUUA GCcgaaagGCGaGuCaaGGuCu UUCCCACC 4450
2140 AAGAUCCA G CAUCCAGG 1771 CCUGGAUG GCcgaaagGCGaGuCaaGGuCu UGGAUCUU 4451
2155 GGGAAUUA G UAGUCAGC 1772 GCUGACUA GCcgaaagGCGaGuCaaGGuCu UAAUUCCC 4452
2158 AAUUAGUA G UCAGCUAU 1773 AUAGCUGA GCcgaaagGCGaGuCaaGGuCu UACUAAUU 4453
2162 AGUAGUCA G CUAUGUCA 1774 UGACAUAG GCcgaaagGCGaGuCaaGGuCu UGACUACU 4454
2173 AUGUCAAC G UUAAUAUG 1775 CAUAUUAA GCcgaaagGCGaGuCaaGGuCu GUUGACAU 4455
2183 UAAUAUGG G CCUAAAAA 1776 UUUUUAGG GCcgaaagGCGaGuCaaGGuCu CCAUAUUA 4456
2208 CUAUUGUG G UUUCACAU 1777 AUGUGAAA GCcgaaagGCGaGuCaaGGuCu CACAAUAG 4457
2235 ACUUUUGG G CGAGAAAC 1778 GUUUCUCG GCcgaaagGCGaGuCaaGGuCu CCAAAAGU 4458
2260 AAUAUUUG G UGUCUUUU 1779 AAAAGACA GCcgaaagGCGaGuCaaGGuCu CAAAUAUU 4459
2272 CUUUUGGA G UGUGGAUU 1780 AAUCCACA GCcgaaagGCGaGuCaaGGuCu UCCAAAAG 4460
2360 ACGAAGAG G CAGGUCCC 1781 GGGACCUG GCcgaaagGCGaGuCaaGGuCu CUCUUCGU 4461
2364 AGAGGCAG G UCCCCUAG 1782 CUAGGGGA GCcgaaagGCGaGuCaaGGuCu CUGCCUCU 4462
2403 AGACGAAG G UCUCAAUC 1783 GAUUGAGA GCcgaaagGCGaGuCaaGGuCu CUUCGUCU 4463
2417 AUCGCCGC G UCGCAGAA 1784 UUCUGCGA GCcgaaagGCGaGuCaaGGuCu GCGGCGAU 4464
2454 CAAUGUUA G UAUUCCUU 1785 AAGGAAUA GCcgaaagGCGaGuCaaGGuCu UAACAUUG 4465
2474 CACAUAAG G UGGGAAAC 1786 GUUUCCCA GCcgaaagGCGaGuCaaGGuCu CUUAUGUG 4466
Table 40
2491 UUUACGGG G CUUUAUUC 1787 GAAUAAAG GCcgaaagGCGaGuCaaGGuCu CCCGUAAA 4467
2507 CUUCUACG G UACCUUGC 1788 GCAAGGUA GCcgaaagGCGaGuCaaGGuCu CGUAGAAG 4468
2530 CCUAAAUG G CAAACUCC 1789 GGAGUUUG GCcgaaagGCGaGuCaaGGuCu CAUUUAGG 4469
2587 AGAUGUAA G CAAUUUGU 1790 ACAAAUUG GCcgaaagGCGaGuCaaGGuCu UUACAUCU 4470
2599 UUUGUGGG G CCCCUUAC 1791 GUAAGGGG GCcgaaagGCGaGuCaaGGuCu CCCACAAA 4471
2609 CCCUUACA G UAAAUGAA 1792 UUCAUUUA GCcgaaagGCGaGuCaaGGuCu UGUAAGGG 4472
2650 CCUGCUAG G UUUUAUCC 1793 GGAUAAAA GCcgaaagGCGaGuCaaGGuCu CUAGCAGG 4473
2701 AUCAAACC G UAUUAUCC 1794 GGAUAAUA GCcgaaagGCGaGuCaaGGuCu GGUUUGAU 4474
2713 UAUCCAGA G UAUGUAGU 1795 ACUACAUA GCcgaaagGCGaGuCaaGGuCu UCUGGAUA 4475
2720 AGUAUGUA G UUAAUCAU 1796 AUGAUUAA GCcgaaagGCGaGuCaaGGuCu UACAUACU 4476
2768 UUUGGAAG G CGGGGAUC 1797 GAUCCCCG GCcgaaagGCGaGuCaaGGuCu CUUCCAAA 4477
2791 AAAAGAGA G UCCACACG 1798 CGUGUGGA GCcgaaagGCGaGuCaaGGuCu UCUCUUUU 4478
2799 GUCCACAC G UAGCGCCU 1799 AGGCGCUA GCcgaaagGCGaGuCaaGGuCu GUGUGGAC 4479
2802 CACACGUA G CGCCUCAU 1800 AUGAGGCG GCcgaaagGCGaGuCaaGGuCu UACGUGUG 4480
2818 UUUUGCGG G UCACCAUA 1801 UAUGGUGA GCcgaaagGCGaGuCaaGGuCu CCGCAAAA 4481
2848 GAUCUACA G CAUGGGAG 1802 CUCCCAUG GCcgaaagGCGaGuCaaGGuCu UGUAGAUC 4482
2857 CAUGGGAG G UUGGUCUU 1803 AAGACCAA GCcgaaagGCGaGuCaaGGuCu CUCCCAUG 4483
2861 GGAGGUUG G UCUUCCAA 1804 UUGGAAGA GCcgaaagGCGaGuCaaGGuCu CAACCUCC 4484
2881 UCGAAAAG G CAUGGGGA 1805 UCCCCAUG GCcgaaagGCGaGuCaaGGuCu CUUUUCGA 4485
2936 GAUCAUCA G UUGGACCC 1806 GGGUCCAA GCcgaaagGCGaGuCaaGGuCu UGAUGAUC 4486
2955 CAUUCAAA G CCAACUCA 1807 UGAGUUGG GCcgaaagGCGaGuCaaGGuCu UUUGAAUG 4487
2964 CCAACUCA G UAAAUCCA 1808 UGGAUUUA GCcgaaagGCGaGuCaaGGuCu UGAGUUGG 4488
3005 GACAACUG G CCGGACGC 1809 GCGUCCGG GCcgaaagGCGaGuCaaGGuCu CAGUUGUC 4489
3021 CCAACAAG G UGGGAGUG 1810 CACUCCCA GCcgaaagGCGaGuCaaGGuCu CUUGUUGG 4490
3027 AGGUGGGA G UGGGAGCA 1811 UGCUCCCA GCcgaaagGCGaGuCaaGGuCu UCCCACCU 4491
3033 GAGUGGGA G CAUUCGGG 1812 CCCGAAUG GCcgaaagGCGaGuCaaGGuCu UCCCACUC 4492
3041 GCAUUCGG G CCAGGGUU 1813 AACCCUGG GCcgaaagGCGaGuCaaGGuCu CCGAAUGC 4493
3047 GGGCCAGG G UUCACCCC 1814 GGGGUGAA GCcgaaagGCGaGuCaaGGuCu CCUGGCCC 4494
3077 CUGUUGGG G UGGAGCCC 1815 GGGCUCCA GCcgaaagGCGaGuCaaGGuCu CCCAACAG 4495
3082 GGGGUGGA G CCCUCACG 1816 CGUGAGGG GCcgaaagGCGaGuCaaGGuCu UCCACCCC 4496
3097 CGCUCAGG G CCUACUCA 1817 UGAGUAGG GCcgaaagGCGaGuCaaGGuCu CCUGAGCG 4497
3117 CUGUGCCA G CAGCUCCU 1818 AGGAGCUG GCcgaaagGCGaGuCaaGGuCu UGGCACAG 4498
3120 UGCCAGCA G CUCCUCCU 1819 AGGAGGAG GCcgaaagGCGaGuCaaGGuCu UGCUGGCA 4499
3146 ACCAAUCG G CAGUCAGG 1820 CCUGACUG GCcgaaagGCGaGuCaaGGuCu CGAUUGGU 4500
Table 40
Input Sequence = AF100308. Cut Site = YG/M or UG/U.
Stem Length = 8 . Core Sequence = GCcgaaagGCGaGuCaaGGuCu
AF100308 (Hepatitis B virus strain 2-18, 3215 bp)
Table 41 Table 41: Human HBV DNAzyme and Substrate Sequence
Pos Substrate Seq ID DNAzyme Rz Seq ID
508 CAACCAGC A CCGGACCA 833 TGGTCCGG GGCTAGCTACAACGA GCTGGTTG 4505
1632 GAACGCCC A CAGGAACC 1096 GGTTCCTG GGCTAGCTACAACGA GGGCGTTC 4506
2992 CAACCCGC A CAAGGACA 1376 TGTCCTTG GGCTAGCTACAACGA GCGGGTTG 4507
61 ACUUUCCU G CUGGUGGC 1448 GCCACCAG GGCTAGCTACAACGA AGGAAAGT 4508
94 UGAGCCCU G CUCAGAAU 1450 ATTCTGAG GGCTAGCTACAACGA AGGGCTCA 4509
112 CUGUCUCU G CCAUAUCG 1451 CGATATGG GGCTAGCTACAACGA AGAGACAG 4510
169 AGAACAUC G CAUCAGGA 1454 TCCTGATG GGCTAGCTACAACGA GATGTTCT 4511
192 GGACCCCU G CUCGUGUU 1455 AACACGAG GGCTAGCTACAACGA AGGGGTCC 4512
315 CAAAAUUC G CAGUCCCA 1457 TGGGACTG GGCTAGCTACAACGA GAATTTTG 4513
374 UGGUUAUC G CUGGAUGU 1458 ACATCCAG GGCTAGCTACAACGA GATAACCA 4514
387 AUGUGUCU G CGGCGUUU 1459 AAACGCCG GGCTAGCTACAACGA AGACACAT 4515
410 CUUCCUCU G CAUCCUGC 1460 GCAGGATG GGCTAGCTACAACGA AGAGGAAG 4516
417 UGCAUCCU G CUGCUAUG 1461 CATAGCAG GGCTAGCTACAACGA AGGATGCA 4517
420 AUCCUGCU G CUAUGCCU 1462 AGGCATAG GGCTAGCTACAACGA AGCAGGAT 4518
425 GCUGCUAU G CCUCAUCU 1463 AGATGAGG GGCTAGCTACAACGA ATAGCAGC 4519
468 GGUAUGUU G CCCGUUUG 1464 CAAACGGG GGCTAGCTACAACGA AACATACC 4520
518 CGGACCAU G CAAAACCU 1465 AGGTTTTG GGCTAGCTACAACGA ATGGTCCG 4521
527 CAAAACCU G CACAACUC 1466 GAGTTGTG GGCTAGCTACAACGA AGGTTTTG 4522
538 CAACUCCU G CUCAAGGA 1467 TCCTTGAG GGCTAGCTACAACGA AGGAGTTG 4523
569 CUCAUGUU G CUGUACAA 1468 TTGTACAG GGCTAGCTACAACGA AACATGAG 4524
596 CGGAAACU G CACCUGUA 1469 TACAGGTG GGCTAGCTACAACGA AGTTTCCG 4525
631 GGGCUUUC G CAAAAUAC 1470 GTATTTTG GGCTAGCTACAACGA GAAAGCCC 4526
687 UUACUAGU G CCAUUUGU 1471 ACAAATGG GGCTAGCTACAACGA ACTAGTAA 4527
795 CCCUUUAU G CCGCUGUU 1474 AACAGCGG GGCTAGCTACAACGA ATAAAGGG 4528
798 UUUAUGCC G CUGUUACC 1475 GGTAACAG GGCTAGCTACAACGA GGCATAAA 4529
911 GGCACAUU G CCACAGGA 1476 TCCTGTGG GGCTAGCTACAACGA AATGTGCC 4530
1020 UGGGGUUU G CCGCCCCU 1479 AGGGGCGG GGCTAGCTACAACGA AAACCCCA 4531
1023 GGUUUGCC G CCCCUUUC 1480 GAAAGGGG GGCTAGCTACAACGA GGCAAACC 4532
1034 CCUUUCAC G CAAUGUGG 1481 CCACATTG GGCTAGCTACAACGA GTGAAAGG 4533
1050 GAUAUUCU G CUUUAAUG 1482 CATTAAAG GGCTAGCTACAACGA AGAATATC 4534
1058 GCUUUAAU G CCUUUAUA 1483 TATAAAGG GGCTAGCTACAACGA ATTAAAGC 4535
1068 CUUUAUAU G CAUGCAUA 1484 TATGCATG GGCTAGCTACAACGA ATATAAAG 4536
1072 AUAUGCAU G CAUACAAG 1485 CTTGTATG GGCTAGCTACAACGA ATGCATAT 4537
1103 ACUUUCUC G CCAACUUA 1486 TAAGTTGG GGCTAGCTACAACGA GAGAAAGT 4538
1155 ACCCCGUU G CUCGGCAA 1488 TTGCCGAG GGCTAGCTACAACGA AACGGGGT 4539
1177 UGGUCUAU G CCAAGUGU 1489 ACACTTGG GGCTAGCTACAACGA ATAGACCA 4540
1188 AAGUGUUU G CUGACGCA 1490 TGCGTCAG GGCTAGCTACAACGA AAACACTT 4541
1194 UUGCUGAC G CAACCCCC 1492 GGGGGTTG GGCTAGCTACAACGA GTCAGCAA 4542
1234 CCAUCAGC G CAUGCGUG 1493 CACGCATG GGCTAGCTACAACGA GCTGATGG 4543
1238 CAGCGCAU G CGUGGAAC 1494 GTTCCACG GGCTAGCTACAACGA ATGCGCTG 4544
1262 UCUCCUCU G CCGAUCCA 1495 TGGATCGG GGCTAGCTACAACGA AGAGGAGA 4545
1275 UCCAUACC G CGGAACUC 1497 GAGTTCCG GGCTAGCTACAACGA GGTATGGA 4546
1290 UCCUAGCC G CUUGUUUU 1498 AAAACAAG GGCTAGCTACAACGA GGCTAGGA 4547
1299 CUUGUUUU G CUCGCAGC 1499 GCTGCGAG GGCTAGCTACAACGA AAAACAAG 4548
1303 UUUUGCUC G CAGCAGGU 1500 ACCTGCTG GGCTAGCTACAACGA GAGCAAAA 4549
1349 UCUGUCGU G CUCUCCCG 1502 CGGGAGAG GGCTAGCTACAACGA ACGACAGA 4550
1357 GCUCUCCC G CAAAUAUA 1503 TATATTTG GGCTAGCTACAACGA GGGAGAGC 4551
Taσie i
1382 CCAUGGCU G CUAGGCUG 1504 CAGCCTAG GGCTAGCTACAACGA AGCCATGG 4552
1392 UAGGCUGU G CUGCCAAC 1505 GTTGGCAG GGCTAGCTACAACGA ACAGCCTA 4553
1395 GCUGUGCU G CCAACUGG 1506 CCAGTTGG GGCTAGCTACAACGA AGCACAGC 4554
1411 GAUCCUAC G CGGGACGU 1507 ACGTCCCG GGCTAGCTACAACGA GTAGGATC 4555
1442 CCGUCGGC G CUGAAUCC 1508 GGATTCAG GGCTAGCTACAACGA GCCGACGG 4556
1452 UGAAUCCC G CGGACGAC 1510 GTCGTCCG GGCTAGCTACAACGA GGGATTCA 4557
1474 CCGGGGCC G CUUGGGGC 1512 GCCCCAAG GGCTAGCTACAACGA GGCCCCGG 4558
1489 GCUCUACC G CCCGCUUC 1513 GAAGCGGG GGCTAGCTACAACGA GGTAGAGC 4559
1493 UACCGCCC G CUUCUCCG 1514 CGGAGAAG GGCTAGCTACAACGA GGGCGGTA 4560
1501 GCUUCUCC G CCUAUUGU 1515 ACAATAGG GGCTAGCTACAACGA GGAGAAGC 4561
1528 CACGGGGC G CACCUCUC 1517 GAGAGGTG GGCTAGCTACAACGA GCCCCGTG 4562
1542 CUCUUUAC G CGGACUCC 1518 GGAGTCCG GGCTAGCTACAACGA GTAAAGAG 4563
1559 CCGUCUGU G CCUUCUCA 1519 TGAGAAGG GGCTAGCTACAACGA ACAGACGG 4564
1571 UCUCAUCU G CCGGACCG 1520 CGGTCCGG GGCTAGCTACAACGA AGATGAGA 4565
1583 GACCGUGU G CACUUCGC 1521 GCGAAGTG GGCTAGCTACAACGA ACACGGTC 4566
1590 UGCACUUC G CUUCACCU 1522 AGGTGAAG GGCTAGCTACAACGA GAAGTGCA 4567
1601 UCACCUCU G CACGUCGC 1523 GCGACGTG GGCTAGCTACAACGA AGAGGTGA 4568
1608 UGCACGUC G CAUGGAGA 1524 TCTCCATG GGCTAGCTACAACGA GACGTGCA 4569
1628 CCGUGAAC G CCCACAGG 1526 CCTGTGGG GGCTAGCTACAACGA GTTCACGG 4570
1642 AGGAACCU G CCCAAGGU 1527 ACCTTGGG GGCTAGCTACAACGA AGGTTCCT 4571
1654 AAGGUCUU G CAUAAGAG 1528 CTCTTATG GGCTAGCTACAACGA AAGACCTT 4572
1818 AGCACCAU G CAACUUUU 1533 AAAAGTTG GGCTAGCTACAACGA ATGGTGCT 4573
1835 UCACCUCU G CCUAAUCA 1534 TGATTAGG GGCTAGCTACAACGA AGAGGTGA 4574
1883 CAAGCUGU G CCUUGGGU 1535 ACCCAAGG GGCTAGCTACAACGA ACAGCTTG 4575
1959 UCUUUUUU G CCUUCUGA 1537 TCAGAAGG GGCTAGCTACAACGA AAAAAAGA 4576
2002 UCGACACC G CCUCUGCU 1541 AGCAGAGG GGCTAGCTACAACGA GGTGTCGA 4577
2008 CCGCCUCU G CUCUGUAU 1542 ATACAGAG GGCTAGCTACAACGA AGAGGCGG 4578
2282 GUGGAUUC G CACUCCUC 1548 GAGGAGTG GGCTAGCTACAACGA GAATCCAC 4579
2293 CUCCUCCU G CAUAUAGA 1549 TCTATATG GGCTAGCTACAACGA AGGAGGAG 4580
2311 CACCAAAU G CCCCUAUC 1550 GATAGGGG GGCTAGCTACAACGA ATTTGGTG 4581
2388 ACUCCCUC G CCUCGCAG 1552 CTGCGAGG GGCTAGCTACAACGA GAGGGAGT 4582
2393 CUCGCCUC G CAGACGAA 1553 TTCGTCTG GGCTAGCTACAACGA GAGGCGAG 4583
2412 UCUCAAUC G CCGCGUCG 1555 CGACGCGG GGCTAGCTACAACGA GATTGAGA 4584
2415 CAAUCGCC G CGUCGCAG 1556 CTGCGACG GGCTAGCTACAACGA GGCGATTG 4585
2420 GCCGCGUC G CAGAAGAU 1557 ATCTTCTG GGCTAGCTACAACGA GACGCGGC 4586
2514 GGUACCUU G CUUUAAUC 1558 GATTAAAG GGCTAGCTACAACGA AAGGTACC 4587
2560 AUUCAUUU G CAGGAGGA 1560 TCCTCCTG GGCTAGCTACAACGA AAATGAAT 4588
2641 UUAACUAU G CCUGCUAG 1563 CTAGCAGG GGCTAGCTACAACGA ATAGTTAA 4589
2645 CUAUGCCU G CUAGGUUU 1564 AAACCTAG GGCTAGCTACAACGA AGGCATAG 4590
2677 AAAUAUUU G CCCUUAGA 1565 TCTAAGGG GGCTAGCTACAACGA AAATATTT 4591
2740 UUCCAGAC G CGACAUUA 1566 TAATGTCG GGCTAGCTACAACGA GTCTGGAA 4592
2804 CACGUAGC G CCUCAUUU 1568 AAATGAGG GGCTAGCTACAACGA GCTACGTG 4593
2814 CUCAUUUU G CGGGUCAC 1569 GTGACCCG GGCTAGCTACAACGA AAAATGAG 4594
2946 UGGACCCU G CAUUCAAA 1572 TTTGAATG GGCTAGCTACAACGA AGGGTCCA 4595
2990 CUCAACCC G CACAAGGA 1573 TCCTTGTG GGCTAGCTACAACGA GGGTTGAG 4596
3012 GGCCGGAC G CCAACAAG 1574 CTTGTTGG GGCTAGCTACAACGA GTCCGGCC 4597
3090 GCCCUCAC G CUCAGGGC 1575 GCCCTGAG GGCTAGCTACAACGA GTGAGGGC 4598
3113 ACAACUGU G CCAGCAGC 1576 GCTGCTGG GGCTAGCTACAACGA ACAGTTGT 4599
3132 CUCCUCCU G CCUCCACC 1577 GGTGGAGG GGCTAGCTACAACGA AGGAGGAG 4600
51 AGGGCCCU G UACUUUCC 1578 GGAAAGTA GGCTAGCTACAACGA AGGGCCCT 4601
106 AGAAUACU G UCUCUGCC 1579 GGCAGAGA GGCTAGCTACAACGA AGTATTCT 4602
TaDie i
148 GGGACCCU G UACCGAAC 1580 GTTCGGTA GGCTAGCTACAACGA AGGGTCCC 4603
198 CUGCUCGU G UUACAGGC 1581 GCCTGTAA GGCTAGCTACAACGA ACGAGCAG 4604
219 UUUUUCUU G UUGACAAA 1582 TTTGTCAA GGCTAGCTACAACGA AAGAAAAA 4605
297 ACACCCGU G UGUCUUGG 1583 CCAAGACA GGCTAGCTACAACGA ACGGGTGT 4606
299 ACCCGUGU G UCUUGGCC 1584 GGCCAAGA GGCTAGCTACAACGA ACACGGGT 4607
347 ACCAACCU G UUGUCCUC 1585 GAGGACAA GGCTAGCTACAACGA AGGTTGGT 4608
350 AACCUGUU G UCCUCCAA 1586 TTGGAGGA GGCTAGCTACAACGA AACAGGTT 4609
362 UCCAAUUU G UCCUGGUU 1587 AACCAGGA GGCTAGCTACAACGA AAATTGGA 4610
381 CGCUGGAU G UGUCUGCG 1588 CGCAGACA GGCTAGCTACAACGA ATCCAGCG 4611
383 CUGGAUGU G UCUGCGGC 1589 GCCGCAGA GGCTAGCTACAACGA ACATCCAG 4612
438 AUCUUCUU G UUGGUUCU 1590 AGAACCAA GGCTAGCTACAACGA AAGAAGAT 4613
465 CAAGGUAU G UUGCCCGU 1591 ACGGGCAA GGCTAGCTACAACGA ATACCTTG 4614
476 GCCCGUUU G UCCUCUAA 1592 TTAGAGGA GGCTAGCTACAACGA AAACGGGC 4615
555 ACCUCUAU G UUUCCCUC 1593 GAGGGAAA GGCTAGCTACAACGA ATAGAGGT 4616
566 UCCCUCAU G UUGCUGUA 1594 TACAGCAA GGCTAGCTACAACGA ATGAGGGA 4617
572 AUGUUGCU G UACAAAAC 1595 GTTTTGTA GGCTAGCTACAACGA AGCAACAT 4618
602 CUGCACCU G UAUUCCCA 1596 TGGGAATA GGCTAGCTACAACGA AGGTGCAG 4619
694 UGCCAUUU G UUCAGUGG 1597 CCACTGAA GGCTAGCTACAACGA AAATGGCA 4620
724 CCCCCACU G UCUGGCUU 1598 AAGCCAGA GGCTAGCTACAACGA AGTGGGGG 4621
750 UGGAUGAU G UGGUUUUG 1599 CAAAACCA GGCTAGCTACAACGA ATCATCCA 4622
771 CCAAGUCU G UACAACAU 1600 ATGTTGTA GGCTAGCTACAACGA AGACTTGG 4623
801 AUGCCGCU G UUACCAAU 1601 ATTGGTAA GGCTAGCTACAACGA AGCGGCAT 4624
818 UUUCUUUU G UCUUUGGG 1602 CCCAAAGA GGCTAGCTACAACGA AAAAGAAA 4625
888 UGGGAUAU G UAAUUGGG 1603 CCCAATTA GGCTAGCTACAACGA ATATCCCA 4626
927 AACAUAUU G UACAAAAA 1604 TTTTTGTA GGCTAGCTACAACGA AATATGTT 4627
944 AUCAAAAU G UGUUUUAG 1605 CTAAAACA GGCTAGCTACAACGA ATTTTGAT 4628
946 CAAAAUGU G UUUUAGGA 1606 TCCTAAAA GGCTAGCTACAACGA ACATTTTG 4629
963 AACUUCCU G UAAACAGG 1607 CCTGTTTA GGCTAGCTACAACGA AGGAAGTT 4630
991 GAAAGUAU G UCAACGAA 1608 TTCGTTGA GGCTAGCTACAACGA ATACTTTC 4631
1002 AACGAAUU G UGGGUCUU 1609 AAGACCCA GGCTAGCTACAACGA AATTCGTT 4632
1039 CACGCAAU G UGGAUAUU 1610 AATATCCA GGCTAGCTACAACGA ATTGCGTG 4633
1137 AACAGUAU G UGAACCUU 1611 AAGGTTCA GGCTAGCTACAACGA ATACTGTT 4634
1184 UGCCAAGU G UUUGCUGA 1612 TCAGCAAA GGCTAGCTACAACGA ACTTGGCA 4635
1251 GAACCUUU G UGUCUCCU 1613 AGGAGACA GGCTAGCTACAACGA AAAGGTTC 4636
1253 ACCUUUGU G UCUCCUCU 1614 AGAGGAGA GGCTAGCTACAACGA ACAAAGGT 4637
1294 AGCCGCUU G UUUUGCUC 1615 GAGCAAAA GGCTAGCTACAACGA AAGCGGCT 4638
1344 ACAAUUCU G UCGUGCUC 1616 GAGCACGA GGCTAGCTACAACGA AGAATTGT 4639
1390 GCUAGGCU G UGCUGCCA 1617 TGGCAGCA GGCTAGCTACAACGA AGCCTAGC 4640
1425 CGUCCUUU G UUUACGUC 1618 GACGTAAA GGCTAGCTACAACGA AAAGGACG 4641
1508 CGCCUAUU G UACCGACC 1619 GGTCGGTA GGCTAGCTACAACGA AATAGGCG 4642
1557 CCCCGUCU G UGCCUUCU 1620 AGAAGGCA GGCTAGCTACAACGA AGACGGGG 4643
1581 CGGACCGU G UGCACUUC 1621 GAAGTGCA GGCTAGCTACAACGA ACGGTCCG 4644
1684 UCAGCAAU G UCAACGAC 1622 GTCGTTGA GGCTAGCTACAACGA ATTGCTGA 4645
1719 CAAAGACU G UGUGUUUA 1623 TAAACACA GGCTAGCTACAACGA AGTCTTTG 4646
1721 AAGACUGU G UGUUUAAU 1624 ATTAAACA GGCTAGCTACAACGA ACAGTCTT 4647
1723 GACUGUGU G UUUAAUGA 1625 TCATTAAA GGCTAGCTACAACGA ACACAGTC 4648
1772 AGGUCUUU G UACUAGGA 1626 TCCTAGTA GGCTAGCTACAACGA AAAGACCT 4649
1785 AGGAGGCU G UAGGCAUA 1627 TATGCCTA GGCTAGCTACAACGA AGCCTCCT 4650
1801 AAAUUGGU G UGUUCACC 1628 GGTGAACA GGCTAGCTACAACGA ACCAATTT 4651
1803 AUUGGUGU G UUCACCAG 1629 CTGGTGAA GGCTAGCTACAACGA ACACCAAT 4652
1850 CAUCUCAU G UUCAUGUC 1630 GACATGAA GGCTAGCTACAACGA ATGAGATG 4653
Taσie 4 1
1856 AUGUUCAU G UCCUACUG 1631 CAGTAGGA GGCTAGCTACAACGA ATGAACAT 4654
1864 GUCCUACU G UUCAAGCC 1632 GGCTTGAA GGCTAGCTACAACGA AGTAGGAC 4655
1881 UCCAAGCU G UGCCUUGG 1633 CCAAGGCA GGCTAGCTACAACGA AGCTTGGA 4656
1939 GAGCUUCU G UGGAGUUA 1634 TAACTCCA GGCTAGCTACAACGA AGAAGCTC 4657
2013 UCUGCUCU G UAUCGGGG 1635 CCCCGATA GGCTAGCTACAACGA AGAGCAGA 4658
2045 GGAACAUU G UUCACCUC 1636 GAGGTGAA GGCTAGCTACAACGA AATGTTCC 4659
2082 GCUAUUCU G UGUUGGGG 1637 CCCCAACA GGCTAGCTACAACGA AGAATAGC 4660
2084 UAUUCUGU G UUGGGGUG 1638 CACCCCAA GGCTAGCTACAACGA ACAGAATA 4661
2167 UCAGCUAU G UCAACGUU 1639 AACGTTGA GGCTAGCTACAACGA ATAGCTGA 4662
2205 CAACUAUU G UGGUUUCA 1640 TGAAACCA GGCTAGCTACAACGA AATAGTTG 4663
2222 CAUUUCCU G UCUUACUU 1641 AAGTAAGA GGCTAGCTACAACGA AGGAAATG 4664
2245 GAGAAACU G UUCUUGAA 1642 TTCAAGAA GGCTAGCTACAACGA AGTTTCTC 4665
2262 UAUUUGGU G UCUUUUGG 1643 CCAAAAGA GGCTAGCTACAACGA ACCAAATA 4666
2274 UUUGGAGU G UGGAUUCG 1644 CGAATCCA GGCTAGCTACAACGA ACTCCAAA 4667
2344 AAACUACU G UUGUUAGA 1645 TCTAACAA GGCTAGCTACAACGA AGTAGTTT 4668
2347 CUACUGUU G UUAGACGA 1646 TCGTCTAA GGCTAGCTACAACGA AACAGTAG 4669
2450 AUCUCAAU G UUAGUAUU 1647 AATACTAA GGCTAGCTACAACGA ATTGAGAT 4670
2573 AGGACAUU G UUGAUAGA 1648 TCTATCAA GGCTAGCTACAACGA AATGTCCT 4671
2583 UGAUAGAU G UAAGCAAU 1649 ATTGCTTA GGCTAGCTACAACGA ATCTATCA 4672
2594 AGCAAUUU G UGGGGCCC 1650 GGGCCCCA GGCTAGCTACAACGA AAATTGCT 4673
2663 AUCCCAAU G UUACUAAA 1651 TTTAGTAA GGCTAGCTACAACGA ATTGGGAT 4674
2717 CAGAGUAU G UAGUUAAU 1652 ATTAACTA GGCTAGCTACAACGA ATACTCTG 4675
2901 AUCUUUCU G UCCCCAAU 1653 ATTGGGGA GGCTAGCTACAACGA AGAAAGAT 4676
3071 GGGGGACU G UUGGGGUG 1654 CACCCCAA GGCTAGCTACAACGA AGTCCCCC 4677
3111 UCACAACU G UGCCAGCA 1655 TGCTGGCA GGCTAGCTACAACGA AGTTGTGA 4678
40 AUCCCAGA G UCAGGGCC 1656 GGCCCTGA GGCTAGCTACAACGA TCTGGGAT 4679
46 GAGUCAGG G CCCUGUAC 1657 GTACAGGG GGCTAGCTACAACGA CCTGACTC 4680
65 UCCUGCUG G UGGCUCCA 1658 TGGAGCCA GGCTAGCTACAACGA CAGCAGGA 4681
68 UGCUGGUG G CUCCAGUU 1659 AACTGGAG GGCTAGCTACAACGA CACCAGCA 4682
74 UGGCUCCA G UUCAGGAA 1660 TTCCTGAA GGCTAGCTACAACGA TGGAGCCA 4683
85 CAGGAACA G UGAGCCCU 1661 AGGGCTCA GGCTAGCTACAACGA TGTTCCTG 4684
89 AACAGUGA G CCCUGCUC 1662 GAGCAGGG GGCTAGCTACAACGA TCACTGTT 4685
120 GCCAUAUC G UCAAUCUU 1663 AAGATTGA GGCTAGCTACAACGA GATATGGC 4686
196 CCCUGCUC G UGUUACAG 1664 CTGTAACA GGCTAGCTACAACGA GAGCAGGG 4687
205 UGUUACAG G CGGGGUUU 1665 AAACCCCG GGCTAGCTACAACGA CTGTAACA 4688
210 CAGGCGGG G UUUUUCUU 1666 AAGAAAAA GGCTAGCTACAACGA CCCGCCTG 4689
248 ACCACAGA G UCUAGACU 1667 AGTCTAGA GGCTAGCTACAACGA TCTGTGGT 4690
258 CUAGACUC G UGGUGGAC 1668 GTCCACCA GGCTAGCTACAACGA GAGTCTAG 4691
261 GACUCGUG G UGGACUUC 1669 GAAGTCCA GGCTAGCTACAACGA CACGAGTC 4692
295 GAACACCC G UGUGUCUU 1670 AAGACACA GGCTAGCTACAACGA GGGTGTTC 4693
305 GUGUCUUG G CCAAAAUU 1671 AATTTTGG GGCTAGCTACAACGA CAAGACAC 4694
318 AAUUCGCA G UCCCAAAU 1672 ATTTGGGA GGCTAGCTACAACGA TGCGAATT 4695
332 AAUCUCCA G UCACUCAC 1673 GTGAGTGA GGCTAGCTACAACGA TGGAGATT 4696
368 UUGUCCUG G UUAUCGCU 1674 AGCGATAA GGCTAGCTACAACGA CAGGACAA 4697
390 UGUCUGCG G CGUUUUAU 1675 ATAAAACG GGCTAGCTACAACGA CGCAGACA 4698
392 UCUGCGGC G UUUUAUCA 1676 TGATAAAA GGCTAGCTACAACGA GCCGCAGA 4699
442 UCUUGUUG G UUCUUCUG 1677 CAGAAGAA GGCTAGCTACAACGA CAACAAGA 4700
461 CUAUCAAG G UAUGUUGC 1678 GCAACATA GGCTAGCTACAACGA CTTGATAG 4701
472 UGUUGCCC G UUUGUCCU 1679 AGGACAAA GGCTAGCTACAACGA GGGCAACA 4702
506 AACAACCA G CACCGGAC 1680 GTCCGGTG GGCTAGCTACAACGA TGGTTGTT 4703
625 CAUCUUGG G CUUUCGCA 1681 TGCGAAAG GGCTAGCTACAACGA CCAAGATG 4704
Taoie 1
648 CUAUGGGA G UGGGCCUC 1682 GAGGCCCA GGCTAGCTACAACGA TCCCATAG 4705
652 GGGAGUGG G CCUCAGUC 1683 GACTGAGG GGCTAGCTACAACGA CCACTCCC 4706
658 GGGCCUCA G UCCGUUUC 1684 GAAACGGA GGCTAGCTACAACGA TGAGGCCC 4707
662 CUCAGUCC G UUUCUCUU 1685 AAGAGAAA GGCTAGCTACAACGA GGACTGAG 4708
672 UUCUCUUG G CUCAGUUU 1686 AAACTGAG GGCTAGCTACAACGA CAAGAGAA 4709
677 UUGGCUCA G UUUACUAG 1687 CTAGTAAA GGCTAGCTACAACGA TGAGCCAA 4710
685 GUUUACUA G UGCCAUUU 1688 AAATGGCA GGCTAGCTACAACGA TAGTAAAC 4711
699 UUUGUUCA G UGGUUCGU 1689 ACGAACCA GGCTAGCTACAACGA TGAACAAA 4712
702 GUUCAGUG G UUCGUAGG 1690 CCTACGAA GGCTAGCTACAACGA CACTGAAC 4713
706 AGUGGUUC G UAGGGCUU 1691 AAGCCCTA GGCTAGCTACAACGA GAACCACT 4714
711 UUCGUAGG G CUUUCCCC 1692 GGGGAAAG GGCTAGCTACAACGA CCTACGAA 4715
729 ACUGUCUG G CUUUCAGU 1693 ACTGAAAG GGCTAGCTACAACGA CAGACAGT 4716
736 GGCUUUCA G UUAUAUGG 1694 CCATATAA GGCTAGCTACAACGA TGAAAGCC 4717
753 AUGAUGUG G UUUUGGGG 1695 CCCCAAAA GGCTAGCTACAACGA CACATCAT 4718
762 UUUUGGGG G CCAAGUCU 1696 AGACTTGG GGCTAGCTACAACGA CCCCAAAA 4719
767 GGGGCCAA G UCUGUACA 1697 TGTACAGA GGCTAGCTACAACGA TTGGCCCC 4720
785 CAUCUUGA G UCCCUUUA 1698 TAAAGGGA GGCTAGCTACAACGA TCAAGATG 4721
826 GUCUUUGG G UAUACAUU 1699 AATGTATA GGCTAGCTACAACGA CCAAAGAC 4722
898 AAUUGGGA G UUGGGGCA 1700 TGCCCCAA GGCTAGCTACAACGA TCCCAATT 4723
904 GAGUUGGG G CACAUUGC 1701 GCAATGTG GGCTAGCTACAACGA CCCAACTC 4724
971 GUAAACAG G CCUAUUGA 1702 TCAATAGG GGCTAGCTACAACGA CTGTTTAC 4725
987 AUUGGAAA G UAUGUCAA 1703 TTGACATA GGCTAGCTACAACGA TTTCCAAT 4726
1006 AAUUGUGG G UCUUUUGG 1704 CCAAAAGA GGCTAGCTACAACGA CCACAATT 4727
1016 CUUUUGGG G UUUGCCGC 1705 GCGGCAAA GGCTAGCTACAACGA CCCAAAAG 4728
1080 GCAUACAA G CAAAACAG 1706 CTGTTTTG GGCTAGCTACAACGA TTGTATGC 4729
1089 CAAAACAG G CUUUUACU 1707 AGTAAAAG GGCTAGCTACAACGA CTGTTTTG 4730
1116 CUUACAAG G CCUUUCUA 1708 TAGAAAGG GGCTAGCTACAACGA CTTGTAAG 4731
1126 CUUUCUAA G UAAACAGU 1709 ACTGTTTA GGCTAGCTACAACGA TTAGAAAG 4732
1133 AGUAAACA G UAUGUGAA 1710 TTCACATA GGCTAGCTACAACGA TGTTTACT 4733
1152 UUUACCCC G UUGCUCGG 1711 CCGAGCAA GGCTAGCTACAACGA GGGGTAAA 4734
1160 GUUGCUCG G CAACGGCC 1712 GGCCGTTG GGCTAGCTACAACGA CGAGCAAC 4735
1166 CGGCAACG G CCUGGUCU 1713 AGACCAGG GGCTAGCTACAACGA CGTTGCCG 4736
1171 ACGGCCUG G UCUAUGCC 1714 GGCATAGA GGCTAGCTACAACGA CAGGCCGT 4737
1182 UAUGCCAA G UGUUUGCU 1715 AGCAAACA GGCTAGCTACAACGA TTGGCATA 4738
1207 CCCCACUG G UUGGGGCU 1716 AGCCCCAA GGCTAGCTACAACGA CAGTGGGG 4739
1213 UGGUUGGG G CUUGGCCA 1717 TGGCCAAG GGCTAGCTACAACGA CCCAACCA 4740
1218 GGGGCUUG G CCAUAGGC 1718 GCCTATGG GGCTAGCTACAACGA CAAGCCCC 4741
1225 GGCCAUAG G CCAUCAGC 1719 GCTGATGG GGCTAGCTACAACGA CTATGGCC 4742
1232 GGCCAUCA G CGCAUGCG 1720 CGCATGCG GGCTAGCTACAACGA TGATGGCC 4743
1240 GCGCAUGC G UGGAACCU 1721 AGGTTCCA GGCTAGCTACAACGA GCATGCGC 4744
1287 AACUCCUA G CCGCUUGU 1722 ACAAGCGG GGCTAGCTACAACGA TAGGAGTT 4745
1306 UGCUCGCA G CAGGUCUG 1723 CAGACCTG GGCTAGCTACAACGA TGCGAGCA 4746
1310 CGCAGCAG G UCUGGGGC 1724 GCCCCAGA GGCTAGCTACAACGA CTGCTGCG 4747
1317 GGUCUGGG G CAAAACUC 1725 GAGTTTTG GGCTAGCTACAACGA CCCAGACC 4748
1347 AUUCUGUC G UGCUCUCC 1726 GGAGAGCA GGCTAGCTACAACGA GACAGAAT 4749
1379 UUUCCAUG G CUGCUAGG 1727 CCTAGCAG GGCTAGCTACAACGA CATGGAAA 4750
1387 GCUGCUAG G CUGUGCUG 1728 CAGCACAG GGCTAGCTACAACGA CTAGCAGC 4751
1418 CGCGGGAC G UCCUUUGU 1729 ACAAAGGA GGCTAGCTACAACGA GTCCCGCG 4752
1431 UUGUUUAC G UCCCGUCG 1730 CGACGGGA GGCTAGCTACAACGA GTAAACAA 4753
1436 UACGUCCC G UCGGCGCU 1731 AGCGCCGA GGCTAGCTACAACGA GGGACGTA 4754
1440 UCCCGUCG G CGCUGAAU 1732 ATTCAGCG GGCTAGCTACAACGA CGACGGGA 4755
Taoie 4 1
1471 CUCCCGGG G CCGCUUGG 1733 CCAAGCGG GGCTAGCTACAACGA CCCGGGAG 4756
1481 CGCUUGGG G CUCUACCG 1734 CGGTAGAG GGCTAGCTACAACGA CCCAAGCG 4757
1517 UACCGACC G UCCACGGG 1735 CCCGTGGA GGCTAGCTACAACGA GGTCGGTA 4758
1526 UCCACGGG G CGCACCUC 1736 GAGGTGCG GGCTAGCTACAACGA CCCGTGGA 4759
1553 GACUCCCC G UCUGUGCC 1737 GGCACAGA GGCTAGCTACAACGA GGGGAGTC 4760
1579 GCCGGACC G UGUGCACU 1738 AGTGCACA GGCTAGCTACAACGA GGTCCGGC 4761
1605 CUCUGCAC G UCGCAUGG 1739 CCATGCGA GGCTAGCTACAACGA GTGCAGAG 4762
1622 AGACCACC G UGAACGCC 1740 GGCGTTCA GGCTAGCTACAACGA GGTGGTCT 4763
1649 UGCCCAAG G UCUUGCAU 1741 ATGCAAGA GGCTAGCTACAACGA CTTGGGCA 4764
1679 GACUUUCA G CAAUGUCA 1742 TGACATTG GGCTAGCTACAACGA TGAAAGTC 4765
1703 ACCUUGAG G CAUACUUC 1743 GAAGTATG GGCTAGCTACAACGA CTCAAGGT 4766
1732 UUUAAUGA G UGGGAGGA 1744 TCCTCCCA GGCTAGCTACAACGA TCATTAAA 4767
1741 UGGGAGGA G UUGGGGGA 1745 TCCCCCAA GGCTAGCTACAACGA TCCTCCCA 4768
1754 GGGAGGAG G UUAGGUUA 1746 TAACCTAA GGCTAGCTACAACGA CTCCTCCC 4769
1759 GAGGUUAG G UUAAAGGU 1747 ACCTTTAA GGCTAGCTACAACGA CTAACCTC 4770
1766 GGUUAAAG G UCUUUGUA 1748 TACAAAGA GGCTAGCTACAACGA CTTTAACC 4771
1782 ACUAGGAG G CUGUAGGC 1749 GCCTACAG GGCTAGCTACAACGA CTCCTAGT 4772
1789 GGCUGUAG G CAUAAAUU 1750 AATTTATG GGCTAGCTACAACGA CTACAGCC 4773
1799 AUAAAUUG G UGUGUUCA 1751 TGAACACA GGCTAGCTACAACGA CAATTTAT 4774
1811 GUUCACCA G CACCAUGC 1752 GCATGGTG GGCTAGCTACAACGA TGGTGAAC 4775
1870 CUGUUCAA G CCUCCAAG 1753 CTTGGAGG GGCTAGCTACAACGA TTGAACAG 4776
1878 GCCUCCAA G CUGUGCCU 1754 AGGCACAG GGCTAGCTACAACGA TTGGAGGC 4777
1890 UGCCUUGG G UGGCUUUG 1755 CAAAGCCA GGCTAGCTACAACGA CCAAGGCA 4778
1893 CUUGGGUG G CUUUGGGG 1756 CCCCAAAG GGCTAGCTACAACGA CACCCAAG 4779
1901 GCUUUGGG G CAUGGACA 1757 TGTCCATG GGCTAGCTACAACGA CCCAAAGC 4780
1917 AUUGACCC G UAUAAAGA 1758 TCTTTATA GGCTAGCTACAACGA GGGTCAAT 4781
1933 AAUUUGGA G CUUCUGUG 1759 CACAGAAG GGCTAGCTACAACGA TCCAAATT 4782
1944 UCUGUGGA G UUACUCUC 1760 GAGAGTAA GGCTAGCTACAACGA TCCACAGA 4783
2023 AUCGGGGG G CCUUAGAG 1761 CTCTAAGG GGCTAGCTACAACGA CCCCCGAT 4784
2031 GCCUUAGA G UCUCCGGA 1762 TCCGGAGA GGCTAGCTACAACGA TCTAAGGC 4785
2062 ACCAUACG G CACUCAGG 1763 CCTGAGTG GGCTAGCTACAACGA CGTATGGT 4786
2070 GCACUCAG G CAAGCUAU 1764 ATAGCTTG GGCTAGCTACAACGA CTGAGTGC 4787
2074 UCAGGCAA G CUAUUCUG 1765 CAGAATAG GGCTAGCTACAACGA TTGCCTGA 4788
2090 GUGUUGGG G UGAGUUGA 1766 TCAACTCA GGCTAGCTACAACGA CCCAACAC 4789
2094 UGGGGUGA G UUGAUGAA 1767 TTCATCAA GGCTAGCTACAACGA TCACCCCA 4790
2107 UGAAUCUA G CCACCUGG 1768 CCAGGTGG GGCTAGCTACAACGA TAGATTCA 4791
2116 CCACCUGG G UGGGAAGU 1769 ACTTCCCA GGCTAGCTACAACGA CCAGGTGG 4792
2123 GGUGGGAA G UAAUUUGG 1770 CCAAATTA GGCTAGCTACAACGA TTCCCACC 4793
2140 AAGAUCCA G CAUCCAGG 1771 CCTGGATG GGCTAGCTACAACGA TGGATCTT 4794
2155 GGGAAUUA G UAGUCAGC 1772 GCTGACTA GGCTAGCTACAACGA TAATTCCC 4795
2158 AAUUAGUA G UCAGCUAU 1773 ATAGCTGA GGCTAGCTACAACGA TACTAATT 4796
2162 AGUAGUCA G CUAUGUCA 1774 TGACATAG GGCTAGCTACAACGA TGACTACT 4797
2173 AUGUCAAC G UUAAUAUG 1775 CATATTAA GGCTAGCTACAACGA GTTGACAT 4798
2183 UAAUAUGG G CCUAAAAA 1776 TTTTTAGG GGCTAGCTACAACGA CCATATTA 4799
2208 CUAUUGUG G UUUCACAU 1777 ATGTGAAA GGCTAGCTACAACGA CACAATAG 4800
2235 ACUUUUGG G CGAGAAAC 1778 GTTTCTCG GGCTAGCTACAACGA CCAAAAGT 4801
2260 AAUAUUUG G UGUCUUUU 1779 AAAAGACA GGCTAGCTACAACGA CAAATATT 4802
2272 CUUUUGGA G UGUGGAUU 1780 AATCCACA GGCTAGCTACAACGA TCCAAAAG 4803
2360 ACGAAGAG G CAGGUCCC 1781 GGGACCTG GGCTAGCTACAACGA CTCTTCGT 4804
2364 AGAGGCAG G UCCCCUAG 1782 CTAGGGGA GGCTAGCTACAACGA CTGCCTCT 4805
2403 AGACGAAG G UCUCAAUC 1783 GATTGAGA GGCTAGCTACAACGA CTTCGTCT 4806
Taoie 4 1
2417 AUCGCCGC G UCGCAGAA 1784 TTCTGCGA GGCTAGCTACAACGA GCGGCGAT 4807
2454 CAAUGUUA G UAUUCCUU 1785 AAGGAATA GGCTAGCTACAACGA TAACATTG 4808
2474 CACAUAAG G UGGGAAAC 1786 GTTTCCCA GGCTAGCTACAACGA CTTATGTG 4809
2491 UUUACGGG G CUUUAUUC 1787 GAATAAAG GGCTAGCTACAACGA CCCGTAAA 4810
2507 CUUCUACG G UACCUUGC 1788 GCAAGGTA GGCTAGCTACAACGA CGTAGAAG 4811
2530 CCUAAAUG G CAAACUCC 1789 GGAGTTTG GGCTAGCTACAACGA CATTTAGG 4812
2587 AGAUGUAA G CAAUUUGU 1790 ACAAATTG GGCTAGCTACAACGA TTACATCT 4813
2599 UUUGUGGG G CCCCUUAC 1791 GTAAGGGG GGCTAGCTACAACGA CCCACAAA 4814
2609 CCCUUACA G UAAAUGAA 1792 TTCATTTA GGCTAGCTACAACGA TGTAAGGG 4815
2650 CCUGCUAG G UUUUAUCC 1793 GGATAAAA GGCTAGCTACAACGA CTAGCAGG 4816
2701 AUCAAACC G UAUUAUCC 1794 GGATAATA GGCTAGCTACAACGA GGTTTGAT 4817
2713 UAUCCAGA G UAUGUAGU 1795 ACTACATA GGCTAGCTACAACGA TCTGGATA 4818
2720 AGUAUGUA G UUAAUCAU 1796 ATGATTAA GGCTAGCTACAACGA TACATACT 4819
2768 UUUGGAAG G CGGGGAUC 1797 GATCCCCG GGCTAGCTACAACGA CTTCCAAA 4820
2791 AAAAGAGA G UCCACACG 1798 CGTGTGGA GGCTAGCTACAACGA TCTCTTTT 4821
2799 GUCCACAC G UAGCGCCU 1799 AGGCGCTA GGCTAGCTACAACGA GTGTGGAC 4822
2802 CACACGUA G CGCCUCAU 1800 ATGAGGCG GGCTAGCTACAACGA TACGTGTG 4823
2818 UUUUGCGG G UCACCAUA 1801 TATGGTGA GGCTAGCTACAACGA CCGCAAAA 4824
2848 GAUCUACA G CAUGGGAG 1802 CTCCCATG GGCTAGCTACAACGA TGTAGATC 4825
2857 CAUGGGAG G UUGGUCUU 1803 AAGACCAA GGCTAGCTACAACGA CTCCCATG 4826
2861 GGAGGUUG G UCUUCCAA 1804 TTGGAAGA GGCTAGCTACAACGA CAACCTCC 4827
2881 UCGAAAAG G CAUGGGGA 1805 TCCCCATG GGCTAGCTACAACGA CTTTTCGA 4828
2936 GAUCAUCA G UUGGACCC 1806 GGGTCCAA GGCTAGCTACAACGA TGATGATC 4829
2955 CAUUCAAA G CCAACUCA 1807 TGAGTTGG GGCTAGCTACAACGA TTTGAATG 4830
2964 CCAACUCA G UAAAUCCA 1808 TGGATTTA GGCTAGCTACAACGA TGAGTTGG 4831
3005 GACAACUG G CCGGACGC 1809 GCGTCCGG GGCTAGCTACAACGA CAGTTGTC 4832
3021 CCAACAAG G UGGGAGUG 1810 CACTCCCA GGCTAGCTACAACGA CTTGTTGG 4833
3027 AGGUGGGA G UGGGAGCA 1811 TGCTCCCA GGCTAGCTACAACGA TCCCACCT 4834
3033 GAGUGGGA G CAUUCGGG 1812 CCCGAATG GGCTAGCTACAACGA TCCCACTC 4835
3041 GCAUUCGG G CCAGGGUU 1813 AACCCTGG GGCTAGCTACAACGA CCGAATGC 4836
3047 GGGCCAGG G UUCACCCC 1814 GGGGTGAA GGCTAGCTACAACGA CCTGGCCC 4837
3077 CUGUUGGG G UGGAGCCC 1815 GGGCTCCA GGCTAGCTACAACGA CCCAACAG 4838
3082 GGGGUGGA G CCCUCACG 1816 CGTGAGGG GGCTAGCTACAACGA TCCACCCC 4839
3097 CGCUCAGG G CCUACUCA 1817 TGAGTAGG GGCTAGCTACAACGA CCTGAGCG 4840
3117 CUGUGCCA G CAGCUCCU 1818 AGGAGCTG GGCTAGCTACAACGA TGGCACAG 4841
3120 UGCCAGCA G CUCCUCCU 1819 AGGAGGAG GGCTAGCTACAACGA TGCTGGCA 4842
3146 ACCAAUCG G CAGUCAGG 1820 CCTGACTG GGCTAGCTACAACGA CGATTGGT 4843
3149 AAUCGGCA G UCAGGAAG 1821 CTTCCTGA GGCTAGCTACAACGA TGCCGATT 4844
3158 UCAGGAAG G CAGCCUAC 1822 GTAGGCTG GGCTAGCTACAACGA CTTCCTGA 4845
3161 GGAAGGCA G CCUACUCC 1823 GGAGTAGG GGCTAGCTACAACGA TGCCTTCC 4846
3204 AUCCUCAG G CCAUGCAG 1824 CTGCATGG GGCTAGCTACAACGA CTGAGGAT 4847
10 ACUCCACC A CUUUCCAC 1825 GTGGAAAG GGCTAGCTACAACGA GGTGGAGT 4848
17 CACUUUCC A CCAAACUC 1826 GAGTTTGG GGCTAGCTACAACGA GGAAAGTG 4849
22 UCCACCAA A CUCUUCAA 1827 TTGAAGAG GGCTAGCTACAACGA TTGGTGGA 4850
32 UCUUCAAG A UCCCAGAG 1828 CTCTGGGA GGCTAGCTACAACGA CTTGAAGA 4851
53 GGCCCUGU A CUUUCCUG 1829 CAGGAAAG GGCTAGCTACAACGA ACAGGGCC 4852
82 GUUCAGGA A CAGUGAGC 1830 GCTCACTG GGCTAGCTACAACGA TCCTGAAC 4853
101 UGCUCAGA A UACUGUCU 1831 AGACAGTA GGCTAGCTACAACGA TCTGAGCA 4854
103 CUCAGAAU A CUGUCUCU 1832 AGAGACAG GGCTAGCTACAACGA ATTCTGAG 4855
115 UCUCUGCC A UAUCGUCA 1833 TGACGATA GGCTAGCTACAACGA GGCAGAGA 4856
117 UCUGCCAU A UCGUCAAU 1834 ATTGACGA GGCTAGCTACAACGA ATGGCAGA 4857
Taoie 4 1
124 UAUCGUCA A UCUUAUCG 1835 CGATAAGA GGCTAGCTACAACGA TGACGATA 4858
129 UCAAUCUU A UCGAAGAC 1836 GTCTTCGA GGCTAGCTACAACGA AAGATTGA 4859
136 UAUCGAAG A CUGGGGAC 1837 GTCCCCAG GGCTAGCTACAACGA CTTCGATA 4860
143 GACUGGGG A CCCUGUAC 1838 GTACAGGG GGCTAGCTACAACGA CCCCAGTC 4861
150 GACCCUGU A CCGAACAU 1839 ATGTTCGG GGCTAGCTACAACGA ACAGGGTC 4862
155 UGUACCGA A CAUGGAGA 1840 TCTCCATG GGCTAGCTACAACGA TCGGTACA 4863
157 UACCGAAC A UGGAGAAC 1841 GTTCTCCA GGCTAGCTACAACGA GTTCGGTA 4864
164 CAUGGAGA A CAUCGCAU 1842 ATGCGATG GGCTAGCTACAACGA TCTCCATG 4865
166 UGGAGAAC A UCGCAUCA 1843 TGATGCGA GGCTAGCTACAACGA GTTCTCCA 4866
171 AACAUCGC A UCAGGACU 1844 AGTCCTGA GGCTAGCTACAACGA GCGATGTT 4867
177 GCAUCAGG A CUCCUAGG 1845 CCTAGGAG GGCTAGCTACAACGA CCTGATGC 4868
186 CUCCUAGG A CCCCUGCU 1846 AGCAGGGG GGCTAGCTACAACGA CCTAGGAG 4869
201 CUCGUGUU A CAGGCGGG 1847 CCCGCCTG GGCTAGCTACAACGA AACACGAG 4870
223 UCUUGUUG A CAAAAAUC 1848 GATTTTTG GGCTAGCTACAACGA CAACAAGA 4871
229 UGACAAAA A UCCUCACA 1849 TGTGAGGA GGCTAGCTACAACGA TTTTGTCA 4872
235 AAAUCCUC A CAAUACCA 1850 TGGTATTG GGCTAGCTACAACGA GAGGATTT 4873
238 UCCUCACA A UACCACAG 1851 CTGTGGTA GGCTAGCTACAACGA TGTGAGGA 4874
240 CUCACAAU A CCACAGAG 1852 CTCTGTGG GGCTAGCTACAACGA ATTGTGAG 4875
243 ACAAUACC A CAGAGUCU 1853 AGACTCTG GGCTAGCTACAACGA GGTATTGT 4876
254 GAGUCUAG A CUCGUGGU 1854 ACCACGAG GGCTAGCTACAACGA CTAGACTC 4877
265 CGUGGUGG A CUUCUCUC 1855 GAGAGAAG GGCTAGCTACAACGA CCACCACG 4878
275 UUCUCUCA A UUUUCUAG 1856 CTAGAAAA GGCTAGCTACAACGA TGAGAGAA 4879
289 UAGGGGGA A CACCCGUG 1857 CACGGGTG GGCTAGCTACAACGA TCCCCCTA 4880
291 GGGGGAAC A CCCGUGUG 1858 CACACGGG GGCTAGCTACAACGA GTTCCCCC 4881
311 UGGCCAAA A UUCGCAGU 1859 ACTGCGAA GGCTAGCTACAACGA TTTGGCCA 4882
325 AGUCCCAA A UCUCCAGU 1860 ACTGGAGA GGCTAGCTACAACGA TTGGGACT 4883
335 CUCCAGUC A CUCACCAA 1861 TTGGTGAG GGCTAGCTACAACGA GACTGGAG 4884
339 AGUCACUC A CCAACCUG 1862 CAGGTTGG GGCTAGCTACAACGA GAGTGACT 4885
343 ACUCACCA A CCUGUUGU 1863 ACAACAGG GGCTAGCTACAACGA TGGTGAGT 4886
358 GUCCUCCA A UUUGUCCU 1864 AGGACAAA GGCTAGCTACAACGA TGGAGGAC 4887
371 UCCUGGUU A UCGCUGGA 1865 TCCAGCGA GGCTAGCTACAACGA AACCAGGA 4888
379 AUCGCUGG A UGUGUCUG 1866 CAGACACA GGCTAGCTACAACGA CCAGCGAT 4889
397 GGCGUUUU A UCAUCUUC 1867 GAAGATGA GGCTAGCTACAACGA AAAACGCC 4890
400 GUUUUAUC A UCUUCCUC 1868 GAGGAAGA GGCTAGCTACAACGA GATAAAAC 4891
412 UCCUCUGC A UCCUGCUG 1869 CAGCAGGA GGCTAGCTACAACGA GCAGAGGA 4892
423 CUGCUGCU A UGCCUCAU 1870 ATGAGGCA GGCTAGCTACAACGA AGCAGCAG 4893
430 UAUGCCUC A UCUUCUUG 1871 CAAGAAGA GGCTAGCTACAACGA GAGGCATA 4894
452 UCUUCUGG A CUAUCAAG 1872 CTTGATAG GGCTAGCTACAACGA CCAGAAGA 4895
455 UCUGGACU A UCAAGGUA 1873 TACCTTGA GGCTAGCTACAACGA AGTCCAGA 4896
463 AUCAAGGU A UGUUGCCC 1874 GGGCAACA GGCTAGCTACAACGA ACCTTGAT 4897
484 GUCCUCUA A UUCCAGGA 1875 TCCTGGAA GGCTAGCTACAACGA TAGAGGAC 4898
492 AUUCCAGG A UCAUCAAC 1876 GTTGATGA GGCTAGCTACAACGA CCTGGAAT 4899
495 CCAGGAUC A UCAACAAC 1877 GTTGTTGA GGCTAGCTACAACGA GATCCTGG 4900
499 GAUCAUCA A CAACCAGC 1878 GCTGGTTG GGCTAGCTACAACGA TGATGATC 4901
502 CAUCAACA A CCAGCACC 1879 GGTGCTGG GGCTAGCTACAACGA TGTTGATG 4902
513 AGCACCGG A CCAUGCAA 1880 TTGCATGG GGCTAGCTACAACGA CCGGTGCT 4903
516 ACCGGACC A UGCAAAAC 1881 GTTTTGCA GGCTAGCTACAACGA GGTCCGGT 4904
523 CAUGCAAA A CCUGCACA 1882 TGTGCAGG GGCTAGCTACAACGA TTTGCATG 4905
529 AAACCUGC A CAACUCCU 1883 AGGAGTTG GGCTAGCTACAACGA GCAGGTTT 4906
532 CCUGCACA A CUCCUGCU 1884 AGCAGGAG GGCTAGCTACAACGA TGTGCAGG 4907
547 CUCAAGGA A CCUCUAUG 1885 CATAGAGG GGCTAGCTACAACGA TCCTTGAG 4908
Taoie 4 i
553 GAACCUCU A UGUUUCCC 1886 GGGAAACA GGCTAGCTACAACGA AGAGGTTC 4909
564 UUUCCCUC A UGUUGCUG 1887 CAGCAACA GGCTAGCTACAACGA GAGGGAAA 4910
574 GUUGCUGU A CAAAACCU 1888 AGGTTTTG GGCTAGCTACAACGA ACAGCAAC 4911
579 UGUACAAA A CCUACGGA 1889 TCCGTAGG GGCTAGCTACAACGA TTTGTACA 4912
583 CAAAACCU A CGGACGGA 1890 TCCGTCCG GGCTAGCTACAACGA AGGTTTTG 4913
587 ACCUACGG A CGGAAACU 1891 AGTTTCCG GGCTAGCTACAACGA CCGTAGGT 4914
593 GGACGGAA A CUGCACCU 1892 AGGTGCAG GGCTAGCTACAACGA TTCCGTCC 4915
598 GAAACUGC A CCUGUAUU 1893 AATACAGG GGCTAGCTACAACGA GCAGTTTC 4916
604 GCACCUGU A UUCCCAUC 1894 GATGGGAA GGCTAGCTACAACGA ACAGGTGC 4917
610 GUAUUCCC A UCCCAUCA 1895 TGATGGGA GGCTAGCTACAACGA GGGAATAC 4918
615 CCCAUCCC A UCAUCUUG 1896 CAAGATGA GGCTAGCTACAACGA GGGATGGG 4919
618 AUCCCAUC A UCUUGGGC 1897 GCCCAAGA GGCTAGCTACAACGA GATGGGAT 4920
636 UUCGCAAA A UACCUAUG 1898 CATAGGTA GGCTAGCTACAACGA TTTGCGAA 4921
638 CGCAAAAU A CCUAUGGG 1899 CCCATAGG GGCTAGCTACAACGA ATTTTGCG 4922
642 AAAUACCU A UGGGAGUG 1900 CACTCCCA GGCTAGCTACAACGA AGGTATTT 4923
681 CUCAGUUU A CUAGUGCC 1901 GGCACTAG GGCTAGCTACAACGA AAACTGAG 4924
690 CUAGUGCC A UUUGUUCA 1902 TGAACAAA GGCTAGCTACAACGA GGCACTAG 4925
721 UUUCCCCC A CUGUCUGG 1903 CCAGACAG GGCTAGCTACAACGA GGGGGAAA 4926
739 UUUCAGUU A UAUGGAUG 1904 CATCCATA GGCTAGCTACAACGA AACTGAAA 4927
741 UCAGUUAU A UGGAUGAU 1905 ATCATCCA GGCTAGCTACAACGA ATAACTGA 4928
745 UUAUAUGG A UGAUGUGG 1906 CCACATCA GGCTAGCTACAACGA CCATATAA 4929
748 UAUGGAUG A UGUGGUUU 1907 AAACCACA GGCTAGCTACAACGA CATCCATA 4930
773 AAGUCUGU A CAACAUCU 1908 AGATGTTG GGCTAGCTACAACGA ACAGACTT 4931
776 UCUGUACA A CAUCUUGA 1909 TCAAGATG GGCTAGCTACAACGA TGTACAGA 4932
778 UGUACAAC A UCUUGAGU 1910 ACTCAAGA GGCTAGCTACAACGA GTTGTACA 4933
793 GUCCCUUU A UGCCGCUG 1911 CAGCGGCA GGCTAGCTACAACGA AAAGGGAC 4934
804 CCGCUGUU A CCAAUUUU 1912 AAAATTGG GGCTAGCTACAACGA AACAGCGG 4935
808 UGUUACCA A UUUUCUUU 1913 AAAGAAAA GGCTAGCTACAACGA TGGTAACA 4936
828 CUUUGGGU A UACAUUUA 1914 TAAATGTA GGCTAGCTACAACGA ACCCAAAG 4937
830 UUGGGUAU A CAUUUAAA 1915 TTTAAATG GGCTAGCTACAACGA ATACCCAA 4938
832 GGGUAUAC A UUUAAACC 1916 GGTTTAAA GGCTAGCTACAACGA GTATACCC 4939
838 ACAUUUAA A CCCUCACA 1917 TGTGAGGG GGCTAGCTACAACGA TTAAATGT 4940
844 AAACCCUC A CAAAACAA 1918 TTGTTTTG GGCTAGCTACAACGA GAGGGTTT 4941
849 CUCACAAA A CAAAAAGA 1919 TCTTTTTG GGCTAGCTACAACGA TTTGTGAG 4942
857 ACAAAAAG A UGGGGAUA 1920 TATCCCCA GGCTAGCTACAACGA CTTTTTGT 4943
863 AGAUGGGG A UAUUCCCU 1921 AGGGAATA GGCTAGCTACAACGA CCCCATCT 4944
865 AUGGGGAU A UUCCCUUA 1922 TAAGGGAA GGCTAGCTACAACGA ATCCCCAT 4945
874 UUCCCUUA A CUUCAUGG 1923 CCATGAAG GGCTAGCTACAACGA TAAGGGAA 4946
879 UUAACUUC A UGGGAUAU 1924 ATATCCCA GGCTAGCTACAACGA GAAGTTAA 4947
884 UUCAUGGG A UAUGUAAU 1925 ATTACATA GGCTAGCTACAACGA CCCATGAA 4948
886 CAUGGGAU A UGUAAUUG 1926 CAATTACA GGCTAGCTACAACGA ATCCCATG 4949
891 GAUAUGUA A UUGGGAGU 1927 ACTCCCAA GGCTAGCTACAACGA TACATATC 4950
906 GUUGGGGC A CAUUGCCA 1928 TGGCAATG GGCTAGCTACAACGA GCCCCAAC 4951
908 UGGGGCAC A UUGCCACA 1929 TGTGGCAA GGCTAGCTACAACGA GTGCCCCA 4952
914 ACAUUGCC A CAGGAACA 1930 TGTTCCTG GGCTAGCTACAACGA GGCAATGT 4953
920 CCACAGGA A CAUAUUGU 1931 ACAATATG GGCTAGCTACAACGA TCCTGTGG 4954
922 ACAGGAAC A UAUUGUAC 1932 GTACAATA GGCTAGCTACAACGA GTTCCTGT 4955
924 AGGAACAU A UUGUACAA 1933 TTGTACAA GGCTAGCTACAACGA ATGTTCCT 4956
929 CAUAUUGU A CAAAAAAU 1934 ATTTTTTG GGCTAGCTACAACGA ACAATATG 4957
936 UACAAAAA A UCAAAAUG 1935 CATTTTGA GGCTAGCTACAACGA TTTTTGTA 4958
942 AAAUCAAA A UGUGUUUU 1936 AAAACACA GGCTAGCTACAACGA TTTGATTT 4959
Taoie 4 i
956 UUUAGGAA A CUUCCUGU 1937 ACAGGAAG GGCTAGCTACAACGA TTCCTAAA 4960
967 UCCUGUAA A CAGGCCUA 1938 TAGGCCTG GGCTAGCTACAACGA TTACAGGA 4961
975 ACAGGCCU A UUGAUUGG 1939 CCAATCAA GGCTAGCTACAACGA AGGCCTGT 4962
979 GCCUAUUG A UUGGAAAG 1940 CTTTCCAA GGCTAGCTACAACGA CAATAGGC 4963
989 UGGAAAGU A UGUCAACG 1941 CGTTGACA GGCTAGCTACAACGA ACTTTCCA 4964
995 GUAUGUCA A CGAAUUGU 1942 ACAATTCG GGCTAGCTACAACGA TGACATAC 4965
999 GUCAACGA A UUGUGGGU 1943 ACCCACAA GGCTAGCTACAACGA TCGTTGAC 4966
1032 CCCCUUUC A CGCAAUGU 1944 ACATTGCG GGCTAGCTACAACGA GAAAGGGG 4967
1037 UUCACGCA A UGUGGAUA 1945 TATCCACA GGCTAGCTACAACGA TGCGTGAA 4968
1043 CAAUGUGG A UAUUCUGC 1946 GCAGAATA GGCTAGCTACAACGA CCACATTG 4969
1045 AUGUGGAU A UUCUGCUU 1947 AAGCAGAA GGCTAGCTACAACGA ATCCACAT 4970
1056 CUGCUUUA A UGCCUUUA 1948 TAAAGGCA GGCTAGCTACAACGA TAAAGCAG 4971
1064 AUGCCUUU A UAUGCAUG 1949 CATGCATA GGCTAGCTACAACGA AAAGGCAT 4972
1066 GCCUUUAU A UGCAUGCA 1950 TGCATGCA GGCTAGCTACAACGA ATAAAGGC 4973
1070 UUAUAUGC A UGCAUACA 1951 TGTATGCA GGCTAGCTACAACGA GCATATAA 4974
1074 AUGCAUGC A UACAAGCA 1952 TGCTTGTA GGCTAGCTACAACGA GCATGCAT 4975
1076 GCAUGCAU A CAAGCAAA 1953 TTTGCTTG GGCTAGCTACAACGA ATGCATGC 4976
1085 CAAGCAAA A CAGGCUUU 1954 AAAGCCTG GGCTAGCTACAACGA TTTGCTTG 4977
1095 AGGCUUUU A CUUUCUCG 1955 CGAGAAAG GGCTAGCTACAACGA AAAAGCCT 4978
1107 UCUCGCCA A CUUACAAG 1956 CTTGTAAG GGCTAGCTACAACGA TGGCGAGA 4979 llll GCCAACUU A CAAGGCCU 1957 AGGCCTTG GGCTAGCTACAACGA AAGTTGGC 4980
1130 CUAAGUAA A CAGUAUGU 1958 ACATACTG GGCTAGCTACAACGA TTACTTAG 4981
1135 UAAACAGU A UGUGAACC 1959 GGTTCACA GGCTAGCTACAACGA ACTGTTTA 4982
1141 GUAUGUGA A CCUUUACC 1960 GGTAAAGG GGCTAGCTACAACGA TCACATAC 4983
1147 GAACCUUU A CCCCGUUG 1961 CAACGGGG GGCTAGCTACAACGA AAAGGTTC 4984
1163 GCUCGGCA A CGGCCUGG 1962 CCAGGCCG GGCTAGCTACAACGA TGCCGAGC 4985
1175 CCUGGUCU A UGCCAAGU 1963 ACTTGGCA GGCTAGCTACAACGA AGACCAGG 4986
1192 GUUUGCUG A CGCAACCC 1964 GGGTTGCG GGCTAGCTACAACGA CAGCAAAC 4987
1197 CUGACGCA A CCCCCACU 1965 AGTGGGGG GGCTAGCTACAACGA TGCGTCAG 4988
1203 CAACCCCC A CUGGUUGG 1966 CCAACCAG GGCTAGCTACAACGA GGGGGTTG 4989
1221 GCUUGGCC A UAGGCCAU 1967 ATGGCCTA GGCTAGCTACAACGA GGCCAAGC 4990
1228 CAUAGGCC A UCAGCGCA 1968 TGCGCTGA GGCTAGCTACAACGA GGCCTATG 4991
1236 AUCAGCGC A UGCGUGGA 1969 TCCACGCA GGCTAGCTACAACGA GCGCTGAT 4992
1245 UGCGUGGA A CCUUUGUG 1970 CACAAAGG GGCTAGCTACAACGA TCCACGCA 4993
1266 CUCUGCCG A UCCAUACC 1971 GGTATGGA GGCTAGCTACAACGA CGGCAGAG 4994
1270 GCCGAUCC A UACCGCGG 1972 CCGCGGTA GGCTAGCTACAACGA GGATCGGC 4995
1272 CGAUCCAU A CCGCGGAA 1973 TTCCGCGG GGCTAGCTACAACGA ATGGATCG 4996
1280 ACCGCGGA A CUCCUAGC 1974 GCTAGGAG GGCTAGCTACAACGA TCCGCGGT 4997
1322 GGGGCAAA A CUCAUCGG 1975 CCGATGAG GGCTAGCTACAACGA TTTGCCCC 4998
1326 CAAAACUC A UCGGGACU 1976 AGTCCCGA GGCTAGCTACAACGA GAGTTTTG 4999
1332 UCAUCGGG A CUGACAAU 1977 ATTGTCAG GGCTAGCTACAACGA CCCGATGA 5000
1336 CGGGACUG A CAAUUCUG 1978 CAGAATTG GGCTAGCTACAACGA CAGTCCCG 5001
1339 GACUGACA A UUCUGUCG 1979 CGACAGAA GGCTAGCTACAACGA TGTCAGTC 5002
1361 UCCCGCAA A UAUACAUC 1980 GATGTATA GGCTAGCTACAACGA TTGCGGGA 5003
1363 CCGCAAAU A UACAUCAU 1981 ATGATGTA GGCTAGCTACAACGA ATTTGCGG 5004
1365 GCAAAUAU A CAUCAUUU 1982 AAATGATG GGCTAGCTACAACGA ATATTTGC 5005
1367 AAAUAUAC A UCAUUUCC 1983 GGAAATGA GGCTAGCTACAACGA GTATATTT 5006
1370 UAUACAUC A UUUCCAUG 1984 CATGGAAA GGCTAGCTACAACGA GATGTATA 5007
1376 UCAUUUCC A UGGCUGCU 1985 AGCAGCCA GGCTAGCTACAACGA GGAAATGA 5008
1399 UGCUGCCA A CUGGAUCC 1986 GGATCCAG GGCTAGCTACAACGA TGGCAGCA 5009
1404 CCAACUGG A UCCUACGC 1987 GCGTAGGA GGCTAGCTACAACGA CCAGTTGG 5010
Table 41
1409 UGGAUCCU A CGCGGGAC 1988 GTCCCGCG GGCTAGCTACAACGA AGGATCCA 5011
1416 UACGCGGG A CGUCCUUU 1989 AAAGGACG GGCTAGCTACAACGA CCCGCGTA 5012
1429 CUUUGUUU A CGUCCCGU 1990 ACGGGACG GGCTAGCTACAACGA AAACAAAG 5013
1447 GGCGCUGA A UCCCGCGG 1991 CCGCGGGA GGCTAGCTACAACGA TCAGCGCC 5014
1456 UCCCGCGG A CGACCCCU 1992 AGGGGTCG GGCTAGCTACAACGA CCGCGGGA 5015
1459 CGCGGACG A CCCCUCCC 1993 GGGAGGGG GGCTAGCTACAACGA CGTCCGCG 5016
1486 GGGGCUCU A CCGCCCGC 1994 GCGGGCGG GGCTAGCTACAACGA AGAGCCCC 5017
1505 CUCCGCCU A UUGUACCG 1995 CGGTACAA GGCTAGCTACAACGA AGGCGGAG 5018
1510 CCUAUUGU A CCGACCGU 1996 ACGGTCGG GGCTAGCTACAACGA ACAATAGG 5019
1514 UUGUACCG A CCGUCCAC 1997 GTGGACGG GGCTAGCTACAACGA CGGTACAA 5020
1521 GACCGUCC A CGGGGCGC 1998 GCGCCCCG GGCTAGCTACAACGA GGACGGTC 5021
1530 CGGGGCGC A CCUCUCUU 1999 AAGAGAGG GGCTAGCTACAACGA GCGCCCCG 5022
1540 CUCUCUUU A CGCGGACU 2000 AGTCCGCG GGCTAGCTACAACGA AAAGAGAG 5023
1546 UUACGCGG A CUCCCCGU 2001 ACGGGGAG GGCTAGCTACAACGA CCGCGTAA 5024
1567 GCCUUCUC A UCUGCCGG 2002 CCGGCAGA GGCTAGCTACAACGA GAGAAGGC 5025
1576 UCUGCCGG A CCGUGUGC 2003 GCACACGG GGCTAGCTACAACGA CCGGCAGA 5026
1585 CCGUGUGC A CUUCGCUU 2004 AAGCGAAG GGCTAGCTACAACGA GCACACGG 5027
1595 UUCGCUUC A CCUCUGCA 2005 TGCAGAGG GGCTAGCTACAACGA GAAGCGAA 5028
1603 ACCUCUGC A CGUCGCAU 2006 ATGCGACG GGCTAGCTACAACGA GCAGAGGT 5029
1610 CACGUCGC A UGGAGACC 2007 GGTCTCCA GGCTAGCTACAACGA GCGACGTG 5030
1616 GCAUGGAG A CCACCGUG 2008 CACGGTGG GGCTAGCTACAACGA CTCCATGC 5031
1619 UGGAGACC A CCGUGAAC 2009 GTTCACGG GGCTAGCTACAACGA GGTCTCCA 5032
1626 CACCGUGA A CGCCCACA 2010 TGTGGGCG GGCTAGCTACAACGA TCACGGTG 5033
1638 CCACAGGA A CCUGCCCA 2011 TGGGCAGG GGCTAGCTACAACGA TCCTGTGG 5034
1656 GGUCUUGC A UAAGAGGA 2012 TCCTCTTA GGCTAGCTACAACGA GCAAGACC 5035
1664 AUAAGAGG A CUCUUGGA 2013 TCCAAGAG GGCTAGCTACAACGA CCTCTTAT 5036
1672 ACUCUUGG A CUUUCAGC 2014 GCTGAAAG GGCTAGCTACAACGA CCAAGAGT 5037
1682 UUUCAGCA A UGUCAACG 2015 CGTTGACA GGCTAGCTACAACGA TGCTGAAA 5038
1688 CAAUGUCA A CGACCGAC 2016 GTCGGTCG GGCTAGCTACAACGA TGACATTG 5039
1691 UGUCAACG A CCGACCUU 2017 AAGGTCGG GGCTAGCTACAACGA CGTTGACA 5040
1695 AACGACCG A CCUUGAGG 2018 CCTCAAGG GGCTAGCTACAACGA CGGTCGTT 5041
1705 CUUGAGGC A UACUUCAA 2019 TTGAAGTA GGCTAGCTACAACGA GCCTCAAG 5042
1707 UGAGGCAU A CUUCAAAG 2020 CTTTGAAG GGCTAGCTACAACGA ATGCCTCA 5043
1716 CUUCAAAG A CUGUGUGU 2021 ACACACAG GGCTAGCTACAACGA CTTTGAAG 5044
1728 UGUGUUUA A UGAGUGGG 2022 CCCACTCA GGCTAGCTACAACGA TAAACACA 5045
1774 GUCUUUGU A CUAGGAGG 2023 CCTCCTAG GGCTAGCTACAACGA ACAAAGAC 5046
1791 CUGUAGGC A UAAAUUGG 2024 CCAATTTA GGCTAGCTACAACGA GCCTACAG 5047
1795 AGGCAUAA A UUGGUGUG 2025 CACACCAA GGCTAGCTACAACGA TTATGCCT 5048
1807 GUGUGUUC A CCAGCACC 2026 GGTGCTGG GGCTAGCTACAACGA GAACACAC 5049
1813 UCACCAGC A CCAUGCAA 2027 TTGCATGG GGCTAGCTACAACGA GCTGGTGA 5050
1816 CCAGCACC A UGCAACUU 2028 AAGTTGCA GGCTAGCTACAACGA GGTGCTGG 5051
1821 ACCAUGCA A CUUUUUCA 2029 TGAAAAAG GGCTAGCTACAACGA TGCATGGT 5052
1829 ACUUUUUC A CCUCUGCC 2030 GGCAGAGG GGCTAGCTACAACGA GAAAAAGT 5053
1840 UCUGCCUA A UCAUCUCA 2031 TGAGATGA GGCTAGCTACAACGA TAGGCAGA 5054
1843 GCCUAAUC A UCUCAUGU 2032 ACATGAGA GGCTAGCTACAACGA GATTAGGC 5055
1848 AUCAUCUC A UGUUCAUG 2033 CATGAACA GGCTAGCTACAACGA GAGATGAT 5056
1854 UCAUGUUC A UGUCCUAC 2034 GTAGGACA GGCTAGCTACAACGA GAACATGA 5057
1861 CAUGUCCU A CUGUUCAA 2035 TTGAACAG GGCTAGCTACAACGA AGGACATG 5058
1903 UUUGGGGC A UGGACAUU 2036 AATGTCCA GGCTAGCTACAACGA GCCCCAAA 5059
1907 GGGCAUGG A CAUUGACC 2037 GGTCAATG GGCTAGCTACAACGA CCATGCCC 5060
1909 GCAUGGAC A UUGACCCG 2038 CGGGTCAA GGCTAGCTACAACGA GTCCATGC 5061
TaDie 4 1
1913 GGACAUUG A CCCGUAUA 2039 TATACGGG GGCTAGCTACAACGA CAATGTCC 5062
1919 UGACCCGU A UAAAGAAU 2040 ATTCTTTA GGCTAGCTACAACGA ACGGGTCA 5063
1926 UAUAAAGA A UUUGGAGC 2041 GCTCCAAA GGCTAGCTACAACGA TCTTTATA 5064
1947 GUGGAGUU A CUCUCUUU 2042 AAAGAGAG GGCTAGCTACAACGA AACTCCAC 5065
1967 GCCUUCUG A CUUCUUUC 2043 GAAAGAAG GGCTAGCTACAACGA CAGAAGGC 5066
1981 UUCCUUCU A UUCGAGAU 2044 ATCTCGAA GGCTAGCTACAACGA AGAAGGAA 5067
1988 UAUUCGAG A UCUCCUCG 2045 CGAGGAGA GGCTAGCTACAACGA CTCGAATA 5068
1997 UCUCCUCG A CACCGCCU 2046 AGGCGGTG GGCTAGCTACAACGA CGAGGAGA 5069
1999 UCCUCGAC A CCGCCUCU 2047 AGAGGCGG GGCTAGCTACAACGA GTCGAGGA 5070
2015 UGCUCUGU A UCGGGGGG 2048 CCCCCCGA GGCTAGCTACAACGA ACAGAGCA 5071
2040 UCUCCGGA A CAUUGUUC 2049 GAACAATG GGCTAGCTACAACGA TCCGGAGA 5072
2042 UCCGGAAC A UUGUUCAC 2050 GTGAACAA GGCTAGCTACAACGA GTTCCGGA 5073
2049 CAUUGUUC A CCUCACCA 2051 TGGTGAGG GGCTAGCTACAACGA GAACAATG 5074
2054 UUCACCUC A CCAUACGG 2052 CCGTATGG GGCTAGCTACAACGA GAGGTGAA 5075
2057 ACCUCACC A UACGGCAC 2053 GTGCCGTA GGCTAGCTACAACGA GGTGAGGT 5076
2059 CUCACCAU A CGGCACUC 2054 GAGTGCCG GGCTAGCTACAACGA ATGGTGAG 5077
2064 CAUACGGC A CUCAGGCA 2055 TGCCTGAG GGCTAGCTACAACGA GCCGTATG 5078
2077 GGCAAGCU A UUCUGUGU 2056 ACACAGAA GGCTAGCTACAACGA AGCTTGCC 5079
2098 GUGAGUUG A UGAAUCUA 2057 TAGATTCA GGCTAGCTACAACGA CAACTCAC 5080
2102 GUUGAUGA A UCUAGCCA 2058 TGGCTAGA GGCTAGCTACAACGA TCATCAAC 5081
2110 AUCUAGCC A CCUGGGUG 2059 CACCCAGG GGCTAGCTACAACGA GGCTAGAT 5082
2126 GGGAAGUA A UUUGGAAG 2060 CTTCCAAA GGCTAGCTACAACGA TACTTCCC 5083
2135 UUUGGAAG A UCCAGCAU 2061 ATGCTGGA GGCTAGCTACAACGA CTTCCAAA 5084
2142 GAUCCAGC A UCCAGGGA 2062 TCCCTGGA GGCTAGCTACAACGA GCTGGATC 5085
2151 UCCAGGGA A UUAGUAGU 2063 ACTACTAA GGCTAGCTACAACGA TCCCTGGA 5086
2165 AGUCAGCU A UGUCAACG 2064 CGTTGACA GGCTAGCTACAACGA AGCTGACT 5087
2171 CUAUGUCA A CGUUAAUA 2065 TATTAACG GGCTAGCTACAACGA TGACATAG 5088
2177 CAACGUUA A UAUGGGCC 2066 GGCCCATA GGCTAGCTACAACGA TAACGTTG 5089
2179 ACGUUAAU A UGGGCCUA 2067 TAGGCCCA GGCTAGCTACAACGA ATTAACGT 5090
2191 GCCUAAAA A UCAGACAA 2068 TTGTCTGA GGCTAGCTACAACGA TTTTAGGC 5091
2196 AAAAUCAG A CAACUAUU 2069 AATAGTTG GGCTAGCTACAACGA CTGATTTT 5092
2199 AUCAGACA A CUAUUGUG 2070 CACAATAG GGCTAGCTACAACGA TGTCTGAT 5093
2202 AGACAACU A UUGUGGUU 2071 AACCACAA GGCTAGCTACAACGA AGTTGTCT 5094
2213 GUGGUUUC A CAUUUCCU 2072 AGGAAATG GGCTAGCTACAACGA GAAACCAC 5095
2215 GGUUUCAC A UUUCCUGU 2073 ACAGGAAA GGCTAGCTACAACGA GTGAAACC 5096
2227 CCUGUCUU A CUUUUGGG 2074 CCCAAAAG GGCTAGCTACAACGA AAGACAGG 5097
2242 GGCGAGAA A CUGUUCUU 2075 AAGAACAG GGCTAGCTACAACGA TTCTCGCC 5098
2253 GUUCUUGA A UAUUUGGU 2076 ACCAAATA GGCTAGCTACAACGA TCAAGAAC 5099
2255 UCUUGAAU A UUUGGUGU 2077 ACACCAAA GGCTAGCTACAACGA ATTCAAGA 5100
2278 GAGUGUGG A UUCGCACU 2078 AGTGCGAA GGCTAGCTACAACGA CCACACTC 5101
2284 GGAUUCGC A CUCCUCCU 2079 AGGAGGAG GGCTAGCTACAACGA GCGAATCC 5102
2295 CCUCCUGC A UAUAGACC 2080 GGTCTATA GGCTAGCTACAACGA GCAGGAGG 5103
2297 UCCUGCAU A UAGACCAC 2081 GTGGTCTA GGCTAGCTACAACGA ATGCAGGA 5104
2301 GCAUAUAG A CCACCAAA 2082 TTTGGTGG GGCTAGCTACAACGA CTATATGC 5105
2304 UAUAGACC A CCAAAUGC 2083 GCATTTGG GGCTAGCTACAACGA GGTCTATA 5106
2309 ACCACCAA A UGCCCCUA 2084 TAGGGGCA GGCTAGCTACAACGA TTGGTGGT 5107
2317 AUGCCCCU A UCUUAUCA 2085 TGATAAGA GGCTAGCTACAACGA AGGGGCAT 5108
2322 CCUAUCUU A UCAACACU 2086 AGTGTTGA GGCTAGCTACAACGA AAGATAGG 5109
2326 UCUUAUCA A CACUUCCG 2087 CGGAAGTG GGCTAGCTACAACGA TGATAAGA 5110
2328 UUAUCAAC A CUUCCGGA 2088 TCCGGAAG GGCTAGCTACAACGA GTTGATAA 5111
2338 UUCCGGAA A CUACUGUU 2089 AACAGTAG GGCTAGCTACAACGA TTCCGGAA 5112
TaDie 4 i
2341 CGGAAACU A CUGUUGUU 2090 AACAACAG GGCTAGCTACAACGA AGTTTCCG 5113
2352 GUUGUUAG A CGAAGAGG 2091 CCTCTTCG GGCTAGCTACAACGA CTAACAAC 5114
2380 GAAGAAGA A CUCCCUCG 2092 CGAGGGAG GGCTAGCTACAACGA TCTTCTTC 5115
2397 CCUCGCAG A CGAAGGUC 2093 GACCTTCG GGCTAGCTACAACGA CTGCGAGG 5116
2409 AGGUCUCA A UCGCCGCG 2094 CGCGGCGA GGCTAGCTACAACGA TGAGACCT 5117
2427 CGCAGAAG A UCUCAAUC 2095 GATTGAGA GGCTAGCTACAACGA CTTCTGCG 5118
2433 AGAUCUCA A UCUCGGGA 2096 TCCCGAGA GGCTAGCTACAACGA TGAGATCT 5119
2442 UCUCGGGA A UCUCAAUG 2097 CATTGAGA GGCTAGCTACAACGA TCCCGAGA 5120
2448 GAAUCUCA A UGUUAGUA 2098 TACTAACA GGCTAGCTACAACGA TGAGATTC 5121
2456 AUGUUAGU A UUCCUUGG 2099 CCAAGGAA GGCTAGCTACAACGA ACTAACAT 5122
2465 UUCCUUGG A CACAUAAG 2100 CTTATGTG GGCTAGCTACAACGA CCAAGGAA 5123
2467 CCUUGGAC A CAUAAGGU 2101 ACCTTATG GGCTAGCTACAACGA GTCCAAGG 5124
2469 UUGGACAC A UAAGGUGG 2102 CCACCTTA GGCTAGCTACAACGA GTGTCCAA 5125
2481 GGUGGGAA A CUUUACGG 2103 CCGTAAAG GGCTAGCTACAACGA TTCCCACC 5126
2486 GAAACUUU A CGGGGCUU 2104 AAGCCCCG GGCTAGCTACAACGA AAAGTTTC 5127
2496 GGGGCUUU A UUCUUCUA 2105 TAGAAGAA GGCTAGCTACAACGA AAAGCCCC 5128
2504 AUUCUUCU A CGGUACCU 2106 AGGTACCG GGCTAGCTACAACGA AGAAGAAT 5129
2509 UCUACGGU A CCUUGCUU 2107 AAGCAAGG GGCTAGCTACAACGA ACCGTAGA 5130
2520 UUGCUUUA A UCCUAAAU 2108 ATTTAGGA GGCTAGCTACAACGA TAAAGCAA 5131
2527 AAUCCUAA A UGGCAAAC 2109 GTTTGCCA GGCTAGCTACAACGA TTAGGATT 5132
2534 AAUGGCAA A CUCCUUCU 2110 AGAAGGAG GGCTAGCTACAACGA TTGCCATT 5133
2550 UUUUCCUG A CAUUCAUU 2111 AATGAATG GGCTAGCTACAACGA CAGGAAAA 5134
2552 UUCCUGAC A UUCAUUUG 2112 CAAATGAA GGCTAGCTACAACGA GTCAGGAA 5135
2556 UGACAUUC A UUUGCAGG 2113 CCTGCAAA GGCTAGCTACAACGA GAATGTCA 5136
2568 GCAGGAGG A CAUUGUUG 2114 CAACAATG GGCTAGCTACAACGA CCTCCTGC 5137
2570 AGGAGGAC A UUGUUGAU 2115 ATCAACAA GGCTAGCTACAACGA GTCCTCCT 5138
2577 CAUUGUUG A UAGAUGUA 2116 TACATCTA GGCTAGCTACAACGA CAACAATG 5139
2581 GUUGAUAG A UGUAAGCA 2117 TGCTTACA GGCTAGCTACAACGA CTATCAAC 5140
2590 UGUAAGCA A UUUGUGGG 2118 CCCACAAA GGCTAGCTACAACGA TGCTTACA 5141
2606 GGCCCCUU A CAGUAAAU 2119 ATTTACTG GGCTAGCTACAACGA AAGGGGCC 5142
2613 UACAGUAA A UGAAAACA 2120 TGTTTTCA GGCTAGCTACAACGA TTACTGTA 5143
2619 AAAUGAAA A CAGGAGAC 2121 GTCTCCTG GGCTAGCTACAACGA TTTCATTT 5144
2626 AACAGGAG A CUUAAAUU 2122 AATTTAAG GGCTAGCTACAACGA CTCCTGTT 5145
2632 AGACUUAA A UUAACUAU 2123 ATAGTTAA GGCTAGCTACAACGA TTAAGTCT 5146
2636 UUAAAUUA A CUAUGCCU 2124 AGGCATAG GGCTAGCTACAACGA TAATTTAA 5147
2639 AAUUAACU A UGCCUGCU 2125 AGCAGGCA GGCTAGCTACAACGA AGTTAATT 5148
2655 UAGGUUUU A UCCCAAUG 2126 CATTGGGA GGCTAGCTACAACGA AAAACCTA 5149
2661 UUAUCCCA A UGUUACUA 2127 TAGTAACA GGCTAGCTACAACGA TGGGATAA 5150
2666 CCAAUGUU A CUAAAUAU 2128 ATATTTAG GGCTAGCTACAACGA AACATTGG 5151
2671 GUUACUAA A UAUUUGCC 2129 GGCAAATA GGCTAGCTACAACGA TTAGTAAC 5152
2673 UACUAAAU A UUUGCCCU 2130 AGGGCAAA GGCTAGCTACAACGA ATTTAGTA 5153
2685 GCCCUUAG A UAAAGGGA 2131 TCCCTTTA GGCTAGCTACAACGA CTAAGGGC 5154
2693 AUAAAGGG A UCAAACCG 2132 CGGTTTGA GGCTAGCTACAACGA CCCTTTAT 5155
2698 GGGAUCAA A CCGUAUUA 2133 TAATACGG GGCTAGCTACAACGA TTGATCCC 5156
2703 CAAACCGU A UUAUCCAG 2134 CTGGATAA GGCTAGCTACAACGA ACGGTTTG 5157
2706 ACCGUAUU A UCCAGAGU 2135 ACTCTGGA GGCTAGCTACAACGA AATACGGT 5158
2715 UCCAGAGU A UGUAGUUA 2136 TAACTACA GGCTAGCTACAACGA ACTCTGGA 5159
2724 UGUAGUUA A UCAUUACU 2137 AGTAATGA GGCTAGCTACAACGA TAACTACA 5160
2727 AGUUAAUC A UUACUUCC 2138 GGAAGTAA GGCTAGCTACAACGA GATTAACT 5161
2730 UAAUCAUU A CUUCCAGA 2139 TCTGGAAG GGCTAGCTACAACGA AATGATTA 5162
2738 ACUUCCAG A CGCGACAU 2140 ATGTCGCG GGCTAGCTACAACGA CTGGAAGT 5163
Taoie 4 1
2743 CAGACGCG A CAUUAUUU 2141 AAATAATG GGCTAGCTACAACGA CGCGTCTG 5164
2745 GACGCGAC A UUAUUUAC 2142 GTAAATAA GGCTAGCTACAACGA GTCGCGTC 5165
2748 GCGACAUU A UUUACACA 2143 TGTGTAAA GGCTAGCTACAACGA AATGTCGC 5166
2752 CAUUAUUU A CACACUCU 2144 AGAGTGTG GGCTAGCTACAACGA AAATAATG 5167
2754 UUAUUUAC A CACUCUUU 2145 AAAGAGTG GGCTAGCTACAACGA GTAAATAA 5168
2756 AUUUACAC A CUCUUUGG 2146 CCAAAGAG GGCTAGCTACAACGA GTGTAAAT 5169
2774 AGGCGGGG A UCUUAUAU 2147 ATATAAGA GGCTAGCTACAACGA CCCCGCCT 5170
2779 GGGAUCUU A UAUAAAAG 2148 CTTTTATA GGCTAGCTACAACGA AAGATCCC 5171
2781 GAUCUUAU A UAAAAGAG 2149 CTCTTTTA GGCTAGCTACAACGA ATAAGATC 5172
2795 GAGAGUCC A CACGUAGC 2150 GCTACGTG GGCTAGCTACAACGA GGACTCTC 5173
2797 GAGUCCAC A CGUAGCGC 2151 GCGCTACG GGCTAGCTACAACGA GTGGACTC 5174
2809 AGCGCCUC A UUUUGCGG 2152 CCGCAAAA GGCTAGCTACAACGA GAGGCGCT 5175
2821 UGCGGGUC A CCAUAUUC 2153 GAATATGG GGCTAGCTACAACGA GACCCGCA 5176
2824 GGGUCACC A UAUUCUUG 2154 CAAGAATA GGCTAGCTACAACGA GGTGACCC 5177
2826 GUCACCAU A UUCUUGGG 2155 CCCAAGAA GGCTAGCTACAACGA ATGGTGAC 5178
2836 UCUUGGGA A CAAGAUCU 2156 AGATCTTG GGCTAGCTACAACGA TCCCAAGA 5179
2841 GGAACAAG A UCUACAGC 2157 GCTGTAGA GGCTAGCTACAACGA CTTGTTCC 5180
2845 CAAGAUCU A CAGCAUGG 2158 CCATGCTG GGCTAGCTACAACGA AGATCTTG 5181
2850 UCUACAGC A UGGGAGGU 2159 ACCTCCCA GGCTAGCTACAACGA GCTGTAGA 5182
2870 UCUUCCAA A CCUCGAAA 2160 TTTCGAGG GGCTAGCTACAACGA TTGGAAGA 5183
2883 GAAAAGGC A UGGGGACA 2161 TGTCCCCA GGCTAGCTACAACGA GCCTTTTC 5184
2889 GCAUGGGG A CAAAUCUU 2162 AAGATTTG GGCTAGCTACAACGA CCCCATGC 5185
2893 GGGGACAA A UCUUUCUG 2163 CAGAAAGA GGCTAGCTACAACGA TTGTCCCC 5186
2908 UGUCCCCA A UCCCCUGG 2164 CCAGGGGA GGCTAGCTACAACGA TGGGGACA 5187
2918 CCCCUGGG A UUCUUCCC 2165 GGGAAGAA GGCTAGCTACAACGA CCCAGGGG 5188
2929 CUUCCCCG A UCAUCAGU 2166 ACTGATGA GGCTAGCTACAACGA CGGGGAAG 5189
2932 CCCCGAUC A UCAGUUGG 2167 CCAACTGA GGCTAGCTACAACGA GATCGGGG 5190
2941 UCAGUUGG A CCCUGCAU 2168 ATGCAGGG GGCTAGCTACAACGA CCAACTGA 5191
2948 GACCCUGC A UUCAAAGC 2169 GCTTTGAA GGCTAGCTACAACGA GCAGGGTC 5192
2959 CAAAGCCA A CUCAGUAA 2170 TTACTGAG GGCTAGCTACAACGA TGGCTTTG 5193
2968 CUCAGUAA A UCCAGAUU 2171 AATCTGGA GGCTAGCTACAACGA TTACTGAG 5194
2974 AAAUCCAG A UUGGGACC 2172 GGTCCCAA GGCTAGCTACAACGA CTGGATTT 5195
2980 AGAUUGGG A CCUCAACC 2173 GGTTGAGG GGCTAGCTACAACGA CCCAATCT 5196
2986 GGACCUCA A CCCGCACA 2174 TGTGCGGG GGCTAGCTACAACGA TGAGGTCC 5197
2998 GCACAAGG A CAACUGGC 2175 GCCAGTTG GGCTAGCTACAACGA CCTTGTGC 5198
3001 CAAGGACA A CUGGCCGG 2176 CCGGCCAG GGCTAGCTACAACGA TGTCCTTG 5199
3010 CUGGCCGG A CGCCAACA 2177 TGTTGGCG GGCTAGCTACAACGA CCGGCCAG 5200
3016 GGACGCCA A CAAGGUGG 2178 CCACCTTG GGCTAGCTACAACGA TGGCGTCC 5201
3035 GUGGGAGC A UUCGGGCC 2179 GGCCCGAA GGCTAGCTACAACGA GCTCCCAC 5202
3051 CAGGGUUC A CCCCUCCC 2180 GGGAGGGG GGCTAGCTACAACGA GAACCCTG 5203
3061 CCCUCCCC A UGGGGGAC 2181 GTCCCCCA GGCTAGCTACAACGA GGGGAGGG 5204
3068 CAUGGGGG A CUGUUGGG 2182 CCCAACAG GGCTAGCTACAACGA CCCCCATG 5205
3088 GAGCCCUC A CGCUCAGG 2183 CCTGAGCG GGCTAGCTACAACGA GAGGGCTC 5206
3101 CAGGGCCU A CUCACAAC 2184 GTTGTGAG GGCTAGCTACAACGA AGGCCCTG 5207
3105 GCCUACUC A CAACUGUG 2185 CACAGTTG GGCTAGCTACAACGA GAGTAGGC 5208
3108 UACUCACA A CUGUGCCA 2186 TGGCACAG GGCTAGCTACAACGA TGTGAGTA 5209
3138 CUGCCUCC A CCAAUCGG 2187 CCGATTGG GGCTAGCTACAACGA GGAGGCAG 5210
3142 CUCCACCA A UCGGCAGU 2188 ACTGCCGA GGCTAGCTACAACGA TGGTGGAG 5211
3165 GGCAGCCU A CUCCCUUA 2189 TAAGGGAG GGCTAGCTACAACGA AGGCTGCC 5212
3173 ACUCCCUU A UCUCCACC 2190 GGTGGAGA GGCTAGCTACAACGA AAGGGAGT 5213
3179 UUAUCUCC A CCUCUAAG 2191 CTTAGAGG GGCTAGCTACAACGA GGAGATAA 5214
Taσie 4
Input Sequence = AF100308. Cut Site = YG/M or UG/U. Stem Length = 8 . Core Sequence = GGCTAGCTACAACGA AF100308 (Hepatitis B virus strain 2-18, 3215 bp)
Table 42
Table 42: Human HBV Amberzyme Ribozyme and Substrate Sequence
Pos Substrate Seq ID Ribozyme Rz Seq ID
61 ACUUUCCU G CUGGUGGC 1448 GCCACCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGAAAGU 5219
87 GGAACAGU G AGCCCUGC 1449 GCAGGGCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACUGUUCC 5220
94 UGAGCCCU G CUCAGAAU 1450 AUUCUGAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGGCUCA 5221
112 CUGUCUCU G CCAUAUCG 1451 CGAUAUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGAGACAG 5222
132 AUCUUAUC G AAGACUGG 1452 CCAGUCUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAUAAGAU 5223
153 CCUGUACC G AACAUGGA 1453 UCCAUGUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGUACAGG 5224
169 AGAACAUC G CAUCAGGA 1454 UCCUGAUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAUGUUCU 5225
192 GGACCCCU G CUCGUGUU 1455 AACACGAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGGGUCC 5226
222 UUCUUGUU G ACAAAAAU 1456 AUUUUUGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AACAAGAA 5227
315 CAAAAUUC G CAGUCCCA 1457 UGGGACUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAAUUUUG 5228
374 UGGUUAUC G CUGGAUGU 1458 ACAUCCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAUAACCA 5229
387 AUGUGUCU G CGGCGUUU 1459 AAACGCCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGACACAU 5230
410 CUUCCUCU G CAUCCUGC 1460 GCAGGAUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAGGAAG 5231
417 UGCAUCCU G CUGCUAUG 1461 CAUAGCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGAUGCA 5232
420 AUCCUGCU G CUAUGCCU 1462 AGGCAUAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCAGGAU 5233
425 GCUGCUAU G CCUCAUCU 1463 AGAUGAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUAGCAGC 5234
468 GGUAUGUU G CCCGUUUG 1464 CAAACGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AACAUACC 5235
518 CGGACCAU G CAAAACCU 1465 AGGUUUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGGUCCG 5236
527 CAAAACCU G CACAACUC 1466 GAGUUGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGUUUUG 5237
538 CAACUCCU G CUCAAGGA 1467 UCCUUGAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGAGUUG 5238
569 CUCAUGUU G CUGUACAA 1468 UUGUACAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AACAUGAG 5239
596 CGGAAACU G CACCUGUA 1469 UACAGGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUUUCCG 5240
631 GGGCUUUC G CAAAAUAC 1470 GUAUUUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GAAAGCCC 5241
687 UUACUAGU G CCAUUUGU 1471 ACAAAUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACUAGUAA 5242
747 AUAUGGAU G AUGUGGUU 1472 AACCACAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCCAUAU 5243
783 AACAUCUU G AGUCCCUU 1473 AAGGGACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGAUGUU 5244
795 CCCUUUAU G CCGCUGUU 1474 AACAGCGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUAAAGGG 5245
798 UUUAUGCC G CUGUUACC 1475 GGUAACAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGCAUAAA 5246
911 GGCACAUU G CCACAGGA 1476 UCCUGUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUGUGCC 5247
978 GGCCUAUU G AUUGGAAA 1477 UUUCCAAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUAGGCC 5248
Table 42
997 AUGUCAAC G AAUUGUGG 1478 CCACAAUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUUGACAU 5249
1020 UGGGGUUU G CCGCCCCU 1479 AGGGGCGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAACCCCA 5250
1023 GGUUUGCC G CCCCUUUC 1480 GAAAGGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGCAAACC 5251
1034 CCUUUCAC G CAAUGUGG 1481 CCACAUUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUGAAAGG 5252
1050 GAUAUUCU G CUUUAAUG 1482 CAUUAAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGAAUAUC 5253
1058 GCUUUAAU G CCUUUAUA 1483 UAUAAAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUUAAAGC 5254
1068 CUUUAUAU G CAUGCAUA 1484 UAUGCAUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUAUAAAG 5255
1072 AUAUGCAU G CAUACAAG 1485 CUUGUAUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUGCAUAU 5256
1103 ACUUUCUC G CCAACUUA 1486 UAAGUUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAGAAAGU 5257
1139 CAGUAUGU G AACCUUUA 1487 UAAAGGUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACAUACUG 5258
1155 ACCCCGUU G CUCGGCAA 1488 UUGCCGAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AACGGGGU 5259
1177 UGGUCUAU G CCAAGUGU 1489 ACACUUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUAGACCA 5260
1188 AAGUGUUU G CUGACGCA 1490 UGCGUCAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAACACUU 5261
1191 UGUUUGCU G ACGCAACC 1491 GGUUGCGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCAAACA 5262
1194 UUGCUGAC G CAACCCCC 1492 GGGGGUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUCAGCAA 5263
1234 CCAUCAGC G CAUGCGUG 1493 CACGCAUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCUGAUGG 5264
1238 CAGCGCAU G CGUGGAAC 1494 GUUCCACG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGCGCUG 5265
1262 UCUCCUCU G CCGAUCCA 1495 UGGAUCGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAGGAGA 5266
1265 CCUCUGCC G AUCCAUAC 1496 GUAUGGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGCAGAGG 5267
1275 UCCAUACC G CGGAACUC 1497 GAGUUCCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGUAUGGA 5268
1290 UCCUAGCC G CUUGUUUU 1498 AAAACAAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGCUAGGA 5269
1299 CUUGUUUU G CUCGCAGC 1499 GCUGCGAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAACAAG 5270
1303 UUUUGCUC G CAGCAGGU 1500 ACCUGCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GAGCAAAA 5271
1335 UCGGGACU G AGAAUUCU 1501 AGAAUUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUCCCGA 5272
1349 UCUGUCGU G CUCUCCCG 1502 CGGGAGAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACGACAGA 5273
1357 GCUCUCCC G CAAAUAUA 1503 UAUAUUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGAGAGC 5274
1382 CCAUGGCU G CUAGGCUG 1504 CAGCCUAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCCAUGG 5275
1392 UAGGCUGU G CUGCCAAC 1505 GUUGGCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAGCCUA 5276
1395 GCUGUGCU G CCAACUGG 1506 CCAGUUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCACAGC 5277
1411 GAUCCUAC G CGGGACGU 1507 ACGUCCCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUAGGAUC 5278
1442 CCGUCGGC G CUGAAUCC 1508 GGAUUCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCCGACGG 5279
1445 UCGGCGCU G AAUCCCGC 1509 GCGGGAUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCGCCGA 5280
1452 UGAAUCCC G CGGACGAC 1510 GUCGUCCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGAUUCA 5281
1458 CCGCGGAC G ACCCCUCC 1511 GGAGGGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUCCGCGG 5282
Table 42
1474 CCGGGGCC G CUUGGGGC 1512 GCCCCAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGCCCCGG 5283
1489 GCUCUACC G CCCGCUUC 1513 GAAGCGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGUAGAGC 5284
1493 UACCGCCC G CUUCUCCG 1514 CGGAGAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGGCGGUA 5285
1501 GCUUCUCC G CCUAUUGU 1515 ACAAUAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGAGAAGC 5286
1513 AUUGUACC G ACCGUCCA 1516 UGGACGGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGUACAAU 5287
1528 CACGGGGC G CACCUCUC 1517 GAGAGGUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCCCCGUG 5288
1542 CUCUUUAC G CGGACUCC 1518 GGAGUCCG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUAAAGAG 5289
1559 CCGUCUGU G CCUUCUCA 1519 UGAGAAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACAGACGG 5290
1571 UCUCAUCU G CCGGACCG 1520 CGGUCCGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGAUGAGA 5291
1583 GACCGUGU G CACUUCGC 1521 GCGAAGUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACACGGUC 5292
1590 UGCACUUC G CUUCACCU 1522 AGGUGAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAAGUGCA 5293
1601 UCACCUCU G CACGUCGC 1523 GCGACGUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGAGGUGA 5294
1608 UGCACGUC G CAUGGAGA 1524 UCUCCAUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GACGUGCA 5295
1624 ACCACCGU G AACGCCCA 1525 UGGGCGUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACGGUGGU 5296
1628 CCGUGAAC G CCCACAGG 1526 CCUGUGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUUCACGG 5297
1642 AGGAACCU G CCCAAGGU 1527 ACCUUGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGUUCCU 5298
1654 AAGGUCUU G CAUAAGAG 1528 CUCUUAUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGACCUU 5299
1690 AUGUCAAC G ACCGACCU 1529 AGGUCGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUUGACAU 5300
1694 CAACGACC G ACCUUGAG 1530 CUCAAGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGUCGUUG 5301
1700 CCGACCUU G AGGCAUAC 1531 GUAUGCCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGGUCGG 5302
1730 UGUUUAAU G AGUGGGAG 1532 CUCCCACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUUAAACA 5303
1818 AGCACCAU G CAACUUUU 1533 AAAAGUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGGUGCU 5304
1835 UCACCUCU G CCUAAUCA 1534 UGAUUAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAGGUGA 5305
1883 CAAGCUGU G CCUUGGGU 1535 ACCCAAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAGCUUG 5306
1912 UGGACAUU G ACCCGUAU 1536 AUACGGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUGUCCA 5307
1959 UCUUUUUU G CCUUCUGA 1537 UCAGAAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAAAAGA 5308
1966 UGCCUUCU G ACUUCUUU 1538 AAAGAAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAAGGCA 5309
1985 UUCUAUUC G AGAUCUCC 1539 GGAGAUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GAAUAGAA 5310
1996 AUCUCCUC G ACACCGCC 1540 GGCGGUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GAGGAGAU 5311
2002 UCGACACC G CCUCUGCU 1541 AGCAGAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGUGUCGA 5312
2008 CCGCCUCU G CUCUGUAU 1542 AUACAGAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAGGCGG 5313
2092 GUUGGGGU G AGUUGAUG 1543 CAUCAACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCCCAAC 5314
2097 GGUGAGUU G AUGAAUCU 1544 AGAUUCAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AACUCACC 5315
2100 GAGUUGAU G AAUCUAGC 1545 GCUAGAUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCAACUC 5316
Table 42
2237 UUUUGGGC G AGAAACUG 1546 CAGUUUCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCCCAAAA 5317
2251 CUGUUCUU G AAUAUUUG 1547 CAAAUAUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAGAACAG 5318
2282 GUGGAUUC G CACUCCUC 1548 GAGGAGUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAAUCCAC 5319
2293 CUCCUCCU G CAUAUAGA 1549 UCUAUAUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGAGGAG 5320
2311 CACCAAAU G CCCCUAUC 1550 GAUAGGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUUUGGUG 5321
2354 UGUUAGAC G AAGAGGCA 1551 UGCCUCUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUCUAACA 5322
2388 ACUCCCUC G CCUCGCAG 1552 CUGCGAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAGGGAGU 5323
2393 CUCGCCUC G CAGACGAA 1553 UUCGUCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAGGCGAG 5324
2399 UCGCAGAC G AAGGUCUC 1554 GAGACCUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUCUGCGA 5325
2412 UCUCAAUC G CCGCGUCG 1555 CGACGCGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAUUGAGA 5326
2415 CAAUCGCC G CGUCGCAG 1556 CUGCGACG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGCGAUUG 5327
2420 GCCGCGUC G CAGAAGAU 1557 AUCUUCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GACGCGGC 5328
2514 GGUACCUU G CUUUAAUC 1558 GAUUAAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAGGUACC 5329
2549 CUUUUCCU G ACAUUCAU 1559 AUGAAUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGAAAAG 5330
2560 AUUCAUUU G CAGGAGGA 1560 UCCUCCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAUGAAU 5331
2576 ACAUUGUU G AUAGAUGU 1561 ACAUCUAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AACAAUGU 5332
2615 CAGUAAAU G AAAACAGG 1562 CCUGUUUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUUUACUG 5333
2641 UUAACUAU G CCUGCUAG 1563 CUAGCAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUAGUUAA 5334
2645 CUAUGCCU G CUAGGUUU 1564 AAACCUAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGCAUAG 5335
2677 AAAUAUUU G CCCUUAGA 1565 UCUAAGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAUAUUU 5336
2740 UUCCAGAC G CGACAUUA 1566 UAAUGUCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUCUGGAA 5337
2742 CCAGACGC G ACAUUAUU 1567 AAUAAUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCGUCUGG 5338
2804 CACGUAGC G CCUCAUUU 1568 AAAUGAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCUACGUG 5339
2814 CUCAUUUU G CGGGUCAC 1569 GUGACCCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAAUGAG 5340
2875 CAAACCUC G AAAAGGCA 1570 UGCCUUUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GAGGUUUG 5341
2928 UCUUCCCC G AUCAUCAG 1571 CUGAUGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGGAAGA 5342
2946 UGGACCCU G CAUUCAAA 1572 UUUGAAUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGGUCCA 5343
2990 CUCAACCC G CACAAGGA 1573 UCCUUGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGUUGAG 5344
3012 GGCCGGAC G CCAACAAG 1574 CUUGUUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUCCGGCC 5345
3090 GCCCUCAC G CUCAGGGC 1575 GCCCUGAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUGAGGGC 5346
3113 ACAACUGU G CCAGCAGC 1576 GCUGCUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAGUUGU 5347
3132 CUCCUCCU G CCUCCACC 1577 GGUGGAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGAGGAG 5348
51 AGGGCCCU G UACUUUCC 1578 GGAAAGUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGGCCCU 5349
106 AGAAUACU G UCUCUGCC 1579 GGCAGAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUAUUCU 5350
Table 42
148 GGGACCCU G UACCGAAC 1580 GUUCGGUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGGUCCC 5351
198 CUGCUCGU G UUACAGGC 1581 GCCUGUAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACGAGCAG 5352
219 UUUUUCUU G UUGACAAA 1582 UUUGUCAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAGAAAAA 5353
297 ACACCCGU G UGUCUUGG 1583 CCAAGACA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACGGGUGU 5354
299 ACCCGUGU G UCUUGGCC 1584 GGCCAAGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACACGGGU 5355
347 ACCAACCU G UUGUCCUC 1585 GAGGACAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGUUGGU 5356
350 AACCUGUU G UCCUCCAA 1586 UUGGAGGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AACAGGUU 5357
362 UCCAAUUU G UCCUGGUU 1587 AACCAGGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAUUGGA 5358
381 CGCUGGAU G UGUCUGCG 1588 CGCAGACA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUCCAGCG 5359
383 CUGGAUGU G UCUGCGGC 1589 GCCGCAGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACAUCCAG 5360
438 AUCUUCUU G UUGGUUCU 1590 AGAACCAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAGAAGAU 5361
465 CAAGGUAU G UUGCCCGU 1591 ACGGGCAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUACCUUG 5362
476 GCCCGUUU G UCCUCUAA 1592 UUAGAGGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAACGGGC 5363
555 ACCUCUAU G UUUCCCUC 1593 GAGGGAAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUAGAGGU 5364
566 UCCCUCAU G UUGCUGUA 1594 UACAGCAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGAGGGA 5365
572 AUGUUGCU G UACAAAAC 1595 GUUUUGUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCAACAU 5366
602 CUGCACCU G UAUUCCCA 1596 UGGGAAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGUGCAG 5367
694 UGCCAUUU G UUCAGUGG 1597 CCACUGAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAUGGCA 5368
724 CCCCCACU G UCUGGCUU 1598 AAGCCAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUGGGGG 5369
750 UGGAUGAU G UGGUUUUG 1599 CAAAACCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCAUCCA 5370
771 CCAAGUCU G UACAACAU 1600 AUGUUGUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGACUUGG 5371
801 AUGCCGCU G UUACCAAU 1601 AUUGGUAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCGGCAU 5372
818 UUUCUUUU G UCUUUGGG 1602 CCCAAAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAAGAAA 5373
888 UGGGAUAU G UAAUUGGG 1603 CCCAAUUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUAUCCCA 5374
927 AACAUAUU G UACAAAAA 1604 UUUUUGUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUAUGUU 5375
944 AUCAAAAU G UGUUUUAG 1605 CUAAAACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUUUUGAU 5376
946 CAAAAUGU G UUUUAGGA 1606 UCCUAAAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAUUUUG 5377
963 AACUUCCU G UAAACAGG 1607 CCUGUUUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGAAGUU 5378
991 GAAAGUAU G UCAACGAA 1608 UUCGUUGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUACUUUC 5379
1002 AACGAAUU G UGGGUCUU 1609 AAGACCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUUCGUU 5380
1039 CACGCAAU G UGGAUAUU 1610 AAUAUCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUUGCGUG 5381
1137 AACAGUAU G UGAACCUU 1611 AAGGUUCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUACUGUU 5382
1184 UGCCAAGU G UUUGCUGA 1612 UCAGCAAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACUUGGCA 5383
1251 GAACCUUU G UGUCUCCU 1613 AGGAGACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAGGUUC 5384
Table 42
1253 ACCUUUGU G UCUCCUCU 1614 AGAGGAGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACAAAGGU 5385
1294 AGCCGCUU G UUUUGCUC 1615 GAGCAAAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAGCGGCU 5386
1344 ACAAUUCU G UCGUGCUC 1616 GAGCACGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGAAUUGU 5387
1390 GCUAGGCU G UGCUGCCA 1617 UGGCAGCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCCUAGC 5388
1425 CGUCCUUU G UUUACGUC 1618 GACGUAAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAGGACG 5389
1508 CGCCUAUU G UACCGACC 1619 GGUCGGUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAUAGGCG 5390
1557 CCCCGUCU G UGCCUUCU 1620 AGAAGGCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGACGGGG 5391
1581 CGGACCGU G UGCACUUC 1621 GAAGUGCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACGGUCCG 5392
1684 UCAGCAAU G UCAACGAC 1622 GUCGUUGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUUGCUGA 5393
1719 CAAAGACU G UGUGUUUA 1623 UAAACACA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGUCUUUG 5394
1721 AAGACUGU G UGUUUAAU 1624 AUUAAACA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACAGUCUU 5395
1723 GACUGUGU G UUUAAUGA 1625 UCAUUAAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACACAGUC 5396
1772 AGGUCUUU G UACUAGGA 1626 UCCUAGUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAGACCU 5397
1785 AGGAGGCU G UAGGCAUA 1627 UAUGCCUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCCUCCU 5398
1801 AAAUUGGU G UGUUCACC 1628 GGUGAACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCAAUUU 5399
1803 AUUGGUGU G UUCACCAG 1629 CUGGUGAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACACCAAU 5400
1850 CAUCUCAU G UUCAUGUC 1630 GACAUGAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGAGAUG 5401
1856 AUGUUCAU G UCCUACUG 1631 CAGUAGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGAACAU 5402
1864 GUCCUACU G UUCAAGCC 1632 GGCUUGAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUAGGAC 5403
1881 UCCAAGCU G UGCCUUGG 1633 CCAAGGCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGCUUGGA 5404
1939 GAGCUUCU G UGGAGUUA 1634 UAACUCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAAGCUC 5405
2013 UCUGCUCU G UAUCGGGG 1635 CCCCGAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAGCAGA 5406
2045 GGAACAUU G UUCACCUC 1636 GAGGUGAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUGUUCC 5407
2082 GCUAUUCU G UGUUGGGG 1637 CCCCAACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAAUAGC 5408
2084 UAUUCUGU G UUGGGGUG 1638 CACCCCAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAGAAUA 5409
2167 UCAGCUAU G UCAACGUU 1639 AACGUUGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUAGCUGA 5410
2205 CAACUAUU G UGGUUUCA 1640 UGAAACCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUAGUUG 5411
2222 CAUUUCCU G UCUUACUU 1641 AAGUAAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGGAAAUG 5412
2245 GAGAAACU G UUCUUGAA 1642 UUCAAGAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUUUCUC 5413
2262 UAUUUGGU G UCUUUUGG 1643 CCAAAAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCAAAUA 5414
2274 UUUGGAGU G UGGAUUCG 1644 CGAAUCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACUCCAAA 5415
2344 AAACUACU G UUGUUAGA 1645 UCUAACAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUAGUUU 5416
2347 CUACUGUU G UUAGACGA 1646 UCGUCUAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AACAGUAG 5417
2450 AUCUCAAU G UUAGUAUU 1647 AAUACUAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUUGAGAU 5418
Table 42
2573 AGGACAUU G UUGAUAGA 1648 UCUAUCAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAUGUCCU 5419
2583 UGAUAGAU G UAAGCAAU 1649 AUUGCUUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUCUAUCA 5420
2594 AGCAAUUU G UGGGGCCC 1650 GGGCCCCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAUUGCU 5421
2663 AUCCCAAU G UUACUAAA 1651 UUUAGUAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUUGGGAU 5422
2717 CAGAGUAU G UAGUUAAU 1652 AUUAACUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUACUCUG 5423
2901 AUCUUUCU G UCCCCAAU 1653 AUUGGGGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGAAAGAU 5424
3071 GGGGGACU G UUGGGGUG 1654 CACCCCAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGUCCCCC 5425
3111 UCACAACU G UGCCAGCA 1655 UGCUGGCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGUUGUGA 5426
40 AUCCCAGA G UCAGGGCC 1656 GGCCCUGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCUGGGAU 5427
46 GAGUCAGG G CCCUGUAC 1657 GUACAGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCUGACUC 5428
65 UCCUGCUG G UGGCUCCA 1658 UGGAGCCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGCAGGA 5429
68 UGCUGGUG G CUCCAGUU 1659 AACUGGAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACCAGCA 5430
74 UGGCUCCA G UUCAGGAA 1660 UUCCUGAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGGAGCCA 5431
85 CAGGAACA G UGAGCCCU 1661 AGGGCUCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUUCCUG 5432
89 AACAGUGA G CCCUGCUC 1662 GAGCAGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCACUGUU 5433
120 GCCAUAUC G UCAAUCUU 1663 AAGAUUGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GAUAUGGC 5434
196 CCCUGCUC G UGUUACAG 1664 CUGUAACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GAGCAGGG 5435
205 UGUUACAG G CGGGGUUU 1665 AAACCCCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGUAACA 5436
210 CAGGCGGG G UUUUUCUU 1666 AAGAAAAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCGCCUG 5437
248 ACCACAGA G UCUAGACU 1667 AGUCUAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCUGUGGU 5438
258 CUAGACUC G UGGUGGAC 1668 GUCCACCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GAGUCUAG 5439
261 GACUCGUG G UGGACUUC 1669 GAAGUCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACGAGUC 5440
295 GAACACCC G UGUGUCUU 1670 AAGACACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGUGUUC 5441
305 GUGUCUUG G CCAAAAUU 1671 AAUUUUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAGACAC 5442
318 AAUUCGCA G UCCCAAAU 1672 AUUUGGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCGAAUU 5443
332 AAUCUCCA G UCACUCAC 1673 GUGAGUGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGAGAUU 5444
368 UUGUCCUG G UUAUCGCU 1674 AGCGAUAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGGACAA 5445
390 UGUCUGCG G CGUUUUAU 1675 AUAAAACG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGCAGACA 5446
392 UCUGCGGC G UUUUAUCA 1676 UGAUAAAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCCGCAGA 5447
442 UCUUGUUG G UUCUUCUG 1677 CAGAAGAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAACAAGA 5448
461 CUAUCAAG G UAUGUUGC 1678 GCAACAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUGAUAG 5449
472 UGUUGCCC G UUUGUCCU 1679 AGGACAAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGCAACA 5450
506 AACAACCA G CACCGGAC 1680 GUCCGGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGUUGUU 5451
625 CAUCUUGG G CUUUCGCA 1681 UGCGAAAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAAGAUG 5452
Table 42
648 CUAUGGGA G UGGGCCUC 1682 GAGGCCCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCCCAUAG 5453
652 GGGAGUGG G CCUCAGUC 1683 GACUGAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCACUCCC 5454
658 GGGCCUCA G UCCGUUUC 1684 GAAACGGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGAGGCCC 5455
662 CUCAGUCC G UUUCUCUU 1685 AAGAGAAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGACUGAG 5456
672 UUCUCUUG G CUCAGUUU 1686 AAACUGAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAAGAGAA 5457
677 UUGGCUCA G UUUACUAG 1687 CUAGUAAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGAGCCAA 5458
685 GUUUACUA G UGCCAUUU 1688 AAAUGGCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAGUAAAC 5459
699 UUUGUUCA G UGGUUCGU 1689 ACGAACCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGAACAAA 5460
702 GUUCAGUG G UUCGUAGG 1690 CCUACGAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACUGAAC 5461
706 AGUGGUUC G UAGGGCUU 1691 AAGCCCUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAACCACU 5462
711 UUCGUAGG G CUUUCCCC 1692 GGGGAAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCUACGAA 5463
729 ACUGUCUG G CUUUCAGU 1693 ACUGAAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGACAGU 5464
736 GGCUUUCA G UUAUAUGG 1694 CCAUAUAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGAAAGCC 5465
753 AUGAUGUG G UUUUGGGG 1695 CCCCAAAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACAUCAU 5466
762 UUUUGGGG G CCAAGUCU 1696 AGACUUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCCAAAA 5467
767 GGGGCCAA G UCUGUACA 1697 UGUACAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGGCCCC 5468
785 CAUCUUGA G UCCCUUUA 1698 UAAAGGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCAAGAUG 5469
826 GUCUUUGG G UAUACAUU 1699 AAUGUAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAAAGAC 5470
898 AAUUGGGA G UUGGGGCA 1700 UGCCCCAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCCAAUU 5471
904 GAGUUGGG G CACAUUGC 1701 GCAAUGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCAACUC 5472
971 GUAAACAG G CCUAUUGA 1702 UCAAUAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGUUUAC 5473
987 AUUGGAAA G UAUGUCAA 1703 UUGACAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUCCAAU 5474
1006 AAUUGUGG G UCUUUUGG 1704 CCAAAAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCACAAUU 5475
1016 CUUUUGGG G UUUGCCGC 1705 GCGGCAAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCAAAAG 5476
1080 GCAUACAA G CAAAACAG 1706 CUGUUUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGUAUGC 5477
1089 CAAAACAG G CUUUUACU 1707 AGUAAAAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGUUUUG 5478
1116 CUUACAAG G CCUUUCUA 1708 UAGAAAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUGUAAG 5479
1126 CUUUCUAA G UAAACAGU 1709 ACUGUUUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUAGAAAG 5480
1133 AGUAAACA G UAUGUGAA 1710 UUCACAUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUUUACU 5481
1152 UUUACCCC G UUGCUCGG 1711 CCGAGCAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGGUAAA 5482
1160 GUUGCUCG G CAACGGCC 1712 GGCCGUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGAGCAAC 5483
1166 CGGCAACG G CCUGGUCU 1713 AGACCAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGUUGCCG 5484
1171 ACGGCCUG G UCUAUGCC 1714 GGCAUAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGGCCGU 5485
1182 UAUGCCAA G UGUUUGCU 1715 AGCAAACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGGCAUA 5486
Table 42
1207 CCCCACUG G UUGGGGCU 1716 AGCCCCAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGUGGGG 5487
1213 UGGUUGGG G CUUGGCCA 1717 UGGCCAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCAACCA 5488
1218 GGGGCUUG G CCAUAGGC 1718 GCCUAUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAAGCCCC 5489
1225 GGCCAUAG G CCAUCAGC 1719 GCUGAUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUAUGGCC 5490
1232 GGCCAUCA G CGCAUGCG 1720 CGCAUGCG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGAUGGCC 5491
1240 GCGCAUGC G UGGAACCU 1721 AGGUUCCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCAUGCGC 5492
1287 AACUCCUA G CCGCUUGU 1722 ACAAGCGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAGGAGUU 5493
1306 UGCUCGCA G CAGGUCUG 1723 CAGACCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCGAGCA 5494
1310 CGCAGCAG G UCUGGGGC 1724 GCCCCAGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUGCUGCG 5495
1317 GGUCUGGG G CAAAACUC 1725 GAGUUUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCAGACC 5496
1347 AUUCUGUC G UGCUCUCC 1726 GGAGAGCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GACAGAAU 5497
1379 UUUCCAUG G CUGCUAGG 1727 CCUAGCAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUGGAAA 5498
1387 GCUGCUAG G CUGUGCUG 1728 CAGCACAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUAGCAGC 5499
1418 CGCGGGAC G UCCUUUGU 1729 ACAAAGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUCCCGCG 5500
1431 UUGUUUAC G UCCCGUCG 1730 CGACGGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUAAACAA 5501
1436 UACGUCCC G UCGGCGCU 1731 AGCGCCGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGACGUA 5502
1440 UCCCGUCG G CGCUGAAU 1732 AUUCAGCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGACGGGA 5503
1471 CUCCCGGG G CCGCUUGG 1733 CCAAGCGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCGGGAG 5504
1481 CGCUUGGG G CUCUACCG 1734 CGGUAGAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCAAGCG 5505
1517 UACCGACC G UCCACGGG 1735 CCCGUGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGUCGGUA 5506
1526 UCCACGGG G CGCACCUC 1736 GAGGUGCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCGUGGA 5507
1553 GACUCCCC G UCUGUGCC 1737 GGCACAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGGGAGUC 5508
1579 GCCGGACC G UGUGCACU 1738 AGUGCACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGUCCGGC 5509
1605 CUCUGCAC G UCGCAUGG 1739 CCAUGCGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUGCAGAG 5510
1622 AGACCACC G UGAACGCC 1740 GGCGUUCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGUGGUCU 5511
1649 UGCCCAAG G UCUUGCAU 1741 AUGCAAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUGGGCA 5512
1679 GACUUUCA G CAAUGUCA 1742 UGACAUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGAAAGUC 5513
1703 ACCUUGAG G CAUACUUC 1743 GAAGUAUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCAAGGU 5514
1732 UUUAAUGA G UGGGAGGA 1744 UCCUCCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCAUUAAA 5515
1741 UGGGAGGA G UUGGGGGA 1745 UCCCCCAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCUCCCA 5516
1754 GGGAGGAG G UUAGGUUA 1746 UAACCUAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCCUCCC 5517
1759 GAGGUUAG G UUAAAGGU 1747 ACCUUUAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUAACCUC 5518
1766 GGUUAAAG G UCUUUGUA 1748 UACAAAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUUAACC 5519
1782 ACUAGGAG G CUGUAGGC 1749 GCCUACAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCCUAGU 5520
Table 42
1789 GGCUGUAG G CAUAAAUU 1750 AAUUUAUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUACAGCC 5521
1799 AUAAAUUG G UGUGUUCA 1751 UGAACACA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAAUUUAU 5522
1811 GUUCACCA G CACCAUGC 1752 GCAUGGUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGGUGAAC 5523
1870 CUGUUCAA G CCUCCAAG 1753 CUUGGAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUGAACAG 5524
1878 GCCUCCAA G CUGUGCCU 1754 AGGCACAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUGGAGGC 5525
1890 UGCCUUGG G UGGCUUUG 1755 CAAAGCCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCAAGGCA 5526
1893 CUUGGGUG G CUUUGGGG 1756 CCCCAAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACCCAAG 5527
1901 GCUUUGGG G CAUGGACA 1757 UGUCCAUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCAAAGC 5528
1917 AUUGACCC G UAUAAAGA 1758 UCUUUAUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGGUCAAU 5529
1933 AAUUUGGA G CUUCUGUG 1759 CACAGAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCCAAAUU 5530
1944 UCUGUGGA G UUACUCUC 1760 GAGAGUAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCCACAGA 5531
2023 AUCGGGGG G CCUUAGAG 1761 CUCUAAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCCCGAU 5532
2031 GCCUUAGA G UCUCCGGA 1762 UCCGGAGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCUAAGGC 5533
2062 ACCAUACG G CACUCAGG 1763 CCUGAGUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGUAUGGU 5534
2070 GCACUCAG G CAAGCUAU 1764 AUAGCUUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGAGUGC 5535
2074 UCAGGCAA G CUAUUCUG 1765 CAGAAUAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGCCUGA 5536
2090 GUGUUGGG G UGAGUUGA 1766 UCAACUCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCAACAC 5537
2094 UGGGGUGA G UUGAUGAA 1767 UUCAUCAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCACCCCA 5538
2107 UGAAUCUA G CCACCUGG 1768 CCAGGUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAGAUUCA 5539
2116 CCACCUGG G UGGGAAGU 1769 ACUUCCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAGGUGG 5540
2123 GGUGGGAA G UAAUUUGG 1770 CCAAAUUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCCCACC 5541
2140 AAGAUCCA G CAUCCAGG 1771 CCUGGAUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGAUCUU 5542
2155 GGGAAUUA G UAGUCAGC 1772 GCUGACUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAAUUCCC 5543
2158 AAUUAGUA G UCAGCUAU 1773 AUAGCUGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UACUAAUU 5544
2162 AGUAGUCA G CUAUGUCA 1774 UGACAUAG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGACUACU 5545
2173 AUGUCAAC G UUAAUAUG 1775 CAUAUUAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUUGACAU 5546
2183 UAAUAUGG G CCUAAAAA 1776 UUUUUAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAUAUUA 5547
2208 CUAUUGUG G UUUCACAU 1777 AUGUGAAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACAAUAG 5548
2235 ACUUUUGG G CGAGAAAC 1778 GUUUCUCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAAAAGU 5549
2260 AAUAUUUG G UGUCUUUU 1779 AAAAGACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAAUAUU 5550
2272 CUUUUGGA G UGUGGAUU 1780 AAUCCACA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCAAAAG 5551
2360 ACGAAGAG G CAGGUCCC 1781 GGGACCUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCUUCGU 5552
2364 AGAGGCAG G UCCCCUAG 1782 CUAGGGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGCCUCU 5553
2403 AGACGAAG G UCUCAAUC 1783 GAUUGAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUCGUCU 5554
Table 42
2417 AUCGCCGC G UCGCAGAA 1784 UUCUGCGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCGGCGAU 5555
2454 CAAUGUUA G UAUUCCUU 1785 AAGGAAUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAACAUUG 5556
2474 CACAUAAG G UGGGAAAC 1786 GUUUCCCA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUUAUGUG 5557
2491 UUUACGGG G CUUUAUUC 1787 GAAUAAAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCGUAAA 5558
2507 CUUCUACG G UACCUUGC 1788 GCAAGGUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGUAGAAG 5559
2530 CCUAAAUG G CAAACUCC 1789 GGAGUUUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUUUAGG 5560
2587 AGAUGUAA G CAAUUUGU 1790 ACAAAUUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUACAUCU 5561
2599 UUUGUGGG G CCCCUUAC 1791 GUAAGGGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCACAAA 5562
2609 CCCUUACA G UAAAUGAA 1792 UUCAUUUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGUAAGGG 5563
2650 CCUGCUAG G UUUUAUCC 1793 GGAUAAAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUAGCAGG 5564
2701 AUCAAACC G UAUUAUCC 1794 GGAUAAUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGUUUGAU 5565
2713 UAUCCAGA G UAUGUAGU 1795 ACUACAUA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCUGGAUA 5566
2720 AGUAUGUA G UUAAUCAU 1796 AUGAUUAA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UACAUACU 5567
2768 UUUGGAAG G CGGGGAUC 1797 GAUCCCCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUCCAAA 5568
2791 AAAAGAGA G UCCACACG 1798 CGUGUGGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCUCUUUU 5569
2799 GUCCACAC G UAGCGCCU 1799 AGGCGCUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUGUGGAC 5570
2802 CACACGUA G CGCCUCAU 1800 AUGAGGCG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UACGUGUG 5571
2818 UUUUGCGG G UCACCAUA 1801 UAUGGUGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGCAAAA 5572
2848 GAUCUACA G CAUGGGAG 1802 CUCCCAUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUAGAUC 5573
2857 CAUGGGAG G UUGGUCUU 1803 AAGACCAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCCCAUG 5574
2861 GGAGGUUG G UCUUCCAA 1804 UUGGAAGA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAACCUCC 5575
2881 UCGAAAAG G CAUGGGGA 1805 UCCCCAUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUUUCGA 5576
2936 GAUCAUCA G UUGGACCC 1806 GGGUCCAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGAUGAUC 5577
2955 CAUUCAAA G CCAACUCA 1807 UGAGUUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUGAAUG 5578
2964 CCAACUCA G UAAAUCCA 1808 UGGAUUUA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGAGUUGG 5579
3005 GACAACUG G CCGGACGC 1809 GCGUCCGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGUUGUC 5580
3021 CCAACAAG G UGGGAGUG 1810 CACUCCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUGUUGG 5581
3027 AGGUGGGA G UGGGAGCA 1811 UGCUCCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCCACCU 5582
3033 GAGUGGGA G CAUUCGGG 1812 CCCGAAUG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCCACUC 5583
3041 GCAUUCGG G CCAGGGUU 1813 AACCCUGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGAAUGC 5584
3047 GGGCCAGG G UUCACCCC 1814 GGGGUGAA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCUGGCCC 5585
3077 CUGUUGGG G UGGAGCCC 1815 GGGCUCCA GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCAACAG 5586
3082 GGGGUGGA G CCCUCACG 1816 CGUGAGGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCACCCC 5587
3097 CGCUCAGG G CCUACUCA 1817 UGAGUAGG GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCUGAGCG 5588
Table 42
3117 CUGUGCCA G CAGCUCCU 1818 AGGAGCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGGCACAG 5589
3120 UGCCAGCA G CUCCUCCU 1819 AGGAGGAG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCUGGCA 5590
3146 ACCAAUCG G CAGUCAGG 1820 CCUGACUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGAUUGGU 5591
3149 AAUCGGCA G UCAGGAAG 1821 CUUCCUGA GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCCGAUU 5592
3158 UCAGGAAG G CAGCCUAC 1822 GUAGGCUG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUUCCUGA 5593
3161 GGAAGGCA G CCUACUCC 1823 GGAGUAGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCCUUCC 5594
3204 AUCCUCAG G CCAUGCAG 1824 CUGCAUGG GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUGAGGAU 5595
31 CUCUUCAA G AUCCCAGA 2196 UCUGGGAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUGAAGAG 5596
38 AGAUCCCA G AGUCAGGG 2197 CCCUGACU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGGGAUCU 5597
44 CAGAGUCA G GGCCCUGU 2198 ACAGGGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGACUCUG 5598
45 AGAGUCAG G GCCCUGUA 2199 UACAGGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUGACUCU 5599
64 UUCCUGCU G GUGGCUCC 2200 GGAGCCAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCAGGAA 5600
67 CUGCUGGU G GCUCCAGU 2201 ACUGGAGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACCAGCAG 5601
79 CCAGUUCA G GAACAGUG 2202 CACUGUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGAACUGG 5602
80 CAGUUCAG G AACAGUGA 2203 UCACUGUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGAACUG 5603
99 CCUGCUCA G AAUACUGU 2204 ACAGUAUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGAGCAGG 5604
135 UUAUCGAA G ACUGGGGA 2205 UCCCCAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCGAUAA 5605
139 CGAAGACU G GGGACCCU 2206 AGGGUCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUCUUCG 5606
140 GAAGACUG G GGACCCUG 2207 CAGGGUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGUCUUC 5607
141 AAGACUGG G GACCCUGU 2208 ACAGGGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAGUCUU 5608
142 AGACUGGG G ACCCUGUA 2209 UACAGGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCAGUCU 5609
159 CCGAACAU G GAGAACAU 2210 AUGUUCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGUUCGG 5610
160 CGAACAUG G AGAACAUC 2211 GAUGUUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUGUUCG 5611
162 AACAUGGA G AACAUCGC 2212 GCGAUGUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCAUGUU 5612
175 UCGCAUCA G GACUCCUA 2213 UAGGAGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGAUGCGA 5613
176 CGCAUCAG G ACUCCUAG 2214 CUAGGAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGAUGCG 5614
184 GACUCCUA G GACCCCUG 2215 CAGGGGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAGGAGUC 5615
185 ACUCCUAG G ACCCCUGC 2216 GCAGGGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUAGGAGU 5616
204 GUGUUACA G GCGGGGUU 2217 AACCCCGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUAACAC 5617
207 UUACAGGC G GGGUUUUU 2218 AAAAACCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCCUGUAA 5618
208 UACAGGCG G GGUUUUUC 2219 GAAAAACC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGCCUGUA 5619
209 ACAGGCGG G GUUUUUCU 2220 AGAAAAAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGCCUGU 5620
246 AUACCACA G AGUCUAGA 2221 UCUAGACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUGGUAU 5621
253 AGAGUCUA G ACUCGUGG 2222 CCACGAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAGACUCU 5622
Table 42
260 AGACUCGU G GUGGACUU 2223 AAGUCCAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACGAGUCU 5623
263 CUCGUGGU G GACUUCUC 2224 GAGAAGUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACCACGAG 5624
264 UCGUGGUG G ACUUCUCU 2225 AGAGAAGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACCACGA 5625
283 AUUUUCUA G GGGGAACA 2226 UGUUCCCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAGAAAAU 5626
284 UUUUCUAG G GGGAACAC 2227 GUGUUCCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUAGAAAA 5627
285 UUUCUAGG G GGAACACC 2228 GGUGUUCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCUAGAAA 5628
286 UUCUAGGG G GAACACCC 2229 GGGUGUUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCUAGAA 5629
287 UCUAGGGG G AACACCCG 2230 CGGGUGUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCCUAGA 5630
304 UGUGUCUU G GCCAAAAU 2231 AUUUUGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAGACACA 5631
367 UUUGUCCU G GUUAUCGC 2232 GCGAUAAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGACAAA 5632
377 UUAUCGCU G GAUGUGUC 2233 GACACAUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGCGAUAA 5633
378 UAUCGCUG G AUGUGUCU 2234 AGACACAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGCGAUA 5634
389 GUGUCUGC G GCGUUUUA 2235 UAAAACGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCAGACAC 5635
441 UUCUUGUU G GUUCUUCU 2236 AGAAGAAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AACAAGAA 5636
450 GUUCUUCU G GACUAUCA 2237 UGAUAGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGAAGAAC 5637
451 UUCUUCUG G ACUAUCAA 2238 UUGAUAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGAAGAA 5638
460 ACUAUCAA G GUAUGUUG 2239 CAACAUAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGAUAGU 5639
490 UAAUUCCA G GAUCAUCA 2240 UGAUGAUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGAAUUA 5640
491 AAUUCCAG G AUCAUCAA 2241 UUGAUGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGGAAUU 5641
511 CCAGCACC G GACCAUGC 2242 GCAUGGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGUGCUGG 5642
512 CAGCACCG G ACCAUGCA 2243 UGCAUGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGGUGCUG 5643
544 CUGCUCAA G GAACCUCU 2244 AGAGGUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGAGCAG 5644
545 UGCUCAAG G AACCUCUA 2245 UAGAGGUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUGAGCA 5645
585 AAACCUAC G GACGGAAA 2246 UUUCCGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUAGGUUU 5646
586 AACCUACG G ACGGAAAC 2247 GUUUCCGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGUAGGUU 5647
589 CUACGGAC G GAAACUGC 2248 GCAGUUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUCCGUAG 5648
590 UACGGACG G AAACUGCA 2249 UGCAGUUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGUCCGUA 5649
623 AUCAUCUU G GGCUUUCG 2250 CGAAAGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGAUGAU 5650
624 UCAUCUUG G GCUUUCGC 2251 GCGAAAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAGAUGA 5651
644 AUACCUAU G GGAGUGGG 2252 CCCACUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUAGGUAU 5652
645 UACCUAUG G GAGUGGGC 2253 GCCCACUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUAGGUA 5653
646 ACCUAUGG G AGUGGGCC 2254 GGCCCACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAUAGGU 5654
650 AUGGGAGU G GGCCUCAG 2255 CUGAGGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACUCCCAU 5655
651 UGGGAGUG G GCCUCAGU 2256 ACUGAGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACUCCCA 5656
Table 42
671 UUUCUCUU G GCUCAGUU 2257 AACUGAGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAGAGAAA 5657
701 UGUUCAGU G GUUCGUAG 2258 CUACGAAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACUGAACA 5658
709 GGUUCGUA G GGCUUUCC 2259 GGAAAGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UACGAACC 5659
710 GUUCGUAG G GCUUUCCC 2260 GGGAAAGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUACGAAC 5660
728 CACUGUCU G GCUUUCAG 2261 CUGAAAGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGACAGUG 5661
743 AGUUAUAU G GAUGAUGU 2262 ACAUCAUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AUAUAACU 5662
744 GUUAUAUG G AUGAUGUG 2263 CACAUCAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUAUAAC 5663
752 GAUGAUGU G GUUUUGGG 2264 CCCAAAAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACAUCAUC 5664
758 GUGGUUUU G GGGGCCAA 2265 UUGGCCCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAACCAC 5665
759 UGGUUUUG G GGGCCAAG 2266 CUUGGCCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAAAACCA 5666
760 GGUUUUGG G GGCCAAGU 2267 ACUUGGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCAAAACC 5667
761 GUUUUGGG G GCCAAGUC 2268 GACUUGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCAAAAC 5668
824 UUGUCUUU G GGUAUACA 2269 UGUAUACC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAGACAA 5669
825 UGUCUUUG G GUAUACAU 2270 AUGUAUAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAAGACA 5670
856 AACAAAAA G AUGGGGAU 2271 AUCCCCAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUUUGUU 5671
859 AAAAAGAU G GGGAUAUU 2272 AAUAUCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUCUUUUU 5672
860 AAAAGAUG G GGAUAUUC 2273 GAAUAUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUCUUUU 5673
861 AAAGAUGG G GAUAUUCC 2274 GGAAUAUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAUCUUU 5674
862 AAGAUGGG G AUAUUCCC 2275 GGGAAUAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCAUCUU 5675
881 AACUUCAU G GGAUAUGU 2276 ACAUAUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGAAGUU 5676
882 ACUUCAUG G GAUAUGUA 2277 UACAUAUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUGAAGU 5677
883 CUUCAUGG G AUAUGUAA 2278 UUACAUAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAUGAAG 5678
894 AUGUAAUU G GGAGUUGG 2279 CCAACUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUUACAU 5679
895 UGUAAUUG G GAGUUGGG 2280 CCCAACUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAUUACA 5680
896 GUAAUUGG G AGUUGGGG 2281 CCCCAACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAAUUAC 5681
901 UGGGAGUU G GGGCACAU 2282 AUGUGCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AACUCCCA 5682
902 GGGAGUUG G GGCACAUU 2283 AAUGUGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAACUCCC 5683
903 GGAGUUGG G GCACAUUG 2284 CAAUGUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAACUCC 5684
917 UUGCCACA G GAACAUAU 2285 AUAUGUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUGGCAA 5685
918 UGCCACAG G AACAUAUU 2286 AAUAUGUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGUGGCA 5686
952 GUGUUUUA G GAAACUUC 2287 GAAGUUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAAAACAC 5687
953 UGUUUUAG G AAACUUCC 2288 GGAAGUUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUAAAACA 5688
970 UGUAAACA G GCCUAUUG 2289 CAAUAGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUUUACA 5689
982 UAUUGAUU G GAAAGUAU 2290 AUACUUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUCAAUA 5690
Table 42
983 AUUGAUUG G AAAGUAUG 2291 CAUACUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAAUCAAU 5691
1004 CGAAUUGU G GGUCUUUU 2292 AAAAGACC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACAAUUCG 5692
1005 GAAUUGUG G GUCUUUUG 2293 CAAAAGAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACAAUUC 5693
1013 GGUCUUUU G GGGUUUGC 2294 GCAAACCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAAGACC 5694
1014 GUCUUUUG G GGUUUGCC 2295 GGCAAACC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAAAAGAC 5695
1015 UCUUUUGG G GUUUGCCG 2296 CGGCAAAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCAAAAGA 5696
1041 CGCAAUGU G GAUAUUCU 2297 AGAAUAUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACAUUGCG 5697
1042 GCAAUGUG G AUAUUCUG 2298 CAGAAUAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACAUUGC 5698
1088 GCAAAACA G GCUUUUAC 2299 GUAAAAGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGUUUUGC 5699
1115 ACUUACAA G GCCUUUCU 2300 AGAAAGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUGUAAGU 5700
1159 CGUUGCUC G GCAACGGC 2301 GCCGUUGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAGCAACG 5701
1165 UCGGCAAC G GCCUGGUC 2302 GACCAGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUUGCCGA 5702
1170 AACGGCCU G GUCUAUGC 2303 GCAUAGAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGCCGUU 5703
1206 CCCCCACU G GUUGGGGC 2304 GCCCCAAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUGGGGG 5704
1210 CACUGGUU G GGGCUUGG 2305 CCAAGCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AACCAGUG 5705
1211 ACUGGUUG G GGCUUGGC 2306 GCCAAGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAACCAGU 5706
1212 CUGGUUGG G GCUUGGCC 2307 GGCCAAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAACCAG 5707
1217 UGGGGCUU G GCCAUAGG 2308 CCUAUGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGCCCCA 5708
1224 UGGCCAUA G GCCAUCAG 2309 CUGAUGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAUGGCCA 5709
1242 GCAUGCGU G GAACCUUU 2310 AAAGGUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACGCAUGC 5710
1243 CAUGCGUG G AACCUUUG 2311 CAAAGGUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACGCAUG 5711
1277 CAUACCGC G GAACUCCU 2312 AGGAGUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCGGUAUG 5712
1278 AUACCGCG G AACUCCUA 2313 UAGGAGUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGCGGUAU 5713
1309 UCGCAGCA G GUCUGGGG 2314 CCCCAGAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCUGCGA 5714
1314 GCAGGUCU G GGGCAAAA 2315 UUUUGCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGACCUGC 5715
1315 CAGGUCUG G GGCAAAAC 2316 GUUUUGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGACCUG 5716
1316 AGGUCUGG G GCAAAACU 2317 AGUUUUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAGACCU 5717
1329 AACUCAUC G GGACUGAC 2318 GUCAGUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GAUGAGUU 5718
1330 ACUCAUCG G GACUGACA 2319 UGUCAGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGAUGAGU 5719
1331 CUCAUCGG G ACUGACAA 2320 UUGUCAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGAUGAG 5720
1378 AUUUCCAU G GCUGCUAG 2321 CUAGCAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGGAAAU 5721
1386 GGCUGCUA G GCUGUGCU 2322 AGCACAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAGCAGCC 5722
1402 UGCCAACU G GAUCCUAC 2323 GUAGGAUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUUGGCA 5723
1403 GCCAACUG G AUCCUACG 2324 CGUAGGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAGUUGGC 5724
Table 42
1413 UCCUACGC G GGACGUCC 2325 GGACGUCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCGUAGGA 5725
1414 CCUACGCG G GACGUCCU 2326 AGGACGUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGCGUAGG 5726
1415 CUACGCGG G ACGUCCUU 2327 AAGGACGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCGCGUAG 5727
1439 GUCCCGUC G GCGCUGAA 2328 UUCAGCGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GACGGGAC 5728
1454 AAUCCCGC G GACGACCC 2329 GGGUCGUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GCGGGAUU 5729
1455 AUCCCGCG G ACGACCCC 2330 GGGGUCGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGCGGGAU 5730
1468 CCCCUCCC G GGGCCGCU 2331 AGCGGCCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGGAGGGG 5731
1469 CCCUCCCG G GGCCGCUU 2332 AAGCGGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGGGAGGG 5732
1470 CCUCCCGG G GCCGCUUG 2333 CAAGCGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCGGGAGG 5733
1478 GGCCGCUU G GGGCUCUA 2334 UAGAGCCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAGCGGCC 5734
1479 GCCGCUUG G GGCUCUAC 2335 GUAGAGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAAGCGGC 5735
1480 CCGCUUGG G GCUCUACC 2336 GGUAGAGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCAAGCGG 5736
1523 CCGUCCAC G GGGCGCAC 2337 GUGCGCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUGGACGG 5737
1524 CGUCCACG G GGCGCACC 2338 GGUGCGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGUGGACG 5738
1525 GUCCACGG G GCGCACCU 2339 AGGUGCGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGUGGAC 5739
1544 CUUUACGC G GACUCCCC 2340 GGGGAGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCGUAAAG 5740
1545 UUUACGCG G ACUCCCCG 2341 CGGGGAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGCGUAAA 5741
1574 CAUCUGCC G GACCGUGU 2342 ACACGGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGCAGAUG 5742
1575 AUCUGCCG G ACCGUGUG 2343 CACACGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGGCAGAU 5743
1612 CGUCGCAU G GAGACCAC 2344 GUGGUCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGCGACG 5744
1613 GUCGCAUG G AGACCACC 2345 GGUGGUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUGCGAC 5745
1615 CGCAUGGA G ACCACCGU 2346 ACGGUGGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCAUGCG 5746
1635 CGCCCACA G GAACCUGC 2347 GCAGGUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGUGGGCG 5747
1636 GCCCACAG G AACCUGCC 2348 GGCAGGUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGUGGGC 5748
1648 CUGCCCAA G GUCUUGCA 2349 UGCAAGAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGGGCAG 5749
1660 UUGCAUAA G AGGACUCU 2350 AGAGUCCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUAUGCAA 5750
1662 GCAUAAGA G GACUCUUG 2351 CAAGAGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCUUAUGC 5751
1663 CAUAAGAG G ACUCUUGG 2352 CCAAGAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCUUAUG 5752
1670 GGACUCUU G GACUUUCA 2353 UGAAAGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGAGUCC 5753
1671 GACUCUUG G ACUUUCAG 2354 CUGAAAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAGAGUC 5754
1702 GACCUUGA G GCAUACUU 2355 AAGUAUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCAAGGUC 5755
1715 ACUUCAAA G ACUGUGUG 2356 CACACAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUGAAGU 5756
1734 UAAUGAGU G GGAGGAGU 2357 ACUCCUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACUCAUUA 5757
1735 AAUGAGUG G GAGGAGUU 2358 AACUCCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACUCAUU 5758
Table 42
1736 AUGAGUGG G AGGAGUUG 2359 CAACUCCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCACUCAU 5759
1738 GAGUGGGA G GAGUUGGG 2360 CCCAACUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCCCACUC 5760
1739 AGUGGGAG G AGUUGGGG 2361 CCCCAACU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUCCCACU 5761
1744 GAGGAGUU G GGGGAGGA 2362 UCCUCCCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AACUCCUC 5762
1745 AGGAGUUG G GGGAGGAG 2363 CUCCUCCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAACUCCU 5763
1746 GGAGUUGG G GGAGGAGG 2364 ccuccucc GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCAACUCC 5764
1747 GAGUUGGG G GAGGAGGU 2365 ACCUCCUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCAACUC 5765
1748 AGUUGGGG G AGGAGGUU 2366 AACCUCCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCCAACU 5766
1750 UUGGGGGA G GAGGUUAG 2367 CUAACCUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCCCCCAA 5767
1751 UGGGGGAG G AGGUUAGG 2368 CCUAACCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUCCCCCA 5768
1753 GGGGAGGA G GUUAGGUU 2369 AACCUAAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCCUCCCC 5769
1758 GGAGGUUA G GUUAAAGG 2370 CCUUUAAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAACCUCC 5770
1765 AGGUUAAA G GUCUUUGU 2371 ACAAAGAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUUAACCU 5771
1778 UUGUACUA G GAGGCUGU 2372 ACAGCCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAGUACAA 5772
1779 UGUACUAG G AGGCUGUA 2373 UACAGCCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUAGUACA 5773
1781 UACUAGGA G GCUGUAGG 2374 CCUACAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCUAGUA 5774
1788 AGGCUGUA G GCAUAAAU 2375 AUUUAUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UACAGCCU 5775
1798 CAUAAAUU G GUGUGUUC 2376 GAACACAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAUUUAUG 5776
1888 UGUGCCUU G GGUGGCUU 2377 AAGCCACC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGGCACA 5777
1889 GUGCCUUG G GUGGCUUU 2378 AAAGCCAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAGGCAC 5778
1892 CCUUGGGU G GCUUUGGG 2379 CCCAAAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCCAAGG 5779
1898 GUGGCUUU G GGGCAUGG 2380 CCAUGCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAGCCAC 5780
1899 UGGCUUUG G GGCAUGGA 2381 UCCAUGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAAGCCA 5781
1900 GGCUUUGG G GCAUGGAC 2382 GUCCAUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAAAGCC 5782
1905 UGGGGCAU G GACAUUGA 2383 UCAAUGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGCCCCA 5783
1906 GGGGCAUG G ACAUUGAC 2384 GUCAAUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUGCCCC 5784
1924 CGUAUAAA G AAUUUGGA 2385 UCCAAAUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUAUACG 5785
1930 AAGAAUUU G GAGCUUCU 2386 AGAAGCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAUUCUU 5786
1931 AGAAUUUG G AGCUUCUG 2387 CAGAAGCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAAUUCU 5787
1941 GCUUCUGU G GAGUUACU 2388 AGUAACUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAGAAGC 5788
1942 CUUCUGUG G AGUUACUC 2389 GAGUAACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACAGAAG 5789
1987 CUAUUCGA G AUCUCCUC 2390 GAGGAGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCGAAUAG 5790
2018 UCUGUAUC G GGGGGCCU 2391 AGGCCCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GAUACAGA 5791
2019 CUGUAUCG G GGGGCCUU 2392 AAGGCCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGAUACAG 5792
Table 42
2020 UGUAUCGG G GGGCCUUA 2393 UAAGGCCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCGAUACA 5793
2021 GUAUCGGG G GGCCUUAG 2394 CUAAGGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCGAUAC 5794
2022 UAUCGGGG G GCCUUAGA 2395 UCUAAGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCCGAUA 5795
2029 GGGCCUUA G AGUCUCCG 2396 CGGAGACU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAAGGCCC 5796
2037 GAGUCUCC G GAACAUUG 2397 CAAUGUUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGAGACUC 5797
2038 AGUCUCCG G AACAUUGU 2398 ACAAUGUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGGAGACU 5798
2061 CACCAUAC G GCACUCAG 2399 CUGAGUGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GUAUGGUG 5799
2069 GGCACUCA G GCAAGCUA 2400 UAGCUUGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGAGUGCC 5800
2087 UCUGUGUU G GGGUGAGU 2401 ACUCACCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AACACAGA 5801
2088 CUGUGUUG G GGUGAGUU 2402 AACUCACC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAACACAG 5802
2089 UGUGUUGG G GUGAGUUG 2403 CAACUCAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCAACACA 5803
2114 AGCCACCU G GGUGGGAA 2404 UUCCCACC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGUGGCU 5804
2115 GCCACCUG G GUGGGAAG 2405 CUUCCCAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGGUGGC 5805
2118 ACCUGGGU G GGAAGUAA 2406 UUACUUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCCAGGU 5806
2119 CCUGGGUG G GAAGUAAU 2407 AUUACUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACCCAGG 5807
2120 CUGGGUGG G AAGUAAUU 2408 AAUUACUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCACCCAG 5808
2130 AGUAAUUU G GAAGAUCC 2409 GGAUCUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAUUACU 5809
2131 GUAAUUUG G AAGAUCCA 2410 UGGAUCUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAAUUAC 5810
2134 AUUUGGAA G AUCCAGCA 2411 UGCUGGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCCAAAU 5811
2147 AGCAUCCA G GGAAUUAG 2412 CUAAUUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGAUGCU 5812
2148 GCAUCCAG G GAAUUAGU 2413 ACUAAUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGGAUGC 5813
2149 CAUCCAGG G AAUUAGUA 2414 UACUAAUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCUGGAUG 5814
2181 GUUAAUAU G GGCCUAAA 2415 UUUAGGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUAUUAAC 5815
2182 UUAAUAUG G GCCUAAAA 2416 UUUUAGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUAUUAA 5816
2195 AAAAAUCA G ACAACUAU 2417 AUAGUUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGAUUUUU 5817
2207 ACUAUUGU G GUUUCACA 2418 UGUGAAAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAAUAGU 5818
2233 UUACUUUU G GGCGAGAA 2419 UUCUCGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAAGUAA 5819
2234 UACUUUUG G GCGAGAAA 2420 UUUCUCGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAAAGUA 5820
2239 UUGGGCGA G AAACUGUU 2421 AACAGUUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCGCCCAA 5821
2259 GAAUAUUU G GUGUCUUU 2422 AAAGACAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAUAUUC 5822
2269 UGUCUUUU G GAGUGUGG 2423 CCACACUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAAAGACA 5823
2270 GUCUUUUG G AGUGUGGA 2424 UCCACACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAAAGAC 5824
2276 UGGAGUGU G GAUUCGCA 2425 UGCGAAUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACACUCCA 5825
2277 GGAGUGUG G AUUCGCAC 2426 GUGCGAAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACACUCC 5826
Table 42
2300 UGCAUAUA G ACCACCAA 2427 UUGGUGGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAUAUGCA 5827
2334 ACACUUCC G GAAACUAC 2428 GUAGUUUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GGAAGUGU 5828
2335 CACUUCCG G AAACUACU 2429 AGUAGUUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CGGAAGUG 5829
2351 UGUUGUUA G ACGAAGAG 2430 CUCUUCGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAACAACA 5830
2357 UAGACGAA G AGGCAGGU 2431 ACCUGCCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUCGUCUA 5831
2359 GACGAAGA G GCAGGUCC 2432 GGACCUGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCUUCGUC 5832
2363 AAGAGGCA G GUCCCCUA 2433 UAGGGGAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCCUCUU 5833
2372 GUCCCCUA G AAGAAGAA 2434 UUCUUCUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAGGGGAC 5834
2375 CCCUAGAA G AAGAACUC 2435 GAGUUCUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUCUAGGG 5835
2378 UAGAAGAA G AACUCCCU 2436 AGGGAGUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUCUUCUA 5836
2396 GCCUCGCA G ACGAAGGU 2437 ACCUUCGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCGAGGC 5837
2402 CAGACGAA G GUCUCAAU 2438 AUUGAGAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUCGUCUG 5838
2423 GCGUCGCA G AAGAUCUC 2439 GAGAUCUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGCGACGC 5839
2426 UCGCAGAA G AUCUCAAU 2440 AUUGAGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCUGCGA 5840
2438 UCAAUCUC G GGAAUCUC 2441 GAGAUUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GAGAUUGA 5841
2439 CAAUCUCG G GAAUCUCA 2442 UGAGAUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGAGAUUG 5842
2440 AAUCUCGG G AAUCUCAA 2443 UUGAGAUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGAGAUU 5843
2463 UAUUCCUU G GACACAUA 2444 UAUGUGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGGAAUA 5844
2464 AUUCCUUG G ACACAUAA 2445 UUAUGUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAGGAAU 5845
2473 ACACAUAA G GUGGGAAA 2446 UUUCCCAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUAUGUGU 5846
2476 CAUAAGGU G GGAAACUU 2447 AAGUUUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCUUAUG 5847
2477 AUAAGGUG G GAAACUUU 2448 AAAGUUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACCUUAU 5848
2478 UAAGGUGG G AAACUUUA 2449 UAAAGUUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCACCUUA 5849
2488 AACUUUAC G GGGCUUUA 2450 UAAAGCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUAAAGUU 5850
2489 ACUUUACG G GGCUUUAU 2451 AUAAAGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGUAAAGU 5851
2490 CUUUACGG G GCUUUAUU 2452 AAUAAAGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGUAAAG 5852
2506 UCUUCUAC G GUACCUUG 2453 CAAGGUAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GUAGAAGA 5853
2529 UCCUAAAU G GCAAACUC 2454 GAGUUUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUUUAGGA 5854
2563 CAUUUGCA G GAGGACAU 2455 AUGUCCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGCAAAUG 5855
2564 AUUUGCAG G AGGACAUU 2456 AAUGUCCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGCAAAU 5856
2566 UUGCAGGA G GACAUUGU 2457 ACAAUGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCUGCAA 5857
2567 UGCAGGAG G ACAUUGUU 2458 AACAAUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUCCUGCA 5858
2580 UGUUGAUA G AUGUAAGC 2459 GCUUACAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UAUCAACA 5859
2596 CAAUUUGU G GGGCCCCU 2460 AGGGGCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACAAAUUG 5860
Table 42
2597 AAUUUGUG G GGCCCCUU 2461 AAGGGGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACAAAUU 5861
2598 AUUUGUGG G GCCCCUUA 2462 UAAGGGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCACAAAU 5862
2622 UGAAAACA G GAGACUUA 2463 UAAGUCUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGUUUUCA 5863
2623 GAAAACAG G AGACUUAA 2464 UUAAGUCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUGUUUUC 5864
2625 AAACAGGA G ACUUAAAU 2465 AUUUAAGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UCCUGUUU 5865
2649 GCCUGCUA G GUUUUAUC 2466 GAUAAAAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAGCAGGC 5866
2684 UGCCCUUA G AUAAAGGG 2467 CCCUUUAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UAAGGGCA 5867
2690 UAGAUAAA G GGAUCAAA 2468 UUUGAUCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUUAUCUA 5868
2691 AGAUAAAG G GAUCAAAC 2469 GUUUGAUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUUUAUCU 5869
2692 GAUAAAGG G AUCAAACC 2470 GGUUUGAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCUUUAUC 5870
2711 AUUAUCCA G AGUAUGUA 2471 UACAUACU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGGAUAAU 5871
2737 UACUUCCA G ACGCGACA 2472 UGUCGCGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGGAAGUA 5872
2763 CACUCUUU G GAAGGCGG 2473 CCGCCUUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAAGAGUG 5873
2764 ACUCUUUG G AAGGCGGG 2474 CCCGCCUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAAGAGU 5874
2767 CUUUGGAA G GCGGGGAU 2475 AUCCCCGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUCCAAAG 5875
2770 UGGAAGGC G GGGAUCUU 2476 AAGAUCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCCUUCCA 5876
2771 GGAAGGCG G GGAUCUUA 2477 UAAGAUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGCCUUCC 5877
2772 GAAGGCGG G GAUCUUAU 2478 AUAAGAUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCGCCUUC 5878
2773 AAGGCGGG G AUCUUAUA 2479 UAUAAGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCGCCUU 5879
2787 AUAUAAAA G AGAGUCCA 2480 UGGACUCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUUAUAU 5880
2789 AUAAAAGA G AGUCCACA 2481 UGUGGACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCUUUUAU 5881
2816 CAUUUUGC G GGUCACCA 2482 UGGUGACC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GCAAAAUG 5882
2817 AUUUUGCG G GUCACCAU 2483 AUGGUGAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGCAAAAU 5883
2832 AUAUUCUU G GGAACAAG 2484 CUUGUUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AAGAAUAU 5884
2833 UAUUCUUG G GAACAAGA 2485 UCUUGUUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAAGAAUA 5885
2834 AUUCUUGG G AACAAGAU 2486 AUCUUGUU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAAGAAU 5886
2840 GGGAACAA G AUCUACAG 2487 CUGUAGAU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGUUCCC 5887
2852 UACAGCAU G GGAGGUUG 2488 CAACCUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGCUGUA 5888
2853 ACAGCAUG G GAGGUUGG 2489 CCAACCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUGCUGU 5889
2854 CAGCAUGG G AGGUUGGU 2490 ACCAACCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAUGCUG 5890
2856 GCAUGGGA G GUUGGUCU 2491 AGACCAAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UCCCAUGC 5891
2860 GGGAGGUU G GUCUUCCA 2492 UGGAAGAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AACCUCCC 5892
2880 CUCGAAAA G GCAUGGGG 2493 CCCCAUGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUUUCGAG 5893
2885 AAAGGCAU G GGGACAAA 2494 UUUGUCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGCCUUU 5894
Table 42
2886 AAGGCAUG G GGACAAAU 2495 AUUUGUCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAUGCCUU 5895
2887 AGGCAUGG G GACAAAUC 2496 GAUUUGUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCAUGCCU 5896
2888 GGCAUGGG G ACAAAUCU 2497 AGAUUUGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCCAUGCC 5897
2915 AAUCCCCU G GGAUUCUU 2498 AAGAAUCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AGGGGAUU 5898
2916 AUCCCCUG G GAUUCUUC 2499 GAAGAAUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAGGGGAU 5899
2917 UCCCCUGG G AUUCUUCC 2500 GGAAGAAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCAGGGGA 5900
2939 CAUCAGUU G GACCCUGC 2501 GCAGGGUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AACUGAUG 5901
2940 AUCAGUUG G ACCCUGCA 2502 UGCAGGGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAACUGAU 5902
2973 UAAAUCCA G AUUGGGAC 2503 GUCCCAAU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGGAUUUA 5903
2977 UCCAGAUU G GGACCUCA 2504 UGAGGUCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG AAUCUGGA 5904
2978 CGAGAUUG G GACCUCAA 2505 UUGAGGUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAAUCUGG 5905
2979 CAGAUUGG G ACCUCAAC 2506 GUUGAGGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCAAUCUG 5906
2996 CCGCACAA G GACAACUG 2507 CAGUUGUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUGUGCGG 5907
2997 CGCACAAG G ACAACUGG 2508 CCAGUUGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUUGUGCG 5908
3004 GGACAACU G GCCGGACG 2509 CGUCCGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AGUUGUCC 5909
3008 AACUGGCC G GACGCCAA 2510 UUGGCGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GGCCAGUU 5910
3009 ACUGGCCG G ACGCCAAC 2511 GUUGGCGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGGCCAGU 5911
3020 GCCAACAA G GUGGGAGU 2512 ACUCCCAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UUGUUGGC 5912
3023 AACAAGGU G GGAGUGGG 2513 CCCACUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACCUUGUU 5913
3024 ACAAGGUG G GAGUGGGA 2514 UCCCACUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACCUUGU 5914
3025 CAAGGUGG G AGUGGGAG 2515 CUCCCACU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCACCUUG 5915
3029 GUGGGAGU G GGAGCAUU 2516 AAUGCUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG ACUCCCAC 5916
3030 UGGGAGUG G GAGCAUUC 2517 GAAUGCUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CACUCCCA 5917
3031 GGGAGUGG G AGCAUUCG 2518 CGAAUGCU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCACUCCC 5918
3039 GAGCAUUC G GGCCAGGG 2519 CCCUGGCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG GAAUGCUC 5919
3040 AGCAUUCG G GCCAGGGU 2520 ACCCUGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CGAAUGCU 5920
3045 UCGGGCCA G GGUUCACC 2521 GGUGAACC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGGCCCGA 5921
3046 CGGGCCAG G GUUCACCC 2522 GGGUGAAC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CUGGCCCG 5922
3063 CUCCCCAU G GGGGACUG 2523 CAGUCCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AUGGGGAG 5923
3064 UCCCCAUG G GGGACUGU 2524 ACAGUCCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CAUGGGGA 5924
3065 CCCCAUGG G GGACUGUU 2525 AACAGUCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCAUGGGG 5925
3066 CCCAUGGG G GACUGUUG 2526 CAACAGUC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCAUGGG 5926
3067 CCAUGGGG G ACUGUUGG 2527 CCAACAGU GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG CCCCAUGG 5927
3074 GGACUGUU G GGGUGGAG 2528 CUCCACCC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG AACAGUCC 5928
Table 42
3075 GACUGUUG G GGUGGAGC 2529 GCUCCACC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CAACAGUC 5929
3076 ACUGUUGG G GUGGAGCC 2530 GGCUCCAC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCAACAGU 5930
3079 GUUGGGGU G GAGCCCUC 2531 GAGGGCUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG ACCCCAAC 5931
3080 UUGGGGUG G AGCCCUCA 2532 UGAGGGCU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CACCCCAA 5932
3095 CACGCUCA G GGCCUACU 2533 AGUAGGCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGAGCGUG 5933
3096 ACGCUCAG G GCCUACUC 2534 GAGUAGGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUGAGCGU 5934
3145 CACCAAUC G GCAGUCAG 2535 CUGACUGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG GAUUGGUG 5935
3153 GGCAGUCA G GAAGGCAG 2536 CUGCCUUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UGACUGCC 5936
3154 GCAGUCAG G AAGGCAGC 2537 GCUGCCUU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUGACUGC 5937
3157 GUCAGGAA G GCAGCCUA 2538 UAGGCUGC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUCCUGAC 5938
3187 ACCUCUAA G GGACACUC 2539 GAGUGUCC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG UUAGAGGU 5939
3188 CCUCUAAG G GACACUCA 2540 UGAGUGUC GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CUUAGAGG 5940
3189 CUCUAAGG G ACACUCAU 2541 AUGAGUGU GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG CCUUAGAG 5941
3203 CAUCCUCA G GCCAUGCA 2542 UGCAUGGC GGAGGAAACUCC cu UCAAGGACAUCGUCCGGG UGAGGAUG 5942
Input Sequence = AF100308. Cut Site = YG/M or UG/U.
Stem Length = 8. Core Sequence = GGAGGAAACUCC CU UCAAGGACAUCGUCCGGG
AF100308 (Hepatitis B virus strain 2-18, 3215 bp)
Table 43
Table 43: Human HBV Ribozyme and Target Sequence
Table 43
Table 43
Table 43
Table 43
Table 43
Table 43
Table 43
Table 43
Table 43
Table 43
Table 43
UPPER CASE = RIBO lower case = 2'-0-methyl s = phosphorothioate linkage B = inverted deoxyabasic residue U = 2'-deoxy-2'-C-allyl Uridine U= 2 '-deoxy-2 '-amino Uridine C = 2'-deoxy-2'-amino Cytidine
Table 44 Table 44: Group Designation and Dosage levels for HBV transgenic mouse study
*administered via sc infusion using Alzet mini-osmotic pumps
Table 45
TABLE 45. NUCLEOSIDES USED FOR CHEMICAL SYNTHESIS OF MODIFIED NUCLEOTIDE TRIPHOSPHATES
Table 45
Table 46
Table 46: PHOSPHORYLATION OF URIDINE IN THE PRESENCE OF DMAP
Table 48: INCORPORATION OF MODIFIED NUCLEOTIDE TRIPHOSPHATES
Table 48
Table 49
Table 49: INCORPORATION OF MODIFIED NUCLEOTIDE TRIPHOSPHATES USING WILD TYPE BACTERIOPHAGE T7 POLYMERASE
Table 50 a: Incorporation of 2'-his-UTP and Modified CTP's
Table 50 b: Incorporation of 2'-his-UTP, 2-amino CTP, and Modified ATP's
Numbers shown are a percentage of incorporation compared to the all-RNA control
* -Bold number indicates best observed rate of modified nucleotide triphosphate incorporation
Table 51
Table 51: INCORPORATION OF 2'-his-UTP, 2'-NH2-CTP, 2'-NH2-DAP, and rGTP USING VARIOUS REACTION CONDITIONS
Numbers shown are a percentage of incorporation compared to the all-RNA control
* Two highest levels of incorporation contained both methanol and LiCl
Table 52
Table 52: Selection of Oligonucleotides with Ribozyme Activity
Table 53
Table 53: Kinetic Activity of Combinatorial Libraries
Table 54
Table 54: Kinetic Activity of Clones within N60 and N40 Combinatorial Libraries
Table 55 Table 55: Effect of Magnesium Concentration of the Cleavage Rate of N20
I able 56
Enzymatic Nucleic Acid Motifs Targeting HCV
Seq ID Al Las Sequence Rz Seq ID
:ucc 1 HCV.R1A-6 A b.Rz-10/5 ggagugucgc GgaggaaacucC CU UCAAGGACAUCGUCCGGG cccau B 39
3CGU 2 HCV.R1A-56 Amb.Rz-10/5 acgcuuucug GgaggaaacucC CU UCAAGGACAUCGUCCGGG gugaa B 40
3UAU 3 HCV.R1A-75 Amb.Rz-10/5 auacuaacgc GgaggaaacucC CU UCAAGGACAUCGUCCGGG auggc B 41
JAUG 4 HCV.R1A-76 Amb.Rz-10/5 cauacuaacg GgaggaaacucC cu UCAAGGACAUCGUCCGGG caugg B 42
:CAG 5 HCV.R1A-95 Amb.Rz-10/5 cuggaggcug GgaggaaacucC cu UCAAGGACAUCGUCCGGG acgac B 43
:GGU 6 HCV.R1A-138 Amb.Rz-10/5 accgguuccg GgaggaaacucC cu UCAAGGACAUCGUCCGGG agacc B 44
^CAC 7 HCV.R1A-146 Amb.Rz-10/5 guguacucac GgaggaaacucC cu UCAAGGACAUCGUCCGGG gguuc B 45 CAG 8 HCV.R1A-158 Amb.Rz-10/5 cuggcaauuc GgaggaaacucC cu UCAAGGACAUCGUCCGGG ggugu B 46
ΓGAC 9 HCV.R1A-164 Amb.Rz-10/5 gucguccugg GgaggaaacucC cu UCAAGGACAUCGUCCGGG aauuc B 47
JUCU 10 HCV.R1A-176 Amb.Rz-10/5 agaaaggacc GgaggaaacucC cu UCAAGGACAUCGUCCGGG ggucg B 48
JCUU 11 HCV.RlA-177 Amb.Rz-10/5 aagaaaggac GgaggaaacucC cu UCAAGGACAUCGUCCGGG cgguc B 49
JGGG 12 HCV.R1A-209 Amb.Rz-10/5 cccaaaucuc GgaggaaacucC cu UCAAGGACAUCGUCCGGG aggca B 50
3AGU 13 HCV.R1A-237 Amb.Rz-10/5 acucggcuag GgaggaaacucC cu UCAAGGACAUCGUCCGGG agucu B 51
3AAA 14 HCV.R1A-254 Amb.Rz-10/5 uuucgcgacc GgaggaaacucC cu UCAAGGACAUCGUCCGGG aacac B 52
!yyiG 15 HCV. RIA-255 Amb.Rz-10/5 cuuucgcgac GgaggaaacucC cu UCAAGGACAUCGUCCGGG caaca B 53
3CCU 16 HCV. RIA-259 Amb.Rz-10/5 aggccuuucg GgaggaaacucC cu UCAAGGACAUCGUCCGGG gaccc B 54
3GUA 17 HCV.R1A-266 Amb.Rz-10/5 uaccacaagg GgaggaaacucC cu UCAAGGACAUCGUCCGGG cuuuc B 55
_CUG 18 HCV.R1A-273 Amb.Rz-10/5 caggcaguac GgaggaaacucC cu UCAAGGACAUCGUCCGGG acaag B 56
3CGA 19 HCV.R1A-288 Amb.Rz-10/5 ucgcaagcac GgaggaaacucC cu UCAAGGACAUCGUCCGGG cuauc B 57
>-GUG 20 HCV.R1A-291 Amb.Rz-10/5 cacucgcaag GgaggaaacucC cu UCAAGGACAUCGUCCGGG acccu B 58
JCCA 21 HCV.R1A-7 Amb.Rz-10/5 uggagugucg GgaggaaacucC cu UCAAGGACAUCGUCCGGG cccca B 59
"CAU 22 HCV.R1A-119 Amb.Rz-10/5 auggcucucc GgaggaaacucC cu UCAAGGACAUCGUCCGGG gggag B 60 AUA 23 HCV.R1A-120 Amb.Rz-10/5 uauggcucuc GgaggaaacucC cu UCAAGGACAUCGUCCGGG cggga B 61
3GAA 24 HCV.R1A-133 Amb.Rz-10/5 uuccgcagac GgaggaaacucC cu UCAAGGACAUCGUCCGGG acuau B 62
3UGA 25 HCV.R1A-140 Amb.Rz-10/5 ucaccgguuc GgaggaaacucC cu UCAAGGACAUCGUCCGGG gcaga B 63
_CCG 26 HCV.R1A-188 Amb.Rz-10/5 cgggguuauc GgaggaaacucC cu UCAAGGACAUCGUCCGGG aagaa B 64
3CCU 27 HCV.R1A-198 Amb.Rz-10/5 aggcauugag GgaggaaacucC cu UCAAGGACAUCGUCCGGG 9999U B 65
3AUU 28 HCV.R1A-205 Amb.Rz-10/5 aaucuccagg GgaggaaacucC cu UCAAGGACAUCGUCCGGG auuga B 66
ZCCC 29 HCV. RIA-217 Amb.Rz-10/5 ggggcacgcc GgaggaaacucC cu UCAAGGACAUCGUCCGGG aaauc B 67 CCC 30 HCV.R1A-218 Amb.Rz-10/5 gggggcacgc GgaggaaacucC cu UCAAGGACAUCGUCCGGG caaau B 68
:CCG 31 HCV.R1A-219 Amb.Rz-10/5 cgggggcacg GgaggaaacucC cu UCAAGGACAUCGUCCGGG ccaaa B 69
"AAG 32 HCV.R1A-223 Amb.Rz-10/5 cuugcggggg GgaggaaacucC cu UCAAGGACAUCGUCCGGG acgcc B 70
JGCU 33 HCV.R1A-229 Amb.Rz-10/5 agcagucuug GgaggaaacucC cu UCAAGGACAUCGUCCGGG ggggg B 71
^GGG 34 HCV.R1A-279 Amb.Rz-10/5 cccuaucagg GgaggaaacucC cu UCAAGGACAUCGUCCGGG aguac B 72
Table 56
lower case = 2'-O-methyl TJ, C = 2 '-deoxy-2 '-amino U, 2 '-deoxy-2 '-amino C
B = inverted deoxyabasic
Table 57 Table 57. Additional Class II enzymatic nucleic acid Motifs
Table 58
Table 58: Human Her2 Class II Ribozyme and Target Sequen
RPI# NTPos Substrate Seq ID Ribozyme Alias Ribozyme Sequence Seq ID
#
18722 180 CAUGGA G CUGGCG 85 erbB2-180 Zin Rz-6 amino stabl cs9scscsa9 GccgaaagGCGaGucaaGGuCu uccaug B 194
18835 184 GAGCUG G CGGCCU 86 erbB2-184 Zin Rz-6 ammo stabl as9S9scs°9 GccgaaagGCGaGucaaGGuCu cagcuc B 195
18828 276 AGCUGCG G CUCCCUG 87 erbB2-276 Zin Rz-7 amino stabl caaagsgggag GccgaaagGCGaGucaaGGuCu cgcagcu B 196
18653 314 UGCUCC G CCACCU 88 erbB2-314 Zin Rz-6 ammo stabl as9s9sus99 GccgaaagGCGaGucaaGGuCu ggagca B 197
18825 314 AUGCUCC G CCACCUC 89 erbB2-314 Zin Rz-7 amino stabl 9sas9S s u99 GccgaaagGCGaGucaaGGuCu ggagcau B 198
18831 379 ACCAAU G CCAGCC 90 erbB2-379 Zin Rz-6 amino stabl s9scsus99 GccgaaagGCGaGucaaGGuCu auuggu B 199
18680 433 GCUCAUC G CUCACAA 91 erbB2-433 Zin Rz-7 ammo stabl usus9sus9a9 GccgaaagGCGaGucaaGGuCu gaugagc B 200
18711 594 GGAGCU G CAGCUU 92 erbB2-594 Zin Rz-6 amino stabl asas9scsu9 GccgaaagGCGaGucaaGGuCu agcucc B 201
18681 594 GGGAGCU G CAGCUUC 93 erbB2-594 Zm Rz-7 amino stabl 9s asas9scu9 GccgaaagGCGaGucaaGGuCu agcuccc B 202
18697 597 GCUGCA G CUUCGA 94 erbB2-597 Zin Rz-6 ammo stabl uscs9sasa9 GccgaaagGCGaGucaaGGuCu ugcagc B 203
18665 597 AGCUGCA G CUUCGAA 95 erbB2-597 Zin Rz-7 amino stabl ususc S9S aa9 GccgaaagGCGaGucaaGGuCu ugcagcu B 204
18712 659 AGCUCU G CUACCA 96 erbB2-659 Zin Rz-6 ammo stabl us9s9susa GccgaaagGCGaGucaaGGuCu agagcu B 205
18682 659 CAGCUCU G CUACCAG 97 erbB2-659 Zin Rz-7 amino stabl csus9s9sua9 GccgaaagGCGaGucaaGGuCu agagcug B 206
18683 878 CUGACU G CUGCCA 98 erbB2-878 Zin Rz-6 amino stabl us9S9scsa9 GccgaaagGCGaGucaaGGuCu agucag B 207
18654 878 ACUGACU G CUGCCAU 99 erbB2-878 Zin Rz-7 amino stabl asus9a s ca9 GccgaaagGCGaGucaaGGuCu agucagu B 208
18685 881 ACUGCU G CCAUGA 100 erbB2-881 Zin Rz-6 ammo stabl usc S asus99 GccgaaagGCGaGucaaGGuCu agcagu B 209
18684 881 GACUGCU G CCAUGAG 101 erbB2-881 Zin Rz-7 ammo stabl csuscsasu99 GccgaaagGCGaGucaaGGuCu agcaguc B 210
18723 888 GCCAUGA G CAGUGUG 102 erbB2-888 Zm Rz-7 amino stabl csascsa3CU9 GccgaaagGCGaGucaaGGuCu ucauggc B 211
18686 929 CUGACU G CCUGGC 103 erbB2-929 Zin Rz-6 amino stabl 9scscsas99 GccgaaagGCGaGucaaGGuCu agucag B 212
18648 929 UCUGACU G CCUGGCC 104 erbB2-929 Zin Rz-7 amino stabl 9s93 cscsa99 GccgaaagGCGaGucaaGGuCu agucaga B 213
18666 934 UGCCUG G CCUGCC 105 erbB2-934 Zin Rz-6 ammo stabl 939scsas99 GccgaaagGCGaGucaaGGuCu caggca B 214
18651 934 CUGCCUG G CCUGCCU 106 erbB2-934 Zm Rz-7 amino stabl a S9s9scsa99 GccgaaagGCGaGucaaGGuCu caggcag B 215
18655 938 UGGCCU G CCUCCA 107 erbB2-938 Zin Rz-6 ammo stabl us9s9s as99 GccgaaagGCGaGucaaGGuCu aggcca B 216
18649 938 CUGGCCU G CCUCCAC 108 erbB2-938 Zin Rz 7 amino stabl 9sus9S9sa99 GccgaaagGCGaGucaaGGuCu aggccag B 217
18667 969 CUGUGA G CUGCAC 109 erbB2-969 Zin Rz-6 amino stabl 9sus9scsa9 GccgaaagGCGaGucaaGGuCu ucacag B 218
18668 969 UCUGUGA G CUGCACU 110 erbB2-969 Zm Rz-7 amino stabl as9a u 39sca9 GccgaaagGCGaGucaaGGuCu ucacaga B 219
18656 972 UGAGCU G CACUGC 111 erbB2-972 Zin Rz-6 amino stabl 9scsas9su9 GccgaaagGCGaGucaaGGuCu agcuca B 220
18657 972 GUGAGCU G CACUGCC 112 erbB2-972 Zin Rz-7 amino stabl 9s9scsas9u9 GccgaaagGCGaGucaaGGuCu agcucac B 221
19294 972 erbB2-972 Zm Rz-6 ammo stabl g c a g ug GccaauuugugGCGaGucaaGGuCu agcuca B 222
Table 58
19295 972 erbB2-972 Zm Rz-6 amino stabl ggcgasg3ug GccAAuuuGuGGCGaGucaaGGuCu agcuca B 223
19293 972 erbB2-972 Zm Rz-6 amino stabl 9scsa S9su9 GccgaaagGCGaGuGaGGuCu agcuca B 224
19292 972 erbB2-972 Zin Rz-6 amino stabl 9scsas9su9 GccgaaagGCGaGuGaGGuCu agcuca B 225
19296 972 erbB2-972 Zin Rz-6 amino stabl g c a g ug GccacAAuuuGuGGcagGCGaGucaaGGuCu agcuca B 226
19727 972 erbB2-972 Zm Rz-6 amino stabl 9s sas9su9 gccgaaaggCgagugagguCu agcuca B 227
19728 972 erbB2-972 Zm Rz-6 amino stabl 9scsas9su9 gccgaaaggCgagugagGuCu agcuca B 228
18659 1199 GAGUGU G CUAUGG 113 erbB2-1199 Zin Rz-6 amino stabl cscsa3Usa9 GccgaaagGCGaGucaaGGuCu acacuc B 229
18658 1199 CGAGUGU G CUAUGGU 114 erbB2-1199 Zm Rz-7 ammo stabl ascscsasua9 GccgaaagGCGaGucaaGGuCu acacucg B 230
18724 1205 GCUAUG G UCUGGG 115 erbB2-1205 Zin Rz-6 ammo stabl cscscsas9a GccgaaagGCGaGucaaGGuCu cauagc B 231
18669 1205 UGCUAUG G UCUGGGC 116 erbB2-1205 Zin Rz-7 amino stabl 9sc S cscsa9a GccgaaagGCGaGucaaGGuCu cauagca B 232
18725 1211 GUCUGG G CAUGGA 117 erbB2-1211 Z Rz-6 amino stabl uscscsasu9 GccgaaagGCGaGucaaGGuCu ccagac B 233
18726 1292 UUGGGA G CCUGGC 118 erbB2-1292 Zin Rz-6 amino stabl 9scscsas99 GccgaaagGCGaGucaaGGuCu ucccaa B 234
18698 1292 UUUGGGA G CCUGGCA 119 erbB2-1292 Zin Rz-7 amino stabl us9scscsa99 GccgaaagGCGaGucaaGGuCu ucccaaa B 235
18727 1313 CCGGAGA G CUUUGAU 120 erbB2-1313 Zin Rz-7 amino stabl asuscsasaa9 GccgaaagGCGaGucaaGGuCu ucuccgg B 236
18699 1397 UCACAG G UUACCU 121 erbB2-1397 Zin Rz-6 ammo stabl as9s9su S aa GccgaaagGCGaGucaaGGuCu cuguga B 237
18728 1414 AUCUCA G CAUGGC 122 erbB2-1414 Zin Rz-6 ammo stabl 9scs sasu9 GccgaaagGCGaGucaaGGuCu ugagau B 238
18670 1414 CAUCUCA G CAUGGCC 123 erbB2-1414 Zin Rz-7 amino stabl 9s9s cscsau9 GccgaaagGCGaGucaaGGuCu ugagaug B 239
18671 1536 GCUGGG G CUGCGC 124 erbB2-1536 Zin Rz-6 amino stabl 9scs9scsa9 GccgaaagGCGaGucaaGGuCu cccagc B 240
18687 1541 GGCUGC G CUCACU 125 erbB2-1541 Zin Rz-6 amino stabl as9sus9sa9 GccgaaagGCGaGucaaGGuCu gcagcc B 241
18829 1562 CUGGGCA G UGGACUG 126 erbB2-1562 Zin Rz-7 ammo stabl csas9suscca GccgaaagGCGaGucaaGGuCu ugcccag B 242
18830 1626 GGGACCA G CUCUUUC 127 erbB2-1626 Zin Rz-7 amino stabl 9sasas s9a9 GccgaaagGCGaGucaaGGuCu ugguccc B 243
18700 1755 CACCCA G UGUGUC 128 erbB2-1755 Zin Rz-6 amino stabl 9sascsasca GccgaaagGCGaGucaaGGuCu ugggug B 244
18672 1755 CCACCCA G UGUGUCA 129 erbB2-1755 Zin Rz-7 ammo stabl us9a ascsaca GccgaaagGCGaGucaaGGuCu ugggugg B 245
18688 1757 CCCAGU G UGUCAA 130 erbB2-1757 Zm Rz-6 ammo stabl usus9s asca GccgaaagGCGaGucaaGGuCu acuggg B 246
18660 1757 ACCCAGU G UGUCAAC 131 erbB2-1757 Zin Rz-7 amino stabl 9s usus9s aca GccgaaagGCGaGucaaGGuCu acugggu B 247
18689 1759 CAGUGU G UCAACU 132 erbB2-1759 Zin Rz-6 amino stabl as9s usu__9a GccgaaagGCGaGucaaGGuCu acacug B 248
18690 1759 CCAGUGU G UCAACUG 133 erbB2-1759 Zin Rz-7 amino stabl csas9susu9a GccgaaagGCGaGucaaGGuCu acacugg B 249
18701 1784 UUCGGG G CCAGGA 134 erbB2-1784 Zin Rz-6 ammo stabl u S cscsus99 GccgaaagGCGaGucaaGGuCu cccgaa B 250
18673 1784 CUUCGGG G CCAGGAG 135 erbB2-1784 Zm Rz-7 amino stabl csuscscsu99 GccgaaagGCGaGucaaGGuCu cccgaag B 251
18691 2063 UCAACU G CACCCA 136 erbB2-2063 Zin Rz-6 ami o stabl u S9s9s9su9 GccgaaagGCGaGucaaGGuCu aguuga B 252
18661 2063 AUCAACU G CACCCAC 137 erbB2-2063 Zin Rz-7 amino stabl 9sus9sg s9u9 GccgaaagGCGaGucaaGGuCu aguugau B 253
Table 58
18692 2075 ACUCCU G UGUGGA 138 erbB2-2075 Zin Rz 6 ammo stabl uscscsasca GccgaaagGCGaGucaaGGuCu aggagu B 254
18729 2116 CAGAGA G CCAGCC 139 erbB2-2116 Zin Rz-6 ammo stabl 9s9a sus99 GccgaaagGCGaGucaaGGuCu ucucug B 255
18832 2247 GACUGCU G CAGGAAA 140 erbB2-2247 Zm Rz-7 amino stabl u3ususc a cu9 GccgaaagGCGaGucaaGGuCu agcaguc B 256
18833 2271 UGGAGCC G CUGACAC 141 erbB2-2271 Zin Rz-7 amino stabl 9__u a9a u a ca9 GccgaaagGCGaGucaaGGuCu ggcucca B 257
18702 2341 AGGAAG G UGAAGG 142 erbB2-2341 Zin Rz-6 amino stabl c„ 8c3„u„8u„Sca GccgaaagGCGaGucaaGGuCu cuuccu B 258
18730 2347 GUGAAG G UGCUUG 143 erbB2-2347 Zin Rz-6 amino stabl c3a a a a93 ca GccgaaagGCGaGucaaGGuCu cuucac B 259
18674 2347 GGUGAAG G UGCUUGG 144 erbB2-2347 Zm Rz-7 amino stabl cscsasasgca GccgaaagGCGaGucaaGGuCu cuucacc B 260
18713 2349 GAAGGU G CUUGGA 145 erbB2-2349 Zm Rz-6 amino stabl u„ 3c_3c„3a_8ag3 GccgaaagGCGaGucaaGGuCu accuuc B 261
18693 2349 UGAAGGU G CUUGGAU 146 erbB2-2349 Zm Rz-7 amino stabl a3uscsc a aa9 GccgaaagGCGaGucaaGGuCu accuuca B 262
18731 2384 UACAAGG G CAUCUGG 147 erbB2-2384 Zin Rz 7 amino stabl C8C3as9sau9 GccgaaagGCGaGucaaGGuCu ccuugua B 263
18714 2410 GGAGAAU G UGAAAAU 148 erbB2-2410 Zin Rz-7 ammo stabl a 8__u„3u_3_u,3_uca GccgaaagGCGaGucaaGGuCu auucucc B 264
18732 2497 GUGAUG G CUGGUG 149 erbB2-2497 Zin Rz-6 amino stabl c_ 8a_S_c8__c_3agJ GccgaaagGCGaGucaaGGuCu caucac B 265
18703 2501 UGGCUG G UGUGGG 150 erbB2-2501 Zin Rz-6 amino stabl c_ 3c=3c„8a„8ca GccgaaagGCGaGucaaGGuCu cagcca B 266
18715 2540 GCAUCU G CCUGAC 151 erbB2-2540 Zin Rz-6 amino stabl 98 uscsas99 GccgaaagGCGaGucaaGGuCu agaugc B 267
18733 2563 CAGCUG G UGACAC 152 erbB2-2563 Zin Rz-6 ammo stabl 9sus9susca GccgaaagGCGaGucaaGGuCu cagcug B 268
18734 2571 GACACA G CUUAUG 153 erbB2 2571 Zin Rz-6 ammo stabl c_s_a„sus_,a_sag3 GccgaaagGCGaGucaaGGuCu uguguc B 269
18675 2571 UGACACA G CUUAUGC 154 erbB2-2571 Zm Rz-7 amino stabl 9scsasusaa9 GccgaaagGCGaGucaaGGuCu uguguca B 270
18716 2662 CAGAUU G CCAAGG 155 erbB2-2662 Zin Rz-6 amino stabl c3Csusus99 GccgaaagGCGaGucaaGGuCu aaucug B 271
18704 2675 GGAUGA G CUACCU 156 erbB2-2675 Zin Rz-6 amino stabl as9s9ausa9 GccgaaagGCGaGucaaGGuCu ucaucc B 272
18676 2675 GGGAUGA G CUACCUG 157 erbB2 2675 Zin Rz-7 ammo stabl csa S9s9sua GccgaaagGCGaGucaaGGuCu ucauccc B 273
18735 2738 GUCAAGA G UCCCAAC 158 erbB2 2738 Zin Rz-7 ammo stabl 9susus9S99a GccgaaagGCGaGucaaGGuCu ucuugac B 274
18705 2773 GGGCUG G CUCGGC 159 erbB2-2773 Zm Rz-6 amino stabl 9scscs9sa9 GccgaaagGCGaGucaaGGuCu cagccc B 275
18836 2778 UGGCUCG G CUGCUGG 160 erbB2-2778 Zin Rz-7 amino stabl cscsa S9s ca9 GccgaaagGCGaGucaaGGuCu cgagcca B 276
18694 2781 UCGGCU G CUGGAC 161 erbB2-2781 Zin Rz-6 amino stabl 9suscscsa9 GccgaaagGCGaGucaaGGuCu agccga B 277
18662 2781 CUCGGCU G CUGGACA 162 erbB2 2781 Zin Rz-7 ammo stabl us9suscsca9 GccgaaagGCGaGucaaGGuCu agccgag B 278
18737 2802 GACAGA G UACCAU 163 erbB2-2802 Zm Rz-6 amino stabl 8Us9a9sua GccgaaagGCGaGucaaGGuCu ucuguc B 279
18736 2802 AGACAGA G UACCAUG 164 erbB2-2802 Zin Rz-7 ammo stabl c 3 asusgs9ua GccgaaagGCGaGucaaGGuCu ucugucu B 280
18717 2809 GUACCAU G CAGAUGG 165 erbB2-2809 Zin Rz-7 ammo stabl cscsasuscug GccgaaagGCGaGucaaGGuCu augguac B 281
18738 2819 AUGGGG G CAAGGU 166 erbB2-2819 Zin Rz-6 ammo stabl ascscsu3u9 GccgaaagGCGaGucaaGGuCu ccccau B 282
18706 2819 GAUGGGG G CAAGGUG 167 erbB2-2819 Zin Rz-7 amino stabl csa3cscsuu9 GccgaaagGCGaGucaaGGuCu ccccauc B 283
18695 2887 GAGUGAU G UGUGGAG 168 erbB2-2887 Zin Rz-7 amino stabl C3USC a C a aCa GccgaaagGCGaGucaaGGuCu aucacuc B 284
Table 58
18663 2908 GUGACU G UGUGGG 169 erbB2-2908 Zin Rz-6 amino stabl c„ 3c3cΛ3a„3ca GccgaaagGCGaGucaaGGuCu agucac B 285
18826 2908 UGUGACU G UGUGGGA 170 erbB2-2908 Zm Rz-7 ammo stabl u 8_c_ScS_c_3aca GccgaaagGCGaGucaaGGuCu agucaca B 286
18664 2910 GACUGU G UGGGAG 171 erbB2-2910 Zin Rz-6 amino stabl C8U8C8C a C GccgaaagGCGaGucaaGGuCu acaguc B 287
18650 2910 UGACUGU G UGGGAGC 172 erbB2-2910 Zin Rz-7 ammo stabl 9a csuscscca GccgaaagGCGaGucaaGGuCu acaguca B 288
18677 2916 GUGGGA G CUGAUG 173 erbB2-2916 Zin Rz-6 amino stabl C3a8U8C8a9 GccgaaagGCGaGucaaGGuCu ucccac B 289
18652 2916 UGUGGGA G CUGAUGA 174 erbB2-2916 Zm Rz-7 ammo stabl uscsasusca9 GccgaaagGCGaGucaaGGuCu ucccaca B 290
18707 2932 UUUGGG G CCAAAC 175 erbB2-2932 Zin Rz-6 ammo stabl 93 uβusu899 GccgaaagGCGaGucaaGGuCu cccaaa B 291
18678 2932 UUUUGGG G CCAAACC 176 erbB2-2932 Z Rz-7 amino stabl 9s9s usuau99 GccgaaagGCGaGucaaGGuCu cccaaaa B 292
18719 3025 AUUGAU G UCUACA 177 erbB2-3025 Zin Rz-6 amino stabl us9susas9a GccgaaagGCGaGucaaGGuCu aucaau B 293
18718 3025 CAUUGAU G UCUACAU 178 erbB2-3025 Zm Rz-7 amino stabl asus9ausa9a GccgaaagGCGaGucaaGGuCu aucaaug B 294
18720 3047 UCAAAU G UUGGAU 179 erbB2-3047 Zm Rz-6 ammo stabl a_ 3_u_8_c3__c_3aa GccgaaagGCGaGucaaGGuCu auuuga B 295
18696 3047 GUCAAAU G UUGGAUG 180 erbB2-3047 Zm Rz-7 amino stabl c 8_a_8u_Sc_8caa GccgaaagGCGaGucaaGGuCu auuugac B 296
18739 3087 CCGGGA G UUGGUG 181 erbB2-3087 Zin Rz-6 amino stabl c 3a3c8c8aa GccgaaagGCGaGucaaGGuCu ucccgg B 297
18708 3087 UCCGGGA G UUGGUGU 182 erbB2-3087 Zin Rz-7 amino stabl ascsascscaa GccgaaagGCGaGucaaGGuCu ucccgga B 298
18740 3415 GAAGGG G CUGGCU 183 erbB2-3415 Zin Rz-6 am o stabl as9s cscsa9 GccgaaagGCGaGucaaGGuCu cccuuc B 299
18741 3419 GGGCUG G CUCCGA 184 erbB2-3419 Zin Rz-6 ammo stabl uscs9s9sa9 GccgaaagGCGaGucaaGGuCu cagccc B 300
18837 3419 GGGGCUG G CUCCGAU 185 erbB2-3419 Zm Rz-7 amino stabl asuscs9s9a9 GccgaaagGCGaGucaaGGuCu cagcccc B 301
18709 3437 UUGAUG G UGACCU 186 erbB2-3437 Zin Rz-6 amino stabl as9s9s usca GccgaaagGCGaGucaaGGuCu caucaa B 302
18679 3437 UUUGAUG G UGACCUG 187 erbB2-3437 Zin Rz-7 ammo stabl csas9a9suca GccgaaagGCGaGucaaGGuCu caucaaa B 303
18823 3504 UCUACA G CGGUAC 188 erbB2-3504 Zin Rz-6 amino stabl 9s u 8 a 8 Csc9 GccgaaagGCGaGucaaGGuCu uguaga B 304
18710 3504 CUCUACA G CGGUACA 189 erbB2-3504 Zm Rz-7 amino stabl u S9a u 8 a a cc9 GccgaaagGCGaGucaaGGuCu uguagag B 305
18721 3724 CAAAGAC G UUUUUGC 190 erbB2-3724 Zin Rz-7 amino stabl 9a cs 8a a aa GccgaaagGCGaGucaaGGuCu gucuuug B 306
18834 3808 CCUCCU G CCUUCA 191 erbB2-3808 Zin Rz-6 amino stabl u a9a a a as99 GccgaaagGCGaGucaaGGuCu aggagg B 307
18827 3808 UCCUCCU G CCUUCAG 192 erbB2-3808 Zin Rz-7 amino stabl c 8 u S9sasa99 GccgaaagGCGaGucaaGGuCu aggagga B 308
18824 3996 GGGAAG G CCUGAC 193 erbB2-3996 Zin Rz-6 amino stabl 9susc 8 a S99 GccgaaagGCGaGucaaGGuCu cuuccc B 309
UPPER CASE = RIBO Lower case = 2'-0-methyl C = 2'-deoxy-2'-amino Cytidine s = phosphorothioate B = inverted deoxyabasic
Table 59 Table 59: Human HER2 Class II (zinzyme) Ribozyme and Target Sequece
Pos Substrate Seq ID Ribozyme RzSeq ID
46 GGGCAGCC G CGCGCCCC 310 GGGGCGCG GCCGAAAGGCGAGUCAAGGUCU GGCUGCCC 895
48 GCAGCCGC G CGCCCCUU 311 AAGGGGCG GCCGAAAGGCGAGUCAAGGUCU GCGGCUGC 896
50 AGCCGCGC G CCCCUUCC 312 GGAAGGGG GCCGAAAGGCGAGUCAAGGUCU GCGCGGCU B97
75 CCUUUACU G CGCCGCGC 313 GCGCGGCG GCCGAAAGGCGAGUCAAGGUCU AGUAAAGG 898
77 UUUACUGC G CCGCGCGC 314 GCGCGCGG GCCGAAAGGCGAGUCAAGGUCU GCAGUAAA 899
80 ACUGCGCC G CGCGCCCG 315 CGGGCGCG GCCGAAAGGCGAGUCAAGGUCU GGCGCAGU 900
82 UGCGCCGC G CGCCCGGC 316 GCCGGGCG GCCGAAAGGCGAGUCAAGGUCU GCGGCGCA 901
84 CGCCGCGC G CCCGGCCC 317 GGGCCGGG GCCGAAAGGCGAGUCAAGGUCU GCGCGGCG 902
102 CACCCCUC G CAGCACCC 318 GGGUGCUG GCCGAAAGGCGAGUCAAGGUCU GAGGGGUG 903
112 AGCACCCC G CGCCCCGC 319 GCGGGGCG GCCGAAAGGCGAGUCAAGGUCU GGGGUGCU 904
114 CACCCCGC G CCCCGCGC 320 GCGCGGGG GCCGAAAGGCGAGUCAAGGUCU GCGGGGUG 905
119 cGcσcccc G CGCCCUCC 321 GGAGGGCG GCCGAAAGGCGAGUCAAGGUCU GGGGCGCG 906
121 CGCCCCGC G CCCUCCCA 322 UGGGAGGG GCCGAAAGGCGAGUCAAGGUCU GCGGGGCG 907
163 CCGGAGCC G CAGUGAGC 323 GCUCACUG GCCGAAAGGCGAGUCAAGGUCU GGCUCCGG 908
194 GGCCUUGU G CCGCUGGG 324 CCCAGCGG GCCGAAAGGCGAGUCAAGGUCU ACAAGGCC 909
197 CUUGUGCC G CUGGGGGC 325 GCCCCCAG GCCGAAAGGCGAGUCAAGGUCU GGCACAAG 910
214 UCCUCCUC G CCCUCUUG 326 CAAGAGGG GCCGAAAGGCGAGUCAAGGUCU GAGGAGGA 911
222 GCCCUCUU G CCCCCCGG 327 CCGGGGGG GCCGAAAGGCGAGUCAAGGUCU AAGAGGGC 912
235 CCGGAGCC G CGAGCACC 328 GGUGCUCG GCCGAAAGGCGAGUCAAGGUCU GGCUCCGG 913
251 CCAAGUGU G CACCGGCA 329 UGCCGGUG GCCGAAAGGCGAGUCAAGGUCU ACACUUGG 914
273 AUGAAGCU G CGGCUCCC 330 GGGAGCCG GCCGAAAGGCGAGUCAAGGUCU AGCUUCAU 915
283 GGCUCCCU G CCAGUCCC 331 GGGACUGG GCCGAAAGGCGAGUCAAGGUCU AGGGAGCC 916
309 CUGGACAU G CUCCGCCA 332 UGGCGGAG GCCGAAAGGCGAGUCAAGGUCU AUGUCCAG 917
314 CAUGCUCC G CCACCUCU 333 AGAGGUGG GCCGAAAGGCGAGUCAAGGUCU GGAGCAUG 918
332 CCAGGGCU G CCAGGUGG 334 CCACCUGG GCCGAAAGGCGAGUCAAGGUCU AGCCCUGG 919
342 CAGGUGGU G CAGGGAAA 335 UUUCCCUG GCCGAAAGGCGAGUCAAGGUCU ACCACCUG 920
369 ACCUACCU G CCCACCAA 336 UUGGUGGG GCCGAAAGGCGAGUCAAGGUCU AGGUAGGU 921
379 CCACCAAU G CCAGCCUG 337 CAGGCUGG GCCGAAAGGCGAGUCAAGGUCU AUUGGUGG 922
396 UCCUUCCU G CAGGAUAU 338 AUAUCCUG GCCGAAAGGCGAGUCAAGGUCU AGGAAGGA 923
414 CAGGAGGU G CAGGGCUA 339 UAGCCCUG GCCGAAAGGCGAGUCAAGGUCU ACCUCCUG 924
426 GGCUACGU G CUCAUCGC 340 GCGAUGAG GCCGAAAGGCGAGUCAAGGUCU ACGUAGCC 925
433 UGCUCAUC G CUCACAAC 341 GUUGUGAG GCCGAAAGGCGAGUCAAGGUCU GAUGAGCA 926
462 GUCCCACU G CAGAGGCU 342 AGCCUCUG GCCGAAAGGCGAGUCAAGGUCU AGUGGGAC 927
471 CAGAGGCU G CGGAUUGU 343 ACAAUCCG GCCGAAAGGCGAGUCAAGGUCU AGCCUCUG 928
480 CGGAUUGU G CGAGGCAC 344 GUGCCUCG GCCGAAAGGCGAGUCAAGGUCU ACAAUCCG 929
511 ACAACUAU G CCCUGGCC 345 GGCCAGGG GCCGAAAGGCGAGUCAAGGUCU AUAGUUGU 930
522 CUGGCCGU G CUAGACAA 346 UUGUCUAG GCCGAAAGGCGAGUCAAGGUCU ACGGCCAG 931
540 GGAGACCC G CUGAACAA 347 UUGUUCAG GCCGAAAGGCGAGUCAAGGUCU GGGUCUCC 932
585 GGAGGCCU G CGGGAGCU 348 AGCUCCCG GCCGAAAGGCGAGUCAAGGUCU AGGCCUCC 933
594 CGGGAGCU G CAGCUUCG 349 CGAAGCUG GCCGAAAGGCGAGUCAAGGUCU AGCUCCCG 934
659 CCAGCUCU G CUACCAGG 350 CCUGGUAG GCCGAAAGGCGAGUCAAGGUCU AGAGCUGG 935
737 CACCAACC G CUCUCGGG 351 CCCGAGAG GCCGAAAGGCGAGUCAAGGUCU GGUUGGUG 936
749 UCGGGCCU G CCACCCCU 352 AGGGGUGG GCCGAAAGGCGAGUCAAGGUCU AGGCCCGA 937
Table 59
782 GGGCUCCC G CUGCUGGG 353 CCCAGCAG GCCGAAAGGCGAGUCAAGGUCU GGGAGCCC 938
785 CUCCCGCU G CUGGGGAG 354 CUCCCCAG GCCGAAAGGCGAGUCAAGGUCU AGCGGGAG 939
822 AGCCUGAC G CGCACUGU 355 ACAGUGCG GCCGAAAGGCGAGUCAAGGUCU GUCAGGCU 940
824 CCUGACGC G CACUGUCU 356 AGACAGUG GCCGAAAGGCGAGUCAAGGUCU GCGUCAGG 941
835 CUGUCUGU G CCGGUGGC 357 GCCACCGG GCCGAAAGGCGAGUCAAGGUCU ACAGACAG 942
847 GUGGCUGU G CCCGCUGC 358 GCAGCGGG GCCGAAAGGCGAGUCAAGGUCU ACAGCCAC 943
851 CUGUGCCC G CUGCAAGG 359 CCUUGCAG GCCGAAAGGCGAGUCAAGGUCU GGGCACAG 944
854 UGCCCGCU G CAAGGGGC 360 GCCCCUUG GCCGAAAGGCGAGUCAAGGUCU AGCGGGCA 945
867 GGGCCACU G CCCACUGA 361 UCAGUGGG GCCGAAAGGCGAGUCAAGGUCU AGUGGCCC 946
878 CACUGACU G CUGCCAUG 362 CAUGGCAG GCCGAAAGGCGAGUCAAGGUCU AGUCAGUG 947
881 UGACUGCU G CCAUGAGC 363 GCUCAUGG GCCGAAAGGCGAGUCAAGGUCU AGCAGUCA 948
895 AGCAGUGU G CUGCCGGC 364 GCCGGCAG GCCGAAAGGCGAGUCAAGGUCU ACACUGCU 949
898 AGUGUGCU G CCGGCUGC 365 GCAGCCGG GCCGAAAGGCGAGUCAAGGUCU AGCACACU 950
905 UGCCGGCU G CACGGGCC 366 GGCCCGUG GCCGAAAGGCGAGUCAAGGUCU AGCCGGCA 951
929 CUCUGACU G CCUGGCCU 367 AGGCCAGG GCCGAAAGGCGAGUCAAGGUCU AGUCAGAG 952
938 CCUGGCCU G CCUCCACU 368 AGUGGAGG GCCGAAAGGCGAGUCAAGGUCU AGGCCAGG 953
972 UGUGAGCU G CACUGCCC 369 GGGCAGUG GCCGAAAGGCGAGUCAAGGUCU AGCUCACA 954
977 GCUGCACU G CCCAGCCC 370 GGGCUGGG GCCGAAAGGCGAGUCAAGGUCU AGUGCAGC 955
1020 GAGUCCAU G CCCAAUCC 371 GGAUUGGG GCCGAAAGGCGAGUCAAGGUCU AUGGACUC 956
1051 CAUUCGGC G CCAGCUGU 372 ACAGCUGG GCCGAAAGGCGAGUCAAGGUCU GCCGAAUG 957
1066 GUGUGACU G CCUGUCCC 373 GGGACAGG GCCGAAAGGCGAGUCAAGGUCU AGUCACAC 958
1106 GGGAUCCU G CACCCUCG 374 CGAGGGUG GCCGAAAGGCGAGUCAAGGUCU AGGAUCCC 959
1118 CCUCGUCU G CCCCCUGC 375 GCAGGGGG GCCGAAAGGCGAGUCAAGGUCU AGACGAGG 960
1125 UGCCCCCU G CACAACCA 376 UGGUUGUG GCCGAAAGGCGAGUCAAGGUCU AGGGGGCA 961
1175 UGAGAAGU G CAGCAAGC 377 GCUUGCUG GCCGAAAGGCGAGUCAAGGUCU ACUUCUCA 962
1189 AGCCCUGU G CCCGAGUG 378 CACUCGGG GCCGAAAGGCGAGUCAAGGUCU ACAGGGCU 963
1199 CCGAGUGU G CUAUGGUC 379 GACCAUAG GCCGAAAGGCGAGUCAAGGUCU ACACUCGG 964
1224 GAGCACUU G CGAGAGGU 380 ACCUCUCG GCCGAAAGGCGAGUCAAGGUCU AAGUGCUC 965
1249 UUACCAGU G CCAAUAUC 381 GAUAUUGG GCCGAAAGGCGAGUCAAGGUCU ACUGGUAA 966
1267 AGGAGUUU G CUGGCUGC 382 GCAGCCAG GCCGAAAGGCGAGUCAAGGUCU AAACUCCU 967
1274 UGCUGGCU G CAAGAAGA 383 UCUUCUUG GCCGAAAGGCGAGUCAAGGUCU AGCCAGCA 968
1305 GCAUUUCU G CCGGAGAG 384 CUCUCCGG GCCGAAAGGCGAGUCAAGGUCU AGAAAUGC 969
1342 CCAACACU G CCCCGCUC 385 GAGCGGGG GCCGAAAGGCGAGUCAAGGUCU AGUGUUGG 970
1347 ACUGCCCC G CUCCAGCC 386 GGCUGGAG GCCGAAAGGCGAGUCAAGGUCU GGGGCAGU 971
1431 GACAGCCU G CCUGACCU 387 AGGUCAGG GCCGAAAGGCGAGUCAAGGUCU AGGCUGUC 972
1458 CAGAACCU G CAAGUAAU 388 AUUACUUG GCCGAAAGGCGAGUCAAGGUCU AGGUUCUG 973
1482 CGAAUUCU G CACAAUGG 389 CCAUUGUG GCCGAAAGGCGAGUCAAGGUCU AGAAUUCG 974
1492 ACAAUGGC G CCUACUCG 390 CGAGUAGG GCCGAAAGGCGAGUCAAGGUCU GCCAUUGU 975
1500 GCCUACUC G CUGACCCU 391 AGGGUCAG GCCGAAAGGCGAGUCAAGGUCU GAGUAGGC 976
1509 CUGACCCU G CAAGGGCU 392 AGCCCUUG GCCGAAAGGCGAGUCAAGGUCU AGGGUCAG 977
1539 CUGGGGCU G CGCUCACU 393 AGUGAGCG GCCGAAAGGCGAGUCAAGGUCU AGCCCCAG 978
1541 GGGGCUGC G CUCACUGA 394 UCAGUGAG GCCGAAAGGCGAGUCAAGGUCU GCAGCCCC 979
1598 CCACCUCU G CUUCGUGC 395 GCACGAAG GCCGAAAGGCGAGUCAAGGUCU AGAGGUGG 980
1605 UGCUUCGU G CACACGGU 396 ACCGUGUG GCCGAAAGGCGAGUCAAGGUCU ACGAAGCA 981
1614 CACAcσσu G CCCUGGGA 397 UCCCAGGG GCCGAAAGGCGAGUCAAGGUCU ACCGUGUG 982
1641 CGGAACCC G CACCAAGC 398 GCUUGGUG GCCGAAAGGCGAGUCAAGGUCU GGGUUCCG 983
1653 CAAGCUCU G CUCCACAC 399 GUGUGGAG GCCGAAAGGCGAGUCAAGGUCU AGAGCUUG 984
Table 59
1663 UCCACACU G CCAACCGG 400 CCGGUUGG GCCGAAAGGCGAGUCAAGGUCU AGUGUGGA 985
1706 CCUGGCCU G CCACCAGC 401 GCUGGUGG GCCGAAAGGCGAGUCAAGGUCU AGGCCAGG 986
1718 CCAGCUGU G CGCCCGAG 402 CUCGGGCG GCCGAAAGGCGAGUCAAGGUCU ACAGCUGG 987
1720 AGCUGUGC G CCCGAGGG 403 CCCUCGGG GCCGAAAGGCGAGUCAAGGUCU GCACAGCU 988
1733 AGGGCACU G CUGGGGUC 404 GACCCCAG GCCGAAAGGCGAGUCAAGGUCU AGUGCCCU 989
1766 UGUCAACU G CAGCCAGU 405 ACUGGCUG GCCGAAAGGCGAGUCAAGGUCU AGUUGACA 990
1793 CCAGGAGU G CGUGGAGG 406 CCUCCACG GCCGAAAGGCGAGUCAAGGUCU ACUCCUGG 991
1805 GGAGGAAU G CCGAGUAC 407 GUACUCGG GCCGAAAGGCGAGUCAAGGUCU AUUCCUCC 992
1815 CGAGUACU G CAGGGGCU 408 AGCCCCUG GCCGAAAGGCGAGUCAAGGUCU AGUACUCG 993
1843 AUGUGAAU G CCAGGCAC 409 GUGCCUGG GCCGAAAGGCGAGUCAAGGUCU AUUCACAU 994
1857 CACUGUUU G CCGUGCCA 410 UGGCACGG GCCGAAAGGCGAGUCAAGGUCU AAACAGUG 995
1862 UUUGCCGU G CCACCCUG 411 CAGGGUGG GCCGAAAGGCGAGUCAAGGUCU ACGGCAAA 996
1936 UGGCCUGU G CCCACUAU 412 AUAGUGGG GCCGAAAGGCGAGUCAAGGUCU ACAGGCCA 997
1961 UCCCUUCU G CGUGGCCC 413 GGGCCACG GCCGAAAGGCGAGUCAAGGUCU AGAAGGGA 998
1970 CGUGGCCC G CUGCCCCA 414 UGGGGCAG GCCGAAAGGCGAGUCAAGGUCU GGGCCACG 999
1973 GGCCCGCU G CCCCAGCG 415 CGCUGGGG GCCGAAAGGCGAGUCAAGGUCU AGCGGGCC 1000
2007 UCCUACAU G CCCAUCUG 416 CAGAUGGG GCCGAAAGGCGAGUCAAGGUCU AUGUAGGA 1001
2038 AGGAGGGC G CAUGCCAG 417 CUGGCAUG GCCGAAAGGCGAGUCAAGGUCU GCCCUCCU 1002
2042 GGGCGCAU G CCAGCCUU 418 AAGGCUGG GCCGAAAGGCGAGUCAAGGUCU AUGCGCCC 1003
2051 CCAGCCUU G CCCCAUCA 419 UGAUGGGG GCCGAAAGGCGAGUCAAGGUCU AAGGCUGG 1004
2063 CAUCAACU G CACCCACU 420 AGUGGGUG GCCGAAAGGCGAGUCAAGGUCU AGUUGAUG 1005
2099 CAAGGGCU G CCCCGCCG 421 CGGCGGGG GCCGAAAGGCGAGUCAAGGUCU AGCCCUUG 1006
2104 GCUGCCCC G CCGAGCAG 422 CUGCUCGG GCCGAAAGGCGAGUCAAGGUCU GGGGCAGC 1007
2143 UCAUCUCU G CGGUGGUU 423 AACCACCG GCCGAAAGGCGAGUCAAGGUCU AGAGAUGA 1008
2160 GGCAUUCU G CUGGUCGU 424 ACGACCAG GCCGAAAGGCGAGUCAAGGUCU AGAAUGCC 1009
2235 UACACGAU G CGGAGACU 425 AGUCUCCG GCCGAAAGGCGAGUCAAGGUCU AUCGUGUA 1010
2244 CGGAGACU G CUGCAGGA 426 UCCUGCAG GCCGAAAGGCGAGUCAAGGUCU AGUCUCCG 1011
2247 AGACUGCU G CAGGAAAC 427 GUUUCCUG GCCGAAAGGCGAGUCAAGGUCU AGCAGUCU 1012
2271 GUGGAGCC G CUGACACC 428 GGUGUCAG GCCGAAAGGCGAGUCAAGGUCU GGCUCCAC 1013
2292 GGAGCGAU G CCCAACCA 429 UGGUUGGG GCCGAAAGGCGAGUCAAGGUCU AUCGCUCC 1014
2304 AACCAGGC G CAGAUGCG 430 CGCAUCUG GCCGAAAGGCGAGUCAAGGUCU GCCUGGUU 1015
2310 GCGCAGAU G CGGAUCCU 431 AGGAUCCG GCCGAAAGGCGAGUCAAGGUCU AUCUGCGC 1016
2349 GUGAAGGU G CUUGGAUC 432 GAUCCAAG GCCGAAAGGCGAGUCAAGGUCU ACCUUCAC 1017
2362 GAUCUGGC G CUUUUGGC 433 GCCAAAAG GCCGAAAGGCGAGUCAAGGUCU GCCAGAUC 1018
2525 UGUCUCCC G CCUUCUGG 434 CCAGAAGG GCCGAAAGGCGAGUCAAGGUCU GGGAGACA 1019
2540 GGGCAUCU G CCUGACAU 435 AUGUCAGG GCCGAAAGGCGAGUCAAGGUCU AGAUGCCC 1020
2556 UCCACGGU G CAGCUGGU 436 ACCAGCUG GCCGAAAGGCGAGUCAAGGUCU ACCGUGGA 1021
2577 CAGCUUAU G CCCUAUGG 437 CCAUAGGG GCCGAAAGGCGAGUCAAGGUCU AUAAGCUG 1022
2588 CUAUGGCU G CCUCUUAG 438 CUAAGAGG GCCGAAAGGCGAGUCAAGGUCU AGCCAUAG 1023
2615 GGAAAACC G CGGACGCC 439 GGCGUCCG GCCGAAAGGCGAGUCAAGGUCU GGUUUUCC 1024
2621 CCGCGGAC G CCUGGGCU 440 AGCCCAGG GCCGAAAGGCGAGUCAAGGUCU GUCCGCGG 1025
2640 CAGGACCU G CUGAACUG 441 CAGUUCAG GCCGAAAGGCGAGUCAAGGUCU AGGUCCUG 1026
2655 UGGUGUAU G CAGAUUGC 442 GCAAUCUG GCCGAAAGGCGAGUCAAGGUCU AUACACCA 1027
2662 UGCAGAUU G CCAAGGGG 443 CCCCUUGG GCCGAAAGGCGAGUCAAGGUCU AAUCUGCA 1028
2691 GAGGAUGU G CGGCUCGU 444 ACGAGCCG GCCGAAAGGCGAGUCAAGGUCU ACAUCCUC 1029
2716 ACUUGGCC G CUCGGAAC 445 GUUCCGAG GCCGAAAGGCGAGUCAAGGUCU GGCCAAGU 1030
2727 CGGAACGU G CUGGUCAA 446 UUGACCAG GCCGAAAGGCGAGUCAAGGUCU ACGUUCCG 1031
Table 59
2781 GCUCGGCU G CUGGACAU 447 AUGUCCAG GCCGAAAGGCGAGUCAAGGUCU AGCCGAGC 1032
2809 AGUACCAU G CAGAUGGG 448 CCCAUCUG GCCGAAAGGCGAGUCAAGGUCU AUGGUACU 1033
2826 GGCAAGGU G CCCAUCAA 449 UUGAUGGG GCCGAAAGGCGAGUCAAGGUCU ACCUUGCC 1034
2844 UGGAUGGC G CUGGAGUC 450 GACUCCAG GCCGAAAGGCGAGUCAAGGUCU GCCAUCCA 1035
2861 CAUUCUCC G CCGGCGGU 451 ACCGCCGG GCCGAAAGGCGAGUCAAGGUCU GGAGAAUG 1036
2976 CCUGACCU G CUGGAAAA 452 UUUUCCAG GCCGAAAGGCGAGUCAAGGUCU AGGUCAGG 1037
2997 GAGCGGCU G CCCCAGCC 453 GGCUGGGG GCCGAAAGGCGAGUCAAGGUCU AGCCGCUC 1038
3014 CCCCAUCU G CACCAUUG 454 CAAUGGUG GCCGAAAGGCGAGUCAAGGUCU AGAUGGGG 1039
3107 AUUCUCCC G CAUGGCCA 455 UGGCCAUG GCCGAAAGGCGAGUCAAGGUCU GGGAGAAU 1040
3128 CCCCCAGC G CUUUGUGG 456 CCACAAAG GCCGAAAGGCGAGUCAAGGUCU GCUGGGGG 1041
3191 CUUCUACC G CUCACUGC 457 GCAGUGAG GCCGAAAGGCGAGUCAAGGUCU GGUAGAAG 1042
3198 CGCUCACU G CUGGAGGA 458 UCCUCCAG GCCGAAAGGCGAGUCAAGGUCU AGUGAGCG 1043
3232 UGGUGGAU G CUGAGGAG 459 CUCCUCAG GCCGAAAGGCGAGUCAAGGUCU AUCCACCA 1044
3280 CAGACCCU G CCCCGGGC 460 GCCCGGGG GCCGAAAGGCGAGUCAAGGUCU AGGGUCUG 1045
3289 CCCCGGGC G CUGGGGGC 461 GCCCCCAG GCCGAAAGGCGAGUCAAGGUCU GCCCGGGG 1046
3317 CAGGCACC G CAGCUCAU 462 AUGAGCUG GCCGAAAGGCGAGUCAAGGUCU GGUGCCUG 1047
3468 AAGGGGCU G CAAAGCCU 463 AGGCUUUG GCCGAAAGGCGAGUCAAGGUCU AGCCCCUU 1048
3534 GUACCCCU G CCCUCUGA 464 UCAGAGGG GCCGAAAGGCGAGUCAAGGUCU AGGGGUAC 1049
3559 GCUACGUU G CCCCCCUG 465 CAGGGGGG GCCGAAAGGCGAGUCAAGGUCU AACGUAGC 1050
3572 CCUGACCU G CAGCCCCC 466 GGGGGCUG GCCGAAAGGCGAGUCAAGGUCU AGGUCAGG 1051
3627 CCCCCUUC G CCCCGAGA 467 UCUCGGGG GCCGAAAGGCGAGUCAAGGUCU GAAGGGGG 1052
3645 GGCCCUCU G CCUGCUGC 468 GCAGCAGG GCCGAAAGGCGAGUCAAGGUCU AGAGGGCC 1053
3649 CUCUGCCU G CUGCCCGA 469 UCGGGCAG GCCGAAAGGCGAGUCAAGGUCU AGGCAGAG 1054
3652 UGCCUGCU G CCCGACCU 470 AGGUCGGG GCCGAAAGGCGAGUCAAGGUCU AGCAGGCA 1055
3661 CCCGACCU G CUGGUGCC 471 GGCACCAG GCCGAAAGGCGAGUCAAGGUCU AGGUCGGG 1056
3667 CUGCUGGU G CCACUCUG 472 CAGAGUGG GCCGAAAGGCGAGUCAAGGUCU ACCAGCAG 1057
3730 ACGUUUUU G CCUUUGGG 473 CCCAAAGG GCCGAAAGGCGAGUCAAGGUCU AAAAACGU 1058
3742 UUGGGGGU G CCGUGGAG 474 CUCCACGG GCCGAAAGGCGAGUCAAGGUCU ACCCCCAA 1059
3784 GAGGAGCU G CCCCUCAG 475 CUGAGGGG GCCGAAAGGCGAGUCAAGGUCU AGCUCCUC 1060
3808 CUCCUCCU G CCUUCAGC 476 GCUGAAGG GCCGAAAGGCGAGUCAAGGUCU AGGAGGAG 1061
3933 CUGGACGU G CCAGUGUG 477 CACACUGG GCCGAAAGGCGAGUCAAGGUCU ACGUCCAG 1062
3960 CCAAGUCC G CAGAAGCC 478 GGCUUCUG GCCGAAAGGCGAGUCAAGGUCU GGACUUGG 1063
4007 UGACUUCU G CUGGCAUC 479 GAUGCCAG GCCGAAAGGCGAGUCAAGGUCU AGAAGUCA 1064
4056 GGGAACCU G CCAUGCCA 480 UGGCAUGG GCCGAAAGGCGAGUCAAGGUCU AGGUUCCC 1065
4061 CCUGCCAU G CCAGGAAC 481 GUUCCUGG GCCGAAAGGCGAGUCAAGGUCU AUGGCAGG 1066
4094 UCCUUCCU G CUUGAGUU 482 AACUCAAG GCCGAAAGGCGAGUCAAGGUCU AGGAAGGA 1067
4179 GAGGCCCU G CCCAAUGA 483 UCAUUGGG GCCGAAAGGCGAGUCAAGGUCU AGGGCCUC 1068
4208 CAGUGGAU G CCACAGCC 484 GGCUGUGG GCCGAAAGGCGAGUCAAGGUCU AUCCACUG 1069
4351 CUAGUACU G CCCCCCAU 485 AUGGGGGG GCCGAAAGGCGAGUCAAGGUCU AGUACUAG 1070
4406 UACAGAGU G CUUUUCUG 486 CAGAAAAG GCCGAAAGGCGAGUCAAGGUCU ACUCUGUA 1071
192 GCGGCCUU G UGCCGCUG 487 CAGCGGCA GCCGAAAGGCGAGUCAAGGUCU AAGGCCGC 1072
249 ACCCAAGU G UGCACCGG 488 CCGGUGCA GCCGAAAGGCGAGUCAAGGUCU ACUUGGGU 1073
387 GCCAGCCU G UCCUUCCU 489 AGGAAGGA GCCGAAAGGCGAGUCAAGGUCU AGGCUGGC 1074
478 UGCGGAUU G UGCGAGGC 490 GCCUCGCA GCCGAAAGGCGAGUCAAGGUCU AAUCCGCA 1075
559 CCACCCCU G UCACAGGG 491 CCCUGUGA GCCGAAAGGCGAGUCAAGGUCU AGGGGUGG 1076
678 ACGAUUUU G UGGAAGGA 492 UCCUUCCA GCCGAAAGGCGAGUCAAGGUCU AAAAUCGU 1077
758 CCACCCCU G UUCUCCGA 493 UCGGAGAA GCCGAAAGGCGAGUCAAGGUCU AGGGGUGG 1078
Table 59
768 UCUCCGAU G UGUAAGGG 494 CCCUUACA GCCGAAAGGCGAGUCAAGGUCU AUCGGAGA 1079
770 UCCGAUGU G UAAGGGCU 495 AGCCCUUA GCCGAAAGGCGAGUCAAGGUCU ACAUCGGA 1080
809 UGAGGAUU G UCAGAGCC 496 GGCUCUGA GCCGAAAGGCGAGUCAAGGUCU AAUCCUCA 1081
829 CGCGCACU G UCUGUGCC 497 GGCACAGA GCCGAAAGGCGAGUCAAGGUCU AGUGCGCG 1082
833 CACUGUCU G UGCCGGUG 498 CACCGGCA GCCGAAAGGCGAGUCAAGGUCU AGACAGUG 1083
845 CGGUGGCU G UGCCCGCU 499 AGCGGGCA GCCGAAAGGCGAGUCAAGGUCU AGCCACCG 1084
893 UGAGCAGU G UGCUGCCG 500 CGGCAGCA GCCGAAAGGCGAGUCAAGGUCU ACUGCUCA 1085
965 UGGCAUCU G UGAGCUGC 501 GCAGCUCA GCCGAAAGGCGAGUCAAGGUCU AGAUGCCA 1086
1058 CGCCAGCU G UGUGACUG 502 CAGUCACA GCCGAAAGGCGAGUCAAGGUCU AGCUGGCG 1087
1060 CCAGCUGU G UGACUGCC 503 GGCAGUCA GCCGAAAGGCGAGUCAAGGUCU ACAGCUGG 1088
1070 GACUGCCU G UCCCUACA 504 UGUAGGGA GCCGAAAGGCGAGUCAAGGUCU AGGCAGUC 1089
1166 ACAGCGGU G UGAGAAGU 505 ACUUCUCA GCCGAAAGGCGAGUCAAGGUCU ACCGCUGU 1090
1187 CAAGCCCU G UGCCCGAG 506 CUCGGGCA GCCGAAAGGCGAGUCAAGGUCU AGGGCUUG 1091
1197 GCCCGAGU G UGCUAUGG 507 CCAUAGCA GCCGAAAGGCGAGUCAAGGUCU ACUCGGGC 1092
1371 CUCCAAGU G UUUGAGAC 508 GUCUCAAA GCCGAAAGGCGAGUCAAGGUCU ACUUGGAG 1093
1685 GGACGAGU G UGUGGGCG 509 CGCCCACA GCCGAAAGGCGAGUCAAGGUCU ACUCGUCC 1094
1687 ACGAGUGU G UGGGCGAG 510 CUCGCCCA GCCGAAAGGCGAGUCAAGGUCU ACACUCGU 1095
1716 CACCAGCU G UGCGCCCG 511 CGGGCGCA GCCGAAAGGCGAGUCAAGGUCU AGCUGGUG 1096
1757 CACCCAGU G UGUCAACU 512 AGUUGACA GCCGAAAGGCGAGUCAAGGUCU ACUGGGUG 1097
1759 CCCAGUGU G UCAACUGC 513 GCAGUUGA GCCGAAAGGCGAGUCAAGGUCU ACACUGGG 1098
1837 GGGAGUAU G UGAAUGCC 514 GGCAUUCA GCCGAAAGGCGAGUCAAGGUCU AUACUCCC 1099
1853 CAGGCACU G UUUGCCGU 515 ACGGCAAA GCCGAAAGGCGAGUCAAGGUCU AGUGCCUG 1100
1874 CCCUGAGU G UCAGCCCC 516 GGGGCUGA GCCGAAAGGCGAGUCAAGGUCU ACUCAGGG 1101
1901 AGUGACCU G UUUUGGAC 517 GUCCAAAA GCCGAAAGGCGAGUCAAGGUCU AGGUCACU 1102
1925 UGACCAGU G UGUGGCCU 518 AGGCCACA GCCGAAAGGCGAGUCAAGGUCU ACUGGUCA 1103
1927 ACCAGUGU G UGGCCUGU 519 ACAGGCCA GCCGAAAGGCGAGUCAAGGUCU ACACUGGU 1104
1934 UGUGGCCU G UGCCCACU 520 AGUGGGCA GCCGAAAGGCGAGUCAAGGUCU AGGCCACA 1105
1984 CCAGCGGU G UGAAACCU 521 AGGUUUCA GCCGAAAGGCGAGUCAAGGUCU ACCGCUGG 1106
2075 CCACUCCU G UGUGGACC 522 GGUCCACA GCCGAAAGGCGAGUCAAGGUCU AGGAGUGG 1107
2077 ACUCCUGU G UGGACCUG 523 CAGGUCCA GCCGAAAGGCGAGUCAAGGUCU ACAGGAGU 1108
2410 GGGAGAAU G UGAAAAUU 524 AAUUUUCA GCCGAAAGGCGAGUCAAGGUCU AUUCUCCC 1109
2436 AUCAAAGU G UUGAGGGA 525 UCCCUCAA GCCGAAAGGCGAGUCAAGGUCU ACUUUGAU 1110
2503 UGGCUGGU G UGGGCUCC 526 GGAGCCCA GCCGAAAGGCGAGUCAAGGUCU ACCAGCCA llll
2518 CCCCAUAU G UCUCCCGC 527 GCGGGAGA GCCGAAAGGCGAGUCAAGGUCU AUAUGGGG 1112
2602 UAGACCAU G UCCGGGAA 528 UUCCCGGA GCCGAAAGGCGAGUCAAGGUCU AUGGUCUA 1113
2651 GAACUGGU G UAUGCAGA 529 UCUGCAUA GCCGAAAGGCGAGUCAAGGUCU ACCAGUUC 1114
2689 UGGAGGAU G UGCGGCUC 530 GAGCCGCA GCCGAAAGGCGAGUCAAGGUCU AUCCUCCA 1115
2749 CCAACCAU G UCAAAAUU 531 AAUUUUGA GCCGAAAGGCGAGUCAAGGUCU AUGGUUGG 1116
2887 AGAGUGAU G UGUGGAGU 532 ACUCCACA GCCGAAAGGCGAGUCAAGGUCU AUCACUCU 1117
2889 AGUGAUGU G UGGAGUUA 533 UAACUCCA GCCGAAAGGCGAGUCAAGGUCU ACAUCACU 1118
2902 GUUAUGGU G UGACUGUG 534 CACAGUCA GCCGAAAGGCGAGUCAAGGUCU ACCAUAAC 1119
2908 GUGUGACU G UGUGGGAG 535 CUCCCACA GCCGAAAGGCGAGUCAAGGUCU AGUCACAC 1120
2910 GUGACUGU G UGGGAGCU 536 AGCUCCCA GCCGAAAGGCGAGUCAAGGUCU ACAGUCAC 1121
3025 CCAUUGAU G UCUACAUG 537 CAUGUAGA GCCGAAAGGCGAGUCAAGGUCU AUCAAUGG 1122
3047 GGUCAAAU G UUGGAUGA 538 UCAUCCAA GCCGAAAGGCGAGUCAAGGUCU AUUUGACC 1123
3068 CUCUGAAU G UCGGCCAA 539 UUGGCCGA GCCGAAAGGCGAGUCAAGGUCU AUUCAGAG 1124
3093 GAGUUGGU G UCUGAAUU 540 AAUUCAGA GCCGAAAGGCGAGUCAAGGUCU ACCAACUC 1125
Table 59
3133 AGCGCUUU G UGGUCAUC 541 GAUGACCA GCCGAAAGGCGAGUCAAGGUCU AAAGCGCU 1126
3269 CUUCUUCU G UCCAGACC 542 GGUCUGGA GCCGAAAGGCGAGUCAAGGUCU AGAAGAAG 1127
3427 GCUCCGAU G UAUUUGAU 543 AUCAAAUA GCCGAAAGGCGAGUCAAGGUCU AUCGGAGC 1128
3592 CUGAAUAU G UGAACCAG 544 CUGGUUCA GCCGAAAGGCGAGUCAAGGUCU AUAUUCAG 1129
3607 AGCCAGAU G UUCGGCCC 545 GGGCCGAA GCCGAAAGGCGAGUCAAGGUCU AUCUGGCU 1130
3939 GUGCCAGU G UGAACCAG 546 CUGGUUCA GCCGAAAGGCGAGUCAAGGUCU ACUGGCAC 1131
3974 GCCCUGAU G UGUCCUCA 547 UGAGGACA GCCGAAAGGCGAGUCAAGGUCU AUCAGGGC 1132
3976 CCUGAUGU G UCCUCAGG 548 CCUGAGGA GCCGAAAGGCGAGUCAAGGUCU ACAUCAGG 1133
4072 AGGAACCU G UCCUAAGG 549 CCUUAGGA GCCGAAAGGCGAGUCAAGGUCU AGGUUCCU 1134
4162 GAGUCUUU G UGGAUUCU 550 AGAAUCCA GCCGAAAGGCGAGUCAAGGUCU AAAGACUC 1135
4300 AAGGGAGU G UCUAAGAA 551 UUCUUAGA GCCGAAAGGCGAGUCAAGGUCU ACUCCCUU 1136
4332 CAGAGACU G UCCCUGAA 552 UUCAGGGA GCCGAAAGGCGAGUCAAGGUCU AGUCUCUG 1137
4380 GCAAUGGU G UCAGUAUC 553 GAUACUGA GCCGAAAGGCGAGUCAAGGUCU ACCAUUGC 1138
4397 CAGGCUUU G UACAGAGU 554 ACUCUGUA GCCGAAAGGCGAGUCAAGGUCU AAAGCCUG 1139
4414 GCUUUUCU G UUUAGUUU 555 AAACUAAA GCCGAAAGGCGAGUCAAGGUCU AGAAAAGC 1140
4434 CUUUUUUU G UUUUGUUU 556 AAACAAAA GCCGAAAGGCGAGUCAAGGUCU AAAAAAAG 1141
4439 UUUGUUUU G UUUUUUUA 557 UAAAAAAA GCCGAAAGGCGAGUCAAGGUCU AAAACAAA 1142
9 AAGGGGAG G UAACCCUG 558 CAGGGUUA GCCGAAAGGCGAGUCAAGGUCU CUCCCCUU 1143
18 UAACCCUG G CCCCUUUG 559 CAAAGGGG GCCGAAAGGCGAGUCAAGGUCU CAGGGUUA 1144
27 CCCCUUUG G UCGGGGCC 560 GGCCCCGA GCCGAAAGGCGAGUCAAGGUCU CAAAGGGG 1145
33 UGGUCGGG G CCCCGGGC 561 GCCCGGGG GCCGAAAGGCGAGUCAAGGUCU CCCGACCA 1146
40 GGCCCCGG G CAGCCGCG 562 CGCGGCUG GCCGAAAGGCGAGUCAAGGUCU CCGGGGCC 1147
43 CCCGGGCA G CCGCGCGC 563 GCGCGCGG GCCGAAAGGCGAGUCAAGGUCU UGCCCGGG 1148
65 CCCACGGG G CCCUUUAC 564 GUAAAGGG GCCGAAAGGCGAGUCAAGGUCU CCCGUGGG 1149
89 CGCGCCCG G CCCCCACC 565 GGUGGGGG GCCGAAAGGCGAGUCAAGGUCU CGGGCGCG 1150
105 CCCUCGCA G CACCCCGC 566 GCGGGGUG GCCGAAAGGCGAGUCAAGGUCU UGCGAGGG 1151
130 CCCUCCCA G CCGGGUCC 567 GGACCCGG GCCGAAAGGCGAGUCAAGGUCU UGGGAGGG 1152
135 CCAGCCGG G UCCAGCCG 568 CGGCUGGA GCCGAAAGGCGAGUCAAGGUCU CCGGCUGG 1153
140 CGGGUCCA G CCGGAGCC 569 GGCUCCGG GCCGAAAGGCGAGUCAAGGUCU UGGACCCG 1154
146 CAGCCGGA G CCAUGGGG 570 CCCCAUGG GCCGAAAGGCGAGUCAAGGUCU UCCGGCUG 1155
154 GCCAUGGG G CCGGAGCC 571 GGCUCCGG GCCGAAAGGCGAGUCAAGGUCU CCCAUGGC 1156
160 GGGCCGGA G CCGCAGUG 572 CACUGCGG GCCGAAAGGCGAGUCAAGGUCU UCCGGCCC 1157
166 GAGCCGCA G UGAGCACC 573 GGUGCUCA GCCGAAAGGCGAGUCAAGGUCU UGCGGCUC 1158
170 CGCAGUGA G CACCAUGG 574 CCAUGGUG GCCGAAAGGCGAGUCAAGGUCU UCACUGCG 1159
180 ACCAUGGA G CUGGCGGC 575 GCCGCCAG GCCGAAAGGCGAGUCAAGGUCU UCCAUGGU 1160
184 UGGAGCUG G CGGCCUUG 576 CAAGGCCG GCCGAAAGGCGAGUCAAGGUCU CAGCUCCA 1161
187 AGCUGGCG G CCUUGUGC 577 GCACAAGG GCCGAAAGGCGAGUCAAGGUCU CGCCAGCU 1162
204 CGCUGGGG G CUCCUCCU 578 AGGAGGAG GCCGAAAGGCGAGUCAAGGUCU CCCCAGCG 1163
232 CCCCCGGA G CCGCGAGC 579 GCUCGCGG GCCGAAAGGCGAGUCAAGGUCU UCCGGGGG 1164
239 AGCCGCGA G CACCCAAG 580 CUUGGGUG GCCGAAAGGCGAGUCAAGGUCU UCGCGGCU 1165
247 GCACCCAA G UGUGCACC 581 GGUGCACA GCCGAAAGGCGAGUCAAGGUCU UUGGGUGC 1166
257 GUGCACCG G CACAGACA 582 UGUCUGUG GCCGAAAGGCGAGUCAAGGUCU CGGUGCAC 1167
270 GACAUGAA G CUGCGGCU 583 AGCCGCAG GCCGAAAGGCGAGUCAAGGUCU UUCAUGUC 1168
276 AAGCUGCG G CUCCCUGC 584 GCAGGGAG GCCGAAAGGCGAGUCAAGGUCU CGCAGCUU 1169
287 CCCUGCCA G UCCCGAGA 585 UCUCGGGA GCCGAAAGGCGAGUCAAGGUCU UGGCAGGG 1170
329 CUACCAGG G CUGCCAGG 586 CCUGGCAG GCCGAAAGGCGAGUCAAGGUCU CCUGGUAG 1171
337 GCUGCCAG G UGGUGCAG 587 CUGCACCA GCCGAAAGGCGAGUCAAGGUCU CUGGCAGC 1172
Table 59
340 GCCAGGUG G UGCAGGGA 588 UCCCUGCA GCCGAAAGGCGAGUCAAGGUCU CACCUGGC 1173
383 CAAUGCCA G CCUGUCCU 589 AGGACAGG GCCGAAAGGCGAGUCAAGGUCU UGGCAUUG 1174
412 UCCAGGAG G UGCAGGGC 590 GCCCUGCA GCCGAAAGGCGAGUCAAGGUCU CUCCUGGA 1175
419 GGUGCAGG G CUACGUGC 591 GCACGUAG GCCGAAAGGCGAGUCAAGGUCU CCUGCACC 1176
424 AGGGCUAC G UGCUCAUC 592 GAUGAGCA GCCGAAAGGCGAGUCAAGGUCU GUAGCCCU 1177
445 ACAACCAA G UGAGGCAG 593 CUGCCUCA GCCGAAAGGCGAGUCAAGGUCU UUGGUUGU 1178
450 CAAGUGAG G CAGGUCCC 594 GGGACCUG GCCGAAAGGCGAGUCAAGGUCU CUCACUUG 1179
454 UGAGGCAG G UCCCACUG 595 CAGUGGGA GCCGAAAGGCGAGUCAAGGUCU CUGCCUCA 1180
468 CUGCAGAG G CUGCGGAU 596 AUCCGCAG GCCGAAAGGCGAGUCAAGGUCU CUCUGCAG 1181
485 UGUGCGAG G CACCCAGC 597 GCUGGGUG GCCGAAAGGCGAGUCAAGGUCU CUCGCACA 1182
492 GGCACCCA G CUCUUUGA 598 UCAAAGAG GCCGAAAGGCGAGUCAAGGUCU UGGGUGCC 1183
517 AUGCCCUG G CCGUGCUA 599 UAGCACGG GCCGAAAGGCGAGUCAAGGUCU CAGGGCAU 1184
520 CCCUGGCC G UGCUAGAC 600 GUCUAGCA GCCGAAAGGCGAGUCAAGGUCU GGCCAGGG 1185
568 UCACAGGG G CCUCCCCA 601 UGGGGAGG GCCGAAAGGCGAGUCAAGGUCU CCCUGUGA 1186
581 CCCAGGAG G CCUGCGGG 602 CCCGCAGG GCCGAAAGGCGAGUCAAGGUCU CUCCUGGG 1187
591 CUGCGGGA G CUGCAGCU 603 AGCUGCAG GCCGAAAGGCGAGUCAAGGUCU UCCCGCAG 1188
597 GAGCUGCA G CUUCGAAG 604 CUUCGAAG GCCGAAAGGCGAGUCAAGGUCU UGCAGCUC 1189
605 GCUUCGAA G CCUCACAG 605 CUGUGAGG GCCGAAAGGCGAGUCAAGGUCU UUCGAAGC 1190
631 AAGGAGGG G UCUUGAUC 606 GAUCAAGA GCCGAAAGGCGAGUCAAGGUCU CCCUCCUU 1191
642 UUGAUCCA G CGGAACCC 607 GGGUUCCG GCCGAAAGGCGAGUCAAGGUCU UGGAUCAA 1192
654 AACCCCCA G CUCUGCUA 608 UAGCAGAG GCCGAAAGGCGAGUCAAGGUCU UGGGGGUU 1193
708 AACAACCA G CUGGCUCU 609 AGAGCCAG GCCGAAAGGCGAGUCAAGGUCU UGGUUGUU 1194
712 ACCAGCUG G CUCUCACA 610 UGUGAGAG GCCGAAAGGCGAGUCAAGGUCU CAGCUGGU 1195
745 GCUCUCGG G CCUGCCAC 611 GUGGCAGG GCCGAAAGGCGAGUCAAGGUCU CCGAGAGC 1196
776 GUGUAAGG G CUCCCGCU 612 AGCGGGAG GCCGAAAGGCGAGUCAAGGUCU CCUUACAC 1197
797 GGGAGAGA G UUCUGAGG 613 CCUCAGAA GCCGAAAGGCGAGUCAAGGUCU UCUCUCCC 1198
815 UUGUCAGA G CCUGACGC 614 GCGUCAGG GCCGAAAGGCGAGUCAAGGUCU UCUGACAA 1199
839 CUGUGCCG G UGGCUGUG 615 CACAGCCA GCCGAAAGGCGAGUCAAGGUCU CGGCACAG 1200
842 UGCCGGUG G CUGUGCCC 616 GGGCACAG GCCGAAAGGCGAGUCAAGGUCU CACCGGCA 1201
861 UGCAAGGG G CCACUGCC 617 GGCAGUGG GCCGAAAGGCGAGUCAAGGUCU CCCUUGCA 1202
888 UGCCAUGA G CAGUGUGC 618 GCACACUG GCCGAAAGGCGAGUCAAGGUCU UCAUGGCA 1203
891 CAUGAGCA G UGUGCUGC 619 GCAGCACA GCCGAAAGGCGAGUCAAGGUCU UGCUCAUG 1204
902 UGCUGCCG G CUGCACGG 620 CCGUGCAG GCCGAAAGGCGAGUCAAGGUCU CGGCAGCA 1205
911 CUGCACGG G CCCCAAGC 621 GCUUGGGG GCCGAAAGGCGAGUCAAGGUCU CCGUGCAG 1206
918 GGCCCCAA G CACUCUGA 622 UCAGAGUG GCCGAAAGGCGAGUCAAGGUCU UUGGGGCC 1207
934 ACUGCCUG G CCUGCCUC 623 GAGGCAGG GCCGAAAGGCGAGUCAAGGUCU CAGGCAGU 1208
956 CAACCACA G UGGCAUCU 624 AGAUGCCA GCCGAAAGGCGAGUCAAGGUCU UGUGGUUG 1209
959 CCACAGUG G CAUCUGUG 625 CACAGAUG GCCGAAAGGCGAGUCAAGGUCU CACUGUGG 1210
969 AUCUGUGA G CUGCACUG 626 CAGUGCAG GCCGAAAGGCGAGUCAAGGUCU UCACAGAU 1211
982 ACUGCCCA G CCCUGGUC 627 GACCAGGG GCCGAAAGGCGAGUCAAGGUCU UGGGCAGU 1212
988 CAGCCCUG G UCACCUAC 628 GUAGGUGA GCCGAAAGGCGAGUCAAGGUCU CAGGGCUG 1213
1008 ACAGACAC G UUUGAGUC 629 GACUCAAA GCCGAAAGGCGAGUCAAGGUCU GUGUCUGU 1214
1014 ACGUUUGA G UCCAUGCC 630 GGCAUGGA GCCGAAAGGCGAGUCAAGGUCU UCAAACGU 1215
1034 UCCCGAGG G CCGGUAUA 631 UAUACCGG GCCGAAAGGCGAGUCAAGGUCU CCUCGGGA 1216
1038 GAGGGCCG G UAUACAUU 632 AAUGUAUA GCCGAAAGGCGAGUCAAGGUCU CGGCCCUC 1217
1049 UACAUUCG G CGCCAGCU 633 AGCUGGCG GCCGAAAGGCGAGUCAAGGUCU CGAAUGUA 1218
1055 CGGCGCCA G CUGUGUGA 634 UCACACAG GCCGAAAGGCGAGUCAAGGUCU UGGCGCCG 1219
Table 59
1096 CUACGGAC G UGGGAUCC 635 GGAUCCCA GCCGAAAGGCGAGUCAAGGUCU GUCCGUAG 1220
1114 GCACCCUC G UCUGCCCC 636 GGGGCAGA GCCGAAAGGCGAGUCAAGGUCU GAGGGUGC 1221
1138 ACCAAGAG G UGACAGCA 637 UGCUGUCA GCCGAAAGGCGAGUCAAGGUCU CUCUUGGU 1222
1144 AGGUGACA G CAGAGGAU 638 AUCCUCUG GCCGAAAGGCGAGUCAAGGUCU UGUCACCU 1223
1161 GGAACACA G CGGUGUGA 639 UCACACCG GCCGAAAGGCGAGUCAAGGUCU UGUGUUCC 1224
1164 ACACAGCG G UGUGAGAA 640 UUCUCACA GCCGAAAGGCGAGUCAAGGUCU CGCUGUGU 1225
1173 UGUGAGAA G UGCAGCAA 641 UUGCUGCA GCCGAAAGGCGAGUCAAGGUCU UUCUCACA 1226
1178 GAAGUGCA G CAAGCCCU 642 AGGGCUUG GCCGAAAGGCGAGUCAAGGUCU UGCACUUC 1227
1182 UGCAGCAA G CCCUGUGC 643 GCACAGGG GCCGAAAGGCGAGUCAAGGUCU UUGCUGCA 1228
1195 GUGCCCGA G UGUGCUAU 644 AUAGCACA GCCGAAAGGCGAGUCAAGGUCU UCGGGCAC 1229
1205 GUGCUAUG G UCUGGGCA 645 UGCCCAGA GCCGAAAGGCGAGUCAAGGUCU CAUAGCAC 1230
1211 UGGUCUGG G CAUGGAGC 646 GCUCCAUG GCCGAAAGGCGAGUCAAGGUCU CCAGACCA 1231
1218 GGCAUGGA G CACUUGCG 647 CGCAAGUG GCCGAAAGGCGAGUCAAGGUCU UCCAUGCC 1232
1231 UGCGAGAG G UGAGGGCA 648 UGCCCUCA GCCGAAAGGCGAGUCAAGGUCU CUCUCGCA 1233
1237 AGGUGAGG G CAGUUACC 649 GGUAACUG GCCGAAAGGCGAGUCAAGGUCU CCUCACCU 1234
1240 UGAGGGCA G UUACCAGU 650 ACUGGUAA GCCGAAAGGCGAGUCAAGGUCU UGCCCUCA 1235
1247 AGUUACCA G UGCCAAUA 651 UAUUGGCA GCCGAAAGGCGAGUCAAGGUCU UGGUAACU 1236
1263 AUCCAGGA G UUUGCUGG 652 CCAGCAAA GCCGAAAGGCGAGUCAAGGUCU UCCUGGAU 1237
1271 GUUUGCUG G CUGCAAGA 653 UCUUGCAG GCCGAAAGGCGAGUCAAGGUCU CAGCAAAC 1238
1292 CUUUGGGA G CCUGGCAU 654 AUGCCAGG GCCGAAAGGCGAGUCAAGGUCU UCCCAAAG 1239
1297 GGAGCCUG G CAUUUCUG 655 CAGAAAUG GCCGAAAGGCGAGUCAAGGUCU CAGGCUCC 1240
1313 GCCGGAGA G CUUUGAUG 656 CAUCAAAG GCCGAAAGGCGAGUCAAGGUCU UCUCCGGC 1241
1330 GGGACCCA G CCUCCAAC 657 GUUGGAGG GCCGAAAGGCGAGUCAAGGUCU UGGGUCCC 1242
1353 CCGCUCCA G CCAGAGCA 658 UGCUCUGG GCCGAAAGGCGAGUCAAGGUCU UGGAGCGG 1243
1359 CAGCCAGA G CAGCUCCA 659 UGGAGCUG GCCGAAAGGCGAGUCAAGGUCU UCUGGCUG 1244
1362 CCAGAGCA G CUCCAAGU 660 ACUUGGAG GCCGAAAGGCGAGUCAAGGUCU UGCUCUGG 1245
1369 AGCUCCAA G UGUUUGAG 661 CUCAAACA GCCGAAAGGCGAGUCAAGGUCU UUGGAGCU 1246
1397 GAUCACAG G UUACCUAU 662 AUAGGUAA GCCGAAAGGCGAGUCAAGGUCU CUGUGAUC 1247
1414 ACAUCUCA G CAUGGCCG 663 CGGCCAUG GCCGAAAGGCGAGUCAAGGUCU UGAGAUGU 1248
1419 UCAGCAUG G CCGGACAG 664 CUGUCCGG GCCGAAAGGCGAGUCAAGGUCU CAUGCUGA 1249
1427 GCCGGACA G CCUGCCUG 665 CAGGCAGG GCCGAAAGGCGAGUCAAGGUCU UGUCCGGC 1250
1442 UGACCUCA G CGUCUUCC 666 GGAAGACG GCCGAAAGGCGAGUCAAGGUCU UGAGGUCA 1251
1444 ACCUCAGC G UCUUCCAG 667 CUGGAAOA GCCGAAAGGCGAGUCAAGGUCU GCUGAGGU 1252
1462 ACCUGCAA G UAAUCCGG 668 CCGGAUUA GCCGAAAGGCGAGUCAAGGUCU UUGCAGGU 1253
1490 GCACAAUG G CGCCUACU 669 AGUAGGCG GCCGAAAGGCGAGUCAAGGUCU CAUUGUGC 1254
1515 CUGCAAGG G CUGGGCAU 670 AUGCCCAG GCCGAAAGGCGAGUCAAGGUCU CCUUGCAG 1255
1520 AGGGCUGG G CAUCAGCU 671 AGCUGAUG GCCGAAAGGCGAGUCAAGGUCU CCAGCCCU 1256
1526 GGGCAUCA G CUGGCUGG 672 CCAGCCAG GCCGAAAGGCGAGUCAAGGUCU UGAUGCCC 1257
1530 AUCAGCUG G CUGGGGCU 673 AGCCCCAG GCCGAAAGGCGAGUCAAGGUCU CAGCUGAU 1258
1536 UGGCUGGG G CUGCGCUC 674 GAGCGCAG GCCGAAAGGCGAGUCAAGGUCU CCCAGCCA 1259
1559 GGAACUGG G CAGUGGAC 675 GUCCACUG GCCGAAAGGCGAGUCAAGGUCU CCAGUUCC 1260
1562 ACUGGGCA G UGGACUGG 676 CCAGUCCA GCCGAAAGGCGAGUCAAGGUCU UGCCCAGU 1261
1570 GUGGACUG G CCCUCAUC 677 GAUGAGGG GCCGAAAGGCGAGUCAAGGUCU CAGUCCAC 1262
1603 UCUGCUUC G UGCACACG 678 CGUGUGCA GCCGAAAGGCGAGUCAAGGUCU GAAGCAGA 1263
1612 UGCACACG G UGCCCUGG 679 CCAGGGCA GCCGAAAGGCGAGUCAAGGUCU CGUGUGCA 1264
1626 UGGGACCA G CUCUUUCG 680 CGAAAGAG GCCGAAAGGCGAGUCAAGGUCU UGGUCCCA 1265
1648 CGCACCAA G CUCUGCUC 681 GAGCAGAG GCCGAAAGGCGAGUCAAGGUCU UUGGUGCG 1266
Table 59
1671 GCCAACCG G CCAGAGGA 682 UCCUCUGG GCCGAAAGGCGAGUCAAGGUCU CGGUUGGC 1267
1683 GAGGACGA G UGUGUGGG 683 CCCACACA GCCGAAAGGCGAGUCAAGGUCU UCGUCCUC 1268
1691 GUGUGUGG G CGAGGGCC 684 GGCCCUCG GCCGAAAGGCGAGUCAAGGUCU CCACACAC 1269
1697 GGGCGAGG G CCUGGCCU 685 AGGCCAGG GCCGAAAGGCGAGUCAAGGUCU CCUCGCCC 1270
1702 AGGGCCUG G CCUGCCAC 686 GUGGCAGG GCCGAAAGGCGAGUCAAGGUCU CAGGCCCU 1271
1713 UGCCACCA G CUGUGCGC 687 GCGCACAG GCCGAAAGGCGAGUCAAGGUCU UGGUGGCA 1272
1728 GCCCGAGG G CACUGCUG 688 CAGCAGUG GCCGAAAGGCGAGUCAAGGUCU CCUCGGGC 1273
1739 CUGCUGGG G UCCAGGGC 689 GCCCUGGA GCCGAAAGGCGAGUCAAGGUCU CCCAGCAG 1274
1746 GGUCCAGG G CCCACCCA 690 UGGGUGGG GCCGAAAGGCGAGUCAAGGUCU CCUGGACC 1275
1755 CCCACCCA G UGUGUCAA 691 UUGACACA GCCGAAAGGCGAGUCAAGGUCU UGGGUGGG 1276
1769 CAACUGCA G CCAGUUCC 692 GGAACUGG GCCGAAAGGCGAGUCAAGGUCU UGCAGUUG 1277
1773 UGCAGCCA G UUCCUUCG 693 CGAAGGAA GCCGAAAGGCGAGUCAAGGUCU UGGCUGCA 1278
1784 CCUUCGGG G CCAGGAGU 694 ACUCCUGG GCCGAAAGGCGAGUCAAGGUCU CCCGAAGG 1279
1791 GGCCAGGA G UGCGUGGA 695 UCCACGCA GCCGAAAGGCGAGUCAAGGUCU UCCUGGCC 1280
1795 AGGAGUGC G UGGAGGAA 696 UUCCUCCA GCCGAAAGGCGAGUCAAGGUCU GCACUCCU 1281
1810 AAUGCCGA G UACUGCAG 697 CUGCAGUA GCCGAAAGGCGAGUCAAGGUCU UCGGCAUU 1282
1821 CUGCAGGG G CUCCCCAG 698 CUGGGGAG GCCGAAAGGCGAGUCAAGGUCU CCCUGCAG 1283
1833 CCCAGGGA G UAUGUGAA 699 UUCACAUA GCCGAAAGGCGAGUCAAGGUCU UCCCUGGG 1284
1848 AAUGCCAG G CACUGUUU 700 AAACAGUG GCCGAAAGGCGAGUCAAGGUCU CUGGCAUU 1285
1860 UGUUUGCC G UGCCACCC 701 GGGUGGCA GCCGAAAGGCGAGUCAAGGUCU GGCAAACA 1286
1872 CACCCUGA G UGUCAGCC 702 GGCUGACA GCCGAAAGGCGAGUCAAGGUCU UCAGGGUG 1287
1878 GAGUGUCA G CCCCAGAA 703 UUCUGGGG GCCGAAAGGCGAGUCAAGGUCU UGACACUC 1288
1889 CCAGAAUG G CUCAGUGA 704 UCACUGAG GCCGAAAGGCGAGUCAAGGUCU CAUUCUGG 1289
1894 AUGGCUCA G UGACCUGU 705 ACAGGUCA GCCGAAAGGCGAGUCAAGGUCU UGAGCCAU 1290
1915 GACCGGAG G CUGACCAG 706 CUGGUCAG GCCGAAAGGCGAGUCAAGGUCU CUCCGGUC 1291
1923 GCUGACCA G UGUGUGGC 707 GCCACACA GCCGAAAGGCGAGUCAAGGUCU UGGUCAGC 1292
1930 AGUGUGUG G CCUGUGCC 708 GGCACAGG GCCGAAAGGCGAGUCAAGGUCU CACACACU 1293
1963 CCUUCUGC G UGGCCCGC 709 GCGGGCCA GCCGAAAGGCGAGUCAAGGUCU GCAGAAGG 1294
1966 UCUGCGUG G CCCGCUGC 710 GCAGCGGG GCCGAAAGGCGAGUCAAGGUCU CACGCAGA 1295
1979 CUGCCCCA G CGGUGUGA 711 UCACACCG GCCGAAAGGCGAGUCAAGGUCU UGGGGCAG 1296
1982 CCCCAGCG G UGUGAAAC 712 GUUUCACA GCCGAAAGGCGAGUCAAGGUCU CGCUGGGG 1297
2019 AUCUGGAA G UUUCCAGA 713 UCUGGAAA GCCGAAAGGCGAGUCAAGGUCU UUCCAGAU 1298
2036 UGAGGAGG G CGCAUGCC 714 GGCAUGCG GCCGAAAGGCGAGUCAAGGUCU CCUCCUCA 1299
2046 GCAUGCCA G CCUUGCCC 715 GGGCAAGG GCCGAAAGGCGAGUCAAGGUCU UGGCAUGC 1300
2096 UGACAAGG G CUGCCCCG 716 CGGGGCAG GCCGAAAGGCGAGUCAAGGUCU CCUUGUCA 1301
2109 CCCGCCGA G CAGAGAGC 717 GCUCUCUG GCCGAAAGGCGAGUCAAGGUCU UCGGCGGG 1302
2116 AGCAGAGA G CCAGCCCU 718 AGGGCUGG GCCGAAAGGCGAGUCAAGGUCU UCUCUGCU 1303
2120 GAGAGCCA G CCCUCUGA 719 UCAGAGGG GCCGAAAGGCGAGUCAAGGUCU UGGCUCUC 1304
2130 CCUCUGAC G UCCAUCAU 720 AUGAUGGA GCCGAAAGGCGAGUCAAGGUCU GUCAGAGG 1305
2146 UCUCUGCG G UGGUUGGC 721 GCCAACCA GCCGAAAGGCGAGUCAAGGUCU CGCAGAGA 1306
2149 CUGCGGUG G UUGGCAUU 722 AAUGCCAA GCCGAAAGGCGAGUCAAGGUCU CACCGCAG 1307
2153 GGUGGUUG G CAUUCUGC 723 GCAGAAUG GCCGAAAGGCGAGUCAAGGUCU CAACCACC 1308
2164 UUCUGCUG G UCGUGGUC 724 GACCACGA GCCGAAAGGCGAGUCAAGGUCU CAGCAGAA 1309
2167 UGCUGGUC G UGGUCUUG 725 CAAGACCA GCCGAAAGGCGAGUCAAGGUCU GACCAGCA 1310
2170 UGGUCGUG G UCUUGGGG 726 CCCCAAGA GCCGAAAGGCGAGUCAAGGUCU CACGACCA 1311
2179 UCUUGGGG G UGGUCUUU 727 AAAGACCA GCCGAAAGGCGAGUCAAGGUCU CCCCAAGA 1312
2182 UGGGGGUG G UCUUUGGG 728 CCCAAAGA GCCGAAAGGCGAGUCAAGGUCU CACCCCCA 1313
Table 59
2202 CUCAUCAA G CGACGGCA 729 UGCCGUCG GCCGAAAGGCGAGUCAAGGUCU UUGAUGAG 1314
2208 AAGCGACG G CAGCAGAA 730 UUCUGCUG GCCGAAAGGCGAGUCAAGGUCU CGUCGCUU 1315
2211 CGACGGCA G CAGAAGAU 731 AUCUUCUG GCCGAAAGGCGAGUCAAGGUCU UGCCGUCG 1316
2226 AUCCGGAA G UACACGAU 732 AUCGUGUA GCCGAAAGGCGAGUCAAGGUCU UUCCGGAU 1317
2259 GAAACGGA G CUGGUGGA 733 UCCACCAG GCCGAAAGGCGAGUCAAGGUCU UCCGUUUC 1318
2263 CGGAGCUG G UGGAGCCG 734 CGGCUCCA GCCGAAAGGCGAGUCAAGGUCU CAGCUCCG 1319
2268 CUGGUGGA G CCGCUGAC 735 GUCAGCGG GCCGAAAGGCGAGUCAAGGUCU UCCACCAG 1320
2282 GACACCUA G CGGAGCGA 736 UCGCUCCG GCCGAAAGGCGAGUCAAGGUCU UAGGUGUC 1321
2287 CUAGCGGA G CGAUGCCC 737 GGGCAUCG GCCGAAAGGCGAGUCAAGGUCU UCCGCUAG 1322
2302 CCAACCAG G CGCAGAUG 738 CAUCUGCG GCCGAAAGGCGAGUCAAGGUCU CUGGUUGG 1323
2331 GAGACGGA G CUGAGGAA 739 UUCCUCAG GCCGAAAGGCGAGUCAAGGUCU UCCGUCUC 1324
2341 UGAGGAAG G UGAAGGUG 740 CACCUUCA GCCGAAAGGCGAGUCAAGGUCU CUUCCUCA 1325
2347 AGGUGAAG G UGCUUGGA 741 UCCAAGCA GCCGAAAGGCGAGUCAAGGUCU CUUCACCU 1326
2360 UGGAUCUG G CGCUUUUG 742 CAAAAGCG GCCGAAAGGCGAGUCAAGGUCU CAGAUCCA 1327
2369 CGCUUUUG G CACAGUCU 743 AGACUGUG GCCGAAAGGCGAGUCAAGGUCU CAAAAGCG 1328
2374 UUGGCACA G UCUACAAG 744 CUUGUAGA GCCGAAAGGCGAGUCAAGGUCU UGUGCCAA 1329
2384 CUACAAGG G CAUCUGGA 745 UCCAGAUG GCCGAAAGGCGAGUCAAGGUCU CCUUGUAG 1330
2422 AAAUUCCA G UGGCCAUC 746 GAUGGCCA GCCGAAAGGCGAGUCAAGGUCU UGGAAUUU 1331
2425 UUCCAGUG G CCAUCAAA 747 UUUGAUGG GCCGAAAGGCGAGUCAAGGUCU CACUGGAA 1332
2434 CCAUCAAA G UGUUGAGG 748 CCUCAACA GCCGAAAGGCGAGUCAAGGUCU UUUGAUGG 1333
2461 CCCCCAAA G CCAACAAA 749 UUUGUUGG GCCGAAAGGCGAGUCAAGGUCU UUUGGGGG 1334
2485 UAGACGAA G CAUACGUG 750 CACGUAUG GCCGAAAGGCGAGUCAAGGUCU UUCGUCUA 1335
2491 AAGCAUAC G UGAUGGCU 751 AGCCAUCA GCCGAAAGGCGAGUCAAGGUCU GUAUGCUU 1336
2497 ACGUGAUG G CUGGUGUG 752 CACACCAG GCCGAAAGGCGAGUCAAGGUCU CAUCACGU 1337
2501 GAUGGCUG G UGUGGGCU 753 AGCCCACA GCCGAAAGGCGAGUCAAGGUCU CAGCCAUC 1338
2507 UGGUGUGG G CUCCCCAU 754 AUGGGGAG GCCGAAAGGCGAGUCAAGGUCU CCACACCA 1339
2534 CCUUCUGG G CAUCUGCC 755 GGCAGAUG GCCGAAAGGCGAGUCAAGGUCU CCAGAAGG 1340
2554 CAUCCACG G UGCAGCUG 756 CAGCUGCA GCCGAAAGGCGAGUCAAGGUCU CGUGGAUG 1341
2559 ACGGUGCA G CUGGUGAC 757 GUCACCAG GCCGAAAGGCGAGUCAAGGUCU UGCACCGU 1342
2563 UGCAGCUG G UGACACAG 758 CUGUGUCA GCCGAAAGGCGAGUCAAGGUCU CAGCUGCA 1343
2571 GUGACACA G CUUAUGCC 759 GGCAUAAG GCCGAAAGGCGAGUCAAGGUCU UGUGUCAC 1344
2585 GCCCUAUG G CUGCCUCU 760 AGAGGCAG GCCGAAAGGCGAGUCAAGGUCU CAUAGGGC 1345
2627 ACGCCUGG G CUCCCAGG 761 CCUGGGAG GCCGAAAGGCGAGUCAAGGUCU CCAGGCGU 1346
2649 CUGAACUG G UGUAUGCA 762 UGCAUACA GCCGAAAGGCGAGUCAAGGUCU CAGUUCAG 1347
2675 GGGGAUGA G CUACCUGG 763 CCAGGUAG GCCGAAAGGCGAGUCAAGGUCU UCAUCCCC 1348
2694 GAUGUGCG G CUCGUACA 764 UGUACGAG GCCGAAAGGCGAGUCAAGGUCU CGCACAUC 1349
2698 UGCGGCUC G UACACAGG 765 CCUGUGUA GCCGAAAGGCGAGUCAAGGUCU GAGCCGCA 1350
2713 GGGACUUG G CCGCUCGG 766 CCGAGCGG GCCGAAAGGCGAGUCAAGGUCU CAAGUCCC 1351
2725 CUCGGAAC G UGCUGGUC 767 GACCAGCA GCCGAAAGGCGAGUCAAGGUCU GUUCCGAG 1352
2731 ACGUGCUG G UCAAGAGU 768 ACUCUUGA GCCGAAAGGCGAGUCAAGGUCU CAGCACGU 1353
2738 GGUCAAGA G UCCCAACC 769 GGUUGGGA GCCGAAAGGCGAGUCAAGGUCU UCUUGACC 1354
2769 GACUUCGG G CUGGCUCG 770 CGAGCCAG GCCGAAAGGCGAGUCAAGGUCU CCGAAGUC 1355
2773 UCGGGCUG G CUCGGCUG 771 CAGCCGAG GCCGAAAGGCGAGUCAAGGUCU CAGCCCGA 1356
2778 CUGGCUCG G CUGCUGGA 772 UCCAGCAG GCCGAAAGGCGAGUCAAGGUCU CGAGCCAG 1357
2802 GAGACAGA G UACCAUGC 773 GCAUGGUA GCCGAAAGGCGAGUCAAGGUCU UCUGUCUC 1358
2819 AGAUGGGG G CAAGGUGC 774 GCACCUUG GCCGAAAGGCGAGUCAAGGUCU CCCCAUCU 1359
2824 GGGGCAAG G UGCCCAUC 775 GAUGGGCA GCCGAAAGGCGAGUCAAGGUCU CUUGCCCC 1360
Table 59
2835 CCCAUCAA G UGGAUGGC 776 GCCAUCCA GCCGAAAGGCGAGUCAAGGUCU UUGAUGGG 1361
2842 AGUGGAUG G CGCUGGAG 777 CUCCAGCG GCCGAAAGGCGAGUCAAGGUCU CAUCCACU 1362
2850 GCGCUGGA G UCCAUUCU 778 AGAAUGGA GCCGAAAGGCGAGUCAAGGUCU UCCAGCGC 1363
2865 CUCCGCCG G CGGUUCAC 779 GUGAACCG GCCGAAAGGCGAGUCAAGGUCU CGGCGGAG 1364
2868 CGCCGGCG G UUCACCCA 780 UGGGUGAA GCCGAAAGGCGAGUCAAGGUCU CGCCGGCG 1365
2882 CCACCAGA G UGAUGUGU 781 ACACAUCA GCCGAAAGGCGAGUCAAGGUCU UCUGGUGG 1366
2894 UGUGUGGA G UUAUGGUG 782 CACCAUAA GCCGAAAGGCGAGUCAAGGUCU UCCACACA 1367
2900 GAGUUAUG G UGUGACUG 783 CAGUCACA GCCGAAAGGCGAGUCAAGGUCU CAUAACUC 1368
2916 GUGUGGGA G CUGAUGAC 784 GUCAUCAG GCCGAAAGGCGAGUCAAGGUCU UCCCACAC 1369
2932 CUUUUGGG G CCAAACCU 785 AGGUUUGG GCCGAAAGGCGAGUCAAGGUCU CCCAAAAG 1370
2956 GGAUCCCA G CCCGGGAG 786 CUCCCGGG GCCGAAAGGCGAGUCAAGGUCU UGGGAUCC 1371
2991 AAGGGGGA G CGGCUGCC 787 GGCAGCCG GCCGAAAGGCGAGUCAAGGUCU UCCCCCUU 1372
2994 GGGGAGCG G CUGCCCCA 788 UGGGGCAG GCCGAAAGGCGAGUCAAGGUCU CGCUCCCC 1373
3003 CUGCCCCA G CCCCCCAU 789 AUGGGGGG GCCGAAAGGCGAGUCAAGGUCU UGGGGCAG 1374
3040 UGAUCAUG G UCAAAUGU 790 ACAUUUGA GCCGAAAGGCGAGUCAAGGUCU CAUGAUCA 1375
3072 GAAUGUCG G CCAAGAUU 791 AAUCUUGG GCCGAAAGGCGAGUCAAGGUCU CGACAUUC 1376
3087 UUCCGGGA G UUGGUGUC 792 GACACCAA GCCGAAAGGCGAGUCAAGGUCU UCCCGGAA 1377
3091 GGGAGUUG G UGUCUGAA 793 UUCAGACA GCCGAAAGGCGAGUCAAGGUCU CAACUCCC 1378
3112 CCCGCAUG G CCAGGGAC 794 GUCCCUGG GCCGAAAGGCGAGUCAAGGUCU CAUGCGGG 1379
3126 GACCCCCA G CGCUUUGU 795 ACAAAGCG GCCGAAAGGCGAGUCAAGGUCU UGGGGGUC 1380
3136 GCUUUGUG G UCAUCCAG 796 CUGGAUGA GCCGAAAGGCGAGUCAAGGUCU CACAAAGC 1381
3158 GGACUUGG G CCCAGCCA 797 UGGCUGGG GCCGAAAGGCGAGUCAAGGUCU CCAAGUCC 1382
3163 UGGGCCCA G CCAGUCCC 798 GGGACUGG GCCGAAAGGCGAGUCAAGGUCU UGGGCCCA 1383
3167 CCCAGCCA G UCCCUUGG 799 CCAAGGGA GCCGAAAGGCGAGUCAAGGUCU UGGCUGGG 1384
3179 CUUGGACA G CACCUUCU 800 AGAAGGUG GCCGAAAGGCGAGUCAAGGUCU UGUCCAAG 1385
3226 GGGACCUG G UGGAUGCU 801 AGCAUCCA GCCGAAAGGCGAGUCAAGGUCU CAGGUCCC 1386
3240 GCUGAGGA G UAUCUGGU 802 ACCAGAUA GCCGAAAGGCGAGUCAAGGUCU UCCUCAGC 1387
3247 AGUAUCUG G UACCCCAG 803 CUGGGGUA GCCGAAAGGCGAGUCAAGGUCU CAGAUACU 1388
3255 GUACCCCA G CAGGGCUU 804 AAGCCCUG GCCGAAAGGCGAGUCAAGGUCU UGGGGUAC 1389
3260 CCAGCAGG G CUUCUUCU 805 AGAAGAAG GCCGAAAGGCGAGUCAAGGUCU CCUGCUGG 1390
3287 UGCCCCGG G CGCUGGGG 806 CCCCAGCG GCCGAAAGGCGAGUCAAGGUCU CCGGGGCA 1391
3296 CGCUGGGG G CAUGGUCC 807 GGACCAUG GCCGAAAGGCGAGUCAAGGUCU CCCCAGCG 1392
3301 GGGGCAUG G UCCACCAC 808 GUGGUGGA GCCGAAAGGCGAGUCAAGGUCU CAUGCCCC 1393
3312 CACCACAG G CACCGCAG 809 CUGCGGUG GCCGAAAGGCGAGUCAAGGUCU CUGUGGUG 1394
3320 GCACCGCA G CUCAUCUA 810 UAGAUGAG GCCGAAAGGCGAGUCAAGGUCU UGCGGUGC 1395
3335 UACCAGGA G UGGCGGUG 811 CACCGCCA GCCGAAAGGCGAGUCAAGGUCU UCCUGGUA 1396
3338 CAGGAGUG G CGGUGGGG 812 CCCCACCG GCCGAAAGGCGAGUCAAGGUCU CACUCCUG 1397
3341 GAGUGGCG G UGGGGACC 813 GGUCCCCA GCCGAAAGGCGAGUCAAGGUCU CGCCACUC 1398
3360 ACACUAGG G CUGGAGCC 814 GGCUCCAG GCCGAAAGGCGAGUCAAGGUCU CCUAGUGU 1399
3366 GGGCUGGA G CCCUCUGA 815 UCAGAGGG GCCGAAAGGCGAGUCAAGGUCU UCCAGCCC 1400
3382 AAGAGGAG G CCCCCAGG 816 CCUGGGGG GCCGAAAGGCGAGUCAAGGUCU CUCCUCUU 1401
3390 GCCCCCAG G UCUCCACU 817 AGUGGAGA GCCGAAAGGCGAGUCAAGGUCU CUGGGGGC 1402
3400 CUCCACUG G CACCCUCC 818 GGAGGGUG GCCGAAAGGCGAGUCAAGGUCU CAGUGGAG 1403
3415 CCGAAGGG G CUGGCUCC 819 GGAGCCAG GCCGAAAGGCGAGUCAAGGUCU CCCUUCGG 1404
3419 AGGGGCUG G CUCCGAUG 820 CAUCGGAG GCCGAAAGGCGAGUCAAGGUCU CAGCCCCU 1405
3437 AUUUGAUG G UGACCUGG 821 CCAGGUCA GCCGAAAGGCGAGUCAAGGUCU CAUCAAAU 1406
3454 GAAUGGGG G CAGCCAAG 822 CUUGGCUG GCCGAAAGGCGAGUCAAGGUCU CCCCAUUC 1407
Table 59
3457 UGGGGGCA G CCAAGGGG 823 CCCCUUGG GCCGAAAGGCGAGUCAAGGUCU UGCCCCCA 1408
3465 GCCAAGGG G CUGCAAAG 824 CUUUGCAG GCCGAAAGGCGAGUCAAGGUCU CCCUUGGC 1409
3473 GCUGCAAA G CCUCCCCA 825 UGGGGAGG GCCGAAAGGCGAGUCAAGGUCU UUUGCAGC 1410
3494 UGACCCCA G CCCUCUAC 826 GUAGAGGG GCCGAAAGGCGAGUCAAGGUCU UGGGGUCA 1411
3504 CCUCUACA G CGGUACAG 827 CUGUACCG GCCGAAAGGCGAGUCAAGGUCU UGUAGAGG 1412
3507 CUACAGCG G UACAGUGA 828 UCACUGUA GCCGAAAGGCGAGUCAAGGUCU CGCUGUAG 1413
3512 GCGGUACA G UGAGGACC 829 GGUCCUCA GCCGAAAGGCGAGUCAAGGUCU UGUACCGC 1414
3526 ACCCCACA G UACCCCUG 830 CAGGGGUA GCCGAAAGGCGAGUCAAGGUCU UGUGGGGU 1415
3551 GACUGAUG G CUACGUUG 831 CAACGUAG GCCGAAAGGCGAGUCAAGGUCU CAUCAGUC 1416
3556 AUGGCUAC G UUGCCCCC 832 GGGGGCAA GCCGAAAGGCGAGUCAAGGUCU GUAGCCAU 1417
3575 GACCUGCA G CCCCCAGC 833 GCUGGGGG GCCGAAAGGCGAGUCAAGGUCU UGCAGGUC 1418
3582 AGCCCCCA G CCUGAAUA 834 UAUUCAGG GCCGAAAGGCGAGUCAAGGUCU UGGGGGCU 1419
3600 GUGAACCA G CCAGAUGU 835 ACAUCUGG GCCGAAAGGCGAGUCAAGGUCU UGGUUCAC 1420
3612 GAUGUUCG G CCCCAGCC 836 GGCUGGGG GCCGAAAGGCGAGUCAAGGUCU CGAACAUC 1421
3618 CGGCCCCA G CCCCCUUC 837 GAAGGGGG GCCGAAAGGCGAGUCAAGGUCU UGGGGCCG 1422
3638 CCGAGAGG G CCCUCUGC 838 GCAGAGGG GCCGAAAGGCGAGUCAAGGUCU CCUCUCGG 1423
3665 ACCUGCUG G UGCCACUC 839 GAGUGGCA GCCGAAAGGCGAGUCAAGGUCU CAGCAGGU 1424
3681 CUGGAAAG G CCCAAGAC 840 GUCUUGGG GCCGAAAGGCGAGUCAAGGUCU CUUUCCAG 1425
3712 AGAAUGGG G UCGUCAAA 841 UUUGACGA GCCGAAAGGCGAGUCAAGGUCU CCCAUUCU 1426
3715 AUGGGGUC G UCAAAGAC 842 GUCUUUGA GCCGAAAGGCGAGUCAAGGUCU GACCCCAU 1427
3724 UCAAAGAC G UUUUUGCC 843 GGCAAAAA GCCGAAAGGCGAGUCAAGGUCU GUCUUUGA 1428
3740 CUUUGGGG G UGCCGUGG 844 CCACGGCA GCCGAAAGGCGAGUCAAGGUCU CCCCAAAG 1429
3745 GGGGUGCC G UGGAGAAC 845 GUUCUCCA GCCGAAAGGCGAGUCAAGGUCU GGCACCCC 1430
3759 AACCCCGA G UACUUGAC 846 GUCAAGUA GCCGAAAGGCGAGUCAAGGUCU UCGGGGUU 1431
3781 AGGGAGGA G CUGCCCCU 847 AGGGGCAG GCCGAAAGGCGAGUCAAGGUCU UCCUCCCU 1432
3792 GCCCCUCA G CCCCACCC 848 GGGUGGGG GCCGAAAGGCGAGUCAAGGUCU UGAGGGGC 1433
3815 UGCCUUCA G CCCAGCCU 849 AGGCUGGG GCCGAAAGGCGAGUCAAGGUCU UGAAGGCA 1434
3820 UCAGCCCA G CCUUCGAC 850 GUCGAAGG GCCGAAAGGCGAGUCAAGGUCU UGGGCUGA 1435
3861 CCACCAGA G CGGGGGGC 851 GCCCCCCG GCCGAAAGGCGAGUCAAGGUCU UCUGGUGG 1436
3868 AGCGGGGG G CUCCACCC 852 GGGUGGAG GCCGAAAGGCGAGUCAAGGUCU CCCCCGCU 1437
3878 UCCACCCA G CACCUUCA 853 UGAAGGUG GCCGAAAGGCGAGUCAAGGUCU UGGGUGGA 1438
3901 CACCUACG G CAGAGAAC 854 GUUCUCUG GCCGAAAGGCGAGUCAAGGUCU CGUAGGUG 1439
3915 AACCCAGA G UACCUGGG 855 CCCAGGUA GCCGAAAGGCGAGUCAAGGUCU UCUGGGUU 1440
3923 GUACCUGG G UCUGGACG 856 CGUCCAGA GCCGAAAGGCGAGUCAAGGUCU CCAGGUAC 1441
3931 GUCUGGAC G UGCCAGUG 857 CACUGGCA GCCGAAAGGCGAGUCAAGGUCU GUCCAGAC 1442
3937 ACGUGCCA G UGUGAACC 858 GGUUCACA GCCGAAAGGCGAGUCAAGGUCU UGGCACGU 1443
3951 ACCAGAAG G CCAAGUCC 859 GGACUUGG GCCGAAAGGCGAGUCAAGGUCU CUUCUGGU 1444
3956 AAGGCCAA G UCCGCAGA 860 UCUGCGGA GCCGAAAGGCGAGUCAAGGUCU UUGGCCUU 1445
3966 CCGCAGAA G CCCUGAUG 861 CAUCAGGG GCCGAAAGGCGAGUCAAGGUCU UUCUGCGG 1446
3987 CUCAGGGA G CAGGGAAG 862 CUUCCCUG GCCGAAAGGCGAGUCAAGGUCU UCCCUGAG 1447
3996 CAGGGAAG G CCUGACUU 863 AAGUCAGG GCCGAAAGGCGAGUCAAGGUCU CUUCCCUG 1448
4011 UUCUGCUG G CAUCAAGA 864 UCUUGAUG GCCGAAAGGCGAGUCAAGGUCU CAGCAGAA 1449
4021 AUCAAGAG G UGGGAGGG 865 CCCUCCCA GCCGAAAGGCGAGUCAAGGUCU CUCUUGAU 1450
4029 GUGGGAGG G CCCUCCGA 866 UCGGAGGG GCCGAAAGGCGAGUCAAGGUCU CCUCCCAC 1451
4100 CUGCUUGA G UUCCCAGA 867 UCUGGGAA GCCGAAAGGCGAGUCAAGGUCU UCAAGCAG 1452
4111 CCCAGAUG G CUGGAAGG 868 CCUUCCAG GCCGAAAGGCGAGUCAAGGUCU CAUCUGGG 1453
4121 UGGAAGGG G UCCAGCCU 869 AGGCUGGA GCCGAAAGGCGAGUCAAGGUCU CCCUUCCA 1454
Table 59
4126 GGGGUCCA G CCUCGUUG 870 CAACGAGG GCCGAAAGGCGAGUCAAGGUCU UGGACCCC 1455
4131 CCAGCCUC G UUGGAAGA 871 UCUUCCAA GCCGAAAGGCGAGUCAAGGUCU GAGGCUGG 1456
4146 GAGGAACA G CACUGGGG 872 CCCCAGUG GCCGAAAGGCGAGUCAAGGUCU UGUUCCUC 1457
4156 ACUGGGGA G UCUUUGUG 873 CACAAAGA GCCGAAAGGCGAGUCAAGGUCU UCCCCAGU 1458
4174 AUUCUGAG G CCCUGCCC 874 GGGCAGGG GCCGAAAGGCGAGUCAAGGUCU CUCAGAAU 1459
4197 ACUCUAGG G UCCAGUGG 875 CCACUGGA GCCGAAAGGCGAGUCAAGGUCU CCUAGAGU 1460
4202 AGGGUCCA G UGGAUGCC 876 GGCAUCCA GCCGAAAGGCGAGUCAAGGUCU UGGACCCU 1461
4214 AUGCCACA G CCCAGCUU 877 AAGCUGGG GCCGAAAGGCGAGUCAAGGUCU UGUGGCAU 1462
4219 ACAGCCCA G CUUGGCCC 878 GGGCCAAG GCCGAAAGGCGAGUCAAGGUCU UGGGCUGU 1463
4224 CCAGCUUG G CCCUUUCC 879 GGAAAGGG GCCGAAAGGCGAGUCAAGGUCU CAAGCUGG 1464
4246 GAUCCUGG G UACUGAAA 880 UUUCAGUA GCCGAAAGGCGAGUCAAGGUCU CCAGGAUC 1465
4255 UACUGAAA G CCUUAGGG 881 CCCUAAGG GCCGAAAGGCGAGUCAAGGUCU UUUCAGUA 1466
4266 UUAGGGAA G CUGGCCUG 882 CAGGCCAG GCCGAAAGGCGAGUCAAGGUCU UUCCCUAA 1467
4270 GGAAGCUG G CCUGAGAG 883 CUCUCAGG GCCGAAAGGCGAGUCAAGGUCU CAGCUUCC 1468
4284 GAGGGGAA G CGGCCCUA 884 UAGGGCCG GCCGAAAGGCGAGUCAAGGUCU UUCCCCUC 1469
4287 GGGAAGCG G CCCUAAGG 885 CCUUAGGG GCCGAAAGGCGAGUCAAGGUCU CGCUUCCC 1470
4298 CUAAGGGA G UGUCUAAG 886 CUUAGACA GCCGAAAGGCGAGUCAAGGUCU UCCCUUAG 1471
4314 GAACAAAA G CGACCCAU 887 AUGGGUCG GCCGAAAGGCGAGUCAAGGUCU UUUUGUUC 1472
4346 GAAACCUA G UACUGCCC 888 GGGCAGUA GCCGAAAGGCGAGUCAAGGUCU UAGGUUUC 1473
4372 AAGGAACA G CAAUGGUG 889 CACCAUUG GCCGAAAGGCGAGUCAAGGUCU UGUUCCUU 1474
4378 CAGCAAUG G UGUCAGUA 890 UACUGACA GCCGAAAGGCGAGUCAAGGUCU CAUUGCUG 1475
4384 UGGUGUCA G UAUCCAGG 891 CCUGGAUA GCCGAAAGGCGAGUCAAGGUCU UGACACCA 1476
4392 GUAUCCAG G CUUUGUAC 892 GUACAAAG GCCGAAAGGCGAGUCAAGGUCU CUGGAUAC 1477
4404 UGUACAGA G UGCUUUUC 893 GAAAAGCA GCCGAAAGGCGAGUCAAGGUCU UCUGUACA 1478
4419 UCUGUUUA G UUUUUACU 894 AGUAAAAA GCCGAAAGGCGAGUCAAGGUCU UAAACAGA 1479
Input Sequence = HSE B2 . Cut Site = G/Y
Stem Length = 8 . Core Sequence = GCcgaaagGCGaGuCaaGGuCu
HSERB2R (Human c-erb-B-2 mRNA; 4473 bp)
Table 60
Table 60: Substrate Specificity for Class I Ribozymes
Table 61
Table 61: Random region alignments/mutations for Class I ribozyme
Table 62
Table 62: Human Her2 Class II Ribozyme and Target Sequence
Table 62
U, C = 2'-deoxy-2'-amino U, = 2'-deoxy-2'-amino C
G,A = ribo G, A
B = inverted deoxyabasic
P= polyethylene glycol 18 (PEG 18) linker
Table 63 Table 63: Human PKCα NCH Ribozyme and Substrate Sequence
224 AGGCUUCC A GUGCCAAG CUUGGCAC CUGAUGAG X CGAA IGAAGCCU
229 UCCAGUGC C AAGUUUGC GCAAACUU CUGAUGAG X CGAA ICACUGGA
230 CCAGUGCC A AGUUUGCU AGCAAACU CUGAUGAG X CGAA IGCACUGG
238 AAGUUUGC U GUUUUGUG CACAAAAC CUGAUGAG X CGAA ICAAACUU
250 UUGUGGUC C ACAAGAGG CCUCUUGU CUGAUGAG X CGAA IACCACAA
251 UGUGGUCC A CAAGAGGU ACCUCUUG CUGAUGAG X CGAA IGACCACA
253 UGGUCCAC A AGAGGUGC GCACCUCU CUGAUGAG X CGAA IUGGACCA
262 AGAGGUGC C AUGAAUUU AAAUUCAU CUGAUGAG X CGAA ICACCUCU
263 GAGGUGCC A UGAAUUUG CAAAUUCA CUGAUGAG X CGAA IGCACCUC
276 UUUGUUAC U UUUUCUUG CAAGAAAA CUGAUGAG X CGAA IUAACAAA
282 ACUUUUUC U UGUCCGGG CCCGGACA CUGAUGAG X CGAA IAAAAAGU
287 UUCUUGUC C GGGUGCGG CCGCACCC CUGAUGAG X CGAA IACAAGAA
305 UAAGGGAC C CGACACUG CAGUGUCG CUGAUGAG X CGAA IUCCCUUA
306 AAGGGACC C GACACUGA UCAGUGUC CUGAUGAG X CGAA IGUCCCUU
310 GACCCGAC A CUGAUGAC GUCAUCAG CUGAUGAG X CGAA IUCGGGUC
312 CCCGACAC U GAUGACCC GGGUCAUC CUGAUGAG X CGAA IUGUCGGG
319 CUGAUGAC C CCAGGAGC GCUCCUGG CUGAUGAG X CGAA IUCAUCAG
320 UGAUGACC C CAGGAGCA UGCUCCUG CUGAUGAG X CGAA IGUCAUCA
321 GAUGACCC C AGGAGCAA UUGCUCCU CUGAUGAG X CGAA IGGUCAUC
322 AUGACCCC A GGAGCAAG CUUGCUCC CUGAUGAG X CGAA IGGGUCAU
328 CCAGGAGC A AGCACAAG CUUGUGCU CUGAUGAG X CGAA ICUCCUGG
332 GAGCAAGC A CAAGUUCA UGAACUUG CUGAUGAG X CGAA ICUUGCUC
334 GCAAGCAC A AGUUCAAA UUUGAACU CUGAUGAG X CGAA IUGCUUGC
340 ACAAGUUC A AAAUCCAC GUGGAUUU CUGAUGAG X CGAA IAACUUGU
346 UCAAAAUC C ACACUUAC GUAAGUGU CUGAUGAG X CGAA lAUUUUGA
347 CAAAAUCC A CACUUACG CGUAAGUG CUGAUGAG X CGAA IGAUUUUG
349 AAAUCCAC A CUUACGGA UCCGUAAG CUGAUGAG X CGAA IUGGAUUU
351 AUCCACAC U UACGGAAG CUUCCGUA CUGAUGAG X CGAA IUGUGGAU
361 ACGGAAGC C CCACCUUC GAAGGUGG CUGAUGAG X CGAA ICUUCCGU
362 CGGAAGCC C CACCUUCU AGAAGGUG CUGAUGAG X CGAA IGCUUCCG
363 GGAAGCCC C ACCUUCUG CAGAAGGU CUGAUGAG X CGAA IGGCUUCC
364 GAAGCCCC A CCUUCUGC GCAGAAGG CUGAUGAG X CGAA IGGGCUUC
366 AGCCCCAC C UUCUGCGA UCGCAGAA CUGAUGAG X CGAA IUGGGGCU
367 GCCCCACC U UCUGCGAU AUCGCAGA CUGAUGAG X CGAA IGUGGGGC
370 CCACCUUC U GCGAUCAC GUGAUCGC CUGAUGAG X CGAA IAAGGUGG
377 CUGCGAUC A CUGUGGGU ACCCACAG CUGAUGAG X CGAA IAUCGCAG
379 GCGAUCAC U GUGGGUCA UGACCCAC CUGAUGAG X CGAA IUGAUCGC
387 UGUGGGUC A CUGCUCUA UAGAGCAG CUGAUGAG X CGAA lACCCACA
389 UGGGUCAC U GCUCUAUG CAUAGAGC CUGAUGAG X CGAA IUGACCCA
392 GUCACUGC U CUAUGGAC GUCCAUAG CUGAUGAG X CGAA ICAGUGAC
394 CACUGCUC U AUGGACUU AAGUCCAU CUGAUGAG X CGAA IAGCAGUG
401 CUAUGGAC U UAUCCAUC GAUGGAUA CUGAUGAG X CGAA IUCCAUAG
406 GACUUAUC C AUCAAGGG CCCUUGAU CUGAUGAG X CGAA IAUAAGUC
407 ACUUAUCC A UCAAGGGA UCCCUUGA CUGAUGAG X CGAA IGAUAAGU
410 UAUCCAUC A AGGGAUGA UCAUCCCU CUGAUGAG X CGAA lAUGGAUA
427 AAUGUGAC A CCUGCGAU AUCGCAGG CUGAUGAG X CGAA IUCACAUU
429 UGUGACAC C UGCGAUAU AUAUCGCA CUGAUGAG X CGAA lUGUCACA
430 GUGACACC U GCGAUAUG CAUAUCGC CUGAUGAG X CGAA IGUGUCAC
446 GAACGUUC A CAAGCAAU AUUGCUUG CUGAUGAG X CGAA IAACGUUC
448 ACGUUCAC A AGCAAUGC GCAUUGCU CUGAUGAG X CGAA IUGAACGU
452 UCACAAGC A AUGCGUCA UGACGCAU CUGAUGAG X CGAA ICUUGUGA
Table 63
460 AAUGCGUC A UCAAUGUC GACAUUGA CUGAUGAG X CGAA IACGCAUU
463 GCGUCAUC A AUGUCCCC GGGGACAU CUGAUGAG X CGAA IAUGACGC
469 UCAAUGUC C CCAGCCUC GAGGCUGG CUGAUGAG X CGAA IACAUUGA
470 CAAUGUCC C CAGCCUCU AGAGGCUG CUGAUGAG X CGAA IGACAUUG
471 AAUGUCCC C AGCCUCUG CAGAGGCU CUGAUGAG X CGAA IGGACAUU
472 AUGUCCCC A GCCUCUGC GCAGAGGC CUGAUGAG X CGAA IGGGACAU
475 UCCCCAGC C UCUGCGGA UCCGCAGA CUGAUGAG X CGAA ICUGGGGA
476 CCCCAGCC U CUGCGGAA UUCCGCAG CUGAUGAG X CGAA IGCUGGGG
478 CCAGCCUC U GCGGAAUG CAUUCCGC CUGAUGAG X CGAA lAGGCUGG
491 AAUGGAUC A CACUGAGA UCUCAGUG CUGAUGAG X CGAA IAUCCAUU
493 UGGAUCAC A CUGAGAAG CUUCUCAG CUGAUGAG X CGAA IUGAUCCA
495 GAUCACAC U GAGAAGAG CUCUUCUC CUGAUGAG X CGAA IUGUGAUC
517 GGAUUUAC C UAAAGGCU AGCCUUUA CUGAUGAG X CGAA IUAAAUCC
518 GAUUUACC U AAAGGCUG CAGCCUUU CUGAUGAG X CGAA IGUAAAUC
525 CUAAAGGC U GAGGUUGC GCAACCUC CUGAUGAG X CGAA ICCUUUAG
534 GAGGUUGC U GAUGAAAA UUUUCAUC CUGAUGAG X CGAA ICAACCUC
545 UGAAAAGC U CCAUGUCA UGACAUGG CUGAUGAG X CGAA ICUUUUCA
547 AAAAGCUC C AUGUCACA UGUGACAU CUGAUGAG X CGAA IAGCUUUU
548 AAAGCUCC A UGUCACAG CUGUGACA CUGAUGAG X CGAA IGAGCUUU
553 UCCAUGUC A CAGUACGA UCGUACUG CUGAUGAG X CGAA IACAUGGA
555 CAUGUCAC A GUACGAGA UCUCGUAC CUGAUGAG X CGAA IUGACAUG
567 CGAGAUGC A AAAAAUCU AGAUUUUU CUGAUGAG X CGAA ICAUCUCG
575 AAAAAAUC U AAUCCCUA UAGGGAUU CUGAUGAG X CGAA IAUUUUUU
580 AUCUAAUC C CUAUGGAU AUCCAUAG CUGAUGAG X CGAA IAUUAGAU
581 UCUAAUCC C UAUGGAUC GAUCCAUA CUGAUGAG X CGAA IGAUUAGA
582 CUAAUCCC U AUGGAUCC GGAUCCAU CUGAUGAG X CGAA IGGAUUAG
590 UAUGGAUC C AAACGGGC GCCCGUUU CUGAUGAG X CGAA IAUCCAUA
591 AUGGAUCC A AACGGGCU AGCCCGUU CUGAUGAG X CGAA IGAUCCAU
599 AAACGGGC U UUCAGAUC GAUCUGAA CUGAUGAG X CGAA ICCCGUUU
603 GGGCUUUC A GAUCCUUA UAAGGAUC CUGAUGAG X CGAA IAAAGCCC
608 UUCAGAUC C UUAUGUGA UCACAUAA CUGAUGAG X CGAA IAUCUGAA
609 UCAGAUCC U UAUGUGAA UUCACAUA CUGAUGAG X CGAA IGAUCUGA
620 UGUGAAGC U GAAACUUA UAAGUUUC CUGAUGAG X CGAA ICUUCACA
626 GCUGAAAC U UAUUCCUG CAGGAAUA CUGAUGAG X CGAA IUUUCAGC
632 ACUUAUUC C UGAUCCCA UGGGAUCA CUGAUGAG X CGAA IAAUAAGU
633 CUUAUUCC U GAUCCCAA UUGGGAUC CUGAUGAG X CGAA IGAAUAAG
638 UCCUGAUC C CAAGAAUG CAUUCUUG CUGAUGAG X CGAA IAUCAGGA
639 CCUGAUCC C AAGAAUGA UCAUUCUU CUGAUGAG X CGAA IGAUCAGG
640 CUGAUCCC A AGAAUGAA UUCAUUCU CUGAUGAG X CGAA IGGAUCAG
652 AUGAAAGC A AGCAAAAA UUUUUGCU CUGAUGAG X CGAA ICUUUCAU
656 AAGCAAGC A AAAAACCA UGGUUUUU CUGAUGAG X CGAA ICUUGCUU
663 CAAAAAAC C AAAACCAU AUGGUUUU CUGAUGAG X CGAA IUUUUUUG
664 AAAAAACC A AAACCAUC GAUGGUUU CUGAUGAG X CGAA IGUUUUUU
669 ACCAAAAC C AUCCGCUC GAGCGGAU CUGAUGAG X CGAA IUUUUGGU
670 CCAAAACC A UCCGCUCC GGAGCGGA CUGAUGAG X CGAA IGUUUUGG
673 AAACCAUC C GCUCCACA UGUGGAGC CUGAUGAG X CGAA IAUGGUUU
676 CCAUCCGC U CCACACUA UAGUGUGG CUGAUGAG X CGAA ICGGAUGG
678 AUCCGCUC C ACACUAAA UUUAGUGU CUGAUGAG X CGAA IAGCGGAU
679 UCCGCUCC A CACUAAAU AUUUAGUG CUGAUGAG X CGAA IGAGCGGA
681 CGCUCCAC A CUAAAUCC GGAUUUAG CUGAUGAG X CGAA IUGGAGCG
683 CUCCACAC U AAAUCCGC GCGGAUUU CUGAUGAG X CGAA IUGUGGAG
Table 63
689 ACUAAAUC C GCAGUGGA UCCACUGC CUGAUGAG X CGAA IAUUUAGU
692 AAAUCCGC A GUGGAAUG CAUUCCAC CUGAUGAG X CGAA ICGGAUUU
705 AAUGAGUC C UUUACAUU AAUGUAAA CUGAUGAG X CGAA IACUCAUU
706 AUGAGUCC U UUACAUUC GAAUGUAA CUGAUGAG X CGAA IGACUCAU
711 UCCUUUAC A UUCAAAUU AAUUUGAA CUGAUGAG X CGAA IUAAAGGA
715 UUACAUUC A AAUUGAAA UUUCAAUU CUGAUGAG X CGAA IAAUGUAA
725 AUUGAAAC C UUCAGACA UGUCUGAA CUGAUGAG X CGAA IUUUCAAU
726 UUGAAACC U UCAGACAA UUGUCUGA CUGAUGAG X CGAA IGUUUCAA
729 AAACCUUC A GACAAAGA UCUUUGUC CUGAUGAG X CGAA IAAGGUUU
733 CUUCAGAC A AAGACCGA UCGGUCUU CUGAUGAG X CGAA IUCUGAAG
739 ACAAAGAC C GACGACUG CAGUCGUC CUGAUGAG X CGAA IUCUUUGU
746 CCGACGAC U GUCUGUAG CUACAGAC CUGAUGAG X CGAA IUCGUCGG
750 CGACUGUC U GUAGAAAU AUUUCUAC CUGAUGAG X CGAA IACAGUCG
760 UAGAAAUC U GGGACUGG CCAGUCCC CUGAUGAG X CGAA IAUUUCUA
766 UCUGGGAC U GGGAUCGA UCGAUCCC CUGAUGAG X CGAA IUCCCAGA
777 GAUCGAAC A ACAAGGAA UUCCUUGU CUGAUGAG X CGAA IUUCGAUC
780 CGAACAAC A AGGAAUGA UCAUUCCU CUGAUGAG X CGAA IUUGUUCG
790 GGAAUGAC U UCAUGGGA UCCCAUGA CUGAUGAG X CGAA IUCAUUCC
793 AUGACUUC A UGGGAUCC GGAUCCCA CUGAUGAG X CGAA IAAGUCAU
801 AUGGGAUC C CUUUCCUU AAGGAAAG CUGAUGAG X CGAA IAUCCCAU
802 UGGGAUCC C UUUCCUUU AAAGGAAA CUGAUGAG X CGAA IGAUCCCA
803 GGGAUCCC U UUCCUUUG CAAAGGAA CUGAUGAG X CGAA IGGAUCCC
807 UCCCUUUC C UUUGGAGU ACUCCAAA CUGAUGAG X CGAA lAAAGGGA
808 CCCUUUCC U UUGGAGUU AACUCCAA CUGAUGAG X CGAA IGAAAGGG
824 UUCGGAGC U GAUGAAGA UCUUCAUC CUGAUGAG X CGAA ICUCCGAA
836 GAAGAUGC C GGCCAGUG CACUGGCC CUGAUGAG X CGAA ICAUCUUC
840 AUGCCGGC C AGUGGAUG CAUCCACU CUGAUGAG X CGAA ICCGGCAU
841 UGCCGGCC A GUGGAUGG CCAUCCAC CUGAUGAG X CGAA IGCCGGCA
853 GAUGGUAC A AGUUGCUU AAGCAACU CUGAUGAG X CGAA IUACCAUC
860 CAAGUUGC U UAACCAAG CUUGGUUA CUGAUGAG X CGAA ICAACUUG
865 UGCUUAAC C AAGAAGAA UUCUUCUU CUGAUGAG X CGAA lUUAAGCA
866 GCUUAACC A AGAAGAAG CUUCUUCU CUGAUGAG X CGAA IGUUAAGC
883 GUGAGUAC U ACAACGUA UACGUUGU CUGAUGAG X CGAA IUACUCAC
886 AGUACUAC A ACGUACCC GGGUACGU CUGAUGAG X CGAA IUAGUACU
893 CAACGUAC C CAUUCCGG CCGGAAUG CUGAUGAG X CGAA IUACGUUG
894 AACGUACC C AUUCCGGA UCCGGAAU CUGAUGAG X CGAA IGUACGUU
895 ACGUACCC A UUCCGGAA UUCCGGAA CUGAUGAG X CGAA IGGUACGU
899 ACCCAUUC C GGAAGGGG CCCCUUCC CUGAUGAG X CGAA IAAUGGGU
922 AAGGAAAC A UGGAACUC GAGUUCCA CUGAUGAG X CGAA IUUUCCUU
929 CAUGGAAC U CAGGCAGA UCUGCCUG CUGAUGAG X CGAA IUUCCAUG
931 UGGAACUC A GGCAGAAA UUUCUGCC CUGAUGAG X CGAA IAGUUCCA
935 ACUCAGGC A GAAAUUCG CGAAUUUC CUGAUGAG X CGAA ICCUGAGU
951 GAGAAAGC C AAACUUGG CCAAGUUU CUGAUGAG X CGAA ICUUUCUC
952 AGAAAGCC A AACUUGGC GCCAAGUU CUGAUGAG X CGAA IGCUUUCU
956 AGCCAAAC U UGGCCCUG CAGGGCCA CUGAUGAG X CGAA UUUGGCU
961 AACUUGGC C CUGCUGGC GCCAGCAG CUGAUGAG X CGAA ICCAAGUU
962 ACUUGGCC C UGCUGGCA UGCCAGCA CUGAUGAG X CGAA IGCCAAGU
963 CUUGGCCC u GCUGGCAA UUGCCAGC CUGAUGAG X CGAA IGGCCAAG
966 GGCCCUGC u GGCAACAA UUGUUGCC CUGAUGAG X CGAA ICAGGGCC
970 CUGCUGGC A ACAAAGUC GACUUUGU CUGAUGAG X CGAA ICCAGCAG
973 CUGGCAAC A AAGUCAUC GAUGACUU CUGAUGAG X CGAA IUUGCCAG
Table 63
979 ACAAAGUC A UCAGUCCC GGGACUGA CUGAUGAG X CGAA IACUUUGU
982 AAGUCAUC A GUCCCUCU AGAGGGAC CUGAUGAG X CGAA IAUGACUU
986 CAUCAGUC C CUCUGAAG CUUCAGAG CUGAUGAG X CGAA IACUGAUG
987 AUCAGUCC C UCUGAAGA UCUUCAGA CUGAUGAG X CGAA IGACUGAU
988 UCAGUCCC U CUGAAGAC GUCUUCAG CUGAUGAG X CGAA IGGACUGA
990 AGUCCCUC U GAAGACAG CUGUCUUC CUGAUGAG X CGAA IAGGGACU
997 CUGAAGAC A GGAAACAA UUGUUUCC CUGAUGAG X CGAA IUCUUCAG
1004 CAGGAAAC A ACCUUCCA UGGAAGGU CUGAUGAG X CGAA IUUUCCUG
1007 GAAACAAC C UUCCAACA UGUUGGAA CUGAUGAG X CGAA IUUGUUUC
1008 AAACAACC U UCCAACAA UUGUUGGA CUGAUGAG X CGAA IGUUGUUU
1011 CAACCUUC C AACAACCU AGGUUGUU CUGAUGAG X CGAA IAAGGUUG
1012 AACCUUCC A ACAACCUU AAGGUUGU CUGAUGAG X CGAA IGAAGGUU
1015 CUUCCAAC A ACCUUGAC GUCAAGGU CUGAUGAG X CGAA IUUGGAAG
1018 CCAACAAC C UUGACCGA UCGGUCAA CUGAUGAG X CGAA IUUGUUGG
1019 CAACAACC U UGACCGAG CUCGGUCA CUGAUGAG X CGAA IGUUGUUG
1024 ACCUUGAC C GAGUGAAA UUUCACUC CUGAUGAG X CGAA IUCAAGGU
1034 AGUGAAAC U CACGGACU AGUCCGUG CUGAUGAG X CGAA IUUUCACU
1036 UGAAACUC A CGGACUUC GAAGUCCG CUGAUGAG X CGAA IAGUUUCA
1042 UCACGGAC U UCAAUUUC GAAAUUGA CUGAUGAG X CGAA IUCCGUGA
1045 CGGACUUC A AUUUCCUC GAGGAAAU CUGAUGAG X CGAA IAAGUCCG
1051 UCAAUUUC C UCAUGGUG CACCAUGA CUGAUGAG X CGAA IAAAUUGA
1052 CAAUUUCC U CAUGGUGU ACACCAUG CUGAUGAG X CGAA IGAAAUUG
1054 AUUUCCUC A UGGUGUUG CAACACCA CUGAUGAG X CGAA IAGGAAAU
■ 1091 GGUGAUGC U UGCCGACA UGUCGGCA CUGAUGAG X CGAA ICAUCACC
1095 AUGCUUGC C GACAGGAA UUCCUGUC CUGAUGAG X CGAA ICAAGCAU
1099 UUGCCGAC A GGAAGGGC GCCCUUCC CUGAUGAG X CGAA IUCGGCAA
1108 GGAAGGGC A CAGAAGAA UUCUUCUG CUGAUGAG X CGAA ICCCUUCC
1110 AAGGGCAC A GAAGAACU AGUUCUUC CUGAUGAG X CGAA IUGCCCUU
1118 AGAAGAAC U GUAUGCAA UUGCAUAC CUGAUGAG X CGAA lUUCUUCU
1125 CUGUAUGC A AUCAAAAU AUUUUGAU CUGAUGAG X CGAA ICAUACAG
1129 AUGCAAUC A AAAUCCUG CAGGAUUU CUGAUGAG X CGAA IAUUGCAU
1135 UCAAAAUC C UGAAGAAG CUUCUUCA CUGAUGAG X CGAA lAUUUUGA
1136 CAAAAUCC U GAAGAAGG CCUUCUUC CUGAUGAG X CGAA IGAUUUUG
1157 GGUGAUUC A GGAUGAUG CAUCAUCC CUGAUGAG X CGAA IAAUCACC
1177 UGGAGUGC A CCAUGGUA UACCAUGG CUGAUGAG X CGAA ICACUCCA
1179 GAGUGCAC C AUGGUAGA UCUACCAU CUGAUGAG X CGAA IUGCACUC
1180 AGUGCACC A UGGUAGAA UUCUACCA CUGAUGAG X CGAA IGUGCACU
1198 AGCGAGUC U UGGCCCUG CAGGGCCA CUGAUGAG X CGAA IACUCGCU
1203 GUCUUGGC C CUGCUUGA UCAAGCAG CUGAUGAG X CGAA ICCAAGAC
1204 UCUUGGCC C UGCUUGAC GUCAAGCA CUGAUGAG X CGAA IGCCAAGA
1205 CUUGGCCC U GCUUGACA UGUCAAGC CUGAUGAG X CGAA IGGCCAAG
1208 GGCCCUGC U UGACAAAC GUUUGUCA CUGAUGAG X CGAA ICAGGGCC
1213 UGCUUGAC A AACCCCCG CGGGGGUU CUGAUGAG X CGAA IUCAAGCA
1217 UGACAAAC C CCCGUUCU AGAACGGG CUGAUGAG X CGAA IUUUGUCA
1218 GACAAACC C CCGUUCUU AAGAACGG CUGAUGAG X CGAA IGUUUGUC
1219 ACAAACCC C CGUUCUUG CAAGAACG CUGAUGAG X CGAA IGGUUUGU
1220 CAAACCCC C GUUCUUGA UCAAGAAC CUGAUGAG X CGAA IGGGUUUG
1225 CCCCGUUC U UGACGCAG CUGCGUCA CUGAUGAG X CGAA IAACGGGG
1232 CUUGACGC A GCUGCACU AGUGCAGC CUGAUGAG X CGAA ICGUCAAG
1235 GACGCAGC U GCACUCCU AGGAGUGC CUGAUGAG X CGAA ICUGCGUC
1238 GCAGCUGC A CUCCUGCU AGCAGGAG CUGAUGAG X CGAA ICAGCUGC
Table 63
1240 AGCUGCAC U CCUGCUUC GAAGCAGG CUGAUGAG X CGAA IUGCAGCU
1242 CUGCACUC C UGCUUCCA UGGAAGCA CUGAUGAG X CGAA IAGUGCAG
1243 UGCACUCC U GCUUCCAG CUGGAAGC CUGAUGAG X CGAA IGAGUGCA
1246 ACUCCUGC U UCCAGACA UGUCUGGA CUGAUGAG X CGAA ICAGGAGU
1249 CCUGCUUC C AGACAGUG CACUGUCU CUGAUGAG X CGAA IAAGCAGG
1250 CUGCUUCC A GACAGUGG CCACUGUC CUGAUGAG X CGAA IGAAGCAG
1254 UUCCAGAC A GUGGAUCG CGAUCCAC CUGAUGAG X CGAA IUCUGGAA
1265 GGAUCGGC U GUACUUCG CGAAGUAC CUGAUGAG X CGAA ICCGAUCC
1270 GGCUGUAC U UCGUCAUG CAUGACGA CUGAUGAG X CGAA IUACAGCC
1276 ACUUCGUC A UGGAAUAU AUAUUCCA CUGAUGAG X CGAA IACGAAGU
1288 AAUAUGUC A ACGGUGGG CCCACCGU CUGAUGAG X CGAA IACAUAUU
1300 GUGGGGAC C UCAUGUAC GUACAUGA CUGAUGAG X CGAA IUCCCCAC
1301 UGGGGACC U CAUGUACC GGUACAUG CUGAUGAG X CGAA IGUCCCCA
1303 GGGACCUC A UGUACCAC GUGGUACA CUGAUGAG X CGAA IAGGUCCC
1309 UCAUGUAC C ACAUUCAG CUGAAUGU CUGAUGAG X CGAA IUACAUGA
1310 CAUGUACC A CAUUCAGC GCUGAAUG CUGAUGAG X CGAA IGUACAUG
1312 UGUACCAC A UUCAGCAA UUGCUGAA CUGAUGAG X CGAA lUGGUACA
1316 CCACAUUC A GCAAGUAG CUACUUGC CUGAUGAG X CGAA IAAUGUGG
1319 CAUUCAGC A AGUAGGAA UUCCUACU CUGAUGAG X CGAA ICUGAAUG
1340 UAAGGAAC C ACAAGCAG CUGCUUGU CUGAUGAG X CGAA IUUCCUUA
1341 AAGGAACC A CAAGCAGU ACUGCUUG CUGAUGAG X CGAA IGUUCCUU
1343 GGAACCAC A AGCAGUAU AUACUGCU CUGAUGAG X CGAA IUGGUUCC
1347 CCACAAGC A GUAUUCUA UAGAAUAC CUGAUGAG X CGAA ICUUGUGG
1354 CAGUAUUC U AUGCGGCA UGCCGCAU CUGAUGAG X CGAA IAAUACUG
1362 UAUGCGGC A GAGAUUUC GAAAUCUC CUGAUGAG X CGAA ICCGCAUA
1371 GAGAUUUC C AUCGGAUU AAUCCGAU CUGAUGAG X CGAA IAAAUCUC
1372 AGAUUUCC A UCGGAUUG CAAUCCGA CUGAUGAG X CGAA IGAAAUCU
1384 GAUUGUUC U UUCUUCAU AUGAAGAA CUGAUGAG X CGAA IAACAAUC
1388 GUUCUUUC U UCAUAAAA UUUUAUGA CUGAUGAG X CGAA IAAAGAAC
1391 CUUUCUUC A UAAAAGAG CUCUUUUA CUGAUGAG X CGAA IAAGAAAG
1405 GAGGAAUC A UUUAUAGG CCUAUAAA CUGAUGAG X CGAA IAUUCCUC
1418 UAGGGAUC U GAAGUUAG CUAACUUC CUGAUGAG X CGAA IAUCCCUA
1435 AUAACGUC A UGUUGGAU AUCCAACA CUGAUGAG X CGAA IACGUUAU
1446 UUGGAUUC A GAAGGACA UGUCCUUC CUGAUGAG X CGAA IAAUCCAA
1454 AGAAGGAC A UAUCAAAA UUUUGAUA CUGAUGAG X CGAA IUCCUUCU
1459 GACAUAUC A AAAUUGCU AGCAAUUU CUGAUGAG X CGAA IAUAUGUC
1467 AAAAUUGC U GACUUUGG CCAAAGUC CUGAUGAG X CGAA ICAAUUUU
1471 UUGCUGAC U UUGGGAUG CAUCCCAA CUGAUGAG X CGAA IUCAGCAA
1483 GGAUGUGC A AGGAACAC GUGUUCCU CUGAUGAG X CGAA ICACAUCC
1490 CAAGGAAC A CAUGAUGG CCAUCAUG CUGAUGAG X CGAA IUUCCUUG
1492 AGGAACAC A UGAUGGAU AUCCAUCA CUGAUGAG X CGAA lUGUUCCU
1507 AUGGAGUC A CGACCAGG CCUGGUCG CUGAUGAG X CGAA IACUCCAU
1512 GUCACGAC C AGGACCUU AAGGUCCU CUGAUGAG X CGAA IUCGUGAC
1513 UCACGACC A GGACCUUC GAAGGUCC CUGAUGAG X CGAA IGUCGUGA
1518 ACCAGGAC C UUCUGUGG CCACAGAA CUGAUGAG X CGAA IUCCUGGU
1519 CCAGGACC U UCUGUGGG CCCACAGA CUGAUGAG X CGAA IGUCCUGG
1522 GGACCUUC U GUGGGACU AGUCCCAC CUGAUGAG X CGAA IAAGGUCC
1530 UGUGGGAC U CCAGAUUA UAAUCUGG CUGAUGAG x- CGAA IUCCCACA
1532 UGGGACUC C AGAUUAUA UAUAAUCU CUGAUGAG X CGAA IAGUCCCA
1533 GGGACUCC A GAUUAUAU AUAUAAUC CUGAUGAG X CGAA IGAGUCCC
1545 UAUAUCGC C CCAGAGAU AUCUCUGG CUGAUGAG X CGAA ICGAUAUA
Table 63
1546 AUAUCGCC C CAGAGAUA UAUCUCUG CUGAUGAG X CGAA IGCGAUAU
1547 UAUCGCCC C AGAGAUAA UUAUCUCU CUGAUGAG X CGAA IGGCGAUA
1548 AUCGCCCC A GAGAUAAU AUUAUCUC CUGAUGAG X CGAA IGGGCGAU
1560 AUAAUCGC U UAUCAGCC GGCUGAUA CUGAUGAG X CGAA ICGAUUAU
1565 CGCUUAUC A GCCGUAUG CAUACGGC CUGAUGAG X CGAA IAUAAGCG
1568 UUAUCAGC C GUAUGGAA UUCCAUAC CUGAUGAG X CGAA ICUGAUAA
1581 GGAAAAUC U GUGGACUG CAGUCCAC CUGAUGAG X CGAA IAUUUUCC
1588 CUGUGGAC U GGUGGGCC GGCCCACC CUGAUGAG X CGAA IUCCACAG
1596 UGGUGGGC C UAUGGCGU ACGCCAUA CUGAUGAG X CGAA ICCCACCA
1597 GGUGGGCC U AUGGCGUC GACGCCAU CUGAUGAG X CGAA IGCCCACC
1606 AUGGCGUC C UGUUGUAU AUACAACA CUGAUGAG X CGAA IACGCCAU
1607 UGGCGUCC U GUUGUAUG CAUACAAC CUGAUGAG X CGAA IGACGCCA
1622 UGAAAUGC u UGCCGGGC GCCCGGCA CUGAUGAG X CGAA ICAUUUCA
1626 AUGCUUGC c GGGCAGCC GGCUGCCC CUGAUGAG X CGAA ICAAGCAU
1631 UGCCGGGC A GCCUCCAU AUGGAGGC CUGAUGAG X CGAA ICCCGGCA
1634 CGGGCAGC C UCCAUUUG CAAAUGGA CUGAUGAG X CGAA ICUGCCCG
1635 GGGCAGCC U CCAUUUGA UCAAAUGG CUGAUGAG X CGAA IGCUGCCC
1637 GCAGCCUC C AUUUGAUG CAUCAAAU CUGAUGAG X CGAA IAGGCUGC
1638 CAGCCUCC A UUUGAUGG CCAUCAAA CUGAUGAG X CGAA IGAGGCUG
1664 AGACGAGC U AUUUCAGU ACUGAAAU CUGAUGAG X CGAA ICUCGUCU
1670 GCUAUUUC A GUCUAUCA UGAUAGAC CUGAUGAG X CGAA IAAAUAGC
1674 UUUCAGUC U AUCAUGGA UCCAUGAU CUGAUGAG X CGAA lACUGAAA
1678 AGUCUAUC A UGGAGCAC GUGCUCCA CUGAUGAG X CGAA IAUAGACU
1685 CAUGGAGC A CAACGUUU AAACGUUG CUGAUGAG X CGAA ICUCCAUG
1687 UGGAGCAC A ACGUUUCC GGAAACGU CUGAUGAG X CGAA IUGCUCCA
1695 AACGUUUC C UAUCCAAA UUUGGAUA CUGAUGAG X CGAA IAAACGUU
1696 ACGUUUCC U AUCCAAAA UUUUGGAU CUGAUGAG X CGAA IGAAACGU
1700 UUCCUAUC C AAAAUCCU AGGAUUUU CUGAUGAG X CGAA IAUAGGAA
1701 UCCUAUCC A AAAUCCUU AAGGAUUU CUGAUGAG X CGAA IGAUAGGA
1707 CCAAAAUC C UUGUCCAA UUGGACAA CUGAUGAG X CGAA IAUUUUGG
1708 CAAAAUCC U UGUCCAAG CUUGGACA CUGAUGAG X CGAA IGAUUUUG
1713 UCCUUGUC C AAGGAGGC GCCUCCUU CUGAUGAG X CGAA IACAAGGA
1714 CCUUGUCC A AGGAGGCU AGCCUCCU CUGAUGAG X CGAA IGACAAGG
1722 AAGGAGGC U GUUUCUAU AUAGAAAC CUGAUGAG X CGAA ICCUCCUU
1728 GCUGUUUC U AUCUGCAA UUGCAGAU CUGAUGAG X CGAA IAAACAGC
1732 UUUCUAUC U GCAAAGGA UCCUUUGC CUGAUGAG X CGAA lAUAGAAA
1735 CUAUCUGC A AAGGACUG CAGUCCUU CUGAUGAG X CGAA ICAGAUAG
1742 CAAAGGAC U GAUGACCA UGGUCAUC CUGAUGAG X CGAA IUCCUUUG
1749 CUGAUGAC C AAACACCC GGGUGUUU CUGAUGAG X CGAA IUCAUCAG
1750 UGAUGACC A AACACCCA UGGGUGUU CUGAUGAG X CGAA IGUCAUCA
1754 GACCAAAC A CCCAGCCA UGGCUGGG CUGAUGAG X CGAA IUUUGGUC
1756 CCAAACAC C CAGCCAAG CUUGGCUG CUGAUGAG X CGAA IUGUUUGG
1757 CAAACACC C AGCCAAGC GCUUGGCU CUGAUGAG X CGAA IGUGUUUG
1758 AAACACCC A GCCAAGCG CGCUUGGC CUGAUGAG X CGAA IGGUGUUU
1761 CACCCAGC C AAGCGGCU AGCCGCUU CUGAUGAG X CGAA ICUGGGUG
1762 ACCCAGCC A AGCGGCUG CAGCCGCU CUGAUGAG X CGAA IGCUGGGU
1769 CAAGCGGC U GGGCUGUG CACAGCCC CUGAUGAG X CGAA ICCGCUUG
1774 GGCUGGGC U GUGGGCCU AGGCCCAC CUGAUGAG X CGAA ICCCAGCC
1781 CUGUGGGC C UGAGGGGG CCCCCUCA CUGAUGAG X CGAA ICCCACAG
1782 UGUGGGCC U GAGGGGGA UCCCCCUC CUGAUGAG X CGAA IGCCCACA
1808 GAGAGAGC A UGCCUUCU AGAAGGCA CUGAUGAG X CGAA ICUCUCUC
Table 63
1812 GAGCAUGC C UUCUUCCG CGGAAGAA CUGAUGAG X CGAA ICAUGCUC
1813 AGCAUGCC U UCUUCCGG CCGGAAGA CUGAUGAG X CGAA IGCAUGCU
1816 AUGCCUUC U UCCGGAGG CCUCCGGA CUGAUGAG X CGAA IAAGGCAU
1819 CCUUCUUC C GGAGGAUC GAUCCUCC CUGAUGAG X CGAA IAAGAAGG
1831 GGAUCGAC U GGGAAAAA UUUUUCCC CUGAUGAG X CGAA IUCGAUCC
1841 GGAAAAAC U GGAGAACA UGUUCUCC CUGAUGAG X CGAA IUUUUUCC
1849 UGGAGAAC A GGGAGAUC GAUCUCCC CUGAUGAG X CGAA lUUCUCCA
1858 GGGAGAUC C AGCCACCA UGGUGGCU CUGAUGAG X CGAA IAUCUCCC
1859 GGAGAUCC A GCCACCAU AUGGUGGC CUGAUGAG X CGAA IGAUCUCC
1862 GAUCCAGC C ACCAUUCA UGAAUGGU CUGAUGAG X CGAA ICUGGAUC
1863 AUCCAGCC A CCAUUCAA UUGAAUGG CUGAUGAG X CGAA IGCUGGAU
1865 CCAGCCAC C AUUCAAGC GCUUGAAU CUGAUGAG X CGAA IUGGCUGG
1866 CAGCCACC A UUCAAGCC GGCUUGAA CUGAUGAG X CGAA IGUGGCUG
1870 CACCAUUC A AGCCCAAA UUUGGGCU CUGAUGAG X CGAA IAAUGGUG
1874 AUUCAAGC C CAAAGUGU ACACUUUG CUGAUGAG X CGAA ICUUGAAU
1875 UUCAAGCC C AAAGUGUG CACACUUU CUGAUGAG X CGAA IGCUUGAA
1876 UCAAGCCC A AAGUGUGU ACACACUU CUGAUGAG X CGAA IGGCUUGA
1888 UGUGUGGC A AAGGAGCA UGCUCCUU CUGAUGAG X CGAA ICCACACA
1896 AAAGGAGC A GAGAACUU AAGUUCUC CUGAUGAG X CGAA ICUCCUUU
1903 CAGAGAAC U UUGACAAG CUUGUCAA CUGAUGAG X CGAA IUUCUCUG
1909 ACUUUGAC A AGUUCUUC GAAGAACU CUGAUGAG X CGAA IUCAAAGU
1915 ACAAGUUC U UCACACGA UCGUGUGA CUGAUGAG X CGAA IAACUUGU
1918 AGUUCUUC A CACGAGGA UCCUCGUG CUGAUGAG X CGAA IAAGAACU
1920 UUCUUCAC A CGAGGACA UGUCCUCG CUGAUGAG X CGAA IUGAAGAA
1928 ACGAGGAC A GCCCGUCU AGACGGGC CUGAUGAG X CGAA IUCCUCGU
1931 AGGACAGC C CGUCUUAA UUAAGACG CUGAUGAG X CGAA ICUGUCCU
1932 GGACAGCC C GUCUUAAC GUUAAGAC CUGAUGAG X CGAA IGCUGUCC
1936 AGCCCGUC U UAACACCA UGGUGUUA CUGAUGAG X CGAA IACGGGCU
1941 GUCUUAAC A CCACCUGA UCAGGUGG CUGAUGAG X CGAA IUUAAGAC
1943 CUUAACAC C ACCUGAUC GAUCAGGU CUGAUGAG X CGAA IUGUUAAG
1944 UUAACACC A CCUGAUCA UGAUCAGG CUGAUGAG X CGAA IGUGUUAA
1946 AACACCAC C UGAUCAGC GCUGAUCA CUGAUGAG X CGAA IUGGUGUU
1947 ACACCACC U GAUCAGCU AGCUGAUC CUGAUGAG X CGAA IGUGGUGU
1952 ACCUGAUC A GCUGGUUA UAACCAGC CUGAUGAG X CGAA IAUCAGGU
1955 UGAUCAGC U GGUUAUUG CAAUAACC CUGAUGAG X CGAA ICUGAUCA
1965 GUUAUUGC U AACAUAGA UCUAUGUU CUGAUGAG X CGAA ICAAUAAC
1969 UUGCUAAC A UAGACCAG CUGGUCUA CUGAUGAG X CGAA IUUAGCAA
1975 ACAUAGAC C AGUCUGAU AUCAGACU CUGAUGAG X CGAA IUCUAUGU
1976 CAUAGACC A GUCUGAUU AAUCAGAC CUGAUGAG X CGAA IGUCUAUG
1980 GACCAGUC U GAUUUUGA UCAAAAUC CUGAUGAG X CGAA IACUGGUC
1996 AAGGGUUC U CGUAUGUC GACAUACG CUGAUGAG X CGAA IAACCCUU
2005 CGUAUGUC A ACCCCCAG CUGGGGGU CUGAUGAG X CGAA IACAUACG
2008 AUGUCAAC C CCCAGUUU AAACUGGG CUGAUGAG X CGAA IUUGACAU
2009 UGUCAACC C CCAGUUUG CAAACUGG CUGAUGAG X CGAA IGUUGACA
2010 GUCAACCC C CAGUUUGU ACAAACUG CUGAUGAG X CGAA IGGUUGAC
2011 UCAACCCC C AGUUUGUG CACAAACU CUGAUGAG X CGAA IGGGUUGA
2012 CAACCCCC A GUUUGUGC GCACAAAC CUGAUGAG X CGAA IGGGGUUG
2021 GUUUGUGC A CCCCAUCU AGAUGGGG CUGAUGAG X CGAA ICACAAAC
2023 UUGUGCAC C CCAUCUUA UAAGAUGG CUGAUGAG X CGAA IUGCACAA
2024 UGUGCACC C CAUCUUAC GUAAGAUG CUGAUGAG X CGAA IGUGCACA
2025 GUGCACCC C AUCUUACA UGUAAGAU CUGAUGAG X CGAA IGGUGCAC
Table 63
2026 UGCACCCC A UCUUACAG CUGUAAGA CUGAUGAG X CGAA IGGGUGCA
2029 ACCCCAUC U UACAGAGU ACUCUGUA CUGAUGAG X CGAA IAUGGGGU
2033 CAUCUUAC A GAGUGCAG CUGCACUC CUGAUGAG X CGAA IUAAGAUG
2040 CAGAGUGC A GUAUGAAA UUUCAUAC CUGAUGAG X CGAA ICACUCUG
2050 UAUGAAAC U CACCAGCG CGCUGGUG CUGAUGAG X CGAA IUUUCAUA
2052 UGAAACUC A CCAGCGAG CUCGCUGG CUGAUGAG X CGAA IAGUUUCA
2054 AAACUCAC C AGCGAGAA UUCUCGCU CUGAUGAG X CGAA IUGAGUUU
2055 AACUCACC A GCGAGAAC GUUCUCGC CUGAUGAG X CGAA IGUGAGUU
2064 GCGAGAAC A AACACCUC GAGGUGUU CUGAUGAG X CGAA IUUCUCGC
2068 GAACAAAC A CCUCCCCA UGGGGAGG CUGAUGAG X CGAA IUUUGUUC
2070 ACAAACAC C UCCCCAGC GCUGGGGA CUGAUGAG X CGAA IUGUUUGU
2071 CAAACACC U CCCCAGCC GGCUGGGG CUGAUGAG X CGAA IGUGUUUG
2073 AACACCUC C CCAGCCCC GGGGCUGG CUGAUGAG X CGAA IAGGUGUU
2074 ACACCUCC C CAGCCCCC GGGGGCUG CUGAUGAG X CGAA IGAGGUGU
2075 CACCUCCC C AGCCCCCA UGGGGGCU CUGAUGAG X CGAA IGGAGGUG
2076 ACCUCCCC A GCCCCCAG CUGGGGGC CUGAUGAG X CGAA IGGGAGGU
2079 UCCCCAGC C CCCAGCCC GGGCUGGG CUGAUGAG X CGAA ICUGGGGA
2080 CCCCAGCC C CCAGCCCU AGGGCUGG CUGAUGAG X CGAA IGCUGGGG
2081 CCCAGCCC C CAGCCCUC GAGGGCUG CUGAUGAG X CGAA IGGCUGGG
2082 CCAGCCCC C AGCCCUCC GGAGGGCU CUGAUGAG X CGAA IGGGCUGG
2083 CAGCCCCC A GCCCUCCC GGGAGGGC CUGAUGAG X CGAA IGGGGCUG
2086 CCCCCAGC C CUCCCCGC GCGGGGAG CUGAUGAG X CGAA ICUGGGGG
2087 CCCCAGCC C UCCCCGCA UGCGGGGA CUGAUGAG X CGAA IGCUGGGG
2088 CCCAGCCC U CCCCGCAG CUGCGGGG CUGAUGAG X CGAA IGGCUGGG
2090 CAGCCCUC C CCGCAGUG CACUGCGG CUGAUGAG X CGAA IAGGGCUG
2091 AGCCCUCC C CGCAGUGG CCACUGCG CUGAUGAG X CGAA IGAGGGCU
2092 GCCCUCCC C GCAGUGGA UCCACUGC CUGAUGAG X CGAA IGGAGGGC
2095 CUCCCCGC A GUGGAAGU ACUUCCAC CUGAUGAG X CGAA ICGGGGAG
2109 AGUGAAUC C UUAACCCU AGGGUUAA CUGAUGAG X CGAA IAUUCACU
2110 GUGAAUCC U UAACCCUA UAGGGUUA CUGAUGAG X CGAA IGAUUCAC
2115 UCCUUAAC C CUAAAAUU AAUUUUAG CUGAUGAG X CGAA lUUAAGGA
2116 CCUUAACC C UAAAAUUU AAAUUUUA CUGAUGAG X CGAA IGUUAAGG
2117 CUUAACCC U AAAAUUUU AAAAUUUU CUGAUGAG X CGAA IGGUUAAG
2131 UUUAAGGC C ACGGCUUG CAAGCCGU CUGAUGAG X CGAA ICCUUAAA
2132 UUAAGGCC A CGGCUUGU ACAAGCCG CUGAUGAG X CGAA IGCCUUAA
2137 GCCACGGC U UGUGUCUG CAGACACA CUGAUGAG X CGAA ICCGUGGC
2144 CUUGUGUC U GAUUCCAU AUGGAAUC CUGAUGAG X CGAA IACACAAG
2150 UCUGAUUC C AUAUGGAG CUCCAUAU CUGAUGAG X CGAA IAAUCAGA
2151 CUGAUUCC A UAUGGAGG CCUCCAUA CUGAUGAG X CGAA IGAAUCAG
2161 AUGGAGGC C UGAAAAUU AAUUUUCA CUGAUGAG X CGAA ICCUCCAU
2162 UGGAGGCC U GAAAAUUG CAAUUUUC CUGAUGAG X CGAA IGCCUCCA
2185 UAUUAGUC C AAAUGUGA UCACAUUU CUGAUGAG X CGAA IACUAAUA
2186 AUUAGUCC A AAUGUGAU AUCACAUU CUGAUGAG X CGAA IGACUAAU
2196 AUGUGAUC A ACUGUUCA UGAACAGU CUGAUGAG X CGAA IAUCACAU
2199 UGAUCAAC U GUUCAGGG CCCUGAAC CUGAUGAG X CGAA IUUGAUCA
2204 AACUGUUC A GGGUCUCU AGAGACCC CUGAUGAG X CGAA IAACAGUU
2210 UCAGGGUC U CUCUCUUA UAAGAGAG CUGAUGAG X CGAA lACCCUGA
2212 AGGGUCUC U CUCUUACA UGUAAGAG CUGAUGAG X CGAA IAGACCCU
2214 GGUCUCUC U CUUACAAC GUUGUAAG CUGAUGAG X CGAA IAGAGACC
2216 UCUCUCUC U UACAACCA UGGUUGUA CUGAUGAG X CGAA IAGAGAGA
2220 UCUCUUAC A ACCAAGAA UUCUUGGU CUGAUGAG X CGAA IUAAGAGA
Table 63
Input Sequence = PRKCA. Cut Site = CH/ .
Stem Length = 8 . Core Sequence = CUGAUGAG X CGAA (X = GCCGUUAGGC or other stem II)
PRKCA (Homo sapiens protein kinase C, alpha (PRKCA) mRNA. ; 2245 bp)
Table 64
Table 64: Activity of ribozyme core substituted analogues
B
Comparison of single turnover cleavage rates for GCH and GUH substrates with 1-15.1 and A-15.1 ribozymes and ribozyme analogs. Conditions: Single turnover (250 nM substrate, 2.5 μM ribozyme) pH 6.0, 37 C, 10 mM Mg++